late cretaceous (santonian-maastrichtian) serial...
TRANSCRIPT

Late Cretaceous (Santonian-Maastrichtian) serial foraminiferawith pore mounds or pore mound-based ornamentation
structures
Marius Dan Georgescu1, José Antonio Arz2, Ryan Victor Macauley1,
Ross Brian Kukulski1, Ignacio Arenillas2 and Irene Pérez-Rodríguez2
1 Department of Geoscience, University of Calgary, 2500 University Drive N.W., Calgary, Alberta, T2N 1N4, Canada. [email protected]
2 Departamento de Ciencias de la Tierra (Paleontología) & Instituto Universitario de Ciencias Ambientales de Aragón (IUCA), Universidad de Zaragoza, Pedro Cerbuna 12, E-50009 Zaragoza, Spain.
Resumen
La ornamentación de poros en túmulo ha sido identificada en foraminíferos planctónicos y bentónicos del Cretácico Superior (San-toniense-Maastrichtiense) con disposición de las cámaras biseriada y/o triseriada. El estudio de imágenes de alta resolución conSEM/ESEM muestra que los poros en túmulo varían tanto en su forma (circular y elíptica) como en su grado de desarrollo (incipienteo bien desarrollado). Los patrones de distribución de los poros en túmulo muestran gran variabilidad, siendo el más frecuente el re-presentado por poros en túmulo dispersos. Los poros en túmulo pueden fusionarse para formar estructuras como quillas [p. ej., Pseu-douvigerina cristata (Marsson, 1878)] y plataformas ornamentadas [p.ej., Trachelinella watersi (Cushman, 1927a)]. Se propone unnuevo género, Elongateporeia, para un grupo de foraminíferos bentónicos biseriados del Santoniente-Campaniense superior con porosen túmulo elípticos, de incipientes a bien desarrollados, y poro de alargado a lineal. Se describen tres nuevas especies: Bolivinoidesmirabilis, Elongateporeia elongatoporosa y Neobulimina newjerseyensis.
Palabras clave: Foraminíferos bentónicos, foraminíferos planctónicos, Cretácico Superior, nuevo género, nueva especie.
Abstract
Pore mounded ornamentation has been reported from Late Cretaceous (Santonian-Maastrichtian) benthic and planktic foraminifera withbiserial and/or triserial chamber arrangement. This high resolution SEM/ESEM-based study shows that the pore mounds are morpho-logically diverse in shape (circular and elliptical) and degree of development (incipient and well-developed). Pore mound distributionpatterns show significant variability, with scattered pore mounds observed most often. Pore mounds can fuse to form pore mound-basedstructures, such as keels [i.e., Pseudouvigerina cristata (Marsson, 1878)] and ornamented platforms [i.e., Trachelinella watersi (Cush-man, 1927a)]. A new genus, Elongateporeia, is identified for a group of late Santonian-late Campanian biserial benthics with elongateto slit-like pores ornamented with incipient to well-developed elliptical pore mounds. Three new species are described: Bolivinoidesmirabilis, Elongateporeia elongatoporosa and Neobulimina newjerseyensis.
Keywords: Benthic foraminifera, planktic foraminifera, Late Cretaceous, new genus, new species.
109
Revista Española de Micropaleontología, 43 (1-2), 2011, pp. 109-139©Instituto Geológico y Minero de EspañaISSN: 0556-655X

110
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
1. INTRODUCTION
Pore mounds are relatively rare ornamentation featuresfound on some foraminiferal tests. They are small struc-tures with a diameter rarely exceeding five microns andfor this reason they are rarely observed under the classicaloptical stereomicroscope. The Cretaceous record showsthat pore mound ornamentation occurs in only a smallnumber of planktic foraminifera: Guembelitria cretaceaCushman, 1933, G. blowi Arz et al., 2010, G. dammulaVoloshina, 1961, Archaeoguembelitria harrisi (Tappan,1940), Laeviheterohelix pulchra (Brotzen, 1936), L. reni-formis (Marie, 1941), Steineckia steinecki Georgescu,2009a and Hedbergella infracretacea (Glaessner, 1937).Pore mounds were also reported by Gawor-Biedowa(1992) in some benthic foraminiferal species recoveredfrom Campanian-Maastrichtian aged sediments of Poland:Bolivina praecrenulata Gawor-Biedowa, 1992, B. aleksan-drae Gawor-Biedowa, 1992, Pyramidina rudita (Cushman& Parker, 1936), Pseudouvigerina cristata (Marsson, 1878),Biedafrancizkina beaumonti (Cushman & Renz, 1946)and Pseudouvigerina rugosa Brotzen, 1945 [generic as-signments are from Gawor-Biedowa (1992)].
Scanning Electron Microscope (SEM) and EnvironmentalScanning Electron Microscope (ESEM) has provided insightinto foraminiferal taxonomy by allowing observations ondetailed test morphological features. It allowed the defini-tion of two heterohelicid genera based primarily on poremound ornamentation: Laeviheterohelix Nederbragt, 1991– emended Georgescu 2009a and Steineckia Georgescu,2009a. Systematic study of the test ornamentation using theSEM/ESEM technology has helped to better define the poremound development processes illustrated in Georgescu(2009b, pl. 6, fig. 6) where “incipient pore mounds” weredescribed in the triserial planktic species A. harrisi of theuppermost Albian-lowermost Turonian. In this case, incip-ient pore mounds were formed through the gradual eleva-tion of the test wall. A planktic habitat for A. harrisi was in-ferred based on this ornamentation (Georgescu, 2009b, p.138). An alternate process of pore mound formationthrough the modification of costae was recognized byGeorgescu (2009a, p. 331) in the biserial planktic L. pul-chra. These studies document the incremental developmentof our knowledge base and provide justification for adopt-ing an evolutionary classification framework for taxa withpore mound ornamentation (Georgescu, 2009a, 2009b).
This study focuses on some benthic and plankticforaminifera with biserial and/or triserial chamber arrange-ment and continues the effort to examine the evolutionaryimplications of pore mound occurrence and developmentin Cretaceous foraminifera. It represents the first step inthe development of a lineage-based classification systemfor small Cretaceous benthic foraminifera and in under-standing of the origin of Guembelitria Cushman, 1933.
2. ORNAMENTATION AND POROSITYTERMINOLOGY ANDMEASUREMENTS
This high resolution study of pore mound shapes revealedthe complexity of this ornamentation feature. Two typesof pore mounds were recognized and classified accord-ing to the degree of development: incipient (Fig. 1A, 1C)and well-developed (Fig. 1B, 1D). The incipient poremounds represent an early stage of formation and canonly be observed with the aid of the ESE or ESEM. Well-developed pore mounds are larger than incipient poremounds, sharply separated from the test wall and can oc-casionally be observed using an optical stereomicroscope.A parallel pore mound classification can be made accord-ing to their shape: circular (Fig. 1A-B) and elliptical (Fig.1C-D). In order to increase the precision of description,the terminology used herein combines the two classifica-tions into a four-fold one: incipient circular (Fig. 1A), well-developed circular (1B), incipient elliptical (1C) andwell-developed elliptical (Fig. 1D). Measurements andcorresponding symbols of the previously discussed fea-tures are outlined in Figure 1 and Table 1A.
The species included in this study also exhibit two otherornamentation elements, pustules and tubercle-like struc-tures. Their dimensions are recorded as pustule diameter(Øp) and maximum dimension (Dmax) respectively.
A classification of the pores according to their shape isproposed herein: circular (Fig. 2A), elliptical (Fig. 2B) andslit-like (Fig. 2C), with slit-like pores showing straight orundulated forms. The dimensions measured for each ofthese pore types are differentially labelled (Fig. 2, Table1B).

111
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Figure 1. Pore mound terminology and measurements. A, incipient circular. B, well-developed circular. C, incipient elliptical. D, well-developed el-liptical. Measurement symbols as in Table 1.
Table 1. Pore mound and pore terminology, measured parametersand their corresponding symbols.

112
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
3. MATERIAL PROVENANCE
Most of the material examined in this study comes fromupper Santonian-Maastrichtian sediments of the UnitedStates of America (New Jersey and Texas) and Deep SeaDrilling Project Hole 111A (Orphan Knoll, North AtlanticOcean) (Fig. 3). Sample labelling is different for each lo-cation and is presented in the corresponding sections ofthis chapter. The original sample labelling system wasused to remain consistent with the previous studies.
The holotypes of Trachelinella watersi (Cushman) andPseudouvigerina plummerae Cushman from the CushmanCollection at the National Museum of Natural History(NMNH), Washington, D.C. were studied in order to com-pare observations made on newly acquired material withprevious benthic descriptions. The holotypes of these twospecies are re-illustrated with the aid of the ESEM. Topo-types of Guembelitria cretacea Cushman were examinedin the Loeblich and Tappan Topotype Collection (NMNH).Material from the McGugan Collection of late Campan-ian-early Maastrichtian age (University of Calgary, Alberta,Canada) was also examined; it includes samples collectedby Dr. Alan McGugan during the Sixth Annual Field Tripof the Alabama Geological Society, December 6-7, 1968(Scott et al., 1968). Two lithostratigraphic units, the RipleyFormation and the Prairie Bluff Chalk, were sampled fromfour outcrops located in the Lowndes and Montgomerycounties of Alabama. The foraminiferal assemblages indi-cate a late Campanian-early Maastrichtian age, consistent
with the age assignments by Mancini et al. (1996),Mancini & Puckett (2005) and Puckett (2005). Two ben-thic foraminiferal species ornamented with pore moundsand pore mound-based structures are found in these sed-iments: Pseudouvigerina rudita (Cushman & Parker, 1936)and P. cristata (Marsson, 1878). The planktic triserial genusGuembelitria is also frequent to abundant in most of thestudied samples. Additional material was collected fromthe Upper Cretaceous El Kef and Assila sections of Tunisia(northern Africa) where the three species of CretaceousGuembelitria were identified (Arz et al., 2010).
The type material of the new species is deposited in theWilli Karl Braun Collection (University of Calgary, Alberta,Canada). They are labelled with the standard collectionacronym (WKB) followed by the inventory number.
The biostratigraphic data referred to in this study adhere tothe standard planktic foraminiferal framework set out byRobaszynski & Caron (1995). Local planktic foraminiferalframeworks [e.g., Georgescu (2006) for the Santonian-Campanian sediments of the New Jersey coastal plain,etc] were used whenever necessary due to the frequentabsence of deep-water planktic foraminiferal species inthe inner and middle shelf environments.
3.1. ODP Leg 174AX, New Jersey coastalplain (USA)
Well-preserved benthic foraminifera with pore mound or-namentation were collected from the Merchantville and
Figure 2. Pore terminology and measurements. A, circular. B, elliptical. C, slit-like. Measurement symbols as in Table 1.

113
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Marshalltown formations of the Bass River borehole of theODP Leg 174AX (geographical coordinates: 39o 36` 42”N and 74o 26` 12” W). Sample depths are given in metersbelow surface (mbsf). The Merchantville and Marshall-town formations are two transgressive units of late San-tonian and late Campanian age respectively (Petters,1976, 1977a, b; Olsson & Nyong, 1984; Miller et al.,1998; Georgescu, 2006, 2010; Georgescu & Abramovich,2008). They preserve the transgressive systems tracts oftwo sedimentary cycles, namely the Cheesequake-Mer-chantville of late Santonian-Campanian age and the Mar-shalltown, of late Campanian age.
Foraminiferal preservation in the two claystone dominatedtransgressive units is variable. Complete well preservedtests are commonly recovered from the Merchantville For-mation due to deposition in shelf deeper waters wherecurrent energy is low. The tests in the upper Marshalltownpresent little diagenetic alteration but are strongly frag-mented reflecting the higher current energy and shallowernature of the Marshalltown sedimentary cycle when com-pared to the underlying Cheesequake-Merchantville suc-cession.
Benthic foraminifera with pore mounds have only beenobserved in the two transgressive units. Elongateporeiaculverensis (Barr, 1967) is abundant in the upper part of
the Merchantville Formation and apparently is the onlyspecies described with pore mounds below the Santon-ian/Campanian boundary. The uppermost part of the Mar-shalltown Formation yielded a diverse assemblage ofbenthic foraminifera with pore mound ornamentation: Bo-livinoides mirabilis – new species, Elongateporeia elon-gatoporosa – new species and Neobuliminanewjerseyensis – new species. In addition, the sedimentsof the upper Marshalltown Formation produced B. sp., ex.gr. B. decorata (Jones in Wright, 1886) specimens, whichpresent irregular elevated areas that occasionally resemblepore mounds. Species with pore mound ornamentation inupper Marshalltown sediments are rare and diminutiveand for this reason they are easily overlooked when as-semblages are examined by optical microscope alone.
3.2. Mullinax-1 and Mullinax-3 wells, Texas(USA)
Abundant, well-preserved upper Maastrichtianforaminifera with pore mounds and pore mound-basedstructures were examined from the Corsicana Formationof two wells from Texas, Mullinax-1 (geographical coordi-nates: 31o 07’ N and 96o 49’ W) and Mullinax-3 (geo-graphical coordinates: 31o 06’ N and 96o 50’ W). Samplesare labelled as follows: well name, sample number-corenumber-section number, sample top depth. The
Figure 3. Provenance of the studied material: 1 – Texas (USA), 2 – Alabama (USA), 3 – New Jersey (USA), 4 – Orphan Knoll (North Atlantic Ocean) and5 – Tunisia (northern Africa). The paleogeographic map at 100 m.y. is redrawn and simplified after Hay et al. (1999).

