increased inflammatory markers identified in the dorsolateral prefrontal cortex of individuals with...
TRANSCRIPT

ORIGINAL ARTICLE
Increased inflammatory markers identified in the dorsolateralprefrontal cortex of individuals with schizophreniaSG Fillman1,2,3, N Cloonan4, VS Catts1,2,3, LC Miller5, J Wong6, T McCrossin7, M Cairns1,8 and CS Weickert,1,2,3
Upregulation of the immune response may be involved in the pathogenesis of schizophrenia with changes occurring inboth peripheral blood and brain tissue. To date, microarray technology has provided a limited view of specificinflammatory transcripts in brain perhaps due to sensitivity issues. Here we used SOLiD Next Generation Sequencing toquantify neuroimmune mRNA expression levels in the dorsolateral prefrontal cortex of 20 individuals with schizophreniaand their matched controls. We detected 798 differentially regulated transcripts present in people with schizophreniacompared with controls. Ingenuity pathway analysis identified the inflammatory response as a key change. Using quantitativereal-time PCR we confirmed the changes in candidate cytokines and immune modulators, including interleukin (IL)-6,IL-8, IL-1b and SERPINA3. The density of major histocompatibility complex-II-positive cells morphologically resemblingmicroglia was significantly increased in schizophrenia and correlated with IL-1b expression. A group of individuals,most of whom had schizophrenia, were found to have increased inflammatory mRNA expression. In summary, we havedemonstrated changes in an inflammatory response pathway that are present in B40% of people diagnosed withschizophrenia. This suggests that therapies aimed at immune system attenuation in schizophrenia may be of directbenefit in the brain.
Molecular Psychiatry advance online publication, 7 August 2012; doi:10.1038/mp.2012.110
Keywords: immune; inflammatory; microglia; next generation sequencing; schizophrenia
INTRODUCTIONSchizophrenia is a severe neuropsychiatric disorder affectingaround 1% of the world’s population. Current treatments areineffective at addressing the full spectrum of symptoms. Under-standing schizophrenia’s causes will allow us to better tailor thetreatments to specific biological subgroups, which may be presentunder the broad diagnostic group of schizophrenia. Previousstudies have used microarrays to examine the brain transcriptomeof individuals with schizophrenia to establish patterns of geneexpression changes implicating GABAergic, synaptic and myelinpathways, as well as inflammatory/immune response.1--6 However,microarray studies have been limited by target specificity,restricted linear range and background interference.7,8 Next-generation sequencing (NGS), specifically RNA-Seq, overcomesthese problems through the use of massively parallel nucleotidesequencing of small sections of the transcriptome and digitalreassembly of the reads, resulting in no upper limit to quantifica-tion and a very sensitive detection threshold.7
The immune system’s role in schizophrenia is somewhatcontroversial but evidence from genomic, blood, postmortem andin vivo imaging studies are leading toward a greater consensus thatimmune activation is involved.9--12 A significant positive geneticassociation between schizophrenia and the human leukocyteantigen (HLA) locus responsible for major histocompatibility complex(MHC) expression and the genetic region containing cytokines
interleukin (IL)-1a, IL-1b and IL1RA have been found.13--15 Serumcytokines are also significantly increased in schizophrenia (IL-6, IL-8).10,16--21 Elevations in expression of other immune-related genes,but not cytokines, have been found in the dorsolateral prefrontalcortex (DLPFC) of individuals with schizophrenia.1,5 In brain, cytokinesare synthesized and secreted by the resident immune cells andmicroglia, among others. Microglia have diverse functions includingserving as antigen-presenting cells through the MHC-II receptor.22--24
Brain microglia have been measured using in vivo positron emissiontomography and in vitro binding with significant increases in(11C)PK-11195 binding observed in people with schizophreniacompared with controls in two out of three studies.25--27 Thisevidence would indicate that there is a subset of living individualswith schizophrenia who may have increased immune responses intheir brain. Thus, we hypothesized that we would detect increasedmicroglia density and increased mRNA levels of pro-inflammatorycytokines in the brains of people with schizophrenia.
The current study is among the first to use NGS to examine thetranscriptome of DLPFC from 20 individuals with schizophreniaand 20 matched controls, providing the most sensitive examina-tion of gene expression changes in schizophrenia available todate. In addition, the RNA-Seq results were confirmed byquantitative PCR (qPCR) in an expanded cohort (n¼ 74) in whichmicroglial density was also measured through immunohistochem-istry, cell counting, and western blotting.
Received 6 March 2012; revised 22 May 2012; accepted 12 June 2012
1Schizophrenia Research Institute, Sydney, New South Wales, Australia; 2Schizophrenia Research Laboratory, Neuroscience Research Australia, Sydney, New South Wales,Australia; 3School of Psychiatry, University of New South Wales, Sydney, New South Wales, Australia; 4Institute for Molecular Bioscience, University of Queensland, Brisbane,Queensland, Australia; 5Children’s Medical Research Institute, Westmead, New South Wales, Australia; 6Illawarra Health and Medical Research Institute and the School ofBiological Sciences, University of Wollongong, Wollongong, New South Wales, Australia; 7Discipline of Pathology, The University of Sydney, Sydney, New South Wales, Australiaand 8School of Biomedical Sciences and Pharmacy, University of Newcastle, Newcastle, New South Wales, Australia. Correspondence: Professor CS Weickert, Macquarie GroupFoundation Chair of Schizophrenia Research, Neuroscience Research Australia, Barker Street, Randwick, NSW 2031 Australia.E-mail: [email protected]
Molecular Psychiatry (2012) 1 -- 9& 2012 Macmillan Publishers Limited All rights reserved 1359-4184/12
www.nature.com/mp

MATERIALS AND METHODSNew South Wales (NSW) Tissue Resource Center postmortem braintissue samplesTissue from the DLPFC (BA46) was obtained for individuals withschizophrenia and controls (demographic summary Table 1, detailsSupplementary Table 1). Details on tissue preparation and RNA extractioncan be found in Weickert et al.28 This human study was carried out inaccordance with the latest version of the Declaration of Helsinki after reviewby the Human Research Ethics Committee at the University of New SouthWales (HREC #07261). Samples for sequencing were chosen to reducesubject variability by being male and having a primary diagnosis ofschizophrenia (excluding schizoaffective). The tissue from control (n¼ 20)and schizophrenia (n¼ 20) groups remained; age, pH, PMI, RIN and durationof time in freezer matched to controls.
NGS library preparation and sequencingTwelve micrograms of RNA from each of the 40 samples was used increation of the library for sequencing. Briefly the RNA was DNase treatedusing TURBO DNA-free (Ambion, Austin TX, USA) before the depletion ofribosomal RNA using the Ribominus kit (Invitrogen, Carlsbad, CA, USA). Theresulting RNA was fragmented and the SOLiD library prepared using theTotal RNA-Seq kit (Invitrogen). Quality control of the library was assessedby size (480% and within 200--300 bp) using a bioanalyser. Samples wererun with four individuals per slide on the SOLiDv4 (Life Technologies,Carlsbad, CA, USA). Reads were unpaired and 50 nt in length. Averagereads per individual were 135 355 990 (±32 081 820) and did not differbetween diagnostic groups.
MappingRNA-Seq reads were aligned to the genome using the X-MATErecursive mapping pipeline29 and the following parameters: (i) strand-specific mapping, where tags are expected to align on the sense strand; (ii)raw tag length of 50 nucleotides; (iii) genome and junction mappingperformed with the ISAS mapping engine;30 (iv) recursive mappingN, M where N¼ length of tag and M¼number of allowable mismatcheswas 50, 5 then 45, 5 then 40, 5 then 35, 3; (v) tags aligning to multiplegenomic locations were discarded; (vi) exhaustive alignments wereenabled; (vii) quality filtering was disabled; and (viii) mapping to junctionswas enabled. During the alignment to junctions, 10 nucleotides wererequired to overlap the exon--exon boundary for tag lengths X40 nt, andfive nucleotides were required to overlap for tag lengths o40 nt. Exon--exon junction libraries were constructed using Ref-Seq hg19 (April 2011)annotations using the scripts distributed with X-MATE. Average readsmapped per individual were 59 418 524 (±20 208 906) or 43% (min: 11%max: 52%) of total.
