inactivation of escherichia coli o157:h7 during thermophilic anaerobic digestion of manure from...
TRANSCRIPT

ARTICLE IN PRESS
Available at www.sciencedirect.com
WAT E R R E S E A R C H 4 1 ( 2 0 0 7 ) 1 6 5 9 – 1 6 6 6
0043-1354/$ - see frodoi:10.1016/j.watres
�Corresponding auE-mail address:
journal homepage: www.elsevier.com/locate/watres
Inactivation of Escherichia coli O157:H7 during thermophilicanaerobic digestion of manure from dairy cattle
Michael D. Aitken�, Mark D. Sobsey, Nicole A. Van Abel, Kimberly E. Blauth,David R. Singleton, Phillip L. Crunk, Cora Nichols, Glenn W. Walters, Maria Schneider
Department of Environmental Sciences and Engineering, School of Public Health, University of North Carolina, Chapel Hill,
NC 27599-7431, USA
a r t i c l e i n f o
Article history:
Received 14 August 2006
Received in revised form
19 January 2007
Accepted 28 January 2007
Available online 13 March 2007
Keywords:
Thermophilic anaerobic digestion
Animal waste
Dairy cattle
E. coli O157:H7
Inactivation
nt matter & 2007 Elsevie.2007.01.034
thor. Tel.: +1 919 966 1024;[email protected] (M
a b s t r a c t
Inactivation of the pathogenic Escherichia coli serotype O157:H7 and a non-pathogenic E. coli
strain isolated from dairy cattle manure was evaluated with batch tests at 50 and 55 1C in
biosolids from a thermophilic anaerobic digester treating the manure. Using differential-
selective plating on sorbitol-MacConkey (SMAC) agar to quantify E. coli, the decline in
concentrations of both the sorbitol-negative (putative E. coli O157:H7) and sorbitol-positive
(putative non-pathogenic E. coli) organisms followed a model that assumed there was a
heat-sensitive fraction and a heat-resistant fraction. Inactivation rates of the heat-sensitive
fractions were similar for both colony types at each temperature, suggesting that wild-type
E. coli can be used as an indicator of inactivation of serotype O157:H7. The decimal
reduction time for the heat-sensitive fractions was in the order of 10 min at 55 1C and
ranged from approximately 1–3 h at 50 1C. Concentrations of heat-resistant organisms at
55 1C were 1.4–1.7 log10 cfu/mL. Confirmatory analyses conducted on 30 randomly selected
colonies of heat-resistant sorbitol-negative cells from treatment at 55 1C indicated that
none were serotype O157:H7, nor were they E. coli. Similar analyses on 10 sorbitol-negative
isolates from untreated manure indicated that none were serotype O157:H7, although 16S
rRNA gene sequence analysis indicated that eight were E. coli or closely related enteric
bacteria. These findings suggest that plating on differential-selective media to quantify E.
coli, including serotype O157:H7, in effluent samples from thermophilic anaerobic digestion
can lead to false positive results. Therefore, more specific methods should be used to
evaluate the extent of thermal inactivation of both pathogenic and non-pathogenic E. coli in
manure treatment systems.
& 2007 Elsevier Ltd. All rights reserved.
1. Introduction
Escherichia coli serotype O157:H7 is a widespread human
pathogen responsible for a high incidence of gastrointestinal
illnesses, including hemorrhagic colitis (Armstrong et al.,
1996; Mead and Griffin, 1998; Duffy, 2003; Muniesa et al., 2006).
Serotype O157:H7 and other enterohemorrhagic strains of
E. coli are believed to be involved in 90% of cases of post-
r Ltd. All rights reserved.
fax: +1 919 966 7911..D. Aitken).
diarrheal hemolytic uremic syndrome, which can lead to
severe renal injury that is potentially fatal (Mead and Griffin,
1998; Todd and Dundas, 2001). Both diseased and healthy
cattle as well as other farm animals are reservoirs of this
pathogen (Armstrong et al., 1996; Todd and Dundas, 2001;
Djordjevic et al., 2004). In addition to transmission through
contaminated meat and milk of infected animals, E. coli
O157:H7 can contaminate vegetable crops through the use of

ARTICLE IN PRESS
WAT E R R E S E A R C H 4 1 ( 2 0 0 7 ) 1 6 5 9 – 1 6 6 61660
contaminated manure or irrigation water on agricultural
fields (Mead and Griffin, 1998; Todd and Dundas, 2001). Direct
contact with the edible parts of plants is not the only basis for
contamination, as the organism can be taken up from
contaminated water through the plant roots (Solomon et al.,
2002). The feces of infected animals can contain concentra-
tions of serotype O157:H7 as high as 104–105 cfu or MPN/g
(Zhao et al., 1995; Omisakin et al., 2003; Fegan et al., 2003). The
organism can survive in manure for 2–3 months in the field
(Duffy, 2003; Nicholson et al., 2005), and recent data suggest
that it can grow in animal bedding and feedlot soils (Davis et
al., 2005; Berry and Miller, 2005).
The prevalence of E. coli O157:H7 in animal feces, its ability
to survive in manure during storage, and the widespread
practice of using manure for fertilizer on agricultural fields,
suggest that methods of treating manure to inactivate this
and other human or animal pathogens should be investi-
gated. The objective of this study was to evaluate the
inactivation of serotype O157:H7 under conditions relevant
to the thermophilic anaerobic digestion of manure from dairy
cattle. The rates of inactivation of E. coli O157:H7 and a non-
pathogenic indigenous strain of E. coli isolated from the
manure were measured at 50 and 55 1C in a batch reactor
containing digested manure from a laboratory scale, contin-
uous-flow digester.
