growth hormone induced alterations of leptin serum concentrations in dairy cows as measured by a...
TRANSCRIPT

www.elsevier.com/locate/livprodsci
Livestock Production Science 87 (2004) 189–195
Growth hormone induced alterations of leptin serum concentrations
in dairy cows as measured by a novel enzyme immunoassay
H. Sauerweina,*, U. Heintgesa, M. Henniesa, T. Selhorstb, A. Daxenbergerc,1
a Institute of Physiology, Biochemistry and Animal Hygiene, Bonn University, Katzenburgweg 7-9, 53115 Bonn, GermanybFederal Research Center for Virus Diseases, Institute of Epidemiology, Seestrasse 55, 16868 Wusterhausen, Germany
c Institute of Physiology, Technical University Munich, Weihenstephaner Berg 3, 85354 Freising, Germany
Received 6 June 2002; received in revised form 29 July 2003; accepted 7 August 2003
Abstract
The present knowledge on the interrelationship between leptin and the somatotropic axis in livestock, in particular the effects
of growth hormone (GH) treatment on leptin secretion is inconsistent. We therefore aimed to characterise GH-induced
alterations of blood leptin concentrations in dairy cows and first developed and validated a competitive enzyme immunoassay
(EIA). The assay is valid in cattle, sheep, goat, pig and in horses. Lactating cows were injected once with 500 mg depot-
formulated bovine GH and blood samples were drawn every second day 2 weeks before and 4 weeks after the GH injections
and were assayed for leptin and for insulin-like growth factor-1 (IGF-1). Leptin concentrations were decreased in response to
the GH treatment solely in pregnant cows ( p < 0.05), whereas IGF-1 concentrations were increased independently of pregnancy
status. In conclusion, the response of leptin blood concentrations towards GH treatment in lactating cows might depend from
endocrine and metabolic cues related to pregnancy.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Leptin; Enzyme immunoassay; Growth hormone; IGF-1; Ruminant; Pig; Horse
1. Introduction and energy expenditure (Houseknecht and Portocar-
Leptin, the first of several recently discovered
adipocyte proteins, putatively signals the extent of
fat energy reserves to the hypothalamus and is thus
involved in the coordination of the metabolic, endo-
crine and behavioural systems regulating food intake
0301-6226/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/j.livprodsci.2003.08.001
* Corresponding author. Tel.: +49-228-732804; fax: +49-228-
737938.
E-mail address: [email protected] (H. Sauerwein).1 Present address: VITACERT GmbH, Westendstrasse 199,
80686 Munchen, Germany.
rero, 1998; Ahima and Flier, 2000; Vernon et al.,
2001; Ingvartsen and Boisclair, 2001). Besides its
central effects, leptin might also exert peripheral
effects in a number of different tissues, both acting
in an endocrine or a paracrine/autocrine manner, e.g.
in skeletal muscle (Bates et al., 2002), ovary (Spicer,
2001) or adipose tissue itself (Newby et al., 2001).
The functions demonstrated in man and in rodents for
leptin in appetite regulation, lean and fat deposition
and also reproduction are of crucial interest not only
for human medicine but also for animal science. For
dairy as well as for meat production, most of the

H. Sauerwein et al. / Livestock Production Science 87 (2004) 189–195190
research on the endocrine regulatory mechanisms
involved was focused on steroid hormones, somato-
tropic and lactogenic hormones. The exogenous ap-
plication of some of these hormones has evolved a
relatively detailed understanding of their respective
physiological role during the past decades. For live-
stock species, the position of leptin within these
regulative systems could not be investigated properly
until the first species specific assay became available
in 2000 (Delavaud et al., 2000). Since then, the
number of publications dealing with leptin in farm
animals has rapidly increased and several groups have
developed adequate assay systems by now (for review
see Chilliard et al., 2001). Most of the studies on
leptin have focused on the effects of various levels of
energy intake; less is known about the role of leptin
within the hormonal system of the somatotropic axis,
i.e. the growth hormone/insulin-like growth factor
axis. There are several in vivo as well as in vitro
studies on the effects of leptin on growth hormone
(GH) release (e.g. Chen et al., 2001; Henry et al.,
2001; Morrison et al., 2001). Vice versa, studies on
the effects of GH on leptin secretion in farm animals
are presently limited to leptin mRNA quantification in
white adipose tissue (Raymond et al., 1997; House-
knecht et al., 2000). We therefore aimed to further
characterise the hormonal regulation of leptin, in
particular the effects of growth hormone on blood
leptin concentrations in cows of different physiolog-
ical status. A specific enzyme immunoassay was
developed, validated and applied for this purpose.
