in vitro fermentation of beet fibre and barley bran, of their insoluble residues after digestion and...
TRANSCRIPT

J Sci Food Agric 1997, 75, 315È325
In Vitro Fermentation of Beet Fibre and BarleyBran, of their Insoluble Residues after Digestionand of Ileal EffluentsAnthony Fardet, Fabienne Guillon,* Christine Hoebler and Jean-Luc Barry
Human Nutrition Research Centre, Station of Technology Applied to Nutrition, INRA, BP 71627, 44316Nantes Ce� dex 03, France
(Received 8 July 1996 ; revised version received 2 September 1996 ; accepted 9 April 1997)
Abstract : The main objective of this study was to determine the form in whichbeet and barley bran Ðbres reach the colon, and to evaluate the inÑuence ofendogeneous compounds on their patterns of fermentation. Raw Ðbres (RF), cor-responding ileal effluents (IE) from pigs, and insoluble Ðbre residues (IR)extracted from IE, were fermented with human faecal inoculum for 24 h in an invitro batch system. For beet Ðbre, rate but not extent of cell wall sugars degrada-tion was increased (]34% at 6 h, P\ 0É05) after oroileal transit, due to a moreporous structure. For barley bran, oroileal conditions degraded endosperm com-pounds such as b-glucans, leading to a lower extent of cell wall glucose fermenta-tion compared with RF ([22% at 24 h, P\ 0É05). In the presence ofendogeneous substances, degradation of beet Ðbre polysaccharides was delayed(P\ 0É05) at each incubation time but that of barley bran Ðbre was unaltered.Compared to RF, IR and IE signiÐcantly exhibited lower acetate production forbeet Ðbre, and higher propionate and lower butyrate production for barley branafter 24 h. It is concluded that in vivo digestion modiÐed fermentation patterns ofboth Ðbres in a manner depending on botanical structure.
J Sci Food Agric 75, 315È325 (1997)No. of Figures : 2. No. of Tables : 4. No. of References : 45
Key words : dietary Ðbre, beet Ðbre, barley bran, ileal effluents, insoluble Ðbreresidues, in vitro fermentation, human faecal bacteria
INTRODUCTION
Colonic fermentation in humans has important implica-tions for health. Short-chain fatty acids (SCFAs) are themain end-products of dietary Ðbre fermentation, andthey are extensively absorbed by the colonic mucosabefore undergoing a variety of metabolic fates withinthe body (Cummings and McFarlene 1991), with beneÐ-cial physiological e†ects. Non-fermented substrates andthe bacterial biomass, through their ability to holdwater, may exert a mechanical physical action whichhelps regulate transit and faecal output (Cherbut et al1991).
The quantity and nature of substrates available forcolonic fermentation depends on the amount and type
* To whom correspondence should be addressed.
of diet eaten and on the ability of the small intestine todigest food components. Dietary Ðbre unabsorbed inthe small intestine may provide a large proportion ofthe available substrates for bacteria (Nordgaard andMortensen 1995). Not all dietary Ðbres are fermented tothe same extent or lead to the same SCFA proÐle.
The identity of factors inÑuencing fermentability ofÐbre substrates has been the subject of much research.They include monosaccharide composition (Salvador etal 1993), hydration properties (McBurney et al 1985),particle size and porosity (Au†ret et al 1993). Fibre withhigh initial water-retention capacity is fermented moreby faecal bacteria than Ðbre with low water-retentioncapacity (Stephen and Cummings 1979 ; McBurney et al1985). Particle size and porosity determine the surfacearea in contact with bacteria (Au†ret et al 1993). Poro-sity at the level of bacteria is related essentially to the
3151997 SCI. J Sci Food Agric 0022-5142/97/$17.50. Printed in Great Britain(

316 A Fardet et al
anatomical structure of dietary Ðbre. The three-dimensional arrangement of polysaccharides within thecell wall, and interactions with other substances e†ectdietary Ðbre fermentability (Bourquin et al 1992 ;Guillon et al 1992).
Most published results have been obtained from invitro batch systems with raw and pure products, andprovide data that are relevant but not, however, suffi-cient to extrapolate to the in vivo situation. The form inwhich Ðbre reaches the large intestine must be docu-mented. It has been shown that under the conditionsprevailing throughout the small intestine of pigs(Millard and Chesson 1984 ; Graham et al 1986 ;Robertson et al 1992 ; Bach Knudsen et al 1993) andhumans, especially ileostomates (Holloway et al 1978 ;
et al 1994), dietary Ðbre may undergo physi-A� mancochemical modiÐcations. It is partially degraded, sothat signiÐcant changes in particle size (Robertson et al1992), porosity and water hydration properties mayoccur. Thus, in vitro fermentation proÐles from Ðbre aseaten cannot be simply extrapolated to the in vivo situ-ation, and could be misleading.
Moreover, non-dietary Ðbre compounds such as pro-teins, pancreatic and gut secretions, mucus andsloughed epithelial cells, all of which being available forfermentation, are probably signiÐcant substrates forbacteria. Non-cell-wall compounds could account for50% of dry matter entering the colon (Cummings andMacFarlane 1991). The fate of these compounds in thecolon and their interactions with dietary Ðbre fermenta-tion has been poorly documented (McBurney et al1988 ; Nordgaard et al 1995 ; Silvester et al 1995). Thepurpose of the present study was to investigate the invitro fermentation of two dietary Ðbres, in the form inwhich they reach the colon, and under physiologicalconditions. Firstly, the fermentation patterns of rawÐbre (RF) and of their insoluble residues (IR) after gas-trointestinal digestion were compared ; secondly, fer-mentations of IR and of corresponding ileal effluents(IE) were compared to study inÑuence of non-cell-wallcompounds on fermentation parameters.
EXPERIMENTAL
Raw material (RF)
Sugar-beet Ðbre (by-product of sucrose extraction,average particle size 510 km) was obtained fromAgroindustrie-Recherche-De� veloppement (Arthenay,France). Barley bran (average particle size 520 km) wasfrom Whestove (Arques, France).
Collection of ileal effluents (IE)
Three Large White female pigs (45È50 kg) housed indi-vidually in pens with an ambient temperature of 20¡C
were Ðtted with a permanent cannula in the terminalileum and successively given a basal diet and two Ðbre-enriched diets. Four days before IE collection, theyreceived a basal diet twice daily consisting of 360 g ofpregelatinised wheat starch, 88É5 g of casein, 25 g ofmaize oil and 26É5 g of a vitamin/mineral mixture(2019 kcal). The feed was thoroughly mixed with waterbeforehand. The day before IE collection, 6% of pregel-atinised starch was substituted for Ðbre. On the day ofIE collection, pigs were given a diet with 12% Ðbre.Four hours after the meal, IE were collected for a totalof 10 h. This collection period is considered representa-tive of 24 h collection (Livingstone et al 1980). Betweeneach diet period, pigs were given a Ðbre-free diet for 2days to remove residual Ðbre. An equal quantity of IEcollected from the three pigs was then pooled for eachdiet and divided into two fractions, one immediatelyfreeze-dried for further chemical analysis and in vitrofermentations and the other used to isolate insolubleÐbre residues. Samples were as chemical and physi-cochemical characteristics of IE and IR for each pigwere in the same range. Pooled IE and IR were con-sidered as the mean, representative of collected residuesfrom the three pigs.
Preparation of insoluble Ðbre residues (IR) from IE
A subsample of IE was centrifuged at 4500 ] g for10 min. The supernatant was then treated with fourvolumes of absolute ethanol, and the mixture was cen-trifuged. The supernatant was discarded and the pre-cipitate freeze-dried. From the weight of precipitate,soluble polymer was calculated to account for less than5% of the IE of the two Ðbre diets and it was less thanone-third cell wall sugars. The soluble Ðbre fraction wastherefore not subsequently taken into account in thisexperiment.
