gm polymorphism and the evolutionary history of modern humans
TRANSCRIPT

Original article
GM polymorphism and the evolutionary history of modern humansHassen Chaabani *
Faculté de Pharmacie de Monastir, Université du Centre, 5000 Monastir, Tunisia
Received 5 October 2001; accepted 8 November 2002
Abstract
Present human populations show a complex network of genetic relationships, which reflects mainly their unique origin and their migrationand isolation history since the recent creation of modern man. The scrutiny of their genetic characteristics, according to GM polymorphism,shows that the continuity of the genetic variation between populations from neighbouring continents, assured by intermediate world partpopulations, is against any attempt to divide present human populations into major groups. GM polymorphism analysis also shows threeremarkable levels of genetic differentiation, which would have appeared, respectively, within populations of sub-Saharan Africa, Europe andEast Asia. The first small groups of people that split from the common ancestral population gave the sub-Saharan Africans. On the otherhand, Asians diverged mainly from Europeans and Near East populations during a later period. The confrontation between the phylogenyand the frequency distribution of GM haplotypes shows that the ancestral population of actual South-Arabia people could be a candidatefor a common ancestral population. The first major expansions of modern humans were proposed in a hypothetical scenario, which will opena new track in the research of our geographic origin. © 2002 Editions scientifiques et médicales Elsevier SAS. All rights reserved.
Keywords: Evolutionary relationships of human populations; GM and Cγ polymorphisms
1. Introduction
Using genetic data, many studies were carried out toanalyse genetic variations within and between populations.They have shown a small amount of genetic variationthroughout all present human populations, which stronglysupports the theory of the single recent origin of all of us(e.g., Refs. [15,19]). In other studies, genetic data areanalysed in an attempt to reconstruct the evolutionaryhistory of human populations. In these studies, researchershave followed one of two principal approaches. Some ofthem have analysed nuclear non-DNA and/or DNA poly-morphisms using at once a large number of loci [4,6,31,32].On the other hand, others have analysed particular geneticmaterial as that of maternal (mt DNA) (e.g., Refs. [5,23]) orpaternal (Y chromosome) (e.g., Ref. [20]) lineages or that ofa single nuclear gene group (e.g., Refs. [24,47,48]).
The immunoglobulin GM system is known by its unpar-alleled ability, of a single system, to differentiate humanpopulations. Here, using this single nuclear gene group in a
new approach, I have studied the evolutionary relationshipsof human populations.
2. Materials and methods
In this study, I have chosen representative populationsfrom old continents without neglecting those living inintermediate parts (Table 1). These populations were studiedfor the GM polymorphism in terms of statistically correctgene frequency estimates.
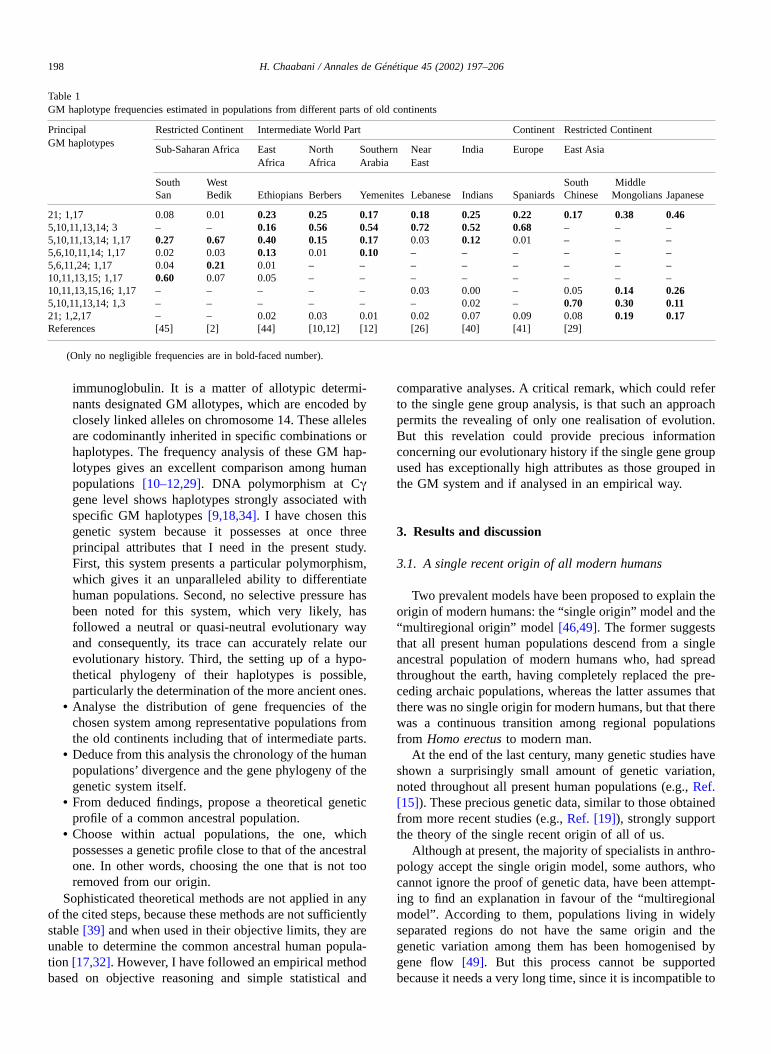
For reinforcing statistically the analysis of each GMhaplotype distribution, many populations of each of thedifferent parts of old continents are taken into considerationin charts of GM haplotype distribution (Fig. 1, Appendix A).
Successive steps of the approach followed in this studycould be summarised as follows:
• Accept the theory of the single and recent origin of allof us and limit its meaning.