114
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
foraminiferal assemblages and biostratigraphic frameworkat these two locations were extensively studied by Keller(1989) and Keller et al. (2007, 2009) and are followedherein. Beds from the Corsicana Formation that yieldedforaminiferal tests accumulated in shallower water, underrelatively quiet conditions, resulting in excellent preser-vation.
Benthic foraminiferal species identified in a succession offour samples from the upper Maastrichtian sediments(Abathomphalus mayaroensis Biozone equivalent) of theMullinax-1 well yielded the following species with poremounds and pore mound-based structures: T. watersi,Pseudouvigerina rudita and P. cristata. Tests of the triserialplanktic species Guembelitria cretacea are frequent toabundant (Keller, 2002; Keller & Pardo, 2004). The samebenthic foraminiferal species ornamented with poremounds and pore mound-based structures are reportedfrom a succession of nine samples from the Corsicana For-mation upper Maastrichtian sediments (Pseudoguembe-lina hariaensis Biozone equivalent) from the Mullinax-3well.
3.3. DSDP Hole 111A, Orphan Knoll (NorthAtlantic Ocean)
The only occurrence of foraminifera with pore moundsand pore mound-based ornamentation structures in deepoceanic sediments is known in the DSDP Hole 111Adrilled in the Orphan Knoll, North Atlantic Ocean (geo-graphical coordinates: 50o 26’ N and 46o 22’ W). Creta-ceous sediments were only recovered in core 11 and theywere assigned to the upper Maastrichtian (Abathomphalusmayaroensis Biozone). The upper Maastrichtian was stud-ied in a succession of 11 samples, all labelled accordingto the DSDP/Ocean Drilling Program/Integrated OceanDrilling Program standards as follows: leg number-site/hole number-core number-core section, Sample incentimetres.
Pseudouvigerina cristata provides a minor component ofthe foraminiferal assemblages throughout this section. Itis the only species with pore mound ornamentation andtest periphery lined by keels from fused pore mounds. Incontrast to the material examined from other occurrences,the tests of P. cristata at DSDP Hole 111A are diageneti-cally altered. Although completely recrystallized, the dis-
crete morphological features are still preserved and over-all the material is considered suitable for a taxonomicalstudy.
4. SYSTEMATIC CLASSIFICATION
Higher classification typological units are after Loeblich& Tappan (1984, 1987).
Order FORAMINIFERIDA Eichwald, 1830Suborder ROTALIINA Delage & Hérouard, 1896
Superfamily BOLIVINACEA Glaessner, 1937Family BOLIVINOITIDAE Loeblich & Tappan, 1984
Genus Bolivinoides Cushman, 1927a – emended
Type species: Bolivina draco Marsson, 1878 (originaldesignation).
1927a Bolivinoides Cushman, p. 89.1927b Bolivinoides Cushman, 1927 – Cushman, p. 61.1927c Bolivinoides Cushman, 1927 – Cushman, p.
158.1929 Bolivinoides Cushman, 1927 – White, p. 44.1950 Bolivinoides Cushman, 1927 – Hiltermann &
Koch, p. 595.1964 Bolivinoides Cushman, 1927 – Beckmann &
Koch, p. 37.1964 Bolivinoides Cushman, 1927 – Loeblich & Tap-
pan, p. C549.1966 Bolivinoides Cushman, 1927 – Barr, p. 227.1987 Bolivinoides Cushman, 1927 – Loeblich & Tap-
pan, p. 500.
Description.– Test is biserial throughout, with rhomboidaloutline. Chambers are broad and low and increase grad-ually in size resulting in a distinct adapical test flaring.Chambers are wider than high, reniform and oblique tothe test growth axis; each chamber presents two or morebackward extensions over the previous suture and cham-ber. Sutures are depressed and oblique to the test growthaxis and often obscured by the successive chamber back-ward extensions. Test compressed in edge view; periph-ery is rounded to subangular, without peripheralstructures. Aperture is an elongate opening on the lastchamber apertural face and provided with a simple or
115
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
curved toothplate; an oblique supplementary suture con-nects the aperture base with the suture between the last-formed and penultimate chambers. Chamber surfacesmooth or, more rarely, ornamented with strong tubercles,pustules and pore mounds. Test wall is calcitic, hyalineand perforate; pores are circular to subpolygonal, occa-sionally elliptical in shape, often concentrated on the an-terior side of the chamber.
Remarks.– Bolivinoides differs from the Cretaceousspecies of Bolivina d’Orbigny, 1839 by having well devel-oped chamber backward extensions, which result in afolded test surface and areal aperture rather than at thebase of the last formed chamber. It differs from TappaninaMontanaro Gallitelli, 1955 and Cretaceous species ofAragonia Finlay, 1939 by having chambers with backwardextensions and depresses sutures, rather than lined by nar-row, blade-like ridges. The test ornamentation in Bolivi-noides is reinterpreted and chamber backward extensionsare considered elements of the gross test architecturerather than ornamentation. Bolivinoides gross test archi-tecture (e.g., rhomboidal and compressed tests, chamberswith backward extensions, aperture provided with a tooth-plate, etc) is maintained throughout group evolution withsmall changes; they were used in the past to deciphergroup evolution (Edgell, 1954; Hofker, 1957; Hiltermann,
1963; Barr, 1966; Petters, 1977b). Notably, no studies fo-cused on the test ultrastructure, porosity characteristicsand ornamentation patterns as potential indicators in rec-ognizing the ancestor-descendant relationships betweenthe species of Bolivinoides. Therefore, Bolivinoidesspecies classification is entirely typological.
Stratigraphic range.– Upper Santonian–Paleocene.
Geographic distribution.– Cosmopolitan.
Bolivinoides mirabilis new speciesPlate 1, Figs 1-10
Holotype.– WKB 010110.
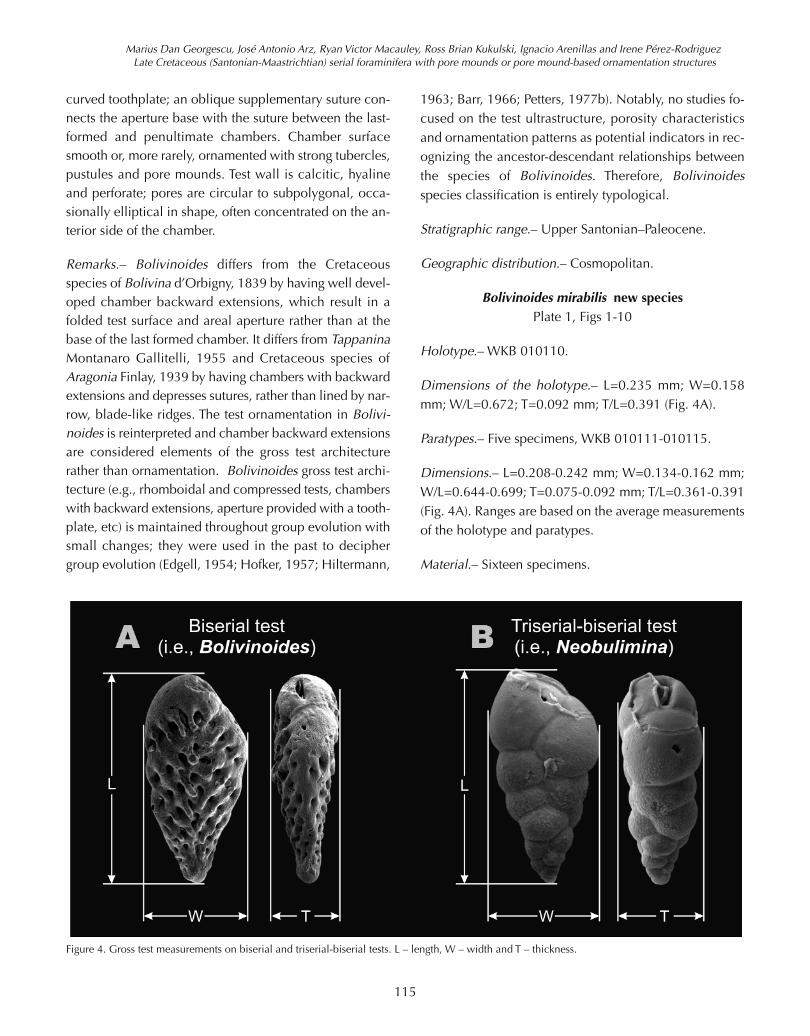
Dimensions of the holotype.– L=0.235 mm; W=0.158mm; W/L=0.672; T=0.092 mm; T/L=0.391 (Fig. 4A).
Paratypes.– Five specimens, WKB 010111-010115.
Dimensions.– L=0.208-0.242 mm; W=0.134-0.162 mm;W/L=0.644-0.699; T=0.075-0.092 mm; T/L=0.361-0.391(Fig. 4A). Ranges are based on the average measurementsof the holotype and paratypes.
Material.– Sixteen specimens.
Figure 4. Gross test measurements on biserial and triserial-biserial tests. L – length, W – width and T – thickness.

116
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
Type locality.– ODP Leg 174AX (Bass River Site); geo-graphical coordinates: 39o 36’ 42”N and 74o 26’ 12” W.
Type level.– The green silts of the upper Campanian Mar-shalltown Formation; sample depth at the type locality:435.85-435.86 cm.
Etymology. – Name derived from the Latin adjective“mirabilis” (=wonderful).
Diagnosis.– Bolivinoides ornamented with pore moundsand circular, more rarely elliptical pores.
Description.– Test biserial throughout, consisting of 13 to15 chambers that increase gradually in size (Pl. 1, Figs 1,3, 6, 8, 10). Chambers are wider than high, reniform andoblique to the test growth axis; chambers of the adult stagepresent three to four slender backward extensions, whichare longer in the central portion of the test, shorter towardsthe peripheral region and separated by wide subrectangu-lar intervals size (Pl. 1, Figs 1, 3, 6, 8-10). Chamber back-ward extensions are less distinct over the earlier chambersand, those of the same row of chambers in the adult stagemay be occasionally aligned. Sutures are distinct, de-pressed and oblique to the test growth axis and often ob-scured by the successive chamber backward extensions.Test compressed in edge view, with quasi-parallel sides;periphery broadly rounded and without peripheral struc-tures (Pl. 1, Figs 2, 7). Aperture has elongate shape and isprovided with a hook-like toothplate, which is occasion-ally preserved (Pl. 1, Figs 2, 4); an oblique supplementarysuture connects the aperture with the suture between thelast-formed and penultimate chambers (Pl. 1, Figs 2, 4, 7).Chamber surface is ornamented with incipient to well-de-veloped circular pore mounds (Øcpm = 3.9-5.6 µm), whichcan be scattered or, more rarely, clustered in small groupsof two or three (Pl. 1, Figs 1-10); pores without poremounds occur only on the last-formed pair of chambers.Test wall is calcitic, hyaline and perforate; pores are circu-lar (Øcp = 1.1-1.7 µm) or elliptical (Lep x Wep = 1.7-1.9µm x 0.5-0.9 µm; Wep/Lep = 0.259-0.542).
Remarks.– Bolivinoides mirabilis differs from any speciesof the genus by the ornamentation consisting of poremounds. There are remarkable similarities between B.mirabilis and the upper Turonian planktic foraminiferalspecies Steineckia steinecki Georgescu 2009a with re-
spect to the size of the pore mounds (3.9-5.6 µm whencompared to 4.0-6.4 µm). However, the two species dif-fer in other morphological features of paramount taxo-nomical significance: chambers with slender backwardextensions, aperture with a toothplate, a supplementarysuture connecting the aperture with the suture betweenthe last-formed and penultimate chambers, variable poreshape (circular to elliptical), larger pores in the former andglobular to reniform chambers without backward exten-sions, simple aperture at the base of the last formed cham-ber and without toothplate, pores completely circular inshape and smaller pores (0.4-0.8 µm) in the latter. An in-teresting morphological feature that cannot be properlyobserved with the aid of the optical microscope is theoblique supplementary suture, which connects the aper-ture and the suture between the last-formed two cham-bers. This structure is reported herein for the first time in aspecies of Bolivinoides (Fig. 5). The only known mecha-nism of formation of this structure is through the “wrap-ping” of the wall with respect to an axis approximatelyparallel to the test grow axis (Georgescu, 2009b). The datasuggest that the development of the oblique supplemen-tary suture must be regarded an iterative process from yetunknown ancestors, most likely Bolivina-like (Fig. 5), tothe Bolivinoides type of test, characterized by develop-ment of backward extensions.
Stratigraphic range.– Upper Campanian.
Geographic distribution.– USA (New Jersey).
Bolivinoides sp., ex. gr. B. decorata (Jones in Wright,1886)
Plate 1, Figs 11-12, Plate 2, Figs 1-2
Material.– Three specimens.
Dimensions.– Species of Bolivinoides with irregular ele-vated areas and elongate pores.
Description.– Test biserial throughout consisting of 17to18 chambers that increase gradually in size (Pl. 1, Fig11). Chambers are wider than high, those of the adultstage reniform and oblique to the test growth axis; cham-bers of the adult stage present four backward extensions,which are longer towards the central part of the test andshorter towards the peripheral region; the backward ex-

