macrocystis pyrifera a preliminary report brian s ... · 114 brian s. anderson, john w. hunt...
TRANSCRIPT

Marine Envirionmental Research 26 (1988) 113-134
Bioassay Methods for Evaluating the Toxicity of Heavy Metals, Biocides and Sewage Etfluent using Microscopic
Stages of Giant Kelp Macrocystis pyrifera (Agardh): A Preliminary Report
Brian S. Anderson & John W. Hunt
Institute for Marine Studies, University of California Santa Cruz, Santa Cruz, CA 95064, USA, and Marine Pollution Studies Laboratory,
Granite Canyon, Coast Rte 1, Monterey, CA 93940, USA
(Received 10 January 1988; revised version received 5 March 1988; accepted 30 July 1988)
A B S T R A C T
Methods for using early life stages of giant kelp Macrocystis pyrifera in toxicity tests intended .for evaluating the toxicity of municipal sewage eff-luen ts are given. Included are methods for both a short-term (48-h) toxicity test, and a long-term (16-day) toxicity test. Preliminary results oJ" experiments with zinc sulfate, sodium pentachlorophenate, and a primary- treated sewage effluent showed that early life stages of Macrocystis have a variable sensitivity to these toxicants. No Observed Effect Concentrations ( NO ECs )for effects of zinc sulfate on germination of Macrocystis zoospores ranged from 1730 lag/liter to 5500 lag/liter in 3 separate 48-h experiments. The NOECs for zinc effects on the growth of Macrocystis zoospore germination tubes ranged from 550 to 1090 lag~liter in three 48-h experiments. The NOEC for zinc effects on sporophyte production by Macrocystis was 1070 lag/liter in a 16-day experiment. The .fungicide and herbicide sodium pentachlorophenate significantly inhibited Macrocystis zoospore germin- ation, and gametophyte reproduction at concentrations of 32lag/liter and greater ( NOEC < 32 lag/liter for both tests), and was more toxic to kelp than zinc. Primary-treated-sewage effluent significantly inhibited zoospore germination at concentrations greater than 1% effluent ( NOEC = 0.56 %
113
Marine Environ. Res. 0141-I 136/88/$03"50 ~ 1988 Elsevier Science Publishers Ltd, England. Printed in Great Britain

114 Brian S. Anderson, John W. Hunt
effluent). The results of these preliminary experiments indicate that early life stages of Macrocystis pyrifera are amenable to evaluating the toxicity of a variety of toxicants, including sewage effluents, and that Macrocystis has a variable sensitivity to toxicants depending on the nature of the toxicant and the endpoint being tested.
I N T R O D U C T I O N
The giant kelp, Macrocystispyrifera is the dominant canopy-forming alga in southern and central California. Giant kelp forms extensive submarine forests along the coast that harbor a rich diversity of marine life. Macrocystis forests are complex ecosystems and therefore provide a valuable economic and ecological resource.
There is concern over the effects of coastal waste discharge on Macrocystis and its associated marine communities as there is reasonable indirect evidence that sewage has contributed to declines in kelp forests in southern California. Wilson et al. (1980) described a correlation between increased solids discharged by the county of Los Angeles at the Whites Point sewage outfall, and the disappearance of kelp forests on the Palos Verdes Peninsula in the late nineteen-sixties. This decline was followed by a reappearance of kelp with the concurrent decrease in suspended solids discharged (see also Meistrell & Montagne 1983, Fig. 7). Explanations for the disappearance of kelp forests in southern California are confounded by the influence of other factors such as warm water temperatures, and sea urchin grazing. Declines in the Point Loma kelp forest have been attributed to these factors (North, 1976).
Several factors related to wastewater discharge have been suggested to have adverse effects on Macrocystis. These include decreased irradiance due to the presence of suspended solids, increased sedimentation, and poisoning by toxic chemicals. While not demonstrating a direct effect on kelp, Eppley et al. (1972) showed that there was increased sedimentation and decreased irradiance around ocean outfalls. Devinny & Volse (1978) demonstrated in laboratory experiments that suspended solids can kill Macrocystis repro- ductive stages. Others (Luning & Neushul, 1978; Deysher & Dean, 1984, 1986a,b; Dean & Jacobsen, 1984) have shown that light is important for kelp reproduction and sporophyte growth, and Grigg (1978) suggested that the potential area for growth of M. pyriJ'era off Palos Verdes was reduced by low light levels associated with the Whites Point sewage outfall. No direct correlations have been demonstrated between concentrations of toxicants associated with sewage discharge and declines in Macrocystis field populations. Clendenning (1958, 1959, 1960) used laboratory experiments to show that elevated concentrations of sewage effluents and various chemical

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 115
compounds associated with them inhibit photosynthesis in Macrocystis blades. Smith & Harrison (1978) found that copper inhibits the growth of Macrocystis gametophytes in the laboratory; copper is found in high concentrations in some sewage effluents (Schafer 1980, 1982, 1984). Because effluent constituents have a demonstrated effect on kelp, it is possible that declines in kelp beds near outfalls may be due to waste discharges.
This paper presents methods and results ofbioassay tests being developed for Macrocystis pyrifera at the California State Water Resources Control Board's Marine Bioassay Project (Anderson et al., 1988). These tests are being designed to evaluate the effect of toxicants on germination, growth, and fertilization of microscopic stages of Macrocystis, and are intended for eventual use in monitoring the toxicity of sewage effluents being discharged into California waters. We selected Macroc3,stis as a macroalgal bioassay species because of its economic and ecological importance, and it can be easily cultured in the laboratory. The life history of Macrocystis has been well described (North 1971, Luning & Neushul 1978, Deysher & Dean 1984, see review by Foster & Schiel 1985), but its use in bioassay tests has been limited (Clendenning 1958-1960, and Smith & Harrison, 1978). There are, however, studies on the trace metal requirements of early life stages of kelp by Kuwabara (1980, 1981), and Kuwabara & North (1980) that are applicable to the interpretation of bioassay results. Early life stage bioassay tests have been developed for another laminarian alga, Laminaria saccharina (Chung & Brinkuis, 1986).
The results of preliminary experiments with two reference toxicants, zinc sulfate and sodium pentachlorophenate, and a sewage treatment plant effluent are presented here.
METHODS
Facilities
All of the experiments were done between June, 1986 and July, 1987 at the California Department of Fish and Game's Marine Pollution Studies Laboratory (MPSL) at Granite Canyon. The laboratory is located on the Big Sur coast in Monterey County. The MPSL seawater intake is located at least 4 km from any known pollution source.
Life cycle
We used the early life stages of Macrocystis pyr(fera for all the experiments. Like all laminarian algae, Macrocvstis alternates between a microscopic

