infants as costly social tools in male barbary macaque networks
TRANSCRIPT

lable at ScienceDirect
Animal Behaviour 79 (2010) 1199e1204
Contents lists avai
Animal Behaviour
journal homepage: www.elsevier .com/locate/anbehav
Infants as costly social tools in male Barbary macaque networks
Stefanie Henkel a, Michael Heistermann b, Julia Fischer a,*aCognitive Ethology Laboratory, German Primate Center, GöttingenbReproductive Biology Unit, German Primate Center, Göttingen
a r t i c l e i n f o
Article history:Received 5 August 2009Initial acceptance 6 October 2009Final acceptance 28 January 2010Available online 1 April 2010MS. number: 09-00519R
Keywords:Barbary macaqueGC levelglucocorticoidgroomingMacaca sylvanusmaleeinfant interactionnonhuman primatesocial network analysis
* Correspondence: J. Fischer, Cognitive EthologyCenter, Kellnerweg 4, 37077 Göttingen, Germany.
E-mail address: [email protected] (J. Fischer).
0003-3472/$38.00 � 2010 The Association for the Studoi:10.1016/j.anbehav.2010.02.005
In mammals, females are the main caretakers of offspring. In a number of species, however, males alsocare for young, and may spend considerable time with infants. In various nonhuman primates, maleinteractions with infants appear to benefit mainly the males, who use them to regulate their relation-ships, particularly with other male group members. We studied whether interactions with infants wererelated to glucocorticoid output in male Barbary macaques, Macaca sylvanus, at the enclosure ‘La Forêtdes Singes’, France. We used a combination of behavioural observations, social network analysis andmeasures of faecal glucocorticoid metabolites as an indicator of physiological stress, and compared theresults for the birth season in spring, during a time of intense interactions with infants, with a period inautumn, when males interacted less frequently with infants. Social network analyses revealed that infantcarriers had stronger ties with other males than noncarriers. Overall, male relationships were stronger inspring than in autumn. Infant care was unevenly distributed among males and not related to rank,suggesting that the motivation to invest in infant carrying depends on the position in the social networkand the current constellation of relationships. Despite the correlation of infant carrying with maleinteractions, the time spent with infants alone explained most of the variation in glucocorticoid levels,suggesting that access to the maleemale network is paid for by an increase in stress hormone levels.Taken together, these results show that Barbary macaque infants can be viewed as costly social tools thatfacilitate networking among males.� 2010 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Owing to gestation and lactation, females bear the largest shareof the cost of reproduction in mammals (Clutton-Brock 1991). Ina number of mammal species, however, males also contribute tothe rearing of young. In cooperatively breeding tamarins, forinstance, males carry infants more frequently than mothers(Wright 1990; Schradin & Anzenberger 2001), while in coopera-tively breedingmeerkats, male groupmembers provide youngwithfood (Doolan & Macdonald 1999; Hodge 2007). In addition to foodprovisioning and carrying, males may defend young againstinfanticide (Pusey & Packer 1994; Palombit et al. 1997; Van Schaik &Kappeler 1997). The occurrence of male care is typically explainedby kin selection (Trivers 1972), but it has also been related to thedirect benefit of increasing one's chances of survival withincreasing group size (Clutton-Brock et al. 2001).
In nonhuman primates, interactions between males and infantsoccur in a number of species (reviewed in Paul et al. 2000),including baboons, Papio spp. (Smuts 1985; Palombit et al. 1997;Buchan et al. 2003) and Tibetan macaques, Macaca thibetana
Laboratory, German Primate
dy of Animal Behaviour. Published
(Zhao 1996). One of themost notable cases is, however, found in theBarbary macaque, M. sylvanus, where males interact extensivelyand intimately with infants soon after birth (Deag & Crook 1971;Deag 1980; Taub 1984). The initial hypothesis that this extensivecaretaking behaviour constitutes paternal investment (Taub 1980)was refuted by DNA fingerprinting (Paul et al. 1992, 1996). Likewise,the ‘male reproductive tactic’ hypothesis, which suggested thatmales increase their mating success by signalling willingness toinvest in a female's offspring, found no support (Paul et al. 1996).Present evidence bolsters the ‘agonistic buffering’ hypothesis,which posits that males use infants to regulate their social rela-tionships and to facilitate maleemale coalitions (Deag & Crook1971; Paul et al. 2000). Coalition formation is a common strategyto increase an individual's competitive ability temporarily(Harcourt 1989) and is found in various primate species (e.g.baboons: Noe & Sluijter 1990; chimpanzees: De Waal 1982). InBarbary macaques, coalition formation was shown to be stronglyrelated to male mating success (Küster & Paul 1992) and to fosterrank maintenance (Widdig et al. 2000) or improvement (Witt et al.1981). Direct evidence linking infant care to male coalition forma-tion and eventually reproductive success is, however, still lacking.
In addition, it remained unclear whether the social conse-quences of infant care, such as increased maleemale interaction or
by Elsevier Ltd. All rights reserved.

