high-throughput determination of free d-aspartic acid in mammals by enzyme immunoassay using...
TRANSCRIPT

ANALYTICALBIOCHEMISTRY
Analytical Biochemistry 357 (2006) 15–20
www.elsevier.com/locate/yabio
High-throughput determination of free D-aspartic acid in mammals by enzyme immunoassay using speciWc monoclonal antibody
Tomohiro Ohgusu, Kenji Hamase, Hiroyuki Tanaka, Yukihiro Shoyama, Kiyoshi Zaitsu ¤
Graduate School of Pharmaceutical Sciences, Kyushu University, 3-1-1 Maidashi, Higashi-ku, Fukuoka 812-8582, Japan
Received 17 April 2006Available online 31 July 2006
Abstract
A method for rapid determination of free D-aspartic acid (D-Asp) in mammals has been established using a highly speciWc mousemonoclonal antibody against D-Asp for the Wrst time. An anti-D-Asp monoclonal antibody was obtained by the immunization of bovine-serum-albumin-conjugated D-Asp to BALB/c mice. The obtained antibody has a high speciWcity toward D-Asp but shows a slight cross-reactivity to all other D- and L- amino acids including L-Asp. The calibration range of the competitive enzyme linked immunosorbentassay (ELISA) is 0.016–16 �mol/mL D-Asp in rat serum samples. The precisions of this method were evaluated by inter-plate and intra-plate assays, and the relative standard deviation values were 4.8% and 4.5%, respectively. The values of D-Asp determined by the presentELISA have a good correlation to those determined by high-performance liquid chromatography with the correlation coeYcient of 0.963.Using this ELISA, the time course of D-Asp in the rat serum after intravenous administration was successfully demonstrated. The presentmethod provides a simple and high-throughput determination of D-Asp in mammals, and is a useful tool for clarifying the physiologicalroles and diagnostic values of this D-amino acid.© 2006 Elsevier Inc. All rights reserved.
Keywords: D-Asp; Monoclonal antibody; ELISA
It has long been believed that only L-amino acids arepresent in higher animals and that their enantiomers, D-amino acids, are present only in the lower species such asmicroorganisms [1,2] and crustaceans [3,4]. However, recentstudies on the enantiomer separation of amino acids in var-ious organisms revealed that even in the higher animalsincluding humans some of the D-amino acids are preset intheir free form [5,6] and in the protein-bound form as D-amino acid residues [7,8]. Free D-aspartic acid (D-Asp) isone of the well-investigated D-amino acids, which is widelyobserved in mammals especially in the endocrine tissuessuch as the pineal gland [9,10], testis [11,12], adrenal gland[13,14], and pituitary gland [15,16]. In these tissues, D-Asphas the function of regulating hormonal secretion, e.g.,stimulating testosterone synthesis in the Leydig cells [12] orinhibiting the norepinephrine-induced melatonin secretion
* Corresponding author. Fax: +81 92 642 6601.E-mail address: [email protected] (K. Zaitsu).
0003-2697/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.ab.2006.06.039
[17]. It is also indicated that, after its intraperitoneal admin-istration, D-Asp is immediately distributed to the pinealgland, pituitary gland, and testis, and the amounts of lutein-izing hormone, testosterone, and progesterone in the bloodare increased [11].
Free D-Asp has also been reported to have close relation-ships to some diseases [5,6,18–21]. In patients of Alzhei-mer’s disease, the concentration of free D-Asp in the brainwhite matter is less than half of that in normal brains [18].It is also suggested that the reduction of the free D-Asp levelin the brain of Alzheimer’s patients results in the decreasedsignal transduction via the N-methyl-D-aspartate receptor[19]. D-Asp is also related to renal disorders; a higheramount of D-Asp was present in the sera of patients com-pared to that in those of healthy volunteers [21]. The con-centration of D-Asp in the serum had a good positivecorrelation to the creatinine levels [21], which is an index ofrenal function. Though various reports concerning freeD-Asp in mammals have been published, many of the

16 ELISA of D-Asp in rat serum / T. Ohgusu et al. / Anal. Biochem. 357 (2006) 15–20
functions, origin, and regulation mechanisms remainunknown, and a rapid analytical method for the quantita-tion of D-Asp in mammals is highly anticipated. The high-throughput assay of D-Asp enables the rapid screening oflarge numbers of clinical samples or those obtained fromlaboratory animals under various physiological conditions;the evaluation of the D-Asp amounts in these samples isimportant to clarify the physiological roles and diagnosticvalues of this D-amino acid.