2. Materials and methods
Manure was obtained as grab samples of fresh barn washings
flowing into a settling pit at the experimental dairy cattle
farm at North Carolina State University (Raleigh, North
Carolina, USA). The samples were combined in a drum
and processed with a submersible grinder pump to reduce
the size of the undigested straw and other large solids that
otherwise interfered with pumping into and out of the
laboratory digester. Processed manure was stored cold
(approximately 5 1C) for up to 8 weeks before feeding to the
digester.
E. coli O157:H7 (ATCC 43895) was obtained from Lee-Ann
Jaykus, Department of Food Science, North Carolina State
University. This strain is positive for both Shiga toxin 1 and
Shiga toxin 2. A non-pathogenic strain of E. coli was isolated
from a sample of manure as follows. The manure was serially
diluted into phosphate-buffered saline and several of the
dilutions were filtered through 0.45mm (pore size) gridded
membrane filters. The filters were placed on plates of mFC
agar (BD Difco; Franklin Lakes, NJ), incubated at 37 1C for 2 h
and then at 44.5 1C for 24 h. Filters were transferred, in the
same orientation, to plates of nutrient agar containing
4-methylumbelliferyl-B-D-glucuronide (MUG) and incubated
at 44.5 1C for 4 h. Colonies that were blue on the mFC plates
and fluoresced blue under long-wavelength UV light on the
nutrient agar-MUG plates were considered putative non-
pathogenic E. coli. Candidate colonies were streaked onto
plates containing tryptic soy agar (TSA) and incubated at
44.5 1C for 24 h. Two more rounds of picking colonies and re-
streaking onto TSA were conducted before selecting several
colonies for further analysis. Selected colonies were analyzed
with Enterotubes II test kits (BD, Franklin Lakes, New Jersey,
USA), and one that exhibited typical E. coli results was
selected for subsequent inactivation experiments. This iso-
late was confirmed not to be serotype O157:H7 by the
immunoassay described below.
2.1. Experimental design and laboratory system
Details of the laboratory thermophilic anaerobic digestion
system are provided elsewhere (Aitken et al., 2005b, c). Briefly,
a 16-L thermophilic anaerobic digester was operated to
simulate continuous flow by pumping influent manure and
effluent biosolids intermittently over short cycles (1 min on
and 15 min off). The digester was operated at 55 1C and for
two different periods at 50 1C (designated 50A and 50B) with a
hydraulic residence time of 7.8 days. For a given operating
condition, only those data obtained after at least one
residence time had elapsed are reported. Digestion perfor-
mance was evaluated by monitoring gas flow and composi-
tion, pH, total solids (TS), volatile solids (VS), volatile fatty
acids (VFAs), and ammonia–nitrogen. Twice at each tempera-
ture, approximately 3.5 L of biosolids was transferred anaero-
bically from the continuous digester to a mixed batch reactor
to evaluate the rate of inactivation of the indigenous E. coli
and serotype O157:H7.
Both the continuous digester and the batch reactor were
suspended in a heated water bath whose temperature was
maintained at the desired level with a precision temperature
controller. The temperature controller and equipment used to
monitor temperature were calibrated against a thermometer
traceable to the US National Institute of Standards and
Technology (NIST); overall temperature accuracy was
70.1 1C (Aitken et al., 2005c).
Batch inactivation experiments in the digested manure
were conducted as described elsewhere (Aitken et al., 2005a).
The E. coli strains were prepared for inactivation experi-
ments by overnight cultivation in trypticase soy broth
(TSB) at 37 1C. Aliquots of each culture were mixed with
manure to a final volume of 105 mL. Concentrations of
each strain in the mixture were approximately 108 cfu/mL.
A 5-mL aliquot of the mixture was removed for analysis
and the remaining 100 mL was warmed to 35 1C while mixing
slowly on a hot-plate stirrer. Once the mixture reached 35 1C,
it was poured into the batch reactor containing the bio-
solids from the continuous digester. Mixing was allowed to
occur for 1.5 min before the first sample was collected;
measured concentrations of putative E. coli types in this
sample are considered to represent the initial (time zero)
concentrations.
Samples were collected anaerobically from the batch
reactor at desired intervals by pressurizing the reactor with
argon and displacing the desired volume of biosolids into
sterile sample bottles. Samples were cooled immediately (to
below 35 1C within 5 min and to 10 1C within 30 min) by
immersing the sample bottles into a chilled bath (�15 1C)
containing a 50:50 (v:v) mixture of isopropanol and water.
Samples were stored cold until analysis, which for most
samples was within 36 h.
Inactivation of each E. coli strain was also evaluated
in a laboratory medium at 5570.5 1C. An overnight culture
grown in TSB at 37 1C was split into 400mL aliquots in

ARTICLE IN PRESS
WAT E R R E S E A R C H 41 (2007) 1659– 1666 1661
1.5-mL microcentrifuge tubes, which were placed in a
heat block. One aliquot was immediately placed on ice
and the remainder were removed at pre-determined in-
tervals and placed on ice for at least 10 min. Cooled samples
were analyzed immediately by the plating method described
below.
2.2. Analytical methods
The residence time in the continuous-flow reactor was
determined by measuring the cumulative effluent volume
over time and dividing the mean volume in the digester
(16.4 L) by the mean flow rate (2.1 L/day; r2¼ 0.996, n ¼ 9). Gas
production was measured with a digital gas-flow meter and
the flow rate was recorded at 15-min intervals. Gas composi-
tion (CH4 and CO2) was analyzed by gas chromatography (GC)
with thermal conductivity detection and autoinjection of gas
routed directly from the headspace of the continuous digester
as described elsewhere (Aitken et al., 2005c). Grab samples
were collected for analysis of pH, TS and VS, VFAs, and
ammonia–N. Total and volatile solids were analyzed in
accordance with method 2540G in Standard Methods for the
Examination of Water and Wastewater (American Public
Health Association and American Water Works Association
and Water Environment Federation, 1999).