Table 1Identity (ID), age, rank in spring and autumn, andweighted degree (male contacts/h)in spring and autumn for the 12 focal males
ID Age(years)
Rankspring
Rankautumn
Degreespring
Degreeautumn
A340 10 1 1 9.25 2.22A337 10 2 4 1.69 2.16O202 19 3 3 7.19 1.08R250 17 4 5 8.06 2.97C361 8 5 2 11.88 3.19C360 8 6 7 2.63 5.24Z321 11 7 9 0.50 0.60M189 20 8 6 7.00 2.43E380 6 9 11 0.81 0.65T265 16 10 10 1.25 0.43D370 7 11 8 0.94 1.03F391 5 12 12 4.81 3.30
S. Henkel et al. / Animal Behaviour 79 (2010) 1199e12041200
a decrease in aggression received, are associated with changes inmale physiology. A number of recent studies have shown thatnonhuman primates respond with elevated glucocorticoid outputto changes in social dynamics in their environment (e.g. Engh et al.2006; Crockford et al. 2008). We therefore determined glucocor-ticoid output in male Barbary macaques, based on analysis of faecalsamples, to study the effects of maleeinfant interactions and theassociated changes in male social dynamics on the adrenal axis,both in spring (intense interactions with infants and other males)and autumn (fewer interactions with infants and other males). Asprevious studies indicated that grooming can influence glucocor-ticoid output in primates (Gust et al. 1993; Shutt et al. 2007; Wittiget al. 2008), we also considered how the amount of grooming givenand received affects glucocorticoid levels. Furthermore, because thefrequency of aggression received could also influence glucocorti-coid levels (e.g. Ostner et al. 2008), this variable was included in themodel as well.
We determined the males' position in the social group and theinfluence of interactions with infants on the number and quality ofsocial relationships using a social network analysis approach. Socialnetwork analyses are attracting increasing attention in studies ofanimal behaviour (Croft et al. 2008; Krause et al. 2009; Lusseau &Conradt 2009). The social network approach extends simplematrix statistics, as it allows one to track connections betweenindividuals to second and higher degrees, and to study the socialorganization of individuals at different levels (Krause et al. 2009). Inaddition, a number of statistics have been developed to charac-terize an animal's position in the social network, the substructure(‘cliquishness’) of a group, as well as population dynamics. In ouranalysis, we applied a statistical measure derived from socialnetwork analysis that captures the centrality of a given individual,based on the number of partners and number of interactions withthese partners. Furthermore, we made use of the graphical repre-sentations derived from the network analysis to provide intuitiveillustrations of the social structure of the group.
If interactions with infants help to alleviate stress, possibly byreducing the amount of aggression received, this should bereflected in a reduced stress response in males that are mainlyinvolved in caretaking. Following the notion that males use infantsto regulate their social relationships, we further expected thatmales who invest heavily in caretaking have stronger ties to othermales than individuals who do not take care of infants.
METHODS
Study Site and Animals
The study was conducted at the outdoor enclosure ‘La Forêt desSinges’ in Rocamadour, France, a visitor park where monkeys rangefreely, while visitors are restricted to a path (Turckheim & Merz1984). Barbary macaques live in multimale multifemale groups, inwhich females remain in the natal group with their offspring(Küster & Paul 1999). They are seasonal breeders with a birthseason in spring and a mating season in autumn (Small 1990). Datawere collected from the beginning of May to the end of June andfrom the end of August until the end of October 2007 in the largestgroup (‘PB’), which comprised 53e54 individuals over 1 year of age(one male died during the study period). All animals of the groupcould be identified individually. During the study, there were 19males between 5 and 27 years of age in the group, from which wechose 12 aged 5e20 years as focal animals (Table 1). Males wereselected with the aim of covering all rank classes (high, middle,low) and the broad range of the age distribution, while excludingvery old males who have a tendency to withdraw from the group (J.Fischer, personal observation). By the start of the observations in
May, two babies (a female born 11 April 2007 to female M189 anda male born 19 May 2007 to female G400) were already intenselycared for by males; another male baby was born 7 July 2007 tofemale L170 after the first period of observations had finished.Neither female G400 nor L170 had male maternal relatives in thegroup. M189 was the mother of A340, the alpha male, who was thebrother of E380, another male in our sample. All other males werematernally unrelated. Relatedness data were obtained from E. Merz(personal communication).
Behavioural Observations
For each focal male, we conducted 69 half-hour protocols (32 inspring and 37 in autumn) with the focal animal sampling method,resulting in a total observation time of 414 h. The order in whichmales were observed was pseudorandomized with the stipulationthat no animalwas observed twice on the sameday, and each animalwas observed equally frequently at the different times of the day.During focal observations, all agonistic (threat, chase away, chaseand attack), submissive (avert, retreat, scream, flee, present, teethchatter), affiliative (grooming, sitting in contact,mount, chatter, hug,support) and maleeinfant interactions (carry baby, triadic interac-tion) were recorded continuously using a Palm handheld computer(Model Zire; Palm Inc., Sunnyvale, CA, U.S.A.) running a customprogram created with Pendragon Forms (Pendragon SoftwareCooperation, Libertyville, IL, U.S.A.). A complete description of theBarbarymacaque ethogram can be found in Hesler & Fischer (2007).We used MatMan 1.1 (Noldus, Wageningen, Netherlands) toconstruct a dominance matrix using ad libitum and focal data ofdyadic agonistic encounters and approacheretreat interactions(N ¼ 369 interactions).
