investigations on the feeding habits of the rocky-shore ... · h. fusca is thought to be a...
TRANSCRIPT

HELGOL,~NDER MEERESUNTERSUCHUNGEN Helgol~nder Meeresunters. 52, 159-177 (1998)
Investigations on the feeding habits of the rocky-shore mite Hyadesia fusca (Acari: Astigmata : Hyadesiidae):
diet range, food preference, food quality, and the implications for distribution patterns
J e n s Bricking
Universit~t Bremen, Institut ffir Okologie und Evolutionsbiologie; Leobener Str. - NW2, D-28359 Bremen, email: jbueck@biologie .uni=bremen.de
ABSTRACT: Within the food web of estuarine and marine rocky shore ecosystems phytophagous mites of terrestrial and marine origin constitute an important part as grazers on algae and as a food source for certain arthropods, especially zoophagous mites. This investigation deals with the feeding biology of Hyadesia fusca taking as an example a population located on an artificial rocky shore of the middle Weser estuary in Northern Germany. The species is characterized by a broad diet range; in feeding experiments diatoms, lichens, detritus as well as blue, red and green algae were accep- ted. Even analyses of faecal pellets produced by field specimen suggest a non-specific feeding ha- bit. However, the influence of certain diets on mortality, offspring number and rearing success showed that the food quality differs significantly. The most suitable food, the Ulvaceae Blidingia, was clearly preferred in a series of pairwise choice tests. These findings correlate with the vertical zonation of the field population i.e.: higher population densities in the vegetation zone dominated by Blidingia. It can be concluded that in addition to abiotic factors food supply could play an impor- tant role for distribution patterns of phytophagous mites.
INTRODUCTION
Rocky shores are inhabi ted by a variety of microarthropods of mar ine as well as
terrestrial origin. In the g reen a lgae belt of the upper intertidal zone and in the lower supralittoral zone covered with l ichens mites often dominate the microfauna with regard
to species diversity and abundance (Schuster, 1965; Bartsch, 1978; Bartsch, 1982). Spe- cies composition, zoogeography, and vert ical distribution of this group are weU known
(e.g. Halbert, 1920; Otto, 1936; Schuster, 1979; Schulte, 1977; Pugh & King, 1985b,c; Ernst et al., 1993), but the functional role of mites in this ecosys tem is only poorly understood.
Especially the in tegrat ion of terrestrial microar thropods in the rocky shore food web and the inf luence of biotic factors such as food supply on distribution patterns needs further investigation.
The mite coenosis of rocky shores comprises a wide spectrum of feed ing habits, e.g. zoophagous (e.g. most Gamasina, Rhagidi idae, Erythraeidae, Bdellidae, Halacarinae) , phytophagous (e.g. most Oribatida, Nanorches t idae , Hyadesi idae, Rhombognathinae) ,
and saprophagous (some Gamasina, Uropodina, and Oribatida) species (Schuster, 1979;
Pugh & King, 1985a).
�9 Biologische Anstalt Helgoland, Hamburg

160 J. Bricking
On artificial ha rdsubs t ra t e s of es tuar ies in nor thern Germany, espec ia l ly the surface- inhabi t ing algivorous Rhombogna th inae and the species Hyadesia [usca (Lohmann) can reach ex t r eme ly h igh popula t ion densi t ies (Siemer, 1996; Bricking, 1995). At the Baltic Sea Hyadesia fusca often m a d e up over 70 % of the total fauna in b r ack i sh -wa te r rock pools with a dense g reen a lgae vege ta t ion (Ganning, 1971) and on artificial sea defences (Kronberg, 1983). The H y a d e s i i d a e have a t ransoceanic dis t r ibut ion and can be found in the in ter t idal and lower supral i t tora l of a lmost every rocky shore. In Europe the distr ibu- tion of the eu ryha l i ne -mar ine H. fusca extends from nor thern Norway (70~ all the way down to the Brit tany coast in Nor thwes te rn France (48~ (Schulte, 1977; Schuster , 1988; Fain & Schuster, 1989). In all known cases they are viviparous; the pos tna ta l d e v e l o p m e n t comprises larva, p ro tonymph, t r i tonymph and the sexual d imorphic adult . Mites of this species can be found (1) epi l i thic and within the a lgae cover (Otto, 1936; Kronberg , 1983; Ernst et aI., 1993), (2) mesol i th ic in the outer region or fr inge of crevices (Schulte, 1977; Pugh, 1985b), (3) among barnac les (BSnard, 1961; Pugh 1985b) and (4) p e r m a n e n t l y sub- mersed in the g reen a lgae vege ta t ion of rock pools (Halbert , 1920; Gann ing , 1970; Ost- mann & R6nnberg, 1991). Moreover Gann ing (1970) desc r ibed the inside of the tubular thalli of Enteromorpha intestinalis (L.) Link (Chlorophyta, Ulvaceae) as be ing a microha- bi tat of, part icularly, the larvae. However , in the Weser es tuary H. fusca l ives only epil i th- ic and within the thin layer of epil i thic g reen algae. During res t ing phase s or per iods of des iccat ion H. fusca wi thdraws into small cavit ies or congrega t e s at the f r inges of crevi- ces (Ernst et al., 1993).
H. fusca is thought to be a spec ia l i sed inhab i t an t and feeder of Enteromorpha (Ganning, 1970; Schuster, 1979) but the f i lamentous g reen a lga Cladophora glomerata Kritzing (Ganning, 1971), Cyanophy te s (Kronberg, 1983) and microphytes (B~nard, 1961) have also been found in their guts or faecal pellets . H. fusca itself is a f requen t p rey for zoophagous mites l ike Abrolophus cf. rubipes (Trouessart) and Bdella septemtrionalis Atyeo et Tux. (Witte, 1972; Alberti , 1973) and has been found in the gut of the omnipha- gous a m p h i p o d Platorchestia ptatensis (Kroyer) (Stueber, pers. comm.), the i sopod Ligia oceanica (L.) (v. d. Heydt , pers. comm.) and of the rock pool - inhabi t ing fish Gasterosteus aculeatus L. (Ganning, 1970).
The ver t ical dis t r ibut ion of H. fusca in the Weser es tuary ranges from just be low the lower dis t r ibut ion limit of the orange l ichen Caloplaca marina (Wedd.) Zahlbr. down to the midd le in ter t ida l (Bricking, in press; Ernst, 1996). Never the less , the popu la t ion is con- cen t ra ted on a na r row band a long M e a n High Water level (MHW), d o m i n a t e d by g reen a lgae of the genus Blidingia. In 1991 a yea r ly mean a b u n d a n c e of 21 ind . / cm z and a maxi- mum of 87 ind . /cm 2 was coun ted in this l i t toral zone, while only few Hyades i i d s have been found in the supral i t tora l fr inge (domina ted by Cyanophytes ) or in the midd le inter- t idal wi th its p r e d o m i n a n t coa t ing with Fucus vesiculosus L. and Enteromorpha/ntesti- nalis. Fur thermore , it was obse rved that, w h e n compar ing ad jacen t sur faces that were different in rock type and therefore a lgae cover (Blidingia on concrete and Enteromor- pha on sands tone) but s i tua ted on the same t idal level, the popula t ion dens i ty of H. fusca was about ten t imes h igher on the Blidingia-dominated area (Bricking, in press).
The objec t of this inves t iga t ion was to answer the quest ion conce rn ing w he the r the diet range, different f eed ing p re fe rences or different qual i t ies of the p r e d o m i n a n t food sources m a y inf luence the ver t ical dis t r ibut ion and popula t ion dens i ty in cer ta in lit toral or vege ta t iona l zones.