117
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
tensions of the successive chambers in the row can bealigned (Pl. 1, Fig 11). Sutures are distinct and depressed,oblique to the test growth axis. The chambers increasegradually in thickness; periphery is broadly rounded andwithout peripheral structures (Pl. 1, Fig. 12). Aperture,with a toothplate, is an elongate opening on the aperturalface of the last-formed chamber: no supplementary sutureoccurs on the apertural face of the last-formed chamber(Pl. 1, Fig. 12, Pl. 2, Fig. 1). Chamber surface is orna-mented with irregular elevated areas, which may occa-sionally resemble incipient pore mounds (Pl. 2, Fig. 2).Test wall is calcitic, hyaline and perforate; pores are ellip-tical (Lep x Wep = 1.9-4.2 µm x 0.4-1.0 µm; Wep/Lep =0.214-0.242) and do not present preferential orientation.
Remarks.– The type material of Bolivinoides decorata wasreviewed by Barr (1966, p. 231-234), who selected a lec-totype for it (ibidem, pl. 36, fig. 1). Unfortunately this re-
view was before the beginnings of the extensive SEM usein the study of fossil and recent foraminiferal tests and theemended description does not contain high resolution in-formation on the test ultrastructure and porosity. There-fore, it is impossible to assign the rare and oftenfragmentary specimens of the Marshalltown Formation toB. decorata with precision. Re-examination of the B. dec-orata type specimens is necessary in order to make directobservations on the pore size and distribution pattern, aswell as the possible occurrence of irregular elevated areason test surface.
Stratigraphic range.– Upper Campanian.
Geographic distribution.– USA (New Jersey).
Genus Elongateporeia – new
Type species: Elongateporeia elongatoporosa – new.
Figure 5. Diagram presenting the iterative pattern in the development of the oblique supplementary suture. The stratigraphic intervals with taxa pre-senting this feature are marked with light grey. Absolute ages are after Gradstein et al. (2004).

118
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
Etymology. – The name results from the combination ofthe two English words “elongate” and “pore”, to whichthe Latin suffix “-ia” is added.
Diagnosis.– Bolivinoitidid genus with elongate, slit-likepores, which can be situated in the central part of an el-liptical pore mound.
Description.– Test is biserial throughout, with rhomboidal-rounded to elliptical outline. Chambers wider than high,oblique to the test growth axis and provided with one ormore backward extensions. Test is compressed in edgeview with periphery rounded and without peripheralstructures. Aperture is an elongate opening situated on theapertural face of the last-formed chamber and providedwith a toothplate. Chambers are smooth or ornamentedwith elongate pore mounds. Test wall is calcitic, hyalineand perforate; pores are elongate to slit-like.
Species included.– Elongateporeia culverensis (Barr, 1967)and E. elongatoporosa – new species.
Remarks.– Elongateporeia differs from Bolivinoides byhaving elongate pores, which are variably present on anelongate pore mound. In general, Bolivinoides presentscircular to subpolygonal pores and B. mirabilis and B. sp.,ex. gr. B. decorata are the only Bolivinoides species withelongate pores.
Stratigraphic range.– Upper Santonian to upper Campan-ian.
Geographic distribution.– Europe (UK, Germany, Roma-nia), USA (New Jersey) and Australia.
Elongateporeia culverensis (Barr, 1967)Plate 2, Figs 3-13, Plate 3, Figs 1-3
1954 Bolivinoides decorata cf. delicatula Cushman –Edgell, pl. 13, fig. 7, pl. 14, fig. 7.
1963 Bolivinoides sp. – Hiltermann, p. 209, pl. 1, figs2-3, 7-9.
1966 Bolivinoides hiltermanni – Barr, p. 229, pl. 36, figs7-8, pl. 37, figs 1-3.
1967 Bolivinoides culverensis – Barr, p. 136.1977b Bolivinoides culverensis Barr – Petters, p. 1031, pl.
1, figs 10-12.1987 Bolivinoides culverensis Barr – Neagu, p. 75, pl.
7, figs 1-11, pl. 8, figs 9-18.
1989 Bolivinoides culverensis Barr – Hart et al., p. 322,pl. 7.4, figs 10-11.
Material.– Five hundred specimens.
Diagnosis.– Elongateporeia with elliptical, elongate to slit-like, rarely circular pores, which are occasionally situatedin the centre of an incipient elongate pore mound.
Description.– Test is biserial throughout, consisting of 13to 17 chambers that increase gradually in size. Chambersare wider than high and oblique to the test growth axis;chambers of the adult stage present two to three wide andclosely-spaced, nearly adjacent backward extensions,which cover the previous chamber and suture (Pl. 2, Figs3-5, 7, 9, 12-13, Pl. 3, Fig. 2). The backward extensionsover the earlier portion of the test are covered by the suc-cessive layers of calcite, resulting in circular tubercle-likestructures (Dmax = 6.7-27.6 µm) (Pl. 2, Figs 3-5, 7, 9, 12-13, Pl. 3, Fig. 2). Sutures are distinct, depressed andoblique to the test growth axis, often obscured by thebackward extensions of the successive chambers. Testcompressed in edge view, with quasi-parallel sides; pe-riphery rounded to subangular, without peripheral struc-tures (Pl. 2, Figs 6, 10, Pl. 3, Fig. 1). Aperture is elongate,situated on the apertural face of the last-formed chamber(Pl. 2, Figs 6, 10, Pl. 3, Fig. 1) and is provided with a tooth-plate (Pl. 2, Fig. 6). Chamber surface is ornamented withscattered incipient pore mounds, which are elliptical inshape (Pl. 2, Figs 8, 11, Pl. 3, Fig. 3) (Liepm x Wiepm = 3.2-5.9 µm x 1.6-3.3 µm; Wiepm/Liepm = 0.462-0.750) and canbe occasionally clustered in groups of two or, more rarely,three. Test wall is calcitic, hyaline and perforate; pores ex-hibit wide variability with respect to shape, orientationand distribution pattern. Pore shape is variable: circular(Øcp = 1.0-1.9 µm), elliptical (Lep x Wep = 1.5-2.2 µm x0.3-0.6 µm; Wep/Lep = 0.156-0.357) and slit-like (Lslp x
Plate 1. Specimens of Bolivinoides mirabilis – new species (1-10) and B.sp., ex.gr. B. decorata (Jones in Wright, 1886) (11-12); all specimens arefrom the upper Campanian Marshalltown Formation of the New Jerseycoastal plain (USA). 1-5, holotype of B. mirabilis (ODP Leg 174AX, Sam-ple 435.85-435.86 cm, G. obliqua Biozone); 4, detail micrograph show-ing the hook-like toothplate (2); 5, detail micrograph on the penultimatechamber as seen in edge view showing the circular pore mounds and cir-cular and elliptical pores (2). 6-9, paratype of B. mirabilis (ODP Leg174AX, Sample 435.85-435.86 cm, G. obliqua Biozone); 9, detail mi-crograph showing the simple pores and pore mounds on the penulti-mate chamber as seen in lateral view (6).10, Paratype of B. mirabilis.11-12, hypotype of B. sp., ex.gr. B. decorata (ODP Leg 174AX, Sample435.57-435.58 cm, G. obliqua Biozone).

119
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Plate 1

120
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
Wslp = 1.8-2.8 µm x 0.07-0.20 µm; Wslp/Lslp = 0.023-0.107); circular pores occur mostly on the earlier portionof the test; there is a distinct trend of the elliptical and slit-like pores to be preferentially aligned parallel to the testgrowth axis (Pl. 2, Fig. 8, Pl. 3, Fig. 3).
Remarks.– Elongateporeia culverensis differs from Bolivi-noides mirabilis by having a more rounded outline, thin-ner tests, apertural face without supplementary suture,incipient pore mound ornamentation rather than fully de-veloped pore mounds and high variability in pore shape,which range from circular to slit-like, rather than circularto elliptical as in the latter. The general Bolivinoides-likeappearance of E. culverensis is probably due to the Bo-livinoides ancestor (Petters, 1977b, p. 1032). However,additional data on the test ultrastructure of B. strigillatusChapman, 1892, its presumed ancestor, are necessary toconfirm or reject this direct phylogenetic relationship.
Stratigraphic range.– Upper Santonian to lower Campan-ian.
Geographic distribution.– Europe (UK, Germany, Roma-nia), USA (New Jersey) and Australia.
Elongateporeia elongatoporosa – new speciesPlate 3, Figs 4-14
Holotype.– WKB 010116.
Dimensions of the holotype.– L=0.322 mm; W=0.145mm; W/L=0.450; T=0.062 mm; T/L=0.193 (Fig. 4A).
Paratypes.– Five specimens, WKB 010117-010121.
Dimensions.– L=0.233=0.322 mm; W=0.113-0.145 mm;W/L=0.430-0.508; T=0.052-0.065; T/L=0.193-0.250 (Fig.4A). Ranges are based on the average measurements ofthe holotype and paratypes.
Material.– Twenty-five specimens.
Type locality.– ODP Leg 174AX (Bass River Site); geo-graphical coordinates: 39o 36’ 42”N and 74o 26’ 12” W.
Type level.– The green silts of the upper Campanian Mar-shalltown Formation; sample depth at the type locality:435.56-435.57 cm.
Etymology. – The name derives from the combination ofthe English words “elongate” and “pore”; the suffix “–osa”is added to it.
Diagnosis.– Elongateporeia ornamented with well-devel-oped elongate pore mounds.
Description.– Test biserial throughout consisting 15-19chambers that increase gradually in size; outline rhom-boidal-rounded to almost elliptical (Pl. 3, Figs 4, 6, 8, 11,14). Chambers are wider than high, those in the adultstage subtriangular-rounded to reniform in shape andoblique to the test growth axis; the chambers of the lastone or two pairs present one wide and short backward ex-tension (Pl. 3, Figs 4, 6, 11, 14). Sutures are flush toslightly depressed, often indistinct between the earlierchambers. Test is compressed in edge view, with quasi-parallel sides; periphery is rounded to subangular andwithout peripheral structures (Pl. 3, Figs 3, 7, 10, 13).Aperture is an elongate to nearly circular opening, situ-ated on the apertural face of the last-formed chamber (Pl.3, Figs 7, 10, 13) and is provided with a thin toothplate,which is rarely preserved (Pl. 3, Fig. 7); no supplementarysuture occurs on the apertural face of the last-formedchamber. Chamber surface is ornamented with scatteredwell-developed elliptical pore mounds (Lepm x Wepm =5.3-9.7 µm x 2.3-3.4 µm; Wepm/Lepm = 0.278-0.529),which are preferentially aligned parallel to the test growthaxis (Pl. 3, Figs 4-11, 13-14); occasionally they can begrouped into cluster of two or, more rarely three. Test wallis calcitic, hyaline and perforate; pores are straight or un-
Plate 2. Specimens of Bolivinoides sp., ex.gr. B. decorata (Jones inWright, 1886) (1-2) and Elongateporeia culverensis (Barr, 1967) (3-13);the illustrated specimens are from the upper Campanian (MarshalltownFormation) and upper Santonian (Merchantville Formation) respectivelyof the New Jersey coastal plain (USA). 1, detail micrograph of the edgeview of B. sp., ex.gr. B. decorata, illustrated in Pl. 1, Fig. 12 showing theaperture with toothplate and ornamentation consisting of elevated irreg-ular areas (ODP Leg 174AX, Sample 435.57-435.58 cm, G. obliqua Bio-zone). 2, detail micrograph of the same specimen showing theornamentation consisting of elevated irregular areas and elliptical poreswithout preferential ornamentation. 3-4, hypotype of E. culverensis(ODP Leg 174AX, Sample 505.05-505.08 cm, G. arca Biozone). 5-8,hypotype of E. culverensis (ODP Leg 174AX, Sample 505.05-505.08 cm,G. arca Biozone). 8, detail micrograph of the lateral view (5) showing theincipient pore mounds and elongate pores with preferential longitudinalorientation. 9-11, hypotype of E. culverensis (ODP Leg 174AX, Sample535.35-505.08 cm, G. arca Biozone). 11, detail micrograph of the lat-eral view (9) showing the incipient pore mounds and elongate pores withpreferential longitudinal orientation. 12-13, hypotype of E. culverensis(ODP Leg 174AX, Sample 505.35-505.38 cm, G. arca Biozone).