• Choose a nuclear gene group in which high exceptionalperformances of anthropological interest were grouped.The immunoglobulin GM system is the nuclear genegroup chosen. It represents the polymorphism of γ1, γ2and γ3 heavy chain constant (C) regions of human
* Corresponding author.E-mail address: [email protected] (H. Chaabani).
Annales de Génétique 45 (2002) 197–206
www.elsevier.com/locate/angen
© 2002 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.PII: S 0 0 0 3 - 3 9 9 5 ( 0 2 ) 0 1 1 4 0 - 1
immunoglobulin. It is a matter of allotypic determi-nants designated GM allotypes, which are encoded byclosely linked alleles on chromosome 14. These allelesare codominantly inherited in specific combinations orhaplotypes. The frequency analysis of these GM hap-lotypes gives an excellent comparison among humanpopulations [10–12,29]. DNA polymorphism at Cγgene level shows haplotypes strongly associated withspecific GM haplotypes [9,18,34]. I have chosen thisgenetic system because it possesses at once threeprincipal attributes that I need in the present study.First, this system presents a particular polymorphism,which gives it an unparalleled ability to differentiatehuman populations. Second, no selective pressure hasbeen noted for this system, which very likely, hasfollowed a neutral or quasi-neutral evolutionary wayand consequently, its trace can accurately relate ourevolutionary history. Third, the setting up of a hypo-thetical phylogeny of their haplotypes is possible,particularly the determination of the more ancient ones.
• Analyse the distribution of gene frequencies of thechosen system among representative populations fromthe old continents including that of intermediate parts.
• Deduce from this analysis the chronology of the humanpopulations’ divergence and the gene phylogeny of thegenetic system itself.
• From deduced findings, propose a theoretical geneticprofile of a common ancestral population.
• Choose within actual populations, the one, whichpossesses a genetic profile close to that of the ancestralone. In other words, choosing the one that is not tooremoved from our origin.
Sophisticated theoretical methods are not applied in anyof the cited steps, because these methods are not sufficientlystable [39] and when used in their objective limits, they areunable to determine the common ancestral human popula-tion [17,32]. However, I have followed an empirical methodbased on objective reasoning and simple statistical and
comparative analyses. A critical remark, which could referto the single gene group analysis, is that such an approachpermits the revealing of only one realisation of evolution.But this revelation could provide precious informationconcerning our evolutionary history if the single gene groupused has exceptionally high attributes as those grouped inthe GM system and if analysed in an empirical way.
3. Results and discussion
3.1. A single recent origin of all modern humans
Two prevalent models have been proposed to explain theorigin of modern humans: the “single origin” model and the“multiregional origin” model [46,49]. The former suggeststhat all present human populations descend from a singleancestral population of modern humans who, had spreadthroughout the earth, having completely replaced the pre-ceding archaic populations, whereas the latter assumes thatthere was no single origin for modern humans, but that therewas a continuous transition among regional populationsfrom Homo erectus to modern man.
At the end of the last century, many genetic studies haveshown a surprisingly small amount of genetic variation,noted throughout all present human populations (e.g., Ref.[15]). These precious genetic data, similar to those obtainedfrom more recent studies (e.g., Ref. [19]), strongly supportthe theory of the single recent origin of all of us.
Although at present, the majority of specialists in anthro-pology accept the single origin model, some authors, whocannot ignore the proof of genetic data, have been attempt-ing to find an explanation in favour of the “multiregionalmodel”. According to them, populations living in widelyseparated regions do not have the same origin and thegenetic variation among them has been homogenised bygene flow [49]. But this process cannot be supportedbecause it needs a very long time, since it is incompatible to
Table 1GM haplotype frequencies estimated in populations from different parts of old continents
PrincipalGM haplotypes
Restricted Continent Intermediate World Part Continent Restricted Continent
Sub-Saharan Africa EastAfrica
NorthAfrica
SouthernArabia
NearEast
India Europe East Asia
South West South MiddleSan Bedik Ethiopians Berbers Yemenites Lebanese Indians Spaniards Chinese Mongolians Japanese
21; 1,17 0.08 0.01 0.23 0.25 0.17 0.18 0.25 0.22 0.17 0.38 0.465,10,11,13,14; 3 – – 0.16 0.56 0.54 0.72 0.52 0.68 – – –5,10,11,13,14; 1,17 0.27 0.67 0.40 0.15 0.17 0.03 0.12 0.01 – – –5,6,10,11,14; 1,17 0.02 0.03 0.13 0.01 0.10 – – – – – –5,6,11,24; 1,17 0.04 0.21 0.01 – – – – – – – –10,11,13,15; 1,17 0.60 0.07 0.05 – – – – – – – –10,11,13,15,16; 1,17 – – – – – 0.03 0.00 – 0.05 0.14 0.265,10,11,13,14; 1,3 – – – – – – 0.02 – 0.70 0.30 0.1121; 1,2,17 – – 0.02 0.03 0.01 0.02 0.07 0.09 0.08 0.19 0.17References [45] [2] [44] [10,12] [12] [26] [40] [41] [29]
(Only no negligible frequencies are in bold-faced number).
198 H. Chaabani / Annales de Génétique 45 (2002) 197–206
the dates proposed for the emergence of the modern man.Indeed, calculations of a population geneticist [36] showthat it would require almost half a million years for anadvantageous gene to travel from South Africa to China bythe normal process of gene flow.
In addition to its evident agreement with genetic data, thesingle recent origin model is in accordance with linguisticdata. In fact, modern linguists put forward the view that all
actual languages have very likely originated from a uniqueand recent ancestral one [37,42,43].