Feeding habits of Hgadesia fusca 161
MATERIALS AND METHODS
S a m p l i n g a n d m a i n t e n a n c e of the m i t e s
The sampling of Hyadesiids assigned for laboratory experiments and field experi- ments was carried out at the location Langli i t jen I. This is the ruin of a fortress, located in the mesohal ine zone of the middle Weser estuary near Nordenham (kin 69.5, 53034 , N. - 8 ~ 31' E., salinity 9-19 PSU), ex tending as a pen insu la into the Wadden Sea (Nationalpark Wattenmeer). The mean tidal rise is about 3.6 m; the bank reinforcement ranges from ca. m e a n sea level up to 2 m above the mean high water line with a slope of about 40 ~ The surface consists of weathered sandstone blocks which are stabilised by cast concrete. The vegetat ion shows a distinct zonation with Enteromorpha sp. and Fucus vesiculosus do- mina t ing in the middle intert idal zone, Enteromorpha sp. and Blidingia sp. in the upper intertidal zone, different Cyanophytes (but no black lichens) in the supralittoral fringe and lichens (Caloplaca spp., Xanthor~a spp., Lecanora spp.) in the supralittoral zone (Ernst, 1995; Haase, 1995).
The Hyadesiids, obtained from Blidingia samples scraped off the rock surface, were separated into the ontogenet ical instars, respectively sexes, and kept in cubic transpa- rent polystyrol chambers (25 x 25 mm surface, 20 mm high) with lids. The bottom of the vessels was covered with piaster, enriched with a small amount of charcoal. The plaster was mois tened with diluted artificial seawater (15 PSU, WIMEX | and the vessels stored in a light thermostat at 20 • 1~ with a 12h/12 h l ight-dark cycle. Saturated air humidi ty was main ta ined by adding fresh water wheneve r necessary.
The algae and lichens ass igned as food were sampled at different locations along the meso- and polyhal ine zone of the Weser estuary, thoroughly c leaned in diluted seawater with the aid of a dissecting microscope and then distributed in a homogenous thin layer onto the plaster bottom. Surplus and even distr ibution of the food obviated the need for searching behaviour. The experiments were started by placing the mites into the feeding vessels.
R e c o r d i n g of the d ie t r a n g e in t he l a b o r a t o r y a n d in the f ie ld
After a period of at least 5 hours without food (complete defecation of food ingested before), adults (all tests) and also larvae, protonymphs, and tr i tonymphs (most tests) were transfered to prepared feeding vessels (one mite per vessel) and checked daily for at least one week. Feed ing was attested, if new faecal pellets, clearly dis t inguishable from food fragments by their peritrophic membrane , were produced.
To examine the diet range in the field, adults from different littoral, or vegetation, zones were collected individually, r insed in c lean sea water, c leaned thoroughly unde r a field dissecting microscope and then transferred into smaU glass tubes filled with diluted artificial seawater. Their faecal pellets were ana lysed for food residues which were com- pared to preparat ions of detritus and plant samples collected at exactly the same places. For these analyses squeeze preparat ions were examined either in vivo or after s taining with Toluidin Blue (1%) or iodin- iodin-potass ium (positive reaction with starch of pyre- noids) with the aid of a DIC microscope (stainings v. Gerlach, 1984). An epifluorescence microscope was used to check the food residues for primary fluorescence and as a test for Cyanophytes (phycocyan-mediated primary f luorescence with green light; see Kronberg,

162 J. B~icking
1983). The identification of food components in faecal pellets produced by field speci- mens was facilitated by comparing them to faecal pellets of laboratory spec imen fed with certain known diets.
E x p e r i m e n t s o n food p r e f e r e n c e
Different plant species were offered to HFadesia fusca in several tests with two food
types each. The plaster bottom of the feeding vessels, similar to that descr ibed above but slightly smaller (18 x 18 mm surface, 15 mm high), was provided with food A and with
food B, each kind covering a third of the total area (Fig. 1). The mites were placed along the midl ine be tween food A and B and their actual distribution was checked after 24
hours. A statistically significant difference in the n u m b e r of test animals on the two food areas wa's interpreted as a preference for the food type concerned. By keep ing the rela-
Fig. 1. Design of the polystyrol vessels used for the experiments on choice of food (18x 18• 15 mm). A = Food A; B = Food B; P = Plaster of Paris-Charcoal Mixture
tive air humidi ty at 100 % and by diffuse i l lumination, humidi ty gradients a nd phototaxis were be ruled out as factors inf luencing the choice behaviour.
The green algae Blidingia minima (N~ig. ex Kfitz.) Kylin, Enteromorpha intestinafis and Rhizoclonium fipaxium (Roth) Harv. as well as the l ichen Caloplaca marina were compared, using 12 vessels per comparison each with 10 females. To take into account possible differences in the behaviour of instars and sexes, the distr ibution of females, males and tr i tonymphs (each 3 x 10) after a period of 24 hours was compared in an addit ional pairwise-choice test with Blidingia and Enteromorpha. In order to study indi- vidual behaviour, 17 females were each placed in the centre of a vessel provided with Blidingia minima and F.nteromorpha intestinalis and observed cont inuously for 6 hours. The intensi ty of feeding was checked by count ing the faecal pellets per area after 24 hours.

Feeding habits of HFadesia fusca 163
T h e ef fec t of d i e t s o n morta l i ty , r ep roduc t iv i ty , a n d d e v e l o p m e n t
After sampling and storage for one day in the light thermostat without food, 20 fe- males per diet were each placed in a feeding vessel of their own and checked every second or third day. New-born larvae were counted and removed at every control. Some of these larvae were transfered to feeding vessels with the same diet, and their further development was checked daily or every second day. Non-feeding mites and specimen that died owing to external factors (e.g. injury) were ignored for data evaluation.
The period of time be tween the first and last appearance of new-born larvae was de- f ined as durat ion of reproduction. The m e a n birth rate for each female is the quot ient of the duration of reproduct ion and the total n u m b e r of larvae. In the case of females with- out offspring be tween start of invest igat ion and death but who posthumously were found with fully developed embryos within their idiosoma (checked by squeezed preparations), the duration of reproduct ion and the birth rate was rated as zero.
Forage used for the food quality experiments was collected at different locations in the Weser estuary; the l ichen Caloplaca marina from the supralittoral zone a nd the Chlo- rophytes Prasiola stipitata Suhr in Jessen from the supralittoral fringe, Blidingia and Rhizoclonium riparium from the upper intertidal and Enteromorpha intestinalis from the middle inter t idal .
S ta t i s t ics
The statistical t rea tment of the preference exper iment is based on the assumption that without any attraction or repulsion of the two food sources the individuals would be distributed homogeneous ly on the vessels bottom. For simplification, the few spe- c imen found on the vessels walls were added to those on the uncovered plaster area. A chi-square test was carried out to compare the expected and observed frequencies of spe-
cimens cumulated for the three areas of the 12 vessels (Ho: EFood A = EFood B = Eplaster/WaUs =
1/3, n = 120, df=2, p < 0.01). If this test indicated an inhomogeneous distribution, the "more attractive" food area was compared to the remain ing area by a chi-square test for
H o : EFood A = 1/3, EFood B § Plaster/Walls = 2/3 (n = 120, df = 1, p < 0.01). The low sample size for the exper iments on food quali ty implied that no rel iable test
on normal distr ibution could be carried out. Therefore, the effect of different diets on mor- tality, reproduction and deve lopment was compared us ing the two-tailed non parametr ic U-test after M a n n & Whi tney (p < 0.05). A possible connect ion be tween long reproduc- tion time and low birth rate was tested us ing the Spearman coefficient of r ank correla- tion. (p for [HI: R r 0] < 0.05).
All statistical tests men t ioned above were calculated according to K6hler et al. (1992).
RESULTS
Die t r a n g e
Table 1 shows the results of the feeding tests with mar ine and habi ta t -ext raneous food.
Except for Fucus vesiculosus and Ulva lactuca L. (which have comparatively firm tissue), all other marine macroalgae, microalgae, lichens and detritus were accepted as