NGS analysisMapping results were converted into an interval format for processing usingGalaxy (main.g2.bx.psu.edu). Intervals were joined to the exon-annotatedgenome (hg19) and counts per exon were calculated before exporting intoR. The counts were aggregated into 33 105 transcripts and analyzed usingEdgeR (www.bioconductor.org/packages/2.3/bioc/html/edgeR.html) to cal-culate differential gene expression between individuals with schizophreniaand controls. Unexpected expression of female-linked sex genes (as shownin Weickert et al.31) was found in one phenotypically male individual(putative XXY), who was excluded from further analysis. Information on foldchange and multiple correction-adjusted P-values according to diagnosticclass was imported into Ingenuity Pathway Analysis (Ingenuity Systems,www.ingenuity.com) for pathway analysis. The gene list was filteredpreserving only those with a false discovery rate o0.05. Remainingsignificant genes were examined with reference to Ingenuity’s pathwayanalysis based on curated literature findings for directly experimentallyobserved results for human. The top biological function was examined and agene list created. A relational network was calculated from these genes andthe top seven candidates were confirmed using qPCR.
Quantitative PCRPreparation of complementary DNA and qPCR experiments were performedas described previously on the entire cohort of 74 individuals.28 The followingprobes were used for the inflammatory genes: SERPINA3 (Hs003153674_m1),IL6 (Hs00174131_m1), IL6ST (Hs01006741_m1), IL8 (HS00174103_m1), IL1B(Hs01555410_m1), NFKB1 (Hs00765730_m1), PTGS2 (Hs00153133_m1), withACTB (Hs99999903_m1), GAPDH (Hs99999905_m1), TBP (Hs00427621_m1)and UBC (Hs00824723_m1) used as normalizing genes. Cycling conditions andreaction amounts were as previously described.28 Some measurements,particularly SERPINA3 and IL8, had qPCR wells, which did not amplify(average: n¼ 1.64, range: 0--6 wells per measurement per group).
Western blottingTo measure HLA-DA/DR/DQ protein expression, western blotting wasperformed as previously described.32 Briefly, 40 mg of frozen tissue fromthe DLPFC was homogenized in 400ml of homogenization buffer. Protein(20mg) was diluted in Laemmli buffer, and separated by SDS-polyacryla-mide gel electrophoresis on 12% Bis/Tris acrylamide gels (Bio-Rad, Sydney,Australia). Proteins were transferred onto nitrocellulose, and blocked.Membranes were incubated with a primary antibody for human HLA-DP/DQ/DR (1: 200 Dako, Campbellfield, Victoria, Australia; M0775, clone CR3/43) or b-actin (1: 10 000 Chemicon, Sydney, Australia; MAB1501) at 4 1Covernight. Membranes were washed, and incubated with anti-mouseperoxidase-conjugated affinity-purified secondary antibodies (1: 1000 inblocking solution, Chemicon) at room temperature for 1 h. After furtherwashing, bound antibodies were incubated with enhanced chemilumines-
Table 1. Demographics of the schizophrenia and matched control subjects in the New South Wales Tissue Resource Center postmortem braintissue cohort
DemographicsWhole cohort Sequencing cohort
Schizophrenia (n¼ 37) Control (n¼ 37) Schizophrenia (n¼ 20) Control (n¼ 20)
Age in years (range) 51.3 (27--75) 51.1 (18--78) 47.3 (27--75) 47.7 (18--74)Gender 13F:24M 7F:30M 20M 20Mhemisphere 17R:20L 23R:14L 7R:13L 11R:9LpH±s.d. 6.61±0.30 6.66±0.29 6.63±0.24 6.67±0.24PMI (hours) ±s.d. 28.46±13.77 24.80±10.97 29.59±16.11 24.23±11.06Months of freezer storage ±s.d. 78.89±37.24 69.62±42.71 91.05±36.03 94.20±40.34RIN ±s.d. 7.28±0.58 7.30±0.57 7.24±0.78 7.36±0.39Age (years) at onset (range) 23.70 (14--40) 23.9 (17--40)Duration of illness (years) ±s.d. 27.62±13.82 23.4±12.41Chlorpromazine median dose (mg) 691.64±502.20 594.94±280.11
Abbreviations: F, female; L, left; M, male; PMI, postmortem interval; R, right; RIN, RNA integrity number.Further demographic detail for individuals is available in Supplementary Table 1.
Increased inflammatory markers in schizophreniaSG Fillman et al
2
Molecular Psychiatry (2012), 1 -- 9 & 2012 Macmillan Publishers Limited

cence reagent (Millipore, Sydney, Australia), and visualized on a ChemidocImaging System (Bio-Rad). The expected B30-kDa band was quantitatedby densitometry using ImageJ (Version 1.4, rsbweb.nih.gov/ij/, U.S. NationalInstitute of Health, Bethesda, MD, USA). Each gel was normalized to aninternal control and b-actin expression, which did not differ betweendiagnostic conditions. The experiment was performed twice and dataaveraged.
Immunohistochemistry3,30-Diaminobenzidine immunohistochemistry was performed as pre-viously described.33 Fresh frozen tissue sections were thawed and fixedin 4% paraformaldehyde and then treated for peroxidase activity. Anti-HLA-DP/DR/DQ primary antibody (1: 1000, Dako; M0775) was appliedovernight at 4 1C, followed by washes and incubation with horse anti-mouse IgG biotinylated secondary antibody (1: 1000, Vector Laboratories,Burlingame, CA, USA). Slides were washed and incubated with avidin--biotin--peroxidase complex (Vectastain ABC kit; Vector Laboratories),treated with 3,30-Diaminobenzidine (Sigma, Castle Hill, NSW, Australia),washed, stained with Nissl and coverslipped.
Immunohistochemistry analysisImages for analysis were obtained on a Nikon Eclipse 80i microscope(Coherent Scientific, Hilton, SA, Australia) with a � 10 lens, numericalaperture (NA) 0.17 and stitched together using Stereoinvestigator software.Layers 3, 5/6 and white matter were identified and marked on each image.Five boxes of 0.3 mm2 were placed randomly 200--400mm apart withineach layer. HLA-DP/DR/DQ stained cells were counted (blind to diagnosis)within the boxes with 2 permissive and 2 non permissive edges, usingImageJ. Immuno-positive cells were defined as darkly 3,30-diaminobenzi-dine stained, with filamentous extensions and a Nissl nuclear-stained core.
Statistical analysisStatistical tests of qPCR data were performed using SPSS statistics (version19, OSX, IBM, Armonk, NY, USA). A Grubbs test was used to identifypopulation outliers in qPCR, immunohistochemistry, and western blotting(all outliers had high expression values, on average: n¼ 1.57, range: 0--5individuals per measurement per diagnostic group were dropped). FurtherqPCR statistical analysis was performed as detailed in Weickert et al.28 Owingto their initial nonnormal distributions, NFKB1, SERPINA3, IL-8, IL-6, IL6ST,
IL-1b and PTGS2 mRNA levels were log10 transformed with HLA-DP/DR/DQwestern blotting data being square root transformed. Pearson’s correlationswere performed between the immunohistochemical cell density of corticallayers in an individual. Stepwise regressions using demographic variables(age, pH, PMI, RIN) were used to identify covariates for analysis of covariance(ANCOVA) of experimentally measured mRNA values. Significantly identifieddemographic variables are indicated in the individual experimental results.Relationships between inflammatory mRNAs and HLA-DP/DR/DQ densitywere examined using stepwise regression and the transformed, normallydistributed, variables with missing values were replaced by standardmeans to enable retention of the greatest number of individuals. Theexistence of an inflammatory subgroup was tested using a recursive two-step cluster analysis on the entire case--control cohort with missing valuesreplaced by means.34 The overall model quality was required to be 40.5,with predictors removed if they did not contribute 40.5 to the model on ascale of 0--1.0. Characteristics of this subgroup were identified by usingt-tests for normally distributed continuous variables and w2 tests forcategorical. Individuals from this category were further examined forcorrelations with brain-derived neurotrophic factor (BDNF) and interneuronmarker mRNA expression using previously reported data.35,36 Spearman’scorrelation tests were performed on clinical variables of duration of diseaseand chlorpromazine dosages.
