linking forest edge structure to edge function: mediation of herbivore damage
TRANSCRIPT

Linking forest edge structure to edge function:
mediation of herbivore damage
M. L. CADENASSO and S. T. A. PICKETT
Institute of Ecosystem Studies, Box AB, Millbrook, NY 12545, USA
Summary
1 Forest edges, which are prominent features in the north-eastern United States
landscape, may control the ¯ux of organisms between forest and non-forest habi-
tats. Previous studies have described edge structure rather than function, as deter-
mined by interaction with such ¯uxes.
2 The function of the forest edge may be linked to the structure of its vegetation.
We tested this hypothesis by experimentally altering the structure of the vegetation
at two deciduous forest edges in Millbrook, New York, USA. Intact and thinned
plots were established at each edge and we determined whether the structure of the
edges in¯uenced the ¯ux of herbivores, as measured by herbivore damage to trans-
planted tree seedlings.
3 Herbivore damage to seedlings at site 1 was a�ected by edge vegetation structure
and by distance from the edge. The edge structure e�ect was due to herbivory by
voles, which was signi®cantly greater in the intact than in the thinned treatment.
Regardless of treatment, voles damaged seedlings only on the edge and 30±40m
from the edge and did no damage in the forest interior (90±100m), whereas deer
damaged signi®cantly more seedlings in the forest interior than on the edge. At site
2, where vole damage was concentrated on the edge, damage to seedlings was
a�ected only by distance from the edge, not edge structure.
4 The two dominant herbivores, white-tailed deer and meadow voles, preferentially
damaged di�erent seedling species. In addition, tree seedlings browsed by deer
resprouted more frequently than those clipped by voles. Our results suggest that
both edge structure and distance from the edge in¯uence herbivore activity and, as
a result, in¯uence the spatial arrangement, density and composition of populations
of tree seedlings during regeneration in forest fragments.
Keywords: edge permeability, fragmentation, meadow voles, tree seedlings, white-
tailed deer
Journal of Ecology (2000) 88, 31±44
Introduction
In the north-eastern United States, as elsewhere,
fragmentation of landscape structure has increased
both the prominence of forest edges and the propor-
tion of forest area that lies close to an edge. While
processes characteristic of the forest interior, such as
tree regeneration, plant±plant interactions and
plant±animal interactions, still occur in forest
patches, their dynamics may be in¯uenced by factors
originating from outside the patch boundary
(Saunders et al. 1991; Angelstam 1992). The impact
of exogenous factors on internal forest dynamics
may depend on the permeability of the edge to those
factors (Wiens et al. 1985; Stamps et al. 1987;
Forman & Moore 1992), and this study represents
the ®rst experimental test of whether the ¯ux of one
such factor, tree seedling herbivores, is indeed
a�ected by the structure of the forest edge.
The importance of forest edges has been acknowl-
edged and investigated by ecologists for a long time,
although the empirical approach has been predomi-
nantly static and descriptive (Murcia 1995). Leopold
(1936) ®rst suggested that edges bene®t game man-
agement. Later, ecologists recognized that there are
both positive and negative e�ects of edges, depend-
ing on the organism or scale of study (Harris 1988).Correspondence: M. L. Cadenasso (fax 914 6775976;
e-mail [email protected]).
Journal of
Ecology 2000,
88, 31±44
# 2000 British
Ecological Society

In the context of the plant community, the focus of
edge research has been on quantifying abiotic gradi-
ents (Geiger 1965; Wales 1967, 1972; Matlack 1993;
Weathers et al. 1995; Cadenasso et al. 1997) and bio-
tic structure (Ranney et al. 1981; Williams-Linera
1990; Brothers & Spingarn 1992) across edges,
allowing comparison between edges that di�er in
age (Matlack 1994; Camargo & Kapos 1995), aspect
(Wales 1972; Matlack 1994) or adjacent land use
(Moran 1984; Chen et al. 1992). Functional signi®-
cance is often inferred from these largely descriptive
patterns of plant community structure: for example,
the use of edge to interior ratios assumes that zones
within a certain distance from the edge have a parti-
cular function (Levenson 1981; Laurance & Yensen
1991). The link is, however, largely untested except
for the e�ect of edges on nest predation and parasit-
ism in birds (Andre n & Angelstam 1988; Andre n
1995), where results are often contradictory (Paton
1994). This research tests the assumption by examin-
ing how edge structure mediates herbivore damage
to tree seedlings, and thus assesses the functional
impact of the edge on the interior forest community
and plant population dynamics.
The call for more research on the functioning of
forest edges has been justi®ed by their potential to
mediate ¯ows between forest and non-forest habitats
(Wiens et al. 1985; Forman & Moore 1992; Hansson
1992; Wiens 1992), depending on the characteristics
of both the two habitats and the edge zone and on
the nature of the ¯ow (Wiens 1992; Wiens et al.
1993). There are several externally originating pro-
cesses that can ¯ow across the landscape and
through forest edges (Saunders et al. 1991; Forman
& Moore 1992; Wiens 1992). These ¯ows of organ-
isms, material and energy link habitat patches in the
landscape and may a�ect landscape pattern, popula-
tion dynamics and nutrient cycling (Wiens 1992).
The further hypothesis that the dynamics within
each patch will be a�ected by the existence of edges
(Wiens et al. 1993) has not been tested experimen-
tally.
We adopted a conceptual model that integrates
structure and function and illustrates the potential
functions of forest edges (Pickett & Cadenasso
1995). This model incorporates ideas from a variety
of sources (Forman & Moore 1992; Wiens 1992;
Wiens et al. 1993; Forman 1995) and comprises
three distinct zones ± the forest interior, the forest
edge and the surrounding non-forest habitat. Most
organisms, material and energy moving from the
forest exterior into the interior must pass through
the forest edge (exceptions include birds or seeds
that can travel over the edge and deposit directly in
the interior, and ¯uxes in ground water). Depending
on the situation, an edge may inhibit or enhance
movement across the boundary or it may have no
e�ect.
Our experimental test of the link between struc-
ture and function of forest edges considered this
model for the particular case of the ¯ux of tree seed-
ling herbivores between deciduous forests and adja-
cent old ®elds released from agriculture. Edges
between these contrasting land covers are structu-
rally and compositionally abrupt, and biotic and
abiotic changes across these edges are steep.
The meadow vole (Microtus pennsylvanicus, Ord
1815) is the primary herbivore in the old ®eld, and
the white-tailed deer (Odocoileus virginianus,
Zimmermann 1780) is the main forest herbivore.
Herbivory is important in determining tree seedling
composition and density in both forests (Alverson
et al. 1988; Tilghman 1989; Gill 1992) and old ®elds
(Rankin & Pickett 1989; Gill & Marks 1991; Ostfeld
& Canham 1993). For example, proximity to the
forest edge is critical to the establishment of woody
species in adjacent old ®elds because it provides a
seed source (Gill & Marks 1991) and in¯uences her-
bivore activity (Williamson & Hirth 1985; Myster &
McCarthy 1989; Ostfeld et al. 1997). Here, we con-
sidered the e�ect of edge on tree regeneration in the
forest, which has been less well studied.
The structure of the edge is hypothesized to in¯u-
ence how it mediates any ¯ux. Edge structure can be
viewed from a geographical perspective: a forest
edge represents a physical disjunction in the land-
scape at a scale of kilometres. We therefore consid-
ered how herbivore activity di�ered with distance
from the forest edge. Alternatively, the ecological
structure of an edge is represented by the complex
of vegetation layers that make up the edge zone and
varies over a scale of metres. We tested the function
of the edge at this scale by experimentally altering
the structure of the vegetation on the edge and mea-
suring how herbivore activity di�ered with vegeta-
tion structure.
Two questions were posed. First, how does the
structure of the vegetation on the forest edge med-
iate herbivory in the forest? To determine the direct
e�ects of edge structure on a key herbivore, small
mammals were trapped. Secondly, what is the net
e�ect of that mediation on tree regeneration in the
forest?
Site description
Two 1-ha sites were located at the Institute of
Ecosystem Studies (IES) in Millbrook, Dutchess
County, New York, USA (41�500N, 73�45
0W).