116 Brian S. Anderson, John W. Hunt
haploid gametophyte stage and a macroscopic diploid sporophyte stage. It is the sporophyte stage that forms the giant kelp forests. These plants produce reproductive blades, called sporophylls, at their base. The sporophylls develop patches (sori) in which biflagellate haploid spores are produced. The spores are called zoospores because they swim. The zoospores are released into the water column and eventually settle onto the bottom and germinate. The spores are either male or female. The male spores develop into male gametophytes and the females develop into female gametophytes. The male gametophytes produce flagellated sperm which swim through the water and may fertilize eggs produced by the female gametophytes. The fertilized eggs develop into sporophytes, completing the life cycle. The entire process from zoospore release to sporophyte production can be completed on a microscope slide in the laboratory in approximately 12-16 days.
Toxicity tests
The two bioassay protocols developed for Macrocystis at MPSL are similar to those developed for Laminaria saccharina by Chung & Brinkuis (1986). There are three endpoints in the assays: (1) germination of the spores, (2) growth of the spores, and (3) sporophyte production through fertilization of female gametophytes (Fig. 1). The germination and growth endpoints are used in a short-term, 48-h bioassay that focuses on the initial stage of spore development and is intended for use in routine monitoring of effluents. The sporophyte production endpoint is used in a long-term, 16-day bioassay. The long-term kelp test was designed to calibrate the sensitivity, and demonstrate the ecological significance of the short-term test endpoints.
embryonic Long-Term Bioassey sporophyte ~, f °sporoph~e production 1
~ ~ 16 dsys I
sperm egg
~ ~ - - 48 h ------* ~ ~ Short-Term Bio~s~y ~' o* 9 "germin~ion
~ with sorus (~ + C~' g~,n'~etophytes "germ-tube length sporophy~e (2N} spores (1N} (newly germinated)
Fig. 1. Schematic diagram of Macrocystis life-cycle and stages at which short-term and long-term bioassays are performed.

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 117
Short-term test methods
Macrocystis reproductive blades (sporophylls) were collected by divers from adult plants located off Granite Canyon or off Monastery Beach, Monterey County, California. The blades were brought back to the laboratory and rinsed with 0.2/~m-filtered, UV-irradiated seawater to remove epiphytic organisms. To induce zoospore release, we blotted the sporophylls with paper towels, let them sit in the air for 1 h, then placed them in 15°C, filtered seawater. Zoospores were released usually within an hour. Swimming zoospores were used for all tests. The zoospore densities were measured with a hemocytometer. Approximately 450000 spores were added to each test container. For every test, we calculated the volume of spore solution necessary to give the required density of spores, then distributed this volume to each of the test containers by pipet. To avoid test-water dilution, we tried to obtain high densities of zoospores by using several ripe sporophylls in 1 liter of filtered seawater. No more than 10 ml of zoospore solution was added to each container. A glass microscope slide was placed on the bot tom of each test container, and this served as a substratum on which the spores settled and developed. At the end of the test the slide was removed and observed under 400 x on a light microscope.
For all tests we used six test concentrations (5 toxicant dilutions and 1 control) each replicated five times. The test concentrations were assigned in a logarithmic sequence: 0, 10, 18, 32, 56, 100pg/liter, within one order of magnitude depending on the toxicity of the toxicant. The dilution water for the short-term tests was filtered 1-#m, UV-irradiated seawater. We used 250ml of test solution for all tests. Water quality (dissolved oxygen, pH, temperature, and salinity) was monitored daily. Although we did not monitor the ammonia concentrations in the test containers during the effluent experiments, their yearly ranges are presented in the results section. The lights used in the short-term tests were cool white flourescent lights adjusted to give lO0~E/m2/s at the test solution surface. This has been shown to be optimal light conditions for sporophyte production (Luning & Neushul 1978; Deysher & Dean 1984). All tests received continuous light. The test duration was 48 h; the solutions were not renewed.
The test containers were either 350-ml-capacity, polyethylene-plastic, food containers (for metal bioassays) or 600-ml-capacity, borosilicate-glass beakers (for complex effluent and organic toxicant bioassays). Plastic test containers were cleaned by rinsing with hot tap water followed by 24-h sequential soaks in solutions of mild detergent, 3N HC1, and deionized water. Glass test containers were cleaned by triple rinsing with hot tap water and reagent-grade acetone, followed by 24-h soaks in 3N HCI, and deionized water.

118 Brian S. Anderson, John W. Hunt
The endpoints measured after 48 h were germination and germ-tube length (Fig. 2). Germination was considered to be unsuccessful if no germ tube was visible. To differentiate between germinated and non-germinated spores we determined whether they were circular (non-germinated) or had a protuberance of at least 1 spore radius (at least 1-5 #m = germinated). The first 100 spores encountered while moving across the microscope slide were counted for each replicate of each treatment. The growth endpoint was the total length of the spore and germination tube. For this endpoint only germinated spores were measured. The spores to be measured were randomly selected by moving the microscope slide to a new field without looking through the ocular lens, spinning the ocular lens, then measuring the length of a spore touching the micrometer. If more than one spore was touching the micrometer, both (or all) were measured. A total of 10 spores for each replicate of each concentration were measured.
Fig. 2. Germinated (upper two) and non-germinated (lower) spores of Macrocystis afler 48 h (original magnification x 400).
Long-term test methods
The test conditions were the same for the short-term and long-term tests except for the following modifications. The dilution water used was 0.2/~m- filtered, autoclaved seawater. The long-term tests were 16-day, static- renewal bioassays with media renewals at 4-day intervals. For supplemental nutrients we added full-strength PES (Prova~oli 1968) enriched growth