To avoid all confusions and vagueness over the model ofsingle recent origin, two remarks must be made. First, thismodel, supported by genetic and linguistic data relating topresent human populations, concerns only modern man,Homo sapiens sapiens, without involving previous Homo asthe archaic Homo sapiens who could be diverged from
Fig. 1. Charts of GM haplotypes distribution in different parts of the old continents. Estimations are based on detection percentages of each GM haplotypein frequency thresholds of 0.05 (in white) and 0.20 (in black). These percentages are been calculated from statistically correct data that reported in theliterature in 47 population studies (see Appendix A): seven populations from restricted sub-Saharan Africa “r. s-S. Af.”, 18 populations from intermediateword parts (including three from middle East Africa “m. E. Af.”, five from North Africa “N. Af.”, five from Arabia and Near East “A.& N. E.” and five fromIndia “Ind.”). Seven from Europe “Eur.” 14 from restricted East Asia (including six from South East Asia “S. E. As.” and nine from middle East Asia “m.E. As.”).
H. Chaabani / Annales de Génétique 45 (2002) 197–206 199

Homo erectus in many regions as well as in a single one.Second, I think that it is improper to link this model with thechoice of sub-Saharan Africans as the population group oforigin, because the single origin model is a general modeland the population group of origin could be another one[11].
3.2. Genetic relationships of present human populations
The GM haplotype distribution estimated in populationsfrom different parts of old continents is presented in Table 1and Fig. 1. I have restricted the geographic part of two of thethree old continents: Asia to East Asia and Africa tosub-Saharan Africa with the exclusion of the extreme eastmiddle Africa. In spite of this restriction, one can note thatpopulations belonging to each of these continents do notpresent quite a genetically homogeneous group. In fact, acontinuous genetic variation appears slightly through neigh-bouring populations and becomes relatively important be-tween distant ones as the San and the Bedik in sub-SaharanAfrica or as Japanese and Chinese in East Asia (Table 1). Inaddition, the population group of each of these continentsbecomes more or less heterogeneous if one tries to arrangethe intermediate world part populations inside it, as the caseof North Africans and sub-Saharan Africans (Table 1).These observations support the idea that geographic dis-tance is a major factor behind population differentiation [7].
Indeed, intermediate world part populations, by theirintermediate geographical situation and particular geneticprofiles, assure the continuity of the genetic variationbetween populations in neighbouring continents (Table 1).This is against any attempt to divide present human popu-lations into major groups. However, in the great part ofpopulations belonging to the same restricted continent,some GM haplotypes are absent or present in slight frequen-cies. But the complete absence of GM haplotypes is ratherhypothetical, as it is difficult to detect GM haplotypes invery slight frequencies. Therefore, it seems reasonable tosuggest that, within populations from the same restrictedcontinent, some GM haplotypes tend to disappear to thedetriment of the increasing frequency of others, while thecomplete absence of these haplotypes could be reached in atheoretical population. As can be seen, within populations ofeach group of the three restricted continents (sub-SaharanAfrica, Europe and East Asia), a genetic profile partiallydistinct from the other two could be reached in a theoreticalpopulation. Continuing in the theoretical vein, these popu-lations do not represent genetically differentiated entitiesbut they correspond to remarkable differentiation stages,which occurred successively during the human evolutionaryhistory.
The chronological order of these stages could be deducedby the following simple reasoning: the GM haplotypedistribution (Table 1, Fig. 1) shows that GM 21,28; 1,17 isfound in common between Asian populations and those ofNear East and Europe. But no GM haplotypes are found, in
no negligible frequency, in common between these popula-tions and those of the restricted sub-Saharan Africa. Thisallows one to suggest that during human evolution ancestralgroups of actual sub-Saharan Africans diverged first from acommon ancestral population, while Near East and Euro-pean populations, on the one hand, and Asians, on the other,diverged from each other at a later date. This obvioussuggestion, deduced from the use of a single preciousgenetic system, is not at variance with those deduced instudies where theoretical methods were used for analysingeither a single gene group [48] or, at once, several non-DNAand/or DNA polymorphisms [6,31]. In fact, these studiessupport a primary division of human populations intosub-Saharan Africans and Eurasian populations, and asecond, which separates Europeans and Asians. But as permy suggestion, I specify that Asians are separated from bothEuropean and Near East populations. This comes as a resultof the fact that the populations of all parts of the oldcontinents were considered in my analysis. In addition, Ispecify that ancestral groups of actual sub-Saharan Africansdiverged first from a common ancestral population, becauseduring the first divergence, the Eurasian population had notyet developed.
In a relatively more recent study [32], using the chim-panzee as an out-group and by employing proper statisticalmethods, Nei and Takezaki constructed phylogenetic treesof human populations for five different sets of gene fre-quency data. Phylogenetic trees thus obtained showed thatsub-Saharan Africans were the first group of people thatsplit from other human populations. In addition, theyshowed that the root is located in the branch connectingsub-Saharan Africans and the rest of the world populationsintroduced in the analyses. From correct construction ofphylogenetic trees, even if the root is located, it is notpossible to determine the common ancestral population.Thus, here I have tried to determine the common ancestralpopulation otherwise.