Both were embedded in large (>40ha) upland
mesic deciduous forests that are approximately 60
years old, and had similar canopy composition
(dominated by several species including Quercus
rubra, Q. alba, Q. prinus, Acer rubrum, A. sac-
charum, Fagus grandifolia, Betula lenta and Carya
spp., with Ostrya virginiana and Carpinus caroliniana
32Linking forest
edge structure to
edge function
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44

as mid-canopy trees and Hammemalis virginiana and
Viburnum acerifolium in the understorey). The ®elds
adjacent to each site had been released from hay
production or grazing in the 1960s. The ®eld at site
1 had been mowed periodically, there was no woody
establishment and it was dominated by several spe-
cies of Solidago, Schizachyrium scoparium, Poa spp.
and other grass and forb species (Armesto et al.
1991). The forest at site 2 was separated from the
®eld by a 3-m wide paved road, accessible to only
the IES. This ®eld had not been mowed except
along the road margins and, in addition to the her-
baceous species found at site 1, had small patches of
Rhus glabra and Cornus racemosa and two indivi-
duals of Acer rubrum approximately 5m tall. Both
sites had relatively straight and structurally abrupt
edges. Site 1 faced north-east (21� from true north)
and site 2 had a west aspect (275� from true north)
(Fig. 1). (Taxonomy is according to Gleason &
Cronquist 1991.)
EXPERIMENTAL MANIPULATION OF EDGE
STRUCTURE
Plots were established at each stand such that the
external boundary approximately followed the
canopy dripline and the sites extended 100m into
the forest interior (Fig. 1). Each of the sites was
divided into two 40-m plots separated by a 20-m
bu�er zone, and intact and thinned treatments were
allocated at random (Fig. 2). In autumn 1994 the
woody vegetation lower than half canopy height
was removed from the outermost 20m of the
thinned plot at each site. This included the lateral
branches of the canopy trees that were lower than
half canopy height, complete trees where they were
shorter than half the canopy height, and shrubs; the
herbaceous layer was not cut. Although we altered
the physical structure of the edges of the thinned
plots, but no canopy trees were removed, the shape
and location of these abrupt edges, as drawn on a
map, were not therefore altered. In other words, we
altered the ecological but not the geographical struc-
ture of the edges. Our manipulation produced a
structure similar to a newly created forest edge
before colonization by shrubs and saplings or devel-
opment of lateral branches by the newly exposed
canopy trees.
Methods
HERBIVORY EXPERIMENTAL DESIGN
Five pairs of caged and uncaged quadrats were
established in a strati®ed random design in each of
three distance bands from the edge of each treat-
ment plot (0±10m, 30±40m and 90±100m) (60
quadrats per site) (Fig. 2). Caging was accomplished
by constructing a four-sided exclosure measuring
1.5� 0.75m and made of 61-cm wide square-weave
`hardware cloth' with 1.27-cm mesh. The perimeter
was buried 4±6 cm to discourage animals from bur-
rowing under, and the cages were regularly checked
for signs of disturbance. During burial, damage to
litter and vegetation was avoided as much as possi-
ble. Twenty-centimetre wide aluminium ¯ashing was
riveted to the top of the hardware cloth around the
Fig. 1 The two ®eld sites used in the experiment. Each site occupied 100m of forest edge and extended 100m into the forest
interior and contained a thinned and a control (intact) plot allocated at random.
33M.L. Cadenasso
& S.T.A. Pickett
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44

entire cage perimeter, to prevent small mammals
from using the mesh to climb up the sides of the
cage and enter the cage through the top. Hexagonal-
weave `chicken wire' (2.54-cm mesh) was secured
over the top of the cage to prevent deer browsing
inside the exclosure. An uncaged quadrat of the
same size was located approximately 3m away from
the caged quadrat with which it was paired.
In the spring of 1995, all (caged and uncaged)
quadrats were planted with 12 seedlings of each of
Betula lenta, Quercus rubra and Ailanthus altissima.
All three species are found in these forests and
represent an early and a late successional species
and a species invasive in the Hudson Valley, respec-
tively. The seedlings were grown from seed either
collected on the IES grounds or bought from F.W.
Schumacher Co. (Sandwich, MA). Seeds were ger-
minated in February and March and seedlings were
maintained in the IES greenhouse until planted in
the ®eld in early June. Immediately after planting
seedlings were watered generously. The three species
were located randomly in the quadrats.
Seedlings were monitored weekly for the ®rst
10weeks then every 2 weeks until leaf drop in the
autumn. They were resurveyed in the spring of 1996.
Seedlings not surviving the ®rst 2 weeks after plant-
ing due to shock or stress were replaced only once.
Each seedling was scored as undamaged, damaged
by herbivores, mechanically damaged or dead from
other causes. Each herbivore damages seedlings in a
unique way, allowing for easy ®eld scoring of
damage by a speci®c herbivore. Voles clip seedlings
at the base, approximately 2 cm from the soil,
cleanly severing the stem. Deer remove biomass
from the top of the seedling and, occasionally,
remove only the apical bud. Due to their lack of
upper incisors, deer leave a very jagged cut on the
remaining seedling stem. Slugs were observed on the
plants and their slime trails were apparent on the
seedling stem. Insect defoliation was incremental
and the defoliation could be tracked until seedling
mortality. Slug and insect damage were combined
into an invertebrate class for the analyses. All seed-
ling damage observed could be attributed to one of
these classes of herbivores.
SMALL MAMMAL TRAPPING
Small mammals were trapped using Sherman live
traps placed in a regular grid extending from 20m
beyond the edge to 50m into the forest. Eight traps
were placed 10m apart along three transects in each
intact and thinned plot. Each site was trapped every
third week for three consecutive nights. Because
traps were moved between the two sites, traps were
placed on the trap station, left closed, and baited at
least 3 days prior to the actual trapping period, to
ensure that animals were familiar with the traps.
This strategy was used to prevent inaccuracies in the
capture data that may have occurred if a lag time
existed for the small mammals to `®nd' the traps.
Each site was trapped on seven occasions between
13 June and 26 October 1995, the period during
which herbivory was measured. A site was not
trapped in the week immediately following seedling
planting so that the herbivores would have access to
Fig. 2 Experimental design. Sites were embedded within deciduous forests (light shading) and a 20-m bu�er zone (hatched)
separated the two experimental plots. At three distances (0±10, 30±40 and 90±100m) on each plot, ®ve pairs of caged (&)
and open (&) quadrats were established randomly.
34Linking forest
edge structure to
edge function
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44

the seedlings used in the experiment. Traps were set
in the late afternoon, baited with rolled oats, and
checked the following morning within 2 h of sunrise.
STATISTICAL ANALYSES
All statistical analyses were done separately for data
from both site 1 and site 2. Because parametric and
non-parametric statistical approaches yielded the
same results, we only present the parametric ana-
lyses.
We used a three-factor nested multivariate analy-
sis of variance (MANOVA) together with univariate
three-factor nested ANOVAs to analyse the e�ects of
edge vegetation structure, distance from the edge
and herbivore exclusion cages on the response vari-
able, and damage to seedlings due to herbivory vs.
damage due to all other causes. Distance from the
edge was nested within edge structure and within
cage e�ect. Damage was calculated as the percen-
tage of the 36 individual seedlings planted in each
caged or uncaged quadrat that was damaged either
by a herbivore or by other means (i.e. mechanical
damage and other causes combined). ANOVA results
for a particular factor (edge structure, cage, distance
nested within edge structure or distance nested
within cage) were not considered unless MANOVA
results for that factor were signi®cant.
The e�ects of edge vegetation structure and dis-
tance from the edge nested within edge structure on
damage to seedlings caused by each of the three her-
bivore classes (deer, voles, invertebrates) were ana-
lysed with a two-factor nested MANOVA. If the
MANOVA results for a factor were signi®cant, then
two-factor nested ANOVAs were used to evaluate
which variable (herbivore class) contributed to the
signi®cant di�erences detected by the multivariate
tests. Damage was calculated for each open quadrat
as a percentage of the 36 planted seedlings that had
been subjected to herbivory by each of the three
classes. Invertebrates did not damage seedlings at
site 2 and were excluded from the analyses for this
site.
We also tested for patterns in damage among
seedling species. A two-factor nested MANOVA was
used to analyse the e�ects of edge vegetation struc-
ture and distance from edge nested within edge
structure on damage to tree seedlings caused by her-
bivory for each of the three tree species used.