Faecal Sample Collection and Glucocorticoid Analysis
We analysed 287 faecal samples (147 in spring, 140 in autumn,5e16 faecal samples per focal animal and season, median ¼ 12).Samples were collected directly after defecation, placed on ice forup to 4 h until transferred to a freezer and stored at �20 �C untilhormone analysis. All analysed samples were collected after noon(1200e1700 hours) as in this time period no effect of collectiontime on glucocorticoid levels was found (data not shown). Prior toanalysis, samples were lyophilized and pulverized and a portion ofthe faecal powder was extracted with 3 ml of 80% methanol(Heistermann et al. 2004). Faecal extracts were analysed forimmunoreactive 11b-hydroxyetiocholanolone (3a,11b-dihydroxy-CM), a group-specific measurement of 5-reduced 3a,11b-dihy-droxylated cortisol metabolites which has been proven to provide

S. Henkel et al. / Animal Behaviour 79 (2010) 1199e1204 1201
reliable information on glucocorticoid output in primates,including the Barbary macaque (Heistermann et al. 2006; Shuttet al. 2007). Hormone measurements were carried out by micro-titerplate enzyme-immunoassay as described in detail byHeistermann et al. (2004). Intra-assay coefficients of variation (CV)of high and low concentrated quality controls (QC) were 6.5%(N ¼ 16) and 7.9% (N ¼ 16), respectively, while interassay CVs were11.0% (QC high, N ¼ 22) and 16.0% (QC low, N ¼ 22). For each male,we calculated the mean 3a,11b-dihydroxy-CM value (‘GC level’hereafter).
Statistical Analysis
We used linear regressions to identify significant relationshipsbetween response and predictor variables (Table 2). Models wereimplemented in the R statistical computing environment (RDevelopment Core Team 2008; library LME4). While all statisticalanalyses were done on the continuous data, we report some of theresults on an aggregate basis for carriers and noncarriers.
To test whether the observed associations in the networkdeviated from a homogeneous distribution, we applied a permu-tation procedure. Specifically, we permuted partners between focalsubjects whereby we restricted the permutation such that inter-actions of the focal animal with itself were impossible. Thispermutation procedure was based on the original data, that is, onthe single interactions between partners, and we assessed whetherboth the frequency and the duration of interactions deviated froma random pattern. As a test statistic we used the standard deviationof the interaction measures per dyad (durations or frequencies). Inother words, we tested whether more dyads showed particularlyhigh or low association measures than expected by chance. Wepermuted the data 1000 times (with the original data included asone permutation) and estimated the P value as the number ofpermutations revealing a test statistic at least as large as the orig-inal one (Adams & Anthony 1996; Manly 1997).
We then established matrices for spring and autumn, respec-tively, which were based on the number of interactions per hourbetween a given dyad.We used UCINET (Borgatti et al. 2002; Vital &Martins 2009) for the network analysis. We examined both theweighted and unweighted (dichotomized) networks. While theformer represent both the number and strength of relationships,the latter are based on the number of partners. Because we foundthe weighted degree more informative, and the two variables werecorrelated, we constructed the network based on the weighteddegree and simply report the results for the unweighted degree. Insocial network analyses parlance, individuals are represented asnodes, while relationships are represented as edges. To reduce the
Table 2Variables used in the statistical analysis
Variable Description
Baby carrying (s) Duration of all dyadic malNumber of male contacts per hour (N/h)
(‘weighted degree’)All instances when two minteractions’ during whichand nuzzling an infant as wwas broken down by dyad
Number of partners (N) (‘degree’) Number of male partnersGrooming given (s) Total duration the focal mGrooming received (s) Total duration a focal maleAggression received (N) Number of times the focal
(>3 m) by another male (sInitiator Ratio of maleemale intera
interactions initiated by thMale rank (ordinal) Established separately forGlucocorticoid levels (mg/g) See Methods for details
effects of noise, we followed the advice of James et al. (2009) andremoved all edges that were based on a single interaction prior tothe calculation of individual measures. We used NETDRAW(Borgatti 2002) to visualize the social network among the focalmales in the two seasons.
RESULTS
Males could be ordered in a linear hierarchy both in spring(h0 ¼ 0.636, c32.23
2 ¼ 163.05, P < 0.001) and in autumn (h0 ¼ 0.604,c32.232 ¼ 152.50, P < 0.001). Several rank reversals occurred
between spring and autumn, particularly among middle-rankingmales (Table 1). Male care for infants (‘baby carrying’) was unevenlydistributed among males. In spring, six of the 12 males wereobserved carrying babies. Three of them accounted for 86% of thetotal time that a baby was observed in the care of a male. Dyadicmaleeinfant interactions amounted to 11.86 min/h on average(range 1.9e29 min/h, N ¼ 6). Male rank did not predict babycarrying (Fig. 1a, Table 3). The three males that carried babies mostfrequently were ranked first, fourth and eighth, respectively(Fig. 1a). In contrast to our predictions, a high amount of babycarrying was associated with high rather than low stress hormonelevels (Fig. 1b). None of the other variables accounted for variationin glucocorticoid levels, except for a marginally significant rela-tionship with the amount of aggression received. Notably, therewas no relation between the amounts of grooming given orreceived and glucocorticoid levels (Table 3).