RESULTSThe inflammatory pathway is upregulated in schizophreniaProcessing of the NGS read counts in EdgeR yielded a total of 798differentially regulated transcripts in schizophrenia DLPFC ascompared with controls (FDRo0.05) (Supplementary Table 2).These transcripts were converted to genes using Ingenuity,yielding 316 genes, whose mRNA expression met the criteria forinclusion into our network analysis. Of these, 115 were core genesused to seed the network analysis. Ingenuity identified fivecustom networks and five biological functions (not mutuallyexclusive) that were significantly differentially regulated in oursample (Supplementary Table 3). The category of inflammatoryresponse reoccurred in these lists. We created a custom pathwayinto which we inserted the seven most changed and mostcommonly listed inflammatory genes (Figure 1). We confirmed thechanges detected by NGS by using qPCR in an expanded cohort
Figure 1. The proposed neuro-inflammatory pathway. An Ingenuity-created inflammatory pathway showing the mRNA expression of sevenmolecules along with HLA-DRA, a marker for antigen-presenting cells, including microglia. Red molecules indicate upregulated and grey nochange in schizophrenia with intensity determined by qPCR/western blotting. Solid lines show direct and dashed lines represent indirectrelationships. Each molecule has its log fold change (Log FC) and q-value (false discovery rate), as identified by EdgeR using Next GenerationSequencing data shown beside.
Increased inflammatory markers in schizophreniaSG Fillman et al
3
Molecular Psychiatry (2012), 1 -- 9& 2012 Macmillan Publishers Limited

(Figure 2). IL-6 and IL-8 mRNAs were significantly upregulated inschizophrenia (IL-6: ANCOVA pH: F(1,63)¼ 36.128, Po0.001, diag-nosis: F(1,63)¼ 3.985, Po0.05, IL-8: ANCOVA pH: F(1,60)¼ 9.751,P¼ 0.003, diagnosis: F(1,60)¼ 4.009, Po0.05, 107% and 39%increase, respectively). IL-1b mRNA showed a 29% (nonsignificant)increase in mRNA expression levels in individuals with schizo-phrenia (Figure 2b). Interestingly, a post hoc ANOVA analysisshowed significantly greater IL-1b mRNA expression levels inindividuals with schizophrenia as compared with those withschizoaffective disorder (F(2,64)¼ 3.436, P¼ 0.038, LSD (SCZ-Schi-zoaffective): P¼ 0.02). SERPINA3, a serine protease inhibitor, was themost significantly upregulated mRNA transcript among the qPCR-verified genes (ANCOVA: age F(1,55)¼ 4.495, P¼ 0.039, pH:F(1,55)¼ 4.085, P¼ 0.048, diagnosis: F(1,55)¼ 6.546, P¼ 0.013,106% increase in schizophrenia). PTGS2 mRNA, also known asCOX-2, was not significantly changed, as opposed to the significantelevation suggested by our NGS results (ANCOVA: freezertime: F(1,71)¼ 4.382, P¼ 0.040, diagnosis: F(1,71)¼ 2.461,P¼ 0.12). Two inflammatory related transcripts suggested byIngenuity, but not differentially regulated in the NGS data; IL6ST(IL-6 signal transducer also known as GP130) and NFKB1, were alsonot found to be significantly changed between our diagnosticgroups by qPCR.
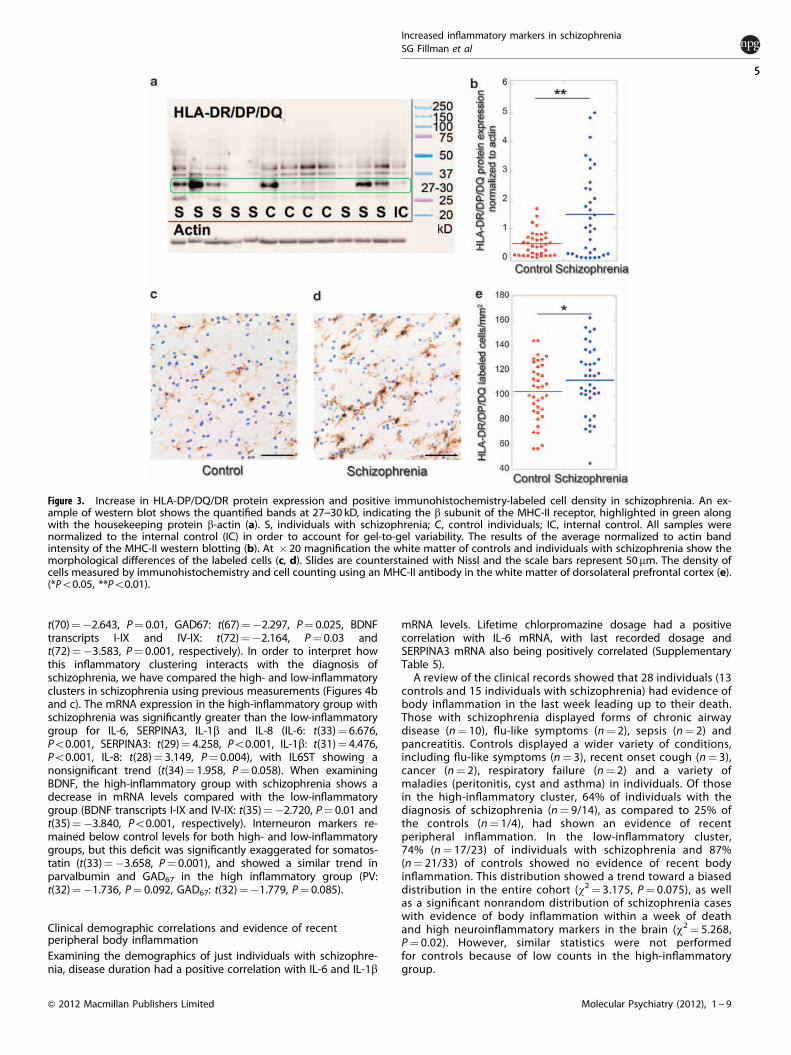
Increased microglial markers in schizophreniaThe HLA-DP/DR/DQ genes encode subunits of the MHC-II receptorexpressed on antigen-presenting cells, including microglia.Western blotting using an antibody specific for the b subunitcommon to the DP/DR/DQ subtypes showed a number of bands,but only one at the predicted 27--30 kD range. The intensity of thisband was highly varied between individuals but showed asignificant increase in schizophrenia (t(67)¼ 3.131, P¼ 0.002)(Figures 3a and b). Additionally, we found an increase in 8/10microglial mRNA markers which were reliably measured by NGSand significantly differentially expressed between individuals withschizophrenia and controls (Supplementary Table 4).
Antigen-presenting cells resembling microglia are increased inschizophreniaIn the white matter, a 9% increase in the density of microglia inpeople with schizophrenia, was found significant when taken in
context of the western blotting results and mRNA markers fromNGS (SCZ: 112.64.±3.68 cells mm�2, CON: 102.94 ± 4.25 cellsmm�2, t(71)¼ 1.930, P¼ 0.03, one-tailed t-test) (Figures 3c--e).There were fewer immuno-positive cells to be counted in the graymatter compared with the white matter, and no significantchanges were detectable in either layer III or V/VI (meancells mm�2 gray matter, SCZ: 50.36, CON: 48.98). However, wefound that microglia densities in both layers were significantlycorrelated with white matter cell densities. (all cases: layer 3:r¼ 0.546, Po0.0001, layer 5/6: r¼ 0.627, Po0.0001).