Damage was calculated as a percentage of the 12
individuals of each species in each open quadrat
that had been a�ected. Again, two-factor nested
ANOVA results were considered for each factor sig-
ni®cant in the MANOVA tests to evaluate which of
the response variables (seedling species) contributed
to the signi®cant di�erences detected by the multi-
variate tests.
To test the net e�ect of herbivore damage on (i)
the mortality and (ii) the composition of the seedling
community, we analysed (i) overall seedling response
to damage caused by each of the herbivores and (ii)
damage caused by each of the herbivores to each
seedling species to determine if a herbivore damaged
seedling species di�erentially. A chi-square fre-
quency analysis was used to test the e�ect of herbiv-
ory by deer and voles on the resprouting frequency
of seedlings following damage. The assumption of
this analysis was that the ability of the damaged
seedling to resprout would be determined by the
identity of the herbivore because of the characteris-
tic manner in which each herbivore damages seed-
lings. Edge structure and distance from the edge
may in¯uence which herbivore encounters the quad-
rat, but were not expected to in¯uence the frequency
of resprout by seedlings following damage.
Therefore, the location of the seedling was not con-
sidered in this analysis. Damage was calculated as
the percentage of seedlings damaged by each herbi-
vore that resprouted in each open quadrat. For
example, if deer and voles damaged seedlings in a
quadrat and all those damaged by deer resprouted
and none damaged by voles resprouted, the response
value for deer was 100% and for voles 0%. Because
the analysis was done to test the net e�ect of herbiv-
ory on seedling community mortality, quadrats with
no herbivore damage were not used in the analysis.
The chi-square test determined whether the fre-
quency of resprout of seedlings damaged by deer
and damaged by voles di�ered from an expected
equal frequency.
A similar test with a separate analysis for each
herbivore class was used to determine whether each
species of seedlings was damaged equally by each
herbivore. Location of the seedling with respect to
edge structure and distance from the edge was again
assumed not to in¯uence the preference of the herbi-
vore. Twelve individuals of each seedling species
were exposed to herbivory in each open quadrat.
The percentage of each species in each quadrat
damaged by a particular herbivore was calculated.
Because the analysis was to test the net e�ect of her-
bivory on the composition of the seedling commu-
nity, quadrats in which the particular herbivore
caused no damage were excluded from the analysis.
A chi-square test was performed for each herbivore
to test whether the herbivore damaged seedling spe-
cies di�erentially. Because invertebrates only
damaged seedlings at site 1, the analysis of inverte-
brate preference was performed only for this site.
Results
HERBIVORE EXCLUSION CAGES AND
DAMAGE TO SEEDLINGS
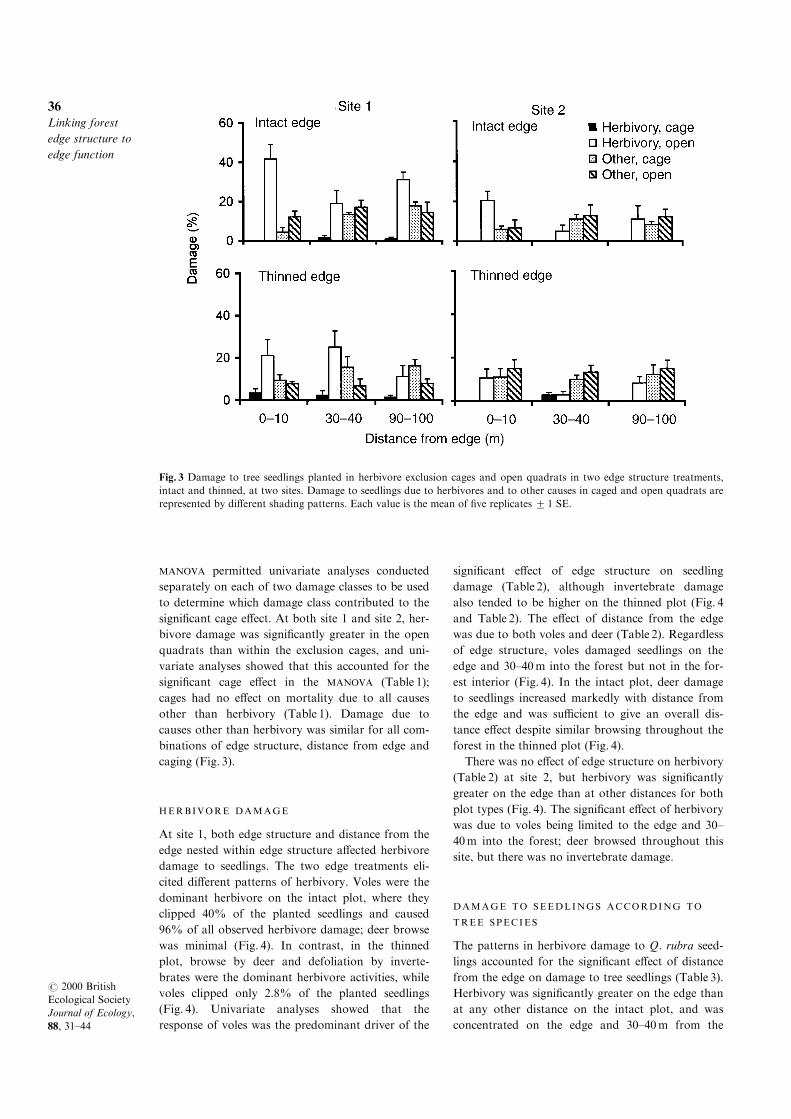
Herbivore exclusion cages a�ected damage to tree
seedlings at both site 1 and site 2 (Fig. 3 and
Table 1). The signi®cant e�ect of cages in the
35M.L. Cadenasso
& S.T.A. Pickett
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44
MANOVA permitted univariate analyses conducted
separately on each of two damage classes to be used
to determine which damage class contributed to the
signi®cant cage e�ect. At both site 1 and site 2, her-
bivore damage was signi®cantly greater in the open
quadrats than within the exclusion cages, and uni-
variate analyses showed that this accounted for the
signi®cant cage e�ect in the MANOVA (Table 1);
cages had no e�ect on mortality due to all causes
other than herbivory (Table 1). Damage due to
causes other than herbivory was similar for all com-
binations of edge structure, distance from edge and
caging (Fig. 3).
HERBIVORE DAMAGE
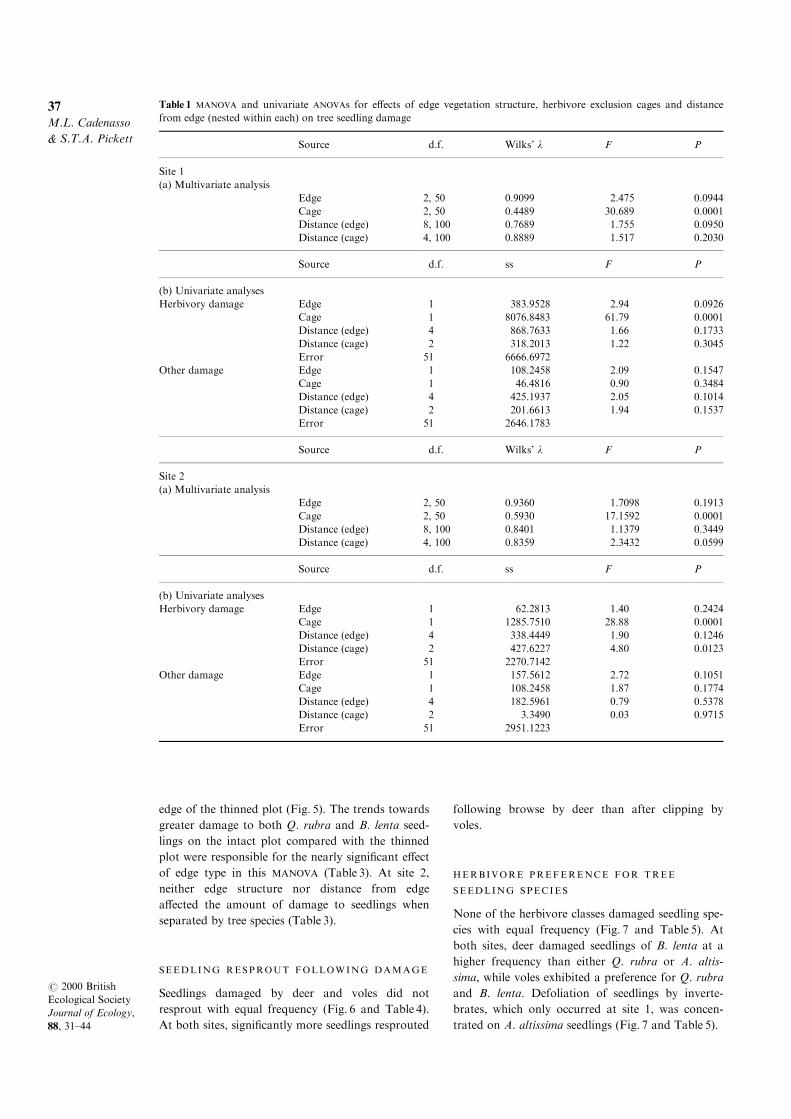
At site 1, both edge structure and distance from the
edge nested within edge structure a�ected herbivore
damage to seedlings. The two edge treatments eli-
cited di�erent patterns of herbivory. Voles were the
dominant herbivore on the intact plot, where they
clipped 40% of the planted seedlings and caused
96% of all observed herbivore damage; deer browse
was minimal (Fig. 4). In contrast, in the thinned
plot, browse by deer and defoliation by inverte-
brates were the dominant herbivore activities, while
voles clipped only 2.8% of the planted seedlings
(Fig. 4). Univariate analyses showed that the
response of voles was the predominant driver of the
signi®cant e�ect of edge structure on seedling
damage (Table 2), although invertebrate damage
also tended to be higher on the thinned plot (Fig. 4
and Table 2). The e�ect of distance from the edge
was due to both voles and deer (Table 2). Regardless
of edge structure, voles damaged seedlings on the
edge and 30±40m into the forest but not in the for-
est interior (Fig. 4). In the intact plot, deer damage
to seedlings increased markedly with distance from
the edge and was su�cient to give an overall dis-
tance e�ect despite similar browsing throughout the
forest in the thinned plot (Fig. 4).