164 J. Bricking
Table 1. Laboratory results of supply experiments with various kinds of food (+ accepted, - not accepted. * only adults tested)
Food source Sample site Result
Marine plants (Weser estuary) Xanthoria parietina (L.) (Lichenophyta) Caloplaca marina (Lichenophyta) Different coccal and filamentous Cyanophyta Prasiola stipitata (Chlorophyta) Blidingia minima (Chlorophyta) Rhizoclonium riparium (Chlorophyta) Detritus floccules (inclusive different microphytes) gnteromorpha intestinalis (Chlorophyta) Rhodochorton sp. (Rhodophyta) with epiphytic diatoms Epiphytic microalgae on Fucus vesiculosus Young Fucus vesiculosus (without epiphytes) (Phaeophyta} Ulva lactuca (Chlorophyta)
Fresh and decayed carrion (Weser estuary) Anurida maritima (Guerin) (Collembola) Ligia oceanica (L.) ([sopoda)
Habitat-extraneous food Ulothrix sp. (Chlorophyta) Pleurococcus sp. (Chlorophyta) Myriophyllum spicatum (L.) (Rosidae) Elodea canadensis (Michx.) (Alismatidae) Sphagnum sp. (Bryophyta) Birch leaves in various degrees of decomposition Laccaria laccata (Skop. ex. Fr.) Berk ex Br. Mycelium (Fungi)
upper supralittoral + lower supralittoral + supralittoral fringe + supralittoral fringe + upper intertidal + upper intertidal + middle intertidal +' middle intertidal + lower intertidal + lower intertidal + * lower intertidal -* lower intertidal -*
upper intertidal -* upper intertidal -"
limnic sublittoral + tree bark -* pond -* pond -" peat-bog - ' edge of a forest -* laboratory culture -*
food. Fresh and decaying carcasses were neve r accepted, even when the spec imen were exposed to longer periods of starvation. Habi ta t -ext raneous food was not accepted , with
one except ion i.e. a limnic species of epilithic growing Ulothrix sp., a sublittoral f i lament- ous g reen alga sampled at a bank re inforcement of the I jsselmeer (0.5 PSU, Nether-
lands). Many species of this genus are also present on marine and es tuar ine hardsub- strates.
The results of these laboratory tests were compared to results of analyses of faecal pellets obta ined from mites sampled in the field (Table 2).
The food is obviously only partially d iges ted by Hyadesia. With Enteromorpha as a
food component , cell wall fragments, parts of chloroplasts, pyrenoids and, in few cases,
also intact cells occurred in the faecal pellets. With Blidingia as food, addit ional ly frag- ments of thalli consisting of up to 10 cells were found. Fine fi lamentous g reen a lgae were
mere ly f ragmented into smaller parts as a result of food processing (Fig. 2). Diatoms were
de tec ted ei ther as intact individuals, empty shells or f ragments of shells. Most of the Cyanophytes were found intact e i ther as dus te r s of coccal species (e.g. Chroococcus) or
as f ragments of f i lamentous species (e.g. Oscillatoria). Many of the a lgae f ragments
found in the faecal pellets showed pr imary f luorescence comparable to that of l iving

F e e d i n g habi t s of Hyadesia fusca 165
Table 2. Food components in faecal pellets of Hyadesia fusca adults, sampled in various littoral and vegetational zones
Sample site Food components (in order of decreasing estimated proportion)
Littoral fringe (sandstone), dominated by tetrasporal Cyanophytes ( Chroococcus s p.)
Upper intertidal (concrete), dominated by Bfidingia sp.
Upper intertidal (sandstone), in the upper part dominated by Blidingia sp., in the lower part dominated by Enteromorpha sp.
Middle intertidal (hardened sediment layer) with diatoms and few filamentous Rhodophyta
Cyanophytes, Blidingla sp., epiphytic diatoms, coccal Chlorophytes, fungal hyphae
Blidingia sp., diatoms, different Cyanophytes, filamentous Chlorophytes
Btidingia sp., Cyanophytes, Enteromorpha sp., benthic diatoms, filamentous Chlorophytes, sediment particles
diatoms
Fig. 2. A: Filamentous green alga, sampled at the ferry landing place Dedesdorf, Weser estuary; B: content of faecal pellets of an adult fed with this alga. Bar = 30 pm

166 J. Bricking
algae. Faecal pellets left on moist plaster for some days often gave rise to a growth of green algae germlings, which penet ra ted the peritrophic membrane .
The analysis of samples taken from the natural habitat confirmed largely the results of the feeding tests. Quanti ty and f requency of the food components from adults of a dis- tinct vegetat ion zone represented to some extent the respective food supply at the sam- ple place. As an exception, the faecal pellets produced by mites sampled in the supralit- total fringe regularly contained Blidingia cells, though this alga occurred in this zone only as patches of few germlings. The occasional appearance of benthic diatoms and of sedi- men t particles within the faecal pellets indicate that detritus is, albeit to a small extent, a part of the food ingested.
F o o d p r e f e r e n c e
Figure 3 shows the results of the pairwise-choice test with 10 females per vessel. A significant inhomogenous distribution of the test populat ion was recorded in all tests, ex- cept in the trial with Rhi zoc lon ium - Caloplaca. The mites showed a strong preference for Blidingia (X 2 > 100). Furthermore, they were more a b u n d a n t on R h i z o c l o n i u m if compa- red with E n t e r o m o r p h a + Plaster/Walls (X 2 = 11), and on E n t e r o m o r p h a if compared with
Fig. 3. Distribution of the test populations in the vessels in which a choice of food was offered. Box & Whisker Plot with median (bold line), interquartfl range (boxes) and range (whiskers); n = 12 with 10 individuals each. B = B1idingia, E = Enteromorpha, R = Rhizoclonium, C = Caloplaca,
P/W = Plaster and Walls