3.3. Genetic profile of the common ancestral population
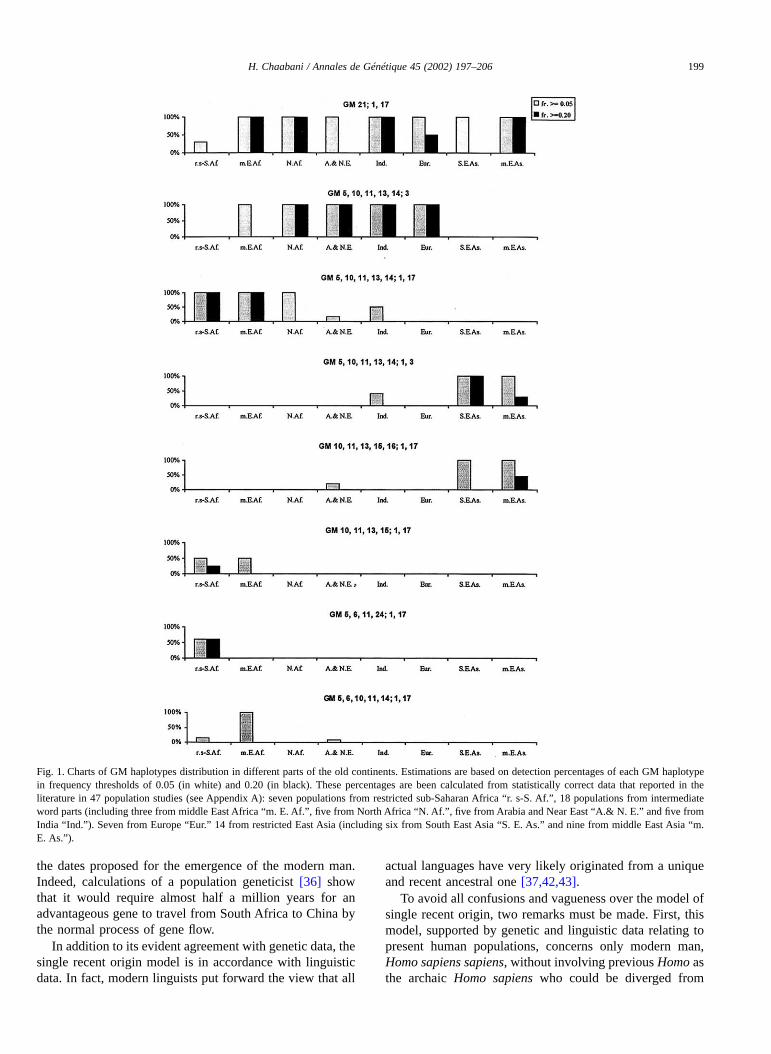
As stated above, it is now accepted that the origin ofmodern humans is recent. Thus, it is reasonable to assumethat the genetic pool of the actual population, whichpresents the continuity of the common ancestral one, muststill possess the principal original genes. I mean by originalgenes, for a given polymorphism, all those have existedsince the development of the common ancestral population.Thus, such original genes are now the most widespread inthe world; the more ancient are mainly those which cannotbe derived from one another by simple molecular evolution-ary mechanisms. Starting from this consideration and usingagain the unique GM system, it is possible to propose atheoretical genetic profile of the common ancestral popula-tion.
From the GM haplotype distribution, presented in Table 1and in Fig. 1, one can note that the GM 21,28; 1,17 is the
200 H. Chaabani / Annales de Génétique 45 (2002) 197–206
most widespread among populations in almost all geo-graphical parts. So it could be considered as an ancientoriginal haplotype. GM 21,28; 1,2,17, very similar to thisancient original haplotype, is likewise disseminated butfound often in negligible frequencies, and therefore could beconsidered as derived later from it by mutation (Fig. 3).Among the remaining haplotypes, GM 5,10,11,13,14; 1,17and GM 5,10,11,13,14; 3 are relatively the most wide-spread. But contrary to GM 5,10,11,13,14; 1,17, GM5,10,11,13,14; 3 is often present in high frequency. So itcould be considered as more ancient and present since the
beginning with GM 21,28; 1,17 in the common ancestralpopulation. This agrees with the fact that GM 21,28; 1,17and GM 5,10,11,13,14; 3 could not have been derived fromone another, whereas, all other GM haplotypes can derivefrom them (Figs. 2 and 3). This suggestion is supported byresults relative to Cγ DNA haplotype studies [1,8,9]. Asshown in Table 2, H1 and H2 Cγ DNA haplotypes areevidently more frequent in the two populations studied andthese Cγ DNA haplotypes are in about 90% of the casesassociated to GM 21,28; 1,17 or GM 5,10,11,13,14; 3. Inaddition, like these two ancient GM haplotypes, the H1 and
Fig. 2. First major expansions of modern humans presented as a hypothetical scenario to be tested in the future. Successive stages of first major migrationsare indicated: I, II, III, and IV. The phylogeny of principal GM haplotypes from the most ancient ones (GM 5 *; 3 and GM 21,28; 1,17) is presented.(*: 10, 11, 13, 14, M: mutation, R: recombination, C: gene conversion).
H. Chaabani / Annales de Génétique 45 (2002) 197–206 201

H2 Cγ DNA haplotypes could not have derived from oneanother. However, all other Cγ DNA haplotypes can derivefrom them.