There was no e�ect of edge structure on herbivory
(Table 2) at site 2, but herbivory was signi®cantly
greater on the edge than at other distances for both
plot types (Fig. 4). The signi®cant e�ect of herbivory
was due to voles being limited to the edge and 30±
40m into the forest; deer browsed throughout this
site, but there was no invertebrate damage.
DAMAGE TO SEEDLINGS ACCORDING TO
TREE SPECIES
The patterns in herbivore damage to Q. rubra seed-
lings accounted for the signi®cant e�ect of distance
from the edge on damage to tree seedlings (Table 3).
Herbivory was signi®cantly greater on the edge than
at any other distance on the intact plot, and was
concentrated on the edge and 30±40m from the
Fig. 3 Damage to tree seedlings planted in herbivore exclusion cages and open quadrats in two edge structure treatments,
intact and thinned, at two sites. Damage to seedlings due to herbivores and to other causes in caged and open quadrats are
represented by di�erent shading patterns. Each value is the mean of ®ve replicates 2 1 SE.
36Linking forest
edge structure to
edge function
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44
edge of the thinned plot (Fig. 5). The trends towards
greater damage to both Q. rubra and B. lenta seed-
lings on the intact plot compared with the thinned
plot were responsible for the nearly signi®cant e�ect
of edge type in this MANOVA (Table 3). At site 2,
neither edge structure nor distance from edge
a�ected the amount of damage to seedlings when
separated by tree species (Table 3).
SEEDLING RESPROUT FOLLOWING DAMAGE
Seedlings damaged by deer and voles did not
resprout with equal frequency (Fig. 6 and Table 4).
At both sites, signi®cantly more seedlings resprouted
following browse by deer than after clipping by
voles.
HERBIVORE PREFERENCE FOR TREE
SEEDLING SPECIES
None of the herbivore classes damaged seedling spe-
cies with equal frequency (Fig. 7 and Table 5). At
both sites, deer damaged seedlings of B. lenta at a
higher frequency than either Q. rubra or A. altis-
sima, while voles exhibited a preference for Q. rubra
and B. lenta. Defoliation of seedlings by inverte-
brates, which only occurred at site 1, was concen-
trated on A. altissima seedlings (Fig. 7 and Table 5).
Table 1 MANOVA and univariate ANOVAs for e�ects of edge vegetation structure, herbivore exclusion cages and distance
from edge (nested within each) on tree seedling damage
Source d.f. Wilks' l F P
Site 1
(a) Multivariate analysis
Edge 2, 50 0.9099 2.475 0.0944
Cage 2, 50 0.4489 30.689 0.0001
Distance (edge) 8, 100 0.7689 1.755 0.0950
Distance (cage) 4, 100 0.8889 1.517 0.2030
Source d.f. ss F P
(b) Univariate analyses
Herbivory damage Edge 1 383.9528 2.94 0.0926
Cage 1 8076.8483 61.79 0.0001
Distance (edge) 4 868.7633 1.66 0.1733
Distance (cage) 2 318.2013 1.22 0.3045
Error 51 6666.6972
Other damage Edge 1 108.2458 2.09 0.1547
Cage 1 46.4816 0.90 0.3484
Distance (edge) 4 425.1937 2.05 0.1014
Distance (cage) 2 201.6613 1.94 0.1537
Error 51 2646.1783
Source d.f. Wilks' l F P
Site 2
(a) Multivariate analysis
Edge 2, 50 0.9360 1.7098 0.1913
Cage 2, 50 0.5930 17.1592 0.0001
Distance (edge) 8, 100 0.8401 1.1379 0.3449
Distance (cage) 4, 100 0.8359 2.3432 0.0599
Source d.f. ss F P
(b) Univariate analyses
Herbivory damage Edge 1 62.2813 1.40 0.2424
Cage 1 1285.7510 28.88 0.0001
Distance (edge) 4 338.4449 1.90 0.1246
Distance (cage) 2 427.6227 4.80 0.0123
Error 51 2270.7142
Other damage Edge 1 157.5612 2.72 0.1051
Cage 1 108.2458 1.87 0.1774
Distance (edge) 4 182.5961 0.79 0.5378
Distance (cage) 2 3.3490 0.03 0.9715
Error 51 2951.1223
37M.L. Cadenasso
& S.T.A. Pickett
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44
Fig. 4 Herbivore damage to tree seedlings at three distances from intact and thinned treatment edges at two sites separated
by herbivore class. Each value is the mean of ®ve replicates 2 1 SE.