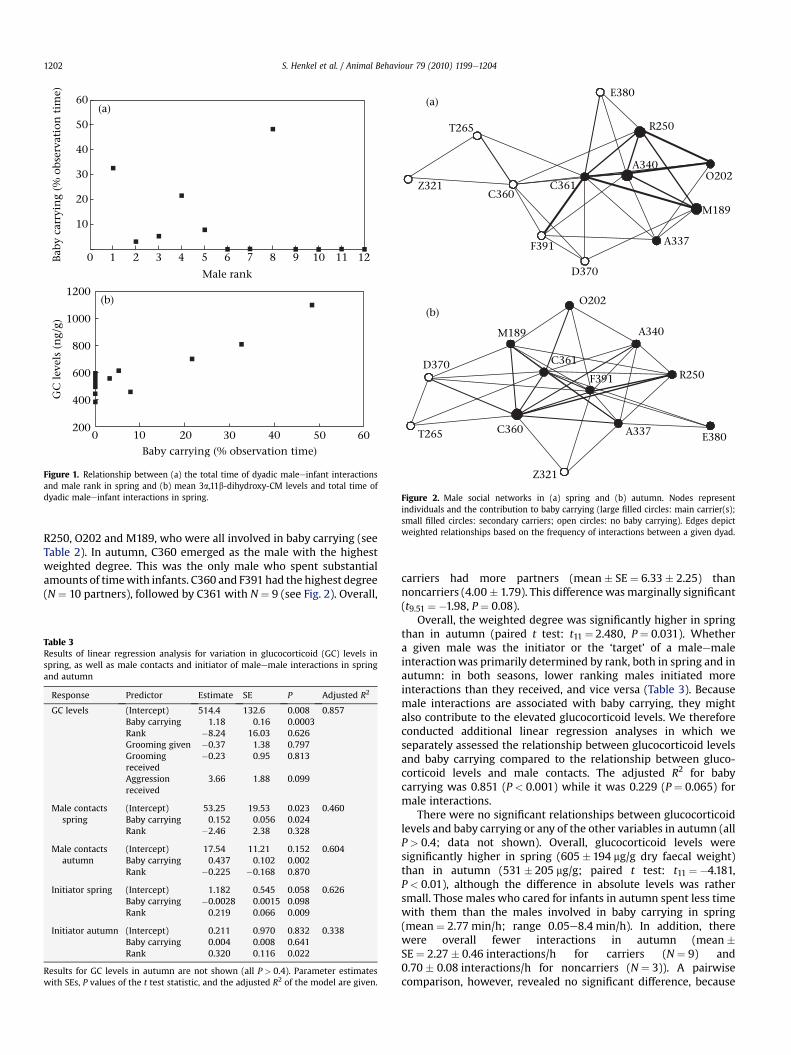
The permutation procedure revealed a nonrandom structure ofthe male network in spring (interaction frequency: SD ¼ 13.63,P ¼ 0.001; for durations: SD ¼ 1015.43, P ¼ 0.001). In autumn, thisdeviation from a random structure was true for the interactionsonly, but not the durations of interactions (frequencies: 4.63,P ¼ 0.001; durations: 340.64, P ¼ 0.405). We found a significantrelationship between baby carrying and the frequency of malecontacts and the structure of the male social network (Fig. 2). Maleswho carried infants had significantly more interactions with othermales than noncarriers (mean � SE ¼ 4.85 � 0.82 interactions/h forcarriers (N ¼ 6) and 1.60 � 0.44 interactions/h for noncarriers(N ¼ 6)). These interactions were significantly shorter for carriers(67.9 � 20.11 s) than for noncarriers (123.4 � 7.6 s; see Table 3 forresults of statistical analyses). In spring, the most central male withthe highest ‘weighted degree’, that is, the highest number of inter-actions was male C361 (see Table 1), who during this periodcontinually challenged other males and rose in rank from 5 to 2 inautumn.He also had thehighest ‘degree’, that is, the highest numberofmale partners (Fig. 2). Themalewith the secondhighestweighteddegree and regular degree was A340, the alpha male, followed by
eeinfant interactions, i.e. carrying, holding or grooming the infantales were sitting in body contact. This included so-called ‘triadicmales engage in an elaborate succession of ritualized greeting, lip smacking,ell as simple contact sitting. For the social network analysis, this
ale was observed grooming another subjectwas observed being groomed by another subjectmale was being threatened, attacked, chased (<3 m), or chased awayee Patzelt et al. 2009 for details)ctions initiated by the focal male divided by the number ofe partner. Interactions initiated by both parties were excluded from this analysisspring and autumn; see Methods for details
(a)
D370
E380
F391 A337
R250T265
Z321C360
C361
A340O202
M189
(b)
F391
A337
R250
T265
Z321
C360
C361
A340
O202
M189
D370
E380
Figure 2. Male social networks in (a) spring and (b) autumn. Nodes representindividuals and the contribution to baby carrying (large filled circles: main carrier(s);small filled circles: secondary carriers; open circles: no baby carrying). Edges depictweighted relationships based on the frequency of interactions between a given dyad.
60
50
6050
40
30
20
10
403020100
0 1 2 3 4 5 6 7 8 9 10 11 12
Male rank
Bab
y ca
rryi
ng
(% o
bser
vati
on t
ime)
Baby carrying (% observation time)
1200
1000
800
600
400
200
GC
lev
els
(ng/
g)
(a)
(b)
Figure 1. Relationship between (a) the total time of dyadic maleeinfant interactionsand male rank in spring and (b) mean 3a,11b-dihydroxy-CM levels and total time ofdyadic maleeinfant interactions in spring.