The pellet was dispersed in distilled water (100 ml)and transferred onto an 80-km sieve. Water spray wasthen injected at 200 ml min~1 for 1 h. The retainedmaterial was divided into two fractions, the Ðrst beingstored at 4¡C in wet form for measurement of water-retention capacity and particle size and the secondfreeze-dried for further chemical analysis and in vitrofermentation. An 80 km pore size sieve was used since apreliminary experiment on raw material had shown thatless than 5% of the total mass for sugar-beet Ðbre andbarley bran was retained. In the case of barley bran, thematerial was composed mainly of starch granules.
In vitro fermentation by human faecal inoculum
In vitro fermentations were conducted in a batch systemby the procedure developed by Guillon et al (1995) andslightly modiÐed. Fresh human faeces were anaero-bically collected from three to four healthy volunteers

Fermentation of dietary Ðbre entering the colon 317
to reduce variability between the fermentation trials(Nyman et al 1986 ; McBurney and Thompson 1987).They had been eating an unspeciÐed Western diet andhad received no antibiotic treatment for 3 months. Noadaptation of donors was considered. Since the Ðbrematerials used in this study are natural components of aEuropean diet, the microÑora was thought to beadapted and to possess the hydrolytic activities neces-sary to the breakdown of the constitutive polysac-charides of the Ðbres. The faeces were immediatelymixed (1 : 3, w/v) with a nutritive bu†erCO2-saturated(38¡C) (Durand and Komisarczuk 1988), then Ðlteredthrough six layers of surgical gauze to remove Ðbre par-ticles. A 49 ml sample of the resulting inoculum wasintroduced into 120 ml serum bottles containing 1É4 gsample of experimental substrate presoaked over-nightat 4¡C with 21 ml of nutritive bu†er and then dispersedto be readily available for fermentation. The dilution ofthe inoculum and the amount of substrate incubatedwere chosen so that no lag phase, due to low initialbacterial population, was observed. Before use, andduring preparation of the inoculum, continuous bub-bling of maintained anaerobiosis and ensured con-CO2stant pH. Each bottle was sealed, then placedhorizontally in a shaking water bath (70 strokes min~1)at 38¡C, and kept for 5 min under a constant Ñow of N2to eliminate As some fermentable material in theO2 .inoculum itself can be present, and contribute toincrease SCFA production during a 24 h incubation, acontrol containing no substrate were also established.Before incubation, pH of the inoculum was 7É0 ^ 0É1and remained in the range 7É0È5É4 on the course ofincubation, which are similar to physiological valuesseen in the large intestine during fermentation. All theseconditions are considered to be compatible with the invivo events as already discussed by Barry et al (1995)and Edwards et al (1996).
Fermentations were stopped after 6, 12 and 24 h byaddition of 0É2 ml of a solution of mercuric chloride(10%, w/v). The medium was centrifuged (5000] g,10 min) ; two 2É5 ml aliquots of supernatant were eachtransferred to a tube containing 250 kl of a solution ofphosphoric acid (5%, w/v) and mercuric chloride (1%,w/v) and then frozen for determination of SCFA con-centration. After fermentation, the Ðbre residues wereseparated from bacteria by the procedure of Guillon etal (1995). Dried unfermented residues were groundbefore neutral and acidic cell wall sugar determinations.Two fermentations (on two di†erent days, ie with di†er-ent inocula) with duplicate for each time were carriedout for RF, IR and IE of beet Ðbre and barley bran.
Analytical
All values were calculated on a moisture-free basis. Drymatter was determined by oven drying at 103¡C to con-
stant weight. Proteins (N ] 6É25) were determined bythe Kjeldahl procedure, and starch by the method ofKarkalas (1985). Ash was determined by incinerationovernight at 550¡C then 1 h at 900¡C.
Neutral sugar composition was determined accordingto Hoebler et al (1989). After two-step acidic hydrolysis(72% at 25¡C for 30 min followed by 4 MH2SO4
at 100¡C for 2 h), neutral sugars were derivedH2SO4into alditol acetates and analysed by gasÈliquid chro-matography. Acidic sugars were quantiÐed as galact-uronic acid by the meta-hydroxydiphenyl method asautomated by Thibault (1979), using galacturonic acidas a standard. The b-glucan contents of brans weredetermined by the enzymatic method of McCleary andCodd (1991). SCFA (acetic acid, C2 ; propionic acid, C3 ;butyric acid, C4 ; isobutyric acid, isoC4 ; valeric acid,C5 ; and isovaleric acid, isoC5) were analysed by gasÈliquid chromatography (mM litre~1) (Jouany 1982).Individual SCFA productions were calculated by sub-tracting the corresponding SCFA concentration at time0 h and are expressed as mmol per gram of dry organicmatter. The rate of apparent disappearance (RAD) ofeach Ðbre component was calculated as follows :
RAD\
(Amount in initial Ðbre[ Amount in unfermented residue
Amount in initial Ðbre] 100
Physicochemical properties
Suction pressure (Robertson and Eastwood 1981) wasused to measure the water-retention capacity of Ðbrefollowing the procedure of Guillon et al (1995). Water-retention capacity was expressed in g of water per g ofdry residue.
Macroporosity was investigated by measuring thewater held within the Ðbrous matrix upon applicationof increasing suction pressure (Robertson 1988). Porediameter (D) is related to suction pressure (*P) andsurface tension (S) by the formula D\ 4S/*P asdescribed by Robertson (1988). *P was generated bydi†erent solutions of polyethylene glycol (PEG : 12É5È180 g of PEG litre~1 of water) and S was equal to50 ] 10~3 N ] m~1. The maximum water-retentioncapacity for a Ðbre source (100% saturation in purewater) was determined by extrapolating of water-retention capacity of zero suction pressure. The totalvolume of macropores, approximated the value ofwater-retention capacity when suction pressure waslowest. Results are expressed in ml of water per gram ofdry residue.
Particle size distribution was measured by wet sieving(Melcion and Delort-Laval 1981) through a series ofeight (barley bran) and nine (sugar-beet Ðbre) sieves ofdecreasing mesh size (1250 to 50 km and 1600 to 50 km,respectively). A sample (2É5 g) was sprinkled on the topof sieves and sieved using a shaker (Retsch, vibration
318 A Fardet et al
amplitude of 3 mm, frequency of 60 Hz) for 7 min. Thelogarithm of the percent cumulative sample weightpassing through each sieve was plotted against the sieveaperture. Particle size distribution was deÐned by geo-metric mean particle size and geometric standard devi-ation (AFNOR 1987).
Statistical analysis
All data from in vitro fermentation measurements areexpressed as the means ^ standard error of the mean(SEM) of the two fermentations conducted in duplicate.All statistics were performed using Stat-View II soft-ware (Abacus Concept Inc, CA, USA) on an AppleMacintosh IIci microcomputer. ANOVA followed bythe least signiÐcant di†erence (LSD) test were used totest the signiÐcance of treatment e†ects for fermentationvariables.