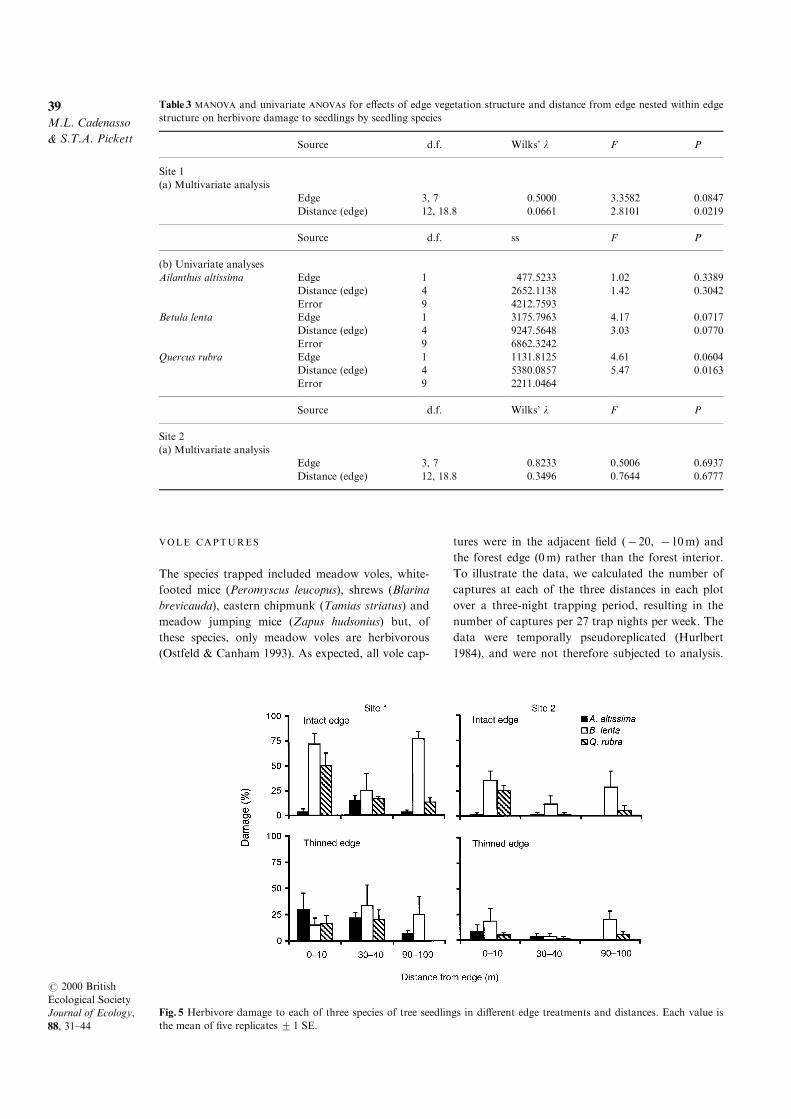
Table 2 MANOVA and univariate ANOVAs for e�ects of edge vegetation structure and distance from edge nested within edge
structure on tree seedling damage by herbivores
Source d.f. Wilks' l F P
Site 1
(a) Multivariate analysis
Edge 3, 22 0.4515 8.9090 0.0005
Distance (edge) 12, 58.5 0.1407 5.3565 0.0001
Source d.f. ss F P
(b) Univariate analyses
Deer Edge 1 50.4663 0.45 0.5067
Distance (edge) 4 2371.7083 5.34 0.0032
Error 24 2666.0335
Voles Edge 1 1408.2231 23.01 0.0001
Distance (edge) 4 4618.0465 18.86 0.0001
Error 24 1469.0088
Invertebrates Edge 1 173.8095 3.69 0.0666
Distance (edge) 4 176.9157 0.94 0.4579
Error 24 1129.4816
Source d.f. Wilks' l F P
Site 2
(a) Multivariate analysis
Edge 2, 23 0.9103 1.1339 0.3391
Distance (edge) 8, 46 0.4414 2.9048 0.0104
Source d.f. ss F P
(b) Univariate analyses
Deer Edge 1 160.7767 1.82 0.1902
Distance (edge) 4 516.4095 1.46 0.2455
Error 24 2123.0323
Voles Edge 1 1.0267 0.27 0.6104
Distance (edge) 4 71.9391 4.67 0.0063
Error 24 92.4742
38Linking forest
edge structure to
edge function
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44
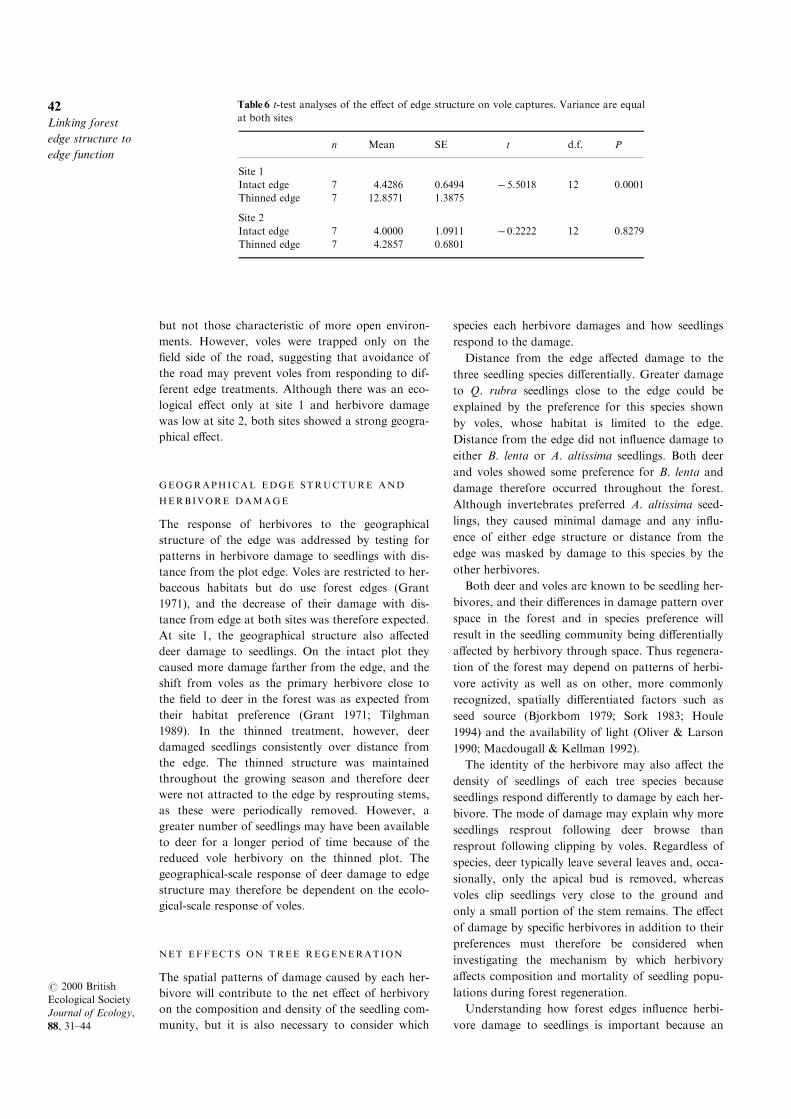
VOLE CAPTURES
The species trapped included meadow voles, white-
footed mice (Peromyscus leucopus), shrews (Blarina
brevicauda), eastern chipmunk (Tamias striatus) and
meadow jumping mice (Zapus hudsonius) but, of
these species, only meadow voles are herbivorous
(Ostfeld & Canham 1993). As expected, all vole cap-
tures were in the adjacent ®eld (ÿ 20, ÿ 10m) andthe forest edge (0m) rather than the forest interior.
To illustrate the data, we calculated the number of
captures at each of the three distances in each plot
over a three-night trapping period, resulting in the
number of captures per 27 trap nights per week. The
data were temporally pseudoreplicated (Hurlbert
1984), and were not therefore subjected to analysis.
Fig. 5 Herbivore damage to each of three species of tree seedlings in di�erent edge treatments and distances. Each value is
the mean of ®ve replicates 2 1 SE.
Table 3 MANOVA and univariate ANOVAs for e�ects of edge vegetation structure and distance from edge nested within edge
structure on herbivore damage to seedlings by seedling species
Source d.f. Wilks' l F P
Site 1
(a) Multivariate analysis
Edge 3, 7 0.5000 3.3582 0.0847
Distance (edge) 12, 18.8 0.0661 2.8101 0.0219
Source d.f. ss F P
(b) Univariate analyses
Ailanthus altissima Edge 1 477.5233 1.02 0.3389
Distance (edge) 4 2652.1138 1.42 0.3042
Error 9 4212.7593
Betula lenta Edge 1 3175.7963 4.17 0.0717
Distance (edge) 4 9247.5648 3.03 0.0770
Error 9 6862.3242
Quercus rubra Edge 1 1131.8125 4.61 0.0604
Distance (edge) 4 5380.0857 5.47 0.0163
Error 9 2211.0464
Source d.f. Wilks' l F P
Site 2
(a) Multivariate analysis
Edge 3, 7 0.8233 0.5006 0.6937
Distance (edge) 12, 18.8 0.3496 0.7644 0.6777
39M.L. Cadenasso
& S.T.A. Pickett
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44
However, at site 1, vole captures were consistently
greater on the thinned than on the intact plot; no
di�erences were apparent between treatments at site
2 (Fig. 8 and Table 6).
Discussion
Is the structure of the vegetation on the edge linked
to its function as re¯ected by herbivore ¯ux?
Herbivore damage to seedlings inside the cages was
minimal, showing that exclusion was successful,
while seedling mortality due to causes other than
herbivory was similar in the caged and open quad-
rats, indicating that cages did not introduce any
additional stresses. Further analyses of the responses
to herbivores were limited to the open quadrats.
ECOLOGICAL EDGE STRUCTURE AND
HERBIVORE DAMAGE
At site 1, the ecological structure of the edge had a
signi®cant e�ect on the amount of herbivore damage
to tree seedlings. This was primarily due to voles,
which were the dominant herbivore on the intact
plot but damaged signi®cantly fewer seedlings on
the thinned plot. We had expected vole activity to
be less on the thinned plot because the experimental
manipulation removed vegetation cover and might
therefore make voles more vulnerable to predation.