IL-1b is linked to MHC-II-expressing cells in the white matterTo establish if there was a link between the cells expressing MHC-IIreceptors and our other inflammatory markers, we performedstepwise regressions. The overall cohort variance in IL-1b mRNAlevels was positively linked to the density of MHC-II-positive cellsin the white matter (b¼ 0.319, F(1, 73)¼ 8.147, P¼ 0.006). Whenexamined separately by diagnostic group, this relationship wasseen only in individuals with schizophrenia and not in controls(b¼ 0.346, F(1, 36)¼ 4.771, P¼ 0.04).
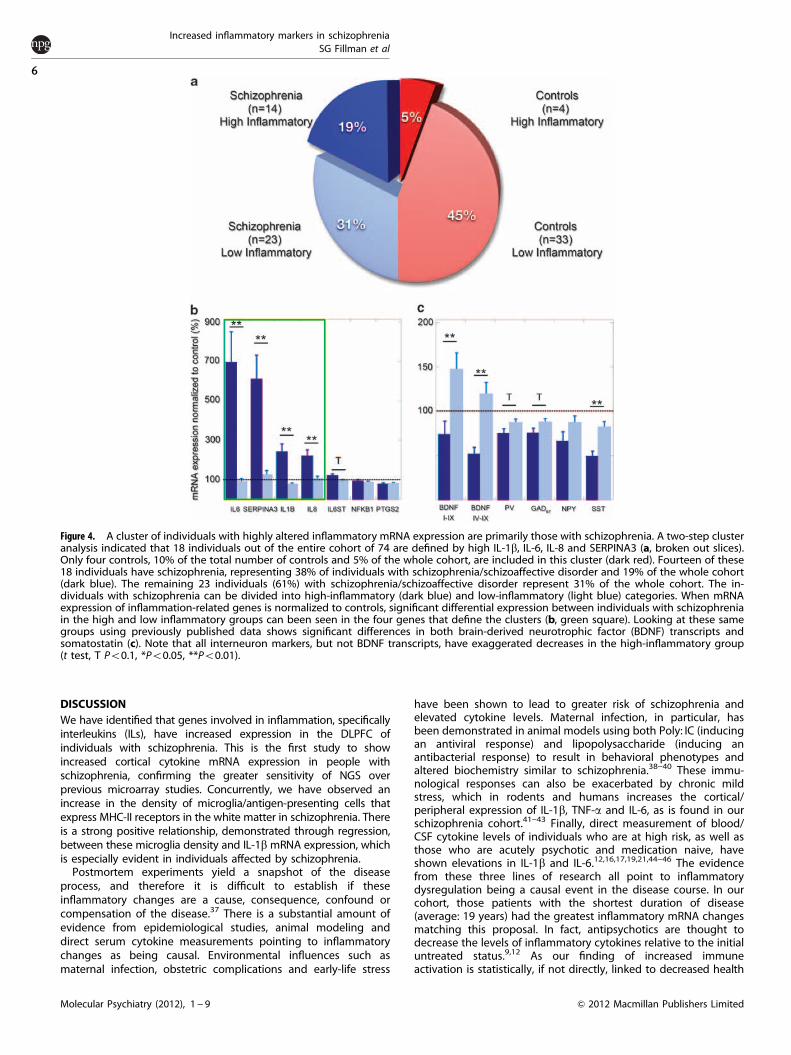
A subgroup of individuals with schizophrenia have elevatedinflammatory markersTwo-step cluster analysis on the entire cohort revealed 18 individualswho represented a high inflammatory factor group (Figure 4a). Theindividuals in this cluster were defined by high IL-1b, IL-6, IL-8 andSERPINA3 mRNA expression (all t44.7, all Po0.001). This group wasprimarily composed of individuals with schizophrenia as opposed tocontrols (n¼ 14 vs 4, w2¼ 7.341, P¼ 0.007). The high-inflammationgroup did not include any of the seven individuals withschizoaffective disorder (w2¼ 5.255, P¼ 0.02), any individuals withthe DSM-IV-defined depressive subtype of schizophrenia (w2¼ 4.359,P¼ 0.037) and had fewer individuals with recorded suicide attempts(w2¼ 5.451, P¼ 0.02). Other characteristics of this group include highmRNA expression of IL6ST (IL6ST t(70)¼ 2.174, P¼ 0.033). Also, whencomparing this high-inflammatory group to the low-inflammatorygroup (including both schizophrenia and controls), using previouslypublished inhibitory interneuron marker and BDNF changes in ourcohort, we find lower somatostatin (SST), parvalbumin, neuropeptideY, glutamate decarboxylase 67 and BDNF I-IX and IV-IX transcripts(SST: t(69)¼�4.370, Po0.001, PV: t(66)¼�2.802, P¼ 0.007, NPY:
Figure 2. Changes in inflammatory mRNA expression. Inflammatory mRNA expression as measured by quantitative PCR (qPCR). Highly variableextracellular ligands SERPINA3 (circles) and interleukin (IL)-6 (diamonds) are shown with a log scale (a). Other extracellular ligands IL-8 (triangles)and IL-1b (inverted triangles) are shown on a linear scale (b). Membrane and intercellular inflammatory proteins PTGS2 (circles), NFKB1 (dia-monds) and IL6ST (triangles) are also on a linear scale (c). Red indicates controls wheras blue are individuals with schizophrenia. (*Po0.05,**Po0.01).
Increased inflammatory markers in schizophreniaSG Fillman et al
4
Molecular Psychiatry (2012), 1 -- 9 & 2012 Macmillan Publishers Limited
t(70)¼�2.643, P¼ 0.01, GAD67: t(67)¼�2.297, P¼ 0.025, BDNFtranscripts I-IX and IV-IX: t(72)¼�2.164, P¼ 0.03 andt(72)¼�3.583, P¼ 0.001, respectively). In order to interpret howthis inflammatory clustering interacts with the diagnosis ofschizophrenia, we have compared the high- and low-inflammatoryclusters in schizophrenia using previous measurements (Figures 4band c). The mRNA expression in the high-inflammatory group withschizophrenia was significantly greater than the low-inflammatorygroup for IL-6, SERPINA3, IL-1b and IL-8 (IL-6: t(33)¼ 6.676,Po0.001, SERPINA3: t(29)¼ 4.258, Po0.001, IL-1b: t(31)¼ 4.476,Po0.001, IL-8: t(28)¼ 3.149, P¼ 0.004), with IL6ST showing anonsignificant trend (t(34)¼ 1.958, P¼ 0.058). When examiningBDNF, the high-inflammatory group with schizophrenia shows adecrease in mRNA levels compared with the low-inflammatorygroup (BDNF transcripts I-IX and IV-IX: t(35)¼�2.720, P¼ 0.01 andt(35)¼�3.840, Po0.001, respectively). Interneuron markers re-mained below control levels for both high- and low-inflammatorygroups, but this deficit was significantly exaggerated for somatos-tatin (t(33)¼�3.658, P¼ 0.001), and showed a similar trend inparvalbumin and GAD67 in the high inflammatory group (PV:t(32)¼�1.736, P¼ 0.092, GAD67: t(32)¼�1.779, P¼ 0.085).
Clinical demographic correlations and evidence of recentperipheral body inflammationExamining the demographics of just individuals with schizophre-nia, disease duration had a positive correlation with IL-6 and IL-1b
mRNA levels. Lifetime chlorpromazine dosage had a positivecorrelation with IL-6 mRNA, with last recorded dosage andSERPINA3 mRNA also being positively correlated (SupplementaryTable 5).