RESULTS
Chemical and physicochemical characteristics ofexperimental substrates (Table 1)
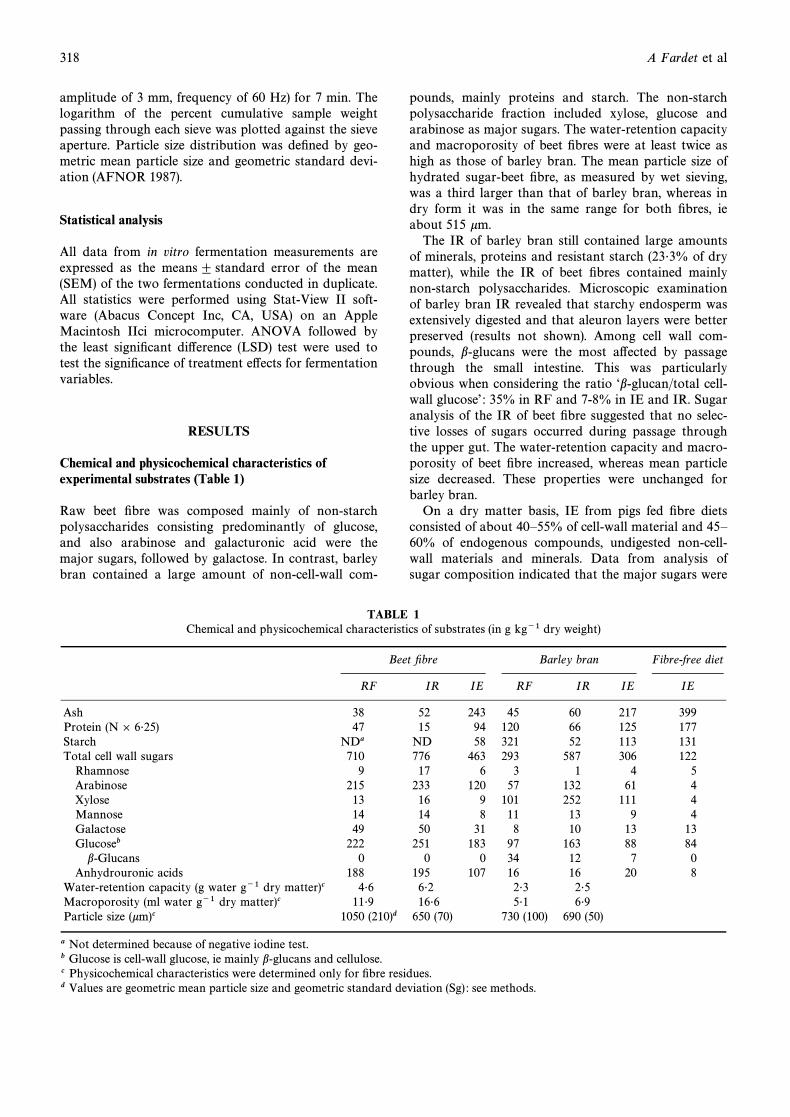
Raw beet Ðbre was composed mainly of non-starchpolysaccharides consisting predominantly of glucose,and also arabinose and galacturonic acid were themajor sugars, followed by galactose. In contrast, barleybran contained a large amount of non-cell-wall com-
pounds, mainly proteins and starch. The non-starchpolysaccharide fraction included xylose, glucose andarabinose as major sugars. The water-retention capacityand macroporosity of beet Ðbres were at least twice ashigh as those of barley bran. The mean particle size ofhydrated sugar-beet Ðbre, as measured by wet sieving,was a third larger than that of barley bran, whereas indry form it was in the same range for both Ðbres, ieabout 515 km.
The IR of barley bran still contained large amountsof minerals, proteins and resistant starch (23É3% of drymatter), while the IR of beet Ðbres contained mainlynon-starch polysaccharides. Microscopic examinationof barley bran IR revealed that starchy endosperm wasextensively digested and that aleuron layers were betterpreserved (results not shown). Among cell wall com-pounds, b-glucans were the most a†ected by passagethrough the small intestine. This was particularlyobvious when considering the ratio “b-glucan/total cell-wall glucoseÏ : 35% in RF and 7-8% in IE and IR. Sugaranalysis of the IR of beet Ðbre suggested that no selec-tive losses of sugars occurred during passage throughthe upper gut. The water-retention capacity and macro-porosity of beet Ðbre increased, whereas mean particlesize decreased. These properties were unchanged forbarley bran.
On a dry matter basis, IE from pigs fed Ðbre dietsconsisted of about 40È55% of cell-wall material and 45È60% of endogenous compounds, undigested non-cell-wall materials and minerals. Data from analysis ofsugar composition indicated that the major sugars were
TABLE 1Chemical and physicochemical characteristics of substrates (in g kg~1 dry weight)
Beet Ðbre Barley bran Fibre-free diet
RF IR IE RF IR IE IE
Ash 38 52 243 45 60 217 399Protein (N ] 6É25) 47 15 94 120 66 125 177Starch NDa ND 58 321 52 113 131Total cell wall sugars 710 776 463 293 587 306 122
Rhamnose 9 17 6 3 1 4 5Arabinose 215 233 120 57 132 61 4Xylose 13 16 9 101 252 111 4Mannose 14 14 8 11 13 9 4Galactose 49 50 31 8 10 13 13Glucoseb 222 251 183 97 163 88 84
b-Glucans 0 0 0 34 12 7 0Anhydrouronic acids 188 195 107 16 16 20 8
Water-retention capacity (g water g~1 dry matter)c 4É6 6É2 2É3 2É5Macroporosity (ml water g~1 dry matter)c 11É9 16É6 5É1 6É9Particle size (km)c 1050 (210)d 650 (70) 730 (100) 690 (50)
a Not determined because of negative iodine test.b Glucose is cell-wall glucose, ie mainly b-glucans and cellulose.c Physicochemical characteristics were determined only for Ðbre residues.d Values are geometric mean particle size and geometric standard deviation (Sg) : see methods.
Fermentation of dietary Ðbre entering the colon 319
those present in the ingested Ðbre. IE collected after thebeet-Ðbre-enriched diet contained more cell-wallmaterial (54% of dry matter) than IE collected after thebarley bran-enriched diet (40% of dry matter). IE recov-ered after Ðbre-free diet contained minerals (48% of drymatter), proteins (21% of dry matter) cell-wall sugars(30É5% of dry matter) and resistant starch (13É1% of drymatter). In addition to glucose, galactose and uronicacid were present in relatively high quantities.
