l-lactate production from seaweed hydrolysate of laminaria japonica using metabolically engineered...
TRANSCRIPT

L-Lactate Production from Seaweed Hydrolysate of Laminariajaponica Using Metabolically Engineered Escherichia coli
Suman Mazumdar & Junho Bang & Min-Kyu Oh
Received: 30 June 2013 /Accepted: 22 November 2013 /Published online: 3 December 2013# Springer Science+Business Media New York 2013
Abstract Renewable and carbon neutral, marine algal biomass could be an attractivealternative substrate for the production of biofuel and various biorefinery products. Thus,the feasibility of brown seaweed (Laminaria japonica) hydrolysate as a carbon source wasinvestigated here for L-lactate production. This work reports the homofermentative route for
L-lactate production by introducing Streptococcus bovis/equinus L-lactate dehydrogenase inan engineered Escherichia coli strain where synthesis of the competing by-product wasblocked. The engineered strain utilized both glucose and mannitol present in the hydroly-sate under microaerobic condition and produced 37.7 g/L of high optical purity L-lactate at80 % of the maximum theoretical value. The result shown in this study implies that algalbiomass would be as competitive with lignocellulosic biomass in terms of lactic acidproduction and that brown seaweed can be used as a feedstock for the industrial productionof other chemicals.
Keywords L-Lactate . Mannitol . Seaweed .Metabolic engineering .Escherichia coli
Introduction
Marine environment comprises over half of the primary production of the global biomass [6].Biomass, being regarded as carbon neutral [42], is an important part of any future renewableenergy mix because it is non-intermittent, can be used to produce chemicals and liquidtransport fuels, and can sequester carbon in addition to providing renewable heat and power.In a densely populated country such as South Korea, the growing of terrestrial crops forbiofuel/chemical production is not feasible due to the limited land availability and high laborcost. Thus, marine biomass here can serve as an excellent alternative source to meet the presentand future fuel/chemical demands.
Marine-derived brown macroalgae/seaweed, Laminaria japonica, is being abundantly usedfor the production of alginate, apart from being a staple food. In 2006, the annual production ofmarine algae was 0.8 million tons in South Korea, corresponding to 10 % of agricultural by-products [32]. Brown seaweed contains up to 67 % carbohydrates by dry weight, which
Appl Biochem Biotechnol (2014) 172:1938–1952DOI 10.1007/s12010-013-0653-9
S. Mazumdar : J. Bang :M.<K. Oh (*)Department of Chemical and Biological Engineering, Korea University, Seoul 136-713, Republic of Koreae-mail: [email protected]

includes materials such as alginate, laminaran, mannitol, etc. [15, 23]. Brown seaweeddoes not contain lignin [31], which is advantageous for hydrolysis. Compared to landcrops, seaweed has a great variety of monomer sugars resulting from polysaccharidehydrolysis [32].
The conversion of biomass from marine algae into biofuel/biochemicals could be econom-ically more feasible than some terrestrial, lignocellulosic biomass feedstock [8, 17, 39]. Studiesto date have been largely focused on the production of biogas [12, 13, 27] and hydrogen [18,37] by anaerobic digestion, while some recent studies have also focused on the utilization ofthe sugars present in seaweed to produce bioethanol by fermentation [1, 11, 15, 24]. However,little research has been done to evaluate the feasibility of L. japonica as a substrate to producebiochemicals, such as L-lactate.
Lactate and its derivatives have many applications in the food, pharmaceutical, and polymerindustries [14, 30]. An example is polylactic acid, a renewable, biodegradable, andenvironment-friendly polymer produced from controlled ratios of the lactate enantiomer[21]. The importance of using pure enantiomers in such applications gives biological processesan advantage over chemical means by producing chirally pure lactate from inexpensive mediacontaining only the carbon source and mineral salts.
While lactic acid bacteria has been traditionally used in the production of L-lactate frompure sugars and carbohydrate-rich alternative substrate, the requirement for complex nutrientmedia and the lack of product selectivity in producing enantiomeric pure L-lactate [14, 30]limit their usage. However, studies have also reported Enterococcus [43, 44] and yeast [16, 40]as possible hosts for producing L-lactate. Recombinant Escherichia coli also possesses strongpotentials for L-lactate production due to its ability to utilize several inexpensive carbonsources, ease of genetic manipulations, and well-studied metabolic pathways; several studieshave reported it as an alternative biocatalysts engineered to produce L-lactate from sugarfeedstocks [7, 9, 46].
Unlike the usage of pure carbohydrates, we focused on the use of algal biomass as a carbonsource for the production of chemicals with high optical and chemical purity. As such, thismanuscript reports the metabolic engineering of E. coli for the efficient conversion of algalhydrolysate to L-lactate in a defined minimal salt medium, a process that has not been reportedyet. The engineered strain holds great promise as it provides a perspective for the conversion ofrenewable resources into higher-valued products such as L-lactate.
Materials and Methods
Source and Preparation of Algal Hydrolysate
Brown seaweed (L. japonica) was obtained from a market in Sanjeong-dong, Mokpo, SouthKorea. After washing five times with distilled water and drying in air, followed by oven dryingat 50 °C for 14 h, the dried seaweed was pulverized, screened through a 35 mesh, and stored inan airtight container at 4 °C until further use. The dried seaweed pellet (30 g) was resuspendedin 300 mL of 0.1 N HCl and heated at 121 °C for 20 min. The resulting lysate was pH-adjustedto 5.5 with citrate phosphate buffer and a mixture of three enzymes—Viscozyme®L,Celluclast® 1.5L, and AMG®300L (Sigma-Aldrich, St. Louis, MO)—was added at 1 % (w/w dried biomass). Enzymatic hydrolysis was carried out at 50 °C in an orbital shaker for 20 h.The feed for fed-batch fermentation was concentrated to one fourth volume by evaporatingwater under vacuum. All biomass hydrolysates were autoclaved after adjusting the pH to 7.2.Centrifugation at 5,000 rpm for 45 min at 4 °C separated the clear algal hydrolysate
Appl Biochem Biotechnol (2014) 172:1938–1952 1939

supernatant, which was transferred into a sterile bottle under aseptic condition and stored at4 °C until further use.