Samples for VFA and ammonia analyses were prepared by
centrifugation and filtration through glass–fiber filters. Sam-
ples for VFA analysis were diluted 1:1 (v:v) with 30 mM oxalic
acid. Samples for ammonia analysis were diluted 1:500 in
reagent water containing enough concentrated sulfuric acid
to reduce the pH to o2. Analysis of VFAs was by GC with
flame ionization detection (Aitken et al., 2005c). Total
ammonia-N (NH4+-N plus NH3-N) was analyzed by the Hach
Nessler Method (Hach Company, 1997) using Hach reagents
(Hach Co., Loveland, CO) and quantified against standards of
reagent-grade ammonium chloride.
A differential-selective plating method was used for routine
analysis and quantification of putative non-pathogenic E. coli
or putative serotype O157:H7 during inactivation experi-
ments. Samples were serially diluted and spread-plated on
MacConkey agar containing sorbitol instead of lactose
(sorbitol-MacConkey agar, SMAC), which is capable of distin-
guishing between E. coli O157:H7 and other E. coli (National
Center for Infectious Diseases, 1994; Armstrong et al., 1996).
On this medium, the wild-type E. coli strain grew as pink
(sorbitol-positive) colonies and serotype O157:H7, which
cannot ferment sorbitol rapidly, grew as clear (sorbitol-
negative) colonies. Serotype O157:H7 measured this way
requires confirmation with more specific analyses (National
Center for Infectious Diseases, 1994; Armstrong et al., 1996).
Accordingly, 10 sorbitol-negative colonies (putative E. coli
O157:H7) from dilutions of each of four samples (feed manure,
effluent from the continuous digester during operation at
55 1C, and samples collected after 4 and 24 h during a batch
inactivation experiment at 55 1C) were selected and stored for
confirmatory analyses. Each isolate was tested by the
Enterotubes II test kit for identification as E. coli and with
the Reveals immunoassay for type O157:H7 (Neogen Corpora-
tion, Lansing, MI). Isolates were also identified by sequencing
their 16S rRNA genes as described below.
2.3. Identification of isolates by 16S rRNA genesequencing
A whole-cell suspension of each isolate was used as template
for polymerase chain reaction (PCR) with general bacterial
primers 8F (Edwards et al., 1989) and 1492R (Lane, 1991) to
obtain nearly complete 16S rRNA genes. The PCR reaction
consisted of 1 mL of cell suspension, 1 mM of each primer, 1�
Eppendorf (Westbury, NY) MasterMixs and sterile water to a
final volume of 20mL. The PCR program included an initial
step of 94 1C for 10 min to lyse cells, followed by 30 cycles of
94 1C for 30 s, 55 1C for 30 s, and 72 1C for 90 s. Amplicons were
purified with a QIAquicks PCR Purification kit (Qiagen,
Valencia, CA) before partial sequencing with primer 8F was
performed by SeqWright (Houston, TX).
Sequences were analyzed and edited using Sequencher
(Gene Codes, Ann Arbor, MI). Sequences with X99% identity
over the region sequenced were grouped into operational
taxonomic units (OTUs), and only the longest sequence in an
OTU was considered in further analyses. The closest relatives
of sequences were determined by BLASTN (Altschul et al.,
1990) searches of GenBank. Sequences from this study and
close relatives were then aligned using the pileup program of
the Genetics Computer Group suite of programs (Wisconsin
Package version 10.3; Accelrys Inc., San Diego, CA) and
imported to CLUSTALW (Thompson et al., 1994) for construc-
tion of a phylogenetic tree. The final neighbor-joining tree
was based on approximately 450 bases (the length of the
shortest considered sequence), created without considering
gaps, and was bootstrapped 1000 times.
Sequences obtained in this study were deposited in
GenBank with accession numbers EF191167–EF191175.
2.4. Data analysis
In each of the four inactivation experiments in digested
manure (two at each temperature), both sorbitol-negative and
sorbitol-positive colonies were observed at sampling times
well beyond those that corresponded to simple first-order
inactivation kinetics. Data from these inactivation experi-
ments therefore were analyzed by assuming that there was a
heat-sensitive population and a heat-resistant population of
each colony type. It was initially assumed that each popula-
tion decayed exponentially but at different rates, such that
the total (heat-sensitive plus heat-resistant) concentration C
of a given colony type at any time t would correspond to:
CC0¼ fe�k1t þ ð1� f Þe�k2t, (1)
where C0 is the concentration at time zero, f is the fraction of
the total concentration that appears to be heat-sensitive, (1�f)
is the fraction of the total concentration that appears to be
heat resistant, and k1 and k2 are the first-order inactivation
rate coefficients for the heat-sensitive and heat resistant
fractions, respectively. Eq. (1) was solved for the three
parameters f, k1 and k2 using ProStat (Poly Software Interna-
tional, Pearl River, NY). If the regression returned for k2 either
a negative value or a very small positive value with a wide 95%
confidence interval, k2 was set to zero and the regression was
re-run to fit the other two parameters, f and k1. Rates of
ARTICLE IN PRESS
WAT E R R E S E A R C H 4 1 ( 2 0 0 7 ) 1 6 5 9 – 1 6 6 61662
inactivation of the indigenous E. coli isolate and the O157:H7
strain in TSB were analyzed by linear regression of ln
concentration vs. time.
The significance of differences between estimates of k1 for
any pair of conditions was determined by calculating the
Z-statistic from the best-fit values and standard errors.
Differences were considered to be different for Z values
corresponding to po0.05.