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 119
Fig. 3. Sporophytes of Macrocystis in 16-day old cultures. The sporophytes are the multicellular structures (upper right and lower left). The other structures are non-
reproductive male and female gametophytes.
solution to the test media, minus EDTA-chelated iron; 20 ml PES/liter test solution. We eliminated E D T A from the nutrient media because we were concerned about the possibility of chelation of divalent cations in the test containers (Smith & Harrison, 1978). One hundred thousand spores were added to each test container at the start of the test. The lights used in the long,term tests were Duro-lite ® Vitalights, a full-spectrum light suitable for inducing gametogenesis. These were adjusted to give 100#E/m2/s.
The endpoint for the long-term test was sporophyte production (Fig. 3). After 16 days the microscope slide was removed from the test container and observed under 100 x magnification on a light microscope. We counted the number of sporophytes visible in three 18mm vertical swaths on each replicate, then took a mean of the 3 swaths for the replicate mean. Only sporophytes with at least 3 or more cells were counted.
Toxicants
We tested the effects of three different toxicants on the early life stages of Macrocystis. We tested one metal salt, one organic toxicant, and one sewage- treatment-plant effluent, f rom a mostly domestic primary-treatment facility. The metal salt tested was zinc sulfate. We conducted 3 short-term zinc tests (at 0-10 000 #g/liter zinc) to assess between-test variability, and 1 long-term zinc test (0-3200~g/liter zinc) to calibrate the short-term tests. Zinc

120 Brian S. Anderson, John W. Hunt
concentrations were verified by analysis of water samples using a Perkin Elmer model 603 atomic absorption spectrometer at the California State Mussel Watch analytical facility. The zinc solutions were sampled from 1 replicate of each treatment concentration at the start and end of each experiment. Unless otherwise noted, all reported zinc values are analytically verified concentrations of total zinc. To assess the toxicity of an organic toxicant to Macrocystis we conducted 1 short-term (48-h) and 1 long-term (15-day) test with the water-soluble sodium salt of pentachlorophenate. A stock solution of 100 ppm was prepared by dissolving 0.1429 g of reagent- grade NaPCP in 1 liter of 0.2 #m-filtered seawater. This stock solution was then diluted to give 0-320pg/liter test concentrations; these test con- centrations were not analytically verified and the results are presented as nominal concentrations. In addition, we did one short-term test with a primary (0-56% effluent). The primary effluent was from a single grab sample. This effluent was not chlorinated. Primary-effluent concentrations greater than 1% had salinities adjusted to 34 ppt using a hypersaline brine. The brine was prepared by aerating and heating (MPSL) seawater to no more than 40°C (as recommended by the US EPA). No brine control was used in the primary effluent test.
Statistics
To compare each concentration to the control all of the data were analyzed using analysis of variance (ANOVA) followed by Dunnett 's multiple comparison test (Zar, 1974). The percentage data (germination and sporophyte production) were first transformed to the arcsine of their square root. The length data were not transformed. For each test, the No Observed Effect Concentration (NOEC) was calculated as the highest concentration not significantly different from the control at p = 0"05.
RESULTS
Kelp short-term zinc tests
The results of the multiple short-term kelp tests show that zinc sulfate has less of an inhibitory effect on germination of Macrocystis spores than it does on the elongation of spore germ tubes (Fig. 4a & b, Table 1). Germination of Macrocyst is spores was significantly inhibited at concentrations above 2030/~g/liter zinc in test No. 1, 5500#g/liter in test No. 2, and 1730pg/liter zinc in test No. 3 (Fig. 4a). These values represent the

Bioassay methods jbr evaluating toxicity using Macrocystis pyrifera 121

122 Brian S. Anderson, John W. Hunt
[,.
t,,,
+i
g,
¢-,
O
~ ~
~=~
0
E
r ~
+1 e . -
2
~, rq eq
~aD ¢'q ¢ q ~ ¢:q ¢q
r-4 ¢q ~
.~" .'q .'4"
m ~
g =
6 6 ~ 6 6 6 ~ z z z
~ e h
to% ~ ¢-,I
,g m r,- m r:- ¢q
6 6 6
! g I -
r : - e q ¢;
6 a b
e'-"
G
3
~5
O
Z