The aim of the present investigation was to establish ahigh-throughput assay of free D-Asp in mammals. Untilnow, various analytical methods have been reported for thedetermination of D-Asp in mammals using gas chromatog-raphy [14,22], high-performance liquid chromatography(HPLC) [23–29], and capillary electrophoresis [30,31]. All ofthese methods include highly selective chromatographic orelectrophoretic separations using various chiral stationaryphases, chiral mobile phases, or chiral derivatizationreagents. Using theses techniques, the distribution of D-Aspin mammalian brain, peripheral tissues, and physiologicalXuids has been elucidated and described [14,23]. The quan-titative variation through the pre- and postnatal life hasalso been demonstrated [27]. Although use of these meth-ods reveals the highly selective determination of D-Asp inmammals, all of them need derivatization with chiral activecompounds or with reagents having strong Xuorescence orabsorption. Laborious sample pretreatment steps are alsoneeded, and the Wnal chromatographic or electrophoreticseparation requires 10–60 min for analyzing one sample.Therefore, a more high-throughput assay of D-Asp isrequired for the rapid screening of its physiological rolesand diagnostic values. In the present study, we have pre-pared a speciWc monoclonal antibody against free D-Aspfor the Wrst time and have established a rapid and simpleenzyme-linked immunosorbent assay (ELISA)1 of D-Asp.Using this method, the time course of D-Asp amounts in ratserum after the intravenous administration of this D-aminoacid has been successfully demonstrated.
Materials and methods
Chemicals
D-Aspartic acid and glutaraldehyde (25% aqueous solu-tion) were purchased from Nacalai Tesque (Kyoto, Japan).Bovine serum albumin (BSA), hypoxantine, aminopterine,and thymidine were obtained from Sigma–Aldrich Co. (St.Louis, MO, USA). Ovalbumin (OVA) was from SeikagakuCorp. (Tokyo, Japan). Freund’s complete adjuvant (FCA)and Freund’s incomplete adjuvant (FIA) were from DifcoLaboratories (Detroit, MI, USA). Polyethylene glycol 1500
1 Abbreviations used: ELISA, enzyme-linked immunosorbent assay;BSA, bovine serum albumin; OVA, ovalbumin; FCA, Freund’s completeadjuvant; FIA, Freund’s incomplete adjuvant; SPF, speciWc pathogen free;PBS, phosphate-buVered saline; HRP, horseradish peroxidase; MALDI-TOF, matrix-assisted laser desorption ionization time of Xight.
was from Roche (Indianapolis, IN, USA), and 2,2�-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)diammoniumsalt (ABTS) was from Wako Pure Chemical Ind., Ltd.(Osaka, Japan). The water was puriWed by a Milli-Q system(Millipore, Bedford, MA, USA). All other reagents and sol-vents were of guaranteed grade.
Animals
Female BALB/c mice (6 weeks of age, speciWc pathogenfree (SPF)) and male Wistar rats (7 weeks of age, SPF) werepurchased from Kyudo (Kumamoto, Japan). They werehoused in an animal center of Kyushu University, Gradu-ate School of Pharmaceutical Sciences, under a 12-h light/12-h dark cycle (lights on at 07:00 am). They had free accessto diet (type NMF; Oriental Yeast, Tokyo, Japan) andwater. The present experiments were performed under thepermission (No. PAE04-64) of the animal care and usecommittee of the Graduate School of Pharmaceutical Sci-ences, Kyushu University.
Preparation of anti-D-Asp mouse monoclonal antibody
An anti-D-Asp mouse monoclonal antibody was raisedfollowing authorized methods [32,33]. BrieXy, BSA (18 mg)was dissolved in 0.1 M sodium-phosphate buVer (pH 7.0)containing 62.5 mM D-Asp (2.4 mL), and 2.5% aqueous glu-taraldehyde solution (0.6 mL) was added and stirred for 2 h.An aqueous sodium borohydride solution (4 mg/mL, 3 mL)was then added and stirred for 1 h. The reaction mixturewas dialyzed twice against phosphate-buVered saline (PBS;0.8% NaCl, 0.02% KCl, 0.024% KH2PO4, 0.14% Na2HPO4in H2O, pH 7.4) at 4 °C, and this D-Asp-GA-BSA solution(50 �L) was injected into the BALB/c mice three times every2 weeks (the Wrst injection was done hypodermically as a1:1 emulsion with FCA, and the second injection was donehypodermically as a 1:1 emulsion with FIA). Three daysafter the third intraperitoneal injection, cell fusion was per-formed with splenocytes and hypoxantine-aminopterin-thymidine-sensitive mouse myeloma cells, sp2/0-Ag14(Riken BioResource Center, Tsukuba, Japan), using poly-ethylene glycol. A hybridoma clone was obtained by thelimited dilution method and a cell line was cultured inserum-free medium. From this culture medium, the mono-clonal antibody was puriWed using a HiTrap Protein G HPcolumn (1 cm i.d.£ 2 cm; Amersham Biosciences, Uppsala,Sweden) at 4 °C. Unless speciWed, all experiments were car-ried out at room temperature.