0 50 100 150 200 250 300
Time, minutes
-5.0
-4.0
-3.0
-2.0
-1.0
0.0Log
10 (
C/C
0)
sorbitol- (first test)
sorbitol- (second test)
sorbitol+ (first test)
sorbitol+ (second test)
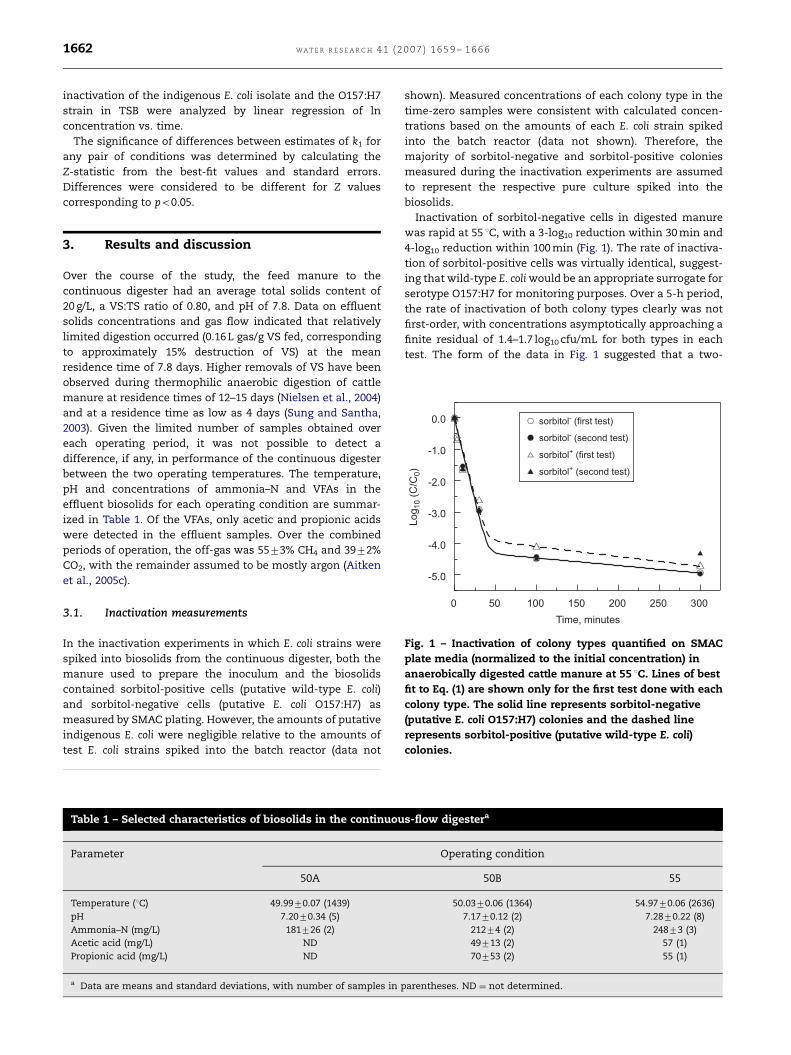
Fig. 1 – Inactivation of colony types quantified on SMAC
plate media (normalized to the initial concentration) in
anaerobically digested cattle manure at 55 1C. Lines of best
fit to Eq. (1) are shown only for the first test done with each
colony type. The solid line represents sorbitol-negative
(putative E. coli O157:H7) colonies and the dashed line
represents sorbitol-positive (putative wild-type E. coli)
colonies.
3. Results and discussion
Over the course of the study, the feed manure to the
continuous digester had an average total solids content of
20 g/L, a VS:TS ratio of 0.80, and pH of 7.8. Data on effluent
solids concentrations and gas flow indicated that relatively
limited digestion occurred (0.16 L gas/g VS fed, corresponding
to approximately 15% destruction of VS) at the mean
residence time of 7.8 days. Higher removals of VS have been
observed during thermophilic anaerobic digestion of cattle
manure at residence times of 12–15 days (Nielsen et al., 2004)
and at a residence time as low as 4 days (Sung and Santha,
2003). Given the limited number of samples obtained over
each operating period, it was not possible to detect a
difference, if any, in performance of the continuous digester
between the two operating temperatures. The temperature,
pH and concentrations of ammonia–N and VFAs in the
effluent biosolids for each operating condition are summar-
ized in Table 1. Of the VFAs, only acetic and propionic acids
were detected in the effluent samples. Over the combined
periods of operation, the off-gas was 5573% CH4 and 3972%
CO2, with the remainder assumed to be mostly argon (Aitken
et al., 2005c).
3.1. Inactivation measurements
In the inactivation experiments in which E. coli strains were
spiked into biosolids from the continuous digester, both the
manure used to prepare the inoculum and the biosolids
contained sorbitol-positive cells (putative wild-type E. coli)
and sorbitol-negative cells (putative E. coli O157:H7) as
measured by SMAC plating. However, the amounts of putative
indigenous E. coli were negligible relative to the amounts of
test E. coli strains spiked into the batch reactor (data not
Table 1 – Selected characteristics of biosolids in the continuou
Parameter
50A
Temperature (1C) 49.9970.07 (1439)
pH 7.2070.34 (5)
Ammonia–N (mg/L) 181726 (2)
Acetic acid (mg/L) ND
Propionic acid (mg/L) ND
a Data are means and standard deviations, with number of samples in
shown). Measured concentrations of each colony type in the
time-zero samples were consistent with calculated concen-
trations based on the amounts of each E. coli strain spiked
into the batch reactor (data not shown). Therefore, the
majority of sorbitol-negative and sorbitol-positive colonies
measured during the inactivation experiments are assumed
to represent the respective pure culture spiked into the
biosolids.