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 123
No Observed Effect Concentrations (NOECs) for tests 3-5, respectively (mean = 3090 + 2090#g/liter sd for 3 tests). The reason for the relatively high variability in germination NOECs between test No. 2 and the other two tests is unknown, but could be related to several factors (see discussion). Tests Nos 1 and 3 had the same nominal NOEC. Although the 560 #g/liter treatment (nominal concentration) was significantly different from the control treatment in test No. 3 (Fig. 4), higher concentrations were not. The NOEC was therefore set at 1730/~g/liter in this test ( = 1800/~g/liter nominal concentration) because all concentrations greater than this were signifi- cantly different from the control.
Elongation of the Macrocystis spore germination tubes was significantly inhibited at the lowest zinc concentrations for all three tests (Fig. 4b). The germination tube length NOECs were < 1090, < 590, and < 550/tg/liter for tests Nos 1-3, respectively (mean = < 570 + 30/~g/liter; note that this is the mean of tests 2 and 3. Test No. 1 did not include a 560/~g/liter nominal concentration).
Long-term zinc test
The production of Macrocystis sporophytes in the long-term test was significantly inhibited at concentrations above the NOEC of 1070~tg/liter zinc (Fig. 4c). The long-term test NOEC was lower than the short-term germination NOEC (1070/~g/liter for sporophyte production vs 3090_+ 2090#g/liter for germination) but higher than short-term germ tube length NOEC (1070 vs < 570 _+ 26 /~g/liter). Although the lowest zinc treatment (320/~g/liter= nominal) was significantly different from the control treatment in this test, higher concentrations were not. The NOEC was therefore set at 1070/tg/liter because, as discussed for the third short- term germination test, all of the treatments greater than this were significantly different from the control.
Kelp NaPCP tests
The organic toxicant sodium pentachlorophenate (NaPCP) significantly inhibited germination of Macrocystis zoospores in the short-term test, and sporophyte production in the long-term test at concentrations much lower than those of zinc. The NOECs for both tests were less than 32/~g/liter NaPCP, the lowest concentration tested. The NOECs for both NaPCP tests (Fig. 5a & b) are from nominal, not analytically verified, concentrations.
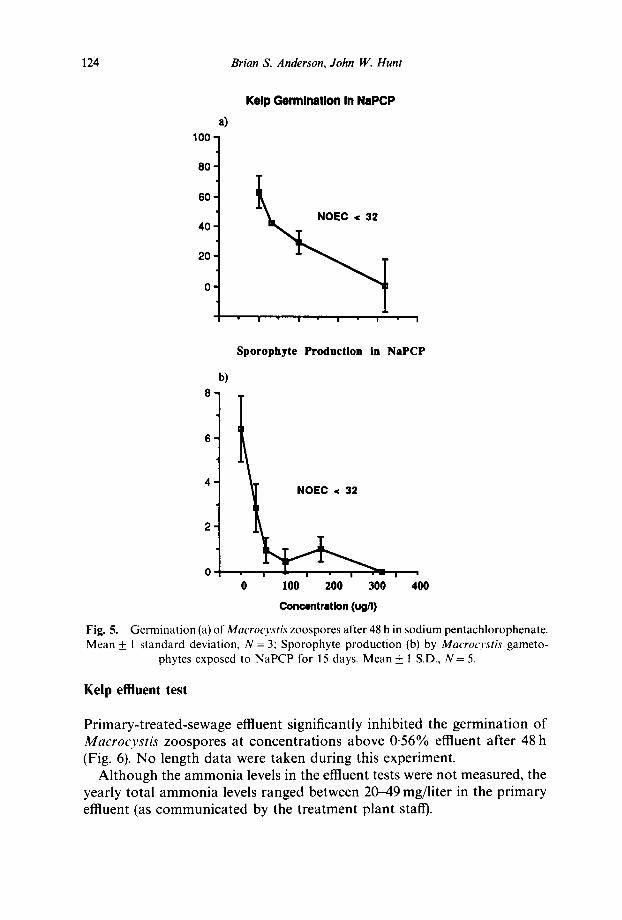
124 Brian S. Anderson, John IV. Hunt
a) 100 -
8o~ 8o~ 4o~ 20
Kelp Germination In NaPCP
| I I I I
Sporophyte Production in NaPCP
b) 8 -
6,
4, NOEC < 32
0' 0 100 200 300 400
Coneamtratlon (ug/I)
Fig. 5. Germination (a) of Macrocystis zoospores after 48 h in sodium pentachlorophenate. Mean _+ 1 standard deviation, N = 3; Sporophyte production (b) by Macrocystis gameto-
phytes exposed to NaPCP for 15 days. Mean _+ 1 S.D., N = 5.
Kelp effluent test
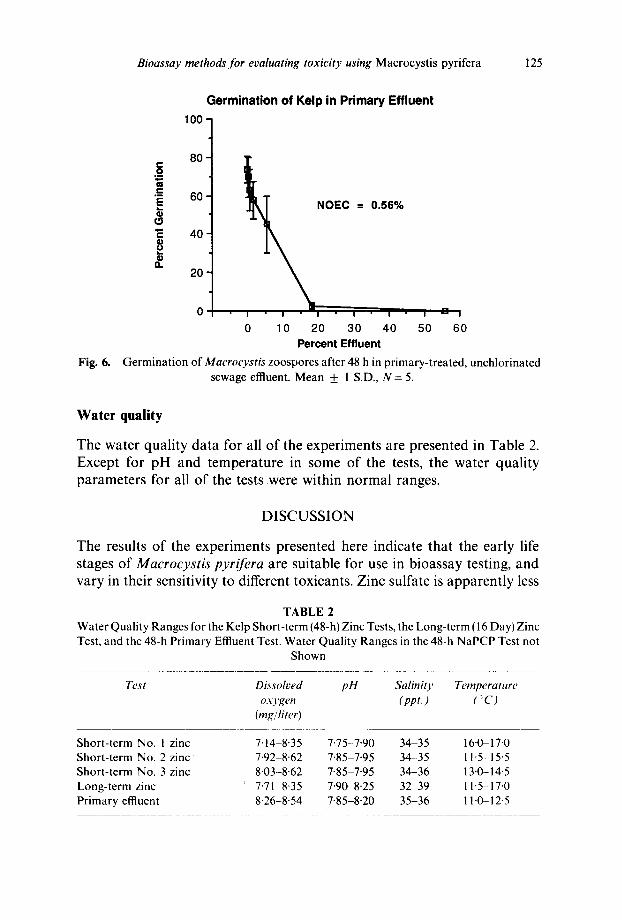
Primary-treated-sewage effluent significantly inhibited the germination of Macrocys t i s zoospores at concentrations above 0-56% effluent after 48 h (Fig. 6). No length data were taken during this experiment.
Although the ammonia levels in the effluent tests were not measured, the yearly total ammonia levels ranged between 20-49 mg/liter in the primary effluent (as communicated by the treatment plant staff).
Bioassay methods for evaluating toxicity using Macrocystis pyrifera 125
Germinat ion of Kelp in Primary Effluent
100 -
NOEC
80' C O
• = - 60'
40'
f t . 2 0
Fig. 6.
= 0.56%
0 I I " I l l I I I I"I I
0 10 20 30 40 50 60 Percent Effluent
Germination of Macrocystis zoospores after 48 h in primary-treated, unchlorinated sewage effluent. Mean ___ 1 S.D., N = 5.
Water quality
The water quality data for all of the experiments are presented in Table 2. Except for pH and temperature in some of the tests, the water quality parameters for all of the tests were within normal ranges.
DISCUSSION
The results of the experiments presented here indicate that the early life stages of Macrocystis pyrifera are suitable for use in bioassay testing, and vary in their sensitivity to different toxicants. Zinc sulfate is apparently less
TABLE 2 Water Quality Ranges for the Kelp Short-term (48-h) Zinc Tests, the Long-term (16 Day) Zinc Test, and the 48-h Primary Effluent Test. Water Quality Ranges in the 48-h NaPCP Test not
Shown
Test Dissolved pH Salinity Temperature oxygen ( ppt. ) (:: C)
(rag/liter)
Short-term No. 1 zinc 7'14-8'35 7.75-7-90 34-35 16-0-17.0 Short-term No. 2 zinc 7.92-8.62 7"85-7-95 34-35 11"5-15"5 Short-term No. 3 zinc 8'03-8"62 7-85-7"95 34-36 13"0-14"5 Long-term zinc 7.71 8"35 7-90-8.25 32 39 11.5-17.0 Primary effluent 8.26-8.54 7.85-8.20 35-36 11.0-12.5