ELISA of D-Asp
D-Asp-GA-OVA (1.5 �g/mL, 100 �L) dissolved in 50 mMsodium-carbonate buVer (pH 9.6) was added to the wells ofa 96-well immunoplate (Nalge Nunc, Roskilde, Denmark)and incubated for 1 h. After washing the plate three timeswith PBS containing 0.05% Tween 20 (T-PBS), PBS con-taining 5% skim milk (300 �L) was added and incubated for
ELISA of D-Asp in rat serum / T. Ohgusu et al. / Anal. Biochem. 357 (2006) 15–20 17
1 h. The plate was washed three times with T-PBS; then20 �L of rat serum and the anti-D-Asp monoclonal anti-body (1 �g/mL in T-PBS, 100 �L) were added. The concen-tration of the anti-D-Asp monoclonal antibody wasdetermined using a control mouse IgG (mouse IgG; ICNPharmaceuticals, Aurora, OH, USA). The plate was incu-bated for 1 h, washed three times with T-PBS, and thenincubated with 100 �L of 1:1000 diluted horseradish-perox-idase (HRP)-conjugated goat IgG fraction to mouse IgG(ICN Pharmaceuticals) for 1 h. After washing the plate withT-PBS three times, substrate solution (0.1 M sodium-citratebuVer (pH 4.0) containing 0.003% H2O2 and 0.3 mg/mLABTS) was added and incubated for 15 min. The absor-bance at 405 nm was then measured by a microplate reader(Model 680 microplate reader, Bio-Rad Japan, Tokyo,Japan).
Results and discussion
Preparation of anti-D-aspartic acid monoclonal antibody
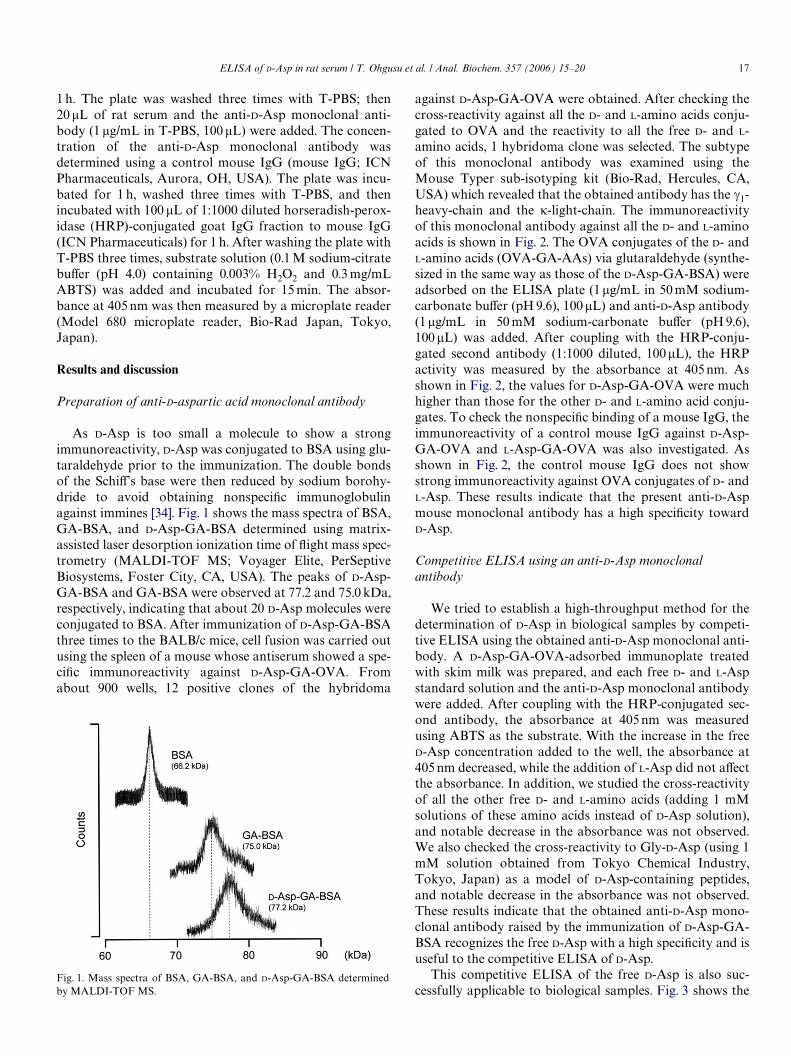
As D-Asp is too small a molecule to show a strongimmunoreactivity, D-Asp was conjugated to BSA using glu-taraldehyde prior to the immunization. The double bondsof the SchiV’s base were then reduced by sodium borohy-dride to avoid obtaining nonspeciWc immunoglobulinagainst immines [34]. Fig. 1 shows the mass spectra of BSA,GA-BSA, and D-Asp-GA-BSA determined using matrix-assisted laser desorption ionization time of Xight mass spec-trometry (MALDI-TOF MS; Voyager Elite, PerSeptiveBiosystems, Foster City, CA, USA). The peaks of D-Asp-GA-BSA and GA-BSA were observed at 77.2 and 75.0 kDa,respectively, indicating that about 20 D-Asp molecules wereconjugated to BSA. After immunization of D-Asp-GA-BSAthree times to the BALB/c mice, cell fusion was carried outusing the spleen of a mouse whose antiserum showed a spe-ciWc immunoreactivity against D-Asp-GA-OVA. Fromabout 900 wells, 12 positive clones of the hybridoma
Fig. 1. Mass spectra of BSA, GA-BSA, and D-Asp-GA-BSA determinedby MALDI-TOF MS.