121
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Plate 2

122
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
dulated slit-like openings (Lslp x Wslp = 3.4-5.9 µm x 0.11-0.29 µm; Wslp/Lslp = 0.002-0.008).
Remarks.– Elongateporeia elongatoporosa differs from E.culverensis by having one rather than three backward ex-tensions on the last pairs of chambers, backward exten-sions restricted to the last one or two pairs of chambers,absence of tubercle-like structures on the earlier portion ofthe test, well-developed pore mound ornamentationrather than incipient elongate pore mounds and slit-likepores rather than a mixture of circular, elliptical and slit-like pores. Elliptical pore mounds ornamentation with slit-like pores makes E. elongatoporosa unique among thebenthic and planktic foraminifera with biserial chamberarrangement and probably the entire foraminiferal group.
Stratigraphic range.– Upper Campanian.
Geographic distribution.– USA (New Jersey).
Superfamily LOXOSTOMACEA Loeblich & Tappan,1962
Family LOXOSTOMATIDAE Loeblich & Tappan, 1962
Genus Trachelinella Montanaro Gallitelli, 1956 –emended
Type species: Bolivina watersi Cushman, 1927a (originaldesignation).
1955 Trakelina Montanaro Gallitelli, p. 215 (nomennudum – without description).
1956 Trachelinella Montanaro Gallitelli, p. 38.1957 Trachelinella Montanaro Gallitelli, 1956 – Mon-
tanaro Gallitelli, p. 150.1964 Trachelinella Montanaro Gallitelli, 1956 – Loe-
blich & Tappan, p. C736.1987 Trachelinella Montanaro Gallitelli, 1956 – Loe-
blich & Tappan, p. 501.
Emended description.– Test is biserial throughout, elon-gate and often twisted around the growth axis. The subrec-tangular chambers increase gradually in size. Eachchamber presents three short backward extensions, one atthe periphery resulting in a fringed outline and two sym-metrically arranged on the two sides of the test, thereforeonly one is visible in lateral view. Sutures are depressedand arched towards the distal part of the test. Test is com-
pressed in edge view, with subangular to angular periph-ery and without peripheral structures. Aperture is termi-nal, elliptical in shape, produced on a short neck,surrounded by an imperforate rim and without a tooth-plate. Chambers are ornamented with circular poremounds, which are concentrated towards the posteriorportion of the chambers forming elevated ornamentedplatforms. Test wall is calcitic, hyaline and perforate; poresare circular or elliptical.
Species included.– Trachelinella watersi (Cushman,1927a).
Remarks.– Trachelinella is emended to accommodate thenew pore mounds ornamentation observations. Fusedpustules ornamentation, which are concentrated and oftenfused towards the posterior portion of the chambers form-ing ornamented platforms, is unique among theforaminiferal group. The origin of Trachelinella is un-known and additional data are needed to clarify the testultrastructure of the bolivinoids with elongate and/ortwisted tests.
Stratigraphic range.– Campanian to Maastrichtian.
Geographic distribution.– USA (Texas, Mississippi, NewYork) and Israel.
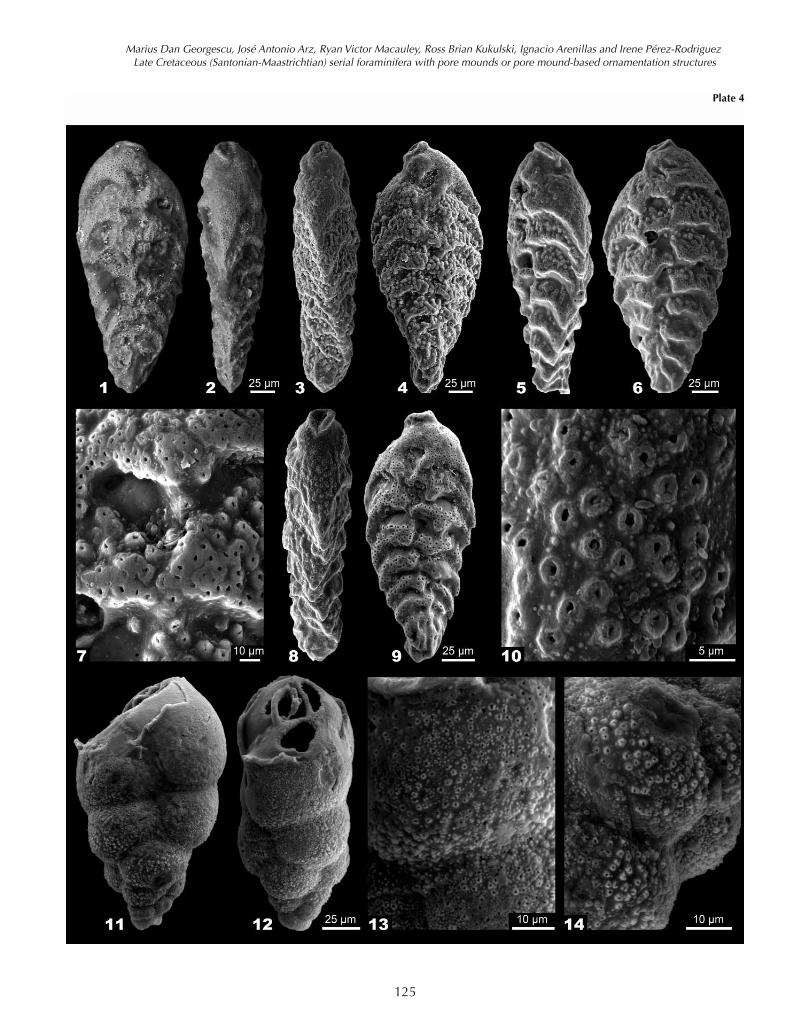
Trachelinella watersi (Cushman, 1927a) – emendedPlate 4, Figs 1-10
1927a Bolivina watersi – Cushman, p. 88, pl. 12, fig. 6.1946 Bolivina watersi Cushman – Cushman, p. 128,
pl. 53, fig. 18.
Plate 3. Specimens of Elongateporeia culverensis (1-3) and E. elongato-porosa – new species (4-14); all the specimens are from the upper Cam-panian Marshalltown Formation of the New Jersey coastal plain (USA).1-3, hypotype of E. culverensis (ODP Leg 174AX, Sample 505.35-505.38cm, G. arca Biozone). 3, detail micrograph of the lateral view (2) show-ing the elongate pores with preferential longitudinal orientation and tu-bercle-like structures. 4-5, holotype of E. elongatoporosa (ODP Leg174AX, Sample 435.56-435.57 m, R. calcarata Biozone). 6-9, paratypeof E. elongatoporosa (ODP Leg 174AX, Sample 435.56-435.57 m, R.calcarata Biozone). 9, detail micrograph of the lateral view (6) showingthe well-developed elongate pore mounds and slit-like pores with pref-erential longitudinal ornamentation. 10-12, paratype of E. elongatopo-rosa (ODP Leg 174AX, Sample 435.56-435.57 m, R. calcarata Biozone).12, detail micrograph of the lateral view (10) showing the aperture withbroken toothplate. 13-14, topotype of E. elongatoporosa (ODP Leg174AX, Sample 435.57-435.58 m, G. obliqua Biozone).

123
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Plate 3

124
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
1954 Bolivinoides watersi (Cushman) – Reiss, p. 156,pl. 29, fig. 8.
1956 Trachelinella watersi (Cushman) – MontanaroGallitelli, 1956, pl. 7, figs 8-10.
1957 Trachelinella watersi (Cushman) – MontanaroGallitelli, 1957, pl. 34, fig. 13.
1965 Trachelinella watersi (Cushman) – Perlmutter &Todd, p. I 15, pl. 2, fig. 12.
1992 Bolivina praecrenulata – Gawor-Biedowa, p. 96,pl. 17, fig. 6.
Material.– Sixty specimens.
Emended description.– Test is biserial throughout, oftentwisted around the growth axis (Pl. 4, Figs 3, 5, 8); the out-line is rhomboidal-rounded, nearly elliptical, with themaximum width in the anterior part. The small proloculusis followed by 17 to 21 chambers that increase graduallyin size; chambers are subrectangular with three backwardextensions: one on each side and a third shorter one atthe periphery, resulting in a fringed outline in the adultportion of the test (Pl. 4, Figs 4, 6, 9). Sutures are de-pressed, curved towards the anterior part and often cov-ered over the earlier chambers by the ornamentationstructures and successive layers of calcite (Pl. 4, Figs 1, 4,6, 9). Test compressed in edge view, with subangular toangular periphery and without peripheral structures (Pl.4, Figs 2, 3, 5, 8). Aperture is terminal in position and el-liptical in shape; it is produced on a short neck and sur-rounded by a rounded, imperforate rim (Pl. 4, Figs 1-6,8-9). Ornamentation consists of well-developed circularpore mounds (Øcpm = 2.9-9.7 µm) ((Pl. 4, Figs 1-10),which are scattered or concentrated towards the posteriorpart of the chambers; pore mounds are often fused to formelevated ornamented platforms (Pl. 4, Fig. 7). Test wall iscalcitic, hyaline and perforate; pores are circular (Øcp =1.2-2.0 µm) or elliptical (Lep x Wep = 2.7-2.9 µm x 0.6-1.0µm; Wep/Lep = 0.210-0.500), those over the earliest pairsof chambers are completely obscured by the successivelayers of calcite added during the ontogeny.
Remarks.– Trachelinella watersi exhibits significant vari-ability in the test twisting around the growth axis (0-90o)and pore shape, which ranges from circular to elliptical.
Stratigraphic range.– Campanian to Maastrichtian.
Geographic distribution.– USA (Texas, Mississippi, NewYork), Europe (Poland) and Israel.
Superfamily TURRILINACEA Cushman, 1927bFamily TURRILINIDAE Cushman, 1927b
Genus Neobulimina Cushman & Wickenden, 1928 –emended
Type species: Neobulimina canadensis Cushman & Wick-enden, 1928 (original designation).
1928 Neobulimina Cushman & Wickenden, p. 12.1964 Neobulimina Cushman & Wickenden, 1928 – Loe-
blich & Tappan, p. C544.1987 Neobulimina Cushman & Wickenden, 1928 – Loe-
blich & Tappan, p. 511.
Emended description.– The test consists of subglobular toglobular chambers that overlap at various rates. Chambersare triserially arranged in the earlier stage; the adult stageconsists of chambers with biserial arrangement. Suturesare distinct and depressed, straight to slightly curved.Aperture is loop shaped, situated at the base of the last-formed chamber or up the apertural face and providedwith a toothplate. Chambers are smooth or ornamentedwith pustules and incipient to well-developed poremounds, which are concentrated over the earlier portionof the test. Test wall is calcitic, hyaline and perforate.
Plate 4. Specimens of Trachelinella watersi (Cushman, 1927a) (1-10) andNeobulimina newjerseyensis – new (11-14). 1-2, holotype of T. watersi,USNM 6119, originally figured by Cushman (1927a, pl. 12, fig. 6). 3-4,hypotype of T. watersi from the Maastrichtian Corsicana Formation ofTexas (Well Mullinax-3, Sample 42, 4-3, 7.15 m, P. hariaensis Biozoneequivalent). 5-7, hypotype of T. watersi from the Maastrichtian Corsi-cana Formation of Texas (Well Mullinax-1, Sample 365, 25-1, 41.07 m,A. mayaroensis Biozone equivalent). 7, detail micrograph of the lateralview (6) showing the fused pore mounds forming ornamented platformsin the posterior portion of the chamber. 8-10, hypotype of T. watersi fromthe Maastrichtian Corsicana Formation of Texas (Well Mullinax-1, Sam-ple 365, 25-1, 41.07 m, A. mayaroensis Biozone equivalent). 10, detailmicrograph of the lateral view (8) showing the scattered pore moundsover the test penultimate chamber. 11-14, holotype of N. newjerseyen-sis from the upper Campanian Marshalltown Formation of the New Jer-sey coastal plain (USA) (ODP Leg 174AX, Sample 435.57-435.58 m, G.obliqua Biozone). 13, detail micrograph of the edge view (12) showingthe circular pore mounds formed through the pustule concentrationaround the circular pores. 14, detail micrograph on the earlier cham-bers of the edge view (12) showing frequent well-developed and small-sized pore mounds.
125
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Plate 4