S. Henkel et al. / Animal Behaviour 79 (2010) 1199e12041202
R250, O202 and M189, who were all involved in baby carrying (seeTable 2). In autumn, C360 emerged as the male with the highestweighted degree. This was the only male who spent substantialamounts of timewith infants. C360 and F391 had the highest degree(N ¼ 10 partners), followed by C361 with N ¼ 9 (see Fig. 2). Overall,
Table 3Results of linear regression analysis for variation in glucocorticoid (GC) levels inspring, as well as male contacts and initiator of maleemale interactions in springand autumn
Response Predictor Estimate SE P Adjusted R2
GC levels (Intercept) 514.4 132.6 0.008 0.857Baby carrying 1.18 0.16 0.0003Rank �8.24 16.03 0.626Grooming given �0.37 1.38 0.797Groomingreceived
�0.23 0.95 0.813
Aggressionreceived
3.66 1.88 0.099
Male contactsspring
(Intercept) 53.25 19.53 0.023 0.460Baby carrying 0.152 0.056 0.024Rank �2.46 2.38 0.328
Male contactsautumn
(Intercept) 17.54 11.21 0.152 0.604Baby carrying 0.437 0.102 0.002Rank �0.225 �0.168 0.870
Initiator spring (Intercept) 1.182 0.545 0.058 0.626Baby carrying �0.0028 0.0015 0.098Rank 0.219 0.066 0.009
Initiator autumn (Intercept) 0.211 0.970 0.832 0.338Baby carrying 0.004 0.008 0.641Rank 0.320 0.116 0.022
Results for GC levels in autumn are not shown (all P > 0.4). Parameter estimateswith SEs, P values of the t test statistic, and the adjusted R2 of the model are given.
carriers had more partners (mean � SE ¼ 6.33 � 2.25) thannoncarriers (4.00 � 1.79). This differencewasmarginally significant(t9.51 ¼ �1.98, P ¼ 0.08).
Overall, the weighted degree was significantly higher in springthan in autumn (paired t test: t11 ¼ 2.480, P ¼ 0.031). Whethera given male was the initiator or the ‘target’ of a maleemaleinteractionwas primarily determined by rank, both in spring and inautumn: in both seasons, lower ranking males initiated moreinteractions than they received, and vice versa (Table 3). Becausemale interactions are associated with baby carrying, they mightalso contribute to the elevated glucocorticoid levels. We thereforeconducted additional linear regression analyses in which weseparately assessed the relationship between glucocorticoid levelsand baby carrying compared to the relationship between gluco-corticoid levels and male contacts. The adjusted R2 for babycarrying was 0.851 (P < 0.001) while it was 0.229 (P ¼ 0.065) formale interactions.
There were no significant relationships between glucocorticoidlevels and baby carrying or any of the other variables in autumn (allP > 0.4; data not shown). Overall, glucocorticoid levels weresignificantly higher in spring (605 � 194 mg/g dry faecal weight)than in autumn (531 � 205 mg/g; paired t test: t11 ¼ �4.181,P < 0.01), although the difference in absolute levels was rathersmall. Those males who cared for infants in autumn spent less timewith them than the males involved in baby carrying in spring(mean ¼ 2.77 min/h; range 0.05e8.4 min/h). In addition, therewere overall fewer interactions in autumn (mean �SE ¼ 2.27 � 0.46 interactions/h for carriers (N ¼ 9) and0.70 � 0.08 interactions/h for noncarriers (N ¼ 3)). A pairwisecomparison, however, revealed no significant difference, because

S. Henkel et al. / Animal Behaviour 79 (2010) 1199e1204 1203
some of the males spent more time with infants in autumn than inspring (paired t test: t11 ¼ �1.272, P ¼ 0.229).
DISCUSSION
Male Barbary macaques who carried infants in the birth seasonin spring had significantly stronger relationships with other malesthan those males who did not carry infants, and they experienceda significantly higher glucocorticoid output thanmales who did notspend time with infants. Because of the collinearity between infantcarrying and the number of interactions with other males, it is notstraightforward to disentangle the relationships between elevatedglucocorticoid levels, the investment in infant carrying and maleinteractions. Males that carried infants had more interactions withother males, but these interactions were significantly shorter. Bydefinition, male interactions without infants involve only ‘contactsitting’ while interactions with infants also involve ‘triadic inter-actions’, including the elaborate greetings and nuzzling proceduretypical for this species (Deag 1980; Taub 1980). In other words,these interactions have an entirely different quality, which mayaffect the average duration of contacts. Despite these difficulties ofseparating the different factors, it is noteworthy that the time spentwith infants alone produced a much better fit in a separate linearregression analysis with glucocorticoid levels than the time spentwith other males. While the strong link between infant carryingand glucocorticoid levels does not establish a causal relationship, itseems unlikely that such a close association would be because‘nervous’ males with high glucocorticoid levels are particularlykeen to hold infants as a means to calm themselves down, or to getaccess to themale network. If this were true, males who spend timewith infants should experience a gradual decrease in glucocorticoidoutput, and thus lower glucocorticoid levels. Instead, it seemsmorelikely that it is the baby carrying that is primarily responsible forelevated glucocorticoid levels. Accordingly, interactions withinfants can be viewed as a costly investment (Sapolsky et al. 2000)to gain access to the male network.