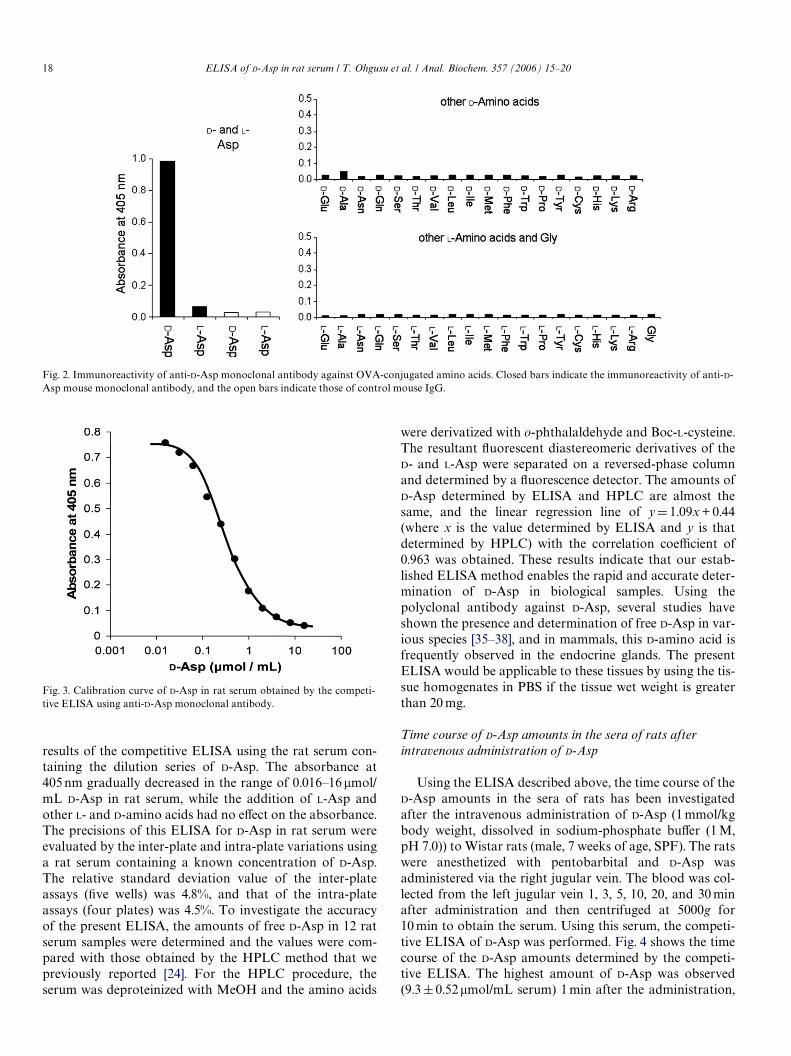
against D-Asp-GA-OVA were obtained. After checking thecross-reactivity against all the D- and L-amino acids conju-gated to OVA and the reactivity to all the free D- and L-amino acids, 1 hybridoma clone was selected. The subtypeof this monoclonal antibody was examined using theMouse Typer sub-isotyping kit (Bio-Rad, Hercules, CA,USA) which revealed that the obtained antibody has the �1-heavy-chain and the �-light-chain. The immunoreactivityof this monoclonal antibody against all the D- and L-aminoacids is shown in Fig. 2. The OVA conjugates of the D- andL-amino acids (OVA-GA-AAs) via glutaraldehyde (synthe-sized in the same way as those of the D-Asp-GA-BSA) wereadsorbed on the ELISA plate (1 �g/mL in 50 mM sodium-carbonate buVer (pH 9.6), 100 �L) and anti-D-Asp antibody(1�g/mL in 50 mM sodium-carbonate buVer (pH 9.6),100 �L) was added. After coupling with the HRP-conju-gated second antibody (1:1000 diluted, 100�L), the HRPactivity was measured by the absorbance at 405 nm. Asshown in Fig. 2, the values for D-Asp-GA-OVA were muchhigher than those for the other D- and L-amino acid conju-gates. To check the nonspeciWc binding of a mouse IgG, theimmunoreactivity of a control mouse IgG against D-Asp-GA-OVA and L-Asp-GA-OVA was also investigated. Asshown in Fig. 2, the control mouse IgG does not showstrong immunoreactivity against OVA conjugates of D- andL-Asp. These results indicate that the present anti-D-Aspmouse monoclonal antibody has a high speciWcity towardD-Asp.