F e e d i n g h a b i t s of Hyadesia fusca 167
Caloplaca + P l a s t e r / W a l l s (X 2 = 14). T h e s e t w o " p r e f e r e n c e s " h a v e to b e t r e a t e d w i t h c a u -
t i o n r e g a r d i n g t h e r e l a t i v e l y h i g h v a r i a b i l i t y (Fig. 3). W i t h t h e e x c e p t i o n of s o m e t r i a l s
w i t h Blidingia s u p p l y , f a e c a l p e l l e t s w e r e f o u n d o n b o t h f o o d a r e a s of t h e v e s s e l s .
T h e d i s t r i b u t i o n of t h e 30 f e m a l e s , m a l e s , a n d t r i t o n y m p h s w a s 24, 27 a n d 26 i n d i v i -
d u a l s , r e s p e c t i v e l y , o n t h e Blidingia a r e a ( c o m p a r e d w i t h Enteromorpha), t h u s s i g n i f i c a n t
d i f f e r e n c e s i n t h e c h o i c e b e h a v i o u r of s e x e s o r i n s t a r s d i d n o t o c c u r .
D u r i n g 6 h of c o n t i n u o u s o b s e r v a t i o n , o n l y 4 of 17 i n d i v i d u a l l y k e p t f e m a l e s m o v e d
to t h e Enteromorpha a r e a , a n d o n l y o n e of t h e s e f e d o n t h i s a l g a , b u t w i t h i n t h e f i r s t h o u r
a l l m i g r a t e d o v e r to t h e Blidingia a r e a . T h e m e a n n u m b e r of f a e c a l p e l l e t s p e r f e m a l e a n d
d a y w a s 18 ( m a x . 36) o n t h e Blidingia a r e a , a n d 1 ( m a x . 13) o n t h e Enteromorpha a r e a .
F o o d q u a l i t y
D i f f e r e n c e s i n t h e n u t r i t i o n a l q u a l i t y of c e r t a i n a l g a e o r l i c h e n s w e r e t e s t e d r e g a r d i n g
t h e i r d i e t a r y i n f l u e n c e o n s u r v i v a l t i m e a n d r e p r o d u c t i o n of f e m a l e s a n d o n t h e p o s t n a t a l
d e v e l o p m e n t of l a r v a e ( T a b l e 3). It m u s t b e n o t e d t h a t t h e f e m a l e s o r i g i n a t e d f r o m f i e l d
s a m p l e s , t h e r e f o r e i n d i v i d u a l a g e , n u m b e r of p r e v i o u s b i r t h s a n d t h e a c t u a l s t o c k of
Table 3. C o m p a r i s o n of var ious food sou rces wi th r e g a r d to the i r effect on survival , la rva l b i r ths a n d d e v e l o p m e n t t ime. Ar i thme t i c mean , s t a n d a r d devia t ion , r a n g e [in bracke ts ) a n d s a m p l e size s ta ted . Different cha r ac t e r s in a c o l u m n indica te s ign i f ican t d i f f e rences (U-test af ter M a n n & Whi tney ,
p < 0.05)
Food Surv iva l Dura t ion of Total n u m b e r Ind iv idua l bir th D e v e l o p m e n t sou rce t ime (d) r eproduc t ion of l a rvae pe r ra te ( larvae pe r t ime (birth to
(d) f e m a l e f e m a l e x d -I) adul t ) (d)
Blidingia
Enterom.
Rh~od.
Prasiola
Caloplaca
U n f e d
79 • 39 a 45 • 17 a 26 • 10 a 0.60 • 0.14 a 44 • 7 a (21 - 136) (7 - 69) (5 - 38) (0.38 - 0.90) (33 - 58)
16 15 15 15 9
2 8 • 1 0 • 7 • 0 . 7 5 • 5 0 • 1 0 a , b (6 - 101) (3 - 26) (3 - 14) (0.29 - 1.75) (39 - 60)
19 17 17 17 3
40 • 25 b 17 • 15 b 9 • 7 b 0.72 • 0.35 a 47 • 8 a ,b (10 - 9 1 ) ( 4 - 4 6 ) ( 3 - 30) (0.17 - 1.50) ( 3 6 - 56)
19 17 17 17 4
28 • 22 b,c 15 • 23 b,c 7 • 8 b,c 0.55 • 0.43 a ,b 101 • 37 b (10 - 87) (0 - 67) (0 - 25) (0.00 - 1.50) (52 - 140)
18 18 18 18 4
34 + 22 b,c 19 • 16 b 6 • 4 b,c 0.43 • 0.31 b 73 • 29 a ,b (5 - 70) (l - 47) (t - 14) (0.10 - 1.25) (53 - 94)
16 14 14 14 2
1 7 + 6 c 4 + 3 c 4 + 3 c 0.80 • 0.68 a,b (10 - 37) (0 - 9) (0 - 12) (0.00 - 2.25)
20 17 17 17

168 J. Bricking
sperm within the receptaculum seminis were unknown. The values given below can be used to show relative differences in the effect of various diets, but do not fully represent the species-specific mean of lifetime, reproduction period or fecundity.
In the following text, differences of m e a n values as shown in Table 3 are pointed out in context with the deve lopment in time.
M o r t a l i t y
Compared to all other diets, a Blidingia diet ensured significantly longer survival. No significant differences were ascer ta ined in the other feeding tests. Compared to
unfed individuals, only those fed on algae had a significantly higher mean survival time. Figure 4 shows the relative portion of survivors in the period after the observat ion began. Every dot in the figure represents a day on which the death of one or more mite(s) was ascertained.
Corresponding data on Blidingia and Rhizoclonium as a food source exist also for adults reared in the laboratory from larvae. The mean survival time was 100 days (SD = 51, range 29-235, n = 15) on Blidingia and 54 days (SD = 48, range 13-129, n = 6) if fed with Rhizoclonium. Even though these values tend to be higher than those for field adults of u n k n o w n age (differences not significant), they show the same relation of ca. 2 : I.
R e p r o d u c t i o n
The durat ion of reproduction and the n u m b e r of births of females fed with Blidingia was about twice as high compared to those of females fed with other diets. Both para- meters are coupled with the individual survival time of the respective females. The birth rate obta ined with the feeding of the various green algae ranged from 0.55 to 0.75 larvae per female/day (no significant differences, see Table 3). The birth rate of females fed with the lichen Caloplaca was significantly lower than that of females fed on other diets ex- cept for Prasiola. While the birth rate of the mites kept on Blidingia varied relatively little, the respective data of mites fed with other food had a high range. The unfed fema- les differed from the fed ones in their very short reproduction period with f requent larvae births, which explains the unexpectedly high birth rate.
Looking at the deve lopment of the birth rate in the course of the experiment , it could be seen that with Blidingia as food larvae births occurred regularly dur ing the entire re- productive period. With all other diets, except for the short-living females with Entero- morpha supply, at the very latest after three weeks of the diets longer periods of up to 29 days (Rhizoclonium) with no larvae product ion occurred. Figure 5 shows such interrup- tions of births comparing Blidingia, Prasiola, and Caloplaca.
A significant negat ive correlation be tween reproduction period and birth rate was found for the females fed with Rhizoclonium, Prasiola, and Caloplaca (R= -0.72 to -0.78, n = 14-17).
D e v e l o p m e n t
Regarding the time period from birth to adult hatch (Table 3), deve lopment was re- tarded in females on a diet of Prasiola and of Caloplaca. Figure 6 shows the deve lopment of individuals for all diets tested (total lifetime of adults is not stated).
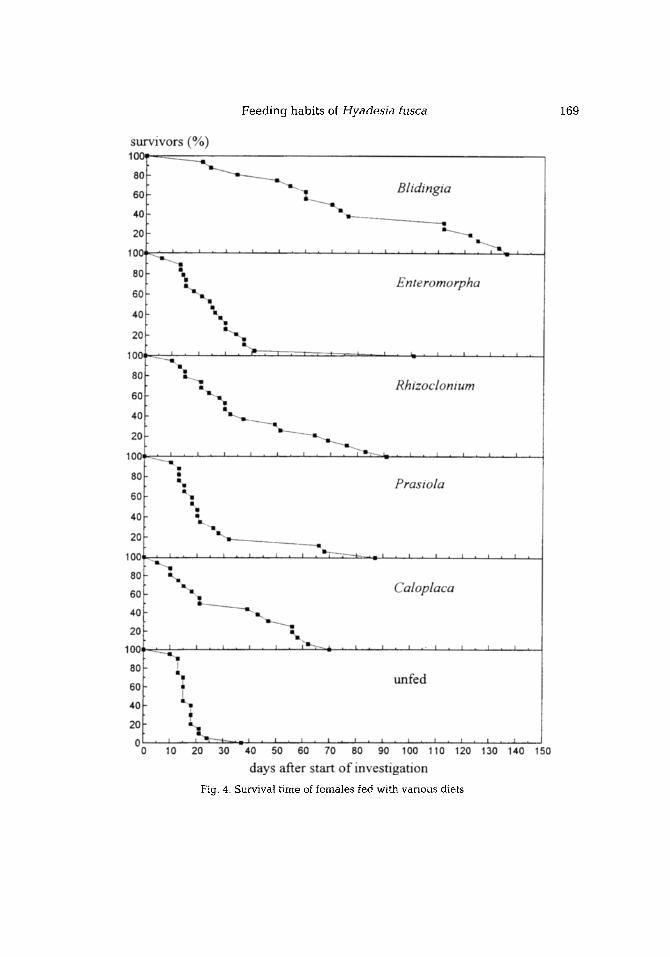
Feeding habits of Hyadesia [usca 169
Fig. 4. Survival time of females fed with various diets