However, although voles reach higher densities in
herbaceous communities when low-statured vegeta-
tion cover is dense, and tend to avoid areas of
decreased vegetation cover (Birney et al. 1976; Peles
& Barrett 1996), the experimental thinning did not
alter ground layer vegetation. In addition, more
voles were captured outside the thinned treatment
but damage was greater in the intact plot. This
apparent contradiction may be explained by a com-
pensatory response of the ground layer to the
experimental manipulation. Removal of lateral
branches of the canopy trees and of small trees and
shrubs may have increased light in the thinned plot.
Although the resulting increase in herbaceous cover
may have encouraged voles, the production of more
herbaceous stems may have made tree seedlings on
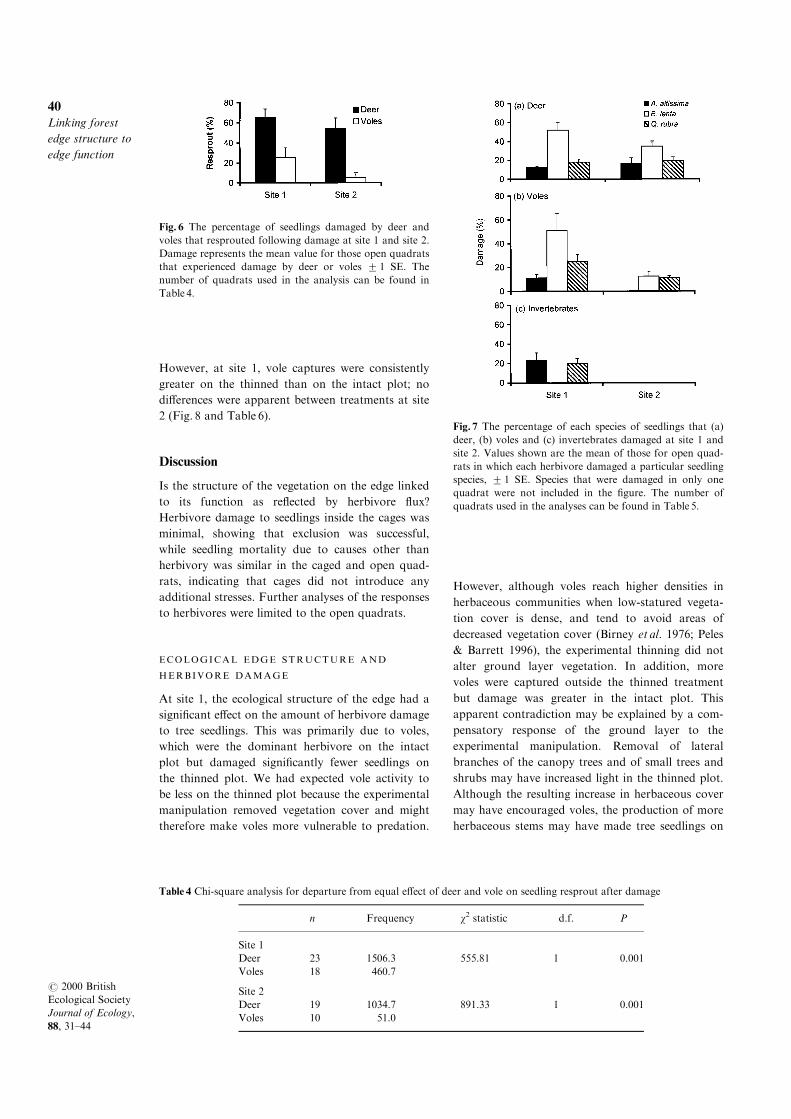
Fig. 6 The percentage of seedlings damaged by deer and
voles that resprouted following damage at site 1 and site 2.
Damage represents the mean value for those open quadrats
that experienced damage by deer or voles 2 1 SE. The
number of quadrats used in the analysis can be found in
Table 4.
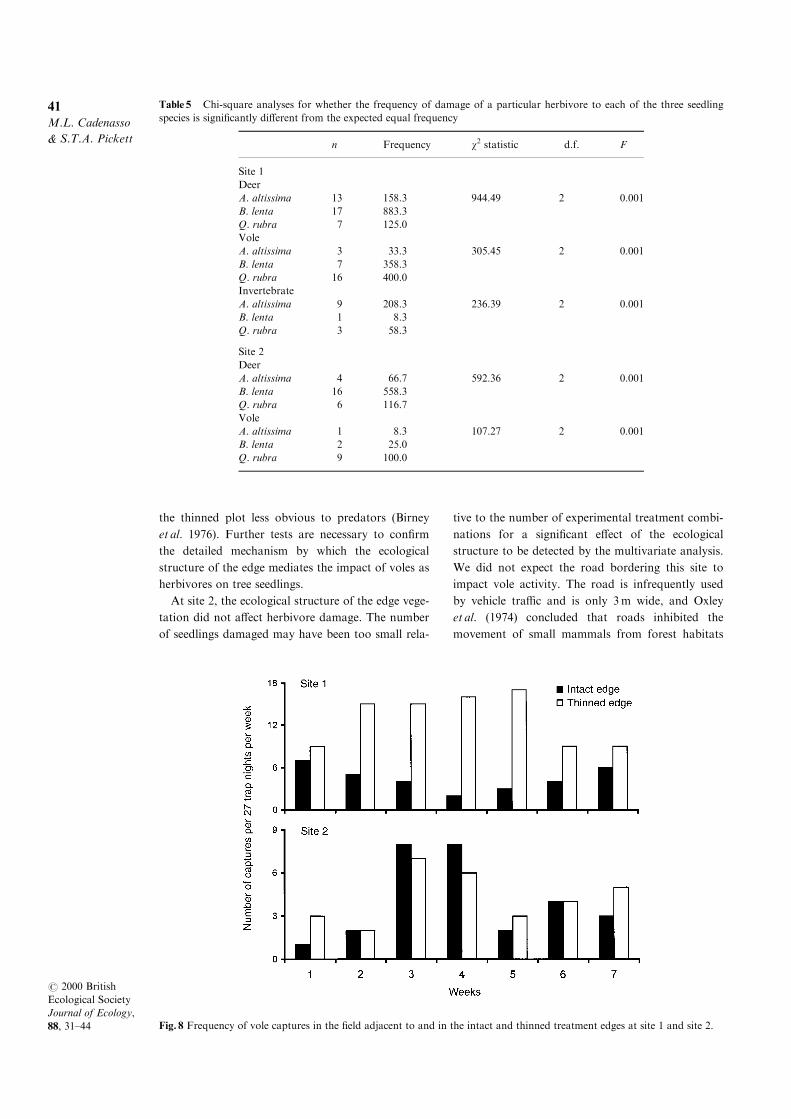
Fig. 7 The percentage of each species of seedlings that (a)
deer, (b) voles and (c) invertebrates damaged at site 1 and
site 2. Values shown are the mean of those for open quad-
rats in which each herbivore damaged a particular seedling
species, 2 1 SE. Species that were damaged in only one
quadrat were not included in the ®gure. The number of
quadrats used in the analyses can be found in Table 5.
Table 4 Chi-square analysis for departure from equal e�ect of deer and vole on seedling resprout after damage
n Frequency w2 statistic d.f. P
Site 1
Deer 23 1506.3 555.81 1 0.001
Voles 18 460.7
Site 2
Deer 19 1034.7 891.33 1 0.001
Voles 10 51.0
40Linking forest
edge structure to
edge function
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44
the thinned plot less obvious to predators (Birney
et al. 1976). Further tests are necessary to con®rm
the detailed mechanism by which the ecological
structure of the edge mediates the impact of voles as
herbivores on tree seedlings.
At site 2, the ecological structure of the edge vege-
tation did not a�ect herbivore damage. The number
of seedlings damaged may have been too small rela-
tive to the number of experimental treatment combi-
nations for a signi®cant e�ect of the ecological
structure to be detected by the multivariate analysis.
We did not expect the road bordering this site to
impact vole activity. The road is infrequently used
by vehicle tra�c and is only 3m wide, and Oxley
et al. (1974) concluded that roads inhibited the
movement of small mammals from forest habitats
Fig. 8 Frequency of vole captures in the ®eld adjacent to and in the intact and thinned treatment edges at site 1 and site 2.