Competitive ELISA using an anti-D-Asp monoclonal antibody
We tried to establish a high-throughput method for thedetermination of D-Asp in biological samples by competi-tive ELISA using the obtained anti-D-Asp monoclonal anti-body. A D-Asp-GA-OVA-adsorbed immunoplate treatedwith skim milk was prepared, and each free D- and L-Aspstandard solution and the anti-D-Asp monoclonal antibodywere added. After coupling with the HRP-conjugated sec-ond antibody, the absorbance at 405 nm was measuredusing ABTS as the substrate. With the increase in the freeD-Asp concentration added to the well, the absorbance at405 nm decreased, while the addition of L-Asp did not aVectthe absorbance. In addition, we studied the cross-reactivityof all the other free D- and L-amino acids (adding 1 mMsolutions of these amino acids instead of D-Asp solution),and notable decrease in the absorbance was not observed.We also checked the cross-reactivity to Gly-D-Asp (using 1mM solution obtained from Tokyo Chemical Industry,Tokyo, Japan) as a model of D-Asp-containing peptides,and notable decrease in the absorbance was not observed.These results indicate that the obtained anti-D-Asp mono-clonal antibody raised by the immunization of D-Asp-GA-BSA recognizes the free D-Asp with a high speciWcity and isuseful to the competitive ELISA of D-Asp.
This competitive ELISA of the free D-Asp is also suc-cessfully applicable to biological samples. Fig. 3 shows the
18 ELISA of D-Asp in rat serum / T. Ohgusu et al. / Anal. Biochem. 357 (2006) 15–20
results of the competitive ELISA using the rat serum con-taining the dilution series of D-Asp. The absorbance at405 nm gradually decreased in the range of 0.016–16 �mol/mL D-Asp in rat serum, while the addition of L-Asp andother L- and D-amino acids had no eVect on the absorbance.The precisions of this ELISA for D-Asp in rat serum wereevaluated by the inter-plate and intra-plate variations usinga rat serum containing a known concentration of D-Asp.The relative standard deviation value of the inter-plateassays (Wve wells) was 4.8%, and that of the intra-plateassays (four plates) was 4.5%. To investigate the accuracyof the present ELISA, the amounts of free D-Asp in 12 ratserum samples were determined and the values were com-pared with those obtained by the HPLC method that wepreviously reported [24]. For the HPLC procedure, theserum was deproteinized with MeOH and the amino acids
Fig. 3. Calibration curve of D-Asp in rat serum obtained by the competi-tive ELISA using anti-D-Asp monoclonal antibody.
were derivatized with o-phthalaldehyde and Boc-L-cysteine.The resultant Xuorescent diastereomeric derivatives of theD- and L-Asp were separated on a reversed-phase columnand determined by a Xuorescence detector. The amounts ofD-Asp determined by ELISA and HPLC are almost thesame, and the linear regression line of yD 1.09x + 0.44(where x is the value determined by ELISA and y is thatdetermined by HPLC) with the correlation coeYcient of0.963 was obtained. These results indicate that our estab-lished ELISA method enables the rapid and accurate deter-mination of D-Asp in biological samples. Using thepolyclonal antibody against D-Asp, several studies haveshown the presence and determination of free D-Asp in var-ious species [35–38], and in mammals, this D-amino acid isfrequently observed in the endocrine glands. The presentELISA would be applicable to these tissues by using the tis-sue homogenates in PBS if the tissue wet weight is greaterthan 20 mg.