Strains, Plasmids, and Genetic Methods
E. coliMG1655, obtained from the Coli Genetic Stock Center (CGSC, Yale University, USA),was used as the host for L-lactate production. Gene knockouts in the E. coli genome wereaccomplished with P1 phage transduction from single-gene knockout mutants (Keio strains)obtained from CGSC as donor strains using a previously described protocol [45]. Multiplegene knockouts were performed sequentially to a common host by the same procedure. Allknockout mutations were confirmed at all disruption sites with PCR using the verificationprimers listed in Table 1. To construct plasmid pZAldh, the ldh gene (BankIt1600304 ldhKC514804) was PCR-amplified using genomic DNA from Streptococcus bovis/equinusKCTC 3959 and ldh primers (Table 1). The resulting PCR product was cloned within theKpnI and BamHI sites of pZA31MCS. The ligated products described above were used totransform E. coli DH5α (Invitrogen, Carlsbad, CA). Positive clones were screened by plasmidisolation and restriction digestion. Manufacturer protocols and standard methods in DNArecombinant technology [36] were used for DNA purification (Qiagen, Valencia, CA), restric-tion endonuclease digestion (NEB, Ipswich, MA), and DNA amplification using PhusionDNA polymerase (NEB). The developed strains were kept in 32.5 % glycerol stocks at−80 °C. All resulting strains, along with the primers and plasmids used in this study, are listedin Table 1.
Culture Medium and Cultivation Conditions
Unless otherwise stated, all fermentations were conducted using 1× MOPS minimalmedium designed by Neidhardt et al. [29], with Na2HPO4 in place of K2HPO4, 5 μMsodium selenite, 3.96 mM Na2HPO4, 5 mM (NH4)2SO4, and 30 mM NH4Cl. The mediawere supplemented with either 20 g/L mannitol or with algal hydrolysate. All thechemicals were obtained from Sigma-Aldrich Co., except for the algal hydrolysate whichwas produced in-house as previously described. Fermentations in shake flask wereperformed in a 100-mL Schott-Duran Erlenmeyer flask (Scott Inc., Elmsford, NY) filledwith 60 mL of 1× MOPS minimal medium supplemented with the appropriate antibioticsor inducer when needed at the following concentrations: chloramphenicol, 34 μg/mL;anhydrotetracycline, 100 ng/mL. Unless otherwise stated, calcium carbonate (5 %, w/v)was added in all the fermentation flasks to buffer the pH. The flasks sealed with cottonplugs were incubated at 37 °C and 200 rpm in an orbital shaker. The fermentations wererun for 48 h (unless otherwise stated) and the samples were collected to determine theoptical density and pH (Hanna Instrument Inc., Woonsocket, RI). To determine the opticaldensities of the cultures with calcium carbonate, the cultures were allowed to stand brieflyuntil calcium carbonate settled to the bottom.
Prior to flask culture, glycerol stocks of the strains at −80 °C were streaked onto agarplates, which were prepared using Luria–Bertani medium containing 1.5 % agar andappropriate antibiotics at the following concentrations: chloramphenicol, 34 μg/mL; am-picillin, 100 μg/mL; and kanamycin, 50 μg/mL. The strains were incubated overnight at37 °C and a single colony was taken and inoculated into 20 mL of 1× MOPS minimalmedium supplemented with 20 g/L of mannitol and appropriate antibiotics and inducer.The flasks were incubated at 37 °C and 150 rpm in an orbital incubating shaker until thebacteria reached a logarithmic growth phase (about OD550=0.7). An appropriate volume
1940 Appl Biochem Biotechnol (2014) 172:1938–1952

of the growing pre-culture was centrifuged and the pellet was washed and used toinoculate into 60 mL of the medium in shake flasks with a target initial optical densityat 550 nm of 0.05.
Table 1 Phage, strains, plasmids, and primers used in the study
Phage/strain /plasmid/primer
Description Source
Phage
P1vir Generalized transducing phage CGSC, Yale University
Strain
MG1655 E. coli K-12, F−λ− ilvG-negative rfb50 rph1 CGSC, Yale University
DH5-α F-ϕ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1hsdR17 (rK−, mK+) phoA supE44 λ− thi-1 gyrA96 relA1
Invitrogen
DSM01 MG1655 ΔpflB::FRT ΔfrdA::FRT ΔldhA::FRT ; sequentialdeletion of pflB, frdA, and ldhA in MG1655
This study
DSM02 MG1655 Δpta::FRT ΔadhE::FRT ΔfrdA::FRT ldhA::FRT;sequential deletion of pta, adhE, frdA, and ldhA in MG1655
This study
DSM03 DSM01 Δ mgsA::FRT This study
DSM04 DSM02 Δ mgsA::FRT This study
DSM05 DSM04 Δ lldD::FRT-Kan-FRT This study
Plasmids
pCP20 Rep psc101ts Apr Cmr cI857 I PR flp+ CGSC, Yale University
pZA31MCS CmR, p15A, PLtetO-1 Expressys, Germany
pZAldh Streptococcus bovis/equinus ldh under the control of PLtetO-1,CmR, p15A
This study
Primersa
v-frdA GGAGCAGTGGAATAGCGTTC This study
TTGCGTCATAAGGCACTTCA
v-ldhA TTCAATATCGCCATAGCTTTCA This study
AATCAGCTCCCCTGGAATG
v-pflB CGTGAAAACGACCACCATTA This study
CCACTTTCGTGGAGCCTTTA
v-adhE CAAATCATCACCGCACTGAC This study
CGTTTATGTTGCCAGACAGC
v-pta CCTGACTGCCTGATTTCACA This study
CAAAGCTGCGGATGATGAC
v-mgsA TGAGATTTTTGTGCCTGTGC This study
TAAGAAACAGGTGGCGTTTG
v-lldD CGAACAACATCAGGCTGTCA This study
GGGGCAGGATAGCAGAAAAT
c-ldh TTAGGTACCATGACTGCAACTAAACAACACAAAAAAGTTATCC
This study
AATGGATCCTTAGTTTTTGCAAGCAGAAGCGAATTCTTCTTTA
a v and c indicate the primer sequences (5′–3′) that were used for the verification purposes during the creation ofdisruption mutants and cloning purposes, respectively. The forward sequence follows the reverse sequence ineach case. Genes deleted or cloned are apparent from the primer names. Restriction sites are marked in bold
Appl Biochem Biotechnol (2014) 172:1938–1952 1941

Analytical Methods
Optical density was measured at 550 nm (Shimadzu UV mini 1240, Tokyo, Japan) and cellmass was calculated assuming that 1 OD equals 0.34 g dry cell weight per liter. Samples takenfrom flask culture were centrifuged at 5,000 rpm for 10 min and the supernatants were storedat −20 °C until HPLC analysis. Mannitol, glucose, organic acids, and ethanol were measuredusing HPLC (ACME-9000, Younglin Instruments, Seoul, South Korea) equipped with sugarSH1011 column (Shodex, Tokyo, Japan) at 75 °C and with a refractive index detector at 45 °C.A mobile phase of 10 mM H2SO4 was used at a flow rate of 0.5 mL/min. The substrates andmetabolites were quantified by their respective standards of known concentrations. Theenantiomeric purity of lactate was determined enzymatically as per manufacturer protocolusing Megazyme’s D- and L-lactic acid assay kit (Megazyme International Ireland, Co.,Wicklow, Ireland).