Inactivation of sorbitol-negative cells in digested manure
was rapid at 55 1C, with a 3-log10 reduction within 30 min and
4-log10 reduction within 100 min (Fig. 1). The rate of inactiva-
tion of sorbitol-positive cells was virtually identical, suggest-
ing that wild-type E. coli would be an appropriate surrogate for
serotype O157:H7 for monitoring purposes. Over a 5-h period,
the rate of inactivation of both colony types clearly was not
first-order, with concentrations asymptotically approaching a
finite residual of 1.4–1.7 log10 cfu/mL for both types in each
test. The form of the data in Fig. 1 suggested that a two-
s-flow digestera
Operating condition
50B 55
50.0370.06 (1364) 54.9770.06 (2636)
7.1770.12 (2) 7.2870.22 (8)
21274 (2) 24873 (3)
49713 (2) 57 (1)
70753 (2) 55 (1)
parentheses. ND ¼ not determined.
ARTICLE IN PRESS
WAT E R R E S E A R C H 41 (2007) 1659– 1666 1663
population model (Eq. (1)), assuming a heat-sensitive fraction
and a heat-resistant fraction of each colony type, would be
more appropriate than a model assuming simple first-order
inactivation of a single population. Results from non-linear
regression analyses are summarized in Tables 2 and 3 for
experiments at 55 and 50 1C, respectively.
In contrast to the observations in digested manure,
inactivation of both the wild-type E. coli strain and the E. coli
O157:H7 strain in TSB at 55 1C followed simple first-order
kinetics (data not shown). The inactivation rate coefficient (k1)
was 0.1470.09 min�1 (r2¼ 0.95, n ¼ 4) for the wild-type isolate
and 0.1170.05 min�1 (r2¼ 0.98, n ¼ 4) for the O157:H7 strain.
The corresponding decimal reduction times (D) were 17 and
21 min, respectively. The difference in inactivation rate
coefficients between the strains is not statistically significant,
but in both cases the rate coefficient for inactivation in
laboratory medium was significantly lower than in digested
manure. These results suggest that there might have been an
effect of the matrix on the inactivation rate in digested
manure, although the matrix component(s) responsible for
such an effect are not known. Of the major components in
digested manure or biosolids, protonated VFAs are known to
affect the inactivation of bacteria (Lee et al., 1989; Cherrington
et al., 1991; Fukushi et al., 2003; Salsali et al., 2006); however,
Table 2 – Summary of non-linear regression results for batch
Parameter Sorbitol-negative cells (putativO157:H7)
First test Secon
C0 (106 cfu/mL) 4.28 2.
k1 (min�1)a 0.2570.10 0.247
D (min)a 9.473.7 9.47
Heat-resistant fraction (cfu/cfu)a 0.671.9�10�4 0.873.5
r2 0.978 0.9
a Best-fit value and 95% confidence interval. There was no significant dif
for either test. D ¼ decimal reduction time; other parameters are defined
Table 3 – Summary of non-linear regression results for batch
Parameter Sorbitol-negative cells (putative E. coO157:H7)
First test Second test
C0 (106 cfu/mL) 3.02 6.76
k1 (min�1)a 0.03470.017 0.01470.004
D (min) 67734 171745
Heat-resistant fraction
(cfu/cfu) 0.771.1� 10�2 4.878.5� 10�
r2 0.956 0.961
a Notes as in Table 2. The difference between k1 values was significant b
difference between colony types for either test.
concentrations of protonated VFAs would have been negli-
gible at the pH of the digested manure.
The rate of inactivation of the heat-sensitive fraction of
both colony types in digested manure was an order of
magnitude higher at 55 1C than at 50 1C (Tables 2 and 3). The
corresponding decimal reduction times were approximately
10 min at 55 1C and ranged from 1 to 3 h at 50 1C. Such a large
difference between rates of inactivation over a 5 1C tempera-
ture interval indicates a steep relationship between inactiva-
tion rate and temperature within this range, as has been
observed for Ascaris suum and poliovirus in digested biosolids
from municipal wastewater treatment (Aitken et al., 2005a).
The decimal reduction times at 55 1C that we measured in
digested manure and in TSB are in the range of values
recently reported for E. coli O157:H7 in deionized water
(3.7–3.9 min; Spinks et al., 2006) and in dilute peptone media
(14–16 min; Sharma and Beuchat, 2004). The values of k2, the
inactivation rate coefficient for the apparent heat-resistant
fraction of each strain, were either 5k1 or near zero for each
strain at each temperature (not shown).
In the batch inactivation experiments at both 50 and 55 1C,
the fraction of apparent heat-resistant cells relative to the
initial population was small but significant (Tables 2 and 3).
Unspiked effluent from the continuous digester operated at
inactivation of E. coli at 55 1C
e E. coli Sorbitol-positive cells (putative non-pathogenicE. coli)
d test First test Second test
48 1.44 0.54
0.12 0.2370.13 0.2470.09
4.7 10.075.5 9.673.5
�10�4 1.778.2�10�4 4.176.6�10�5
85 0.959 0.973
ference between tests for either colony type or between colony types
in Materials and methods.