Time course of D-Asp amounts in the sera of rats after intravenous administration of D-Asp
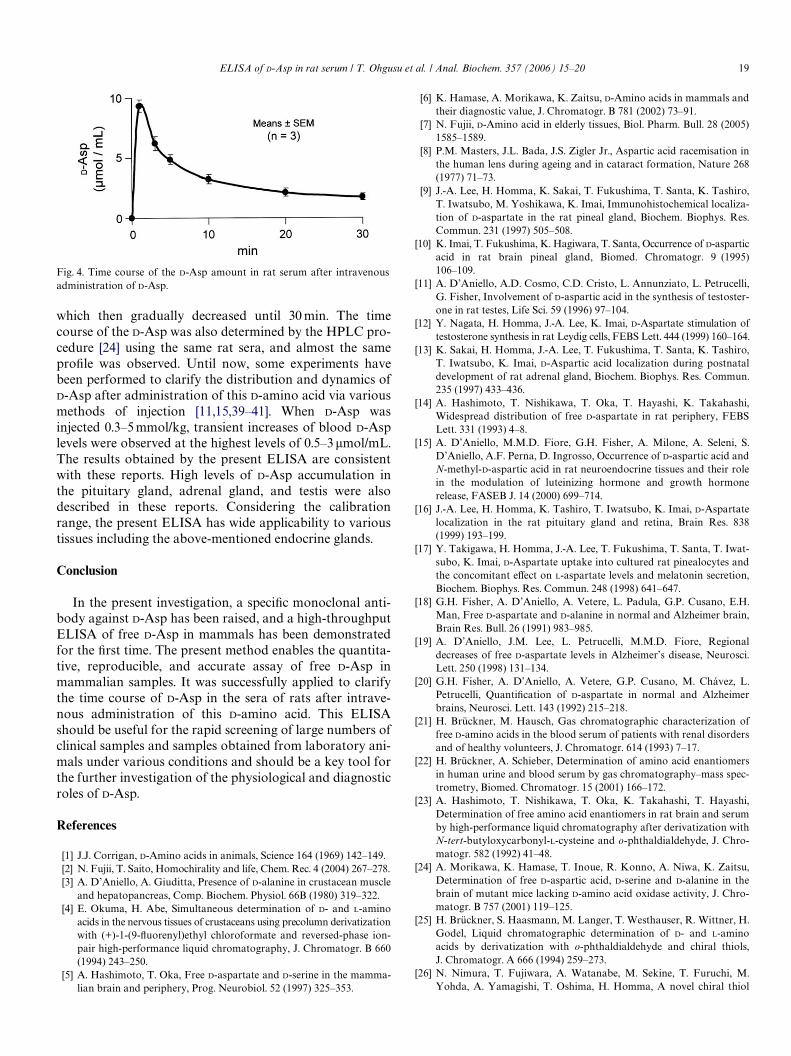
Using the ELISA described above, the time course of theD-Asp amounts in the sera of rats has been investigatedafter the intravenous administration of D-Asp (1 mmol/kgbody weight, dissolved in sodium-phosphate buVer (1 M,pH 7.0)) to Wistar rats (male, 7 weeks of age, SPF). The ratswere anesthetized with pentobarbital and D-Asp wasadministered via the right jugular vein. The blood was col-lected from the left jugular vein 1, 3, 5, 10, 20, and 30 minafter administration and then centrifuged at 5000g for10 min to obtain the serum. Using this serum, the competi-tive ELISA of D-Asp was performed. Fig. 4 shows the timecourse of the D-Asp amounts determined by the competi-tive ELISA. The highest amount of D-Asp was observed(9.3§ 0.52�mol/mL serum) 1 min after the administration,
Fig. 2. Immunoreactivity of anti-D-Asp monoclonal antibody against OVA-conjugated amino acids. Closed bars indicate the immunoreactivity of anti-D-Asp mouse monoclonal antibody, and the open bars indicate those of control mouse IgG.
ELISA of D-Asp in rat serum / T. Ohgusu et al. / Anal. Biochem. 357 (2006) 15–20 19
which then gradually decreased until 30 min. The timecourse of the D-Asp was also determined by the HPLC pro-cedure [24] using the same rat sera, and almost the sameproWle was observed. Until now, some experiments havebeen performed to clarify the distribution and dynamics ofD-Asp after administration of this D-amino acid via variousmethods of injection [11,15,39–41]. When D-Asp wasinjected 0.3–5 mmol/kg, transient increases of blood D-Asplevels were observed at the highest levels of 0.5–3 �mol/mL.The results obtained by the present ELISA are consistentwith these reports. High levels of D-Asp accumulation inthe pituitary gland, adrenal gland, and testis were alsodescribed in these reports. Considering the calibrationrange, the present ELISA has wide applicability to varioustissues including the above-mentioned endocrine glands.
Conclusion
In the present investigation, a speciWc monoclonal anti-body against D-Asp has been raised, and a high-throughputELISA of free D-Asp in mammals has been demonstratedfor the Wrst time. The present method enables the quantita-tive, reproducible, and accurate assay of free D-Asp inmammalian samples. It was successfully applied to clarifythe time course of D-Asp in the sera of rats after intrave-nous administration of this D-amino acid. This ELISAshould be useful for the rapid screening of large numbers ofclinical samples and samples obtained from laboratory ani-mals under various conditions and should be a key tool forthe further investigation of the physiological and diagnosticroles of D-Asp.
References
[1] J.J. Corrigan, D-Amino acids in animals, Science 164 (1969) 142–149.[2] N. Fujii, T. Saito, Homochirality and life, Chem. Rec. 4 (2004) 267–278.[3] A. D’Aniello, A. Giuditta, Presence of D-alanine in crustacean muscle
and hepatopancreas, Comp. Biochem. Physiol. 66B (1980) 319–322.[4] E. Okuma, H. Abe, Simultaneous determination of D- and L-amino
acids in the nervous tissues of crustaceans using precolumn derivatizationwith (+)-1-(9-Xuorenyl)ethyl chloroformate and reversed-phase ion-pair high-performance liquid chromatography, J. Chromatogr. B 660(1994) 243–250.