2. Materials and methods
2.1. Development of a competitive enzyme immuno-
assay (EIA) for leptin in domestic animals
Specific polyclonal antisera against leptin were
raised in five crossbreed rabbits using a mixture of
200 Ag recombinant ovine leptin (roLep; Gertler et
al., 1998) and 100 Ag each of two modified syn-
thetic peptides out of the bovine leptin sequence
(SWISS-PROT protein sequence database; peptide
1: VSSKQRVTGLDFIPGLKY; peptide 2: DLENL-
RDLLHKY; the additional amino acids K and Y
functioning as coupling groups) bound to 200 Agkeyhole limpets hemocyanin (KLH, Sigma-Aldrich,
Taufkirchen, Germany) via the glutardialdehyde re-
action and emulsified in complete Freund’s adjuvant
(Sigma-Aldrich). Nine booster immunisations were
performed with incomplete Freund’s adjuvant (Sig-
ma-Aldrich) in monthly intervals. For the second to
fifth immunisation, half of the amount of antigen
was used and in the last four immunisations 200 Agrecombinant ovine leptin were used omitting the
peptides. The rabbits were bled from an ear vein
1 week after each booster injection and antisera
were stored at � 20 jC.For tracer generation, 40 Ag roLep were biotiny-
lated according to Hennies et al. (2001) using a 100-
fold molar excess of biotinamidocaproate N-hydroxy-
succinimide ester (Sigma-Aldrich).
Microtiter plates (EIA plate 9018, Corning Costar,
Cambridge, MA, USA) were precoated by incubating
150 ng sheep anti-rabbit-Fc fragment antibodies in
100 Al 50 mM sodium hydrogen carbonate, pH 9.6
per well at 4 jC for 20 h. After blocking free binding
sites with 300 Al 2.5% casein in 0.05 M NaCl, pH 7.4
at room temperature for 1.5 h, the plates were washed
five times with 10% PBS, 0.05% Tween 20, filled
with assay buffer and stored at 4 jC up to several
weeks without appreciable loss of sensitivity. The
assay buffer was 0.1% hydrolysed gelatine, 0.12 M
NaCl, 0.02 M Na2HPO4, 0.01 M EDTA, 0.005%
chlorhexidine digluconate (20%), 0.002% phenol red,
200 Al/l proteinase inhibitor cocktail (completek,
Boehringer Mannheim, Germany) and 0.02% ProClin
150R (Supelco, Bellefonte, PA, USA).
For the assay, plates were decanted and leptin
standard or prediluted plasma or serum samples (50
Al) were pipetted in duplicate into the wells. Antise-
rum (50 Al) diluted 1:30,000 with assay buffer con-
taining 2% goose serum were added. All incubation
steps were performed at room temperature. After
preincubation for 16 h, 50 Al of biotinylated leptin
(9 ng/ml assay buffer) were added and incubated for
another 2 h. Bound tracer was quantified via strepta-
vidin-peroxidase as described earlier (Hennies et al.,
2001).
