geo alp - science south tyrol,,natur,21,404.pdfgeo alp - science south tyrol
TRANSCRIPT


Geo Alp A new yearly journal devoted to
· Alpine geology
Neue Jahreszeitschrift zur AI pe·ngeolog ie
La nuova rivista per Ia Geologia delle AI pi

Geo.Aip
Redaktionskomitee: Rainer Brandner, lnnsbruck, Diethard Sanders, lnnsbruck, Volkmar Mair, Bozen, Benno Baumgarten,
Naturmuseum Bozen
Technische Redaktion/Layout: Monika Tessadri-Wackerle
Herausgeber, EigentUmer und Verleger:
lnstitut fur Geologie und PaHiontologie, Universitat lnnsbruck, Naturmuseum Bozen
Chefredakteur Geo.Aip 2: Karl Krainer
Referentlnnen dieser Nummer:
M. Breda, Padova; H. Kerp, Munster; T. Kotsakis, Roma; S.G. Lucas, Albuquerque; D. Nagel, Vienna; Chr. Rupp, Vienna
B. Sala, Ferrara; R. Sardella, Roma; G. Tichy, Salzburg
Erscheinungsweise und Bezug:
Geo.Aip erscheint einmal jahrlich und kann bei beiden herausgebenden lnstitutionen im Abonnement oder einzeln bezogen werden :
lnstitut fUr Geologie und Palaontologie, lnnrain 52, A-6020 lnnsbruck, Austria
Naturmuseum SUdtiroi/Museo Scienze Naturali Alto Adige, Bindergasse/via Bottai 1, 1-39100 Bozen/Bolzano, Italy
© lnstitut fUr Geologie and jlalaontologie, Universitat lnnsbruck; Naturmuseum SUdtiroi/Museo Scienze Naturali Alto Adige
Genehmigung des Landesgerichts Bozen Nr. 12/2004 vim 05/11/2004
Verantwortli~;:her Direktor: Dr. Vito lingerie
ISSN 1824-7741
Umschlagbild: Monika Tessadri-Wackerle, verwendete Abbildung von Evely Kustatscher
Druck: Walser Druck KG

F
Geo.Aip
In halt
Herbert Scholz, Karl-Heinz Bestle & Sebastian Willerich: Ouartargeologische Untersuchungen im Oberetsch
Beitrage zu ,Giornate della Paleontologia der Societa Paleontologica ltaliana 2004", 20-23. Mai 2004:
Raffaele Sardella, Claudia Bedetti, Luca Bellucci, Nicoletta Conti, Danilo Coppola, Emmanuele Di Canzio, Marco Pavia, Carmela Petronio, Mauro Petrucci & Leonardo Salari: The Late Pleistocene vertebrate fauna from Avetrana (Taranto, Apulia, Southern Italy) : preliminary report............. 25
Evelyn Kustatscher & Johanna H.A. van Konijnenburg-van Cittert: The Ladinian Flora (Middle Triassic) of the Dolomites: palaeoenvironmental reconstructions and palaeoclimatic considerations ........................................ 31
Cristiana lata & lassos Kotsakis: Italian fossil chiropteran assemblages: a preliminary report ...................................... 53
Gabriella Mangano: Cervus elaphus siciliae from Pleistocene lacustrine deposits of Acquedolci (North-Eastern Sicily, Italy) and its taphonomic significance......................................................................... 61
Gabriella Mangano, Laura Bonfiglio & Daria Petruso: Excavations of 2003 at the S. Teodoro Cave (north-eastern Sicily, Italy): preliminary faunistic and stratigraphic data 71
Giuseppe Santi: Lower Permian paleoichnology from the Oroboc basin (northern Italy) ..................................... ...... ..... 77
Maria Teresa Curcio, Longino Contoli, Emanuele Di Canzio & lassos Kotsakis: Preliminary analysis of the first lower molar variability in Late Pleistocene and living populations of Terri cola savii (Arvicolidae, Rodentia) ........... 91
Davide Mana: A test application of the SHE method as a biostratigraphical parameter ................ .......... ....... .... .... ....... . 99
Cinzia Galli, Mario Rossi & Giuseppe Santi: Ursus spe/aeus Rosen muller, 1794 from the Venetian region of Northern Italy: Preliminary notes on its evolutionary path .................... .. ........................................... ................................. 107
Alessandro de Carlis, Enrico Alluvione, Alessandro Fonte, Mario Rossi & Giuseppe Santi: Morphometry of the Ursus spelaeus remains from Valstrona (Northern Italy) .................................................. ............. ........................ ......... 115
Abstracts zu ,Giornate della Paleontologia der Societa Paleontologica ltaliana 2004", 20-23. Mai 2004:
Francesco Garofalo, Fabrizio Bizzarini & Federica Ferrieri: The activities of the Ligabue Study Research Centre on the thirtieth anniversary of its foundation ................................................... .. ..... ........ ....................... .. ... ................................. 127
Nicola Daii'Oiio : The origin of the palaeontological fossil concept ................................................................. ........................ 131

..
INSTRUCTIONS TO AUTHORS
Articles may be submitted in English, German or Italian. In case of a German or Italian text, the captions to all figures, plates and tables must be also in English, and an English abridged version (1000-1500 words) and abstract are to be delivered.
Articles shall be submitted in th~e copies to:
Karl Krainer, Diethard Sanders, Institute of Geology and Palaeontology, University of lnnsbruck, lnnrain 52, A-6020 lnnsbruck, Austria. E-mail: [email protected]; [email protected] or to: Benno Baumgarten, Naturmuseum Si.ldtiroi/Museo Scienze Naturali Alto Adige, Bindergasse 1Nia Bottai 1, 1-39100 Bozen/Bolzano, Italy: E-mail: [email protected]
Articles must be typed double-space. The quality of line-drawings must be ready for print. In line-drawings and figures of any sort, all labellings, numbers and letters should be readable upon 50% reduction in size.
Photographs and line-drawings can be submitted in original hardcopy, or in an electronic format. All photographs must be clearly labelled on the backside.
For photographic tables: please note that the page setup of Geo.Aip is 23.47 x 16,70 em (8 em column width).
The final text (Word file) must be submitted on CD or Zip disk. The name of the author, the name and version of the word processing program(s) and type of computer on which the text was prepared must be indicated. To avoid obsolete passages in the manuscript please note that Word files should be saved after using the option "Accept Changes" in the Tools Menu.
Word tables must be on single pages for transformation into pdf format or already in pdf format.
The text should be submitted in single column format, and should be formatted as simple as possible (e.g. no bullets and no automatic numbering). Electronic versions of figures and/or photographs must be submitted as separate files (file format: tif (high -resolution jpg files are also possible), photographs with a resolution of 300 dpi (due to print size), line drawings at least 600 dpi; colour images all in CMYK mode.
Referencing:
Articles: Author 1, X. Y., Author 2, Z. A. (2002): Title of article. - International journal abbreviation (e.g. Sediment. Geol.), vol. : . pp-pp.
Articles in books: Author 1, X. Y., Author 2, Z. A. (2002): Title of article.- In: Person A, Person B. (eds.): Title of book, pp-pp, publisher, place of publication.
Books: Author 1, X. Y., Author 2, Z. A. (2002): Title of book. - no. of pages, publisher, place of publication.
Reprints: 50 reprints are free of charge

Geo.Alp, Vol. 2, S. 1–23, 2005
QQUUAARRTTAARRGGEEOOLLOOGGIISSCCHHEE UUNNTTEERRSSUUCCHHUUNNGGEENN IIMM ÜÜBBEERREETTSSCCHH
Herbert Scholz, Karl-Heinz Bestle & Sebastian Willerich
Mit 8 Abbildungen und 1 FototafelWith 8 figures and 1 plate
ZZuussaammmmeennffaassssuunnggIn der weiten Talung von Eppan-Kaltern im Südtiroler Überetsch bei Bozen ist ein ganzes System von kie-
sigen Lateralmoränen, breiten Kamesterrassen sowie in die mächtige, komplex aufgebaute pleistozäne Tal -füllung erosiv eingeschnittenen, kastenförmigen Schmelzwasserrinnen erkennbar, mit deren Hilfe sich unter -schiedliche Eisstände einer „Kalterer Zunge“ aus dem ausgehenden Hochglazial rekonstruieren lassen, einerTeil zunge des Etschgletschers. An den Osthängen des Mendel-Roèn-Kammes sind zudem zertalte Erosionsre-ste von Murfächern nachweisbar, hier als „Murkames“ bezeichnet, die direkt gegen den absinkendenWestrand dieses Gletschers geschüttet wurden. Sehr kalk- und dolomitreiche Mursedimente, durch Eisauflastüberkonsolidiert und damit vermutlich älter als der letzte Eishöchststand, aber auch Ablagerungen vondeutlich jüngeren Muren, die aus dem Spät- und Postglazial stammen dürften, bedecken große Flächen ander Ostflanke des Mendel-Roèn-Kammes bis ins Tal hinunter, besonders im Gebiet zwischen Margreid, Penonund Kurtatsch. Gerade auf diesen von den Einheimischen „Kampferde“ genannten diamiktischen Ablagerun-gen liegt ein Großteil der Weinberge rund um Tramin und Kurtatsch.
SSuummmmaarryyWithin the wide vale of Eppan-Kaltern (Appiano-Caldaro) at Überetsch (Oltradige, Sella di Appiano-Cal-
daro) close to Bozen (Bolzano) in South Tyrol (Alto Adige) a complicated system of gravelly lateral moraines,large kame terraces as well as erosive fossil meltwater valleys can be identified, which are deeply incised intoa thick and complex sequence of Pleistocene sediments. Most of these structures are remnants of the„Kaltern lobe“, a late Pleniglacial tongue of the Etsch (Adige) valley glacier. Moreover erosional remnants ofdebris fans can be identified at the eastern slopes of Mendola-Roèn-Ridge, obviously deposited in the gapbetween the mountain slope and the western rim of this recessing glacier (“fankame“). The eastern slopes ofthe Mendola-Roèn-Ridge, especially the region between Margreid (Magrè all’Adige), Penon (Penone) andKurtatsch (Cortaccia), are widely covered with debris flow deposits which are extremely rich in limestone anddolomite fragments. They are partly older and partly younger than the “fankame“. Some are obviously over-consolidated by the extra load of glacier ice and therefore presumably older than the Last Glacial Maximum,some are obviously younger and may have a Late to Postglacial age. Many vineyards around Kurtatsch (Cor-taccia) and Tramin (Termeno) are situated right on the surface of these diamictic deposits, called“Kampferde” by the local people.
1
Herbert Scholz, Karl-Heinz Bestle und Sebastian Willerich,Lehrstuhl für Ingenieurgeologie der Technischen Universität München,Arcisstr. 21, D-80290 München; e-mail: [email protected]