Fermentation
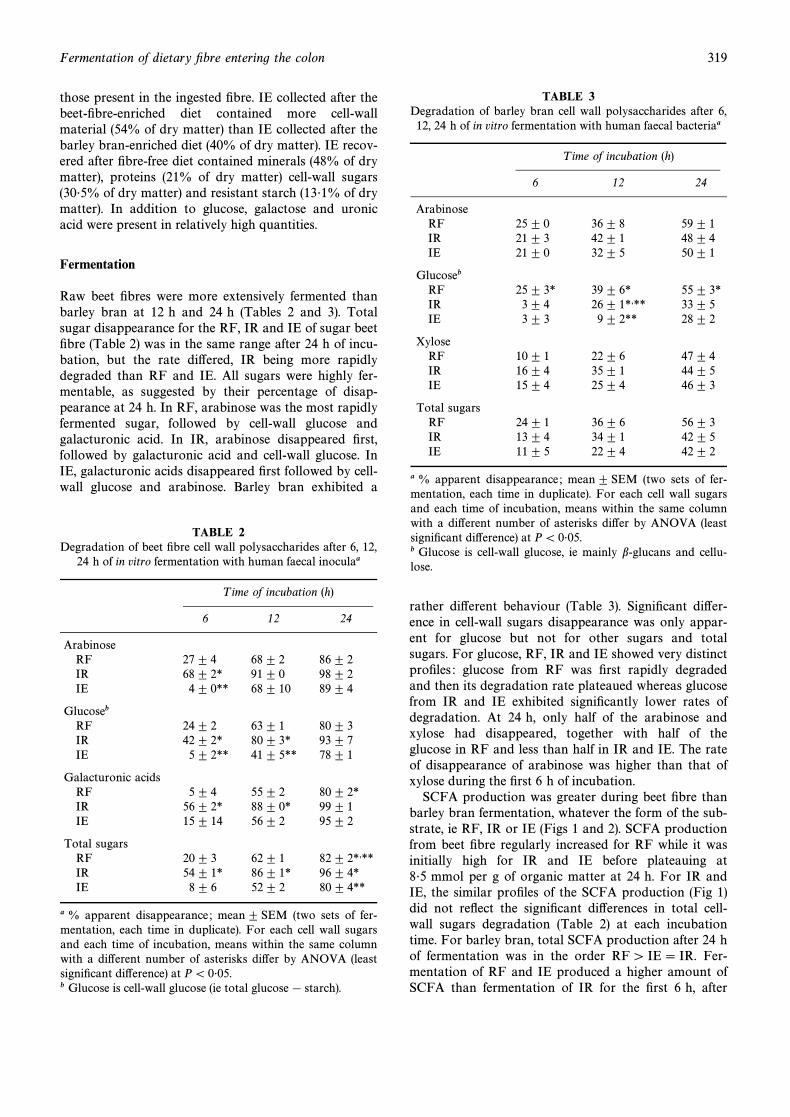
Raw beet Ðbres were more extensively fermented thanbarley bran at 12 h and 24 h (Tables 2 and 3). Totalsugar disappearance for the RF, IR and IE of sugar beetÐbre (Table 2) was in the same range after 24 h of incu-bation, but the rate di†ered, IR being more rapidlydegraded than RF and IE. All sugars were highly fer-mentable, as suggested by their percentage of disap-pearance at 24 h. In RF, arabinose was the most rapidlyfermented sugar, followed by cell-wall glucose andgalacturonic acid. In IR, arabinose disappeared Ðrst,followed by galacturonic acid and cell-wall glucose. InIE, galacturonic acids disappeared Ðrst followed by cell-wall glucose and arabinose. Barley bran exhibited a
TABLE 2Degradation of beet Ðbre cell wall polysaccharides after 6, 12,
24 h of in vitro fermentation with human faecal inoculaa
T ime of incubation (h)
6 12 24
ArabinoseRF 27 ^ 4 68 ^ 2 86 ^ 2IR 68 ^ 2* 91 ^ 0 98 ^ 2IE 4 ^ 0** 68 ^ 10 89 ^ 4
GlucosebRF 24 ^ 2 63 ^ 1 80 ^ 3IR 42 ^ 2* 80 ^ 3* 93 ^ 7IE 5 ^ 2** 41 ^ 5** 78 ^ 1
Galacturonic acidsRF 5 ^ 4 55 ^ 2 80 ^ 2*IR 56 ^ 2* 88 ^ 0* 99 ^ 1IE 15 ^ 14 56 ^ 2 95 ^ 2
Total sugarsRF 20 ^ 3 62 ^ 1 82 ^ 2*,**IR 54 ^ 1* 86 ^ 1* 96 ^ 4*IE 8 ^ 6 52 ^ 2 80 ^ 4**
a % apparent disappearance ; mean ^ SEM (two sets of fer-mentation, each time in duplicate). For each cell wall sugarsand each time of incubation, means within the same columnwith a di†erent number of asterisks di†er by ANOVA (leastsigniÐcant di†erence) at P\ 0É05.b Glucose is cell-wall glucose (ie total glucose[ starch).
TABLE 3Degradation of barley bran cell wall polysaccharides after 6,12, 24 h of in vitro fermentation with human faecal bacteriaa
T ime of incubation (h)
6 12 24
ArabinoseRF 25 ^ 0 36 ^ 8 59 ^ 1IR 21 ^ 3 42 ^ 1 48 ^ 4IE 21 ^ 0 32 ^ 5 50 ^ 1
GlucosebRF 25 ^ 3* 39 ^ 6* 55 ^ 3*IR 3 ^ 4 26 ^ 1*,** 33 ^ 5IE 3 ^ 3 9 ^ 2** 28 ^ 2
XyloseRF 10 ^ 1 22 ^ 6 47 ^ 4IR 16 ^ 4 35 ^ 1 44 ^ 5IE 15 ^ 4 25 ^ 4 46 ^ 3
Total sugarsRF 24 ^ 1 36 ^ 6 56 ^ 3IR 13 ^ 4 34 ^ 1 42 ^ 5IE 11 ^ 5 22 ^ 4 42 ^ 2
a % apparent disappearance ; mean^ SEM (two sets of fer-mentation, each time in duplicate). For each cell wall sugarsand each time of incubation, means within the same columnwith a di†erent number of asterisks di†er by ANOVA (leastsigniÐcant di†erence) at P\ 0É05.b Glucose is cell-wall glucose, ie mainly b-glucans and cellu-lose.
rather di†erent behaviour (Table 3). SigniÐcant di†er-ence in cell-wall sugars disappearance was only appar-ent for glucose but not for other sugars and totalsugars. For glucose, RF, IR and IE showed very distinctproÐles : glucose from RF was Ðrst rapidly degradedand then its degradation rate plateaued whereas glucosefrom IR and IE exhibited signiÐcantly lower rates ofdegradation. At 24 h, only half of the arabinose andxylose had disappeared, together with half of theglucose in RF and less than half in IR and IE. The rateof disappearance of arabinose was higher than that ofxylose during the Ðrst 6 h of incubation.
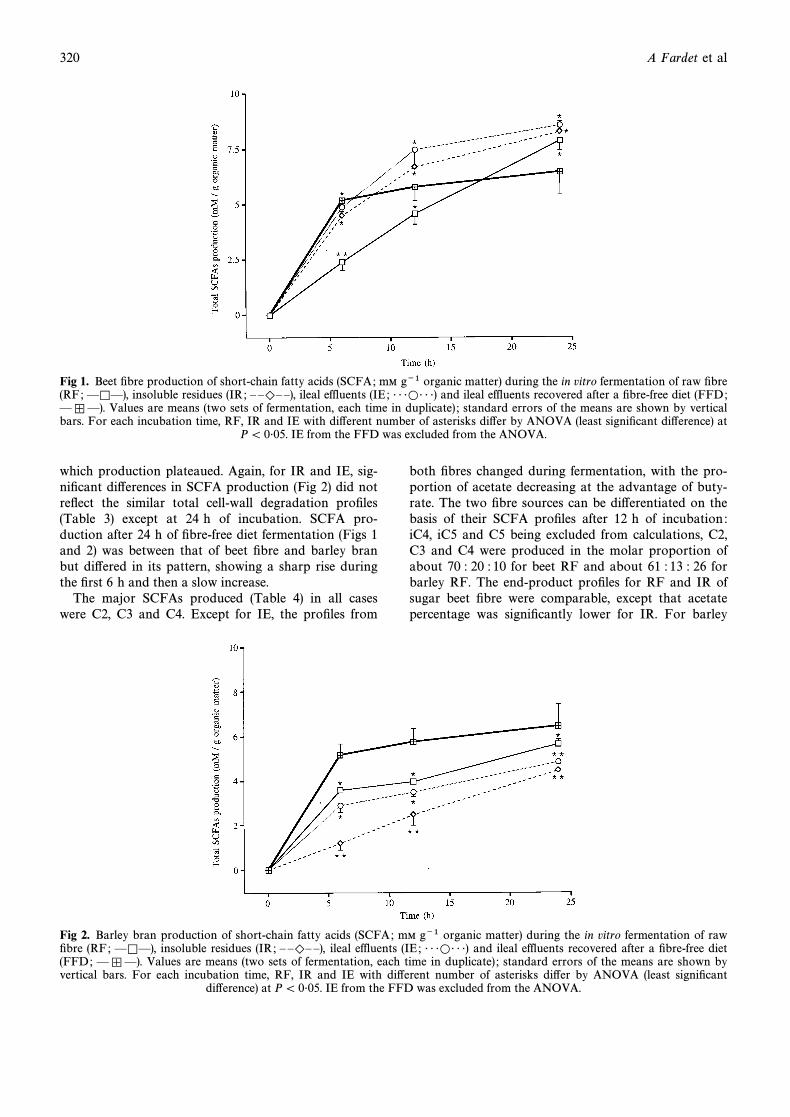
SCFA production was greater during beet Ðbre thanbarley bran fermentation, whatever the form of the sub-strate, ie RF, IR or IE (Figs 1 and 2). SCFA productionfrom beet Ðbre regularly increased for RF while it wasinitially high for IR and IE before plateauing at8É5 mmol per g of organic matter at 24 h. For IR andIE, the similar proÐles of the SCFA production (Fig 1)did not reÑect the signiÐcant di†erences in total cell-wall sugars degradation (Table 2) at each incubationtime. For barley bran, total SCFA production after 24 hof fermentation was in the order RF[ IE \ IR. Fer-mentation of RF and IE produced a higher amount ofSCFA than fermentation of IR for the Ðrst 6 h, after
320 A Fardet et al
Fig 1. Beet Ðbre production of short-chain fatty acids (SCFA; mM g~1 organic matter) during the in vitro fermentation of raw Ðbre(RF; insoluble residues (IR; ileal effluents (IE ; and ileal effluents recovered after a Ðbre-free diet (FFD;ÈKÈ), È È)È È), É É ÉLÉ É É)
Values are means (two sets of fermentation, each time in duplicate) ; standard errors of the means are shown by verticalÈ º È).bars. For each incubation time, RF, IR and IE with di†erent number of asterisks di†er by ANOVA (least signiÐcant di†erence) at
P\ 0É05. IE from the FFD was excluded from the ANOVA.