2.2. Radioimmunoassay for IGF-1
Insulin-like growth factor-1 (IGF-1) was deter-
mined by radioimmunoassay (RIA) as described pre-
viously (Daxenberger et al., 1998) using co-incubation

Table 1
Characterisation of the leptin EIA
Antiserum Rabbit anti-ovine leptin
Final dilution 1:90,000
Standard Recombinant ovine leptin
Tracer Biotinylated recombinant leptin
Maximum sample volume 50 AlMeasuring range 0.3–20 ng/ml
50% Binding 2.1 ng/ml
Recovery
1.7 ng/ml 92.5% (n= 4)
5.1 ng/ml 111.7% (n= 4)
14.9 ng/ml 99.9% (n= 4)
CV
Intra-assay 6.3% (n= 12)
Inter-assay 13.9% (n= 41)
H. Sauerwein et al. / Livestock Production Science 87 (2004) 189–195 191
with IGF-2 to minimise potential interferences of IGF
binding proteins. The cross-reactivity to IGF-2 in this
assay was less than 0.01%. The recovery of five
different concentrations IGF-1 added to a sample with
and high and to one with low IGF-1 were 99.2%
(n = 88). Intra-assay and inter-assay coefficients of
variation were 5.1% (n = 32) and 13.4% (n = 64),
respectively.
2.3. Animals, treatments and sample collection
Nineteen lactating Brown Swiss cows received a
500 mg subcutaneous injection of sustained-release
bovine growth hormone (bGH, PosilacR, Monsanto,
USA). Two weeks before and 4 weeks after the
injections, blood samples were collected every second
day. At the time of the bGH injections, the cows were
either not pregnant (98F 47 days postpartum; range:
57 to 202 days, n = 9) or pregnant (158F 47 days
postpartum, range 106–231 days, mean pregnancy
duration was 66 days post insemination; the range was
21 to 147 days; n = 10) and were in their first to sixth
lactation. Pregnancy status was evaluated from the
insemination time and by milk progesterone determi-
nations performed twice per week after the first
insemination. If progesterone remained above 1 ng/
ml throughout the weeks following insemination, the
cows were characterised as pregnant. Retrospectively,
the classification was also confirmed by the calving.
All animals were kept in a cubicle shed, were milked
twice daily in a milking parlour and were fed accord-
ing to their individual production levels. Average milk
yield before the bGH injections was 27.4F 5.9 kg/
day. Blood samples were collected from the udder
vein and plasma was stored at � 20 jC until assayed
for IGF-1 and leptin.
2.4. Statistical analyses
Data were analysed by using the PROC MIXED
model of the SAS 8.01 package. Four covariance
structures (compound symmetry, first order auto-re-
gressive, ante dependence and Toeplitz) were initially
tested and their suitability was assessed by using the
Akaike’s information criterion (AIC). Toeplitz yielded
the lowest AIC values for both the leptin as well as for
the IGF-1 data sets and was therefore applied for all
the subsequent analyses. The independent variables
(i.e. leptin and IGF-1 plasma concentrations) were
log-transformed before statistical analysis. Specific
contrasts were formulated in order to test the hypoth-
eses that (1) there is no effect of GH treatment on
leptin and IGF-1 plasma concentrations, (2) the effect
of the GH treatment on leptin and IGF-1 plasma
concentrations is similar for pregnant and nonpreg-
nant cows, and (3) there is no difference in leptin and
IGF-1 plasma concentrations determined 1 week be-
fore treatment and 2 weeks after GH treatment.
3. Results
3.1. Characterisation and validation of the leptin EIA
Using a combination of recombinant ovine leptin
and peptides out of the bovine leptin sequence,
specific antisera were successfully raised. The antise-
rum with the highest titer against the recombinant
hormone was used to develop the EIA. The criteria of
assay sensitivity, specificity and precision of the
competitive assay developed herein are summarised
in Table 1. The nonspecific binding was less than 5%
of the maximal binding. Accuracy of the assay was
established by ensuring negligible cross-reactivity
( < 0.01%) with other related proteins (ovine prolactin,
ovine placental lactogen and bovine insulin) and an
excellent parallelism between the standard curve and
dilutions from cattle, goat, mufflon, pig and from
horse serum (Fig. 1).