EEiinnlleeiittuunngg
Die hier vorgestellten Ergebnisse wurden im Rah-men dreier Kartierungsübungen mit fortge -schrittenen Geologiestudenten der TU Münchensowie bei eigenen Geländebegehungen im Ge bietzwischen Oberplanitzing und Margreid in Südtirolerarbeitet. Diese von Prof. Dr. Herbert Scholz be-treuten Kartierungsübungen waren vom 31. Märzbis zum 11. April 2002, vom 24. März bis zum 4.April 2003 und vom 23. März bis zum 3. April 2004durchgeführt worden und hatten vor allem das Ziel,die quartären Sedimente im Überetsch zu erkunden,genetisch zu klas sifizieren sowie zeitlich zu ordnen.Alle Geländeübungen wurden seitens des Amtes fürMaterial prüfung und Geologie in Bozen (Dr. Volk-mar Mair), des Südtiroler Amtes für Gewässerschutzund der Gemeinde sowie der freiwilligen Feuerwehrvon Kurtatsch (Bürgermeister Oswald Schiefer,Kommandant Albert Terzer) unterstützt.An den drei Kartierungsübungen haben folgende
Studenten bzw. Studentinnen teilgenommen: W.Bäumel, I. Baumann, K.-H. Bestle, A. Dargel, M. Döh-ner, M. Elsner, Chr. Gampe, G. Ghon, R. Hohlfeld, J.Kadlcakova, F. Köppl, M. Lammel, F. Meyer, Chr.Minet, Chr. Mögele, I. Mon sorno, S. Suckfüll, I. Thie-le, B. Weiher, Chr. Weber, K. Wendl und S. Willerich. Eingehendere Untersuchungen zur Quartärgeo-
logie des Gemeidegebietes von Kurtatsch wurdenim Rahmen zweier Diplomarbeiten von Karl-HeinzBestle und Sebastian Willerich am Lehr stuhl für In-genieurgeologie der TU München angestellt (Bestle2005 und Willerich 2005). Diese Diplomarbeitenwurden von Prof. Dr. Herbert Scholz im Gebiet zwi-schen Tramin, Graun, Fennberg und Margreid verge-ben und betreut. Sie wurden vom Amt für Geologieund Bau stoffkunde in Kardaun sowie von der Ge-meinde Kurtatsch unterstützt. Allen, die wissen -schaft lich, logistisch oder finanziell zum Gelingendieser Untersuchungen beigetragen haben, sei andieser Stelle herzlich gedankt.
11.. GGeeoollooggiisscchheerr AAuuffbbaauu ddeess ÜÜbbeerreettsscchh
11..11 DDiiee GGeesstteeiinnee ddeess UUnntteerrggrruunnddeess iimm ÜÜbbeerrbblliicckk
Die westliche Talflanke der Etschtalfurche süd-westlich von Bozen, das Gebiet von Überetsch unddem Mendelkamm, wird vor allem von permotriassi-schen Gesteinen aufgebaut. Die Schicht folge, die
mit dem unterpermischen Bozener Quarzporphyr-komplex beginnt, fällt generell leicht nach SW ein.Die Mächtigkeit des Bozener Quarzporphyrs dürfteweit über 1000 m betragen, denn das Gebiet befin-det sich noch innerhalb der permischen „Caldera vonBozen“ (Bosselini 1998: 82), in der besonders mäch-tige Ignimbritfolgen akkumuliert wurden. Über denpermischen Vulkaniten folgt der terrestrische, mit 40bis 70 m vergleichsweise geringmächtige GrödnerSandstein (Perm), eine teilweise kohleführende, buntgefärbte Folge von Sandsteinen und Tonschluff -steinen (Brandner & Mostler 1982, Gwinner 1971). Die Gröden Formation geht zum Hangenden in
die flach-marine Werfen Formation (Skyth) über. Dieoberpermische Bellerophon Formation, die schonwenige Kilometer östlich der Etsch weit verbreitet ist(Heissel 1982: 22, 28), fehlt hier hingegen völlig. Diehier aufgeschlossene, regel mäßig gebankte tonig-schluffige Schichtfolge der Werfe n Formation ent-hält zahlreiche feste Bän ke aus Schluffsteinen,Sandsteinen, Dolomiten, Mergelkalken, Kalken undOolithen. Sie lässt sich nicht ohne weiteres mit demWerfener Standard-Profil im Schlern gebiet oder inder nur wenige Kilometer entfernten Bletterbach-schlucht parallelisieren (vergl. Brandner & Mostler1982, Moser 1996). Das Unteranis wird durch denUnteren Sarldolomit und stellenweise durch bren-nend rote Sandsteine (evtl. Äqui valente des Voltago-Konglomerates) repräsentiert, die sich ohne scharfeGrenze aus der unterla gernden Wer fen Formationentwickeln. Diese bunten Sandsteine werden voneiner grau gefärb ten, kalkig-mergeligen Schichtfol-ge überlagert, bei denen es sich um Äquivalente deroberanisi schen Mor biac-Kalke handeln dürfte. Diesegehen zum Hangenden hin in den Contrindolomitüber, dolo mitischen und gebankten gelblich anwit-ternden Flachwasserkarbonaten des Oberanis. An den steilen Schluchthängen, die vom Mendel-
kamm zum Etschtal hinunterziehen, sind im mer wie-der grobblockige Konglomerate mit sandig-tonigem,rotem Bindemittel, rötliche Sandsteine und Toneaufgeschlossen. Diese konglomeratischen, teilweiseriesige Blöcke enthaltenden Sedi mente stellen of-fensichtlich Füllungen klammartiger fossiler Erosi-onsrinnen dar, die mehrere Dekameter tief in dieSchichtfolge der Werfen Formation eingeschnittensind. Sie werden sicher vom Contrindolomit, teilwei-se wohl auch von den Morbiac-Kalken überlagertund sind z.B. an der Forststraße im Höllental auf -geschlossen, die von Graun nach Söll führt. DieseKonglomerate enthalten vor allem gelblich gefärbte
2 Geo.Alp, Vol. 2, 2005

Komponenten der Werfen Formation, daneben aberauch kleine Geröllchen aus hell grauem Dolomit. Obes sich um Äquivalente des Richthofen- oder desVoltago-Konglomerates handelt, ist unbekannt. Dieoben genannten Konglo me rat vor kom men füllen imHöllental fossile Erosionsrinnen auf, die klammartigbis in den Unteren Sarldolomit ein geschnit ten sind.An der Anis-Ladin-Grenze entwickeln sich im
Hangenden des Contrindolomits zwischen Margreidund Tramin die kalkig-mergeligen, teilweise auchsandigen „Zwischenschichten“, eine bitu minöseBeckenfazies mit Tuffiteinschaltungen. In dieseSchichtfolge sind Dolomite, gebankte Kalksteineund chaotisch gelagerte Brekzien aus Flachwasser-kalken (Olisto strome) eingeschaltet, denn die „Zwi-schenschichten“ verzahnen sich nach S hin offen -sichtlich mit den Flachwas serablagerungen des ba-salen Schlerndolomits. Sie haben sich in einem klei-nen, aber vermutlich recht tiefen, im Norden durchStörungen begrenzten Becken gebildet, im sogen.„Kurtatscher Loch“. Der Mendelkamm selbst wirdvon mächtigem Schlerndolomit (Ladin) und Haupt-dolomit (Karn/Nor) aufgebaut, der im Norden direktdem Contrindolomit, im S auch den „Zwischen -schichten“ auflagert. Südlich von Margreid, wo diegesamte Mittel- und Obertrias in ähnlicher dolomi-tischer Fazies entwickelt ist, lässt sich die Dolomit-folge nur schwer unterglie dern und wird als „Men-deldolomit“ bzw. „Etschtaldolomit“ zusammenge-fasst (Geyer 1993). An der Straße von Penon nach Fennberg und in
Fennberg selbst werden die Dolomite von ge -ringmächtigen, teilweise bunt gefärbten pelagi-schen Kalken überlagert, die schon dem Jura undder Oberkreide angehören.
11..22 TTeekkttoonniikk iimm ÜÜbbeerrbblliicckk
Zwischen Bozen und Neumarkt folgt das Etschtalvermutlich einer N-S-orientierten Störungs zone, ander die östliche Talflanke gegenüber der westlichendeutlich herausgehoben zu sein scheint. Die Aufla-gerungsfläche des Grödner Sandsteins auf demQuarzporphyr liegt bei Kaltern mindestens 500 mtiefer als auf der gegenüberliegenden Seite desEtschtales bei Branzoll. Paral lel dazu dürfte wohlauch – unter mächtigem Quartär verborgen – eineStörungszone durch die weite Talung von Eppan-Kaltern im Überetsch verlaufen, denn der Quarzpor-phyr zwischen Gaid und Tramin passt nicht zu dem
auf der anderen Seite dieser Talung. Die Oberflächedes Quarz porphyrs am Montiggler Wald scheintmehr als 230 m tiefer zu liegen als am gegenüber-liegenden Gandberg bei Oberplanitzing, der amMitterberg knapp 100 m tiefer als am gegenüberlie-genden Seeberg bei Altenburg. Insgesamt scheintdie Etschtalstörung hier also die Struktur eines Gra -bens mit etwas ungleich hoch liegenden Graben-schultern zu besitzen. Außerdem ist die Schichtfolge durch mehrere
quer dazu verlaufende, vor allem E-W- bis SE-NW-orientierte Störungen in einzelne Schollen zerlegt.An solchen Abschiebungen verspringt die Oberkan-te des Quarzporphyres erkennbar, etwa nördlich vonSöll oder unmittelbar nördlich des Bergsturzes vonEppan, wo sich zwischen Matschasch und demSchloss von Englar eine Sprung höhe von rund 480m ergibt! Viele dieser Störungen werden durchgroße Täler und Schluchten nachgezeichnet, dievom Überetsch zum Mendelkamm hinaufziehen,z.B. das Höllental. Oberhalb von Penon und Graun ist eine größere,
über weite Abschnitte subhorizontal verlaufendeÜberschiebungsbahn kartierbar (Vigo-di-Ton-Ter-meno-Linie), entlang der die Dolomite des Mendel-kammes auf unterschiedli che Trias- und Jura-Ge-steine in südöstlicher Richtung über schoben sind.An dieser Überschie bungsbahn sind die Dolomiteder Deckenbasis extrem stark beansprucht und ineiner teilweise dekametermächtigen Zone klein-stückig zerbrochen worden. Diese jedenfalls postju-rassisch ent standene Überschiebungsbahn ist sicheralpidisch. Sie wird von einigen der oben erwähntenQuerstörungen versetzt, die gleichfalls alpidischoder jünger sind. Andere Störungen, etwa die E-W-verlaufende große Abschiebung mit einer Sprung-höhe von mindestens 430 m, die zwischen Graunund Kurtatsch den Nordrand des „Kurtatscher Lo-ches“ markiert, müssen dagegen schon in der Triasaktiv gewesen sein, da sich die Mäch tigkeit der„Zwischenschichten“ an dieser Störung sprunghaftändert. Diese Störung scheint aller dings abschnitts-weise den Charakter einer Aufschiebung zu besit-zen, also wohl durch die alpi dische Einengungstek-tonik überprägt zu sein.
11..33 DDaass QQuuaarrttäärr iimm ÜÜbbeerrbblliicckk
Die permotriassischen Festgesteine des Überetschwerden großflächig von lockeren Ablagerun gen des
3Geo.Alp, Vol. 2, 2005
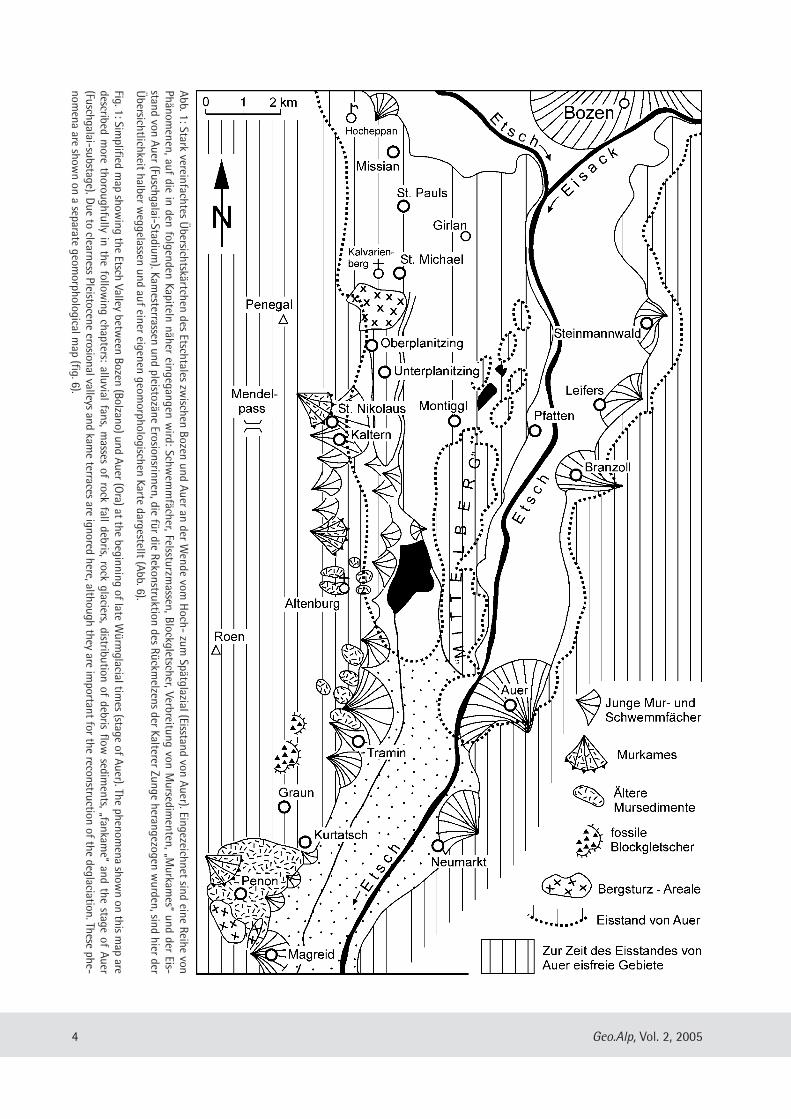
4 Geo.Alp, Vol. 2, 2005
Abb. 1: Stark vereinfachtes Übersichtskärtchen des Etschtales zw
ischen Bozen und Auer an der Wende vom
Hoch- zum
Spätglazial (Eisstand von Auer). Eingezeichnet sind eine Reihe vonPhänom
enen, auf die in den folgenden Kapiteln näher eingegangen wird: Schw
emmfächer, Felssturzm
assen, Blockgletscher, Verbreitung von Mursedim
en ten, „Murkam
es“ und der Eis-stand von Auer (Fuschgalai-Stadium
). Kamesterrassen und pleistozäne Erosionsrin nen, die für die Rekonstruktion des Rückm
elzens der Kalterer Zunge herangezogen wurden, sind hier der
Übersicht lichkeit halber w
eggelassen und auf einer eigenen geomorphologischen Karte dargestellt (Abb. 6).
Fig. 1: Simplified m
ap showing the Etsch Valley betw
een Bozen (Bolzano) und Auer (Ora) at the beginning of late W
ürmglacial tim
es (stage of Auer). The phenomena show
n on this map are
described more thoroughfully in the follow
ing chapters: alluvial fans, masses of rock fall debris, rock glaciers, distribution of debris flow
sediments, „fankam
e“ and the stage of Auer(Fuschgalai-substage). D
ue to clearness Pleistocene erosional valleys and kame terraces are ignored here, although they are im
portant for the reconstruction of the deglaciation. These phe-nom
ena are shown on a separate geom
orphological map (fig. 6).