Our results are in line with the view that males use infants asa ‘social tool’ to establish and/or strengthen social bonds with othergroupmembers (Paul et al. 2000). The uneven distribution of infantcarrying suggests that some males specifically invest in caretakingto maintain strong relations with other males, as revealed by thesocial network analysis. This does not necessarily mean that mainlymales who carried infants initiated maleemale interactions.Instead, the initiation of male contacts was primarily related torank. Male care, in contrast, was not related to rank, refuting theinterpretation that infants are a rare commodity that malescompete for (Small 1990). Instead, it appears more likely that themotivation to invest in infant carrying strongly depends on theposition in the social network and the current constellation ofrelationships.
In autumn, the relationships between the variables underexamination were less clear. Glucocorticoid levels were overallsignificantly lower than in spring, suggesting that some additionalfactor other than infant carrying also affects glucocorticoid levels. Itmay be the case that the rank challenge initiated by one of themales (C361) had led to an overall increase in tension in spring.Alternatively, glucocorticoid levels may vary with the time of year,independently from the social stability in the group. Indeed, manyvertebrate taxa reveal annual rhythms in glucocorticoid concen-trations. In birds and reptiles, glucocorticoid levels peak during thebreeding season, but no consistent pattern has been found inmammals: some studies revealed peaks during and others afterbreeding (Romero 2002). Nevertheless, general shifts in glucocor-ticoid levels are nothing unexpected, although it would be neces-sary to monitor hormone levels over several years to identify the
combination of factors, including annual patterns, that contributeto the physiological stress response.
Our results add to the growing body of evidence that bothpredictable and unpredictable events in the social domain aremajor sources of stress (Crockford et al. 2008). Male red-frontedlemurs, Eulemur fulvus rufus, and male sifakas, Propithecus verrauxi,for instance, experience elevated glucocorticoid levels in the birthseason which are related to the number of infants born, possiblybecause of the imminence of increased male transfers betweengroups and the increased likelihood of infanticide (Ostner et al.2008; Brockman et al. 2009). Similarly, male chacma baboons,Papio ursinus, respond to immigration events that destabilize themale hierarchy with a physiological stress response (Beehner et al.2005; Engh et al. 2006). Our results indicate that in Barbarymacaques, infant carrying is very likely to be one of the factors thatcontribute to the physiological stress response in males. Possibly,male caretakers have elevated glucocorticoid levels because infantsspend considerable time ‘crying’. Previous studies showed thatinfant crying is an aversive stimulus (Todt 1988; Owren & Rendall2001). Because we did not record infant behaviour, we are pres-ently unable to examine this conjecture.
In contrast to previous studies (Gust et al. 1993; Shutt et al.2007), we found no evidence that grooming given or groomingreceived was related to glucocorticoid levels. In female Barbarymacaques, giving rather than receiving grooming is associated withlower stress levels (Shutt et al. 2007). Possibly, this differencebetween the sexes can be accounted for by the difference inmagnitude of time spent grooming: in our study, males spent only54 s/h (1.5% of the total observation time) grooming others,corroborating previous findings (Deag 1980; Paul et al. 1996). Incontrast, in the study by Shutt et al. (2007), females' groomingbouts were almost six times longer.
To summarize, our results indicate that carrying infantsprovides access to the male network and helps to facilitate inter-actions between males. Previous studies indicated that males whoform coalitions with others have higher reproductive success (Wittet al. 1981; Paul et al. 1992), thus conferring a benefit to males whoestablish strong links with others. However, for some males, thisaccess to the male network appears to come with a cost, as evi-denced by the increased stress hormone level in males who spendsignificant amounts of time with young infants.
Acknowledgments
We are grateful to Ellen Merz and Gilbert de Turckheim forpermission to carry out the study at ‘La Forêt des Singes’ inRocamadour, for the continual support of our studies, and to thestaff on site, particularly Gérome, for their cooperation and hospi-tality. Many thanks go to Ellen Merz for kindly providing therelatedness data, Andrea Heistermann for her invaluable assistancewith the hormone analysis, Anke Gutmann for company, RalphPirow for sharing his statistical expertise, Jens Krause for inspiringdiscussions, Stuart Semple for helpful comments on the manu-script, and Roger Mundry for the permutation procedure andfriendship.
References
Adams, D. C. & Anthony, C. D. 1996. Using randomization techniques to analysebehavioural data. Animal Behaviour, 51, 733e738.
Beehner, J. C., Bergman, T. J., Cheney, D. L., Seyfarth, R. M. & Whitten, P. L. 2005.The effect of new alpha males on female stress in free-ranging baboons. AnimalBehaviour, 69, 1211e1221, doi:10.1016/j.anbehav.2004.08.014.
Borgatti, S. P. 2002. NetDraw: Graph Visualization Software. Cambridge, Massa-chusetts: Analytic Technologies.
Borgatti, S. P., Everett, M. G. & Freeman, L. C. 2002. Ucinet for Windows: Softwarefor Social Network Analysis. Cambridge, Massachusetts: Analytic Technologies.

S. Henkel et al. / Animal Behaviour 79 (2010) 1199e12041204
Brockman, D. K., Cobden, A. K. & Whitten, P. L. 2009. Birth season glucocorticoidsare related to the presence of infants in sifaka (Propithecus verreauxi).Proceedings of the Royal Society B, 276, 1855e1863, doi:10.1098/rspb.2008.1912.