[5] A. Hashimoto, T. Oka, Free D-aspartate and D-serine in the mamma-lian brain and periphery, Prog. Neurobiol. 52 (1997) 325–353.
Fig. 4. Time course of the D-Asp amount in rat serum after intravenousadministration of D-Asp.
[6] K. Hamase, A. Morikawa, K. Zaitsu, D-Amino acids in mammals andtheir diagnostic value, J. Chromatogr. B 781 (2002) 73–91.
[7] N. Fujii, D-Amino acid in elderly tissues, Biol. Pharm. Bull. 28 (2005)1585–1589.
[8] P.M. Masters, J.L. Bada, J.S. Zigler Jr., Aspartic acid racemisation inthe human lens during ageing and in cataract formation, Nature 268(1977) 71–73.
[9] J.-A. Lee, H. Homma, K. Sakai, T. Fukushima, T. Santa, K. Tashiro,T. Iwatsubo, M. Yoshikawa, K. Imai, Immunohistochemical localiza-tion of D-aspartate in the rat pineal gland, Biochem. Biophys. Res.Commun. 231 (1997) 505–508.
[10] K. Imai, T. Fukushima, K. Hagiwara, T. Santa, Occurrence of D-asparticacid in rat brain pineal gland, Biomed. Chromatogr. 9 (1995)106–109.
[11] A. D’Aniello, A.D. Cosmo, C.D. Cristo, L. Annunziato, L. Petrucelli,G. Fisher, Involvement of D-aspartic acid in the synthesis of testoster-one in rat testes, Life Sci. 59 (1996) 97–104.
[12] Y. Nagata, H. Homma, J.-A. Lee, K. Imai, D-Aspartate stimulation oftestosterone synthesis in rat Leydig cells, FEBS Lett. 444 (1999) 160–164.
[13] K. Sakai, H. Homma, J.-A. Lee, T. Fukushima, T. Santa, K. Tashiro,T. Iwatsubo, K. Imai, D-Aspartic acid localization during postnataldevelopment of rat adrenal gland, Biochem. Biophys. Res. Commun.235 (1997) 433–436.
[14] A. Hashimoto, T. Nishikawa, T. Oka, T. Hayashi, K. Takahashi,Widespread distribution of free D-aspartate in rat periphery, FEBSLett. 331 (1993) 4–8.
[15] A. D’Aniello, M.M.D. Fiore, G.H. Fisher, A. Milone, A. Seleni, S.D’Aniello, A.F. Perna, D. Ingrosso, Occurrence of D-aspartic acid andN-methyl-D-aspartic acid in rat neuroendocrine tissues and their rolein the modulation of luteinizing hormone and growth hormonerelease, FASEB J. 14 (2000) 699–714.
[16] J.-A. Lee, H. Homma, K. Tashiro, T. Iwatsubo, K. Imai, D-Aspartatelocalization in the rat pituitary gland and retina, Brain Res. 838(1999) 193–199.
[17] Y. Takigawa, H. Homma, J.-A. Lee, T. Fukushima, T. Santa, T. Iwat-subo, K. Imai, D-Aspartate uptake into cultured rat pinealocytes andthe concomitant eVect on L-aspartate levels and melatonin secretion,Biochem. Biophys. Res. Commun. 248 (1998) 641–647.
[18] G.H. Fisher, A. D’Aniello, A. Vetere, L. Padula, G.P. Cusano, E.H.Man, Free D-aspartate and D-alanine in normal and Alzheimer brain,Brain Res. Bull. 26 (1991) 983–985.
[19] A. D’Aniello, J.M. Lee, L. Petrucelli, M.M.D. Fiore, Regionaldecreases of free D-aspartate levels in Alzheimer’s disease, Neurosci.Lett. 250 (1998) 131–134.
[20] G.H. Fisher, A. D’Aniello, A. Vetere, G.P. Cusano, M. Chávez, L.Petrucelli, QuantiWcation of D-aspartate in normal and Alzheimerbrains, Neurosci. Lett. 143 (1992) 215–218.
[21] H. Brückner, M. Hausch, Gas chromatographic characterization offree D-amino acids in the blood serum of patients with renal disordersand of healthy volunteers, J. Chromatogr. 614 (1993) 7–17.
[22] H. Brückner, A. Schieber, Determination of amino acid enantiomersin human urine and blood serum by gas chromatography–mass spec-trometry, Biomed. Chromatogr. 15 (2001) 166–172.
[23] A. Hashimoto, T. Nishikawa, T. Oka, K. Takahashi, T. Hayashi,Determination of free amino acid enantiomers in rat brain and serumby high-performance liquid chromatography after derivatization withN-tert-butyloxycarbonyl-L-cysteine and o-phthaldialdehyde, J. Chro-matogr. 582 (1992) 41–48.