Quartärs verdeckt, die in der weiten Talung vonEppan-Kaltern erhebliche Mächtigkeiten erreichenkönnen. Es handelt sich vor allem um Geschiebeleh-me, Schmelzwasserkiese, Seesedi mente und Murab-lagerungen, deren Mächtigkeiten mit zunehmenderHöhenlage generell abneh men. Der größte Teil die-ser Sedimente ist während des Pleistozäns, vorallem während der Würmeiszeit entstanden undsteht in direktem oder mittelbarem Zusammenhangmit dem Etsch gletscher, der in den kältesten Ab-schnitten des Eiszeitalters zeitweise das ganzeEtschtal aus füllte.Während des Vereisungsmaximums der letzten
Eiszeit, vor ca. 20.000 Jahren, dürfte das Etschtalsüdlich von Bozen bis in Höhen von über 2000 mmit Gletschereis erfüllt gewesen sein (Hantke 1983:197). Nur noch die höchsten Teile des Mendelkam-mes, am Roèn (2116 m), überragten noch die Eis -oberfläche (Klebelsberg 1949, Husen 1982). Südtiroldürfte damals ähnlich im Eis ertrunken gewesensein wie die Gebirge Ostgrönlands (Scholz 1984,1986). Über dem Mendelpass stand das Eis des Et-schgletschers mit dem im Nonstal liegenden Noce-Gletscher in Verbindung (Hantke 1983: 197). DerEtschgletscher stirnte in dieser Zeit noch südlich desGar dasees südlich Solferino (Habbe 1969). Die Ge-schiebe, die der Etschgletscher transportierte, stam-men größtenteils aus den zentralalpinen Nährge-bieten dieses Gletschers, vor allem aus den Ötztal-Stubaier Alpen, der Silvretta, dem Ortler-Gebiet,den Sarntaler Alpen, westlichen Ziller taler Alpenund westlichen Dolomiten. Beim Rückschmelzen des Eises im ausgehenden
Hochglazial sank die Eisoberfläche der großenTalgletscher – natürlich auch die des Etschglet-schers – langsam ab. Dadurch wurden die über -steilten Talhänge freigegeben und waren zuneh-mend der Erosion ausgesetzt. Auf der Höhe vonAuer muss sich der Etschgletscher beim Dünnerwer-den des Eises in zwei Eisloben aufgespalten haben(Abb. 7): eine Eiszunge floss über Bozen und folgtedem Etschtal abwärts (Etschtalzunge), eine zweiteEiszunge drang bei Missian ins Überetsch ein undfolgte der weiten Talung von Kaltern (KaltererZunge). Große Felskuppen, die vom Wilden-Mann-Bühel über den Großen Priol, Jagenberg, Mitterberg,Unterberg und über die Leuchtenburg zum Piglonziehen (insge samt teilweise als „Mittelberg“ be-zeichnet), wirkten dabei als Eisteiler (Abb. 1, 7). DasEis der Etschtalzunge muss um ein Vielfaches mäch-tiger gewesen sein als das der Kalterer Zunge. Be-
sonders die Kalterer Zunge und ihr langsames Rück-schmelzen lässt sich anhand entspre chenderAblagerun gen gut rekonstruieren.Der Etschgletscher hat das Etschtal zum weiten
Trogtal umgeformt, dessen trogähnlicher Talquer -schnitt aber nicht sichtbar ist. Der heutige Talbo-den, die landwirtschaftlich intensiv genutzte Etsch-talebene, ist eine Akkumulationsfläche, die erstwährend und nach dem Rück schmelzen der Glet-scher entstanden ist und bei Andrian rund 240 m,bei Tramin 215 m über dem Meer liegt. Das Etschtalist, wie alle großen Alpentäler, mit mächtigen quar-tären Ablagerungen aufgefüllt, vor allem mit flu-viatilen Kiesen und Seesedimenten. Am Aufbau derquartären Tal füllung sind zwischen Bozen und Sa-lurn entsprechend einer mündl. Mitt. von Herrn Dr.W. Sadgorski (vor mals LfW, München) auch mächti-ge Torfe mit geringmächtigen Auelehm-Zwi-schenlagen beteiligt (insgesamt 30 und 60 m).Randlich dürften auch Rutschmassen und Mursedi-mente am Aufbau der Talfüllung beteiligt sein. DerFelsuntergrund ist bei einer Bohrung südlich vonAndrian erst in einer Teufe von über 670 m unterder Oberfläche erreicht worden (Werth 2003). BeiAuer hat eine Bohrung den Felsuntergrund in einerTiefe von 200 m dagegen noch nicht erreicht(mündl. Mitt. Dr. Volkmar Mair).
22.. LLaannddsscchhaaffttsseelleemmeennttee iimm ÜÜbbeerreettsscchh
22..11 RRuunnddhhööcckkeerr uunndd GGlleettsscchheerrsscchhlliiffffee
Weit verbreitet sind im Überetsch eisüberschlif-fene Rundhöckerlandschaften. Große Felder mitRundhöckern sind fast ausschließlich auf Quarzpor-phyr-Oberflächen ausgebildet, z.B. in der Umge-bung der Montiggler Seen, am Seeberg bei Alten-burg oder am Kalvarienberg in St. Micha el (Eppan).Allerdings scheint die Ausbil dung ideal geformter,walrückenartiger Rundhöcker, mit flachen, ge-schrammten Luv- und stei len, gebrochenen Leesei-ten durch die engständige Klüftung vielfach verhin-dert worden zu sein. Einige ideal geformte Rund-höcker sind am Trimm-dich-Pfad östlich des Sport-platzes von Kaltern zu finden. Schöne Rundhöcker-felder sind auch auf dem Plateau von Unterfenn-berg südlich von Margreid auf Contrin-Dolomitentwickelt. Die anderen Gesteine des untersuchtenGebietes sind offenbar nicht hinreichend isotrop
5Geo.Alp, Vol. 2, 2005