Table 5 Chi-square analyses for whether the frequency of damage of a particular herbivore to each of the three seedling
species is signi®cantly di�erent from the expected equal frequency
n Frequency w2 statistic d.f. F
Site 1
Deer
A. altissima 13 158.3 944.49 2 0.001
B. lenta 17 883.3
Q. rubra 7 125.0
Vole
A. altissima 3 33.3 305.45 2 0.001
B. lenta 7 358.3
Q. rubra 16 400.0
Invertebrate
A. altissima 9 208.3 236.39 2 0.001
B. lenta 1 8.3
Q. rubra 3 58.3
Site 2
Deer
A. altissima 4 66.7 592.36 2 0.001
B. lenta 16 558.3
Q. rubra 6 116.7
Vole
A. altissima 1 8.3 107.27 2 0.001
B. lenta 2 25.0
Q. rubra 9 100.0
41M.L. Cadenasso
& S.T.A. Pickett
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44
but not those characteristic of more open environ-
ments. However, voles were trapped only on the
®eld side of the road, suggesting that avoidance of
the road may prevent voles from responding to dif-
ferent edge treatments. Although there was an eco-
logical e�ect only at site 1 and herbivore damage
was low at site 2, both sites showed a strong geogra-
phical e�ect.
GEOGRAPHICAL EDGE STRUCTURE AND
HERBIVORE DAMAGE
The response of herbivores to the geographical
structure of the edge was addressed by testing for
patterns in herbivore damage to seedlings with dis-
tance from the plot edge. Voles are restricted to her-
baceous habitats but do use forest edges (Grant
1971), and the decrease of their damage with dis-
tance from edge at both sites was therefore expected.
At site 1, the geographical structure also a�ected
deer damage to seedlings. On the intact plot they
caused more damage farther from the edge, and the
shift from voles as the primary herbivore close to
the ®eld to deer in the forest was as expected from
their habitat preference (Grant 1971; Tilghman
1989). In the thinned treatment, however, deer
damaged seedlings consistently over distance from
the edge. The thinned structure was maintained
throughout the growing season and therefore deer
were not attracted to the edge by resprouting stems,
as these were periodically removed. However, a
greater number of seedlings may have been available
to deer for a longer period of time because of the
reduced vole herbivory on the thinned plot. The
geographical-scale response of deer damage to edge
structure may therefore be dependent on the ecolo-
gical-scale response of voles.
NET EFFECTS ON TREE REGENERATION
The spatial patterns of damage caused by each her-
bivore will contribute to the net e�ect of herbivory
on the composition and density of the seedling com-
munity, but it is also necessary to consider which
species each herbivore damages and how seedlings
respond to the damage.
Distance from the edge a�ected damage to the
three seedling species di�erentially. Greater damage
to Q. rubra seedlings close to the edge could be
explained by the preference for this species shown
by voles, whose habitat is limited to the edge.
Distance from the edge did not in¯uence damage to
either B. lenta or A. altissima seedlings. Both deer
and voles showed some preference for B. lenta and
damage therefore occurred throughout the forest.
Although invertebrates preferred A. altissima seed-
lings, they caused minimal damage and any in¯u-
ence of either edge structure or distance from the
edge was masked by damage to this species by the
other herbivores.
Both deer and voles are known to be seedling her-
bivores, and their di�erences in damage pattern over
space in the forest and in species preference will
result in the seedling community being di�erentially
a�ected by herbivory through space. Thus regenera-
tion of the forest may depend on patterns of herbi-
vore activity as well as on other, more commonly
recognized, spatially di�erentiated factors such as
seed source (Bjorkbom 1979; Sork 1983; Houle
1994) and the availability of light (Oliver & Larson
1990; Macdougall & Kellman 1992).
The identity of the herbivore may also a�ect the
density of seedlings of each tree species because
seedlings respond di�erently to damage by each her-
bivore. The mode of damage may explain why more
seedlings resprout following deer browse than
resprout following clipping by voles. Regardless of
species, deer typically leave several leaves and, occa-
sionally, only the apical bud is removed, whereas
voles clip seedlings very close to the ground and
only a small portion of the stem remains. The e�ect
of damage by speci®c herbivores in addition to their
preferences must therefore be considered when
investigating the mechanism by which herbivory
a�ects composition and mortality of seedling popu-
lations during forest regeneration.
Understanding how forest edges in¯uence herbi-
vore damage to seedlings is important because an
Table 6 t-test analyses of the e�ect of edge structure on vole captures. Variance are equal
at both sites
n Mean SE t d.f. P
Site 1
Intact edge 7 4.4286 0.6494 ÿ5.5018 12 0.0001
Thinned edge 7 12.8571 1.3875
Site 2
Intact edge 7 4.0000 1.0911 ÿ0.2222 12 0.8279
Thinned edge 7 4.2857 0.6801
42Linking forest
edge structure to
edge function
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44

increasing proportion of the forested landscape is in
close proximity to edges. Data from sightings of
deer in spotlights has provided a conservative popu-
lation estimate for white-tailed deer on the IES
property at 18 kmÿ 2 (Winchcombe 1993), and
because this is considerably higher than that sug-
gested to inhibit forest regeneration in Wisconsin (4
kmÿ 2) (Alverson et al. 1988), deer may strongly
in¯uence seedling density and composition at our
sites. Deer traverse forest edges every dawn and
dusk (Montgomery 1963) and their interaction with
edge structure and its in¯uence on forest dynamics
may therefore be much greater than expected from
the proportion of space occupied by edges.
This is the ®rst experimental test of the function
of forest edges on plant community regeneration. It
con®rms the suggested importance of forest edge
function as a mediator of herbivore impacts
(Alverson et al. 1988) and suggests hypotheses for
more detailed mechanistic studies. In addition, the
technique of altering the ecological structure of a
forest edge while keeping the geographical scale con-
®guration of the edge intact may be a useful strategy
for studying di�erent landscape ¯uxes and how they
interact with di�erent types of edges to determine
edge functions.
Acknowledgements
We would like to thank Julie E. Hart and M.
Michael Traynor for extensive ®eld assistance in
cage construction, seedling planting and monitoring.
Dave Bulkeley and the IES Greenhouse sta� helped
grow and maintain the seedlings until they were
planted in the ®eld. The cutting of vegetation on the
forest edge was done by Brad Roeller, Alan Kling
and the IES Grounds crew. Peter Morin provided
valuable statistical advice. The manuscript has been
greatly improved by comments from Charlie Nilon,
Rick Ostfeld, John Wiens and Richard Forman.
This research was funded by the NSF, grant DEB
9307252.
References
Alverson, W.S., Waller, D.M. & Solheim, S.L. (1988)
Forests too deer: edge e�ects in northern Wisconsin.
Conservation Biology, 2, 348±358.
Andre n, H. (1995) E�ects of landscape composition on
predation rates at habitat edges. Mosaic Landscapes
and Ecological Processes (eds L. Hansson, L. Fahrig &
G. Merriam), pp. 225±255. Chapman & Hall, London,
UK.
Andre n, H. & Angelstam, P. (1988) Elevated predation
rates as an edge e�ect in habitat islands: experimental
evidence. Ecology, 69, 544±547.
Angelstam, P. (1992) Conservation of communities ± the
importance of edges, surroundings, and landscape
mosaic structure. Ecological Principles of Nature
Conservation (ed. L. Hansson), pp. 9±70. Elsevier
Applied Science, New York, NY.
Armesto, J.J., Pickett, S.T.A. & McDonnell, M.J. (1991)
Spatial heterogeneity during succession: a cyclic model
of invasion and exclusion. Ecological Heterogeneity
(eds J. Kolasa & S.T.A. Pickett), pp. 256±269.
Springer-Verlag, New York, NY.
Birney, E.C., Grant, W.E. & Baird, D.D. (1976)
Importance of vegetative cover to the cycles of
Microtus populations. Ecology, 57, 1043±1051.
Bjorkbom, J.C. (1979) Seed Production and Advance
Regeneration in Allegheny Hardwood Forests. Forest
Service Research Paper NE-435. Forest Service, US
Department of Agriculture, Northeastern Forest
Experiment Station, Radnor, Pennsylvania.
Brothers, T.S. & Spingarn, A. (1992) Forest fragmentation
and alien plant invasion of Central Indiana old-growth
forests. Conservation Biology, 6, 91±100.
Cadenasso, M.L., Traynor, M.M. & Pickett, S.T.A. (1997)
Functional location of forest edges: gradients of multi-
ple physical factors. Canadian Journal of Forest
Research, 27, 774±782.