As I have inferred above, during human evolutionancestral groups of sub-Saharan Africans diverged first froma common ancestral population. In addition, GM5,10,11,13,14; 1,17 is generally the most common haplo-type in sub-Saharan Africans (Table 1). From these twoconsiderations, it seems reasonable to assume that the GM5,10,11,13,14; 1,17 haplotype appeared in the ancestralpopulation before the divergence of sub-Saharan Africans.Thus, in the common ancestral population three GM hap-lotypes are present: GM 21,28; 1,17, GM 5,10,11,13,14; 3and GM 5,10,11,13,14; 1,17. Considering that the latter isformed by a possible recombination of the two more ancientones, its frequency in the common ancestral population
Table 2Cγ DNA haplotype frequencies in Tunisans and North American Cauca-sians
Designation Cγ HaplotypesFragment combinations(sizes in kb)
FrequencyNorthAmericans
Tunisians
N = 58 N = 39
H1 13.5, 12.5, 11.8, 9.4, 8.8 0.43 0.27 ] *H2 25, 12.5, 11.8, 10, 9 0.47 0.40H3 13.5, 12.5, 11.8, 9.0, 8.8 0.02 0.05H4 25, 12.5, 11.8, 10, 9.4 0.09 0.01H5 13.5, 12.5, 11.8, 10, 9 0 0.19H6 13.5, 12.5, 11.8, 10, 9.4 0 0.05References [1] [9]
* In 90% of cases H1 and H2 are associated to GM haplotypes: 21; 1,17 or 5, 10, 11, 13, 14; 3.
Fig. 3. Genetic profile of the hypothetical ancestral population compared with that of actual Yemeni population.
202 H. Chaabani / Annales de Génétique 45 (2002) 197–206

must be non-negligible but not evidently more importantthan that of each one of them (Fig. 3).
On the basis of this theoretical genetic profile thusestablished for a common ancestral population, one canchoose within actual populations the one, which possesses agenetic profile close to it. In other words, choosing the onethat is not too removed from the place of our origin. Asshown in Table 1, the genetic profile of European, MiddleEast and extreme East Africa populations is somewhat akinto that proposed for the common ancestral population witha difference in the frequency of GM 5,10,11,13,14; 1,17. Infact, in European and Middle East populations, this haplo-type is absent or present in negligible frequency, while inpopulations of extreme East Africa, as that of Ethiopia, it ismuch more important than that of the two more ancienthaplotypes. However, the genetic profile of North AfricanBerbers and particularly that of South-Arabian peninsula(Yemen) population tally entirely with that proposed for thecommon ancestral one (Tables 1 and 3). Thus, one of thesetwo population groups could present the continuity of thecommon ancestral population. Prehistoric data, more than10,000 year old, consider that the first inhabitants of NorthAfrica, the ancestors of Berbers, came from Yemen [11,30].Therefore, Yemen could be a probable cradle of the com-mon ancestral population development. This is in agreementwith data obtained in our recent study where, in addition tothe GM system, Rhesus system is also used. An analysis ofthe results of this study shows that present Yemenites wouldmostly be the direct descendants of an original populationnative to Yemen. They have an intermediate genetic com-position including the most widespread and shared Rhesusand GM haplotypes among world populations. This com-position would be close to that of a highly diversifiedancestral population [12].
In spite of the proposition of Southern Arabia as aprobable place of the common ancestral population devel-opment being still preliminary, it could motivate research onour geographic origin which has been relatively stagnantsince the proposition of Cann et al. [5] in 1987. Theseauthors have constructed human mitochondrial DNA typephylogenies and concluded that our origin is from sub-Saharan Africa. Although the biological material and meth-ods used in this study were severely criticised[14,17,22,28,38], some authors, using other DNA polymor-phisms but following the same questionable considerations,have supported this proposition [e.g. Refs. 21,23,47]. Here,with the GM polymorphism, I would also support this thesisif I follow these questionable considerations that can besummarised into three principal ones. First, it is the lack ofpopulation representativeness [11,12,39]. Unfortunately,populations living in regions located somewhere betweenprincipal parts of old continents and having possibly inter-mediate genetic profiles are the most ignored, either due tolack of adequate genetic data and/or considering them as theproducts of gene admixture [6,31] even though, thesepopulations could be original ones as the case of Yemeni
people [12]. Second, the possible establishment of an unrealroot of phylogenetical reconstruction particularly when oneapplies the parsimony analysis: many models of tree con-struction can be used which might lead to contradictoryconclusions [17,22,28]. Third, many authors consider thatpopulation group, genetically the most divergent from therest of the world populations, as that of sub-Saharan Africa,is the first established and the most likely to remain in theplace of origin. Therefore, this population group would bethe original one. But this consideration is not categoricaland seems incompatible with the recent event of theemergence of modern man. Dissenting from this view,Langaney and his collaborators have put forward a morereasonable one. According to them, sub-Saharan Africacould not be the place of our origin. This is due to the factthat the peculiarity of the genetic profile of sub-SaharanAfricans could be the result of their divergence from anancestral population if both intensive selection and geneticdrift occurred after a drastic founder effect and a relativelylong isolation [11,16,25,39]. In addition, as the GM haplo-type distribution shows (Table 1, Fig. 1) and as it has beennoted for other genetic markers [16], populations from therestricted sub-Saharan Africa were devoid of importantgenetic heritage present in the other world populations.Thus, it is impossible to consider their genetic pool as thatof a recent common ancestral population.
Finally, it is interesting to note that my conclusionsrelating to the evolutionary relationships of human popula-tions are not at variance with that of earlier significantfindings. However, they bring out important new precisedetails. On the other hand, the ticklish subject of ourgeographic origin is not yet resolved. I show here question-able considerations that have led to the proposition ofsub-Saharan Africa as a probable place of the commonancestral population development and present a secondproposition that of Southern Arabia. I have advanced thispreliminary proposition, not for dismissing the first but toopen a new track in the research of our geographic origin.However, my proposition must be verified using otherpolymorphisms particularly DNA ones and avoiding thequestionable considerations stated above. Planning of suchstudies is being worked upon.