which production plateaued. Again, for IR and IE, sig-niÐcant di†erences in SCFA production (Fig 2) did notreÑect the similar total cell-wall degradation proÐles(Table 3) except at 24 h of incubation. SCFA pro-duction after 24 h of Ðbre-free diet fermentation (Figs 1and 2) was between that of beet Ðbre and barley branbut di†ered in its pattern, showing a sharp rise duringthe Ðrst 6 h and then a slow increase.
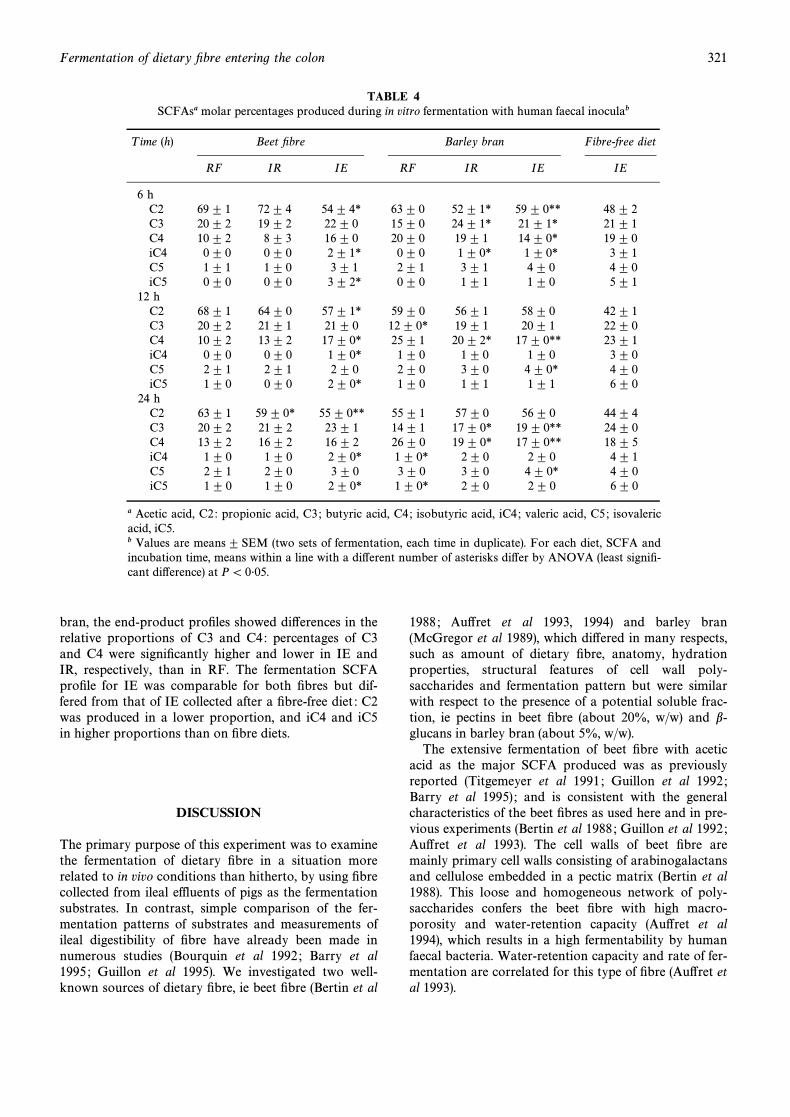
The major SCFAs produced (Table 4) in all caseswere C2, C3 and C4. Except for IE, the proÐles from
both Ðbres changed during fermentation, with the pro-portion of acetate decreasing at the advantage of buty-rate. The two Ðbre sources can be di†erentiated on thebasis of their SCFA proÐles after 12 h of incubation :iC4, iC5 and C5 being excluded from calculations, C2,C3 and C4 were produced in the molar proportion ofabout 70 : 20 : 10 for beet RF and about 61 : 13 : 26 forbarley RF. The end-product proÐles for RF and IR ofsugar beet Ðbre were comparable, except that acetatepercentage was signiÐcantly lower for IR. For barley
Fig 2. Barley bran production of short-chain fatty acids (SCFA; mM g~1 organic matter) during the in vitro fermentation of rawÐbre (RF; insoluble residues (IR; ileal effluents (IE ; and ileal effluents recovered after a Ðbre-free dietÈKÈ), È È)È È), É É ÉLÉ É É)(FFD; Values are means (two sets of fermentation, each time in duplicate) ; standard errors of the means are shown byÈ º È).vertical bars. For each incubation time, RF, IR and IE with di†erent number of asterisks di†er by ANOVA (least signiÐcant
di†erence) at P\ 0É05. IE from the FFD was excluded from the ANOVA.
Fermentation of dietary Ðbre entering the colon 321
TABLE 4SCFAsa molar percentages produced during in vitro fermentation with human faecal inoculab
T ime (h) Beet Ðbre Barley bran Fibre-free diet
RF IR IE RF IR IE IE
6 hC2 69^ 1 72 ^ 4 54 ^ 4* 63 ^ 0 52 ^ 1* 59 ^ 0** 48 ^ 2C3 20^ 2 19 ^ 2 22 ^ 0 15 ^ 0 24 ^ 1* 21 ^ 1* 21 ^ 1C4 10^ 2 8 ^ 3 16 ^ 0 20 ^ 0 19 ^ 1 14 ^ 0* 19 ^ 0iC4 0^ 0 0 ^ 0 2 ^ 1* 0 ^ 0 1 ^ 0* 1 ^ 0* 3 ^ 1C5 1^ 1 1 ^ 0 3 ^ 1 2 ^ 1 3 ^ 1 4 ^ 0 4 ^ 0iC5 0^ 0 0 ^ 0 3 ^ 2* 0 ^ 0 1 ^ 1 1 ^ 0 5 ^ 1
12 hC2 68^ 1 64 ^ 0 57 ^ 1* 59 ^ 0 56 ^ 1 58 ^ 0 42 ^ 1C3 20^ 2 21 ^ 1 21 ^ 0 12 ^ 0* 19 ^ 1 20 ^ 1 22 ^ 0C4 10^ 2 13 ^ 2 17 ^ 0* 25 ^ 1 20 ^ 2* 17 ^ 0** 23 ^ 1iC4 0^ 0 0 ^ 0 1 ^ 0* 1 ^ 0 1 ^ 0 1 ^ 0 3 ^ 0C5 2^ 1 2 ^ 1 2 ^ 0 2 ^ 0 3 ^ 0 4 ^ 0* 4 ^ 0iC5 1^ 0 0 ^ 0 2 ^ 0* 1 ^ 0 1 ^ 1 1 ^ 1 6 ^ 0
24 hC2 63^ 1 59 ^ 0* 55 ^ 0** 55 ^ 1 57 ^ 0 56 ^ 0 44 ^ 4C3 20^ 2 21 ^ 2 23 ^ 1 14 ^ 1 17 ^ 0* 19 ^ 0** 24 ^ 0C4 13^ 2 16 ^ 2 16 ^ 2 26 ^ 0 19 ^ 0* 17 ^ 0** 18 ^ 5iC4 1^ 0 1 ^ 0 2 ^ 0* 1 ^ 0* 2 ^ 0 2 ^ 0 4 ^ 1C5 2^ 1 2 ^ 0 3 ^ 0 3 ^ 0 3 ^ 0 4 ^ 0* 4 ^ 0iC5 1^ 0 1 ^ 0 2 ^ 0* 1 ^ 0* 2 ^ 0 2 ^ 0 6 ^ 0
a Acetic acid, C2 : propionic acid, C3 ; butyric acid, C4 ; isobutyric acid, iC4 ; valeric acid, C5 ; isovalericacid, iC5.b Values are means^ SEM (two sets of fermentation, each time in duplicate). For each diet, SCFA andincubation time, means within a line with a di†erent number of asterisks di†er by ANOVA (least signiÐ-cant di†erence) at P\ 0É05.