126 Brian S. Anderson, John W. Hunt
toxic to kelp zoospores and gametophytes than is the organic toxicant sodium pentachlorophenate. Primary-treated sewage effluent was toxic to kelp zoospores at relatively low concentrations after a 48-h exposure.
Zinc toxicity
The results of the multiple short-term (48-h) zinc tests indicate that the two related endpoints, zoospore germination and germ-tube elongation, are significantly inhibited at different concentrations of zinc (Fig. 4a & b). Although between-replicate variability was low for the percent germination data (Table 1), between-test variability for the 3 tests was relatively high. The NOECs for the 3 tests ranged between 1730 and 5500/~g/liter zinc (2030, 5500 and 1730 #g/liter zinc for tests Nos 1-3, respectively). The NOECs for the first and third tests were similar; both had the same nominal NOEC (1800/~g/liter). The NOEC for test No. 2, however, was more than twice this value (nominal NOEC = 5600 #g/liter). The sources of this variability are not known. All 3 tests were done within one month, so seasonal effects are unlikely. It is possible that there is variability in tolerance to zinc among reproductive stages of different adult kelp plants, but this is an unlikely cause of between-test variability because the sporophylls used in each test came from several different adult plants. Another possible explanation is variability in test methods or test conditions. In particular, the timing of spore release and test duration were slightly different for the 3 tests (spore release + 1 h, test duration + 3 h). It is not clear whether these slight changes affected the results of the experiments. Although the water quality data for the 3 tests were all within normal ranges, there was some variability between the tests (Table 2). Temperature in particular was more variable, and this could have affected the results. Also, it is apparently the free-ion form of trace metals (in this case the divalent cation of zinc) that are biologically more available and therefore toxic to marine algae (Sundra & Guilliard, 1976; Anderson & Morel, 1978; Kuwabara & North, 1980). This might explain some of the variability in the germination data because free-ion concentrations of zinc vary by as much as an order of magnitude from total concentrations (Kuwabara & North, 1980). Differences in complexation of zinc ions might have occurred in the three tests because of different organic ligands present in the dilution water, or possible precipitates due to high concentrations of zinc (Parry & Hayward, 1973). Although our analyses showed good consistency between nominal and analytically verified concentrations, the free ion concentrations were not determined.
It is uncertain what physiological effect high concentrations of zinc have on zoospore germination. Zinc is an important micronutrient for kelp. Kuwabara (1980) suggests that the primary biochemical role of zinc is in

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 127
auxin metabolism. Zinc is also a cofactor in numerous enzyme systems (Lehninger, 1979). It is possible that when zinc ions are present in high enough concentrations they outcompete other divalent cations for ion- specific enzyme cofactor sites. This might disrupt the effectiveness of some enzymes by replacing the appropriate enzyme cofactor (or catalyst) with another ion species; in this case zinc. What effect high concentrations of zinc have on the biochemical processes leading to germination is not known. Jaffe et al. (1974) suggest that cellular polarization and the subsequent development of rhizoids in developing (2N) Fucus zygotes is related to a leakage of cations (Ca 2÷) across the cell membrane. This cation movement generates an electrical field which initiates the movement of negatively charged vessicles to one side of the cell. The movement of these vessicles establishes and maintains localized growth and the beginning of rhizoid formation. It is possible that a similar process occurs during the early stages of germination in (IN) Macrocystis spores, and that the process is inhibited by high concentrations of Zn 2 +. It is also possible that the mechanism of germination inhibition is related to an inhibition of photosynthesis (see following discussion of inhibition of growth).
The results of the first two short-term zinc tests indicate that germination of kelp zoospores increased initially with increasing zinc concentrations, before declining at higher concentrations (Fig. 4a). Although this indicates that zinc, an important micronutrient, was limiting in the dilution/control water, this type of initial increase in response to low concentrations of a toxicant followed by a decline at higher concentrations is typical of dose-response curves (Laughlin et al., 1981).
Germ-tube elongation is apparently a more sensitive and less variable endpoint than germination. The NOECs for length in all of the zinc tests were lower than the lowest concentrations tested (Fig. 4b). Even though germ tube length is a more sensitive and less variable endpoint than zoospore germination, length may not be as relevant as zoospore germination from an ecological perspective. Germination is obviously relevant because if the spore does not germinate it will not develop. The relevance of germ-tube elongation is less clear. The elongation of the germination tube precedes the process of cytoplasm extrusion, in which the contents of the spore are transferred to the first gametophytic cell. The length of the germination tubes in the control spores were noticeably longer, but it is not clear if this means they were more 'healthy' than spores with shorter germ tubes. Growth is an integration of a number of physiological processes, and for many organisms it has been considered an important indication of toxic effect and nutrient deficiency (for references with algae see Bryan, 1984). Growth has been used as an endpoint in previous experiments involving Macrocystis. Kuwabara (1981) used gametophyte growth as an