Calculation of Fermentation Parameters
Data from cell growth, substrate consumption, and product synthesis were used to calculate theproduct yields. L-Lactate yield is expressed in gram per gram and calculated as follows:
Yield : L‐lactate producedð Þ=Substrate sð Þ consumedð Þ
Substrate(s) here refers to either mannitol or total sugar present in the algal hydrolysate.
Results
Composition of Algal Hydrolysate
The contents in L. japonica, such as lipids, protein, ash, and carbohydrates, have been reportedin recent articles [22, 26]. L. japonica used in the current study was acquired from the samegeographic region and the contents varied slightly from previous reports. When treated with0.1 N HCl at a high temperature (121 °C) for 15 min, laminaran (a linear polysaccharide chainof d-glucopyranose) was extracted from the algal biomass with no corresponding sugarmonomers detected in HPLC analysis. After being treated with glucanases, the hydrolyticproducts of laminaran, mainly glucose and mannitol, yielded a total of 31.2±1.4 g/L of sugars.The highest content was glucose (20.0±1.5 g/L), followed by mannitol (10±1.8 g/L). Apartfrom glucose and mannitol, minor amounts (<1.5 g/L) of galactose, mannose, and xylosemixture were also present. At 10 % (w/v) loading of algal biomass, a final yield of 3.1 % (w/v)sugar was obtained.
Elimination of Endogenous Pathway and Substituting E. coli’s D-Lactate Dehydrogenasewith S. bovis/equinus L-Lactate Dehydrogenase
The degree of reduction for mannitol is more as compared to glucose, and its metabolismgenerates an extra NADH (Fig. 1). A previous study [3] had demonstrated the production ofmore reduced metabolites under anaerobic condition while shifting to fermentative conditionunder aerobic condition in the presence of surplus NADH. A similar type of observation wasalso made in the present study, where the wild type (WT) utilizing mannitol produced morereduced products (succinate+lactate+ethanol) than glucose under microaerobic condition
1942 Appl Biochem Biotechnol (2014) 172:1938–1952

(data not shown). Since the algal hydrolysate contains both glucose and mannitol, thisdiscrepancy due to the degree of reduction might affect L-lactate yield and productivity. Asa result, metabolic engineering was applied to streamline the carbon flux to L-lactate.
In order to do so, metabolic targets from the overall pathway leading to the production of L-lactate were looked at: (1) generation of glycolytic intermediate fructose-6-phosphate, (2)conversion of fructose-6-phosphate to pyruvate via the Embden–Meyerhof–Parnas (EMP)pathway, and (3) conversion of pyruvate to L-lactate. Since glucose and mannitol are readilytaken up and the enzymes of EMP are known to support high fluxes, it was assumed thatefficient L-lactate production could be sought by only improving the conversion of pyruvate to
L-lactate. Since wild-type E. coli MG1655 under microaerobic condition exhibited a
Fig. 1 Pathways involved in the microaerobic utilization of mannitol and glucose and the synthesis offermentation products in wild-type and engineered E. coli. Metabolic engineering strategies employed in thiswork are illustrated by cross bars (disruption of pflB, pta, adhE, frdA, ldhA, mgsA, and lldD). Broken linesillustrate multiple steps. Relevant reactions are represented by the names of the gene(s) coding for thecorresponding enzymes from E. coli: frdABCD, fumarate reductase; ldhA, fermentative D-lactate dehydrogenase;aceEF-lpdA, pyruvate dehydrogenase complex; adhE, acetaldehyde/alcohol dehydrogenase; ackA, acetate ki-nase; fdhF, formate dehydrogenase, part of formate hydrogen lyase complex; hycB-I, hydrogenase 3, part offormate hydrogen lyase complex; pflB, pyruvate formate lyase; pta, phosphate acetyltransferase; pykF, pyruvatekinase; fbaA/B, fructose biphosphate aldolase classes I and II; tpi, triose phosphate isomerase; pfkA/B, 6-phosphofructokinases I and II; mtlD, mannitol-1-phosphate 5-dehydrogenase; pgi, phosphoglucose isomerase;mgsA, methylgloxal sythase; gldA, glycerol dehydrogenase; aldA, aldehyde dehydrogenase A; lldD, respiratoryL-lactate dehydrogenase; gloA, glyoxalase I; gloB, glyoxalase II; dld, respiratory D-lactate dehydrogenase; ldh,fermentative L-lactate dehydrogenase from S. bovis/equinus. Mannitiol-1-P mannitol-1-phosphate, Glucose-1-Pglucose-1-phosphate, F-6-P fructose-6-phosphate, F-1,6 BP fructose-1-6-biphosphate,DHAP dihydroxy-acetonephosphate, Gly-3-P glyceraldehyde-3-phosphate, PEP phosphoenolpyruvate, PYR pyruvate, MG methylglyoxal,L-LAL L-lactaldehyde, S-LG S-lactoylglutathione, GSH glutathione, QH2 reduced quinones, NADH nicotinamideadenine dinucleotide—reduced, ATP adenosine triphosphate, NADPH nicotinamide adenine dinucleotidephosphate—reduced, Pi inorganic phosphate; *Glyoxalase III (gene unknown)
Appl Biochem Biotechnol (2014) 172:1938–1952 1943

heterofermentative behavior (Fig. 2), therefore, several metabolic engineering strategies wereapplied to eliminate the production of native D-lactate and other nonessential side products,viz., ethanol, acetate, and succinate. For this purpose, two main strategies were adopted: (1) thedisruptions of pyruvate-formate lyase (pflB), fumarate reductase (frdA), and D-lactatedehydrogense (ldhA) and (2) the disruptions of phosphoacetyltransferase (pta), alcohol/acet-aldehyde dehydrogenase (adhE), D-lactate dehydrogense (ldhA), and fumarate reductase(frdA). The former was for the reduction of carbon fluxes from pyruvate to acetyl CoA aswell as to lactate and succinate, while the latter was for reductions from acetyl CoA to its sideproducts. The resulting triple (ΔpflB ΔfrdA ΔldhA) and quadruple (Δpta ΔadhE ΔfrdAΔldhA) mutants were named DSM01 and DSM02, respectively (Fig. 1).