170 J. Br icking
Fig. 5. Larvae births (cumulative) after start of observation of the respective five females with the longest duratmn of reproduction. The arrows point out the overall survival time of the female (with-
out further larvae births}
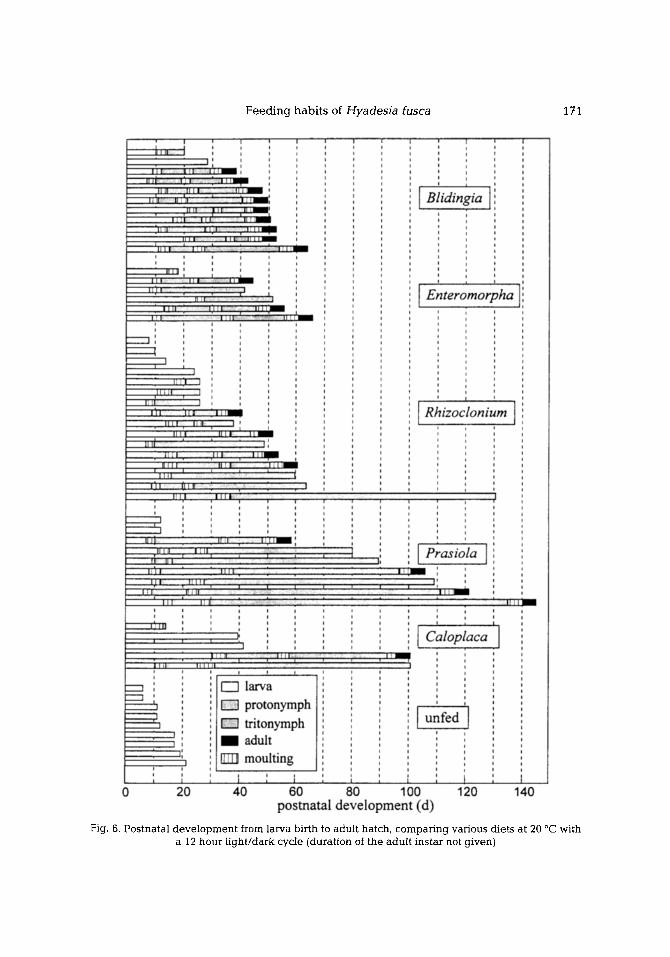
F e e d i n g h a b i t s of Hyadesia fusca 171
Fig. 6. Postnatal development from larva birth to adult hatch, comparing various diets at 20 ~ with a 12 hour light/dark cycle (duration of the adult instar not given)

172 J. Bricking
The various diets differed in their effect on the rearing success. About 80 % of the offspring reared on Bfidingia finally hatched to adults; with other food supplies it was only 20 to 50 %. Unfed larvae showed no further development . The durat ion of deve lopment with different diets showed a high variability, both within groups and be tween groups. While one new-born larva reached adul thood in only 33 days (Blidingia), another took 140 days (Prasiola). The above ment ioned retardat ion of development can obviously oc- cur in all ontogenet ical instars; conspicuous are the prolongation of the protonymphal in- star if fed with Enteromorpha and the prolongat ion of the t r i tonymphal instar ff fed with Prasiola. In some cases, deve lopment stopped though death occurred much later (up to three months). The frequency of such deve lopment retardations was low if reared on Bli- dingia and high with other food supply.
DISCUSSION
F e e d i n g h a b i t s a n d food q u a l i t y
This invest igat ion shows that Hyadesia fusca has a substantial ly broader diet range than was formerly suspected. In Table 4 an overview of the updated results of gut con- tent or faecal pellet analysis carried out in species of the family Hyadesi idae is given.
All Hyadesiids appear to be microphytophagous (in the sense of Schuster, 1956), but some also accept detritus as food. However, it can be assumed that the actual spectrum of food of the species listed in Table 4 is much broader.
The diet range of the observed populat ion is probably more or less d e p e n d e n t on the accessibility of suitable food at the respective sample site. However, food ingest ion does not automatically imply food utilisation. Judg ing by the observations that (1) H. fusca has a clear preference for a distinct green alga with high nutri t ional value and (2) green al- gae constitute a food component of near ly all invest igated Hyadesiids, one may assume that this taxon is mainly algivorous. Nevertheless, the role played by the epiphytic micro- flora of macroalgae in the nutr i t ion of Hyadesi ids is still unknown. However, the bacte- ria-rich detritus constitutes only a small part of the food of H. fusca. A comparison of the nutri t ional quali ty of food samples from the field used in this invest igat ion with that of axenic algae cultures could, perhaps, clarify the issue.
Higher plants and macroalgae with relatively firm tissues e.g. Fucus and Ulva were not accepted in the feeding tests. Tissue structure and therefore the ease with which the mites can manipula te and ingest their food is probably an important factor that influen- ces the diet range of H. fusca. This assumpt ion is further supported by the f inding that Enteromorpha intestinalis, which is closely related to Blidingia minima but has greater cell lumina (12 • 12-20 • 15 pm versus 5-7 pm diameter) and thicker cell wails (Bhding, 1963), is a low-quali ty food. Furthermore, in the course of the supply exper iments the n u m b e r of faecal pellets produced offering Enteromorpha as food was very low compared to that of Blidingia-fed test animals. Schulte (1976) assumed for certain a lgophagous mites of the genus Ameronothrus that the instar specific diet range correlates with the size of the cheticerae and depends mainly on tissue firmness and structure, bu t not on chemical composition of the algae. Larvae and protonymphs accepted only coccal algae and fungi, whereas deutonymphs, t r i tonymphs and adults fed also on macroalgae or lichens. Siemer (in press) found that the t iny larvae of the halacarid mite Metarhom-