A review of the clinical records showed that 28 individuals (13controls and 15 individuals with schizophrenia) had evidence ofbody inflammation in the last week leading up to their death.Those with schizophrenia displayed forms of chronic airwaydisease (n¼ 10), flu-like symptoms (n¼ 2), sepsis (n¼ 2) andpancreatitis. Controls displayed a wider variety of conditions,including flu-like symptoms (n¼ 3), recent onset cough (n¼ 3),cancer (n¼ 2), respiratory failure (n¼ 2) and a variety ofmaladies (peritonitis, cyst and asthma) in individuals. Of thosein the high-inflammatory cluster, 64% of individuals with thediagnosis of schizophrenia (n¼ 9/14), as compared to 25% ofthe controls (n¼ 1/4), had shown an evidence of recentperipheral inflammation. In the low-inflammatory cluster,74% (n¼ 17/23) of individuals with schizophrenia and 87%(n¼ 21/33) of controls showed no evidence of recent bodyinflammation. This distribution showed a trend toward a biaseddistribution in the entire cohort (w2¼ 3.175, P¼ 0.075), as wellas a significant nonrandom distribution of schizophrenia caseswith evidence of body inflammation within a week of deathand high neuroinflammatory markers in the brain (w2¼ 5.268,P¼ 0.02). However, similar statistics were not performedfor controls because of low counts in the high-inflammatorygroup.
Figure 3. Increase in HLA-DP/DQ/DR protein expression and positive immunohistochemistry-labeled cell density in schizophrenia. An ex-ample of western blot shows the quantified bands at 27--30 kD, indicating the b subunit of the MHC-II receptor, highlighted in green alongwith the housekeeping protein b-actin (a). S, individuals with schizophrenia; C, control individuals; IC, internal control. All samples werenormalized to the internal control (IC) in order to account for gel-to-gel variability. The results of the average normalized to actin bandintensity of the MHC-II western blotting (b). At � 20 magnification the white matter of controls and individuals with schizophrenia show themorphological differences of the labeled cells (c, d). Slides are counterstained with Nissl and the scale bars represent 50 mm. The density ofcells measured by immunohistochemistry and cell counting using an MHC-II antibody in the white matter of dorsolateral prefrontal cortex (e).(*Po0.05, **Po0.01).
Increased inflammatory markers in schizophreniaSG Fillman et al
5
Molecular Psychiatry (2012), 1 -- 9& 2012 Macmillan Publishers Limited
DISCUSSIONWe have identified that genes involved in inflammation, specificallyinterleukins (ILs), have increased expression in the DLPFC ofindividuals with schizophrenia. This is the first study to showincreased cortical cytokine mRNA expression in people withschizophrenia, confirming the greater sensitivity of NGS overprevious microarray studies. Concurrently, we have observed anincrease in the density of microglia/antigen-presenting cells thatexpress MHC-II receptors in the white matter in schizophrenia. Thereis a strong positive relationship, demonstrated through regression,between these microglia density and IL-1b mRNA expression, whichis especially evident in individuals affected by schizophrenia.
Postmortem experiments yield a snapshot of the diseaseprocess, and therefore it is difficult to establish if theseinflammatory changes are a cause, consequence, confound orcompensation of the disease.37 There is a substantial amount ofevidence from epidemiological studies, animal modeling anddirect serum cytokine measurements pointing to inflammatorychanges as being causal. Environmental influences such asmaternal infection, obstetric complications and early-life stress
have been shown to lead to greater risk of schizophrenia andelevated cytokine levels. Maternal infection, in particular, hasbeen demonstrated in animal models using both Poly: IC (inducingan antiviral response) and lipopolysaccharide (inducing anantibacterial response) to result in behavioral phenotypes andaltered biochemistry similar to schizophrenia.38--40 These immu-nological responses can also be exacerbated by chronic mildstress, which in rodents and humans increases the cortical/peripheral expression of IL-1b, TNF-a and IL-6, as is found in ourschizophrenia cohort.41--43 Finally, direct measurement of blood/CSF cytokine levels of individuals who are at high risk, as well asthose who are acutely psychotic and medication naive, haveshown elevations in IL-1b and IL-6.12,16,17,19,21,44--46 The evidencefrom these three lines of research all point to inflammatorydysregulation being a causal event in the disease course. In ourcohort, those patients with the shortest duration of disease(average: 19 years) had the greatest inflammatory mRNA changesmatching this proposal. In fact, antipsychotics are thought todecrease the levels of inflammatory cytokines relative to the initialuntreated status.9,12 As our finding of increased immuneactivation is statistically, if not directly, linked to decreased health
Figure 4. A cluster of individuals with highly altered inflammatory mRNA expression are primarily those with schizophrenia. A two-step clusteranalysis indicated that 18 individuals out of the entire cohort of 74 are defined by high IL-1b, IL-6, IL-8 and SERPINA3 (a, broken out slices).Only four controls, 10% of the total number of controls and 5% of the whole cohort, are included in this cluster (dark red). Fourteen of these18 individuals have schizophrenia, representing 38% of individuals with schizophrenia/schizoaffective disorder and 19% of the whole cohort(dark blue). The remaining 23 individuals (61%) with schizophrenia/schizoaffective disorder represent 31% of the whole cohort. The in-dividuals with schizophrenia can be divided into high-inflammatory (dark blue) and low-inflammatory (light blue) categories. When mRNAexpression of inflammation-related genes is normalized to controls, significant differential expression between individuals with schizophreniain the high and low inflammatory groups can been seen in the four genes that define the clusters (b, green square). Looking at these samegroups using previously published data shows significant differences in both brain-derived neurotrophic factor (BDNF) transcripts andsomatostatin (c). Note that all interneuron markers, but not BDNF transcripts, have exaggerated decreases in the high-inflammatory group(t test, T Po0.1, *Po0.05, **Po0.01).
Increased inflammatory markers in schizophreniaSG Fillman et al
6
Molecular Psychiatry (2012), 1 -- 9 & 2012 Macmillan Publishers Limited

and viability of interneurons (specifically somatostatin positive), ourresults suggest that an increased neuroimmune response is linkedto interneuron distress, with further studies needed to confirm thetemporal pathological order.47--49
Microglia, responsible for much of the cortex’s cytokineproduction, have also been shown to be essential for learningand memory in the hippocampus.50,51 Much of this is donethrough modulation of IL-1b expression with increased levelsbeing detrimental to spatial and working memory whenadministered intracerebroventrally or genetically overex-pressed.52--54 As we find increased microglial staining and IL-1bmRNA expression in (a subset of cases with) schizophrenia, thiscould indicate a possible mechanism for the cognitive deficitsobserved in patients. Examining the number and density ofmicroglia cells in schizophrenia has been done using a number ofdifferent markers with varying results.55--60 In our study, theantibody binds to a common epitope on the b-subunit of theMHC-II receptor capturing the combined proteins of HLA-DP/DQ/DR. The MHC-II receptor is constitutively expressed by microgliaand other antigen-presenting cells, but is upregulated in responseto stress/activation.51 Four out of five postmortem studiesexamining HLA-DP/DQ/DR or HLA-DR specifically found quantita-tive or qualitative increases in schizophrenia.55--59 Two in vivo PETand one in vitro binding study used [11C]PK11195 as a ligand forthe translocater protein (formerly peripheral-type benzodiazepinereceptor), a marker for microglia, and found increased gray matterand hippocampus binding, although an in vitro study founddecreases in superior parietal cortex, primary visual cortex andputamen.25--27 It is also notable that only Doorduin et al.25
reported on white matter and observed a 20%, nonsignificantincrease. The small number of subjects in these PET studies(n¼ 16--32) suggests that cohort size may result in theseinconsistent findings, especially when taken in context of theheterogeneity present in our (n¼ 74) cohort.