Buchan, J., Alberts, S., Silk, J. & Altmann, J. 2003. True paternal care in a multi-male primate society. Nature, 425, 179e181, doi:10.1038/nature01866.
Clutton-Brock, T. H. 1991. The Evolution of Parental Care. Princeton, New Jersey:Princeton University Press.
Clutton-Brock, T. H., Brotherton, P. N. M., Russell, A. F., O'Riain, M. J., Gaynor, D.,Kansky, R., Griffin, A., Manser, M., Sharpe, L., McIlrath, G. M., Small, T.,Moss, A. & Monfort, S. 2001. Cooperation, control, and concession in meerkatgroups. Science, 291, 478e481, doi:10.1126/science.291.5503.478.
Crockford, C., Wittig, R. M., Whitten, P. L., Seyfarth, R. M. & Cheney, D. L. 2008.Social stressors and coping mechanisms in wild female baboons (Papiohamadryas ursinus). Hormones and Behavior, 53, 254e265. doi: S0018-506X(07)00245-0 [pii] 10.1016/j.yhbeh.2007.10.007.
Croft, D. P., James, R. A. & Krause, J. 2008. Exploring Animal Social Networks.Princeton, New Jersey: Princeton University Press.
De Waal, F. B. M. 1982. Chimpanzee Politics. New York: Harper & Row.Deag, J. M. 1980. Interactions between males and unweaned Barbary macaques:
testing the agonistic buffering hypothesis. Behaviour, 75, 54e81.Deag, J. M. & Crook, J. H. 1971. Social behaviour and agonistic buffering in the wild
Barbary macaque, Macaca sylvanus. Folia Primatologica, 15, 183e200.Doolan, S. P. & Macdonald, D. W. 1999. Co-operative rearing by slender-tailed
meerkats (Suricata suricatta) in the Southern Kalahari. Ethology, 105, 851e866,doi:10.1046/j.1439-0310.1999.00461.x.
Engh, A. L., Beehner, J. C., Bergman, T. J., Whitten, P. L., Hoffmeyer, R. R.,Seyfarth, R. M. & Cheney, D. L. 2006. Female hierarchy instability, maleimmigration and infanticide increase glucocorticoid levels in female chacmababoons. Animal Behaviour, 71, 1227e1237, doi:10.1016/j.anbehav.2005.11.009.
Gust, D., Gordon, T., Hambright, K. &Wilson, M.1993. Relationship between socialfactors and pituitaryeadrenocortical activity in female rhesus monkeys (Macacamulatta). Hormones and Behavior, 27, 318e331, doi:10.1006/hbeh.1993.1024.
Harcourt, A. H. 1989. Social influences on competitive ability: alliances and theirconsequences. In: Comparative Socioecology: the Behavioural Ecology of Humansand Other Mammals (Ed. by V. Standen & R. A. Foley), pp. 223e242. Oxford:Blackwell Scientific.
Heistermann, M., Ademmer, C. & Kaumanns, W. 2004. Ovarian cycle and effect ofsocial changes on adrenal and ovarian function in Pygatherix nemaeus. Inter-national Journal of Primatology, 25, 689e708.
Heistermann, M., Palme, R. & Ganswindt, A. 2006. Comparison of differentenzymeimmunoassays for assessment of adrenocortical activity in primatesbased on fecal analysis. American Journal of Primatology, 68, 257e273.
Hesler, N. & Fischer, J. 2007. Gestural communication in Barbary macaques (Macacasylvanus): an overview. In: The Gestural Communication of Apes and Monkeys(Ed. by M. Tomasello & J. Call), pp. 159e195. Mahwah, New Jersey: L. Erlbaum.
Hodge, S. J. 2007. Counting the costs: the evolution of male-biased care in thecooperatively breeding banded mongoose. Animal Behaviour, 74, 211e219.
James, R., Croft, D. P. & Krause, J. 2009. Potential banana skins in animal socialnetwork analysis. Behavioral Ecology and Sociobiology, 63, 989e997,doi:10.1007/s00265-009-0742-5.
Krause, J., Lusseau, D. & James, R. 2009. Animal social networks: an introduction.Behavioral Ecology and Sociobiology, 63, 967e973, doi:10.1007/s00265-009-0747-0.
Küster, J. & Paul, A. 1992. Influence of male competition and female mate choice onmale mating success in Barbary macaques (Macaca sylvanus). Behaviour, 120,192e217.
Küster, J. & Paul, A.1999. Male migration in Barbary macaques (Macaca sylvanus) atAffenberg Salem. International Journal of Primatology, 20, 85e106.
Lusseau, D. & Conradt, L. 2009. The emergence of unshared consensus decisions inbottlenose dolphins. Behavioral Ecology and Sociobiology, 63, 1067e1077,doi:10.1007/s00265-009-0740-7.
Manly, B. F. J. 1997. Randomization, Bootstrap and Monte Carlo Methods in Biology.New York: Chapman & Hall.
Noe, R. & Sluijter, A. A. 1990. Reproductive tactics of male savanna baboons.Behaviour, 113, 117e170.
Ostner, J., Kappeler, P. M. & Heistermann, M. 2008. Androgen and glucocorticoidlevels reflect seasonally occurring social challenges in male redfronted lemurs(Eulemur fulvus rufus). Behavioral Ecology and Sociobiology, 62, 627e638,doi:10.1007/s00265-007-0487-y.