3.4. Hypothetical scenario of modern human expansions
In Fig. 2, I have finally depicted a scenario of theexpansion of modern man across the earth since his recentcreation. This scenario is largely a speculation based onconclusions from GM polymorphism analysis. According tothese conclusions, ancestral groups of sub-Saharan Africansdiverged first from a common ancestral population, whichemerged probably in Southern Arabia (Yemen). So in thefirst stage, modern man had chosen, probably under climaticinfluence, to migrate from Yemen to the south going throughthe sound of Bab al-Mandab to extreme East Africa. Thissmall geographic distance between Yemenites and their
H. Chaabani / Annales de Génétique 45 (2002) 197–206 203

neighbouring sub-Saharan Africans, as Ethiopians, corre-sponds to a relatively important variation of GM haplotypefrequencies (Table 1, Fig. 1) estimated in our previous studyby a very important genetic distance [12]. This reflects a lowcorrelation between genetic and geographic distances in thisarea and suggests that the divergence between the originalpopulation of Yemeni people and that of sub-SaharanAfricans occurred by migrations of small groups with afounder effect and a long isolation. From the east middleAfrica, some humans had proceeded to the west and duringtheir expansion, a new haplotype appeared, the GM,5,6,11,24; 1,17, which occurs, particularly in the presentWest Africans. While others had proceeded to the south, andfollowing their expansion, the GM 10,11,13,15; 1,17 ap-peared and consequently has become frequent in actualSouth Africans (Table 1, Figs. 1 and 2).
In our previous study [12] comparing Yemenites withother populations, we show that the genetic distance be-tween Yemenites and Indians is relatively small. So, in thesecond stage, migrations could have been from South-Arabian peninsula toward the east to reach India andprobably to southern East Asia by preliminary expansions.This is not at variance with recent basic results of mito-chondrial DNA study, which shows that the mitochondrialhaplogroup M, first regarded as an ancient marker ofEast-Asian origin, is found in the South-Arabian peninsula,India and Ethiopia [35]. Simultaneously, migrations wouldhave occurred from the north of Yemen up to the Near Eastand, during the third step, from the Near East to Europe andfrom Yemen to northern Africa. In fact, genetic distancesbetween Yemenites and populations from the Near East,Europe and North Africa often vary harmoniously with thecorresponding geographic distances and show, therefore, ahigh correlation amongst themselves [12]. So it is verylikely that these migrations (II and III stages) occurred inrelatively large groups without any serious climatic obstacle
at a period in which the present Saharan regions had anotheraspect [7,11,33]. The remarkable genetic rapprochementbetween Yemenites and North African Berbers suggests thatthey were not subject to a strong genetic drift after theirgeographic differentiation; other less important migrationsfrom Yemen to North Africa have probably followed thefirst one at more recent periods.
As I have inferred, the divergence of East Asians wasmainly from Europeans and neighbouring Middle Eastpopulations during a relatively recent period correspondingin my scenario to stage IV. The presence and the distributionof two GM haplotypes common only in East Asians showthat the expansion to East Asia could have occurred in twomajor directions. One of these expansions to central Asia ismarked by the appearance of the GM 10,11,13,15,16; 1,17present in corresponding populations, the second to thesouth is marked by the appearance of the GM 5,10,11,13,14;1,3 that is frequent in the present Southeast Asians (Table 1,Figs. 1 and 2). From basic results of mitochondrial DNA,similar migrations were proposed. According to these re-sults, radiation through Central Asia, carried among others,the prominent A and B haplogroups, led to its expansion toEast Asia [27]. In the present scenario, no major humanexpansion is proposed either from North Africa to southernEurope or in the opposite direction. This agrees with resultanalysis of Alu insertion polymorphisms [13] and that ofhuman Y-chromosome variation [3], which show a sharpdiscontinuity and limited gene flow between north-westernAfrica and the Iberian Peninsula.
Acknowledgements
The author is grateful to Professor Ferjani Khemais forhis careful reading of the manuscript.
204 H. Chaabani / Annales de Génétique 45 (2002) 197–206

Appendix A
List of population samples used in the estimation of detection percentages of each GM haplotype presented in charts inFig. 1
References
[1] N.T. Bech-Hansen, P.S. Linsley, D.W. Cox, Restriction fragmentlength polymorphisms associated with immunoglobulin Cγ genesreveal linkage disequilibrium and genomic organisation, Proc. Natl.Acad. Sci. USA 80 (1983) 6952–6956.
[2] M. Blanc, A. Sanchez-Mazas, N.H. Van Blyenburgh, A. Sevin,G. Pison, A. Langaney, Interethnic genetic differentiation: GMpolymorphism in Eastern Senegal, Am. J. Hum. Genet. 46 (1990)383–392.
[3] E. Bosch, F. Calafell, D. Comas, P.J. Oefner, P.A. Underhill,J. Bertranpetit, High-resolution analysis of human Y-chromosomevariation shows a sharp discontinuity and limited gene flow betweenNorth-western Africa and the Iberian Peninsula, Am. J. Hum. Genet.68 (2001) 1019–1029.
[4] A.M. Bowcock, A. Ruiz-linares, J. Tomfohrde, E. Minch, J.R. Kidd,L.L. Cavalli-Sforza, High resolution of human evolutionary treeswith polymorphic microsatellites, Nature 368 (1994) 455–457.
[5] R.L. Cann, M. Stoneking, A.C. Wilson, Mitochondrial DNA andhuman evolution, Nature 325 (1987) 31–36.