F e e d i n g h a b i t s of Hyadesia fusca 173
Table 4. Gut content and/or faecal pellet analyses of certain species of Hyadesi idae
Species Reference Sample site Gut/faecal pellet contents
Hyadesia fusca (Lohmann, 1894)
Ganning, 1970 supralittoral rock pools, North sea and Baltic sea
Ganning, 197 l dito
B~nard, 1961" barnacles, Northwest- France (Cotentin, Bretagne)
this estuarine intertidal investigation and lower supralittoral
of embankments (Northern Germany)
Hyadesia tumida B~nard, 1961 Benard, 1961
Hyadesia sellai Schuster, 1979 Viets, 1937
Amhyadesia Fain & Schuster, costaricensis Fain 1984b & Schuster, 1984
Amhyadesia Fain & Schuster, brasiliensis Fain & 1984b Schuster, 1984
Amhyadesia Fain & Schuster, heterophallus Fain 1984a & Schuster, t984
Hyadesia Fain & Schuster, verrucosa Fain & 1985 Schuster, 1985
Hyadesia Trav6, 1988 kerguelensis Lohmann, 1907
Hyadesia zelandica Luxton, 1989 Luxton, 1989
barnacles, Northwest- France (Cotentin, Bretagne)
intertidal rocks and inside the cavity system of the Lithophyllum- trottoir, Italian adria
rocks of the upper intertidal (Costa Rica)
Enteromorpha intestinalis, possibly epiphytic bacter ia
Cladophora glomerata,
mucous layer on barnacles with unicellular algae, bacteria and diatoms
different macroalgae (Ulvales, Prasiolales, Cladophorales), microalgae (Cyanophytes, diatoms, unicellular green algae), detritus (incl. its microflora). In the labora- tory also Rhodophyta and lichens
mucous layer on barnacles with unicellular green algae, bacteria and diatoms
predominant ly g reen algae, few Cyanophytes
green algae (among others)
rocks of the upper intertidal with sessile fauna or epilithic algae turf (Brasil)
intertidal rocks, partly with calcareous algae (Maledives and Philippines)
in depressions and crevices of intertidal rocks, covered with green algae (Istria, former Yugoslavia)
in the Verrucana-belt of the intertidal (Kerguelen islands)
on rocks of the upper intertidal among barnacles, in fissures of encrustated oysters, on lichens and in other places (New Zealand)
leftover of green algae thalli, Cyanophyta, seldom diatoms
presumably, g reen algae (green gut content)
presumably, green algae (green gut content and green faecal pellets)
Verrucarfa sp. (lichens, only the phycobiont)
detritus
�9 B~nard (1961) descr ibed this for F/. [urcilfipes B~nard, 1961, which is according to Fain (1981) a synonym of H. fusca (Lohmann, 1894).

174 J. Bricking
bognathus armatus (Lohmann) cannot p roceed with their pos tembryonic d e v e l o p m e n t if kep t on Enteromorpha as opposed to those kep t on Blidingia. However, f ield adul ts had the same longevi ty on both algae.
The spec t rum of accep t ed food at different localities may be inf luenced by the fact that the shape of thall i and th ickness of cell wall of certain a lgae species m a y vary grea t ly under different abiotic condi t ions (e.g. Bliding, 1963). Young et al. (1987) have shown that Enteromorpha intestinalis from es tuar ine locations has th inner cell walls c o m p a r e d to the same a lga from fully mar ine habitats . On the other hand, the low nutr i t ional qual i ty of cer tain f i lamentous or thin monos t romat ic algae, e.g. Rhizodonium and Prasiola for H. fusca (this observat ion) or Prasiola for M. armatus (Siemer, in press), is e v i d e n c e that a firm tissue cannot be the only reason for a low suitability.
Other poss ible factors inf luencing the suitabil i ty or accep tance of food could be (1) the ene rgy and wa te r content; (2) the ca rbohydra t e and nutr ient composi t ion (e.g. starch, lipid, potyol, amino acids and vi tamin content); (3) the relat ive proport ion of indiges t ib le components and (4) poss ible mechan i sms of the a lgae that protect from be ing fed on. Levinson et al. (1992) r epor ted the s ignif icant inf luence of various v i tamins on lifetime, fecundity, and d e v e l o p m e n t in the flour mite Acarus sire L. (Astigmata: Acar idae) . Bow- man & Childs (1982) found amylase and chi t inase in all of the six inves t iga ted As t igmat id mite species, and cel lulase in two of them. However , the possible role of endosymbion t s and their enzymes in the d iges t ion of cell walls, well known in insects, is not fully un- ders tood in the case of mites.
One possible mechan i sm for a lgae to protect themselves from grazers is chemica l de- fence (Paul, 1992), e.g. the product ion of toxic phenols and tannins as de sc r ibed for brown a lgae (Carefoet, 1977; Roussis et al., 1993; Winter & Estes, 1992). Many g r e e n algae, e.g. Blidingia, Enteromorpha and Ulva, produce a g rea t amount of Dimethylsu l foniumpro- p ionate (DMSP) (Reed, 1983), which is d i scussed to have protec t ing features , poss ib ly due to the re lease of acrylic acid (Glombitzka, 1970; Keller et al., 1989). At leas t for H. fusca with its p re fe rence for Bfidingia the DMSP seems not to be a deterrent .
The exper imen t s on food pre fe rence revea led a search ing behav iour favour ing Bli- dingia, but the causal i ty of this behaviour, e.g. different palatabil i ty, a t t rac tan ts of the pre- fer red food, de te r ren ts or repe l len ts of the avo ided food, mechanica l s t imuli (size and shape of thalli), remains unclear. However , the p re fe rence behav iour of H. [usca is ob- viously cor re la ted to the sui tabi l i ty of the respect ive food. Contradic t ing this, Paine & Va- das (1969) conc luded that avai labi l i ty of seaweeds , and not absolute food value , was a more decis ive factor for the evolut ion of p re fe rences of mar ine herbivores . For example , Carefoot (1973) has shown for the supral i t tora l i sopod Ligia pallasii (Brandt) that feed ing pre fe rence is not necessar i ly l inked to the ene rgy content of the food. On the o ther hand, in some cases food that was in short supp ly or totally absen t in the na tura l hab i t a t of cer- tain mar ine herb ivores was p re fe r red in labora tory tests (Leighton & Boolootian, 1963; Carefoot, 1970, 1973).
T h e i n f l u e n c e of f e e d i n g h a b i t s o n d i s t r i b u t i o n p a t t e r n s
A migra t ion to surfaces covered with Blidingia and a low mortal i ty of juven i l es and adul ts - a c c o m p a n i e d by a h ighe r fecundi ty - would l ead to h igher popu la t ion densi t ies . Evidence that p re fe rence behav iou r and food qual i ty could affect d is t r ibut ion pa t te rns