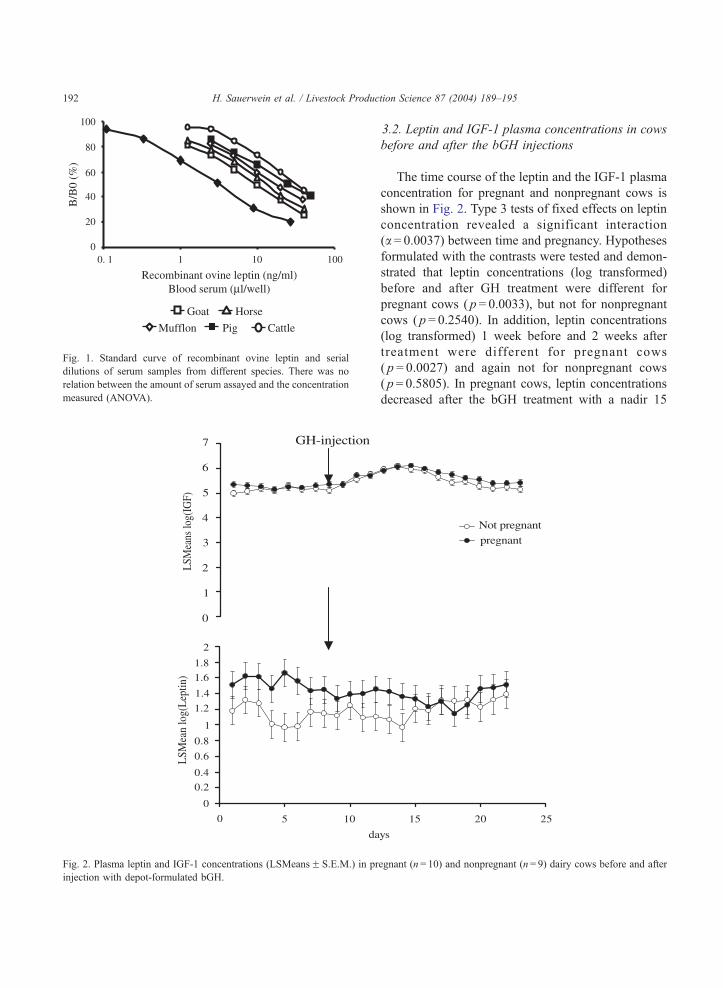
Fig. 1. Standard curve of recombinant ovine leptin and serial
dilutions of serum samples from different species. There was no
relation between the amount of serum assayed and the concentration
measured (ANOVA).
Fig. 2. Plasma leptin and IGF-1 concentrations (LSMeansF S.E.M.) in pr
injection with depot-formulated bGH.
H. Sauerwein et al. / Livestock Production Science 87 (2004) 189–195192
3.2. Leptin and IGF-1 plasma concentrations in cows
before and after the bGH injections
The time course of the leptin and the IGF-1 plasma
concentration for pregnant and nonpregnant cows is
shown in Fig. 2. Type 3 tests of fixed effects on leptin
concentration revealed a significant interaction
(a = 0.0037) between time and pregnancy. Hypotheses
formulated with the contrasts were tested and demon-
strated that leptin concentrations (log transformed)
before and after GH treatment were different for
pregnant cows ( p = 0.0033), but not for nonpregnant
cows ( p = 0.2540). In addition, leptin concentrations
(log transformed) 1 week before and 2 weeks after
treatment were different for pregnant cows
( p = 0.0027) and again not for nonpregnant cows
( p = 0.5805). In pregnant cows, leptin concentrations
decreased after the bGH treatment with a nadir 15
egnant (n= 10) and nonpregnant (n= 9) dairy cows before and after

H. Sauerwein et al. / Livestock Production Science 87 (2004) 189–195 193
days after the injection and returned to pretreatment
levels thereafter. In average, leptin concentrations in
pregnant cows were decreased by 20% after the bGH
injections.