Population Location Reference
Restricted sub-Saharan Africa Bedik Senegal Blanc M. (1990)Fulani Senegal Blanc M. (1990)Mandenka Senegal Blanc M. (1990)Nigerians Nigeria van Loghem E. (1978)San ! Kung South Africa Steinberg A.G. (1975)Khoikhoi South Africa Steinberg A.G. (1975)Bantu South Africa Jenkins T. (1970)
Intermediate world parts East Middle Africa Sidamo Ethiopia Steinberg A.G. (1973)Amhara Ethiopia Fort M. (1997)
North Africa Berbers Tunisia (Chenini) Chaabani H. (1984)Tunisians Tunisia Helal A.N. (1981)Berbers Tunisia (Gallala) Chaabani H. (1988)Tunisians Tunisia (Sfax) Lefranc G. (1979)Tunisians Tunisia (Mehdia) Lefranc G. (1979)
Arabia and Near East Yemenites Southern Arabia Chaabani H. (2000)Iraqis Iraq (Baghdad) Shakib F. (1980)Lebanese Lebanon (Beirut) Lefranc G. (1976)Irani Iran van Loghem E. (1977)Kurds Lebanon, Syria Taleb N. (1973)
India Indian Brahmanes India Schanfield M.S. (1981)Indian Konda Reddy India van Loghem E. (1985)Indian Koya Dora India van Loghem E. (1985)Indian Nalcker India Schanfield M.S. (1981)Indian Hindu India Schanfield M.S. (1981)
Europe Spaniards Spain Schanfield M.S. (1981)Dutch Holland Fraser G.R. (1974)French France Rivat L. (1978)Croatians Croatia Barot N. (1991)Italians Italy Piazza A. (1973)Romanians Romania Johnson W.E. (1977)Finns Finland Steinberg A.G. (1974)
Restricted East Asia Middle East Asia Mongols Mongolia Matsumoto H. 1988Mongols WulanhoubuMongols HuhehoteTibetans Lasa, TibetTibetans W. TibetJapanese MainlandJapanese Naha, OkinawaOlunchums N.E. ChinaKoreans Korea
South East Asia Han (Chinese) S. China (Chengdu)Han (Chinese) S. China (Fuzhou)Takasagos TaiwanPhilippinos LuzonIndonesians JavaMelanesians Bougainville
H. Chaabani / Annales de Génétique 45 (2002) 197–206 205

[6] L.L. Cavalli-Sforza, A. Piazza, P. Menozzi, J. Mountain, Recon-struction of human evolution: bringing together genetic archaeologi-cal and linguistic data, Proc. Natl. Acad. Sci. USA 85 (1988)6002–6006.
[7] L.L. Cavalli-Sforza, P. Menozzi, A. Piazza, The history and geog-raphy of human genes, Princeton University Press, Princeton, NJ,1994.
[8] H. Chaabani, N.T. Bech-Hansen, D.W. Cox, A multigene deletionwithin the immunoglobulin heavy chain region, Am. J. Hum. Genet.37 (1985) 1164–1171.
[9] H. Chaabani, N.T. Bech-Hansen, D.W. Cox, Restriction fragmentlength polymorphisms associated with immunoglobulin heavy chaingamma genes in Tunisians, Hum. Genet. 73 (1986) 110–113.
[10] H. Chaabani, D.W. Cox, Genetic characterisation and origin ofTunisian Berbers, Hum. Hered. 38 (1988) 308–316.
[11] H. Chaabani, The emergence of modern man, Imprimerie Officiellede la République Tunisienne, Tunisia, 1999.
[12] H. Chaabani, A. Sanchez-Mazas, S.F. Sallami, Genetic differentia-tion of Yemeni people according to Rhesus and GM polymorphisms,Ann. Génét. 43 (2000) 155–162.
[13] D. Comas, F. Calafell, N. Benchemsi, A. Helal, G.G. Lefranc,M. Stoneking, M.A. Batser, J. Bertranpetit, A. Sajantila, Aluinsertion polymorphisms in NW Africa and the Iberian Peninsula:evidence for a strong genetic boundary through the Gibraltar Straits,Hum. Genet. 107 (2000) 312–319.
[14] P. Darlu, P. Tassy, Disputed African origin of human populations,Nature 329 (1987) 111.
[15] M. Denaro, H. Blanc, M.J. Johnson, C.H. Chen, E. Wilmsen,L.L. Cavalli-Sforza, D.C. Wallace, Ethnic variation in Hpa Iendonuclease cleavage patterns of human mitochondrial DNA, Proc.Natl. Acad. Sci. USA 78 (1981) 5768–5772.
[16] L. Excoffier, B. Pellegrini, A. Sanchez-Mazas, C. Simon, A. Langa-ney, Genetics and History of sub-Saharan Africa, Yearbook ofPhysical Anthropology 30 (1987) 151–194.
[17] L. Excoffier, A. Langaney, Origin and differentiation of humanmitochondrial DNA, Am. J. Hum. Genet. 44 (1989) 73–85.
[18] N. Ghanem, J.M. Dugoujon, M. Bensmana, S. Huck, M.P. Lefranc,G. Lefranc, Restriction fragment haplotypes in the human immuno-globulin IGHG locus and their correlation with the GM polymor-phism, Eur. J. Immunol. 18 (1988) 1067–1072.
[19] M.F. Hammer, A recent common ancestry for human Y chromo-somes, Nature 378 (1995) 376–378.
[20] M.F. Hammer, S.L. Zegura, The role of the Y chromosome in humanevolutionary studies, Evol. Anthropol. 5 (1996) 116–134.
[21] M.F. Hammer, T. Karafet, A. Rasanayagam, E.T. Wood, T.K. Alth-eide, T. Jenkins, R.C. Griffiths, A.R. Templeton, S.L. Zegura, Out ofAfrica and back again: nested cladistic analysis of human Ychromosome variation, Mol. Biol. Evol. 15 (1998) 427–441.