und fest, um die Entwicklung von Rundhöckern zu-zulassen. Geschrammte Gletscherschliffe sind ge-wöhnlich nur dort erhalten geblieben, wo dieGesteins oberflächen durch eine hinreichend mäch-tige Auflage von Geschiebelehmen vor derVerwitte rung geschützt waren. Trotz einer anzu-nehmenden Eis überlagerung von 1500 bis 1800 mim Überetsch, die an sich zur Ausbildung von Si-chelmarken und Parabelrissen ausreicht, wurden aufden eisüberschliffenen Gesteinsoberflächen keineent sprechenden Strukturen beobachtet.
22..22 TTiillllee ((GGeesscchhiieebbeelleehhmmee,, GGeesscchhiieebbeessaannddee))
Stellenweise treten im Überetsch schluffig-san-dige und stark verdichtete Geschiebelehme auf(lodgement till, „Grundmoräne“), die teilweise sowenig Schluff enthalten, dass sie besser als Geschie-besande bezeichnet werden sollten. Diese Tille ent-halten vor allem Kristallingeschiebe, auch vielQuarzporphyr, aber vergleichsweise wenige undkleine Karbonatkomponenten (Abb. 3). Die westli-che Hälfte des riesigen, fast 10 km breiten Talglet-schers, die den Überetsch erreich te, dürfte vor allemaus Eis bestanden haben, das dem Etschgletsacheraus dem W des Ein zugs gebietes zugeführt wordenist, vor allem aus dem Val Müstair, Martelltal undUltental. Ein Großteil der Geschiebe im Überetschdürfte demnach vor allem aus der relativ nahe gele-genen Ortlergruppe stammen. Gelegentlich sindauch Serpentinit-Komponenten zu finden, die ausdem Oberengadin stammen und über eine Tansflu-enz am Reschenpass ins Etschtal gelangt sein dürf-ten (Ebers 1972: 114).Die in den Tillen enthaltenen Geschiebe sind ge-
wöhnlich recht gut gerundet, aber nur die Karbona-te sind deutlich gekritzt. Lokal dünnen diese Abla-gerungen stark aus und bilden einen geringmächti-gen Geschiebeschleier, doch sind Aufschlüsse selten,in denen sich die Mächtigkeit dieser Geschiebeleh-me ermitteln lässt. Der teilweise ausgezeichneteRundungsgrad der Kristal linkomponenten ließe sichdurch die Annahme erklären, dass das Eis ältere flu-viatile Kiese im Etschtal und im Überetsch aufgear-beitet haben könnte.Oft liegen Geschiebelehme dem eisüberschliffe-
nen Felsuntergrund in wechselnder Mächtigkeit di-rekt auf. Insgesamt sind richtige lodgement tills, diewohl aus Zeiten mit hoher Eisbedeckung stammen,weit verbreitet. Geschiebelehme mit einem eindeu-tig lokalen Geschiebespektrum, also Ablagerungenvon Lokalgletschern des Mendelkammes, warennicht zu finden.
22..33 EEiissrraannddaabbllaaggeerruunnggeenn ((MMoorräänneennwwäällllee uunnddKKaammeess))
Schon Penck (in Penck & Brückner 1909: 924)war am Westhang des Mitter- und Unter berges ge-genüber von Kaltern ein großer Moränenwall auf-gefallen, der südlich von Girlan be ginnt, die Mon-tiggler Seen abdämmt und bis gegen den Kalterer
6 Geo.Alp, Vol. 2, 2005
Abb. 2: Schema der Genese von Kamesterrassen am Westranddes Etschgletschers. Die Kamesterrassen wurden durchSchmelzwässer zwischen Berghang und Eisrand aufgeschüttet,teilweise auch unter Beteiligung von Mur material, das denSchmelzwassersedimenten vom Berghang her seitlich zuge-führt wurde (unten). Nach dem Ab schmelzen des Gletscher -eises wurden die Kamesterrassen zertalt (oben).
Fig. 2: Simplified sketch showing how kame terraces at thewestern rim of the retreating Etsch valley glacier may haveformed. They have been generated by accumulation of melt-water sediments within the gap between the moun tain slopeand the glacier. Gravel derived from the slope above has beenadded by debris flows (below). After the glacier ice has van-ished these kame terraces have been cut by erosional valleys(above).

See hinziehen soll. Nach Penck (in Penck & Brückner1909: 924) markiert er einen längeren Gletscher-halt. Weniger zusammenhängend sieht er die Morä-nenwälle an der Westseite von Eppan. Er gibt an,dass sie sich oberhalb St. Pauls an den Fuß des Buch-berges lehnen, bei Planitzing durch das Trümmer werkeines Bergsturzes und bei Kaltern durch einen großenSchuttkegel unterbrochen sind (Penck in Penck &Brückner 1909: 924 f.). Die Existenz dieser Eisrand -ablagerungen, Moränenwälle und Kamesterrassen,konnte durch die Kartierungen tatsächlich bestätigtwerden. Im E der Talung gibt es am Westhang des Mitter-
und Unterberges gegenüber von Kaltern nicht nureinen einzigen großen Moränenwall, sondern einganzes System von kiesigen Lateralmorä nen undKamesterrassen (Abb. 2, 6), mit deren Hilfe sichmindestens zwei unterschiedliche Eis stände einer„Kalterer Zunge“ rekonstruieren lassen, die in derTalung von Eppan-Kaltern gele gen haben undknapp südlich des heutigen Kalterer Sees gestirnthaben muss (Abb. 1, 7). Die am höchsten gelegeneund deut lichste dieser Strukturen ist ein Wall, denman auf über 1,5 km Länge verfolgen kann. Er hatein deutliches Gefälle in südlicher Richtung undliegt an seinem N-Ende um ca. 60 m höher als anseinem S-Ende (Taf. 1). Ursprünglich scheint es sichwohl eher um eine Kamesterrasse gehandelt zuhaben als um einen Wall. Bei sinkendem Eis standwurde durch ein sich bergseitig eintiefendesSchmelz wassertal (Fuschgalai) ein wallartigerRücken abgetrennt (Abb. 6). Weiter im S lässt sichder Eisstand von Fuschgalai mit Kamesterrassen amFalzig weiterverfolgen, die am Kreithof wieder ineinem deutlichen Wall auslaufen (Abb. 6). DiesesWallstück ist inzwischen größtenteils einem Kiesab-bau zum Opfer gefallen. Obwohl die in den 60erJahren ausgebeutete Grube inzwischen völlig ver-wachsen ist, lässt sich immer noch erken nen, dassdas Material, aus dem der Wall besteht, stark kiesigund sehr kristallinreich ist und zahlreiche meter-große Kristallinblöcke ent hält. Castiglioni & Trevi-san (1973: 6 ff.) rechnen diese groben, auf einerihrer Abbildungen erkennbar geschichteten Kiesefreilich zu den glazi fluvialen Schottern des „Con-glomerato di Caldaro“. Diese Kiese sind aber in un-mittelbare Nähe des Eisrandes entstanden, da siegroße Mengen gekritzter Geschiebe enthalten. Anders als Penck (in Penck & Brückner 1909:
924) glaubt, sind die Wallsysteme in Rich tung Mon-tiggler Seen und Girlan nicht weiter zu verfolgen.
Das auf dem Moränenwall abgreif bare Gefällespricht eher dafür, dass sich der Eisrand der KaltererZunge in der Zeit des Fuschga lai-Stadiums an denNE-Hang des Jagenberges und sich südlich desGroßen Priol mit der Etsch talzunge vereinigt hat.Zwischen dem Wilden-Mann-Bühel und demGroßen Priol müssen damals mehrere Quarzpor-phyr-Kuppen das Eis als Nunatakker knapp überragthaben (Abb. 1, 7). Dieses Sta dium könnte zum Eis-stand von Auer gehören, der nach Hantke (1983:234) demjenigen von Kufstein auf der Alpennord-seite entsprechen soll. Nach Jerz (1993: 95) ent-spricht das einem Alter von etwa 15.000 bis 16.000Jahren vor heute.Im W der Talung Eppan-Kaltern gibt es, anders
als Penck (in Penck & Brückner 1909: 924) vermu-tet, kaum Moränenwälle, wohl aber ein System vonbreiten Kamesterrassen zwischen Kaltern und St.Josef am Kalterer See (Taf. 1, 2), die einen Eisstandnachzeichnen, den wir hier als Stadium von Kalternbezeichnen wollen (Abb. 6, 7). Die ursprünglichwohl zusammenhän genden, bis zu 500 m breitenTerrassen mit ebenen oder leicht welligen Ober-flächen sind durch jüngere, W-E-orientierte Erosi-onstäler, die dem generellen Gefälle des Hanges fol-gen, in mehrere Teilstücke zerlegt worden (Abb. 6).Am Barleitherhof ist ein N-S-orentiertes, wallarti-ges Teil stück der Kamesterrasse durch ein Ero -sionstälchen vom bergwärtigen Rest der Terrasseabge trennt worden (Abb. 6). Die Zertalung mussschon unmittelbar nach der Entstehung dieser Ter-rassen begonnen haben, denn viele der Erosionsrin-nen sind Trockentäler. Ein besonders großes Teil-stück der Kamesterrassen, auf dem der Ortskern vonKaltern steht, ist von der Berg seite her durch denkomplexen Schwem mfächer des Pfusser Bachesüberschüttet worden (Abb. 6). Penck (in Penck &Brückner 1909: 924) glaubt die Kamesterrassen inRichtung Ober planitzing und Eppan weiterverfol-gen zu können, was sich jedoch als unmöglichheraus stellte.Das Gefälle dieser Eisrandterrassen ist etwas ge-
ringer als das des Walles auf der Gegenseite. Sie lie-gen auch deutlich tiefer und entsprechen von ihrerHöhenlage her wohl eher den Kames terrassen anden Bergflanken unterhalb von Fuschgalai (Abb. 7).Mit dem Stadium von Fusch galai der Kalterer Zungedürften wohl eher drei kleine Terrassenreste ober-halb des Barleither Weges korrespondieren (Abb. 6).In den Kamesterrassen gibt es zahlreiche Aufschlüs-se, die Ein blicke in ihren inneren Aufbau erlauben.
7Geo.Alp, Vol. 2, 2005

Zum größten Teil bestehen sie aus gut ausgewa -schenen, geschichteten Kiesen, die teilweise sehrgrob sind und große Mengen gekritzter Geschiebeent halten, also sehr eisrandnah abgelagert wordensind. Daneben spielen geschichtete Sande undSchluffe eine wichtige Rolle. Die Kiesgrube vom Vo-glmeierhof westlich des Kalterer Sees, die bei Casti -glioni & Trevisan (1973: Abb. 7) abgebildet ist, zeigtkeine Schotter, die zum glazi fluvialen „Conglomera-to di Caldaro“ gehören, sondern eisrandnah ent-standene Kameskiese, wie sie in allen Kamesterras-sen auf der Westseite der Kalterer Zunge akkumu-liert worden sind.Verglichen mit den Kiesen innerhalb des Walles
auf der Ostseite der Talung ist das Material hierdeutlich reicher an Karbonatkomponenten. Stellen-weise konnten glazialtektonisch bedingte Schicht-störungen beobachtet werden. Obwohl zahlreichegekritzte Geschiebe zu finden sind, tre ten tillartige
Sedimente stark in den Hintergrund. Dafür sind indie Kamesterrassen stellenweise schluffreiche Sedi-mente mit lokalem Schutt integriert. In diesen Sedi-menten, die als Bestandteile der Kamesterrassen z.B.am Barleither Weg 500 m NNW‘ des Barleitherhofesoder im Tal ober halb von Schloss Kaltenburg aufge-schlossen sind, dominieren eckige Komponenten ausSchlern dolomit sowie aus Karbonaten, Schluff- undSandsteinen der Werfen Formation. Nur ganzunter geordnet finden sich auch Kristallingerölle. Beidiesen Sedimenten handelt es sich definitiv nichtum Lokalmoränen (siehe unten).
22..44 MMuurrsseeddiimmeennttee
Weit verbreitet sind im Untersuchungsgebiet Se-dimente, deren Habitus auf den ersten Blick an Tille(„Moränen“) erinnert, die aber von den Komponen-
8 Geo.Alp, Vol. 2, 2005
Abb. 3: Gegenüberstellung der Texturen genetisch unterschiedlicher Sedimente mit diamiktischer Kornverteilung im Etschtal. Aufden Bildern sind die wichtigsten im Aufschluss sichtbaren Eigenschaften dieser Sedimente sowie deren genetische Deutung schema-tisch dargestellt.
Fig. 3: Comparison of genetically different diamictic sediments in the Etsch valley. Textures and some other important macroscopicvisible features of these sediments are shown here, together with their genetic interpretation.