126
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
Remarks.– Neobulimina is emended to include pustuleand pore mound ornamentation as well as the aperture,which is provided with a toothplate. It differs from Praebu-limina Hofker, 1953 by having two growth stages, whichare apparent in chamber arrangement (triserial in the ju-venile and biserial in the adult) rather than having triser-ial throughout tests.
Stratigraphic range.– Albian to Maastrichtian.
Geographic distribution.– Cosmopolitan.
Neobulimina newjerseyensis – new speciesPlate 4, Figs 11-14, Plate 5, Figs 1-5
Holotype.– WKB 010122.
Dimensions of the holotype.– L=0.201 mm; W=0.105mm; W/L=0.522; T=0.080 mm; T/L=0.398 (Fig. 4B).
Paratypes.– WKB 010123-010127.
Dimensions.– L=0.168-0.203 mm; W=0.091-0.107 mm;W/L=0.517-0.542; T=0.073-0.082 mm; T/L=0.374-0.408(Fig. 4B).
Material.– Twenty-four specimens.
Type locality.– ODP Leg 174AX (Bass River Site); geo-graphical coordinates: 39o 36’ 42”N and 74o 26’ 12” W.
Type level.– The green silts of the upper Campanian Mar-shalltown Formation; sample depth at the type locality:435.57-435.58 cm.
Etymology. – The name is after the State of New Jersey(USA), where the type locality is situated.
Diagnosis.– Neobulimina species ornamented with poremounds.
Description.– Test consists of two growth stages with dif-ferent chamber arrangement. Early stage consists of pro-loculus followed by 8 to 10 chambers with triserialarrangement; adult stage consists of 3 to 4 biseriallyarranged chambers (Pl. 4, Figs 11-12, Pl. 5, Figs 1-3). Thesubglobular to globular chambers increase gradually insize and overlap at various rates. Sutures are distinct, de-pressed, straight to slightly curved. Aperture is loop-
shaped, situated on the apertural face of the last-formedchamber and is provided with a toothplate (Pl. 4, Fig. 12,Pl. 5, Figs 3, 5). Chambers are ornamented with smallscattered pustules (Øp = 0.5-1.5 µm) and incipient to well-developed circular pore mounds (Øcpm = 0.8-1.9 µm) (Pl.4, Figs 13-14, Pl. 5, Figs 4-5); pore mounds are formedthrough the concentration of the pustules around the cir-cular pores (Pl. 4, Fig. 13, Pl. 5, Fig. 4). Ornamentation isconcentrated over the earlier portion of the test and is de-veloped only in the adult individuals; juvenile tests aresmooth or with rare scattered pustules over the earlierchambers (Pl. 5, Fig. 1). Test wall is calcitic, hyaline andperforate; pores are circular (Øcp = 0.3-0.7 µm).
Remarks.– Neobulimina newjerseyensis differs from allthe other species included in the genus mainly by its cir-cular pore mounds formed through pustule concentrationaround the pores; fused pustules result in a solid, well-de-veloped appearance of the pore mounds. The general testappearance of N. newjerseyensis presents remarkablesimilarities to N. subregularis de Klasz, Magné and Rérat,1963 from the Campanian of Gabon and Brazil; the lowerstratigraphic level of the first evolutionary occurrence(upper Santonian) together with the absence of ornamen-tation suggest that N. subregularis is the probable ances-tor of N. newjerseyensis.
Stratigraphic range.– Upper Campanian.
Geographic distribution.– USA (New Jersey).
Genus Pseudouvigerina Cushman, 1927
Type species: Uvigerina cristata Marsson, 1878 (originaldesignation).
1927d Pseudouvigerina Cushman, p. 81.1964 Pseudouvigerina Cushman, 1927 – Loeblich &
Tappan, p. C568.1987 Pseudouvigerina Cushman, 1927 – Loeblich &
Tappan, p. 511.
Diagnosis.– Turrilinids with ornamentation consisting ofpore mounds.
Description.– Test pyramidal to subpyramidal, with trise-rially arranged chambers. Earlier species with broad andlow chambers; chambers of the adult stage in the evolved

127
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
species are high and reniform. Periphery is broadlyrounded and simple in the primitive species and with dou-ble truncation and two keels consisting of aligned andfused pore mounds in the evolved one. Aperture is situ-ated at the base of the last-formed chamber and is pro-vided with a hook-like toothplate in the primitive species;the evolved species has wrapped toothplate, as a result ofthe wall wrapping parallel to the test growth axis. Cham-bers are ornamented with pore mounds with are rare andincipiently developed in the early species and well-devel-oped in the evolved one. Test wall is calcitic, hyaline andperforate; circular to elliptical pores occur in the primi-tive species and the evolved one has only circular pores.
Remarks.– Pseudouvigerina in its emended form includesonly two species: P. rudita (Cushman & Parker, 1936) andP. cristata (Marsson, 1878), grouping the two species in alineage, based on the similarities in the early stage mor-phology and ornamentation. Pseudouvigerina hereinemended accommodates a directional lineage(Georgescu, 2010) in which there is compelling evidencethat all of the morphological features evolve. The changesin two features with paramount taxonomical significance,the apertural position, from basal to terminal and tooth-plate shape from hook-like to wrapped, suggest remark-able changes in the evolutionary path of this lineage.Pseudouvigerina differs from any other lineage of turrilin-ids mainly by developing pore mound ornamentation.Keels composed of aligned and occasionally fused poremounds, are developed in the evolved species of the lin-eage (i.e.., P. cristata) and are unique among the Creta-ceous foraminifera with serial chamber arrangement.
Stratigraphic range.– Campanian to Maastrichtian.
Geographic distribution.– USA (Alabama, Arkansas, Cal-ifornia, Louisiana, Mississippi, New Jersey, Tennessee,Texas), Mexico, Europe (Germany, Holland, Poland, Swe-den, UK), Caribbean region (Trinidad), South America(Brazil) and North Atlantic Ocean (Orphan Knoll).
Pseudouvigerina rudita (Cushman & Parker, 1936) –emended
Plate 5, Figs 6-11
1935 Bulimina ornata – Cushman & Parker (non Egger1895), p. 97, pl. 15, fig. 4.
1936 Bulimina rudita Cushman & Parker – Cushman &Parker, p. 45.
1946 Bulimina rudita Cushman & Parker – Cushman, p.122, pl. 51, fig. 24.
1947 Bulimina rudita Cushman & Parker – Cushman &Parker, p. 82, pl. 19, fig. 26.
1957 Reussella uvigeriniformis – Hofker, p. 217, text-figs265-267.
1957 Reussella rugosa (Brotzen) – Hofker, p. 218, text-figs 268-269.
1968 Pyramidina rudita (Cushman & Parker) – Sliter, p.86, pl. 12, fig. 12.
1992 Pyramidina rudita (Cushman & Parker) – Gawor-Biedowa, p. 122, pl. 23, fig. 6.
2000 Pyramidina rudita (Cushman & Parker) – Kout-soukos & de Klasz, p. 400, figs 6.26-6.27.
Material.– Forty-five specimens.
Diagnosis.– Pseudouvigerina with aperture at the base ofthe last formed chamber and ornamentation consisting ofincipient pore mounds.
Emended description.– Test pyramidal with slightly de-pressed sides, triserial throughout consisting of 13 to 16chambers that increase gradually in size; chambers arebroad and low, those of the adult stage reniform. Suturesare distinct and depressed, straight to curved. Periphery istripartite, broadly rounded and without peripheral struc-tures. Aperture is elongate, at the base of the last-formedchamber and provided with a hook-like toothplate. Cham-bers are ornamented with incipient circular pore mounds(Øicpm = 2.6-5.5 µm), which are more frequent on the ear-lier chambers and often exhibit the pustules from whoseconcentration they apparently formed. Test wall is calcitic,hyaline and perforate; pores are circular (Øcp = 0.6-1.8µm) or elliptical (Lep x Wep = 0.7-1.9 µm x 0.3-0.9 µm;Wep/Lep = 0.214-0.600), simple or situated in the centralpart of an incipient pore mound.
Remarks.– Pseudouvigerina rudita exhibits wide variabil-ity in pore shape, from elliptical to circular; strongly elon-gate, slit-like pores were not observed in this species tests.Pore mound ornamentation was originally de-scribed/proposed by Hofker (1957) and the first SEM illus-

128
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
trations of the pore mounds were given by Gawor-Biedowa (1992, pl. 23, fig 6a-b). However, the Polishspecimen exhibit well-developed pore mounds, whichcontrasts to the specimens included in this study that areonly in incipient stage of development.
Stratigraphic range.– Campanian to Maastrichtian.
Geographic distribution.– USA (Alabama, California, Mis-sissippi, Texas), Mexico, South America (Brazil) and Eu-rope (Germany, Holland, Poland).
Pseudouvigerina cristata (Marsson, 1878) – emendedPlate 5, Fig. 12, Plate 6, Figs 1-11
1878 Uvigerina cristata – Marsson, p. 150, pl. 3, fig. 20.1925 Uvigerina seligi – Cushman, p. 1, pl. 4, fig. 1. 1931 Uvigerina seligi Cushman – Plummer, p. 186, pl.
14, fig. 10.1927e Pseudouvigerina plummerae – Cushman, p. 115,
pl. 23, fig. 8.1932 Pseudouvigerina (?) plummerae Cushman – San-
didge, p. 197, pl. 19, figs 9-11.1936 Uvigerina seligi Cushman – Jennings, p. 190, pl.
3, fig. 24.1943 Pseudouvigerina seligi (Cushman) – Cushman &
Todd, p. 65, pl. 11, fig. 19.1945 Pseudouvigerina cristata (Marsson) – Brotzen, p.
46, figs 8a-b.1946 Pseudouvigerina plummerae Cushman – Cus-
hman, p. 116, pl. 49, figs 14-16.1957 Reussella cristata (Marsson) – Hofker, p. 220, text-
figs 272-274.1957 Pseudouvigerina cristata (Marsson) – Montanaro
Gallitelli, pl. 34, figs 19-22.1960 Pseudouvigerina seligi (Cushman) – Olsson, p. 30,
pl. 4, fig. 23.1965 Pseudouvigerina seligi (Cushman) – Perlmutter &
Todd, p. I15, pl. 2, fig. 19.1968 Pseudouvigerina plummerae Cushman – Sliter, p.
91, pl. 13, fig. 11.1984 Pseudouvigerina seligi (Cushman) – Olsson &
Nyong, pl. 3, fig. 6.1989 Pseudouvigerina cristata (Marsson) – Hart et al., p.
358, pl. 7.22, figs 1-2.1992 Pseudouvigerina cristata (Marsson) – Gawor-Bie-
dowa, p. 117, pl. 24, figs 6-7.
1994 Pseudouvigerina plummerae Cushman – Bolli etal., p. 133, fig. 35.34.
2000 Pseudouvigerina plummerae Cushman – Koutsou-kos & de Klasz, p. 400, figs 6.23-6.25.
Material.– One hundred fifty specimens.
Diagnosis.– Pseudouvigerina with terminal aperture andornamentation characterized by well-developed poremounds, which can be scattered or aligned at the periph-ery to form pore mound-based keels.
Emended description.– Test is subpyramidal, triserialthroughout and consisting of 8 to 10 chambers that in-crease gradually in size; chambers are broad and low inthe early stage and high and reniform in the adult. Suturesare distinct, depressed and curved. Periphery tripartite,with double truncation on each chamber and lined withkeels consisting of aligned and often fused pore mounds.Aperture is terminal, circular to elliptical and producedon a short neck; the wrapped toothplate is a continuationof the test wall and its shape reflects the wall wrappingparallel to the test growth axis. Chamber surface is or-namenented with scattered well-developed circular poremounds (Øcpm = 4.0-6.4 µm), which are more frequent onthe last-formed chambers. Test wall is calcitic, hyaline andperforate; pores circular (Øcp = 1.0-2.0 µm), each of themsituated in the centre of a pore mound.
Remarks.– Pseudouvigerina cristata differs from its pre-sumed ancestor, P. rudita, mainly by the terminal aperturerather than at the base of the last formed chamber, periph-
Plate 5. Specimens of Neobulimina newjerseyensis – new species (1-5),Pseudouvigerina rudita (Cushman & Parker, 1936) (6-11) and P. cristata(Marsson, 1878) (12). 1, paratype of N. newjerseyensis from the upperCampanian Marshalltown Formation of New Jersey (ODP Leg 174AX,Sample 435.57-435.58 m, G. obliqua Biozone). 2-5, paratype of N. new-jerseyensis (ODP Leg 174AX, Sample 435.57-435.58 m, G. obliqua Bio-zone). 4, detail micrograph of the penultimate chamber as seen in edgeview (3) showing the ornamentation consisting of circular pore mounds.5, detail micrograph of the edge view (3) showing the toothplate bor-dered by an imperforate rim. 6-8, hypotype of P. rudita from the Maas-trichtian Corsicana Formation of Texas (Well Mullinax-1, Sample 379,26-1, 42.47 m, A. mayaroensis Biozone equivalent). 8, detail micrographof the edge view (7) showing the circular and elliptical pores. 9-11, hy-potype of P. rudita from the Maastrichtian Corsicana Formation of Texas(Well Mullinax-1, Sample 379, 26-1, 42.47 m, A. mayaroensis Biozoneequivalent). 11, detail micrograph of the edge view (10) showing the in-cipient circular pore mounds. 12, holotype of P. plummerae, USNM(6700), originally figured by Cushman (1927e, pl. 23, fig. 8); this speciesis considered a junior synonym of P. cristata.