Our study is the first to establish mRNA changes in inflamma-tory cytokines in the DLPFC of individuals with schizophrenia. Thegreater sensitivity of NGS has allowed us to form an important linkbetween previously observed peripheral immune-related changesand those in the cortex. Elevated IL-1b expression is known tocause increased secretion of IL-6 from microglia, astrocytes andneurons.61--63 This IL-6 elevation could subsequently lead toproblems in cell migration, which may be one reason for thegreater density of inhibitory interneurons in the white matter ofindividuals with schizophrenia.33,64 The IL-6 receptor has beenshown to exist primarily in neurons, and one study suggestsspecifically in parvalbumin-positive interneurons, although glialexpression has been found as well.65--69 This fits well with themodel proposed by Behrens et al.47--49 of IL-6-mediated produc-tion of reactive oxidative species through the NADPH-oxidasepathway leading to deficits in interneurons.47 Although our resultsshow elevated IL-6, as well as decreased inhibitory interneuronmarkers such as somatostatin, we do not find any significantchanges in NOX2 (CYBB); the models proposed intermediateenzyme, any of the necessary cofactors (CYBA, NCF1 and NCF2) ora change in the IL6ST, required for IL-6 signal transductionbetween schizophrenia and controls (data not shown). It ispossible, however, that posttranscriptional changes such asprotein stability or enzyme activity levels, which we did notmeasure, are responsible for the increased activity of NOX2observed previously with high IL-6 expression.48,49 Others haveobserved a negative relationship between IL-6 and BDNF levels inschizophrenia patients, which may explain why the BDNF levelsfor 50 exons I and IV were significantly decreased in theschizophrenia patients who are members of the high-inflamma-tory cluster compared with those patients in the low-inflammatorycluster.70 The individuals in the low-inflammatory cluster showhigher than control levels of BDNF mRNA (40--50%), which isconsistent with our previously observed overall increase in
schizophrenia BDNF I-IX expression in our cohort.36 However, inexon IV-IX, we previously found no diagnostic change butcurrently observe increased mRNA expression in the low-inflammatory cluster, and a decrease in BDNF mRNA in thehigh-inflammatory cluster when compared with controls suggest-ing a great deal of heterogeneity in cortical BDNF mRNA levelsthat may be related to neuroinflammation.36 The decrease inBDNF mRNA expression in the high-inflammatory patients mayalso account for the exacerbated somatostatin mRNA decrease inthe group with schizophrenia.71 In addition to IL-1bs known,modulation of IL-6, IL-1b increases can also result in elevations ofIL-8.72 Indeed, we find significant increases in IL-8 mRNA, and asIL-8 is a potent chemoattractant for a number of immune-relatedcells including T-cells, neutrophils, basophils and endothelial cells,it is possible that the blood-brain-barrier may be disrupted inschizophrenia.73 This may explain the observed synergisticincreases in brain and blood cytokines of individuals withschizophrenia, and could allow for identification of ‘brain events’through the blood.11,74,75 A greater number of individuals withschizophrenia in our cohort display evidence of concurrentperipheral and central inflammatory response than controls,perhaps indicating a predisposition toward increased inflamma-tory response from a genetic or environmental source.76,77
However, a number of individuals with schizophrenia that showno evidence of peripheral body inflammation still displayindications of an increased brain inflammatory response and anumber of controls with evidence of inflammation in the body donot display elevated levels of brain inflammatory markers. Thiswould suggest that although there may be an interaction betweenbody and brain inflammatory responses, particularly in schizo-phrenia, it does not appear to co-occur in all cases. As a substantialnumber of individuals with schizophrenia seem to have anabnormal expression of inflammatory factors, it should be made apriority to identify these individuals and determine whichtreatments, including prostaglandin synthesis inhibitors andbeyond, will be most beneficial.11,74,78 Targeted supplementalintervention of anti-inflammatory agents may be an effectivetreatment currently available to improve the quality of life ofindividuals with schizophrenia.79
CONFLICT OF INTERESTThe authors declare no conflict of interest.
ACKNOWLEDGEMENTSWe would like to thank Sarah Wagner at Queensland Pathology for performingthe SOLiD library preparation and sequencing. We would also like to thankShan Yuan-Tsai-Chin for her assistance in RNA isolation and Dr Claire Shepherd forher input into the microglia immunohistochemistry analysis. This work wassupported by the Schizophrenia Research Institute (Australia), utilizing funding fromthe Macquarie Group Foundation, New South Wales Ministry of Health, NeuroscienceResearch Australia, and the University of New South Wales. Tissues werereceived from the New South Wales Tissue Resource Center at the University ofSydney, which is supported by the National Health and Medical Research Council ofAustralia, Schizophrenia Research Institute, National Institute of Alcohol Abuse andAlcoholism (NIH (NIAAA) R24AA012725). JW is supported by a National Health andMedical Research Council of Australia (NHMRC) Postdoctoral Training Fellowship(568884).
REFERENCES1 Arion D, Unger T, Lewis DA, Levitt P, Mirnics K. Molecular evidence for increased
expression of genes related to immune and chaperone function in the prefrontalcortex in schizophrenia. Biol Psychiatry 2007; 62: 711--721.
2 Aston C, Jiang L, Sokolov BP. Microarray analysis of postmortem temporal cortexfrom patients with schizophrenia. J Neurosci Res 2004; 77: 858--866.
3 Hakak Y, Walker JR, Li C, Wong WH, Davis KL, Buxbaum JD et al. Genome-wideexpression analysis reveals dysregulation of myelination-related genes in chronicschizophrenia. Proc Natl Acad Sci USA 2001; 98: 4746--4751.
Increased inflammatory markers in schizophreniaSG Fillman et al
7
Molecular Psychiatry (2012), 1 -- 9& 2012 Macmillan Publishers Limited

4 Maycox PR, Kelly F, Taylor A, Bates S, Reid J, Logendra R et al. Analysis of geneexpression in two large schizophrenia cohorts identifies multiple changesassociated with nerve terminal function. Mol Psychiatry 2009; 14: 1083--1094.
5 Saetre P, Emilsson L, Axelsson E, Kreuger J, Lindholm E, Jazin E. Inflammation-related genes up-regulated in schizophrenia brains. BMC Psychiatry 2007; 7: 46.
6 Mirnics K, Middleton FA, Marquez A, Lewis DA, Levitt P. Molecular characterizationof schizophrenia viewed by microarray analysis of gene expression in prefrontalcortex. Neuron 2000; 28: 53--67.
7 Wang Z, Gerstein M, Snyder M. RNA-Seq: a revolutionary tool for transcriptomics.Nat Rev Genet 2009; 10: 57--63.
8 Okoniewski MJ, Miller CJ. Hybridization interactions between probesets in shortoligo microarrays lead to spurious correlations. BMC Bioinformatics 2006; 7: 276.
9 Erbagci AB, Herken H, Koyluoglu O, Yilmaz N, Tarakcioglu M. Serum IL-1beta, sIL-2R, IL-6, IL-8 and TNF-alpha in schizophrenic patients, relation with symptomatol-ogy and responsiveness to risperidone treatment. Mediators Inflamm 2001; 10:109--115.
10 Ganguli R, Yang Z, Shurin G, Chengappa KN, Brar JS, Gubbi AV et al. Seruminterleukin-6 concentration in schizophrenia: elevation associated with durationof illness. Psychiatry Res 1994; 51: 1--10.
11 Laan W, Grobbee DE, Selten J-P, Heijnen CJ, Kahn RS, Burger H. Adjuvant aspirintherapy reduces symptoms of schizophrenia spectrum disorders: results from arandomized, double-blind, placebo-controlled trial. J Clin Psychiatry 2010; 71:520--527.
12 Theodoropoulou S, Spanakos G, Baxevanis CN, Economou M, Gritzapis AD,Papamichail MP et al. Cytokine serum levels, autologous mixed lymphocytereaction and surface marker analysis in never medicated and chronicallymedicated schizophrenic patients. Schizophr Res 2001; 47: 13--25.
13 Xu M, He L. Convergent evidence shows a positive association of interleukin-1gene complex locus with susceptibility to schizophrenia in the Caucasianpopulation. Schizophr Res 2010; 120: 131--142.