tenspektren, den Kornformen und den Kornober-flächen her keine glazigenen Sedimente sein kön-nen. Diese Sedimente haben eine dia miktischeKorn größenverteilung (Taf. 4) und sind von daherTillen ähnlich (Abb. 3). Es handelt sich um matrixge -stützte Sedimente mit einer sandig-schluffigenGrundmasse, in der zahlreiche grobe Komponen tenschwimmen. Die Korngrößen des Grobmaterials lie-gen im Bereich von Kies bis Blockwerk; gelegentlichkommen auch metergroße Blöcke vor. Die grobenKomponenten sind eckig, weisen vielfach scharfeBruchkanten auf, doch sind auch kantengerundeteBruch stücke zu finden. Gut gerundete und/oder ge-kritzte Komponenten, kristallines Material und an-dere Fremdgesteine fehlen oder sind zumindest sel-ten. Die Hauptmasse der Komponenten besteht ausSchlern-, Haupt- bzw. Contrindolomit sowie Bruch-stücken der Hartbänke aus der Werfen Formation.Doch die Zusammensetzung schwankt in weitenGrenzen. Es gibt Bereiche, in denen diese Gesteinefast nur aus Schlern- und Contrindolomit-Bruch-stücken bestehen, an anderen Stellen nur aus Frag-menten der Werfen Formation, manchmal auch auseiner Mischung aus beidem. Die Farbe der feiner-körnigen Matrix ist grau, häufig auch rötlich odergelblich, letzteres vor allem dort, wo viele WerfenerKomponenten in der Grobfraktion zu finden sind.Deutliche Schichtungsgefüge sind meist nicht zuerkennen, selbst dann nicht, wenn man meter hoheAuf schlüsse begutachten kann. Selten kommenaber doch Lagen mit deutlich weniger Grobmaterialoder schluffige, sandige oder kiesige Einschaltun-gen vor. Im Aufschluss sind diese Gesteine überraschend
standfest; fast vertikale Straßen- und Wegan -schnitte erweisen sich seit Jahrzehnten ohne Siche-rungsmaßnamen als standfest (Taf. 4). Diese Gestei-ne finden sich im Untergrund vieler Weinberge zwi-schen Kaltern und Margreid. Die stei nigen Sedi -mente sind auf den Feldern nur schwer zu bearbei-ten, so dass sie die Weinbauern als „Kampf erde“oder „Kampf“ bezeichnen, ein Ausdruck, der ande-renorts in Südtirol auch für lodgement-till („Grund-moräne“) verwendet wird (mündl. Mitt. Dr. VolkmarMair, Bozen). Die Sedimente bilden oft mächtigeDecken über dem Felsuntergrund, deren basaleAuflagerungs flächen oft geneigt sind und parallelzum Hang einfallen. Mitunter kommen sogar fastvertikale Kontakt flächen an Stellen vor, wo die Se-dimente offensichtlich alten, verschütteten Felsstu-fen angelagert sind. Die Mächtigkeiten sind meist
nur schwer abschätzbar. Oft lassen sich aufgrundder Tiefe von Erosionstälern Mächtigkeiten vonmehreren Dekametern schätzen; in Einzelfällenkommt man auf 60 bis 80 m.Im Überetsch sind Sedimente dieses Typs weit
verbreitet (Abb. 1). Als fast geschlossene Deckenvon erheblicher Mächtigkeit treten diese Ablage-rungen an den Hängen oberhalb von Kurtatsch,Entiklar und Margreid auf, wo sie bis über Penonhinauf die tonig-kalkigen „Zwischen schichten“ desUnterladin zusammen mit ihren mächtigen Kalk-und Dolomiteinschaltungen überlagern. Nur in be-sonders tief eingeschnittenen Erosionstälern wirdhier das Quartär durch schnitten. Hier bilden dieseAblagerungen eine fast geschlossene Decke miteiner Gesamtfläche von fast 5 km2. Weiter im N sinddiese Sedimente weniger geschlossen verbreitet,nehmen jeweils kleinere Flächen von immerhinnoch vielen Hektar Größe ein. Auch hier können dieVorkommen mehrere Dekameter mächtig werden.Auffällig ist, dass die Verteilung der Vor kommeneine klare Bezie hung zu den bedeutenderen, tiefeingeschnittenen Rinnen zeigen, die zum Mendel-kamm hinauf ziehen. Ein besonders mächtiges Vor-kommen dieser Sedimente bildet z.B. die markanteKuppe am Ausgang des Höllentales in Tramin, aufder St. Jakob in Kastellaz liegt (Abb. 1). Ein anderesVorkommen ist beispielsweise an der Straße vonKaltern nach Altenburg auf geschlossen, genau un-terhalb des tief eingeschnittenen Val della Lavine.Manche dieser merkwürdigen Sedimente zeigen
eindeutige Beziehungen zu jungen Oberflächen -formen. „Kampferde“-Sedimente, die z.B. NW’Penon, zwischen Altenburg und Kaltern oder ober-halb von Pfuss bei St. Nikolaus in Kaltern vorkom-men (Taf. 3), bauen jeweils mehrere parallel ori -entierte, schmale Rücken auf, die von tief einge-schnittenen Erosionstälern vonein ander getrenntwerden. Die Oberflächen benachbarter Rücken wei-sen ein identisches Gefälle von 15 bis 30° auf (Taf.3). Talwärts sind diese Rücken durch einen Gefälle-knick begrenzt; unterhalb davon hören die Rückenmit einer kräftigen Versteilung des Hanges auf (Taf.3). Dieser Gefälleknick liegt bei benachbar tenRücken ungefähr auf der gleichen Höhe; die Struk-turen erscheinen dadurch wie abgehackt. Bei diesenRücken könnte es sich um Erosionsreste von fächer-artigen Gebilden zu handeln, wohl um die Restealter Murfächer, die von parallel orientierten Tälernzerschnitten worden sind (Abb. 6). Auf grund günsti-ger Aufschlussverhält nisse am anerodierten Mur-
9Geo.Alp, Vol. 2, 2005

fächer von Pfuss ist zu erkennen, dass die Haupt-masse der Höhen rücken tatsächlich aus Ablagerun-gen dieses Typs aufgebaut wird. Schon Penck (inPenck & Brückner 1909: 924) hat diese Vorkommenbei St. Nikolaus in Kaltern gekannt, in ähnlicherWeise als „Schuttkegelrudimente“ gedeutet und sieins „Spätglazial“ gestellt. Am Fuß der Ver steilungenunterhalb des Gefälleknicks scheinen die Mursedi-mente durch eine Zunahme des Kri stallinmaterials,des Rundungsgrades der Komponenten und demvermehrten Auftreten gekritzter Geschiebe in kar-
bonatreiche Geschiebelehme überzugehen, was dieunmittelbare Nähe des Eises am talwärtigen Endeder Strukturen anzeigt. Hier besteht also der begründete Verdacht, dass
es sich um Murfächer handelt, die gegen den Eis -rand des zurückschmelzenden Etschgletschers ge-schüttet worden sind; wir wollen sie hier „Mur -kames“ nennen (Abb. 1, 6). Neben diesen „Murka-mes“ gibt es auch, wie oben schon dargelegt, ge-wöhnliche Kamesterrassen mit ebenen Oberflächen,die außer kiesigen oder schluffig-san digen, gut ge-schichteten Schmelzwassersedimenten auch ab-schittsweise „Kampferde“-Sedi mente enthalten.Solche Kamesterrassen sind z.B. NE‘ von Penon odersüdlich von Kaltern am Barleiter Weg zu finden.Die meisten Vorkommen von Sedimenten dieses
Typs lassen indes keinerlei Beziehungen zu irgend-welchen charakteristischen Oberflächenformen er-kennen. An einigen Stellen ist zu beo bachten, dassderartige Ablagerungen eindeutig von kristallinrei-chen Geschiebelehmen überla gert werden. Das istz.B. an Ablagerungen im Hügel von St. Jakob in Ka-stellaz in Tramin ganz in der Nähe des Bungalowsder Wildbachverbauung zu sehen. Dieses und einigeandere Vorkom men scheinen zudem rundliche,drumlinähnliche Geländeformen zu bilden und soll-ten folglich vom Gletschereis überfahren wordensein. Deshalb muss zumindest ein Teil dieser Sedi-mente vor dem Höchststand des Eises der letztenEiszeit entstanden sein. Ähnlich sieht das auchPenck (in Penck & Brückner 1909: 921). Er argu-mentiert, dass sie zeitlich zwischen zweiaufeinander folgende Vergletscherungen zu stellenwären, da sie gelegent lich auch (umgelagerte)Fremdge schiebe enthalten. Auch bei Meran hatPenck (in Penck & Brückner 1909: 921) solcheSchutt ablagerungen gefunden, zwischen Gardaseeund Meran will er gar Reste von vier verschiedenalten Schuttkegeln nachgewiesen haben. Dafür, dass es sich bei den „Kampferde“-Sedi-
menten um Ablagerungen von debris flows handelt,spricht vor allem die praktisch fehlende Rundungder Komponenten und die äußerst schlechte Sortie-rung des Materials (Johnson & Rodine 1984: 315).Warum sind die „Kampferde“-Abla gerungen, wennman sie als Mursedimente deutet, kaum oder garnicht geschichtet, obwohl post glaziale mudflow-Sedimente, genauso wie rezente Murkegel, immereine wenn auch undeut liche Schichtung aufweisen(Costa 1984, 1988, Davies 1988)? Der typische Auf-bau junger Mursedi mente kann bei spielsweise im
10 Geo.Alp, Vol. 2, 2005
Abb. 4: Schema der Genese von „Murkames“ am Westrand desEtschgletschers. Die Murkames entstanden als Mur fächer undenthalten ausschließlich Material, das aus Erosionsrinnen imHang gegen den Rand des Etschgletschers vorgeschüttetwurde (unten). Nach dem Abschmelzen des Eises wurden dieMurkames, die talwärts primär durch eine steile Sackungskan-te begrenzt sind, erosiv zerschnitten (oben).
Fig. 4: Simplified sketch showing how a "fankame“ at the west-ern rim of the Etsch valley glacier may have been formed.Originally they have been generated as alluvial fans by accu-mulation of debris flows at the glacier rim, the debris derivingentirely from the hillslope above (below). These "fankame“ ex-pose a typical steep edge at their lower part and have been cutby erosional valleys since the glacier ice has vanished (above).