Feeding habits of Hyadesia fusca 175
and populat ion density is given by the fact that a higher abundance of Hyadesia fusca was found on surfaces and in the littoral zone dominated by Bfidingia (Bricking, in press). Food supply is of course only one of the many interact ing factors inf luencing distribution limits and populat ion dynamics. Carefoot (1977, pp. 46-47) remarked, that "The distribution of an organism represents the best compromise be tween its physiological and space require- ments, and the conditions existing in the habitat". In particular the following must be taken into account: (1) al ternating submersion and emersion; (2) high fluctuation of humi- dity, salinity and temperature - increasing with littoral height; (3) surface structure and composition - inclusive vegetat ion and sessile animals;(4) mechanical stress due to wave action, brushing action of algae and sediment-media ted abrasion and (5) interspecific competit ion (cf. Lewis, 1964; Little & Kitching, 1996). Nevertheless, in addition to the great risk of desiccation in the supralittoral zone, the abundance of low-quality food (lichens) would render impossible a bui ld-up of a stable H. fusca population. Even the low nutri- tional quality of algae present in the middle and lower intertidal, e.g. Enteromorpha and red algae, may account for the low populat ion density in this littoral region.
The low species diversity and a b u n d a n c e of mites of terrestrial origin in lower litto- ral zones are thought by many authors to be mainly the result of a restricted tolerance to submersion, a reduced time for foraging and - as previously ment ioned - mechanical im- pact (Pugh & King, 1985b; Schulte, 1977; Schuster, 1979). In view of the fact that (1) H. fusca is a common inhabi tant of Baltic rock pools (Ganning, 1970, 1971; (Dstman & R6nn- berg, 1991) and that (2) specimens of the invest igated intert idal population can live, feed and reproduce totally immersed for several months in the laboratory (Bricking, unpub- lished), foraging and inundat ion time can explain nei ther the absence of this species in the lower intert idal nor their low populat ion densi ty in the middle intertidal of the Weser estuary. The longer time of exposure to wave action and water currents in the lower inter- tidal zones might be an addit ional factor causing lower populat ion densities. This view is supported by results of Ostman & R6nnberg (1991) who observed significantly lower abundances of H. fusca in rock pools subject to regular wave action caused by ships com- pared to undis tu rbed reference pools. According to Pugh & King (1985b), the distribution of Hyadesiids on British rocky shores depends on the prevalence of barnacles which offer protection against wave action. However, both laboratory experiments on the ability of littoral mites to withstand mechanica l stress and measurements of water pres- sure within the microhabitat of the mites, e.g. inside the dense algae cover or within cavi- ties of the rock surface, have yet to be carried out.
Acknowledgements. The author thanks G. Davids for checking the translation and Prof. H. Witte for reading the manuscript.
LITERATURE CITED
Alberti, G., 1973. Ern~hrungsbiologie und SpinnvermSgen der Schnabelmilben (Bdelhdae, Trombi- dfformes). - Z. Morph. Tiere 76, 285-338.
Bartsch, I., 1978. Verbreitung der Halacaridae (Acari) im Gezeitenbereich der Bretagne Kiiste, eine 6kologische Analyse. I. Verbreitung der Halacanden. -Cah. Biol. mar. 19, 363-383.
Bartsch, I., 1982. Halacaridae (Acari) yon der Atlantikkiiste des borealen Nordamerikas. Okotogi- sche und tiergeographische Faunenanalyse. - Helgol/inder Meeresunters. 35, 13-46.
B6nard, F., 1961. Sur deux nouvelles esp~ces d'Acariens matins, Hyadesia tumida et Hyadesia fur- cillipes: sous-ordre des Sarcoptiformes, famille des Hyadesidae. - Cah. Biol. mar. 2, 71-96.

176 J. Bricking
Bliding, C., 1963. A critical survey of European taxa in Ulvales. I.: Capsosiphon, Percursaria, Blidin- gia, Enteromorpha. - Op. bot. Soc. bot. Lund 8, 1-160.
Bowman, C. E. & Childs, M., 1982. Polysaccharidases in astigmatid mites (Arthropoda; Acari). - Comp. Biochem. Physiol. 72B, 551-557.
Bricking, J., 1995. Untersuchungen zur Populations- und ErnShrungsbiologie yon Hyadesia [usca (Lohmann, 1894) (Astigmata: Hyadesiidae), einer phytophagen Milbe des 8stuarinen und mari- nen Felslitorals. Diss., Univ. Bremen, 172 pp.
Biicking, J., 1998. The population biology of a phytophagous mite of marine and estuarine rocky shores (Astigmata, Hyadesiidae). In: Evolution and ecology of Acari. Ed. by J. Bruin, L. P. S. van der Geest & M. W. Sabelis. Kluwer, Dordrecht (in press).
Carefoot, T.. 1970. A comparison of absorption and utilization of food energy in two species of tro- pical Aplysia. - J. exp. mar. Biol. Ecol. 5, 47-62.
Carefoot, T., 1973. Feeding, food preference, and the uptake of food energy by the supralittoral iso- pod Ligia patlasii. - Mar. Biol. 18, 228-236.
Carefoot, T., 1977. Pacific seashores: A guide to intertidal ecology. Univ. of Washington Press, Seattle, 208 pp.
Ernst, H., 1995. Raum-Zeit-Nutzung litoraler Milben terrestrischer Herkunft auf artifiziellen Hart~ substraten norddeutscher Astuare. Diss., Univ. Bremen, 205 pp.
Ernst, H., 1996. The distribution and zonation of terrestrial mites inhabiting artificial rocky shores of estuaries in northern Germany. In: Acarology IX. Ed. by R. Mitchell, D. J. Horn, G. R. Needham & W. C. Welbourn. Ohio Biological Survey, Columbus, Ohio I, 529-532.
Ernst, H., Siemer, F., Bricking, J. & Witte, H., 1993. Die litorale Milbenzbnose auf Uferbefestigungen des Weser~stuars in AbhSngigkeit von Substrat und Salzgehaltsgradient. - Inf. Natursch. Land- schaftspfl. 6, 401-416.
Fain, A., 1981. Notes on the Hyadesiidae Halbert, 1915 and Algophagidae Fain, 1974, nov. tax. (Acari, Astigmata) with a redescription of Hyadesia currasaviensis Viets, 1936 and H. sellai Viets. 1937. - Acarologia 22, 47-61.
Fain, A. & Schuster, R., 1984a. Four new species of the genus Amhyades ia Fain and Ganning, 1979 from the intertidal zone of several Indo-Pacific islands (Acari: Hyadesiidae). - Int. J. Acarol. I0, 103-111.
Fain, A. & Schuster, R., 1984b. Two new species of mites of the family Hyadesiidae (Acari, Astig- mata) from Costa-Rican and Brasilian Coasts. -Acarologia 25, 271-279.
Fain, A. & Schuster, R., 1985. New observations on the Hyadesiidae (Acari, Astigmata). Description of three new species of the genus Hyadesia Megnin, 1891. - Acarologia 26, 67-77.
Fain, A. & Schuster, R., 1989. Geographical distribution of the intertidal mite Hyadesia (Hyadesia) [usca (Lohmann, 1894) (Acari, Hyadesiidae). - Bull. Annls Soc. r. ent. Beige 125, 293-295.
Ganning, B., 1970. Population dynamics and salinity tolerance of Hyadesia fusca (Lohmann) (Aca- rina, Sarcoptiformes) from brackish water rockpools, with notes on the microenvironment inside Enteromorpha tubes. - Oecologia 5, 127-137.
Ganning, B., I971. Studies on chemical, physical and biological conditions in Swedish rockpool eco- sys t ems . - Ophelia 9, 51-105.
Gerlach, D., 1984. Botanische Mikrotechnik. Thieme, Stuttgart, 311 pp. Glombitzka, K.-K., 1970. Antimikrobielle Inhaltsstoffe in Algen. 2. Mitt.: Das Vorkommen yon Acryl-
sSure in verschiedenen Meeresalgen. - Planta reed. 18, 210-221. Haase, R., 1995. Okophysiologische Untersuchungen zur Dimethylsulfid-Emission der eulitoralen
Griinalge Enteromorpha intestinafis (L.) Link. Diss., Univ. Bremen, 210 pp. Halbert, J. N., 1920. The acarina of the seashore. - Proc. R. Ir. Acad. 35B, 106-155. Keller, M. D., Bellows, W. K. & Guillard, R. R. L., 1989. Dimethylsulfide production and marine phyto-
plankton: an additional impact of unusual blooms. - Coast. estuar. Stud. 35, 72-79. K6hler, W., Schachtel, G. & Voleske, P., 1992. Biostatistik. Springer, Berlin, 255 pp. Kronberg, I., 1983. Okologie der schwarzen Zone im marinen Felslitoral. Diss., Christian-Albrechts-
Univ., Kiel, 238 pp. Leighton, D. L. & Boolootian, R. A., 1963. Diet and growth in the black abalone, Haliotis crachero-
dii. - Ecology 44, 227-238. Levinson, H. Z., Levinson, A. R. & Offenberger, M., [992. Effect of dietary antagonists and corre-