bran, the end-product proÐles showed di†erences in therelative proportions of C3 and C4: percentages of C3and C4 were signiÐcantly higher and lower in IE andIR, respectively, than in RF. The fermentation SCFAproÐle for IE was comparable for both Ðbres but dif-fered from that of IE collected after a Ðbre-free diet : C2was produced in a lower proportion, and iC4 and iC5in higher proportions than on Ðbre diets.
DISCUSSION
The primary purpose of this experiment was to examinethe fermentation of dietary Ðbre in a situation morerelated to in vivo conditions than hitherto, by using Ðbrecollected from ileal effluents of pigs as the fermentationsubstrates. In contrast, simple comparison of the fer-mentation patterns of substrates and measurements ofileal digestibility of Ðbre have already been made innumerous studies (Bourquin et al 1992 ; Barry et al1995 ; Guillon et al 1995). We investigated two well-known sources of dietary Ðbre, ie beet Ðbre (Bertin et al
1988 ; Au†ret et al 1993, 1994) and barley bran(McGregor et al 1989), which di†ered in many respects,such as amount of dietary Ðbre, anatomy, hydrationproperties, structural features of cell wall poly-saccharides and fermentation pattern but were similarwith respect to the presence of a potential soluble frac-tion, ie pectins in beet Ðbre (about 20%, w/w) and b-glucans in barley bran (about 5%, w/w).
The extensive fermentation of beet Ðbre with aceticacid as the major SCFA produced was as previouslyreported (Titgemeyer et al 1991 ; Guillon et al 1992 ;Barry et al 1995) ; and is consistent with the generalcharacteristics of the beet Ðbres as used here and in pre-vious experiments (Bertin et al 1988 ; Guillon et al 1992 ;Au†ret et al 1993). The cell walls of beet Ðbre aremainly primary cell walls consisting of arabinogalactansand cellulose embedded in a pectic matrix (Bertin et al1988). This loose and homogeneous network of poly-saccharides confers the beet Ðbre with high macro-porosity and water-retention capacity (Au†ret et al1994), which results in a high fermentability by humanfaecal bacteria. Water-retention capacity and rate of fer-mentation are correlated for this type of Ðbre (Au†ret etal 1993).

322 A Fardet et al
The observation that barley bran was, in contrast tobeet Ðbre, only half-degraded during fermentation isalso consistent with its physical properties. It is an het-erogeneous structure due to a range of tissues includingthe pericarp with the attached testa, aleuron layers andpart of the starchy endosperm (McGregor et al 1989),each tissue being degraded to di†erent extent in fermen-tation. Cell wall sugars of the aleuron layer and of thestarchy endosperm were extensively degraded while thepericarp mostly remained (light micrographs notshown), probably because its fermentability was limitedby the presence of highly branched glucuronoarabin-oxylans and crystalline cellulose, as has been shown forwheat bran (Salvador et al 1993). The pericarp-testalayer is a very rigid structure and constitutes a barrierto bacterial enzymes, thus resisting degradation andalso protection adjacent cells. Endosperm-aleuronlayers possess a looser structure that is more accessibleto bacterial enzymes and also contain most of the seedb-glucans (McMregor et al 1989). Their susceptibility tofermentation explains why fermentation of cell wallglucose is generally high in barley and oat brans com-pared to others of cereal origin such as wheat (Bourquinet al 1992) and maize (Barry et al 1995) brans, whosecellulose is known to be poorly degraded (Jeraci andHorvath 1989).
Barley bran is a by-product containing large amountsof intracellular compounds (57% of dry matter forstarch and protein) which could have been enzymically(Hoebler et al 1991) removed to avoid production ofSCFA from non-Ðbre material. Such a pretreatmentwas, however, omitted because it can lead to partialsolubilisation of cell wall sugars (about 11% of wheatbran carbohydrates, solubilised, Hoebler et al 1991)which would have modiÐed cell wall sugar disap-pearance during fermentation, and also because thematerial would no longer have represented a food com-ponent as consumed. Starch, the more accessible sub-strate, was responsible for the large amount of SCFAproduced as soon as 6 h of fermentation and for thehigh proportion of butyrate obtained at 24 h incubationcompared to beet Ðbre (Englyst and Macfarlane 1987 ;Nordgaard et al 1995).
Apart from showing that intrinsic di†erences betweenÐbre sources have an important inÑuence on the extent,rate and pattern of bacterial digestion, our results alsoshow that it is important to consider the physical formsand physicochemical properties of the Ðbres asexpressed in the conditions of terminal ileum, afterchanges resulting from oroileal transit. The increasedinitial rate of degradation of beet Ðbre from the termin-al ileum is likely to have resulted from physicochemicalmodiÐcations during oroileal transit which loosened thebeet Ðbre matrix, leading to a more rapid di†usion ofbacteria into the substrate, accelerating the breakdownof the Ðbrous structure. Results, presented in Tables 2and 4, are consistant with such changes, as the rate of
sugar disappearance and SCFA production from beetwas increased even though the extent of fermentation ofbeet Ðbre before and after passage through the gut werein the same range. Similar results have been obtainedwhen pectins were solubilised by chemical reagents(Au†ret et al 1993) loosening cell wall material andincreasing water-retention capacity and macroporosityof the Ðbre matrix (Au†ret et al 1993). In our study noselective losses of uronic acids occured, implying that invivo di†erent phenomena, such as the chemical environ-ment, microbial activity and physical activity of the gutmay contribute to these changes. Mastication is usuallythe Ðrst event reducing Ðbre particle size, but in ourexperimental conditions particle size was small enoughnot to require mastication. Disruption of calciumbridges between pectin macromolecules under acidicconditions in the stomach, or b-eliminative degradationof methyl esteriÐed pectins at neutral pH in the ileum,may favour maceration. In association with the mecha-nical activity needed to mix and propel digesta, suchchemical changes could contribute to decreasing parti-cle size hence increasing accessible surface for enzymesand bacteria, and to modiÐcation of the hydrationproperties of the Ðbre matrix. Degradation by bacteriacolonising the pig ileum and possibly the proximalintestine could also cause changes in Ðbre before itsarrival in the caecum (Millard and Chesson 1984 ;Robertson et al 1992). But in the present experiment apH of 7É2 ^ 0É2 and SCFA concentration of29É3 ^ 18É6 mmol litre~1 in the terminal ileum indicatesabsence of intensive fermentation (results not shown),although fermentation of bacteria colonising thestomach can not be excluded. Whatever the cause, ourresults show that sugar-beet Ðbre is fermented at ahigher rate under physiological conditions than can bepredicted from in vitro fermentation with raw material.