Nussental am Hang oberhalb Kuenburg am KaltererSee studiert werden, wo ein steiler Murkegel durcheine kleine Grube angeschnitten ist. Das hieraufgeschlos sene diamikti sche Material, sehr reichan eckigen Quarzporphyr-Komponenten, ist un-deutlich geschichtet. Der geschichtete Eindruckwird durch einen Wechsel in der Korngröße und inder Zusammensetzung der Mursedimente erzeugt,wie sie für Ablagerungen typisch sind, die von de-bris flows aufgebaut werden (Coussot & Meunier1996). Vielleicht hängen die Unterschiede zu den fossi-
len Mursedimenten damit zusammen, dass dieheute noch aktiven, mehrere Dekameter mächtigenMurkegel im Laufe von vielen einzelnen Murereig-nissen akkumuliert worden sind. Bei jedem Mur-gang werden hier jeweils nur wenige Meter Sedi-ment auf einmal abgelagert, da sich die Mure übereinen Teil des Fächers flächenhaft ausbreiten kann.Gleiches gilt auch für die rezenten Beispiele, die beiJohnson & Rodine (1984: 266 ff.) angeführt wer-den. Die viele Dekameter mächtigen „Kampferde“-Sedimente sind im Gegensatz dazu wohl alle kalt-zeitlich und bei sinkenden Eisständen abgelagertworden. In den Kaltzeiten gab es auf den frisch vomEis freigegebenen Steilhängen, wo das Lockermate-rial für die Muren mobilisiert werden konnte, keineVegetation, die den hier liegenden Hangschutt undGeschiebelehme hätte stabilisieren können, undauch der sich nach dem Eisrückzug aufbauende Per-mafrost dürfte bald in der ausgehenden Eiszeit zu-sammengebrochen sein (Haeberli 1996). Dadurch istbei einem einzelnen Ereignis offenbar ungleichmehr Material umgelagert worden als heute. Nochdazu konnten sich die Muren auf den Fächern nichtausbreiten sondern stauten sich am Eisrand (Abb. 4),was schon bei einem einzigen Ereignis zur Akkumu-lation von dekameter mächtigen, intern weitgehendungeschichteten Mursedimenten führte (Abb. 5).
22..55 BBlloocckkgglleettsscchheerr,, LLookkaallgglleettsscchheerr uunnddGGeehhäännggeebbrreekkzziieenn
Seit dem Abschmelzen der Gletscher haben sichvor allem unter den Dolomit-Steilwänden bedeu-tende Hangschuttmassen akkumuliert. GroßeSchuttmassen haben sich vor allem im oberen Teileiner mehr als 1 km breiten Hangverflachung gebil-det, die oberhalb von Kurtatsch zum Tal hin durcheine markante Geländestufe aus Contrindolomit be-
grenzt wird. Diese Hangverfla chung, auf der auchder Ort Graun liegt, ist letztlich durch die hier vor-kommenden kalkig-mer gelig „Zwischenschichten“bedingt, die besonders leicht erodiert werden konn-ten. Etwa 1 km nördlich von Graun, im Oberen Ge-meindewald westlich des Hofes Locherer, liegt einenach drei Seiten steil abfallende, einige hundertMeter breite Hangnase, deren Oberfläche ein auf-fällig unruhiges Relief trägt. Das dicht bewaldeteGelände, dessen höchster Punkt 1018 m hoch liegt,zeigt ein kompliziertes System von Wällen mit tie-fen, abflusslosen Depressionen dazwischen, die anToteislöcher erinnern. Ein Teil der wallartigenRücken scheint sich zu zungenartigen Loben zu-sammenzuschließen. Das Gebiet, das hangaufwärtsin die Schutthalden unter den Schlerndolo mit-Wänden übergeht, besteht selbst ausschließlich aushoch porösem Dolomitschutt. Fremd material undgerundete Komponenten fehlen praktisch völlig. Einetwas kleineres und ca. 50 Höhenmeter tiefer lie-gendes Areal mit morphologisch vergleichbarenStrukturen wird vom Tra miner Höhenweg etwa 1km weiter im N gequert. Bei beiden Strukturen dürfte es sich um Block-
gletscher handeln, also ehemals gefrorene Schutt -
11Geo.Alp, Vol. 2, 2005
Abb. 5: Schematische Schnitte durch moderne Murfächer und„Murkames“, die während des Rückschmelzens des Etschglet-schers entstanden sind. Durch den Rückstau am Rande desTalgletschers waren die Sedimente, die ein ein ziger Murgangbzw. ein einzelnes Murereignis hinterließ, bedeutend mächti-ger (unten) als in heutigen Murfächern (oben). Dadurch er-scheinen die Schichtfolgen in „Murkames“ weitgehend unge-schichtet.
Fig. 5: Schematic cuts through a modern fan in comparison toa late Pleistocene "fankame“, which was generated when theEtsch valley glacier retreated. Due to the damming effect ofthe glacier rim, the sediment succession from a single debrisflow is much thicker within a "fankame“ (below) than in a re-cent alluvial fan (above). Therefore the successions within"fankame“ are poorly stratified.
12 Geo.Alp, Vol. 2, 2005
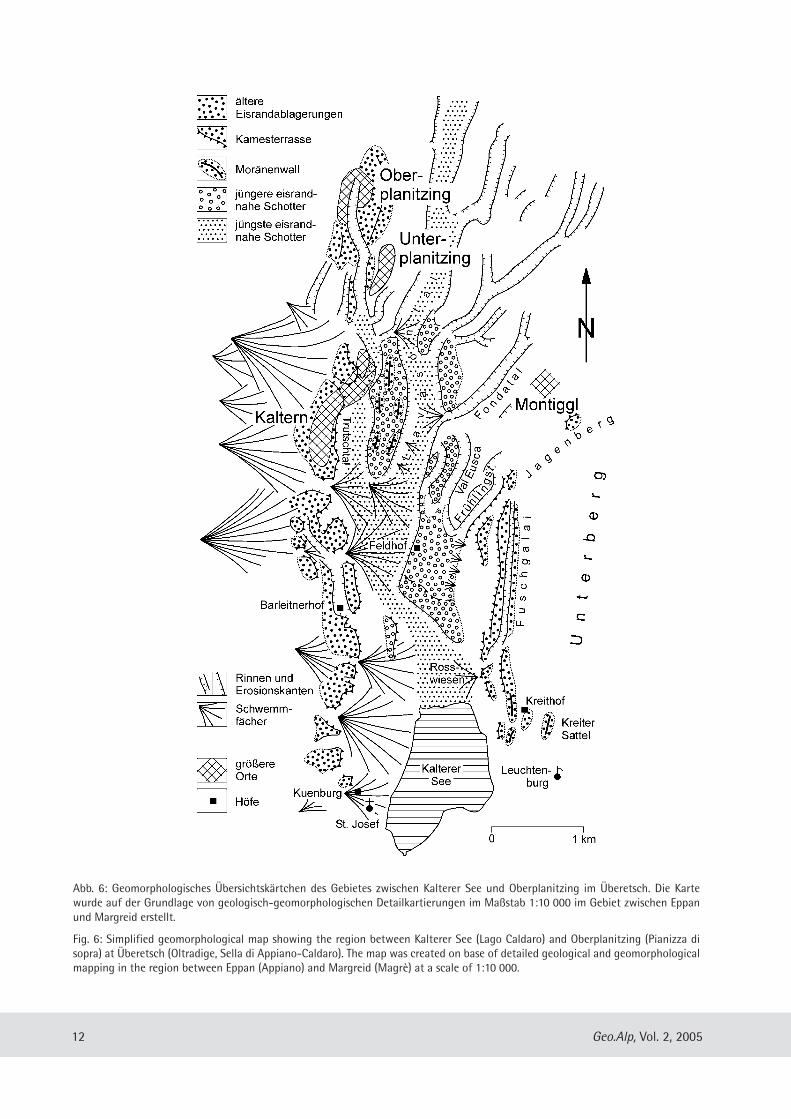
Abb. 6: Geomorphologisches Übersichtskärtchen des Gebietes zwischen Kalterer See und Oberplanitzing im Über etsch. Die Kartewurde auf der Grundlage von geologisch-geomorphologischen Detailkartierungen im Maßstab 1:10 000 im Gebiet zwischen Eppanund Margreid erstellt.
Fig. 6: Simplified geomorphological map showing the region between Kalterer See (Lago Caldaro) and Ober pla nit zing (Pianizza disopra) at Überetsch (Oltradige, Sella di Appiano-Caldaro). The map was created on base of detailed geological and geomorphologicalmapping in the region between Eppan (Appiano) and Margreid (Magrè) at a scale of 1:10 000.

massen, die sich kriechend wie ein Gletscher bewe-gen (Abb. 1). Diese Blockgletscher sind fossil undbewegen sich heute mit Sicherheit nicht mehr aktiv,denn in Höhen um 1000 m ist in den Süd alpenunter den heutigen Klimabedingungen (Weinbaubis in über 800 m Höhe!) mit Sicher heit kein Per-mafrost mehr zu erwarten. Sie dürften sich nachdem Rückschmelzen des Etsch gletschers an derWende vom Hoch- zum Spätglazial gebildet haben,vor allem während der spät glazialen Klimadepres-sionen. Blockgletscher ,aber auch richtige kleine Lo-kalgletscher, die sich gleichzeitig in Karen unterhalbdes Mendelkammes gebildet haben könnten, sinddenkbare Aus löser für große Murgänge, die für dieGenese der oben beschriebenen pleistozänen Mur -sedi mente verantwortlich waren. Am Nordhang des Höllentales oberhalb von Tra-
min liegt ein auffälliger Hangvorsprung, der durchdas Vorkom men einer calcitisch zementierten, hochporösen quartären Brekzie bedingt ist. Diese weit-gehend ungeschichtete Gehängebrekzie, die fastausschließlich aus eckigem Dolomitschutt besteht,lagert der Werfen Formation in einer Mäch tigkeitvon mindestens 10 m auf, in die die Höllental-schlucht eingeschnitten ist. Über das genaue Alterder Brekzie lässt sich nichts aussagen, doch weisenErosion sowie starke Zementierung des Vorkom-mens darauf hin, dass es sich möglicherweise umpräwürmglaziale Bildungen handelt. Weitere Vor-kommen von ähnlichen Gehängebrekzien sind auchnahe dem Hof Steiner am Hang oberhalb des Höl-lentales gegenüber von Tramin oder westlich von St.Nikolaus bei Kaltern zu finden. Stacul (1980) stelltdie Bildung des Karbonatschuttes, aus dem dieGehängebrekzie von St. Nikolaus besteht, in eineKaltzeit, unmittelbar nach dem Rück schmelzen desEtschgletschers. Ihre Verkittung durch „Kalksinter“soll hingegen in einem Inter glazial oder einem In-terstadial erfolgt sein.
22..66 KKaalltteerreerr SScchhootttteerr
Nach Hantke (1983: 233) ist die weite Talung vonEppan-Kaltern mit mächtigen quartären Kie sen er-füllt, die ihrerseits von würmeiszeitlichen Geschie-belehmen bedeckt sein sollen. Die Gesamtmächtig-keit der Schotter von Eppan beträgt nach Blaas(1892) bis zu 200 m. Die Schot ter werden dem„Konglomerat von Kaltern“ gleichgesetzt, obwohlsie größtenteils nicht verfes tigt sind. Nach Ebers
(1972) sind die „Überetscher Schotter“ nicht älterals Eem. Nach Castiglioni & Trevisan (1973) ist das„Conglomerato di Caldaro“ von Schmelzwässern desvorstoßenden Etschgletschers aufgeschüttet wor-den. Seine Aufschüttung soll im Val-Caldaro-Inter-stadial erfolgt sein, das mit einem radiometrisch er-mittelten Alter von rund 30.000 Jahren (Fuchs1969) dem Interstadial von Baumkirchen in denNordalpen entsprechen könnte. Auch Klebelsberg(1926, 1935) und Ebers (1972) gehen davon aus,dass alle größeren Kiesvor kommen im Überetschgenetisch identisch sind, eine einheitliche Be-deckung von Geschiebe lehmen aufweisen und des-halb vor dem Gletscherhöchststand der Würmeis-zeit entstanden sind.So einfach ist die Sache allerdings nicht. Ebers
(1972) und Castiglioni & Trevisan (1973) subsum-mieren unter den Begriffen „Überetscher Schotter“und „Conglomerato di Caldaro“ viele Kiese, die hierzu unterschiedlichen Zeiten und unter ganz unter-schiedlichen Bedingungen ent standen sind. Castig-lioni & Trevisan (1973) stellen beispielsweise diegroben Kiese zum „Conglomerato di Caldaro“, diefrüher am Kreithof („Maso Kreit“) westlich des Kal-terer Sees in einer Kiesgrube abgebaut worden sind(Castiglioni & Trevisan (1973: 6 ff.). Diese Kiese sindaber Teil eines komplexen Systems von Kamester-rassen und Wällen auf der Ostseite der KaltererZunge (siehe oben). Auch die westlich des KaltererSees gelegenen Kiese vom Vogelmeierhof (Castiglio-ni & Trevisan (1973: 6 ff.) gehören zu einem Systemvon komplexen Kamesterrassen, die auf der West-seite der Kalterer Zunge im ausgehenden Hochgla-zial der Würmeiszeit akkumuliert worden sind. Da-neben sind aber auch tatsächlich eindeutig prä-hochglaziale Bildungen zu finden.Tatsächlich ist die weite Talung von Eppan-Kal-
tern von kristallinreichen, teilweise sehr grobkör -nigen, abschnittsweise kaum geschichteten und oftschluffreichen Kiesen erfüllt, die größten teils sehrschlecht aufgeschlossen sind. In den hangendenAbschnitten der Kiese sind gekritzte Geschiebe häu-fig; fleckenweise tragen sie sogar eine Decke vonGeschiebelehmen; östlich von Kaltern sind im Han-genden dieser Kiese sogar wallähnliche Strukturenentwickelt. Da die Kar bonat- und Kristallinkompo-nenten dieser Kiese kaum Verwitterungserscheinun-gen zeigen, dürf ten sie vergleichsweise jung sein.Womöglich handelt es sich wenigstens teilweise umVorstoß schotter, vor allem in der Umgebung derMontiggler Seen, wo die Oberfläche kiesiger Abla -
13Geo.Alp, Vol. 2, 2005