inactivation of E. coli at 50 1C
li Sorbitol-positive cells (putative non-pathogenic E.coli)
First test Second test
7.03 2.34
0.01870.019 0.01870.003
1277132 132721
4 3.777.1� 10�2 1.772.4�10�5
0.896 0.985
etween tests for sorbitol-negative cells but there was no significant

ARTICLE IN PRESS
WAT E R R E S E A R C H 4 1 ( 2 0 0 7 ) 1 6 5 9 – 1 6 6 61664
50 or 55 1C also contained bacteria that grew on SMAC agar at
concentrations ranging from 1.8 to 3.7 log10 cfu/mL (data not
shown). The existence of such heat-resistant cells suggests
that the digested biosolids contained low concentrations of
heat-resistant E. coli or other bacteria that grew on the SMAC
media used to assay E. coli. To evaluate this possibility,
sorbitol-negative isolates from SMAC plates were selected
Table 4 – Confirmatory analyses of sorbitol-negative cells (put
Samplea
Reveals
untreated manure 0
digester effluent 0
batch treatment for 4 h 0
batch treatment for 24 h 0
a Samples of digester effluent and from batch treatment of the effluen
sample, 10 colonies from SMAC plates used to quantify E. coli O157:H7 we
Enterotubes assay (confirmation as E. coli), and partial sequencing of
bacteria). For the 16S rRNA gene analysis, positives are considered to
sequences were not obtained for one isolate from the 4-h batch treatme
Untreated manur
Untreated manure i
Untreated man
24-hour batch treatm
4-hour batch treatme
4-hour batch treatmen
24-hour batch t
4-hour batch treatm
Pseudomonas sp. A
Pseudomonas sp. BW
Pseudomonas thermo
Escherichia col
Escherichia sp.
Escherichia col
Bacillus sp. 171544 (A
Bacillus licheniformis s
Clostridium bifermentaClostridium sp. IBUN 1
Swine manure bacteriuElbe River snow isolate
0.1
Escherichia coShigella sp. BB
Pseudomonas men
Pseudomonas alc
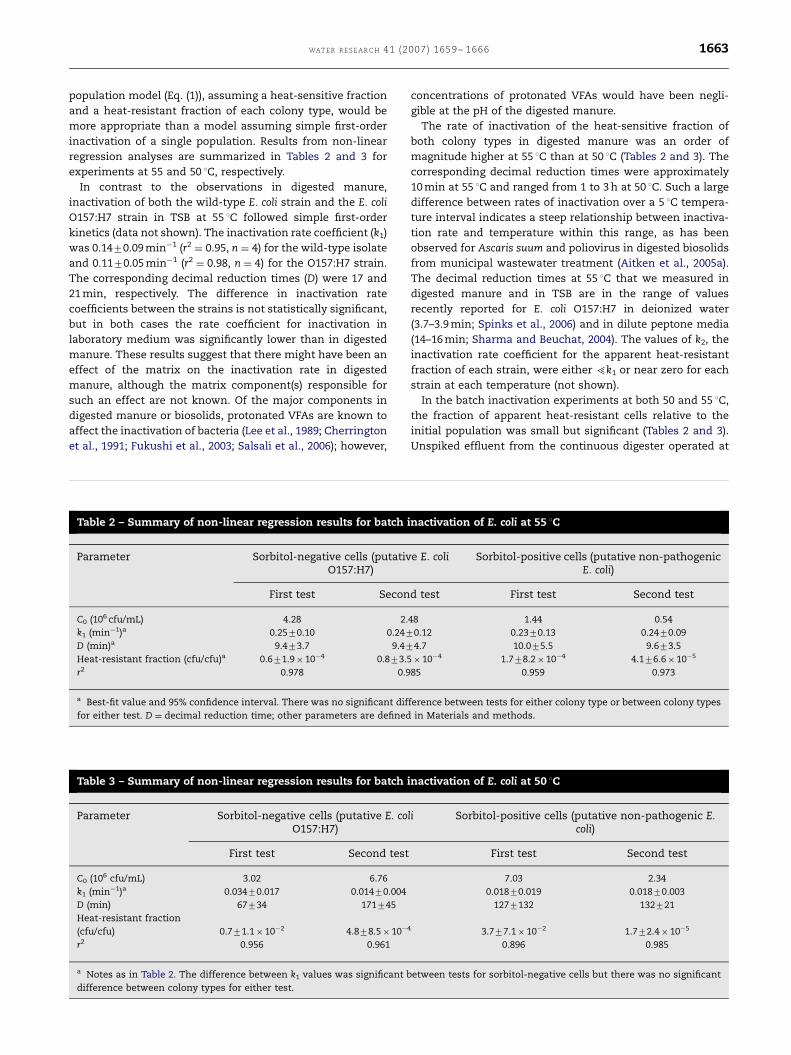
Fig. 2 – Phylogenetic tree based on partial 16S rRNA gene sequen
closest relatives. Sequences from this study are in bold and the n
isolates from each condition (untreated manure/digester effluen
OTU represented by that sequence. Accession numbers of refer
parentheses after the sequence name. Open (J) and closed (K)
support, respectively. The tree was rooted with Mycobacterium va
on tree). The sequence designated ‘‘E. coli isolated from dairy ca
digester in the thermal inactivation measurements.
for followup analyses using the Reveals immunoassay for
confirmation as type O157:H7, as well as the Enterotubes kit
and 16S rRNA gene sequencing for confirmation as E. coli.
None of the 30 heat-resistant, sorbitol-negative isolates
(combined isolates from samples representing treatment at
55 1C) were confirmed to be type O157:H7, and in fact none
were identified as E. coli (Table 4). In a similar analysis of 10
ative isolates of E. coli O157:H7)
Number of positive isolates
Enterotubes 16S rRNA Gene
6 8
0 0
0 0
0 0
t spiked with E. coli strains were from operation at 55 1C. For each
re analyzed by the Reveals assay (confirmation as type O157:H7), the
the 16S rRNA gene (sequence similarity to E. coli or related enteric
be those sequences that clustered with E. coli sequences (Fig. 2);
nt sample and one isolate from the 24-h batch treatment sample.