Bacterial degradation by barley bran di†ered fromthat of beet Ðbre in that cell wall present in IR exhibiteda rather lower rate of degradation than cell wall in RF,even if not statistically signiÐcant. This was mainly dueto a signiÐcant decrease in degradation of cell wallglucose, especially b-glucan, as a result of its partialsolubilisation in the upper gut (Bach Knudsen et al1993). These Ðndings are in good agreement with pre-vious results showing that b-glucans from oat bran weresolubilised and that their degree of polymerizationdecreased in the upper digestive tract (Johansen et al1994). Bach Knudsen et al (1993) estimated the ilealdigestibility of oat bran b-glucans to be 17% in pig ter-minal ileum. He suggested that microbial activity in thestomach and small intestine is responsible for the fer-mentation.
The results from both chemical analysis (Table 1) andfermentation (Tables 2 and 3) that are reported hereindicate that there is a considerable and inÑuentialquantity of material that behaves as dietary Ðbre at theterminal ileum, but that would not be accounted for in

Fermentation of dietary Ðbre entering the colon 323
either a conventional Ðbre analysis or in a fermentabil-ity study of a food before consumption. Fibre poly-saccharides accounted for only one-third to one-half ofthe dry weight of total ileal effluents reaching the largeintestine, which is in agreement with results in the liter-ature (Cummings and McFarlane 1991). The discrep-ancies between cell wall sugar disappearance and SCFAproduction, irrespective of the incubation time, showedthat Ðbres were not the only fermented substrate andthat the non-cell-wall fraction produced signiÐcantquantities of SCFAs. Non-cell wall compounds wererapidly metabolised by bacteria, as shown by the highrate of SCFAs production at 6 h of incubation in IEfrom the Ðbre-free diet, and may be preferred substrates,as the delayed degradation of beet sugars in IE com-pared to IR suggests. Perhaps this could be explainedby the preferential degradation of mucins by strains ofbacteria such as Bacteroides or BiÐdobacteria, whichdegrade mucins as well as pectins (Salyers 1977 ; Mac-farlane and Cummings 1991). Mucins are endogenousand a familiar source of continuous substrate for thisbacteria, which were probably already present at thebeginning of the in vitro fermentation. Therefore, thetwo substrates each competed with the other to bedegraded. In the case of barley bran IR, which wasslowly degraded, endogenous substances did not inÑu-ence the rate of disappearance of cell wall sugars, prob-ably because there was no overlap between utilisation ofnon-cell-wall compounds and cell wall sugars as sub-strates. However, the higher SCFA production in theÐrst 12 h of fermentation of IE again demonstrated asigniÐcant contribution of non-cell-wall compounds tofermentation.
A considerable inÑuence of endogenous substanceson SCFA proÐles was also evident in the results pre-sented here. SCFA proÐles from fermentation of IE dif-fered from those due to IR fermentation. Lower acetateproduction for beet Ðbre IE probably resulted from lossof pectic in the digestive tract. Pectic fermentation char-acteristically produces high levels of acetate (Titgemeyeret al 1991). In barley bran RF, starch fermentation wasresponsible for the high percentage of butyrate pro-duction (Englyst and Macfarlane 1987) ; therefore thereduced production of butyrate during the in vitro fer-mentation of IR may be ascribed to the starch digestionin the small intestine of pigs. With IE, the di†erences ofSCFAs proÐles between beet and barley bran Ðbresources were Ñattened. However, compared to theSCFA proÐle of IE collected after a Ðbre-free diet (basaldiet), a Ðbre e†ect existed. Two explanations are avail-able : in vitro, fermentations were carried out with equiv-alent amount of organic matter. The proportion of ilealeffluent that is Ðbre is lower than in IR, and so the pro-portion of SCFAs derived from Ðbre fermentation islower. Secondly, endogeneous substances are substratesfor bacteria and so produce SCFAs, which will dilutethose arising from Ðbre fermentation. Flattening of
SCFA proÐle in IE was in good agreement with theresults of McBurney et al (1988) for Ðbre extracted fromwhite bread, oat bran, kidney beans and lentils. Theyfound little variability between the di†erent SCFA pro-Ðles of IE, providing evidence for the impact of theupper intestinal tract in modulating ileal effluent com-position. The higher proportions of branched SCFAs inIE, especially IE collected after the Ðbre-free diet, camefrom amino acid fermentation (Rasmussen et al 1988 ;Macfarlane and Cummings, 1987) which were providedby endogenic substances as mucins and digestiveenzymes.
Most in vitro studies of Ðbre fermentation use Ðbre aseaten. Our results, demonstrating that, under physio-logical conditions, Ðbre undergoes a conditioningprocess that changes its properties before arrival at thelarge intestine, show that it is important to take suchchanges into account. The impact of gastrointestinalconditions depends on the Ðbre : for beet Ðbre, due to itshomogeneous structure, the impact is on the initial rateof fermentation, whereas for barely bran, the impact ison SCFA proÐles and rate of SCFA production due toloss of b-glucans and retention of starch and protein.Non-cell-wall compounds must therefore also be takeninto account since they modify the kinetics of Ðbre fer-mentation and provide signiÐcant quantities of iso-SCFAs and valerate. These results suggest that RFcould be used as a good model of IE for ranking Ðbreaccording to their degree of fermentability. However,for quantitatively predicting rate of fermentation orSCFA proÐles, IE should be used to best Ðt the in vivosituation.
ACKNOWLEDGEMENTS
The authors wish to thank C Bonnet and M Rival fortheir valuable assistance with the in vitro fermentationsand physicochemical analyses and J A Monro, from theNew-Zealand Institute for Crop and Food ResearchLtd, for his comments.
REFERENCES
AFNOR (Association de Normalisation) 1987 Stan-FrancÓ aisedards NF X-11-635 and NF X-11-636. In : Recueil deNormes Franaises. AFNOR, Paris, France.
A- man P, Zhang J X, Hallmans G, Lundin E 1994 Excretionand degradation of dietary Ðber constituents in ileostomysubjects consuming a low Ðber diet with and withoutbrewerÏs spent grain. J Nutr 124 359È363.
Au†ret A, Barry J-L, Thibault J-F 1993 E†ect of chemicaltreatments of sugar beet Ðbre on their physico-chemicalproperties and on their in-vitro fermentation. J Sci FoodAgric 61 195È203.

324 A Fardet et al
Au†ret A, Ralet M-C. Guillon F, Barry J-L, Thibault J-F 1994E†ect of grinding and experimental conditions on the mea-surement of hydration properties of dietary Ðbres. L ebensmW iss T echnol 27 166È172.
Bach Knudsen K E, Borg Jensen B, Hansen I 1993 Digestionof polysaccharides and other major components in thesmall and large intestine of pigs fed on diets consisting ofoat fractions rich in b-D-glucan. Br J Nutr 70 537È556.
Barry J-L, Hoebler C, Macfarlane G T, Mathers J C, Reed KA, Mortensen P B, Norgaard I, Rowland I R, Rumney C J1995 Estimation of the fermentability of dietary Ðbre : aEuropean interlaboratory study in-vitro. Br J Nutr 74 303È322.
Bertin C, Rouau X, Thibault J-F 1988 Structure and proper-ties of sugar-beet Ðbre J Sci Food Agric 44 15È29.