Type 3 tests of fixed effects on the IGF-1 plasma
concentration revealed significant effect of time
( p < 0.0001). bGH treatment lead to a gradual increase
of IGF-1 plasma concentrations with a maximum at
125% of the pretreatment levels during the second
week after the bGH injections and a subsequent
decline to pretreatment levels on day 23 and thereafter
(Fig. 2). In general, Spearman correlations between
plasma IGF-1 and leptin were insignificant. Only
during the pretreatment phase, a weak positive rela-
tionship between leptin and IGF-1 was observed
(r = 0.16, p < 0.05). Milk yield was increased by
bGH treatment by 11% in average.
4. Discussion
The EIA developed herein provides a sensitive and
reliable method to quantify leptin concentrations in
blood serum or plasma from ruminant species. The
concentrations we measured in dairy cows are well in
accordance with leptin values measured by RIA as
reviewed by Chilliard et al. (2001). For the only
enzyme immunological assay published which is
applicable for ruminants (Kauter et al., 2000), no data
for cattle are available. In addition to the advantage of
avoiding the use of radioactive labels, our assay is
valid for a broad range of species, i.e. besides porcine
samples it is also applicable for equine samples and
was already successfully used to characterise peripar-
tal leptin concentrations of breeding mares (Heidler et
al., 2002).
Our experiment in dairy cows provides evidence
that the effect of growth hormone on leptin plasma
concentrations is dependent of the physiological status
of the animal since significant GH-induced changes
were limited to pregnant animals. In contrast to
nonpregnant cows, in which leptin concentrations
remained at a constantly low level throughout the
study, leptin concentrations were decreased after the
GH injections in the pregnant animals. The lack of
reaction in nonpregnant cows cannot entirely be
attributed to lactational stage and thus to differences
in energy balance. Although most of the nonpregnant
cows were at a relatively early stage of lactation, i.e.
days 57 to 89 postpartum (n = 6), there were also three
animals at days 117, 134 and 202. In these animals,
the leptin plasma concentrations were as low as in the
other six cows of the nonpregnant cows and there was
no tendency towards higher concentrations at all.
Although pregnancy was not significantly affecting
leptin plasma concentrations in our study, increased
leptin blood concentrations and leptin mRNA levels in
white adipose tissue have been reported in pregnant
versus nonpregnant ewes (Ehrhardt et al., 2001).
Moreover, the lack of differences in IGF-1 plasma
concentrations between pregnant and nonpregnant
cows suggests that the nutritional status of the animals
was comparable. Based on these observations we
speculate that the leptin response towards GH treat-
ment in pregnant animals may at least in part result
from the specific endocrine conditions of pregnancy
rather than from exclusively nutritional cues. The GH-
induced decrease of leptin plasma concentrations in
pregnant cows we observed fits well into these con-
cepts; however, it is in contradiction to the reports on
GH acting stimulative on leptin mRNA expression in
white adipose tissue from growing cattle (House-
knecht et al., 2000) or growing sheep (Raymond et
al., 1997). The discrepancy between our leptin blood
data and the mRNA expression might be due to the
different physiological status of the animals used: in
contrast to growing animals, we used lactating, mostly
full-grown animals for our experiment. In addition,
concomitant changes within the leptin system (free
leptin, bound leptin, and soluble leptin receptor) might
possibly explain for the discrepancy in leptin response
to GH treatment: Randeva et al. (2002) reported that
GH treatment of GH-deficient humans lead to a fall in
free leptin, a rise in bound leptin and soluble leptin
receptor (sLR). For the RIAs and the ELISAs avail-
able for ruminant leptin, including the assay system
described in the present paper, no information is
available as to whether free and/or bound leptin are
equally recognised. If GH treatment increases both
adipose leptin mRNA expression and sLR, a net
decrease of free leptin may result depending on the
molar ratio of leptin and sLR secreted into the
circulation. In monogastric species, GH treatment
has been demonstrated to either decrease or to have
no effect on (free) leptin blood concentrations (Ran-
deva et al., 2002; Marzullo et al., 2002). However, the

H. Sauerwein et al. / Livestock Production Science 87 (2004) 189–195194
results of these studies might not be directly transfer-
able to healthy organisms since the subjects investi-
gated were affected with complex endocrine disorders,
in particular with GH deficiency.