In E. coli, multiple pathways for methylglyoxal (MG) detoxification exist, which can leadto a racemic mixture of lactate (Fig. 1), thereby compromising its optical purity. As a result,mgsA was also deleted from strains DSM01 and DSM02 to block any production of D- or L-lactate, resulting in a strain DSM03 (DSM01ΔmgsA) and DSM04 (DSM02 ΔmgsA). Finally,the L-lactate pathway was introduced into DSM03 and DSM04 by the expression of S. bovis/equinus L-lactate dehydrogenase from pZA plasmid and named accordingly as DSM03-L andDSM04-L, respectively.
Production of L-Lactate from Pure Mannitol
As reported earlier, D-lactate yield was dependent on the reduction state of the substrate asa highly reduced substrate such as glycerol was known to produce more D-lactate [28].Mannitol, being one of the major components in algal hydrolysate and slightly less reducedthan glycerol but more reduced than glucose, was thus used as a primary substrate for theinitial assessment of the constructed strains. As shown in Fig. 2, WT, DSM01, DSM02,DSM03-L, and DSM04-L showed differential growth, with the WT growing the most byconsuming all the available mannitol in the media, followed by the rest of the strains.Growth (0.54 g/L) and mannitol utilization (∼8 g/L) in DSM01 were lower as compared togrowth (0.75 g/L) and mannitol utilization (∼12 g/L) in DSM02. Higher growth wasresumed in strains DSM03-L (1.36 g/L) and DSM04-L (1.67 g/L) only after the L-lactatepathway was introduced.
Fig. 2 Product synthesis, mannitol utilization, and cell growth in 48-h shake flask culture of wild-type MG1655and engineered strains. DSM01(ΔpflBΔfrdAΔldhA), DSM02 (ΔptaΔadhEΔfrdAΔldhA), DSM03-L(DSM01ΔmgsA; pZAldh), DSM04-L (DSM02ΔmgsA; pZAldh). Gene expression from plasmid is indicatedby the gene name next to the plasmid. Error bar represents standard deviations from triplicate measurements
1944 Appl Biochem Biotechnol (2014) 172:1938–1952

Under microaerobic condition, wild-type cells produced more reduced products like succi-nate, ethanol, and lactate as major products in regenerating NAD+, with additional ATPgenerated while producing acetate. D-Lactate produced by WT was racemic in nature, with asmall proportion of L-lactate, indicating its possible production through the MG detoxificationpathway. Deletion of ldhA along with the acetate, ethanol, and succinate pathway abolished theproduction of lactate in strains DSM01 and DSM02 (Fig. 2). Even though these strains weredeleted for succinate, lactate, ethanol, and acetate and cannot readily synthesize any commonfermentative product to acquire ATP and achieve redox balance, still, low-level acetate (1.48–2.0 g/L) production was seen in these strains. Subsequent deletion of mgsA ensured completetermination of D/L-lactate production in DSM03 and DSM04 via the MG detoxification route,while incorporation of L-lactate dehydrogenase (ldh) from S. bovis/equinus in a plasmidpZAldh restored L-lactate production in DSM03-L and DSM04-L, which produced 6.46 and8.78 g/L of L-lactate, respectively.
The enantiomeric purity of the produced L-lactate was high in both strains (>99 %). Theobtained yields were 32 and 44 % of the maximum theoretical yield from mannitol forDSM03-L and DSM04-L, respectively (theoretical lactic acid yield for the conversion of sugaris 1.0 g acid produced per gram of sugar consumed). The above results indicate that the strainsdesigned and engineered as per this strategy was successful in converting mannitol to L-lactate.Because DSM04-L outperformed DSM03-L when compared in terms of yield and volumetricconcentration and DSM03-L showed no additional benefit of using pflB deletion in conjunc-tion with just directly blocking the competitive fermentative products, DSM04-L was chosenfor further metabolic engineering and product enhancement.
Effect of lldD Deletion on Lactate Production
The chosen strain, DSM04-L, as mentioned above was examined for growth kinetics, mannitolutilization, and L-lactate production. As evident from Fig. 3a, this strain produced 7.34 g/L in72 h at a yield of 0.38 g/g. Besides L-lactate, only a small amount of acetate (<1.2 g/L) wasalso produced, demonstrating the near homolactic nature of the fermentation. However, acloser examination of the dynamics of cell growth, mannitol consumption, and productsynthesis at the latter stage of the fermentation revealed that there was a decrease in L-lactateconcentration (peak value, 10 g/L) and yield (peak value, 0.7 g/g), and the culture neverattained the stationary phase even after the exhaustion of mannitol from the media (Fig. 3b).These observations suggested that the accumulation of L-lactate in the medium triggered itsconsumption by the respiratory L-lactate dehydrogenase (lldD), which catalyzes the oxidationof L-lactate to pyruvate [25]. Subsequent deletion of lldD in strain DSM04 and the overex-pression of ldh resulted in a clear stationary phase following the depletion of mannitol from themedium, along with no decrease in yield and volumetric concentration (Fig. 3c). This strain,named DSM05 (pZAldh), produced 13.55 g/L of L-lactate in 72 h, with an overall productyield of 0.7 g/g of mannitol. The obtained volumetric concentration clearly surpasses theperformance of its parent strain, DSM04 (pZAldh), and demonstrates the role of lldD inmaintaining high volumetric concentration and yield.