129
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Plate 5

130
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
ery with double truncation and two keels consisting ofaligned and often fused pore mounds rather than broadlyrounded and without peripheral structures, ornamentationconsisting of well-developed pore mounds on all thechambers rather than incipient pore mounds concentratedon the earlier chambers and larger pores (1.0-2.0 µmrather than 0.6-1.8 µm), which are circular rather than cir-cular to elliptical.
Stratigraphic range.– Upper Campanian-Maastrichtian.
Geographic distribution.– USA (Alabama, Arkansas, Cal-ifornia, Louisiana, Mississippi, New Jersey, Tennessee,Texas), Mexico, Europe (Germany, Holland, Poland, Swe-den, UK), Caribbean region (Trinidad), South America(Brazil) and North Atlantic Ocean (Orphan Knoll).
Superfamily UNCERTAINFamily GUEMBELITRIIDAE Montanaro Gallitelli, 1957
Genus Guembelitria Cushman, 1933
Type species: Guembelitria cretacea Cushman, 1933(original designation).
1933 Guembelitria Cushman, p. 37.1957 Guembelitria Cushman, 1933 – Montanaro Gal-
litelli, p. 136.1964 Guembelitria Cushman, 1933 – Loeblich & Tap-
pan, p. C652.1979 Guembelitria Cushman, 1933 – Blow, p. 1382.1987 Guembelitria Cushman, 1933 – Loeblich & Tap-
pan, p. 453.1990 Guembelitria Cushman, 1933 – Kroon & Neder-
bragt, p. 33.1999 Guembelitria Cushman, 1933 – Olsson et al., p.
79.2007 Guembelitria Cushman, 1933 – Arenillas et al., p.
34.2009b Guembelitria Cushman, 1933 – Georgescu, p.
130.2010 Guembelitria Cushman, 1933 – Arz et al., p. 159.
Description.– Test is triserial throughout, consisting of sub-globular to globular chambers that increase gradually insize and overlap at various rates. Sutures are straight toslightly curved, distinct and depressed. Aperture is a largesubcircular opening at the base of the last formed cham-
ber; it is bordered by a narrow imperforate rim and pres-ents a small, relict toothplate. Chamber surface is orna-mented with pore mounds and scattered dome-likepustules. Test wall is calcitic, hyaline and perforate.
Remarks.– The origin of Guembelitria is reconsideredbased on the new data on the tests with pore mounds ofthe upper Santonian-Maastrichtian stratigraphic interval.Georgescu (2009b, p. 132) considered that G. cretaceaevolved through the complication of the pustulose orna-mentation in G. turrita Kroon & Nederbragt, 1990 fromthe upper Campanian-Maastrichtian. However, the pus-tules in G. turrita are triangular and do not tend to groupinto pore mounds. The description of small pustules andincipient circular pore mounds in the new benthic speciesNeobulimina newjerseyensis represents a major challengein deciphering the origin of Guembelitria. The evolution ofGuembelitria from a species of Neobulimina would besimilar in many respects (e.g., disappearance of the adultbiserial stage, development of ornamentation consistingof pore mounds, reduction of toothplate, etc) to that of thelate Albian from the benthic Praeplanctonia Georgescu,2009b, as ancestor, to Archaeoguembelitria Georgescu,2009b, its descendant. However, the occurrence of clearG. cretacea specimens in the Santonian sediments re-quires additional studies to more adequately define thestratigraphical distribution of N. newjerseyensis, Guem-belitria presumed ancestor. Biometric measurements andmorphostatistical analysis of specimens from El Kef(Tunisia) allowed Arz et al., 2010 to recognize threeGuembelitria species in the upper Maastrichtian: G.blowi, restricted to short-spired morphotypes (Plate 7, Figs7-9), G. cretacea, to medium-spired morphotypes (Plate
Plate 6. Specimens of Pseudouvigerina cristata (Marsson, 1878) (1-11)and Guembelitria dammula Voloshina, 1961 (12-13). 1-4, hypotype ofP. cristata from the upper Campanian Ripley Formation (Braggs, Lown-des County, Alabama, USA, ? R. calcarata Biozone equivalent). 2, detailmicrograph showing the terminal aperture. 3, detail micrograph of thelateral view (1) showing the well-developed pore mounds. 5-7, hypo-type of P. cristata from the lower Maastrichtian Prairie Bluff Chalk(Braggs, Lowndes County, Alabama, USA; ? G. gansseri Biozone equiv-alent). 7, detail micrograph showing the wrapped toothplate, which re-flects the wall wrapping parallel to the test growth axis. 8-9, hypotype ofP. cristata from the lower Maastrichtian Prairie Bluff Chalk (Braggs, Lown-des County, Alabama, USA; ? G. gansseri Biozone equivalent). 10-11,hypotype of P. cristata from the upper Maastrichtian sediments of Or-phan Knoll (North Atlantic Ocean) (DSDP Leg 12, Hole 111A, Sample11-2, 50-52 cm, A. mayaroensis Biozone). 12-13, hypotype of G.dammula from the Maastrichtian Corsicana Formation of Texas (WellMullinax-3, Sample 42, 4-3, 7.15 m, P. hariaensis Biozone equivalent).

131
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Plate 6

132
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
7, Figs 1-6) and G. dammula, to high-spired morphotypes(Plate 6, Figs 12-13). Almost all specimens of these speciespresent typical pore-mounded wall, although other micro-textural variants can occur (Arz et al., 2010). Both G.blowi and G. dammula are descendents of G. cretaceaand probably evolved during the uppermost Campanianor Maastrichtian. For these reasons, we must focus on an-alyzing the wall textural characteristics of G. cretacea andidentifying its first appearance in order to hypothesizeabout the Guembelitria origin.
Stratigraphic range.– Santonian to Paleocene (lower Dan-ian).
Geographic distribution.– Cosmopolitan.
Guembelitria cretacea Cushman, 1933Plate 7, Figs 1-6
1933 Guembelitria cretacea – Cushman, p. 37, pl. 4, fig.12.
1946 Guembelitria cretacea Cushman – Cushman, p.103, pl. 44, fig. 4.
1960 Guembelitria cretacea Cushman – Olsson, p. 27,pl. 4, fig. 8.
1967 Guembelitria cretacea Cushman – Pessagno, p.258, pl. 87, figs 1-3.
1970 Guembelitria cretacea Cushman – Olsson, p. 601,pl. 91, figs 4-5.
1973 Guembelitria cretacea Cushman – Smith & Pes-sagno, p. 15, pl. 1, figs 1-8.
1980 Guembelitria cretacea Cushman – Peryt, p. 33, pl.1, figs 7-8.
1990 Guembelitria cretacea Cushman – Huber, p. 502,pl. 1, fig. 1.
1990 Guembelitria cretacea Cushman – Kroon & Neder-bragt, p. 35, pl. 2, fig. 4 (only).
1991 Guembelitria cretacea Cushman – D’Hondt, p.172, pl. 1, figs 1-6, pl. 2, figs 2-3.
1991 Guembelitria cretacea Cushman – D’Hondt & Ke-ller, p. 93, pl. 3, fig. 1.
1992 Guembelitria cretacea Cushman – Liu & Olsson,p. 341, pl. 1, figs 1-2.
1993 Guembelitria cretacea Cushman – MacLeod, pl. 1,figs 8-10, p[l. 2, figs 1-4, 7-8.
1999 Guembelitria cretacea Cushman – Olsson et al., p.79, pl. 8, figs 1-3, pl. 13, fig. 3, pl. 63, figs 1-12.
2003 Guembelitria cretacea Cushman – Abramovich etal., pl. 1, fig. 1.
2007 Guembelitria cretacea Cushman – Arenillas et al.,p. 37, figs 13.2-13.3.
2009b Guembelitria cretacea Cushman – Georgescu, p.132, fig. 4.
2010 Guembelitria cretacea Cushman – Arz et al., p.161, figs 8.4-8.10.
Material.– Two hundred specimens.
Description.– Test moderately high, triserial throughoutconsisting of 9 to 12 globular to subglobular chambersthat increase moderately in size. Sutures are distinct anddepressed, straight to curved. Periphery is tripartite,broadly rounded and without peripheral structures. Aper-ture is high and rounded, subcircular and situated at thebase of the last formed chamber; it is bordered by a thinimperforate lip and occasionally presents a short, relicttoothplate. Chambers are ornamented with well-devel-oped circular pore mounds (Øcpm = 3.4-6.4 µm), whichcan be absent on the last formed one or two chambers,and scattered dome-like pustules (Øp = 0.9-1.5 µm). Testwall is calcitic, hyaline and perforate; pores are circular(Øcp = 0.4-0.9 µm) and situated in the central part of apore mound.
Remarks.– The emendation of G. cretacea by Arz et al.(2010) is accepted and followed herein. Archaeoguembe-litria harrisi is homeomorphous with G. cretacea and themorphological convergence between the two species isthe result of iterative evolution (Georgescu, 2009b). Therare scattered dome-like pustules in the ornamentation ofG. cretacea resemble those in N. newjerseyensis, suggest-ing a possible phylogenetic relationship between the twospecies; see the ‘Remarks’ section under genus Guembe-
Plate 7. Specimens of Guembelitria cretacea Cushman, 1931 and G.blowi Arz et al., 2010. 1, topotype of G. cretacea from the Loeblich &Tappan Topotype Collection (USNM 473122). 2, hypotype of G. cre-tacea from the lower Paleocene (G. cretacea Biozone, Parvularugoglo-bigerina longiapertura Subzone) of the El Kef section (Tunisia), SampleKF13.5; specimen previously figured by Arz et al. (2010, fig. 8.5). 3, hy-potype of G. cretacea from the Santonian sediments of Assila section(Kalaat Senan region, central Tunisia), Sample AS07. 4-6, hypotype of G.cretacea from the Santonian sediments of Assila section (Kalaat Senan re-gion, central Tunisia), Sample AS07. 7-9, holotype of G. blowi from theupper Maastrichtian (A. mayaroensis Biozone) sediments of the El Kefsection (Tunisia), Sample KF4.50; specimen previously figured by Arz etal. (2010, figs 9.1-9.3).

133
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
Plate 7

134
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
litria. A new early occurrence of G. cretacea is reportedherein from the Santonian sediments of the Assila sectionin the Kalaat Senan region (central Tunisia).
Stratigraphic range.– Santonian to Paleocene (lower Dan-ian).
Geographic distribution.– Cosmopolitan.
5. CONCLUSIONS AND DISCUSSION
High resolution observations made on Late Cretaceous(late Santonian-Maastrichtian) benthic foraminifera withbiserial and/or triserial chamber arrangement from theUnited States and North Atlantic Ocean reveal that a highdiversity of the submicroscopic morphological structuresare present. Pore mounds are morphologically diverse,with two types identified by their shape: circular and el-liptical. These ornamentation structures show two devel-opmental stages: incipient and well-developed. Afour-fold classification for pore mounds is developedherein by considering their shape and degree of develop-ment: incipient circular, well-developed circular, incipi-ent elliptical and well-developed elliptical. Three newornamentation structures are also recognized for the firsttime: irregular elevated areas, ornamented platforms andpore mound-based keels. Pore shape and degree of devel-opment are important features that can be useful for deci-phering the evolutionary history of certain lineages ofplanktic and benthic foraminifera. The taxa ornamentedwith scattered pore mounds and/or pore mound-based or-namentation structures analyzed in this study show threetypes of pores: circular, elliptical and slit-like.
Scattered, well-developed circular pore mounds are re-ported for the first time in a species of the genus Bolivi-noides, namely B. mirabilis – new species, which wasrecovered from the upper Campanian Marshalltown For-mation of the New Jersey coastal plain. Bolivinoidesmirabilis presents an interesting diagnostic morphologicalfeature, an oblique supplementary suture on the aperturalface, which connects the aperture with the suture betweenthe last-formed and penultimate chamber. The occurrenceof such a primitive feature in a late Campanian species(approximately 11 m.y. after the first occurrence of a Bo-
livinoides species in the fossil record-late Santonian) andthe absence of species with similar structures in the upperSantonian-upper Campanian stratigraphical interval mayimply a polyphyletic nature to the genus Bolivinoides.
A numerically minor component of the foraminiferal as-semblages in the upper Campanian Marshalltown Forma-tion (New Jersey coastal plain) is B. sp., ex. gr. B. decorata.This species is ornamented with irregular elevated areas,which occasionally resemble incipient pore mounds. Thepores are elliptical and show no preferential orientationparallel to the test growth axis. This taxon can be easilymisidentified under the optical microscope for B. deco-rata or B. decorata australis Edgell, 1954. Although left inopen nomenclature, this species shows the necessity of amonograph on the genus Bolivinoides, in order to refinethe genus and component species definitions with respectto new observations on wall ultrastructure, ornamentationand porosity.
A new genus, Elongateporeia, is described for two species,E. culverensis and E. elongatoporosa – new species. Elon-gateporeia is characterized by the occurrence of slit-likepores situated in the central part of an elongate poremound. The stratigraphic range of the new genus is upperSantonian-upper Campanian, with the ranges of the twospecies separated by a gap corresponding to theGlobotruncana ventricosa Biozone (middle Campanian inage according to Robaszynski & Caron, 1995). Elon-gateporeia culverensis of the upper Santonian-lower Cam-panian is ornamented with incipient elliptical poremounds and exhibits highly variable pore shape: circular,elliptical and slit-like. The circular pores occur primarilyon the earlier portion of the test and a gradual transitionbetween the elliptical and slit-like pores is observable inthe adult stage of the development. Well-developed ellip-tical pore mounds surround slit-like pores in the upperCampanian E. elongatoporosa. Morphological observa-tions suggest that E. culverensis may represent the ances-tor to E. elongatoporosa; however, further study is requiredto fill the gap between the two species. Elongateporeiaevolved from a Bolivinoides ancestral species [probablyB. strigillatus (Chapman, 1892)] as suggested by the cham-bers backward extensions in the adult stage.
Pore mounds are also reported in the biserial genus Tra-chelinella of the Campanian-Maastrichtian. Its description

135
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
is emended to accommodate the new observations on testornamentation. Pore mounds are fused towards the poste-rior portion of the chambers and form ornamented plat-forms, which are described for the first time in this study.
The discovery of the first Neobulimina with pore moundornamentation is of paramount importance in further un-derstanding the polyphyletic nature of the plankticforaminifera. Neobulimina newjerseyensis – new speciesis reported from the upper Campanian Marshalltown For-mation of the New Jersey coastal plain. Its ornamentationconsists of a combination of small scattered pustules andpore mounds that show a wide variability with respect oftheir degree of development, from incipient to well-de-veloped. Diminutive individuals of this species are orna-mented with only scattered pustules. This ornamentationdescribed in the benthic N. newjerseyensis resembles thatof the triserial planktic Guembelitria cretacea, which sug-
gests a phylogenetic relationship between them as ances-tor and descendant respectively. A similar evolution fromthe triserial-biserial benthics to triserial planktics wasdemonstrated in the late Albian-early Turonian lineagePraeplanctonia-Archaeoguembelitria (Georgescu, 2009b).A similar relationship was suggested using molecular ev-idence from the modern triserial planktic Gallitellia Loe-blich & Tappan, 1986 by Ujiié et al. (2008). However, thefirst occurrence of G. cretacea is documented in the San-tonian and precedes that of N. newjerseyensis, which is inthe upper Campanian (Fig. 6).
The ornamentation of the triserial benthic Pseudouvige-rina consists of circular pore mounds, which can be ei-ther incipient (i.e., P. rudita) or well-developed (i.e., P.cristata). Pseudouvigerina cristata is the evolved speciesin this genus and this is strongly supported by its elaborateornamentation: scattered well-developed pustules over
Figure 6. Diagram summarizing the iterative development of planktic habitat in the Cretaceous planktic foraminifera with serial chamber arrangement.Planktic taxa ranges are given in grey, those of the benthics in black.