5. Conclusion
Our results support an inhibiting effect of GH
treatment on leptin plasma concentrations in dairy
cows depending of their endocrine and/or metabolic
status. In view of the discrepancy with reports from the
literature about stimulating effects of GH on adipose
tissue leptin mRNA expression in growing ruminants,
the underlying mechanisms for the decreased leptin
blood concentrations observed remain to be specifi-
cally addressed in the ruminant considering the entire
leptin system and the physiological status of growing,
pregnant or lactating animals.
Acknowledgements
We thank Prof. A. Gertler, Institute of Biochem-
istry, Food Science and Nutrition, Faculty of Agri-
culture, Food and Environmental Quality Sciences,
The Hebrew University of Jerusalem, Rehovot, Israel
for the kind gift of recombinant ovine leptin. The
animal experiment was supported by the Bavarian
Ministry for Nutrition, Agriculture and Forestry
(Bayerisches Staatsministerium fur Ernahrung, Land-
wirtschaft und Forsten). U. Heintges was recipient of
a grant from the Graduiertenforderung des Landes
Nordrhein Westfalen.
References
Ahima, R.S., Flier, J.S., 2000. Leptin. Annu. Rev. Physiol. 62,
413–437.
Bates, S.H., Gardiner, J.V., Jones, R.B., Bloom, S.R., Bailey, C.J.,
2002. Acute stimulation of glucose uptake by leptin in l6 muscle
cells. Horm. Metab. Res. 34, 111–115.
Chen, C., Roh, S.G., Nie, G.Y., Loneragan, K., Xu, R.W., Ruan,
M., Clarke, L.J., Goding, J.W., Gertler, A., 2001. The in vitro
effect of leptin on growth hormone secretion from primary cul-
tured ovine somatotrophs. Endocrine 14, 73–78.
Chilliard, Y., Bonnet, M., Delavaud, C., Faulconnier, Y., Leroux,
C., Djiane, J., Bocquier, F., 2001. Leptin in ruminants. Gene
expression in adipose tissue and mammary gland, and regu-
lation of plasma concentration. Domest. Anim. Endocrinol.
21, 271–295.
Daxenberger, A., Breier, B.H., Sauerwein, H., 1998. Increased milk
levels of insulin-like growth factor 1 (IGF-1) for the identifica-
tion of bovine somatotropin (bST) treated cows. Analyst 123,
2429–2435.
Delavaud, C., Bocquier, F., Chilliard, Y., Keisler, D.H., Gertler, A.,
Kann, G., 2000. Plasma leptin determination in ruminants: effect
of nutritional status and body fatness on plasma leptin concen-
tration assessed by a specific RIA in sheep. J. Endocrinol. 165,
519–526.
Ehrhardt, R.A., Slepetis, R.M., Bell, A.W., Boisclair, Y.R., 2001.
Maternal leptin is elevated during pregnancy in sheep. Domest.
Anim. Endocrinol. 21, 85–96.
Gertler, A., Simmons, J., Keisler, D.H., 1998. Large-scale prepa-
ration of biologically active recombinant ovine obese protein
(leptin). FEBS Lett. 422, 137–140.
Heidler, B., Sauerwein, H., Aurich, J., Pohl, W., Aurich, C., 2002.