Camargo, J.L.C. & Kapos, V. (1995) Complex edge e�ects
on soil moisture and microclimate in central
Amazonian forest. Journal of Tropical Ecology, 11,
205±221.
Chen, J., Franklin, J.F. & Spies, T.A. (1992) Vegetation
responses to edge environments in old-growth
Douglas-®r forests. Ecological Applications, 2, 387±
396.
Forman, R.T.T. (1995) Land Mosaics: the Ecology of
Landscapes and Regions. Cambridge University Press,
New York, NY.
Forman, R.T.T. & Moore, P.N. (1992) Theoretical founda-
tions for understanding boundaries in landscape
mosaics. Landscape Boundaries. Consequences for
Biotic Diversity and Ecological Flows (eds A.J. Hansen
& F. di Castri), pp. 236±258. Springer-Verlag, New
York, NY.
Geiger, R. (1965) The Climate Near the Ground. Harvard
University Press, Cambridge, MA.
Gill, D.S. & Marks, P.L. (1991) Tree and shrub seedling
colonization of old ®elds in central New York.
Ecological Monographs, 61, 183±205.
Gill, R.M.A. (1992) A review of damage by mammals in
north temperate forests. I. Deer. Forestry, 65, 145±169.
Gleason, H.A. & Cronquist, A. (1991) Manual of Vascular
Plants of Northeastern United States and Adjacent
Canada. The New York Botanical Garden, Bronx,
NY.
Grant, P.R. (1971) The habitat preference of Microtus
pennsylvanicus, and its relevance to the distribution of
this species on islands. Journal of Mammalogy, 52,
351±361.
Hansson, L. (1992) Introduction: applied ecological princi-
ples and their implementation in nature conservation.
Ecological Principles of Nature Conservation (ed.
L. Hansson), pp. 1±8. Elsevier Applied Science,
London, UK.
Harris, L.D. (1988) Edge e�ects and conservation of biotic
diversity. Conservation Biology, 2, 330±332.
Houle, G. (1994) Spatiotemporal patterns in the compo-
nents of regeneration of four sympatric species ± Acer
rubrum, A. saccharum, Betula alleghaniensis and Fagus
grandifolia. Journal of Ecology, 82, 39±53.
Hurlbert, S.H. (1984) Pseudoreplication and the design of
ecological ®eld experiments. Ecological Monographs,
54, 187±211.
Laurance, W.F. & Yensen, E. (1991) Predicting the
impacts of edge e�ects in fragmented habitats.
Biological Conservation, 55, 77±92.
43M.L. Cadenasso
& S.T.A. Pickett
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44

Leopold, A. (1936) Game Management. Charles Scribner's
Sons, New York, NY.
Levenson, J.B. (1981) Woodlots as biogeographic islands
in southeastern Wisconsin. Forest Island Dynamics in
Man-Dominated Landscapes (eds R.L. Burgess &
D.M. Sharpe), pp. 13±39. Springer-Verlag, New York,
NY.
Macdougall, A. & Kellman, M. (1992) The understorey
light regime and patterns of tree seedlings in tropical
riparian forest patches. Journal of Biogeography, 19,
667±675.
Matlack, G.R. (1993) Microenvironment variation within
and among forest edge sites in the eastern United
States. Biological Conservation, 66, 185±194.
Matlack, G.R. (1994) Vegetation dynamics of the forest
edge ± trends in space and successional time. Journal
of Ecology, 82, 113±123.
Montgomery, G.G. (1963) Nocturnal movements and
activity rhythms of white-tailed deer. Journal of
Wildlife Management, 27, 422±427.
Moran, M.A. (1984) In¯uence of adjacent land use on
understorey vegetation of New York forests. Urban
Ecology, 8, 329±340.
Murcia, C. (1995) Edge e�ects in fragmented forests: impli-
cations for conservation. Trends in Ecology and
Evolution, 10, 58±62.
Myster, R.W. & McCarthy, B.C. (1989) E�ects of herbiv-
ory and competition on survival of Carya tomentosa
(Juglandaceae) seedlings. Oikos, 56, 145±148.
Oliver, C.D. & Larson, B.C. (1990) Forest Stand Dynamics.
McGraw-Hill Inc., New York, NY.
Ostfeld, R.S. & Canham, C.D. (1993) E�ects of meadow
vole population density on tree seedling survival in old
®elds. Ecology, 74, 1792±1801.
Ostfeld, R.S., Manson, R.H. & Canham, C.D. (1997)
E�ects of rodents on survival of tree seeds and seed-
lings invading old ®elds. Ecology, 78, 1531±1542.
Oxley, D.J., Fenton, M.B. & Carmody, G.R. (1974) The
e�ects of roads on populations of small mammals.
Journal of Applied Ecology, 11, 51±59.
Paton, P.W.C. (1994) The e�ect of edge on avian nest suc-
cess: how strong is the evidence? Conservation Biology,
8, 17±26.
Peles, J.D. & Barrett, G.W. (1996) E�ects of vegetation
cover on the population dynamics of meadow voles.
Journal of Mammalogy, 77, 857±869.
Pickett, S.T.A. & Cadenasso, M.L. (1995) Landscape ecol-
ogy: spatial heterogeneity in ecological systems.
Science, 269, 331±334.
Rankin, W.T. & Pickett, S.T.A. (1989) Time of establish-
ment of red maple (Acer rubrum) in early old®eld suc-
cession. Bulletin of the Torrey Botanical Club, 116,
182±186.
Ranney, J.W., Bruner, M.C. & Levenson, J.B. (1981) The
importance of edge in the structure and dynamics of
forest islands. Forest Island Dynamics in Man-
Dominated Landscapes (eds R.L. Burgess &
D.M. Sharpe), pp. 67±96. Springer-Verlag, New York,
NY.
Saunders, D.A., Hobbs, R.J. & Margules, C.R. (1991)
Biological consequences of ecosystem fragmentation: a
review. Conservation Biology, 5, 18±32.
Sork, V.L. (1983) Distribution of pignut hickory (Carya
glabra) along a forest to edge transect, and factors
a�ecting seedling recruitment. Bulletin of the Torrey
Botanical Club, 110, 494±506.
Stamps, J.A., Buechner, M. & Krishnan, V.V. (1987) The
e�ects of edge permeability and habitat geometry on
emigration from patches of habitat. American
Naturalist, 129, 533±552.
Tilghman, N.G. (1989) Impacts of white-tailed deer on for-
est regeneration in northwestern Pennsylvania. Journal
of Wildlife Management, 53, 524±532.
Wales, B.A. (1967) Climate, microclimate and vegetation
relationships on north and south forest boundaries in
New Jersey. William L. Hutcheson Memorial Forest
Bulletin, 2, 1±60.
Wales, B.A. (1972) Vegetation analysis of north and south
edges in a mature oak±hickory forest. Ecological
Monographs, 42, 451±471.
Weathers, K.C., Lovett, G.M. & Likens, G.E. (1995)
Cloud deposition to a spruce forest edge. Atmospheric
Environment, 29, 665±672.
Wiens, J.A. (1992) Ecological ¯ows across landscape
boundaries: a conceptual overview. Landscape
Boundaries. Consequences for Biotic Diversity and
Ecological Flows (eds A.J. Hansen & F. di Castri), pp.
217±235. Springer-Verlag, New York, NY.
Wiens, J.A., Crawford, C.S. & Gosz, J.R. (1985) Boundary
dynamics: a conceptual framework for studying land-
scape ecosystems. Oikos, 45, 421±427.
Wiens, J.A., Stenseth, N.C., Van Horne, B. & Ims, R.A.
(1993) Ecological mechanisms and landscape ecology.
Oikos, 66, 369±380.
Williams-Linera, G. (1990) Vegetation structure and envir-
onmental conditions of forest edges in Panama.
Journal of Ecology, 78, 356±373.
Williamson, S.J. & Hirth, D.H. (1985) An evaluation of
edge use by white-tailed deer. Wildlife Society Bulletin,
13, 252±257.
Winchcombe, R.J. (1993) Controlled access hunting for
deer population management: a case study. Northeast
Wildlife, 50, 1±9.
Received 29 October 1998
revision accepted 13 July 1999
44Linking forest
edge structure to
edge function
# 2000 British
Ecological Society
Journal of Ecology,
88, 31±44