128 Brian S. Anderson, John W. Hunt
indicator of trace metal depletion and toxicity. He found significant decreases in gametophytic growth at 250NMol (16-35#g/liter) zinc chloride. The apparent disparity in sensitivity between his Macrocystis cultures and ours might be related to several factors. Because our lowest test concentration was 560#g/liter (nominal concentration), the germ-tube length endpoint could have been considerably more sensitive. We are currently experimenting with lower zinc concentrations to determine the lowest concentration inhibiting germ-tube growth. In addition, we used Granite Canyon seawater as the dilution water in our experiments and Kuwabara used Aquil (a synthetic seawater medium). Kuwabara (1983) found a decrease in free-ion activity in seawater collected from Monterey Bay (near Granite Canyon). He suggested that this might be due to a number of factors including adsorption onto iron hydroxide, and complexation with dissolved organic compounds or particles coated by surface-active organic films. It is possible there was a decrease in Zn 2 ÷ in our seawater relative to Aquil. Because the toxicity of zinc is most likely due to Zn 2 ÷ (Overnell, 1975), the zinc toxicity in our experiments could have been less than in the Aquil medium because ofcomplexation. Also, Kuwabara (1981) used ZnC12 in his experiments while we used ZnSO4. Differences in Zn 2 ÷ availability might have resulted from differences in coprecipitation between these two metal salts. One final factor that could have led to the apparent disparity in sensitivity was the use of different aged gametophytes in the two studies. Kuwabara measured gametophytic growth after 2 weeks in his experiments, while we measured growth after 48 h. Differences in exposure time or relative sensitivities of different aged gametophytes could have contributed to the disparity in results between the two studies.
There is evidence to suggest that elongation of the spore germination tube is a function of the spores' ability to photosynthesize. For example, spores grown under higher light levels (,-~ 100#E/m2/s) have longer germination tubes than those grown under lower light (~ 50~E/m2/s; Anderson and Hunt unpublished data). Toxicant inhibition of germ-tube growth might be linked to the inhibition of the chloroplast. The chloroplast's thylakoid membrane has been described as a fluid mosaic composed of a lipid cont inuum in which protein molecules are imbedded (DeRobertis & DeRobertis, 1980). The distribution of particles within this membrane is related to the presence of cations, in particular Mg 2 ÷ (Ojakian & Satir, 1974; Staehelin, 1976). The presence of high Zn 2 ÷ concentrations could affect the structure of the thylakoid membrane, or the distribution of particles within it. Smith & Harrison (1978) suggested that high Cu 2 ÷ concentrations might disrupt electron transport, or the Na ÷ pump of the chloroplast, or interfere by competing for metal binding sites in cytochrome and chlorophyll. High Zn 2 + concentrations might have similar effects.

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 129
Long-term zinc test
Sporophyte production in the long-term test was significantly reduced above the NOEC of 1070 #g/liter zinc (Fig. 4c). Reproduction in the long- term test was apparently more sensitive to zinc than zoospore germination, but less sensitive than germ-tube elongation. The mean NOEC for the germination endpoint in the 3 short-term zinc tests was 3090 + 2090 #g/liter. The mean NOEC for the germ tube length endpoint in short-term tests Nos 2 and 3 was <570___ 30/~g/liter. Chung & Brinkhuis (1986) found that sporophyte production by Laminaria saccharina was more sensitive to copper than was meiospore settlement and germination. These authors did not measure gametophyte growth.
The variability in the long-term test was relatively high (Fig. 4c). This resulted from differences in sporophyte density among swaths on individual slides. The number of vertical swaths counted on each replicate slide was probably insufficient to eliminate within-replicate variability. Because of this, the statistics used (ANOVA, Dunnett's) did not resolve a significant reduction in sporophyte numbers between the control and 1070pg/liter concentration. This may have reduced the apparent sensitivity (increasing the NOEC) in the long-term test. If this was the case, the short-term germ tube elongation NOECs may correspond more closely to the long-term NOEC. Conversely, the short-term effect of germ-tube elongation might not lead to a long-term effect in reproduction and sporophyte production. A more thorough long-term test with a greater number of swaths read per replicate is planned. By reducing within-replicate variability in the long-term test, we hope to answer this question.
As in the short-term test, it is not clear what toxic effects high concentrations of zinc have on reproduction. As discussed previously for germination and growth, reproductive inhibition could be related to the disrupt ion of photosynthesis. For example, partial impairment of photosynthesis might allow the gametophytes to continue to grow but prevent the accumulation of adequate photosynthate for the construction of gametes. Toxicants might also interfere with fertilization of the female egg by the male sperm. Maier & Muller (1986) have shown that in another laminarian alga (Laminaria digitata) the sperm is attracted to the egg by a pheromone. It is possible that toxicants somehow interfere with the ability of the sperm to find the egg.
Sodium pentachlorophenate toxicity
Not surprisingly, Macrocystis is more sensitive to sodium pentachlorophe- nate than it is to zinc sulfate. Germination in the short-term test was

130 Brian S. Anderson, John W. Hunt
significantly inhibited at or below 32/~g/liter NaPCP, the lowest test concentration (Fig. 5a). Germ tube length was not measured in this test because this endpoint had not yet been developed at the time of this experiment. Clendenning (1959) also found NaPCP to be more toxic to kelp blades than zinc sulfate; he found a significant reduction in Macrocystis photosynthetic activity after a 48-h exposure to 1 mg/liter NaPCP. Thursby et al. (1985) found that 360/1g/liter pentachlorophenol inhibited growth of female Champia parvula (marine red alga) and that 240 #g/liter inhibited growth of Champia tetrasporophytes. Apparently Macrocystis is more sensitive than Champia to pentachlorophenate. The exact mechanism of NaPCP toxicity to kelp spores is not known but pentachlorophenate is an effective fungicide and herbicide that disrupts oxidative phosphorylation. It has been suggested, however, that contaminants in technical grade NaPCP, in particular dioxin, may be more toxic than NaPCP itself (Murphy, 1980). We used reagent-grade chemicals in these tests.
Sporophyte production was also significantly inhibited at or below 32/~g/liter in our 15-day long-term kelp test (Fig. 5b). Germination and sporophyte production exhibited similar sensitivities to NaPCP, suggesting the short-term test is a good indicator of the chronic toxicity of this compound. Replicate short-term tests with an organic toxicant are now being planned to further examine between-test variability, and the sensitivity of germ tube elongation to an organic toxicant.
Effluent toxicity
The results of the kelp bioassays with complex effluents indicate that the short-term bioassay protocol using Macrocystis zoospores can be successfully used for effluent monitoring. Germination of settled zoospores was significantly inhibited by primary effluent at less than 1% effluent (NOEC = 0"56% effluent, Fig. 6). The lengths of the germination tubes were not measured in this test because this endpoint had not yet been developed at the time of this experiment. The settled zoospores were clearly visible in all but the highest concentration of effluent. At 56% effluent, however, particles in the test solution covered the slides and obstructed the spores. Because the toxicity of this effluent was well below this concentration, particle effects were not a factor in establishing a NOEC.
No brine control was used in this test because it was preliminary, and our interest was primarily in evaluating the effects of effluent particulates on the kelp zoospores. Because no brine was needed to adjust the salinity in the lower concentrations (including the lowest effect concentration, 1% effluent), the lack of a brine control did not effect the test results.