Metabolic profiles and plasma leptin concentrations in lactating
and non-lactating mares. Theriogenology 58, 557–561.
Hennies, M., Voglmayr, J.K., Dietrich, E., Stollmann, M., Moeller,
R., Holtz, W., 2001. Hormonal response of female goats to
active immunization against a recombinant human inhibin al-
pha-subunit, and establishment of an enzyme-linked immuno-
sorbent assay for caprine follicle-stimulating hormone. Reprod.
Domest. Anim. 36, 65–71.
Henry, B.A., Goding, J.W., Tilbrook, A.J., Dunshea, F.R., Clarke,
I.J., 2001. Intracerebroventricular infusion of leptin elevates the
secretion of luteinising hormone without affecting food intake in
long-term food-restricted sheep, but increases growth hormone
irrespective of bodyweight. J. Endocrinol. 168, 67–77.
Houseknecht, K.L., Portocarrero, C.P., 1998. Leptin and its recep-
tors: regulators of whole-body energy homeostasis. Domest.
Anim. Endocrinol. 15, 457–475.
Houseknecht, K.L., Portocarrero, C.P., Ji, S., Lemenager, R., Spur-
lock, M.E., 2000. Growth hormone regulates leptin gene expres-
sion in bovine adipose tissue: correlation with adipose IGF-1
expression. J. Endocrinol. 164, 51–57.
Ingvartsen, K.L., Boisclair, Y.R., 2001. Leptin and the regulation of
food intake, energy homeostasis and immunity with special fo-
cus on periparturient ruminants. Domest. Anim. Endocrinol. 21,
215–250.
Kauter, K., Ball, M., Kearney, P., Tellam, R., McFarlane, J.R.,
2000. Adrenaline, insulin and glucagon do not have acute ef-
fects on plasma leptin levels in sheep: development and char-
acterisation of an ovine leptin ELISA. J. Endocrinol. 166,
127–135.
Marzullo, P., Buckway, C., Pratt, K.L., Colao, A., Guevara-Aguirre,
J., Rosenfeld, R.G., 2002. Leptin concentrations in GH defi-
ciency: the effect of GH insensitivity. J. Clin. Endocrinol.
Metab. 87, 540–545.
Morrison, C.D., Daniel, J.A., Holmberg, B.J., Djiane, J., Raver, N.,
Gertler, A., Keisler, D.H., 2001. Central infusion of leptin into
well-fed and undernourished ewe lambs: effects on feed intake
and serum concentrations of growth hormone and luteinizing
hormone. J. Endocrinol. 168, 317–324.

H. Sauerwein et al. / Livestock Production Science 87 (2004) 189–195 195
Newby, D., Gertler, A., Vernon, R.G., 2001. Effects of recombi-
nant ovine leptin on in vitro lipolysis and lipogenesis in sub-
cutaneous adipose tissue from lactating and nonlactating sheep.
J. Anim. Sci. 79, 445–452.
Randeva, H.S., Murray, R.D., Lewandowski, K.C., O’Callaghan,
C.J., Horn, R., O’Hare, P., Brabant, G., Hillhouse, E.W., Shalet,
S.M., 2002. Differential effects of GH replacement on the com-
ponents of the leptin system in GH-deficient individuals. J. Clin.
Endocrinol. Metab. 87, 798–804.
Raymond, S.R., Thomas, M.G., Caroll, J.A., Matteri, R.L., Keisler,
D.H., 1997. Zeranol and growth hormone treatment differen-
tially influenced mRNA levels of the obesity protein, leptin,
and the GH receptor in growth wethers. J. Anim. Sci. 75
(Suppl. 1), 225.
Spicer, L.J., 2001. Leptin: a possible metabolic signal affecting
reproduction. Domest. Anim. Endocrinol. 21, 251–270.
Vernon, R.G., Denis, R.G., Sørensen, A., 2001. Signals of adipo-
sity. Domest. Anim. Endocrinol. 21, 197–214.