136
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
the chamber surface and fused pustules forming poremound-based keels over the double-truncated tripartiteperiphery. The keels comprised of fused pore mounds arereported for the first time in the Cretaceous foraminiferawith triserial chamber arrangement.
The implications of the SEM/ESEM-based studies providesignificant insight into understanding the benthic andplanktic foraminiferal evolution and take the first steps to-wards evolutionary classification. Recognizing the ances-tor-descendant relationships among benthic foraminiferais an important and difficult task for a foraminiferologistinvolved in evolutionary classification. The plankticforaminiferal record shows that tests with intermediatemorphological features between two phylogenetically re-lated species are frequent. In contrast, the benthicforaminiferal record exhibits a different pattern, withspecies and genera often sharply defined in terms of mor-phology. Consequently, tests with intermediate morpho-logical features between two phylogenetically relatedspecies or genera are extremely rare and often absent. Ad-ditional research is required to elucidate the evolutionarylink between the less often studied benthics and the com-monly studied planktics. In this context, the study offoraminifera with pore mounds or pore mound-based or-namentation features provides new and novel researchobjectives in benthic foraminiferal studies.
ACKNOWLEDGMENTS
Drs N. Malumian and C. Náñez (Servicio GeológicoMinero Argentino) are thanked for the throuough reviewsthat substantially increased the manuscript quality. Dr I.Rábano (Editor, Revista Española de Micropaleontología)is thanked for the high-quality editorial process and ac-ceptance for publication. The authors thank Dr B.T. Huber(NMNH, Washington, D.C., USA) for the illustrations ofthe holotypes of Trachelinella watersi and Pseudouvige-rina plummerae. Dr C.M. Henderson (University of Cal-gary, Alberta, Canada) is thanked for granting full accessto the McGugan Collection (University of Calgary). Thegenerosity of Dr S. Abramovich (Ben Gurion University ofthe Negev, Israel) who allowed the study of the Mullinax-1 and Mullinax-3 wells of Texas (USA) is greatly appreci-ated. We warmly thank Dr L.V. Hills (University of
Calgary) for the pre-submittal critical review, which sig-nificantly improved the manuscript. Dr M. Schoel (Micro-scope Imaging Facility) is thanked for the enthusiastic andprofessional help during the SEM operations.DSDP/ODP/IODP headquarters are thanked for allowingus to resample the upper Maastrichtian section at DSDPHole 111A (Orphan Knoll, North Atlantic Ocean). This re-search was partly funded by the Spanish Ministerio deCiencia e Innovación project CGL2007-63724/BTE andthe Aragonian Departamento de Educación y Ciencia(DGA group E05).
REFERENCES
Abramovich, S., Keller, G., Stüben, D. and Berner, Z. 2003. Characteri-zation of late Campanian and Maastrichtian planktic foraminiferaldepth habitats and vital activities based on stable isotopes. Palaeoge-ography, Palaeoclimatology, Palaeoecology, 202, 1-29.
Arenillas, I., Arz, J.A. and Náñez, C. 2007. Morfología, biometría y ta-xonomía de foraminíferos planctónicos del Daniense basal. RevistaEspañola de Paleontología, 22, 21-62.
Arz, J.A., Arenillas, I. and Náñez. C. 2010. Morphostatistical analysis ofMaastrichtian populations of Guembelitria from El Kef, Tunisia. Jour-nal of Foraminiferal Research, 40, 148-164.
Barr, F.T. 1966. The foraminiferal genus Bolivinoides from the Upper Cre-taceous of the British Isles. Palaeontology, 9, 220-243.
Barr, F.T. 1967. Bolivinoides culverensis, new name for the Campanianforaminifer B. hiltermanni Barr. Contributions from the CushmanFoundation for Foraminiferal Research, 18, 136.
Beckmann, J.P. and Koch, W. 1964. Vergleiche von Bolivinoides, Arago-nia und Tappanina (Foraminifera) aus Trinidad (Westindien) und Mit-teleuropa. Geologisches Jahrbuch, 83, 31-64.
Blow, W.H. 1979. The Cainozoic Globigerinidae. A study of the mor-phology, taxonomy, evolutionary relationship and the stratigraphicaldistribution of some Globigerinidae (mainly Globigerinacea). Leiden,E.J. Brill, 1413 pp.
Bolli, H.M., Beckmann, J.P. and Saunders, J.B. 1994. Benthicforaminiferal biostratigraphy in the south Caribbean region. Cam-bridge, Cambridge University Press, 408 pp.
Brotzen, F. 1936. Foraminiferen aus dem schwedischen unterer Senonvon Eriksdal in Schonen. Sveriges Geologiska Untersökning ser. C,396(30), 1-69.
Brotzen, F. 1945. De geologiska resultanten fra° n borrningarna vid Höl-lviken; Preliminär rapport. Del 1. Kritan. Sveriges Geologiska Unter-sökning, ser. C, 465, 1-64.
Chapman, F. 1892. Microzoa from the Phosphatic Chalk of Taplow.Quarterly Journal of the Geological Society of London, 48, 514-518.
Cushman, J.A. 1925. A new Cretaceous Uvigerina from Louisiana. Con-

137
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
tributions from the Cushman Laboratory for Foraminiferal Research,1, 1.
Cushman, J.A. 1927a. American Upper Cretaceous species of Bolivinaand related species. Contributions from the Cushman Laboratory forForaminiferal Research, 2, 85-91.
Cushman, J.A. 1927b. An outline of a re-classification of theforaminifera. Contributions from the Cushman Laboratory forForaminiferal Research, 3, 1-105.
Cushman, J.A. 1927c. Some characteristic Mexican fossil foraminifera.Journal of Paleontology, 1, 147-172.
Cushman, J.A. 1927d. Some new genera of the foraminifera. Contribu-tions from the Cushman Laboratory for Foraminiferal Research, 3, 77-81.
Cushman, J.A. 1927e. New and interesting foraminifera from Mexicoand Texas. Contributions from the Cushman Laboratory forForaminiferal Research, 3, 111-119.
Cushman, J.A. 1933. Some new foraminiferal genera. Contributions fromthe Cushman Laboratory for Foraminiferal Research, 9, 32-38.
Cushman, J.A. 1946. Upper Cretaceous Foraminifera of the Gulf Coastalregion of the United States and adjacent areas. United States Geolog-ical Survey Professional Paper, 206, 1-241.
Cushman, J.A. and Parker, F.L. 1935. Some American Cretaceous bulim-inas. Contributions from the Cushman Laboratory for ForaminiferalResearch, 11, 96-101.
Cushman, J.A. and Parker, F.L. 1936. Some American Eocene buliminas.Contributions from the Cushman Laboratory for Foraminiferal Re-search, 12, 39-45.
Cushman, J.A. and Parker, F.L. 1947. Bulimina and related foraminiferalgenera. United States Geological Survey Professional Paper, 210-D,55-176.
Cushman, J.A. and Renz, H.H. 1946. The foraminiferal fauna of theLizard Springs Formation of Trinidad, B.W.I. Contributions from theCushman Laboratory for Foraminiferal Research, Special Publication,18, 1-48.
Cushman, J.A. and Todd, R. 1943. Foraminifera of the Corsicana Marl.Contributions from the Cushman Laboratory for Foraminiferal Re-search, 19, 49-71.
Cushman, J.A. and Wickenden, R.T.D. 1928. A new foraminiferal genusfrom the Upper Cretaceous. Contributions from the Cushman Labo-ratory for Foraminiferal Research, 4, 12-13.
Delage, Y. and Hérouard, E. 1896. Traité de zoologie concrete. Tome I:La cellule et les protozoaires. Paris, Schleicher Frères, 584 pp.
D’Hondt, S. 1991. Phylogenetic and stratigraphic analysis of Earliest Pa-leocene biserial and triserial planktonic foraminifera. Journal ofForaminiferal Research, 21, 170-183.
D’Hondt, S. and Keller, G. 1991. Some patterns of planktic foraminiferalassemblage turnover at the Cretaceous-Tertiary boundary. Marine Mi-cropaleontology, 17, 77-118.
D’Orbigny, A. d’ 1839. Voyage dans l’Amérique méridionale – Forami-nifères, v. 5, pt. 5. Paris et Strasbourg, P. Bertrand, 86 pp.
Edgell, H.S. 1954. The stratigraphical value of Bolivinoides in the UpperCretaceous of Northwest Australia. Contributions from the CushmanFoundation for Foraminiferal Research, 5, 68-76.
Eichwald, C.E. von 1830. Zoologia specialis II. Vilnae, D.E. Eichwaldus,323 pp.
Finlay, H.J. 1939. New Zealand Foraminifera: Key species in stratigra-phy – No. 3. Transactions of the Royal Society of New Zealand, 69,309-329.
Gawor-Biedowa, E. 1992. Campanian and Maastrichtian foraminiferafrom the Lublin Upland, eastern Poland. Palaeontologia Polonica, 52,1-187.
Georgescu, M.D. 2006. Santonian-Campanian planktonic foraminiferafrom the New Jersey coastal plain and their distribution related to therelative sea-level changes. Canadian Journal of Earth Sciences, 43,101-120.
Georgescu, M.D. 2009a. Taxonomic revision and evolutionary classifi-cation of the biserial Cretaceous planktic foraminiferal genus Laevi-heterohelix Nederbragt, 1991. Revista Mexicana de CienciasGeológicas, 26, 315-334.
Georgescu, M.D. 2009b. On the origins of Superfamily HeterohelicaceaCushman, 1927 and the polyphyletic nature of planktic foraminifera.Revista Española de Micropaleontología, 41, 107-144.
Georgescu, M.D. 2010. Origin, taxonomic revision and evolutionaryclassification of the late Coniacian-early Campanian (Late Creta-ceous) planktic foraminifera with multichamber growth in the adultstage. Revista Española de Micropaleontología, 42, 59-118.
Georgescu, M.D and Abramovich, S. 2008. Taxonomic revision and phy-logenetic classification of the Late Cretaceous (Upper SantonianMaastrichtian) serial planktonic foraminifera (Family HeterohelicidaeCushman, 1927) with peripheral test wall flexure. Revista Españolade Micropaleontología, 40, 97-114.
Georgescu, M.D. and Huber, B.T. 2009. Early evolution of the Creta-ceous serial planktic foraminifera (late Albian-Cenomanian). Journalof Foraminiferal Research, 39, 335-360.
Glaessner, M.F. 1937. Die Entfaltung der Foraminiferenfamilie Bulim-inidae. Paleontologii, Paleontologicheskaya Laboratoriya Moskovs-kogo Gosudarstvennogo Universiteta, 2-3, 411-422.
Gradstein, F.M., Ogg, J.G., Smith, A.G., Agterberg, F.P., Bleeker, W.,Cooper, R.A., Davydov, V., Gibbard, P., Hinnov, L.A., House, M.R.,Lourens, L., Luterbacher, H.P., McArthur, J., Melchin, M.J., Robb, L.J.,Shergpold, J., Villeneuve, M., Wardlaw, B.R., Ali, J., Brinkhuis, H.,Hilgen, F.J., Hooker, J., Hoewath, R.J., Knoll, A.H., Laskar., J., Mon-echi, S., Plumb, K.A., Powell, J., Raffi. I., Röhl, U., Sadler, P., Sanfil-ippo, A., Schmitz, B., Schackelton, N.J., Shields, G.A., Strauss, H.,Van Dam, J., van Kolfschoten, T. and Wilson, D. 2004. A GeologicTime Scale 2004. Cambridge University Press, 1-589.
Hart, M.B., Bailey, H.W., Crittenden, S., Fletcher, B.N., Price, R.J. andSwiecicki, A., 1989. Cretaceous. In: Stratigraphical Atlas of FossilForaminifera, Second Edition (Eds, Jenkins, D.G. and Murray, J.W.),Chichester, Ellis Horwood Limited, 273-371.
Hay, W.H., DeConto, R.M., Wold, C.N., Wilson, K.M., Voigt, S., Schulz,M., Wold, A.R., Dullo, W.C., Ronov, A.B., Balukhovsky, A.N. andSöding, E., 1999, Alternative global Cretaceous paleogeography. In:Evolution of Cretaceous Ocean-Climate System (Eds, Barrera, E. and