Production of L-Lactate from Algal Hydrolysate
The use of abundant algal biomass provides a very novel and economical route for theproduction of biorefinery products. Apart from native lactic acid bacteria, the usage of abiocatalyst developed in this work is of great relevance when cultured in a minimal mediumcontaining only algal hydrolysate as it provides an alternative source of substrate in producing
Appl Biochem Biotechnol (2014) 172:1938–1952 1945

L-lactate. The engineered strain performed very well when algal hydrolysate was used as acarbon source. Due to the presence of glucose in the hydrolysate, cell growth was very prolificand all the available glucose was completely utilized by 36 h, while ∼50 % of the availablemannitol also was utilized during this time. At the end of 72 h, batch culture produced 23.5 g/Lof L-lactate at a yield of 0.77 g/g (Fig. 4). The obtained yield was slightly better yet comparableto that attained with pure mannitol. L-Lactate constitutes 88 % of the total products formed,followed by acetate and cell mass, which constituted 6.5 and 5 %, respectively. However, toassess the potential of this strain for any further improvement in its yield and volumetric
Fig. 3 Kinetics of L-lactate pro-duction by strains DSM04-L (a,b) and DSM05 (pZAldh) (c) inshake flasks containing minimalmedium with 20 g/L of mannitol.Data are shown for the concentra-tion of cells (triangles), mannitol(circles), L-lactate (squares), and L-lactate yield (diamonds). a Fer-mentation profile for strainDSM04-L. b Zoomed out profilefor strain DSM04-L at the latestages of cultivation. c Fermenta-tion profile for strain DSM05(pZAldh). Coefficients of variation(i.e., standard deviation/average×100) were below 5 % in all cases
1946 Appl Biochem Biotechnol (2014) 172:1938–1952
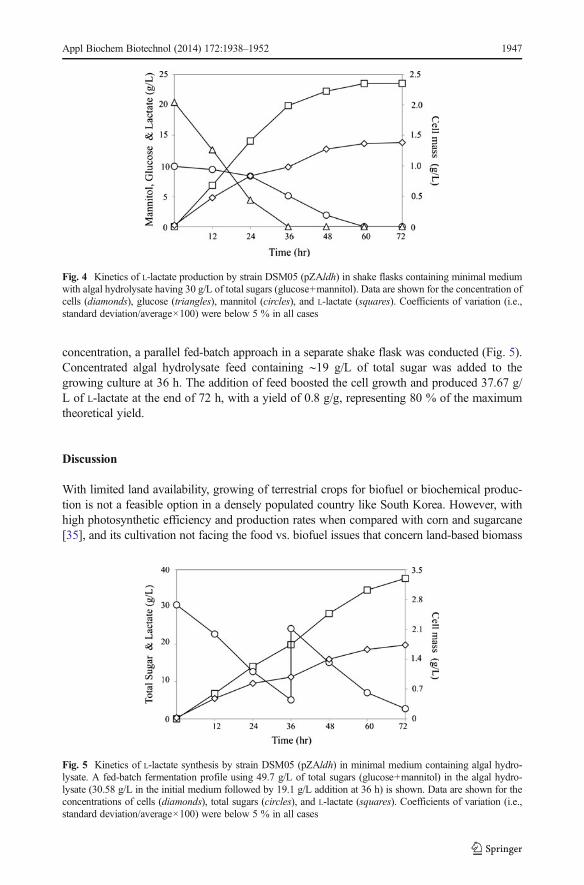
concentration, a parallel fed-batch approach in a separate shake flask was conducted (Fig. 5).Concentrated algal hydrolysate feed containing ∼19 g/L of total sugar was added to thegrowing culture at 36 h. The addition of feed boosted the cell growth and produced 37.67 g/L of L-lactate at the end of 72 h, with a yield of 0.8 g/g, representing 80 % of the maximumtheoretical yield.
Discussion
With limited land availability, growing of terrestrial crops for biofuel or biochemical produc-tion is not a feasible option in a densely populated country like South Korea. However, withhigh photosynthetic efficiency and production rates when compared with corn and sugarcane[35], and its cultivation not facing the food vs. biofuel issues that concern land-based biomass
Fig. 4 Kinetics of L-lactate production by strain DSM05 (pZAldh) in shake flasks containing minimal mediumwith algal hydrolysate having 30 g/L of total sugars (glucose+mannitol). Data are shown for the concentration ofcells (diamonds), glucose (triangles), mannitol (circles), and L-lactate (squares). Coefficients of variation (i.e.,standard deviation/average×100) were below 5 % in all cases
Fig. 5 Kinetics of L-lactate synthesis by strain DSM05 (pZAldh) in minimal medium containing algal hydro-lysate. A fed-batch fermentation profile using 49.7 g/L of total sugars (glucose+mannitol) in the algal hydro-lysate (30.58 g/L in the initial medium followed by 19.1 g/L addition at 36 h) is shown. Data are shown for theconcentrations of cells (diamonds), total sugars (circles), and L-lactate (squares). Coefficients of variation (i.e.,standard deviation/average×100) were below 5 % in all cases
Appl Biochem Biotechnol (2014) 172:1938–1952 1947

production [38], marine algae/seaweeds can truly be an attractive alternative renewableresource. Globally, brown algae accounts for 75 % of the seaweed cultivation, while one ofits species, L. japonica, accounts for 38 % of the South Korean seaweed cultivation. Giventheir abundant production with further scope of expansion, L. japonica can definitely belooked upon as a possible biomass source.
In brown algae, alginic acid is the major component, while laminaran, a polysaccharidecomposed of glucosyl units (∼20–25) terminating either in glucose (G-chain) or in mannitol(M-chain), is the main carbohydrate reserve and constitutes ∼30–50% of the dried mass. In ourexperimental procedure, laminaran was the major extract after acid hydrolysis as the hightemperature coagulated the majority of alginate present in the biomass under acidic condition.However, to our surprise, no monomeric sugars were detected upon HPLC analysis followingacid hydrolysis, which is contradictory to the study by Horn et al. [15], where mannitol wasrecovered under acidic condition at 65 °C. Following enzymatic saccharificationwith glucanasecocktail, monomeric sugars like glucose, galactose, mannose, xylose, and mannitol weredetected. Glucose was the major sugar, followed by mannitol, in the algal hydrolysate asenzymatic activity on laminaran tends to liberate more glucose units than mannitol. Eventhough, this result is contrary to the result reported elsewhere [15, 22], where mannitol wasprojected as a major component, but the yield of total sugar (glucose+mannitol) liberated wascomparable to an earlier report by Kim et al. [22]. The amount of recovered sugars could besufficient enough to be used as a carbon source for the production of biochemicals and biofuels.