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 131
It is unclear what component(s) of the sewage effluent were toxic to the zoospores. This effluent was composed primarily of domestic waste. The reported concentration of copper in a single sample was, for example, 30 #g/liter, while the zinc concentration in the same sample was 60 #g/liter. A variety of organic toxicants were also present in the effluent (gener- ally <10~g/liter), the most conspicuous being phenolic compounds (<40#g/liter). Total ammonia concentrations ranged between 20 and 49mg/liter (20000-49000#g/liter). Based on this range, the ammonia concentration in the effluent concentration that was inhibiting zoospore germination (1%) could have been between 200 and 500/~g/liter. The concentration of unionized ammonia is not known but was certainly far less. Salinity, temperature, pH, and dissolved oxygen were within normal ranges in this test.
The ecological and economic importance of Macrocystispyrifera makes it a valuable resource to the state of California. The discharge of wastes near Macrocystis forests, the ability to routinely culture Macrocystis in the laboratory, and its sensitivity to effluent toxicity make it an appropriate algal species for bioassay testing of complex effluents. The results of the experiments presented here show that bioassays being developed with early life stages of giant kelp can be used for evaluating effluent toxicity. Although the long-term, 16-day reproductive bioassay may not be practical for the routine testing of complex effluents, it can be used to calibrate the short-term test. Some problems need to be addressed before the short-term test is implemented on a widespread basis. The between-test variability of the germination endpoint can hopefully be resolved by doing multiple short- term tests with a reference toxicant other than zinc sulfate. The ecological relevance of the germ tube elongation endpoint can be determined by calibrating this endpoint with a conclusive long-term reproduction bioassay that focuses on sporophyte production.
ACKNOWLEDGEMENTS
We thank Drs Thomas Dean and Lawrence Deysher for contributing substantially to the development of the kelp tests presented here. The manuscript was greatly improved through comments by Dr Francis Palmer, Dr Thomas Dean, Mr Steven Bay and Mr Howard Baily. This project has been funded wholly or in part by the US Environmental Protection Agency using section 205 (j) grant funds under Cooperative Agreement c-060000-23-0 to the State Water Resources Control Board and by State Board Agreement No. 7-120-250-0.

132 Brian S. Anderson, John IV. Hunt
R E F E R E N C E S
Anderson, D. M. & Morel, F. M. M. (1978). Copper sensitivity of Gonyaulax tamarensis. Limnol. Oceanogr., 23, 283-95.
Anderson, B. S., Hunt, J. W., Martin, M., Turpen, S. L. & Palmer, F. H. (1988). Marine Bioassay Project Third Report, protocol development: reference toxicant and initial complex effluent testing. SWRCB, Division of Water Quality report no. 88-7, 1-151.
Bryan, G. W. (1984). Pollution due to heavy metals and their compounds. In Marine Ecology. Vol. 4. ed. O. Kinne, John Wiley & Sons, Chichester, G.B., pp. 1289402.
Chung, I. K. & Brinkhuis, B. H. (1986). Copper effects in early life stages of kelp, Laminaria saccharina. Mar. Poll. Bull., 17, 213-18.
Clendenning, K. A. (1958). Laboratory Investigations. In The Effects of Waste Discharges on Kelp. Water Pollution Control Board Annual Progress Report, July 1958. IMR reference 58-11, 27-38.
Clendenning, K. A. (1959). Laboratory investigations. In The Effects of Waste Discharges on Kelp. Water Pollution Control Board Quarterly Progress Report Oct. Dec. 1958. IMR reference 59-4, 1-13.
Clendenning, K. A. (1960). In The Effects of Waste Discharge on Kelp. Water Pollution Control Board Annual Progress Report, April 1960. IMR reference 60-4, 45.
Dean, T. A. & Jacobsen, F. R. (1984). Growth of juvenile Macrocystis pyrifera (Laminariales) in relation to environmental factors. Mar. Biol., 83, 301-11.
DeRobertis, E. M. & DeRobertis, E. (1980). Cell and Molecular Biology. Saunders College/Holt, Rinehart and Winston, Philadelphia, 1-673.
Devinny, J. S. & Volse, L. A. (1978). Effects of sediments on the development of Macrocystis pyrifera gametophytes. Mar. Biol. ( Berl.), 48, 343-8.
Deysher, L. E. & Dean, T. A. (1984). Critical irradiance levels and the interactive effects on quantum irradiance and quantum dose on gametogenesis in the giant kelp, Macrocystis pyrifera. J. Phycol., 20, 520-4.
Deysher, L. E. & Dean, T. A. (1986a). In situ recruitment of sporophytes of the giant kelp, Macrocystis pyrifera (L.) C.A. Agardh: effects of physical factors. J. Exp. Mar. Biol. Ecol., 103, 41-63.
Deysher, L. E. & Dean, T. A. (1986b). Interactive effects of light and temperature on sporophyte production in the giant kelp, Macrocystis pyrifera. Mar. BioL, 93, 1720-33.
Eppley, R. W., Carlucci, A. F., Holm-Hansen, O., Kiefer, D., McCarthy, J. J. & Williams, P. M. (1972). Evidence for eutrophication in the sea near southern California sewage outfalls, July 1970. Calif. Coop. Oceanic Fish. Invest. Rep. 16, 74-83.
Foster, M. S. & Schiel, D. R. (1985). The ecology of giant kelp forests in California: A community profile. U.S. Fish Wildl. Serv. Biol. Rep., 85(7.2), 1-152.
Grigg, R. W. (1978). Long term changes in rocky bottom communities of Palos Verdes; In Coastal Water Research Project Annual Report, ed. W. Bascom, 157-84.
Jaffe, L. F., Robinson, F. R. & Nucittelli, R. (1974). Local cation entry and self- electrophoresis as an intracellular localization mechanism. Ann. N.Y. Acad. Sci., 238, 372 89.