Owren, M. J. & Rendall, D. 2001. Sound on the rebound: returning signal form andfunction to the forefront in understanding nonhuman primate vocal signaling.Evolutionary Anthropology, 10, 58e71.
Palombit, R. A., Seyfarth, R. M. & Cheney, D. L. 1997. The adaptive value of‘friendships’ to female baboons: experimental and observational evidence.Animal Behaviour, 54, 599e614.
Patzelt, A., Pirow, R. & Fischer, J. 2009. Post-conflict affiliation in Barbarymacaques is influenced by conflict characteristics and relationship quality, butdoes not diminish short-term renewed aggression. Ethology, 115, 658e670,doi:10.1111/j.1439-0310.2009.01649.x.
Paul, A., Küster, J. & Arnemann, J. 1992. DNA fingerprinting reveals that infantcare by male Barbary macaques (Macaca sylvanus) is not paternal investment.Folia Primatologica, 58, 93e98.
Paul, A., Küster, J. & Arnemann, J. 1996. The sociobiology of maleeinfant inter-actions in Barbary macaques (Macaca sylvanus). Animal Behaviour, 51, 155e170.
Paul, A., Preuschoft, S. & Van Schaik, C. 2000. The other side of the coin: infan-ticide and the evolution of affiliative maleeinfant interactions in Old Worldprimates. In: Infanticide by Males and Its Implications (Ed. by C. Van Schaik &C. H. Janson), pp. 269e292. Cambridge: Cambridge University Press.
Pusey, A. & Packer, C. 1994. Infanticide in lions: consequences and counterstrat-egies. In: Infanticide and Parental Care (Ed. by S. Parmigiani & F. S. vom Saal),pp. 277e299. Chur: Harwood Academic.
R Development Core Team 2008. A Language and Environment for StatisticalComputing. Vienna: R Foundation for Statistical Computing.
Romero, L. M. 2002. Seasonal changes in plasma glucocorticoid concentrations infree-living vertebrates. General and Comparative Endocrinology, 128, 1e24.
Sapolsky, R. M., Romero, L. M. & Munck, A. U. 2000. How do glucocorticoidsinfluence stress responses? Integrating permissive, suppressive, stimulatory,and preparative actions. Endocrinology Reviews, 21, 55e89.
Schradin, C. & Anzenberger, G. 2001. Costs of infant carrying in commonmarmosets, Callithrix jacchus: an experimental analysis. Animal Behaviour, 62,289e295.
Shutt, K., MacLarnon, A., Heistermann, M. & Semple, S. 2007. Grooming inBarbary macaques: better to give than to receive? Biology Letters, 3, 231e233.
Small, M. F. 1990. Alloparental behaviour in Barbary macaques, Macaca sylvanus.Animal Behaviour, 39, 297e306.
Smuts, B. B. 1985. Sex and Friendship in Baboons. New York: Aldine.Taub, D. M. 1980. Testing the ‘agonistic buffering’ hypothesis. 1. The dynamics of
participation in the triadic interaction. Behavioral Ecology and Sociobiology, 6,187e197.
Taub, D. M. 1984. Male caretaking behaviour among wild Barbary macaques. In:Primate Paternalism (Ed. by D. Taub), pp. 20e55. New York: Van NostrandReinhold.
Todt, D. 1988. Serial calling as a mediator of interaction processes. In: Primate VocalCommunication (Ed. by D. Todt, P. Goedeking & D. Symmes), pp. 88e107.Berlin: Springer.
Trivers, R. L.1972. Parental investment and sexual selection. In: Sexual Selection andthe Descent of Man (Ed. by B. Campbell), pp. 136e179. Chicago: Aldine-Atherton.
Turckheim, G. D. & Merz, E. 1984. Breeding Barbary macaques in outdoor openenclosures. In: The Barbary Macaque: a Case Study in Conservation (Ed. byJ. F. Fa), pp. 241e261. New York: Plenum.
Van Schaik, C. P. & Kappeler, P. M. 1997. Infanticide risk and the evolution ofmaleefemale association in primates. Proceedings of the Royal Society B, 264,1687e1694.
Vital, C. & Martins, E. P. 2009. UCINET for Windows: software for social networkanalysis. Ethology, 115, 347e355, doi:10.1111/j.1439-0310.2009.01613.x.
Widdig, A., Streich, W. J. & Tembrock, G. 2000. Coalition formation among maleBarbarymacaques (Macaca sylvanus). American Journal of Primatology, 50, 37e51.
Witt, R., Schmidt, C. & Schmitt, J. 1981. Social rank and darwinian fitnessin a multimale group of Barbary macaques (Macaca sylvana Linnaeus, 1758).Folia Primatologica, 36, 201e211.
Wittig, R. M., Crockford, C., Lehmann, J., Whitten, P. L., Seyfarth, R. M. &Cheney, D. L. 2008. Focused grooming networks and stress alleviation in wildfemale baboons. Hormones and Behavior, 54, 170e177, doi:10.1016/j.yhbeh.2008.02.009.
Wright, P. C. 1990. Patterns of paternal care in primates. International Journal ofPrimatology, 11, 89e102.
Zhao, Q. K. 1996. Maleeinfantemale interactions in Tibetan macaques. Primates, 37,135e143.