A heterologous L-lactate pathway can only be incorporated when native D-lactate pathwayalong with nonessential pathway leading to acetate, succinate, and ethanol is blocked (Fig. 1).Two strategies were applied to engineer E. coli by blocking carbon flow either at the pyruvatenode (strain DSM01) or at the pyruvate/acetyl CoA node (strain DSM02). In engineered strainDSM01, growth and substrate utilization, when compared with WT, were lower, which ismainly due to the non-availability of acetyl CoA due to pflB deletion. Since acetyl-CoAoccupies a central role in the regulation of glycolysis, gluconeogenesis, TCA cycle, and fattyacid synthesis, lack of it creates an imbalance and hampers cell growth. Apart from this, acetyl-CoA is a precursor to acetate and ethanol production, and lack of additional ATP from theacetate route might also have contributed to the decreased cell growth and substrate utilization.Even though the growth of DSM02 was less compared to WT, an adequate quantity of acetyl-CoA in the system helped sustain the cells and their growth. However, lack of a NAD+
regeneration route might also have contributed to the decreased growth and mannitol utiliza-tion. The WT, however, grew faster, as expected, due to the lack of any deletions andpromoting pathways which regenerate NAD+ in a balanced way.
Under microaerobic condition, pyruvate-formate lyase is the primary route for pyruvateconversion to acetyl CoA, even though strain DSM03-L and its parent strain DSM01 producedhigher concentrations of acetate than that observed in DSM04-L and its parent strain DSM02.This aspect can be explained by the low-level presence of acetyl CoA, which subsequentlyforms acetate and ATP in turn, and was mainly due to the leakiness of the aerobic pyruvatedehydrogenase complex (AceEF and LpdA). Whereas, acetate formation in DSM04-L and theparent strain DSM02 is directly blocked by a pta deletion, lowering the level of acetate. Sincethe deletion of ldhA and other nonessential pathways in DSM01 and DSM02 made thesestrains not able to synthesize any common fermentative products to produce ATP and redoxbalance, a small increase in acetate accumulation would account for their observed growth asits formation would lead to the generation of additional two ATP molecules per molecule ofmannitol consumed via substrate-level phosphorylation.
Typically, L-lactate is not a native end product of E. coli, but an intermediate produced intwo steps: (1) conversion of MG to L-lactaldehyde and (2) conversion of L-lactaldehyde to L-
1948 Appl Biochem Biotechnol (2014) 172:1938–1952

lactate [2, 4, 5, 41]. However, the synthesis of L-lactate through this route is not desirable dueto the existence of another native MG detoxification pathway in E. coli that can lead to theproduction of D-lactate, thereby compromising the enantiomeric purity of the product. Thus, tomaintain optical purity, methylglyoxal synthase (mgsA) of the MG bypass was deleted toprevent the synthesis of D-lactate. Finally, the incorporated L-lactate dehydrogenase from S.bovis/equinus in DSM03-L and DSM04-L provided a route for NAD+ regeneration, therebyreinstating growth and mannitol utilization and making L-lactate as the primary product.Deletion of mgsA is beneficial, MG being a toxic product, and its accumulation could severelyimpair the metabolism and lead to cell death [5, 41]. However, the production of L-lactate viaMG bypass route is also energy-inefficient as no ATP is generated from its conversion fromDHAP, but rather DHAP formation from mannitol consumes two ATP equivalent (either ATPor PEP)—first its conversion to mannitol-1-phosphate and, second, its conversion to fructose-1-6-biphosphate—whereas the overall conversion of mannitol to lactate through the DHAP-PEP route would lead to the net production of one ATP per molecule of L-lactate produced andone reducing equivalent per molecule of mannitol consumed.
Mannitol→ 2 lactateþ 2 ATPþ NADH ð1ÞE. coli uses NADH:ubiquinone oxidoreductase-I of the aerobic and anaerobic respiratory
chain to oxidize NADH to NAD+, transfer its electrons to the quinone pool, and simultaneouslyexpel 4H+ outside the cell membrane [10], thereby generating the proton motive force (PMF).
NADHþ an Ubiquinoneþ 5Hþ → NADþ þ an Ubiquinolþ 4Hþperiplasmic spaceð Þ ð2Þ
Whereas under microaerobic condition, cytochrome bd oxidase is expressed [34] andcytochrome bd-1 oxidase encoded by cydAB bring about the reduction of oxygen (0.5 mol)to water by utilizing two electrons from ubiquinol. Despite contributing 2H+ to the PMF,cytochrome bd-I oxidase is not a proton pump [33].
An Ubiquinolþ 0:5 O2 þ 2 Hþ → an Ubiquinoneþ H2Oþ 2 Hþperiplasmic spaceð Þ ð3Þ
Since the respiratory chain formed by NADH dehydrogenase and cytochrome bd oxidase iscoupled to the generation of a PMF across the cytoplasmic membrane and with H+/ATPaverage stoichiometry of 3.0 for E. coli F0F1 ATP synthetase under aerobic and anaerobicconditions [19, 20], the mobilized 6H+ can theoretically generate two molecules of ATP. Thus,from Eqs. 2 and 3, the overall reaction can be represented below as
NADHþ 0:5 O2 → NADþ þ H2Oþ 2 ATP ð4ÞHowever, the overall stoichiometry for the conversion of mannitol to L-lactate in strain
DSM05 (pZAldh) can be represented as
Mannitolþ 0:5 O2 → 2 lactateþ 4 ATPþ H2O ð5ÞBased on Eq. 5, it can be inferred that L-lactate production from mannitol can generate two
molecules of ATP per molecule of lactate produced, justifying the importance of the substrate’sdegree of reduction in determining the product profile. Since the algal hydrolysate containsboth glucose and mannitol as a major component, the whole equation can be described as
GlucoseþMannitolþ 0:5 O2 → 4 lactateþ 6 ATPþ H2O ð6Þ
Based on Eq. 6, it can be concluded that dual substrate presence in the algal hydrolysate isboth beneficial for L-lactate production and yet an energetically efficient process.