14 Schwab SG, Hallmayer J, Freimann J, Lerer B, Albus M, Borrmann-Hassenbach Met al. Investigation of linkage and association/linkage disequilibrium of HLA A-,DQA1-, DQB1-, and DRB1-alleles in 69 sib-pair- and 89 trio-families withschizophrenia. Am J Med Genet 2002; 114: 315--320.
15 Ripke S, Sanders AR, Kendler KS, Levinson DF, Sklar P, Holmans PA et al. Genome-wide association study identifies five new schizophrenia loci. Nat Genet 2011; 43:969--976.
16 van Kammen D, McAllister-Sistilli C, Kelley M, Gurklis J, Yao J. Elevated interleukin-6 in schizophrenia. Psychiatry Res 1999; 87: 129--136.
17 Kunz M, Cereser KM, Goi PD, Fries GR, Teixeira AL, Fernandes BS et al. Serum levelsof IL-6, IL-10 and TNF-a in patients with bipolar disorder and schizophrenia:differences in pro- and anti-inflammatory balance. Rev Bras Psiquiatr 2011; e-pubahead of print.
18 Lin A, Kenis G, Bignotti S, Tura GJ, De Jong R, Bosmans E et al. The inflammatoryresponse system in treatment-resistant schizophrenia: increased serum inter-leukin-6. Schizophr Res 1998; 32: 9--15.
19 Naudin J, Mege JL, Azorin JM, Dassa D. Elevated circulating levels of IL-6 inschizophrenia. Schizophr Res 1996; 20: 269--273.
20 Zhang XY, Zhou DF, Cao LY, Zhang PY, Wu GY, Shen YC. Changes in seruminterleukin-2, -6, and -8 levels before and during treatment with risperidone andhaloperidol: relationship to outcome in schizophrenia. J Clin Psychiatry 2004; 65:940--947.
21 Zhang XY, Zhou DF, Zhang PY, Wu GY, Cao LY, Shen YC. Elevated interleukin-2,interleukin-6 and interleukin-8 serum levels in neuroleptic-free schizophrenia:association with psychopathology. Schizophr Res 2002; 57: 247--258.
22 Aloisi F. Immune function of microglia. Glia 2001; 36: 165--179.23 Frank MG, Baratta MV, Sprunger DB, Watkins LR, Maier SF. Microglia serve as a
neuroimmune substrate for stress-induced potentiation of CNS pro-inflammatorycytokine responses. Brain Behav Immun 2007; 21: 47--59.
24 Kettenmann H, Hanisch U-K, Noda M, Verkhratsky A. Physiology of microglia.Physiol Rev 2011; 91: 461--553.
25 Doorduin J, de Vries EFJ, Willemsen ATM, de Groot JC, Dierckx RA, Klein HC.Neuroinflammation in schizophrenia-related psychosis: a PET study. J Nucl Med2009; 50: 1801--1807.
26 van Berckel BN, Bossong MG, Boellaard R, Kloet R, Schuitemaker A, Caspers E et al.Microglia activation in recent-onset schizophrenia: a quantitative (R)-[11C]PK11195positron emission tomography study. Biol Psychiatry 2008; 64: 820--822.
27 Kurumaji A, Wakai T, Toru M. Decreases in peripheral-type benzodiazepinereceptors in postmortem brains of chronic schizophrenics. J Neural Transm 1997;104: 1361--1370.
28 Weickert CS, Sheedy D, Rothmond DA, Dedova I, Fung S, Garrick T et al. Selectionof reference gene expression in a schizophrenia brain cohort. Aust N Z J Psychiatry2010; 44: 59--70.
29 Wood DLA, Xu Q, Pearson JV, Cloonan N, Grimmond SM. X-MATE: a flexiblesystem for mapping short read data. Bioinformatics 2011; 27: 580--581.
30 Koehler R, Issac H, Cloonan N, Grimmond SM. The uniqueome: a mappabilityresource for short-tag sequencing. Bioinformatics 2011; 27: 272--274.
31 Weickert CS, Elashoff M, Richards AB, Sinclair D, Bahn S, Paabo S et al.Transcriptome analysis of male--female differences in prefrontal cortical devel-opment. Mol Psychiatry 2009; 14: 558--561.
32 Wong J, Webster MJ, Cassano H, Weickert CS. Changes in alternative brain-derivedneurotrophic factor transcript expression in the developing human prefrontalcortex. Eur J Neurosci 2009; 29: 1311--1322.
33 Yang Y, Fung SJ, Rothwell A, Tianmei S, Weickert CS. Increased interstitial whitematter neuron density in the dorsolateral prefrontal cortex of people withschizophrenia. Biol Psychiatry 2011; 69: 63--70.
34 Bacher J, Wenzig K, Vogler M. SPSS twostep cluster: A first evaluation. LehrstuhlSoziol 2004.
35 Fung SJ, Webster MJ, Sivagnanasundaram S, Duncan C, Elashoff M, Weickert CS.Expression of interneuron markers in the dorsolateral prefrontal cortex ofthe developing human and in schizophrenia. Am J Psychiatry 2010; 167:1479--1488.
36 Wong J, Hyde TM, Cassano HL, Deep-Soboslay A, Kleinman JE, Weickert CS.Promoter specific alterations of brain-derived neurotrophic factor mRNA inschizophrenia. Neuroscience 2010; 169: 1071--1084.
37 Lewis DA, Gonzalez-Burgos G. Neuroplasticity of neocortical circuits in schizo-phrenia. Neuropsychopharmacology 2008; 33: 141--165.
38 Bilbo SD, Barrientos RM, Eads AS, Northcutt A, Watkins LR, Rudy JW et al. Early-lifeinfection leads to altered BDNF and IL-1beta mRNA expression in rathippocampus following learning in adulthood. Brain Behav Immun 2008; 22:451--455.
39 Bitanihirwe BKY, Peleg-Raibstein D, Mouttet F, Feldon J, Meyer U. Late prenatalimmune activation in mice leads to behavioral and neurochemical abnormalitiesrelevant to the negative symptoms of schizophrenia. Neuropsychopharmacology2010; 35: 2462--2478.
40 Oskvig DB, Elkahloun AG, Johnson KR, Phillips TM, Herkenham M. Maternalimmune activation by LPS selectively alters specific gene expression profiles ofinterneuron migration and oxidative stress in the fetus without triggering a fetalimmune response. Brain Behav Immun 2012.
41 Ackerman KD, Martino M, Heyman R, Moyna NM, Rabin BS. Stressor-inducedalteration of cytokine production in multiple sclerosis patients and controls.Psychosom Med 1998; 60: 484--491.
42 Maes M, Song C, Lin A, De Jongh R, Van Gastel A, Kenis G et al. The effects ofpsychological stress on humans: increased production of pro-inflammatorycytokines and a Th1-like response in stress-induced anxiety. Cytokine 1998; 10:313--318.
43 You Z, Luo C, Zhang W, Chen Y, He J, Zhao Q et al. Pro- and anti-inflammatorycytokines expression in rat0s brain and spleen exposed to chronic mild stress:involvement in depression. Behav Brain Res 2011; 225: 135--141.
44 Sirota P, Schild K, Elizur A, Djaldetti M, Fishman P. Increased interleukin-1 andinterleukin-3 like activity in schizophrenic patients. Prog Neuro-PsychopharmacolBiol Psychiatry 1995; 19: 75--83.
45 Kim Y-K, Myint A-M, Verkerk R, Scharpe S, Steinbusch H, Leonard B. Cytokinechanges and tryptophan metabolites in medication-naıve and medication-freeschizophrenic patients. Neuropsychobiology 2009; 59: 123--129.
46 Kim YK, Kim L, Lee MS. Relationships between interleukins, neurotransmittersand psychopathology in drug-free male schizophrenics. Schizophr Res 2000; 44:165--175.
47 Behrens MM, Ali SS, Dao DN, Lucero J, Shekhtman G, Quick KL et al. Ketamine-induced loss of phenotype of fast-spiking interneurons is mediated by NADPH-oxidase. Science 2007; 318: 1645--1647.