e isolate F-9 [1/0/0/0]
solate F-3 [1/0/0/0]
ure isolate F-8 [8/0/0/0]
ent isolate B24-10 [0/0/0/1]
nt isolate B4-7 [0/0/1/0]
t isolate B4-10 [0/0/1/0]
reatment isolate B24-9 [0/2/1/3]
ent isolate B4-1 [0/7/6/6]
STI (AF118398)
DY-26 (DQ219371)
tolerans strain CM3 (AJ311980)
i O157:H7 strain WAB1892 (AM184233)
BBDP20 (DQ337503)
i (T); ATCC 11775T (X80725)
F071856)
train CICC10085 (AY842869)
ns strain IBUN 188 (DQ680025)84 (DQ680021)
m RT-1A (AY167932) Iso11_3 (AF150692)
li isolated from dairy cattle manureDP15 (DQ337523)
docina strain 174 (AY870674)
aligenes strain HPC 1032 (AY948235)
ces showing the recovered isolates from this study and their
umbers in brackets after each sequence indicate how many
t/batch treatment for 4 h/batch treatment for 24 h) were in the
ence sequences from the GenBank database are in
circles on the nodes represent X95% and X50% bootstrap
nbaalenii (GenBank accession number AY438079; not shown
ttle manure’’ is the wild-type isolate used to spike the

ARTICLE IN PRESS
WAT E R R E S E A R C H 41 (2007) 1659– 1666 1665
sorbitol-negative isolates from the untreated manure, none
were confirmed to be type O157:H7; eight had partial 16S
rRNA gene sequences highly similar to E. coli and other
enteric bacteria (Fig. 2), six of which were identified as
sorbitol-negative E. coli by the Enterotubes assay (Table 4).
Most of the 16S rRNA gene sequences associated with
the heat-resistant isolates clustered with Pseudomonas spp.
(Fig. 2), with the majority highly similar to the recently
discovered Pseudomonas thermotolerans. This species was
originally isolated from the cooking water of a cork-proces-
sing plant and is thought to be the first thermotolerant
Pseudomonas spp. characterized (Manaia and Moore, 2002).
Although we did not quantify this organism in either the feed
manure or the digester, it clearly must have been present in
the untreated manure and is capable of growing on SMAC
plate media. The two isolates from the feed manure whose
partial 16S rRNA gene sequences clustered with Pseudomonas
spp. (Fig. 2) were also identified as species other than E. coli in
the Enterotubes assay.
The findings from this study underscore the importance of
confirming the identity of bacteria tentatively identified as E.
coli by plating methods using differential-selective agar,
particularly in samples representing biosolids from thermal
treatment of animal waste or wastewater sludge. The
measurement of false positives could lead to erroneous
conclusions about the performance of thermal treatment
processes with respect to inactivation of pathogenic strains of
E. coli or the inactivation of other pathogens (such as
Salmonella or Campylobacter) if total E. coli were used as an
indicator for them.
4. Conclusions
E. coli O157:H7, an important human pathogen found in the
manure of cattle and other farm animals, can be rapidly and
extensively inactivated by thermophilic anaerobic digestion.
Its inactivation rate can be predicted by the inactivation of
non-pathogenic E. coli, which are expected to be present at
substantially higher concentrations than type O157:H7 in
cattle manure. However, when plating methods with differ-
ential-selective agars are used to quantify E. coli concentra-
tions, the observable extent of E. coli inactivation might be
limited by the existence of low concentrations of non-target
organisms that can grow on the media. Confirmatory assays
should be conducted when using such media, because false
positives can lead to misinterpretation of inactivation
kinetics and can result in incorrect decisions regarding the
performance of thermal treatment processes.
Acknowledgments
We thank Lee-Ann Jaykus for providing the strain of E. coli
O157:H7, Leonard Bull for facilitating access to the experi-
mental dairy farm at N.C. State University, Wayne McLamb for
assistance with sample collection, and Randall Goodman for
assistance with experimental equipment. A preliminary
description of this work appeared in the proceedings for the
10th IWA World Congress on Anaerobic Digestion (Montreal,
Canada, 2004). This work was supported in part by a Grant
from the US National Science Foundation (Grant BES 0221836).
R E F E R E N C E S
Aitken, M.D., Sobsey, M.D., Blauth, K.E., Shehee, M., Crunk, P.L.,Walters, G.W., 2005a. Inactivation of Ascaris suum and polio-virus in biosolids under thermophilic anaerobic digestionconditions. Environ. Sci. Technol. 39, 5804–5809.
Aitken, M.D., Sobsey, M.D., Shehee, M., Blauth, K.E., Hill, V.R., Farrell,J.B., Nappier, S.P., Walters, G.W., Crunk, P.L., Van Abel, N., 2005b.Laboratory evaluation of thermophilic anaerobic digestion toproduce Class A biosolids. 2. Inactivation of pathogens andindicator organisms in a continuous-flow reactor followed bybatch treatment. Water Environ. Res. 77, 3028–3036.
Aitken, M.D., Walters, G.W., Crunk, P.L., Willis, J.L., Farrell, J.B.,Schafer, P.L., Arnett, C., Turner, B., 2005c. Laboratory evalua-tion of thermophilic anaerobic digestion to produce Class Abiosolids. 1. Stabilization performance of a continuous-flowreactor at low residence time. Water Environ. Res. 77,3019–3027.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990.Basic local alignment search tool. J. Mol. Biol. 215, 403–410.
American Public Health Association, American Water WorksAssociation and Water Environment Federation, 1999. Stan-dard Methods for the Examination of Water and Wastewater,20th ed. American Public Health Association, American WaterWorks Association, and Water Environment Federation, Wa-shington, DC.
Armstrong, G.L., Hollingsworth, J., Morris Jr., J.G., 1996. Emergingfoodborne pathogens: E. coli O157:H7 as a model of entry of anew pathogen into the food supply of the developed world.Epidemiol. Rev. 18, 29–51.