[22] S.B. Hedges, S. Kumar, K. Tamura, M. Stoneking, Human originsand analysis of mitochondrial DNA sequences, Science 255 (1992)737–739.
[23] M. Ingman, H. Kaessmann, S. Paabo, U. Gyllensten, Mitochondrialgenome variation and the origin of modern humans, Nature 408(2000) 652–653.
[24] D. Labuda, E. Zietkiewicz, V. Yotova, Archaic lineages in the historyof modern humans, Genetics 156 (2000) 799–808.
[25] A. Langaney, La nouvelle démographie de l’évolution, Population39 (1984) 587–606.
[26] G. Lefranc, J. Loiselet, L. Rivat, C. Ropartz, Gm, Km and Isfallotypes in the Lebanese population, Acta Anthropogenetica 1(1976) 34–45.
[27] N. Maca-Meyer, A.M. Gonzalez, J.L. Larruga, C. Flores, V. MCabrera, Major genomic mitochondrial lineages delineate earlyhuman expansions, BMC Genetics 2 (2001) 13–21.
[28] D.R. Maddison, M. Ruvolo, D.L. Swofford, Geographic origins ofhuman mitochondrial DNA: phylogenetic evidence from controlregion sequences, Systematic Biol. 41 (1992) 111–124.
[29] H. Matsumoto, Characteristics of Mongoloid and neighbouringpopulation based on the genetic markers of human immunoglobu-lins, Hum. Genet. 80 (1988) 207–218.
[30] R. Nadhouri, The Arabic Maghreb (the ancient eras), ArabicDawning Company for Writing and Publishing, Beyrouth, 1981.
[31] M. Nei, A.K. Roychoudhury, Evolutionary relationships of humanpopulations on a global scale, Mol. Biol. Evol. 10 (1993)927–943.
[32] M. Nei, N. Takezaki, The root of the phylogenetic tree of humanpopulations, Mol. Biol. Evol. 13 (1996) 170–177.
[33] A. Pons, J.L. de Beaulieu, J. Guiot, M. Reille, Le pollen remonte letemps climatique, La Recherche 213 (1991) 518–520.
[34] D.N. Propert, R. Nandan, E.C. Jazwinska, GM and KM allotypesand GM RFLP allogenotypes in Micronesians from Nauru, Hum.Biol. 64 (1992) 17–24.
[35] L. Quintana-Murci, O. Semino, H.J. Bandelt, G. Passarino,K. McElna, A.S. Santachiara-Benerecetti, Genetic evidence of anearly exit of Homo sapiens sapiens from Africa through easternAfrica, Nat. Genet. 4 (1999) 437–441.
[36] S. Rouhani, Molecular genetics and the pattern of human evolution,in: P. Mellars, C.B. Stringer (Eds.), The Human Revolution, Uni-versity Press Princeton, 1989, pp. 47–61.
[37] M. Ruhlen, Toutes parentes, toutes différentes, La Recherche 306(1998) 68–75.
[38] N. Saitou, K. Omoto, Time and place of human origins from mtDNA data, Nature 327 (1987) 288.
[39] A. Sanchez–Mazas, A. Langaney, Common genetic pools betweenhuman populations, Hum. Genet. 78 (1988) 161–166.
[40] M.S. Schanfield, R.L. Kirk, Further studies on the immunoglobulinallotypes (GM Am and Km) in India, Acta Anthropogenet. 5 (1981)21–24.
[41] M.S. Schanfield, R. Baylerian, J. Maiquez, F. Carbonell, Immuno-globulin allotypes in European populations. IV. Gm Am and Kmallotypic markers in Valencia Spain, J. Immunogenet. 8 (1981)529–532.
[42] V. Shevoroshkin, Reconstructing Languages and Cultures, Bochum,Brockmeyer, 1989.
[43] V. Shevoroshkin, The mother tongue, Sciences 3 (1990) 20–27.[44] A.G. Steinberg, GM and Inv allotypes of some Sidamo Ethiopians,
Am. J. Phys. Anthrop. 39 (1973) 403–408.[45] A.G. Steinberg, T. Jenkins, G.T. Nurse, H.C. Harpending, Gamma-
globulin groups of the khoisan peoples of southern Africa: evidencefor polymorphism for a GM 1,5,13,14,21 haplotype among the San,Am. J. Hum. Genet. 27 (1975) 528–542.
[46] C.B. Stringer, P. Andrews, Genetic and fossil evidence for the originof modern humans, Science 239 (1988) 1263–1268.
[47] S.A. Tishkoff, E. Dietzsch, W. Speed, A.J. Pakstis, J.R. Kidd,K. Cheung, B. Bonné-Tamir, A.S. Santachiara-Benerecetti, P. Moral,M. Krings, S. Paabo, E. Watson, N. Risch, T. Jenkins, K.K. Kidd,Global patters of linkage disequilibrium at the CD4 locus andmodern human origins, Science 271 (1996) 1380–1387.
[48] J.S. Wainscoat, A.V.S. Hill, A.L. Boyce, J. Flint, M. Hernandez,S.L. Thein, J.M. Old, J.R. Lynch, A.G. Falusi, D.J. Weatherall,J.B. Clegg, Evolutionary relationships of human populations froman analysis of nuclear DNA polymorphisms, Nature 319 (1986)491–493.
[49] M.H. Wolpoff, Multiregional evolution: the fossil alternative toEden, in: P. Mellars, C.B. Stringer (Eds.), The Human Revolution-Behavioural and Biological Perspective on the Origin of ModernHumans, Princeton University Press, 1989, pp. 62–108.
206 H. Chaabani / Annales de Génétique 45 (2002) 197–206