Johnson, C.C.). Boulder, Colorado, The Geological Society of Amer-ica, Special Publication, 332, 1-47.
Hiltermann, H. 1963. Zur Entwicklung der Benthos-Foraminifere Bolivi-noides. In: Evolutionary trends in foraminifera (Eds, Koenigswald,G.H.R. von, Emeis, J.D., Buning, W.L.and C.W. Wagner). ElsevierPublishing Company, Amsterdam–London–New York, 198-223.
Hiltermann, H. and Koch, W. 1950. Taxonomie und Vertikalverbreitungvon Bolivinoides-Arten im Senon Nordwestdeutschlands. Geologis-ches Jahrbuch, 64, 595-632.
Hofker, J. 1953. Types of genera described in Part III of the “SibogaForaminifera”. Micropaleontologist, 7, 26-28.
Hofker, J. 1957. Foraminiferen der Oberkreide von Nordwestdeutsch-land und Holland. Beihefte zum Geologischen Jahrbuch, 27, 1-464.
Huber, B.T. 1990. Maestrichtian planktonic foraminifer biostratigraphy ofthe Maud Rise (Weddell Sea, Antarctica): ODP Leg 113 Holes 689Band 690C. In: Initial Reports of the Deep Sea Drilling Project, v. 113(Eds. P. F. Baker, J. P. Kennett et al.), Washington, D. C., United StatesGovernment Printing Office, 113, 489-513.
Jennings, P.H. 1936. A Microfauna from the Monmouth and Basal Ran-cocas Group of New Jersey. Bulletins of American Paleontology,23(78), 161-220.
Keller, G. 1989. Extended Cretaceous/Tertiary boundary extinctions anddelayed population change in planktic foraminifera from BrazosRiver, Texas. Paleoceanography, 4, 287-332.
Keller, G. 2002. Guembelitria dominated late Maastrichtian plankticforaminiferal assemblages mimic early Danian in the Eastern Desertof Egypt. Marine Micropaleontology, 47, 71-99.
Keller, G., Abramovich, S., Berner, Z. and Adatte, T. 2009. Biotic effectson the Chicxulub impact, K-T catastrophe and sea-level change inTexas. Palaeogeography, Palaeoclimatology, Palaeoecology, 271, 52-68.
Keller, G., Adatte, T., Berner, Z., Harting, M., Baum, G., Prauss, M.,Tantawy, A.A. and Stüben, D. 2007. Chicxulub impact predates K-Tboundary: New evidence from Brazos, Texas. Earth and PlanetaryScience Letters, 255, 339-356.
Keller, G. and Pardo, A. 2004. Disaster opportunists Guembelitriidae –Index for environmental catastrophes. Marine Micropaleontology,53, 83-116.
Klasz, I. de, Magné, J. and Rérat, D. 1963. Quelques formes nouvellesde Buliminidae caractéristique du Crétacé superieur du Gabon (Afri-que Equatoriale). Revue de Micropaléontologie, 6, 145-152.
Kroon, D. and Nederbragt, A.J. 1990. Ecology and paleoecology of tri-serial planktic foraminifera. Marine Micropaleontology, 16, 25-38.
Koutsoukos, E.A.M. and Klasz, I. de 2000. Late Cretaceous foraminiferalbiogeography (Families Bolivinidae, Buliminidae, Gavelinellidae,Siphogenerinoididae, Turrilinidae) in northeastern Brazilian shelf andcentral West African basins. Cretaceous Research, 21, 381-405.
Liu, C. and Olsson, R.K. 1992. Evolutionary adaptive radiation of mi-croperforate planktic foraminifera following the K/T mass extinctionevent. Journal of Foraminiferal Research, 22, 328-346.
Loeblich, A.R. Jr. and Tappan, H. 1962. Six new generic names in the
Mycetozoida (Trichiidae) and Foraminiferida (Fischerinidae, Bulim-inidae, Caucasinidae and Pleurostomellidae), and a redescription ofLoxostomum (Loxostomidae, new family). Proceedings of the Biolog-ical Society of Washington, 75, 107-113.
Loeblich, A.R. Jr. and Tappan, H. 1964. Sarcodina Chiefly “Thecamoe-bians” and Foraminifera. In: Treatise on Invertebrate Paleontology(Ed., R.C. Moore), The University of Kansas Press and The GeologicalSociety of America, Boulder, Part C, C1-C900.
Loeblich, A.R. Jr. and Tappan, H. 1984. Suprageneric classification ofthe Foraminiferida (Protozoa). Micropaleontology, 30, 1-70.
Loeblich, A.R. Jr. and Tappan, H. 1986. Some new and revised generaand families of hyaline calcareous Foraminiferida (Protozoa). Trans-actions of the American Microscopical Society, 105, 239-265.
Loeblich, A.R. Jr. and Tappan, H. 1987. Foraminiferal Genera and TheirClassification. New York, Van Nostrand Reinhold Company, 970 pp.
MacLeod, N. 1993. The Maastrichtian-Danian radiation of triserial andbiserial planktic foraminifera: testing phylogenetic and adaptationalhypotheses in the (micro)fossil record. Marine Micropaleontology,21, 47-100.
Mancini, E.A. and Puckett, T.M. 2005. Jurassic and Cretaceous Trans-gressive-regressive (T-R) Cycles, Northern Gulf of Mexico, USA.Stratigraphy, 2, 31-48.
Mancini, E.A., Puckett, T.M. and Tew, B.H. 1996. Integrated biostrati-graphic and sequence stratigraphic framework for Upper Cretaceousstrata of the eastern Gulf Coastal Plain, USA. Cretaceous Research,17, 645-669.
Marie, P. 1941. Les foraminifères de la craie a Belemnitella mucronatadu Bassin de Paris. Mémoires du Muséum National d’Histoire Na-turelle, 12, 1-296.
Marsson, T. 1878. Die Foraminiferen der Weissen Schreiberkreide derInseln Rügen. Mitteilungen des Naturwissenschaftlichen Vereins fürNeu-Vorpommern und Rugen in Greifswald, 10, 115-196.
Miller, K.G., Sugarman, P.J., Browning, J.V., Olsson, R.K., Pekar, S.F.,Reilly, T.J., Cramer, B.S., Aubry, M.P., Lawrence, R.P., Curran, J., Stew-art, M., Metzger, J.M., Uptegrove, J., Bukry, D., Burckle, L.H., Wright,J.D., Feigenson, M.D., Brenner, G.J. and Dalton, R.F. 1998. Bass RiverSite. In: Proceedings of the Ocean Drilling Program, Vol. 174AX Sup-plement (Eds, Miller, K.G., Sugarman, P.J. and Browning, J.V.), OceanDrilling Program, National Science Foundation and Joint Oceano-graphic Institutions Publications, 1–39.
Montanaro Gallitelli, E. 1955. Una revisione della famiglia Heteroheli-cidae Cushman. Atti e Memorie Acaademia di Scienze, Lettere edArti, Modena, 13, 213-223.
Montanaro Gallitelli, E. 1956. Bronnimannella, Tappanina, and Trache-linella, three new foraminiferal genera from the Upper Cretaceous.Contributions from the Cushman Foundation for Foraminiferal Re-search, 7, 35-39.
Montanaro Gallitelli, E. 1957. A revision of the Foraminiferal Family Het-erohelicidae. In: Studies in Foraminifera (Ed., Loeblich, A.R.Jr.),United States National Museum Bulletin, 215, 133-154.
Neagu, T. 1987. White Chalk foraminiferal fauna from southern Dobro-gea (Romania). III. Calcareous benthic foraminifera. Revista Españolade Micropaleontología, 24, 59-115.
Revista Española de Micropaleontología / v. 43 / nº 1-2 / 2011
138

Nederbragt, A.J. 1991. Late Cretaceous biostratigraphy and developmentof Heterohelicidae (planktic foraminifera). Micropaleontology, 37,329-372.
Olsson, R.K. 1960. Foraminifera of latest Cretaceous and earliest Tertiaryage in the New Jersey coastal plain. Journal of Paleontology, 34, 1-58.
Olsson, R.K. 1970. Planktic Foraminifera from Base of Tertiary, MillersFerry, Alabama. Journal of Paleontology, 44, 598-604.
Olsson, R.K., Hemleben, C., Berggren, W.A. and Huber, B.T. 1999. Atlasof Paleocene planktonic foraminifera. Smithsonian Contributions toPaleobiology, 85, 1-252.
Olsson, R.K. and Nyong, E.E. 1984. A paleoslope model for Campanian-lower Maestrichtian foraminifera of New Jersey and Delaware. Jour-nal of Foraminiferal Research, 14, 50-68.
Perlmutter, N.M. and Todd, R. 1965. Correlation and Foraminifera of theMonmouth Group (Upper Cretaceous) Long Island, New York. UnitedStates Geological Survey Professional Paper, 483-I, I1-I24.
Peryt, D. 1980. Planktic foraminifera zonation of the Upper Cretaceousin the Middle Vistula River valley, Poland. Palaeontologia Polonica,41, 3-101.
Pessagno, E.A.Jr. 1967. Upper Cretaceous planktic foraminifera from thewestern Gulf Coastal Plain. Palaeontographica Americana, 5(37),245-445.
Petters, S.W. 1976. Upper Cretaceous subsurface stratigraphy in the At-lantic Coastal Plain of New Jersey. The American Association of Pe-troleum Geologists Bulletin, 60, 87-108.
Petters, S.W. 1977a. Upper Cretaceous planktonic foraminifera from thesubsurface of the Atlantic Coastal Plain of New Jersey. Journal ofForaminiferal Research, 7, 165-187.
Petters, S.W. 1977b. Bolivinoides evolution and Upper Cretaceous bios-tratigraphy of the Atlantic Coastal Plain of New Jersey. Journal of Pa-leontology, 51, 1023-1036.
Plummer, H.J. 1931. Some Cretaceous foraminifera in Texas. Universityof Texas Bureau of Economic Geology and Technology Bulletin,3101, 109-203.
Puckett, T.M. 2005. Santonian-Maastrichtian planktonic foraminiferaland ostracode biostratigraphy of the northern Gulf Coastal Plain,USA. Stratigraphy, 2, 117-146.
Reiss, Z. 1954. Upper Cretaceous and Lower Tertiary Bolivinoides fromIsrael. Contributions from the Cushman Foundation for ForaminiferalResearch, 5, 154-164.
Robaszynski, F. and Caron, M. 1995. Foraminifères planctoniques duCrétacé: commentaire de la zonation Europe-Mèditerranée. Bulletinde la Société Géologique de France, 6, 681-692.
Sandidge, J.R. 1932. Significant foraminifera from the Ripley Formationof Alabama. American Midland Naturalist, 13, 190-202.
Scott, J.C., Newton, J.C., Copeland, C.W., Drahovzal, J.A., Monroe,W.H., Cepek, P., Hay, W.W., Masters, B.A. and Worsley, T. W. 1968.Facies changes in the Selma Group in central and eastern Alabama.A guidebook for the Sixth Annual Field Trip of the Alabama Geolog-ical Society, December 6-7, 1968. Alabama Geological Society, Uni-versity of Alabama, 1–69.
Sliter, W.V. 1968. Upper Cretaceous foraminifera from southern Califor-nia and northwestern Baja California, Mexico. The University ofKansas Paleontological Contributions, 49, 1-141.
Smith, C.C. and Pessagno, E.A. Jr. 1973. Planktic foraminifera and stratig-raphy of the Corsicana Formation (Maestrichtian) north-central Texas.Cushman Foundation for Foraminiferal Research, Special Publica-tion, 12, 1-68.
Tappan, H. 1940. Foraminifera from the Grayson Formation of northernTexas. Journal of Paleontology, 14, 93-126.
Ujiié, Y., Kimoto, K. and Pawlowski, J. 2008. Molecular evidence for anindependent origin of modern triserial planktonic foraminifera frombenthic ancestors. Marine Micropaleontology, 69, 334-340.
Voloshina, A.M. 1961. Some new species of Upper Cretaceousforaminifera from the Volhyn-Podol platform. PaleontologicheskiySbornik, 1, 71-81.
White, M.P. 1929. Some index foraminifera of the Tampico EmbaymentArea of Mexico. Part III. Journal of Paleontology, 3, 30-58.
Wright, J. 1886. A list of the Cretaceous Foraminifera of Keady Hill,County Derry. Proceedings of the Belfast Naturalists’ Field Club, 2,327-332.
MANUSCRITO RECIBIDO: 23 de junio, 2010MANUSCRITO ACEPTADO: 6 de septiembre, 2010
Marius Dan Georgescu, José Antonio Arz, Ryan Victor Macauley, Ross Brian Kukulski, Ignacio Arenillas and Irene Pérez-RodriguezLate Cretaceous (Santonian-Maastrichtian) serial foraminifera with pore mounds or pore mound-based ornamentation structures
139