48 Behrens MM, Ali SS, Dugan LL. Interleukin-6 mediates the increase in NADPH-oxidase in the ketamine model of schizophrenia. J Neurosci 2008; 28: 13957--13966.
49 Dugan LL, Ali SS, Shekhtman G, Roberts AJ, Lucero J, Quick KL et al. IL-6 mediateddegeneration of forebrain GABAergic interneurons and cognitive impairment inaged mice through activation of neuronal NADPH oxidase. PloS ONE 2009; 4:e5518.
50 Williamson LL, Sholar PW, Mistry RS, Smith SH, Bilbo SD. Microglia and memory:modulation by early-life infection. J Neurosci 2011; 31: 15511--15521.
51 Tambuyzer BR, Ponsaerts P, Nouwen EJ. Microglia: gatekeepers of central nervoussystem immunology. J Leukoc Biol 2009; 85: 352--370.
52 Gonzalez PV, Schioth HB, Lasaga M, Scimonelli TN. Memory impairment inducedby IL-1beta is reversed by alpha-MSH through central melanocortin-4 receptors.Brain Behav Immun 2009; 23: 817--822.
53 Hein AM, Stasko MR, Matousek SB, Scott-McKean JJ, Maier SF, Olschowka JA et al.Sustained hippocampal IL-1beta overexpression impairs contextual and spatialmemory in transgenic mice. Brain Behav Immun 2010; 24: 243--253.
54 Oitzl MS, van Oers H, Schobitz B, de Kloet ER. Interleukin-1 beta, but notinterleukin-6, impairs spatial navigation learning. Brain Res 1993; 613: 160--163.
Increased inflammatory markers in schizophreniaSG Fillman et al
8
Molecular Psychiatry (2012), 1 -- 9 & 2012 Macmillan Publishers Limited

55 Bayer TA, Buslei R, Havas L, Falkai P. Evidence for activation of microglia inpatients with psychiatric illnesses. Neurosci Lett 1999; 271: 126--128.
56 Radewicz K, Garey LJ, Gentleman SM, Reynolds R. Increase in HLA-DRimmunoreactive microglia in frontal and temporal cortex of chronic schizo-phrenics. J Neuropathol Exp Neurol 2000; 59: 137--150.
57 Steiner J, Bielau H, Brisch R, Danos P, Ullrich O, Mawrin C et al. Immunologicalaspects in the neurobiology of suicide: elevated microglial density in schizophreniaand depression is associated with suicide. J Psychiatr Res 2008; 42: 151--157.
58 Togo T, Akiyama H, Kondo H, Ikeda K, Kato M, Iseki E et al. Expression of CD40 inthe brain of Alzheimer0s disease and other neurological diseases. Brain Res 2000;885: 117--121.
59 Wierzba-Bobrowicz T, Lewandowska E, Lechowicz W, Stepien T, Pasennik E.Quantitative analysis of activated microglia, ramified and damage of processes inthe frontal and temporal lobes of chronic schizophrenics. Folia Neuropathol 2005;43: 81--89.
60 Steiner J, Mawrin C, Ziegeler A, Bielau H, Ullrich O, Bernstein H-G et al. Distributionof HLA-DR-positive microglia in schizophrenia reflects impaired cerebrallateralization. Acta Neuropathol 2006; 112: 305--316.
61 Aloisi F, Care A, Borsellino G, Gallo P, Rosa S, Bassani A et al. Production ofhemolymphopoietic cytokines (IL-6, IL-8, colony-stimulating factors) by normalhuman astrocytes in response to IL-1 beta and tumor necrosis factor-alpha. JImmunol 1992; 149: 2358--2366.
62 Lee SC, Liu W, Dickson DW, Brosnan CF, Berman JW. Cytokine production byhuman fetal microglia and astrocytes. Differential induction by lipopolysaccharideand IL-1 beta. J Immunol 1993; 150: 2659--2667.
63 Ringheim GE, Burgher KL, Heroux JA. Interleukin-6 mRNA expression by corticalneurons in culture: evidence for neuronal sources of interleukin-6 production inthe brain. J Neuroimmunol 1995; 63: 113--123.
64 Wei H, Zou H, Sheikh AM, Malik M, Dobkin C, Brown WT et al. IL-6 is increased inthe cerebellum of autistic brain and alters neural cell adhesion, migration andsynaptic formation. J Neuroinflammation 2011; 8: 52.
65 Burton MD, Sparkman NL, Johnson RW. Inhibition of interleukin-6 trans-signalingin the brain facilitates recovery from lipopolysaccharide-induced sicknessbehavior. J Neuroinflammation 2011; 8: 54.
66 Vollenweider F, Herrmann M, Otten U, Nitsch C. Interleukin-6 receptor expressionand localization after transient global ischemia in gerbil hippocampus. NeurosciLett 2003; 341: 49--52.
67 Gadient RA, Otten U. Interleukin-6 and interleukin-6 receptor mRNA expression inrat central nervous system. Ann NY Acad Sci 1995; 762: 403--406.
68 Islam O, Gong X, Rose-John S, Heese K. Interleukin-6 and neural stem cells: morethan gliogenesis. Mol Biol Cell 2009; 20: 188--199.
69 Schobitz B, de Kloet ER, Sutanto W, Holsboer F. Cellular localization of interleukin6 mRNA and interleukin 6 receptor mRNA in rat brain. Eur J Neurosci 1993; 5:1426--1435.
70 Mondelli V, Cattaneo A, Belvederi Murri M, Di Forti M, Handley R, Hepgul N et al.Stress and inflammation reduce brain-derived neurotrophic factor expression infirst-episode psychosis: a pathway to smaller hippocampal volume. J ClinPsychiatry 2011; 72: 1677--1684.
71 Sanchez-Munoz I, Sanchez-Franco F, Vallejo M, Fernandez A, Palacios N,Fernandez M et al. Regulation of somatostatin gene expression by brainderived neurotrophic factor in fetal rat cerebrocortical cells. Brain Res 2011; 1375:28--40.
72 Ehrlich LC, Hu S, Sheng WS, Sutton RL, Rockswold GL, Peterson PK et al. Cytokineregulation of human microglial cell IL-8 production. J Immunol 1998; 160:1944--1948.
73 Huber AR, Kunkel SL, Todd RF, Weiss SJ. Regulation of transendothelial neutrophilmigration by endogenous interleukin-8. Science 1991; 254: 99--102.
74 Muller N, Krause D, Dehning S, Musil R, Schennach-Wolff R, Obermeier M et al.Celecoxib treatment in an early stage of schizophrenia: results of a randomized,double-blind, placebo-controlled trial of celecoxib augmentation of amisulpridetreatment. Schizophr Res 2010; 121: 118--124.
75 Levin Y, Wang L, Schwarz E, Koethe D, Leweke FM, Bahn S. Global proteomicprofiling reveals altered proteomic signature in schizophrenia serum. MolPsychiatry 2010; 15: 1088--1100.
76 Brown AS. The environment and susceptibility to schizophrenia. Prog Neurobiol2011; 93: 23--58.
77 Zakharyan R, Petrek M, Arakelyan A, Mrazek F, Atshemyan S, Boyajyan A.Interleukin-6 promoter polymorphism and plasma levels in patients withschizophrenia. Tissue Antigens 2012; e-pub ahead of print.
78 Akhondzadeh S, Tabatabaee M, Amini H, Ahmadi Abhari SA, Abbasi SH, BehnamB. Celecoxib as adjunctive therapy in schizophrenia: a double-blind, randomizedand placebo-controlled trial. Schizophr Res 2007; 90: 179--185.
79 Muller N, Schwarz MJ. Immune System and Schizophrenia. Curr Immunol Rev2010; 6: 213--220.
Supplementary Information accompanies the paper on the Molecular Psychiatry website (http://www.nature.com/mp)
Increased inflammatory markers in schizophreniaSG Fillman et al
9
Molecular Psychiatry (2012), 1 -- 9& 2012 Macmillan Publishers Limited