Bourquin L D, Titgemeyer E C, Garleb K A, Fahey G C 1992Short-chain fatty acid production and Ðber degradation byhuman colonic bacteria : e†ects of substrates and cell wallfractionation procedures. J Nutr 122 1508È1520.
Cherbut C, Salvador V, Barry J-L, Doulay F, Delort-Laval J1991 Dietary Ðbre e†ects on intestinal transit in man:involvement of their physicochemical and fermentativeproperties. Food Hydrocoll 5 15È22.
Cummings J-H, Macfarlane G T 1991 A review: the controland consequences of bacterial fermentation in the humancolon. J Appl Bacteriol 70 443.
Durand M, Komisarczuk S 1988 InÑuence of major mineralson rumen microbiota J Nutr 118 249È260.
Edwards C A, Gibson G, Champ M, Jensen B-B, Mathers JC, Nagengast F, Rumney C, Quehl A 1996 In vitro methodfor quantiÐcation of the fermentation of starch by humanfaecal bacteria. J Sci Food Agric 71 209È217.
Englyst H N, Macfarlane G T 1987 Polysaccharide break-down by mixed populations of human faecal bacteria.FEMS Microbiol Ecol 95 163.
Graham H, Hesselman K, A- man P 1986 The inÑuence ofwheat bran and sugar beet pulp on the digestibility ofdietary components in a cereal-based pig diet. J Nutr 116242È251.
Guillon F, Barry J-L, Thibault J-F 1992 E†ect of autoclavingsugar-beet Ðbre on its physico-chemical properties and itsin-vitro degradation by human faecal bacteria. J Sci FoodAgric 60 69È79.
Guillon F, Renard C, Hospers J, Thibault J-F, Barry J L 1995Characterisation of residual Ðbres from fermentation of peaand apple Ðbres by human colonic bacteria. J Sci FoodAgric 68 521È529.
Hoebler C, Barry J-L, David A, Delort-Laval J 1989 Rapidacid hydrolysis of plant cell wall polysaccharides and sim-pliÐed quantitative determination of their neutral monosac-charides by gas-liquid chromatography. J Agric Food Chem37 360È367.
Hoebler C, Barry J-L, David A, Kozlowski F 1991 Enzymaticpreparation and analysis of dietary Ðbre residues, suitablefor in-vitro fermentation studies. Food Hydrocoll 5 35È40.
Holloway W D, Tasman-Jones C, Lee S P 1978 Digestion ofcertain fractions of dietary Ðbre in humans. Am J Clin Nutr31 927È930.
Jeraci J L, Horvath P J 1989 In-vitro fermentation of dietaryÐber by human fecal organisms. Anim Feed Sci T echnol 23121È140.
Johansen H N, Bach Knudsen K E, Wood P J, Fulcher R G1994 Solubilisation and degradation of cell-wall poly-saccharides from oats in the gastro-intestinal tract of pigs.In : Proc ÏEuropean COST 92Ï, ed Amado R, Barry J-L &Frolich W. Carry Le Rouet, Marseille, France, pp 115È118.
Jouany J-P 1982 Dosage des acides gras volatils et des alcoolsdans les contenus digestifs, les jus dÏensilage, les cultures
bacte� riennes et les contenus de fermenteurs anae� robis. SciAlim 2 131È144.
Karkalas J 1985 An improved enzymic method for the deter-mination of the raw and modiÐed starch. J Sci Food Agric36 1019È1027.
Livingstone R-M, Baird B-A, Atkinson T, Crofts R M J 1980Circadian variation in the apparent digestibility of dietsmeasured at the terminal ileum in pigs. J Agric Sci Camb 94339È405.
McBurney M I, Thompson L U 1987 E†ect of human fecalinoculum on in-vitro fermentation variables. Br J Nutr 58223È243.
McBurney M I, Horvath P J, Jeraci J L, Van Soest P J 1985E†ect of in-vitro fermentation using human faecal inoculumon the water-holding capacity of dietary Ðbre. Br J Nutr 5317È24.
McBurney M I, Thompson L U, Cu† D J, Jenkins D J A 1988Comparison of ileal effluents, dietary Ðbers, and wholefoods in predicting the physiological importance of colonicfermentation. Am J Gastroenterol 83 536È540.
McCleary B V, Codd R 1991 Measurement of (1] 3), (1 ] 4)-b-D-glucan in barley and oats : a streamlined enzymatic pro-cedure. J Sci Food Agric 55 303È312.
McGregor A W, Ballance G M, Dushnicky L 1989 Fluores-cence microscopy studies on (1,3)-b-D-glucan in barleyendosperm. Food Microstruct 8 235È244.
Macfarlane G T, Cummings J H 1991 The colonic Ñora, fer-mentation and large bowel digestive function. In : T heL arge Intestine : Physiology, Pathophysiology and Disease,eds Phillips S F, Pemberton J H & Shorter R G. RavenPress, New York, USA, pp 51È92.
Melcion J-P, Delort-Laval J 1981 Me� thodes objectivesdÏappre� ciation des caracte� ristiques physiques des alimentscompose� s. In : de la V aleur Nutritive des AlimentsPre� visiondes Ruminants. INRA, Versailles, France, pp 307È323.
Millard P, Chesson A 1984 ModiÐcations to swede (Brassicanapus L.) anterior to the terminal ileum of pigs : some impli-cations for the analysis of dietary Ðbre. Br J Nutr 52 583È594.
Nordgaard I, Mortensen P B 1995 Digestive processes in thehuman colon. Nutrition 11 37È45.
Nordgaard I, Mortensen P B, Langkilde A M 1995 Smallintestine malabsorption and colonic fermentation of resist-ant starch and resistant peptides to short-chain fatty acids.Nutrition 11 129È137.
Nyman M G, Asp N G, Cummings J, Wiggins 1986 Fermen-tation of dietary Ðbre in the intestinal tract : comparisonbetween man and rat. Br J Nutr 55 487È496.
Rasmussen H S, Holtug K, Mortensen B P 1988 Degradationof amino acids to short-chain fatty acids in humans. Scand.J Gastroenterol 23 178È182.
Robertson J A 1988 Physico-chemical characteristics of foodand the digestion of starch and dietary Ðbre during guttransit. Proc Nutr Soc 47 143È152.
Robertson J A, Eastwood M A 1981 A method to measure thewater-holding properties of dietary Ðbre using suction pres-sure. Br J Nutr 46 247È255.
Robertson J A, Murison S D, Chesson A 1992 Particle sizedistribution and solubility of dietary Ðbre in swede(Brassica Napus)-based and wheat-bran-based diets duringgastrointestinal transit in the pig. J Sci Food Agric 58 197È205.
Salvador V, Cherbut C, Barry J-L, Bertrand D, Bonnet C,Delort-Laval J 1993 Sugar composition of dietary Ðbre andshort-chain fatty acid production during in-vitro fermenta-tion by human bacteria. Br J Nutr 70 189È197.
Salyers A A 1977 Fermentation of mucins and plant poly-saccharides by Bacteroides from the human colon. Appl

Fermentation of dietary Ðbre entering the colon 325
Environ Microbiol 33 319È322.Silvester K R, Englyst H N, Cummings J H 1995 Ileal
recovery of starch from whole diets containing resistantstarch measured in-vitro and fermentation of ileal effluents.Am J Clin Nutr 62 403È411.
Stephen A M, Cummings J H 1979 Water holding by dietaryÐbre in-vitro and its relationship to faecal output in man.
Gut 20 722È729.Thibault J-F 1979 Automatisation du dosage des substances
pectiques par la me� thode au me� ta-hydroxydiphenyl.L ebensm W iss T echnol 12 247È251.
Titgemeyer E C, Bourquin L D, Fahey G C, Garleb K A 1991Fermentability of various Ðber sources by human fecal bac-teria in-vitro. Am J Clin Nutr 53 1418È1424.