[24] A. Morikawa, K. Hamase, T. Inoue, R. Konno, A. Niwa, K. Zaitsu,Determination of free D-aspartic acid, D-serine and D-alanine in thebrain of mutant mice lacking D-amino acid oxidase activity, J. Chro-matogr. B 757 (2001) 119–125.
[25] H. Brückner, S. Haasmann, M. Langer, T. Westhauser, R. Wittner, H.Godel, Liquid chromatographic determination of D- and L-aminoacids by derivatization with o-phthaldialdehyde and chiral thiols,J. Chromatogr. A 666 (1994) 259–273.
[26] N. Nimura, T. Fujiwara, A. Watanabe, M. Sekine, T. Furuchi, M.Yohda, A. Yamagishi, T. Oshima, H. Homma, A novel chiral thiol

20 ELISA of D-Asp in rat serum / T. Ohgusu et al. / Anal. Biochem. 357 (2006) 15–20
reagent for automated precolumn derivatization and high-perfor-mance liquid chromatographic enantioseparation of amino acids andits application to the aspartate racemase assay, Anal. Biochem. 315(2003) 262–269.
[27] K. Hamase, H. Homma, Y. Takigawa, T. Fukushima, T. Santa, K.Imai, Regional distribution and postnatal changes of D-amino acidsin rat brain, Biochim. Biophys. Acta 1334 (1997) 214–222.
[28] Z. Long, N. Nimura, M. Adachi, M. Sekine, T. Hanai, H. Kubo, H.Homma, Determination of D- and L-aspartate in cell culturingmedium, within cells of MPT1 cell line and in rat blood by a column-switching high-performance liquid chromatographic method, J. Chro-matogr. B 761 (2001) 99–106.
[29] A. Neidle, D.S. Dunlop, Developmental changes in free D-asparticacid in the chicken embryo and in the neonatal rat, Life Sci. 46 (1990)1517–1522.
[30] M. Tsunoda, M. Kato, T. Fukushima, T. Santa, H. Homma, H. Yanai,T. Soga, K. Imai, Determination of aspartic acid enantiomers in bio-samples by capillary electrophoresis, Biomed. Chromatogr. 13 (1999)335–339.
[31] S. Zhao, Y. Feng, M.H. LeBlanc, Y.-M. Liu, Determination of freeaspartic acid enantiomers in rat brain by capillary electrophoresiswith laser-induced Xuorescence detection, J. Chromatogr. B 762(2001) 97–101.
[32] J.W. Goding, Antibody production by hybridomas, J. Immunol.Methods 39 (1980) 285–308.
[33] M. Shulman, C.D. Wilde, G. köhler, A better cell line for makinghybridomas secreting speciWc antibodies, Nature 276 (1978) 269–270.
[34] P. Seguela, M. GeVard, R.M. Buijs, M.L. Moal, Antibodies against �-aminobutyric acid: speciWcity studies and immunocytochemicalresults, Proc. Natl. Acad. Sci. USA 81 (1984) 3888–3892.
[35] J.-A. Lee, Z. Long, N. Nimura, T. Iwatsubo, K. Imai, H. Homma,Localization, transport, and uptake of D-aspartate in the ratadrenal and pituitary glands, Arch. Biochem. Biophys. 385 (2001)242–249.
[36] K. Sakai, H. Homma, J.-A. Lee, T. Fukushima, T. Santa, K. Tashiro,T. Iwatsubo, K. Imai, Localization of D-aspartic acid in elongate sper-matids in rat testis, Arch. Biochem. Biophys. 351 (1998) 96–105.
[37] Z. Long, H. Homma, J.-A. Lee, T. Fukushima, T. Santa, T. Iwatsubo,R. Yamada, K. Imai, Biosynthesis of D-aspartate in mammalian cells,FEBS Lett. 434 (1998) 231–235.
[38] J.-A. Lee, H. Homma, K. Sakai, T. Fukushima, T. Santa, K. Tashiro,T. Iwatsubo, M. Yoshikawa, K. Imai, Immunohistochemical localiza-tion of D-aspartate in the rat pineal gland, Biochem. Biophys. Res.Commun. 231 (1997) 505–508.
[39] A. D’Aniello, G. D’Onofrio, M. Pischetola, G. D’Aniello, A. Vetere, L.Petrucelli, G.H. Fisher, Biological role of D-amino acid oxidase and D-aspartate oxidase, J. Biol. Chem. 268 (1993) 26941–26949.
[40] J.-A. Lee, Z. Long, N. Nimura, T. Iwatsubo, K. Imai, H. Homma,Localization, transport, and uptake of D-aspartate in the ratadrenal and pituitary glands, Arch. Biochem. Biophys. 385 (2001)242–249.
[41] R. Boni, R. Santillo, G. Macchia, P. Spinelli, G. Ferrandino, A. D’Ani-ello, D-Aspartate and reproductive activity in sheep, Theriogenology65 (2006) 1265–1278.