Berry, E.D., Miller, D.N., 2005. Cattle feedlot soil moisture andmanure content: II. Impact on E. coli O157. J. Environ. Qual. 34,656–663.
Cherrington, C.A., Hinton, M., Pearson, G.R., Chopra, I., 1991.Short-chain organic acids at pH 5.0 kill E. coli and Salmonellaspp. without causing membrane perturbation. J. Appl. Bacter-iol. 70, 161–165.
Davis, M.A., Cloud-Hansen, K.A., Carpenter, J., Hovde, C.J., 2005.E. coli O157:H7 in environments of culture-positive cattle. Appl.Environ. Microbiol. 71, 6816–6822.
Djordjevic, S.P., Ramachandran, V., Bettelheim, K.A., Vanselow,B.A., Holst, P., Bailey, G., Hornitzky, M.A., 2004. Serotypes andvirulence gene profiles of shiga toxin-producing E. coli strainsisolated from feces of pasture-fed and lot-fed sheep. Appl.Environ. Microbiol. 70, 3910–3917.
Duffy, G., 2003. Verocytoxigenic E. coli in animal faeces, manuresand slurries. J. Appl. Microbiol. 94, 94S–103S.
Edwards, U., Rogall, T., Blocker, H., Emde, M., Bottger, E.C., 1989.Isolation and direct complete nucleotide determination ofentire genes. Characterization of a gene coding for 16Sribosomal RNA. Nucleic Acids Res. 17, 7843–7853.
Fegan, N., Higgs, G., Vanderlinde, P., Desmarchelier, P., 2003.Enumeration of E. coli O157 in cattle faeces using mostprobable number technique and automated immunomagneticseparation. Lett. Appl. Microbiol. 38, 56–59.
Fukushi, K., Babel, S., Burakrai, S., 2003. Survival of Salmonella spp.in a simulated acid-phase anaerobic digester treating sewagesludge. Bioresour. Technol. 86, 53–57.
Hach Company, 1997. Water Analysis Handbook. Hach Company,Loveland, CO.
Lane, D.J., 1991. 16S/23S rRNA sequencing. In: Stackebrandt, E.,Goodfellow, M. (Eds.), Nucleic acid techniques in bacterialsystematics. Wiley, New York, NY, pp. 115–175.

ARTICLE IN PRESS
WAT E R R E S E A R C H 4 1 ( 2 0 0 7 ) 1 6 5 9 – 1 6 6 61666
Lee, K.M., Brunner, C.A., Farrell, J.B., Eralp, A.E., 1989. Destructionof enteric bacteria and viruses during two-phase digestion. J.Water Pollut. Contr. Fed. 61, 1421–1429.
Manaia, C.M., Moore, E.R.B., 2002. Pseudomonas thermotolerans sp.nov., a thermotolerant species of the genus Pseudomonassensu stricto. Int. J. System. Evol. Microbiol. 52, 2203–2209.
Mead, P.S., Griffin, P.M., 1998. E. coli O157:H7. The Lancet 352,1207–1212.
Muniesa, M., Jofre, J., Garcia-Aljaro, C., Blanch, A.R., 2006.Occurrence of E. coli O157:H7 and other enterohemorrhagicE. coli in the environment. Environ. Sci. Technol. 40, 7141–7149.
National Center for Infectious Diseases, 1994. E. coli O157:H7:procedure for isolation and identification from stool speci-mens. Centers for Disease Control and Prevention, Atlanta,GA.
Nicholson, F.A., Groves, S.J., Chambers, B.J., 2005. Pathogensurvival during livestock manure storage and following landapplication. Bioresour. Technol. 96, 135–143.
Nielsen, H.B., Mladenovska, Z., Westermann, P., Ahring, B.K., 2004.Comparison of two-stage thermophilic (68 1C/55 1C) anaerobicdigestion with one-stage thermophilic (55 1C) digestion ofcattle manure. Biotechnol. Bioeng. 86, 291–300.
Omisakin, F., MacRae, M., Ogden, I.D., Strachan, N.J.C., 2003.Concentration and prevalence of E. coli O157 in cattle feces atslaughter. Appl. Environ. Microbiol. 69, 2444–2447.
Salsali, H.R., Parker, W.J., Sattar, S.A., 2006. Impact of concentra-tion, temperature, and pH on inactivation of Salmonella spp. by
volatile fatty acids in anaerobic digestion. Can. J. Microbiol. 52,279–286.
Sharma, M., Beuchat, L.R., 2004. Sensitivity of E. coli O157:H7 tocommercially available alkaline cleaners and subsequentresistance to heat and sanitizers. Appl. Environ. Microbiol. 70,1795–1803.
Solomon, E.B., Yaorn, S., Matthews, K.R., 2002. Transmission of E.coli O157:H7 from contaminated manure and irrigation waterto lettuce plant tissue and its subsequent internalization.Appl. Environ. Microbiol. 68, 397–400.
Spinks, A.T., Dunstan, R.H., Harrison, T., Coombes, P., Kuczera, G.,2006. Thermal inactivation of water-borne pathogenic andindicator bacteria at sub-boiling temperatures. Water Res. 40,1326–1332.
Sung, S., Santha, H., 2003. Performance of temperature-phasedanaerobic digestion (TPAD) system treating dairy cattlewastes. Water Res. 37, 1628–1636.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Res. 22,4673–4680.
Todd, W.T.A., Dundas, S., 2001. The management of VTEC O157infection. Int. J. Food. Microbiol. 66, 103–110.
Zhao, T., Doyle, M.P., Shere, J., Garber, L., 1995. Prevalence ofenterohemorrhagic E. coli O157:H7 in a survey of dairy herds.Appl. Environ. Microbiol. 61, 1290–1293.