Appl Biochem Biotechnol (2014) 172:1938–1952 1949

Kinetic study of DSM04-L revealed that in the absence of a carbon source, the accumulated
L-lactate was reverted back to pyruvate due to the activity of respiratory L-lactate dehydroge-nase (lldD), resulting in a decrease in volumetric concentration and yield. The deletion of lldDand the incorporation of the ldh route led to an increase in L-lactate concentration (13.5 g/L)and yield (0.7 g/g). This deletion was beneficial and yet necessary; otherwise, the pyruvateproduced would eventually lead to acetate production via an available alternative route.
Under batch cultivation using algal hydrolysate, diauxic growth was observed, which wasmainly due to the presence of two different sugars, glucose and mannitol. Due to the presenceof glucose, cell growth was higher and at the end of fermentation 23.5 g/L of L-lactate wasproduced at a yield of 0.77 g/g. However, the yield improved marginally to 0.8 g/g and thevolumetric concentration improved to 37.7 g/L when the fed-batch approach in a shake flaskwas undertaken. The obtained 80 % L-lactate yield from DSM05 (pZAldh) was better than the69 % yield obtained in an earlier study, where an engineered E. coli strain (JP204) produced L-lactate from a plasmid expressing ldh from Lactobacillus casei [7]. As compared to otherprevious studies producing L-lactate from engineered E. coli [9, 46], the reported yield andvolumetric concentration of L-lactate in this study were lower than the preferred values, but thepresent study provided another dimension of using algal hydrolysate rather than pure sugarsfor L-lactate production. Additional improvements through the use of metabolic evolutiontechniques, processes-based modification, including bioreactor-based fed-batch fermentationand high cell density cultures, are also envisioned to further improve the volumetric rate andproductivity of L-lactate. Apart from this, the development of whole-cell biocatalyst is plannedto eliminate the usage of enzymatic hydrolysis.
Acknowledgments This research was supported by the National Research Foundation (NRF) of MEST (grantno. 2012M1A2A2026560) and Korea University.
References
1. Adams, J. M., Gallagher, J. A., & Donnison, I. S. (2009). Fermentation study on Saccharina latissima forbioethanol production considering variable pre-treatments. Journal of Applied Phycology, 21, 569–574.
2. Baldoma, L., & Aguilar, J. (1987). Involvement of lactaldehyde dehydrogenase in several metabolicpathways of Escherichia coli K12. Journal of Biological Chemistry, 262, 13991–13996.
3. Berrı´os-Rivera, S. J., Bennett, G. N., & San, K. Y. (2002). The effect of increasing NADH availability on theredistribution of metabolic fluxes in Escherichia coli chemostat cultures.Metabolic Engineering, 4, 230–237.
4. Booth I. R. (2005). Chapter 3.4.3 Glycerol and methylglyoxal metabolism. In R. Curtis III (Ed.), EcoSal—Escherichia coli and Salmonella: cellular and molecular biology. Washington, D.C.: ASM Press.
5. Booth, I. R., Ferguson, G. P., Miller, S., Li, C., Gunasekera, B., & Kinghorn, S. (2003). Bacterial productionof methylglyoxal: a survival strategy or death by misadventure? Biochemical Society Transactions, 31(6),1406–1408.
6. Carlsson A. S., Van-Beilen J., Moller R., Clayton D. (2007). Micro- and macro-algae: utility for industrialapplications. In D. Bowles (Ed.) Outputs from the EPOBIO project. CNAP, University of York, York, UK.
7. Chang, D. E., Jung, H. C., Rhee, J. S., & Pan, J. G. (1999). Homofermentative production of D- or L-lactate inmetabolically engineered Escherichia coli RR1. Applied and Environmental Microbiology, 65, 1384–1389.
8. Chynoweth, D. P. (2002). Review of biomethane from marine biomass. Gainesville: Department ofAgricultural and Biological Engineering, University of Florida.
9. Dien, B. S., Nichols, N. N., & Bothast, R. J. (2001). Recombinant Escherichia coli engineered for productionof L-lactic acid from hexose and pentose sugars. Journal of Industrial Microbiology and Biotechnology, 27,259–264.
10. Efremov, R. G., Baradaran, R., & Sazanov, L. A. (2010). The architecture of respiratory complex I. Nature,465, 441–445.
11. Ge, L., Wang, P., & Mou, H. (2011). Study on saccharification techniques of seaweed wastes for thetransformation of ethanol. Renewable Energy, 36, 84–89.
1950 Appl Biochem Biotechnol (2014) 172:1938–1952

12. Hanssen, J. F., Indergaard, M., Ostgaard, K., Baevre, O. A., Pedersen, T. A., & Jensen, A. (1987). Anaerobicdigestion of Laminaria spp. and Ascophyllum nodosum and application of end products. Biomass, 14, 1–13.
13. Hansson, G. (1983). Methane production from marine, green macro-algae. Resource Conservation, 8, 185–194.
14. Hofvendahl, K., & Hahn-Hagerdal, B. (2000). Factors affecting the fermentative lactic acid production fromrenewable resources. Enzyme and Microbial Technology, 26, 87–107.
15. Horn, S. J., Aasen, I. M., & Ostgaard, K. (2000). Ethanol production from seaweed extract. Journal ofIndustrial Microbiology and Biotechnology, 25, 249–254.
16. Ishida, N., Saitoh, S., Ohnishi, T., Tokuhiro, K., Nagamori, E., Kitamoto, K., & Takahashi, H. (2006).Metabolic engineering of Saccharomyces cerevisiae for efficient production of pure L-(+)-lactic acid. AppliedBiochemistry and Biotechnology, 129–132, 795–807.
17. John, R. P., Anisha, G. S., Nampoothiri, K. M., & Pandey, A. (2011). Micro and macro-algal biomass: arenewable source for bioethanol. Bioresource Technology, 102, 186–193.
18. Jung, K. W., Kim, D. H., & Shin, H. S. (2011). Fermentative hydrogen production from Laminaria japonicaand optimization of thermal pretreatment conditions. Bioresource Technology, 102, 2745–2750.
19. Kashket, E. R. (1982). Stoichiometry of the H+-ATPase of growing and resting, aerobic Escherichia coli.Biochemistry, 21(22), 5534–5538.
20. Kashket, E. R. (1983). Stoichiometry of the H+-ATPase of Escherichia coli cells during anaerobic growth.FEBS Letters, 154(2), 343–346.