F e e d i n g h a b i t s of Hyadesia fusca 177
s p o n d i n g n u t r i e n t s on g r o w t h a n d r ep roduc t ion of the f lour mi te (Acarus siro L.). - Expe r i en t i a 48, 721-729.
Lewis, J. R., 1964. T h e eco logy of rocky shores . T h e Eng l i sh Univ. Press, London, 323 pp. Little, C. & Ki tching, J.A., 1996. T he biology of rocky shores . Oxford Univ. Press, Oxford, 240 pp. Luxton, M., 1989. Mi tes of t he family H y a d e s i i d a e (Acari: As t igma ta ) f rom N e w Zea l and . - Zool. J.
Linn. Soc. 95, 71-95. C)stman, M. & R 6 n n b e r g , O., 1991. Effects of sh ips ' w a v e s on rock-pools in the ,Zkland a rch ipe lago ,
n o r t h e r n Baltic Sea. - Sars ia 76, 125-132. Otto, G., 1936. Die F a u n a de r E n t e r o m o r p h a z o n e der Kieler Bucht . - Kieler Mee re s fo r sch . 1, 1-48. Paine, R. T. & Vadas , R. L., 1969. Caloric va l ue s of ben th i c m a r i n e a l g a e and thei r p o s t u l a t e d rela-
t ion to i n v e r t e b r a t e food p re fe rence . - Mar. Biol. 4, 79-86. Paul , V. J., 1992. S e a w e e d chemi ca l d e f e n s e s on coral reefs. In: Ecological roles of m a r i n e na tu ra l
p roduc t s . Ed. by V. J. Paul. C o m s t o c k Publ. Ass., London, 24-50. Pugh . P. J. A. & King, P. E., 1985a. F e e d i n g in in ter t ida l Acari. - J. exp. mar. Biol. Ecol. 94, 269-280. Pugh . P. J. A. & King, P. E., 1985b. T h e ver t ical d is t r ibut ion of the British inter t idal Acar i - T h e non
ha l aca r id f a u n a (Arachnida : Acari). - J. Zool., Lond. 207, 21-33. Pugh , P. J. A. & King, P. E., 1985c. Vertical d i s t r ibu t ion a n d subs t r a t e assoc ia t ion of the British Hala-
c a r i d a e . - J. nat . Hist. 19, 961-968. Reed, R. H., 1983. M e a s u r e m e n t a n d osmot ic s ign i f i cance of [3-DMSP in mar ine m a c r o a l g a e . - Mar.
Biol. 4, 173-181. Roussis , V., King, R. L. & Fenical , W., 1993. S e c o n d a r y me tabo l i t e chemis t ry of the Aus t r a l i an b r o w n
a lga Encyothalia cliftonii : e v i d e n c e for he rb ivore chemica l de fence . - P h y t o c h e m i s t r y 34, 107-111.
Schul te , G., 1976. Zur N a h r u n g s b i o l o g i e de r t e r r e s t r i schen u n d m a r i n e n Mi lbenfami l i e A m e r o n o - t h r idae (Acari, Oribatei) . - Pedobio log ia 16, 332-352.
Schul te , G., 1977. Die B i n d u n g de r L a n d a r t h r o p o d e n a n das Felsli toral der M e e r e u n d ihre Ursa- chen . Habi lschr . , Chr i s t i an -Albrech t s -Univ . Kiel., 223 pp.
Schuster , R., 1956. Der Ante i l der Or iba t iden an d e n Z e r s e t z u n g s v o r g ~ n g e n im Boden . - Z. Morphol . Oekol . Tiere 45, 1-33.
Schuster , R., 1965. Die Oko log ie de r t e r r e s t r i s chen Kle in fauna de s M e e r e s s t r a n d e s . - Verh. dt. zool. Ges . 1964, 492-521.
Schus ter , R., 1979. Soil m i t e s in the mar i ne e n v i r o n m e n t . In: Recen t a d v a n c e s in acarology. Ed by J. G. Rodr iguez . Acad. Press, N e w York i, 593-602.
Schuster , R., 1988. T r a n s o c e a n i c d is t r ibut ion of a i r -b rea th ing littoral mites . In: P rogress in acarology. Ed. by G. P. C h a n n a b a s a v a n n a & C. A. V i rak tanna th . Oxford & IBH Publ., N e w Dehl i i, 355-362.
Siemer, F., 1996. T h e dis t r ibut ion, zonat ion , p h e n o l o g y a n d life cycle of the H a l a c a r i d a e (Prostig- mata ) of artificial rocky sho res loca ted at the W ese r e s t u a r y (Germany) . In: Aca ro logy IX. Ed. by R. Mitchel l , D. J. Horn, G. R. N e e d h a m & W. C. Welbourn . Ohio Biological Survey, C o l u m b u s , Oh io I, 547-551 .
Siemer, F., 1998. T h e in f luence of va r ious k inds of food on the d is t r ibut ion a n d life cycle of Meta- rhombognathus arrnatus (Halacar idae: R h o m b o g n a t h i n a e ) . In: Evolut ion a n d eco logy of Acari . Ed. by J. Bruin, L. P. S. v a n der G e e s t & M. W. Sabel is . Kluwer, Dordrech t (in press) .
Trav6, J., 1988. Con t r ibu t ion & la c o n n a i s s a n c e de s a c a r i e n s l i t to reaux de l ' a rch ipe l de Kerque len : q u e l q u e s a s p e c t s des s t ra t6g ies adap t ives . - C N F R A (Com. Nat. Fr. Rech. Antarct . ) 58, 111-127.
Winter , F. C. & Estes , J. A., 1992. E x p e r i m e n t a l e v i d e n c e for t he effects of po lypheno l i c c o m p o u n d s f rom Dictyoneurum californicum R u p r e c h t (Phaeophy ta , Laminar ia les ) on f e e d i n g rate a n d g r o w t h in the r ed a b a l o n e Haliotus rufescens S w a i n s o n - J. exp. mar. Biol. Ecol. i55, 263-277.
Witte, H., 1972. F u n k t i o n s a n a t o m i s c h e u n d e t ho log i s che U n t e r s u c h u n g e n an de r Mi lben fami l i e E ry th rae idae . Diss., Chr i s t i an -Albrech t s -Univ . Kiel, 114 pp.
Young, A. J., Coll ins, J. C. & Russel , G., 1987. Ecotypic var ia t ion in the osmot ic r e s p o n s e s of Entero- morpha intestinalis (L.) Link. - J. exp. Bot. 38, 1309-1324.