ge rungen drumlinisiert ist. Vielfach dürfte es sichaber wohl auch um Schmelzwasser schotter aus derausgehenden Eiszeit handeln, die vor der zurück-schmelzenden Kalterer Zunge akkumulierten undbei einer Eisoszillation nochmals überfahren wur-den. Sie könnten in einem Totraum abgelagert wor-den sein, der sich zwischen der nach Nordenzurückschmelzenden Kalterer Zunge und demBecken des Kalterer Sees befand (Abb. 6). Die Kiese sind gewöhnlich locker und nicht oder
kaum verfestigt und enthalten immer wieder Ein-schaltungen von sandig-schluffigen Laminiten, beidenen es sich um Stillwasserablagerungen handelt.Nur in der kleinen Schlucht zwischen Festplatz undKalvarienberg in Kaltern, über die der Bach aus demTröpfeltal das Lavasontal erreicht, kommen auf derorographisch linken Tal seite durch calcitische Ze-mente fest verbackene, kristallinreiche Konglomera-te heraus. Diese mit Höhlen und Kavernen durch-setzten Ablagerungen sind wohl das „Konglomeratvon Kaltern“ im ursprünglichen Sinne. Es handeltsich um gut sortierte, ausgewaschene Schmelz -wasser sedimente, die zahlreiche Rollkieslagen ent-halten. Die Imbrication der Gerölle weist auf einengenerellen Sedimenttransport von N hin. DeutlicheVerwitterungserscheinungen an den Dolomit kom -po nen ten des Konglomerates lassen Zweifel auf-kommen, ob es mit den weit verbrei teten Kiesen derUmgebung etwas zu tun hat oder ob es nicht dochälter ist. Die fraglichen Vorstoßschotter und die Konglo-
merate sind jedenfalls in der Talung Eppan-Kal ternnur bis zu einer Linie flächenhaft verbreitet, die vonder Kirche von Kaltern nach Montiggl zieht. Weiterim S sind diese und vielleicht auch jüngere Ablage-rungen teilweise ausgeräumt und durch ein Systemvon Kiesterrassen ersetzt, die keine Bedeckung vonGeschiebelehmen tragen und während des Rück -schmelzens der Kalterer Zunge entstanden seinmüssen. Es lassen sich hier zumindest drei unter -schiedliche Terrassenniveaus auskartieren und ei-nerseits miteinander, ande rerseits aber auch miteinem System von Trockentälern in Beziehung brin-gen, aus denen diese Kiese offenbar zu unter -schiedlichen Zeiten herausgeschüttet worden sind(Abb. 6). Es gibt auch eine deutliche Beziehung die-ser Terrassen mit dem Kalterer See: Je höher dieseTerrassen liegen, desto weiter liegen sie vom nördli-chen Seeufer entfernt. Die niedrigsten (und ver-mutlich jüngsten) Terrassen liegen dem See amnächsten (Abb. 6).
22..77 TTrroocckkeennttäälleerr
Die gesamte Talung von Eppan-Kaltern wird voneinem ganzen System von tief eingeschnitte nen,breiten, kastenförmigen Trockentälern durchzogen(Abb. 6). Abschnittsweise werden die Trocken tälerauch von heutigen Gewässern benutzt, die die altenTalböden teilweise durch Schwemm fächer verschüt-tet, in einigen Fällen auch ältere Talgenerationenanerodiert und zerstört haben. Die Trockentäler bilden ein mehrfach verzweigtes
Talsystem, dessen Talachsen größtenteils N-S oderNE-SW-orientiert sind. Das größte und am wenig-sten von jüngeren Schwemmfächern auf gefüllteTrockental, das Lavasontal, lässt sich von den Reit-wiesen am Kalterer See über 6 km nach N verfolgen(Abb. 1, 6). Mehrfach zweigen seitlich einmündendeTrockentäler in nordöst licher Rich tung davon ab(Abb. 6), deren Talböden teilweise vom Haupttal un-terschnitten sind. Nördlich des Feld hofes zweigt einbreites Tal in NNW’ Richtung vom Lavasontal ab, dasdurch junge Schwemm fächer teilweise stark aufge-füllt und dadurch undeutlich geworden ist. DiesesTal lässt sich über den alten Bahnhof von Kalternhinaus nach N verfolgen, wo es sich in mehrere Rin-nen aufspaltet. Diese Verzweigung des Trockentalesist teilweise durch die dichte Bebauung, teilweiseaber auch wegen der Erosion durch den Bach ausdem Tröpfeltal undeutlich geworden. Die am weite-sten nach N verfolgbare Rinne dieses Systems istdiejenige, die von Kaltern nach Oberplanitzingzieht, das Oberplanitzinger Trockental (Abb. 6). Folgt man den Tälern aufwärts, steigen sie mit
meist gleich bleibendem Gefälle an, werden un-deutlich und streichen schließlich in die Luft aus,was für Schmelzwassertäler typisch ist. Wenn dieseRinnen abschnittsweise von modernen Gewässernverwendet werden, fließen diese von der Seite zu;die Quellen liegen niemals am Beginn der Rinnen.Besonders schön ist das am schluchtartig einge-schnittenen Oberplanitzinger Trockental zu sehen,das im Dorfzentrum von Oberplanitzing plötzlichundeutlich wird und verschwindet. Auch im N desLavasontales ist das undeutlich Werden und Ver-schwinden der Rinne sehr gut zu beobachten. Die jüngste Terrasse läuft nach S hin, an den
Reitwiesen, auf Seeniveau aus, setzt sich aber nachN hin ins weithin trockene Lavasontal fort, das sicherst 6 km weiter im N bei St. Michael ver liert. DieTrockentäler, die auf die älteste der drei Terrassenauslaufen, Frühlingstalele und Val Fusca, lassen sich
14 Geo.Alp, Vol. 2, 2005