21. Kharas G. B., Sanchez-Riera F., Severson D. K. (1994). Polymers of lactic acid. In D. P. Mobley (Ed.), Plasticsfrom microbes: microbial synthesis of polymers and polymer precursors (pp. 93–137). Munich: Hanser.
22. Kim, N. J., Li, H., Jung, K., Chang, H. N., & Lee, P. C. (2011). Ethanol production from marine algalhydrolysates using Escherichia coli KO11. Bioresource Technology, 102, 7466–7469.
23. Lee, S. M., & Lee, J. H. (2010). Influence of acid and salt content on the ethanol production from Laminariajaponica. Applied Chemistry for Engineering, 21, 154–161.
24. Lee, S. M., & Lee, J. H. (2011). The isolation and characterization of simultaneous saccharification andfermentation microorganisms for Laminaria japonica utilization. Bioresource Technology, 102, 5962–5967.
25. Luchi, S., Aristarkhov, A., Dong, J. M., Taylor, J. S., & Lin, E. C. (1994). Effects of nitrate respiration onexpression of the Arc-controlled operons encoding succinate dehydrogenase and flavin-linked L-lactatedehydrogenase. Journal of Bacteriology, 176, 1695–1701.
26. Malihan, L. B., Nisola, G. M., & Chung, W. J. (2012). Brown algae hydrolysis in 1-n-butyl-3-methy-limidazolium chloride with mineral acid catalyst system. Bioresource Technology, 118, 545–552.
27. Matsui, T., & Koike, Y. (2010). Methane fermentation of a mixture of seaweed and milk at a pilot-scale plant.Journal of Bioscience and Bioengineering, 110, 558–563.
28. Mazumdar, S., Clomburg, J. M., & Gonzalez, R. (2010). Engineered Escherichia coli strains for thehomofermentative production of D-lactic acid from glycerol. Applied and Environmental Microbiology,76, 4327–4336.
29. Neidhardt, F. C., Bloch, P. L., & Smith, D. F. (1974). Culture medium for enterobacteria. Journal ofBacteriology, 119, 736–747.
30. Okano, K., Tanaka, T., Ogino, C., Fukuda, H., & Kondo, A. (2010). Biotechnological production ofenantiomeric pure lactic acid from renewable resources: recent achievements, perspectives, and limits.Applied Microbiology and Biotechnology, 85, 413–423.
31. Park, J. I., Lee, J. W., Sim, S. J., & Lee, J. H. (2009). Production of hydrogen from marine macro-algaebiomass using anaerobic sewage sludge microflora. Biotechnology and Bioprocess Engineering, 14, 307–315.
32. Park, J. I., Wu, H. C., & Lee, J. H. (2008). Production of bio-energy from marine algae: status andperspectives. Korean Chemical Engineering Research, 46, 833–844 (in Korean).
33. Puustinen, A., Finel, M., Haltia, T., Gennis, R. B., & Wikstrom, M. (1991). Properties of the two terminaloxidases of Escherichia coli. Biochemistry, 30(16), 3936–3942.
34. Reid, G. A., & Ingledew, W. J. (1979). Characterization and phenotypic control of the cytochrome content ofEscherichia coli. Biochemical Journal, 182(2), 465–472.
35. Ross, A. B., Jones, J. M., Kubacki, M. L., & Bridgeman, T. (2008). Classification of macroalgae as fuel andits thermochemical behavior. Bioresource Technology, 99, 6494–6504.
36. Sambrook, J., Fritsch, E. F., & Maniatis, T. (1989).Molecular cloning: a laboratory manual (2nd ed.). ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press.
37. Shi, X., Jung, K. W., Kim, D. H., Ahn, Y. T., & Shin, H. S. (2011). Direct fermentation of Laminariajaponica for biohydrogen production by anaerobic mixed cultures. International Journal of HydrogenEnergy, 36, 5857–5864.
38. Singha, A., Nigamb, P. S., & Murphya, J. D. (2010). Renewable fuels from algae: an answer to debatableland based fuels. Bioresource Technology, 102, 10–16.
Appl Biochem Biotechnol (2014) 172:1938–1952 1951

39. Sluiter A. (2012). Determination of structural carbohydrates and lignin in biomass. Laboratory analyticalprocedure (LAP), Issue date: April 2008, Revision date: August 2012 (version 08-03-2012), NationalRenewable Energy Laboratory, USA. Retrieved from http://www.nrel.gov/biomass/analytical_procedures.html.
40. Tokuhiro, K., Ishida, N., Nagamori, E., Saitoh, S., Onishi, T., Kondo, A., & Takahashi, H. (2009). Doublemutation of the PDC1 and ADH1 genes improves lactate production in the yeast Saccharomyces cerevisiaeexpressing the bovine lactate dehydrogenase gene.AppliedMicrobiology and Biotechnology, 82(5), 883–890.
41. Totemeyer, S., Booth, N. A., Nichols, W. W., Dunbar, B., & Booth, I. R. (1998). From famine to feast: therole of methyl-glyoxal production in Escherichia coli. Molecular Microbiology, 27, 553–562.
42. Ulgiati, U. (2001). A comprehensive energy and economic assessment of biofuels: when “green” is notenough. Critical Reviews in Plant Sciences, 20, 71–106.
43. Wee, Y. J., Kim, J. N., Yun, J. S., & Ryu, H. W. (2004). Utilization of sugar molasses for economical L-(+)-lactic acid production by batch fermentation of Enterococcus faecalis. Enzyme and Microbial Technology,35, 568–573.
44. Wee, Y. J., Yun, J. S., Park, D. H., & Ryu, H. W. (2004). Biotechnological production of L-(+)-lactic acidfrom wood hydrolyzate by batch fermentation of Enterococcus faecallis. Biotechnology Letters, 26, 71–74.
45. Yazdani, S. S., & Gonzalez, R. (2008). Engineering Escherichia coli for the efficient conversion of glycerolto ethanol and co-products. Metabolic Engineering, 10, 340–351.
46. Zhou, S., Shanmugam, K. T., & Ingram, L. O. (2003). Functional replacement of the Escherichia coli D(−)-lactate dehydrogenase gene (ldhA) with the L(+)-lactate dehydrogenase gene (ldhL) from Pediococcusacidilactici. Applied and Environmental Microbiology, 69, 2237–2244.
1952 Appl Biochem Biotechnol (2014) 172:1938–1952