Bioassay methods for evaluating toxicity using Macrocystis pyrifera 133
Kuwabara, J. S. (1980). Micronutrient requirements for Macrocystis pyrifera (L.) C. A. Agardh (giant kelp) gametophytes determined by means of a chemically defined medium, Aquil. Doctoral Dissertation, California Institute of Technology, Pasadena, California, 1-185.
Kuwabara, J. S. (1981). Gametophytic growth by Macrocystis pyrifera (Phaeophyta) in response to various iron and zinc concentrations. J. Phycol., 17, 417-19.
Kuwabara, J. S. (1983). Effects of trace metals and natural organics on algae. In The Effects of Waste Disposal on Kelp Communities, ed. W. Bascom, Southern California Coastal Water Research Project, Long Beach, CA, pp. 136~6.
Kuwabara, J. S. & North, W. J. (1980). Culturing microscopic stages of Macrocystis pyrifera (Phaeophyta) in Aquil, a chemically defined medium. J. Phycol., 16, 546-9.
Laughlin, R. B., Jr., Ng, J. & Guard, H. E. (1981). Hormesis: A response to low environmental concentrations of petroleum hydrocarbons. Science, 211,705 7.
Lehninger, A. L. (1979). Biochemistry. Worth Publishers, New York, pp. 1-1104. Luning, K. & Neushul, M. (1978). Light and temperature demands for growth and
reproduction of laminarian gametophytes in southern and central California. Mar. Biol. ( Berl.), 45, 297-309.
Maier, I. & Muller, D. G. (1986). Sexual pheromones in algae. Biol. Bull., 170, 145 75.
Meistrell, J. C. & Montagne, D. E. (1983). In The Effects of Waste Di.~posal on Kelp Communities, ed. W. Bascom, Southern California Coastal Water Research Project, pp. 84-102.
Murphy, S. D. (1980). In Toxicology, ed. J. Doull, C. D. Klaassen & M. E. Amdur. Macmillan, NY, pp. 393-4.
North, W. J. (ed.) (1971). The biology of giant kelp beds (Macrocvstis) in California. Nova Hedwigia, 32, 1-600.
North, W. J. (1976). Aquacultural techniques for creating and restoring beds of giant kelp, Macrocystis spp. J. Fish. Res. Board Can., 33, 1015 23.
Overnell, J. (1975). The effects of heavy metals on photosynthesis and loss of cell potassium in two species of marine algae, Dunaliella tertiolecta and Phaeodactylum tricornutum. Mar. Biol., 29, 99-103.
Ojakian, G. K. & Satir, P. (1974). Particle movements in chloroplast membranes: Quantitative measurements of membrane fluidity by the freeze fracture technique. Proc. Nat. Acad. Sci., 71, 2052-56.
Parry, G. D. R. & Hayward, J. (1973). The uptake of °SZinc by Dunaliella tertiolecta Butcher. J. Mar. Biol. Ass. U.K., 53, 915-922.
Provasoli, L. (1968). Media and prospects for the cultivation of marine algae. In Cultures and Collections of Algae. Proceedings of the U.S.-Japan Conference, Hakone. Japanese Society of Plant Physiology, ed. A. Watanabe and A. Hattori, pp. 63-75.
Schafer, H. (1980). Characteristics of municipal wastewater. In Southern California Coastal Water Research Biennial Report, ed. W. Bascon. SCCWRP, Long Beach, pp. 236-37.
Schafer, H. (1982). Characteristics of municipal wastewater. In SCCWRP Biennial Report, 1981-1982, ed. W. Bascom, SCCWRP, Long Beach, pp. 12-13.
Schafer, H. (1984). Characteristics of municipal wastewater. In SCCWRP Biennial Report, ed. W. Bascom, SCCWRP, Long Beach, 16-17.

134 Brian S. Anderson, John IV. Hunt
Smith, B. M. & Harrison, F. L. (1978). Sensitivity of Macrocystis gametophytes to copper. Final report to US Nuclear Regulatory Commission, NRC FIN No. A0119, 1-34.
Staehelin, L. A. (1976). Reversible particle movements associated with unstacking and restacking of chloroplast membrane in vitro. J. Cell Bio., 71, 136-58.
Sunda, W. G. & Guilliard, R. R. L. (1976). The relationship between cupric ion activity and the toxicity of copper to phytoplankton. J. Mar. Res., 35, 511-29.
Thursby, G. B., Steele, R. C. & Kane, M. E. (1985). Effect of organic chemicals on growth and reproduction in the marine red alga Champia parvula. Env. Tox. and Chem., 4, 797-805.
Wilson, K. & McPeak, R. (1983). Kelp restoration. In The Effects of Waste Disposal on Kelp Communities, ed. W. Bascom, SCCWRP, Long Beach, pp. 199-216.
Wilson, K. C., Mearns, A. J. & Grant, J. J. (1980). Changes in kelp forests at Palos Verdes. In Southern California Coastal Water Research Project Biennial Rep. SCCWRP, ed. W. Bascom, Long Beach, pp. 77-92.
Zar, J. H. (1974). Biostatistical Analysis. Prentice-Hall Inc., Englewood Cliffs, N J, pp. 1-620.