kaum mehr als 1 km nach N verfolgen. Die dazwi-schen liegende Terrasse kor respondiert mit demFondatal und anderen Trockentälern, die weiter imN enden aber nicht so weit zu verfolgen sind, wiedas Lavasontal (Abb. 6). Um die Gesetzmäßigkeitnoch mal auf den Punkt zu bringen: je älter dieTäler sind, desto weniger weit reichen sie nach N,desto höher lag offensicht lich auch der Vorfluter imBereich des Kalterer Sees. Das zuletzt aktive Tal, dasLavasontal, erhielt sein Wasser auch so weit von Nwie kein anderes, der Vorfluter, der das Wasser auf-nahm, war damals schon fast so tief wie der Kalte-rer See. Penck (in Penck & Brückner 1909: 924) nimmt
an, dass der Überlauf eines Stausees bei St. Paulsüber ein „heute trocken daliegendes Tal, das sichöstlich von Kaltern zum Kalterer See zieht“ erfolgtsein soll, also wohl über das Lavasontal. Bei Kalternsoll dieser Ausfluss nach Cas tiglioni & Trevisan(1973: Abb. 26) in einen weiteren, etwas niedrigerliegenden Stausee gemündet haben, der südlich desKalterer Sees vom Etschgletscher abgedämmt wor-den sein soll, also immer noch deutlich höher gele-gen haben muss, als der heutige Seespiegel. Daskann aber nicht sein, wie oben ausführlich darge-legt wurde. Zudem kann diese Annahme nur dieEntste hung eines der Trockentäler erklären, für alleanderen bleibt sie eine Deutung schuldig.Viel plausibler ließen sich sämtliche Beobachtun-
gen interpretieren, wenn man annimmt, dass dieBildung aller Trockentäler und die Entstehung desTerrassensystems am Kalterer See im Zuge des Rück-schmelzens der Kalterer Zunge entstanden sind. Beiden Trockentälern würde es sich demnach um einSystem peripherer und terminaler Rinnen handeln,über die die Schmelzwässer der zurückschmelzen-den Kalterer Zunge abgeflossen sind (Abb. 8). Mitdem Rückschmelzen waren immer neue Täler inFunktion, während andere trocken fielen. Mit demweiteren Rückzug der Zunge nach N, in Richtung St.Michael, war zuletzt nur noch das tiefst gelegeneund die Achse der Talung nachzeichnende Lavason-tal in Funktion. Als die Gletscherzunge schließlichüber den Sattel bei St. Michael zurückgeschmolzenwar, suchten sich die Schmelzwässer neue Wegeund erreichten den Kalterer See nicht mehr (Abb. 8). Wie groß war die Menge des hier erodierten Ma-
terials? Das hängt unmittelbar mit der Frage nachder Dimension dieser Erosionstäler zusammen. DasLavasontal ist über 6 km lang, auf 5 km Länge ist esum 50 bis 75 m tief in die Umgebung eingeschnit-
ten, mit einer Breite des ebenen Talbodens zwischen50 und 110 m. Man kann abschätzen, dass alleine indieser Rinne mindes tens 50 Mill. m3 erodiert undnach S verfrachtet worden sind. Angesichts derGröße der Ero sionstäler und der Menge des in dentief eingeschnittenen Tälern erodierten Materials istes eigentlich unverständlich, dass der kleine KaltererSee nicht schon während des Eisrückzuges zugefülltworden ist. Das Material, das in allen Rinnen zusam-men erodiert worden ist, dürfte aus reichen, umeinen See, der um ein Vielfaches größer ist als derKalterer See, restlos aufzufüllen. Dabei ist nochnicht einmal berücksichtigt, dass die erodierendenSchmelzwässer sicher nicht nur das in den Tälernerodierte, „alte“ Material transportiert haben, son-dern sicher auch vom Eisrand her mit „frischem“Kies, Sand und Schluff überfrachtet waren.Um erklären zu können, warum das Becken des
Kalterer Sees trotzdem nicht aufgefüllt worden ist,benötigt man eine weitere plausible Annahme: DasSeebecken könnte durch eine im See becken lie -gende große Toteismasse, einem abgetrennten Teilder zurückschmelzenden Kalterer Zunge, solangevor dem Sedimenteintrag geschützt worden sein,bis es nicht mehr durch Schmelzwasser erreichtwerden konnte (Abb. 8). Ursprünglich könnte dieseToteismasse auch die weite Senke nördlich des heu-tigen Sees ausgefüllt haben. Die Annahme einersolchen langsam abschmelzenden und immer klei-ner werdenden Toteismasse würde auch zwangloserklären, warum der Vorfluter sich ständig abge -senkt hat (Abb. 8/ 3-5). Bei dieser Annahme hättendie Schmelzwässer einen Teil der mittransportiertenGrob stoffe seitlich um die Toteismasse herumführen und im Etschtal selbst ablagern müssen. Dasaber sollte sich durch entsprechende Boh rungennachweisen lassen.
22..88 SSeeeesseeddiimmeennttee
An einigen Stellen zwischen Eppan und Kalterntreten geschichtete, sandig-schluffige Ablage run -gen auf, die von Penck (in Penck & Brückner 1909:924) als „glaziale Mehlsande“ von St. Pauls bezeich-net wurden. Sie bedecken vor allem den Nordteildes Überetsch, zwischen Unter rain, Frangart und St.Pauls und überlagern hier ältere quartäre Ablage-rungen bzw. Gesteine der Permotrias. Nach Penck(in Penck & Brückner 1909: 924) wurden diese stel-lenweise viele Dekameter mächtigen Sedimente in
15Geo.Alp, Vol. 2, 2005

einem vom Eis aufgestauten See abgelagert. Glei-ches gilt auch für ähnliche Bildungen, die sich öst-lich des Kreither Sattels beiderseits der Laimburgober halb des Etschtales (am Stadlhof) erhalten ge-blieben sind, ein Vorkommen, das von Castiglioni &Trevisan (1973: 19 f.) als das von „Novale al Varco“oder „Maso Stadio“ bezeichnet wird. Ausführlichwerden diese und die glazilakustrinen Sedimentevon St. Pauls durch Castiglioni & Trevisan (1973: 18ff.) beschrieben. Obwohl die Ablagerungen stellen-weise durch Eisauflast etwas verdichtet und durchdas Eis glazialtektonisch teilweise gestört erschei-nen, müssen sie nach Castiglioni & Trevisan (1973:19) ins Spätglazial, also genauer ins Bühl-Stadiumgestellt werden (Hantke 1983: 234).Der Überlauf des Stausees bei St. Pauls soll nach
Penck (in Penck & Brückner 1909: 924) über ein„heute trocken daliegendes Tal, das sich östlich vonKaltern zum Kalterer See zieht“ erfolgt sein, alsowohl über das Lavasontal. Bei Kaltern soll dieserAusfluss nach Castiglioni & Trevisan (1973, Abb. 26)in einen weiteren, etwas niedriger liegenden Stau-see gemündet haben, der südlich des Kalterer Seesvom Etschgletscher abgedämmt worden sein soll.Wie oben schon dargelegt wurde, ist das Lavasontaleher als normales Schmelzwassertal angelegt wor-den und hat, selbst wenn es später als Überlauf füreinen solchen Schmelzwassersee gedient habensollte, jedenfalls nicht in einen größeren Schmelz-wasserstausee im S des Überetsch gemündet. In derUmgebung des Kalterer Sees gibt es, abgesehen vonden Stauseesedimenten östlich des Kreither Sattels,keine See- oder Deltaablagerungen, die die Annah-me eines solchen Sees rechtfertigen würden. Tatsächlich gibt es Hinweise auf einen Stausee im
Becken des Kalterer Sees, der aber deutlich ältersein muss und eher mit dem frühwürmeiszeitlichenEisaufbau des Etschgletschers als mit dessen Rück-schmelzen im Spätglazial etwas zu tun hat. BeimHotel Leuchtenburg in Kreit am Kalterer See sindoberhalb der Straße Aufschlüsse in schluffig-fein-sandigen, feinschichtigen, etwas eisenschüssigenStillwassersedimenten zu finden, die von kaltzeitli-chen, sehr eisrandnah entstandenen, groben Schot-tern überlagert werden. Die feinkörnigen Sedimentesind überkonsoli diert und deshalb mit Sicherheiteisüberfahren. Bei den überlagernden Schotternkönnte es sich um Vorstoßschotter handeln, viel-leicht sind es aber auch Kiese, die zu den Eisrandab-lagerungen von Fuschgalai gehören und somit alsspätglazial einzustufen sind. Stellenweise sind in
diesen Seeablagerungen schlecht erhaltene Pflan-zenreste zu finden, offenbar Abdrücke von Sten-geln, Zweigen und Blättern. Das Einschwemmenvon Pflanzenresten in glaziale Stauseen erscheintim Zuge des Eisaufbaues eher vorstellbar alswährend des Rückschmelzens der Gletscher. Ver -gleichbare Seeablagerungen wurden übrigens auchin einem künstlichen Aufschluss oberhalb eines Ero-sionstales am Westhang des Lavasontales bei Kal-tern beobachtet.
33.. RRüücckksscchhmmeellzzeenn ddeerr KKaalltteerreerr ZZuunnggee ––eeiinn RReekkoonnssttrruukkttiioonnssvveerrssuucchh
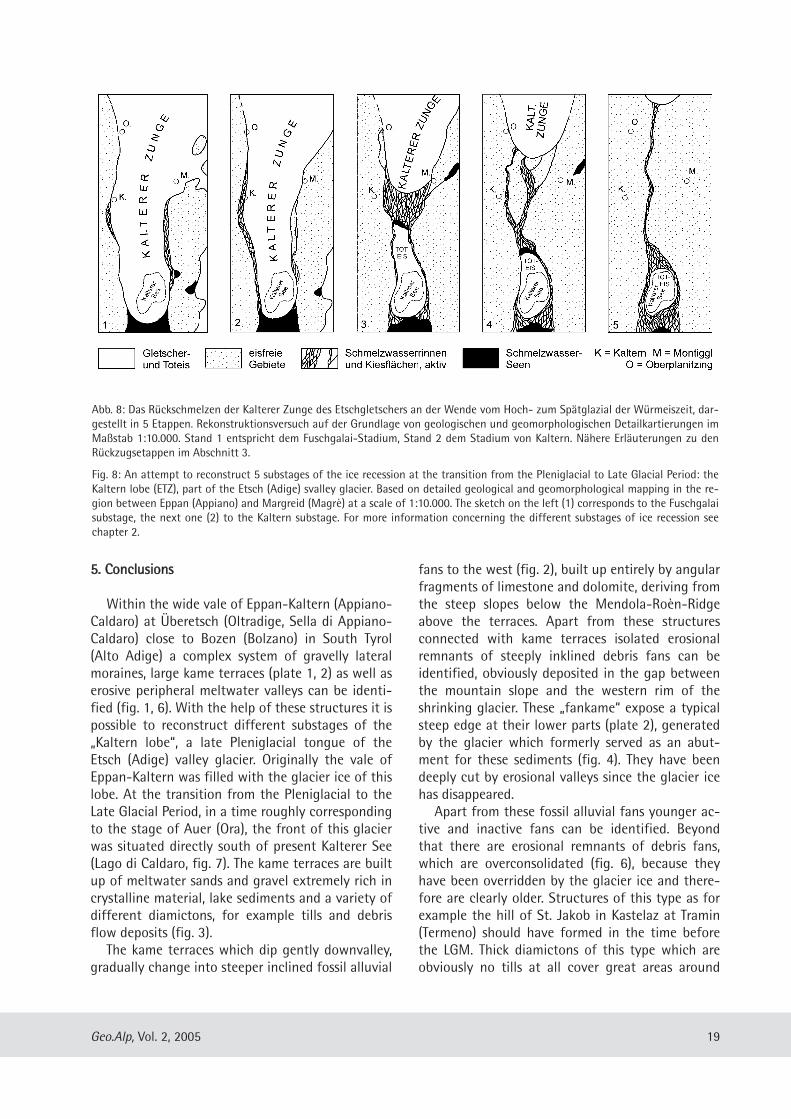
Der hier vorgestellte Rekonstruktionsversuch des„Eisrückzuges“ in der Umgebung von Kaltern(Abb. 8) wurde auf der Grundlage von geologischenDetailkarten erarbeitet, die bei den drei vom Erstau-tor betreuten Kar tierungsübungen mit Geologiestu-denten der TU München in Südtirol entstandenwaren (siehe oben). Die hier dargestellten Rück-zugsstände (Abb. 8/ 1-5) sind wohl mit dem Eis-stand von Auer parallelisierbar, der nach Hantke(1983: 234) demjenigen von Kufstein auf der Al-pennordseite gleichzusetzen sein soll. Nach Jerz(1993: 95) entspricht dies einem Alter von etwa15.000 bis 16.000 Jahren vor heute. Das Rück-schmelzen der Zunge von Kaltern muss also insge-samt im ausgehenden Hochglazial bzw. an derWende zum Spätglazial der Würmeiszeit erfolgtsein. Was man zur Bestätigung der Annahmen undzur Abrundung des Bildes allerdings noch bräuchte,ist die Auswertung von hinreichend tiefen Bohrun-gen in der Talebene südlich des Kalterer Sees.
1. Die Stirn des Etschgletschers ist im Haupttal bisetwa nach Auer zurückgeschmolzen. Ein Seiten ast,die Kalterer Zunge, bedeckt große Teile des Über -etsch, die weite Talung von Eppan-Kaltern undstirnt etwas südlich des Kalterer Sees. Das Etschtalist teilweise von Schmelzwasserseen erfüllt. DieKalterer Zunge wird von Eis genährt, das überTransfluenzen von N her bei Eppan und von NEher über die Montiggler Seen vom Hauptgletscherher überquillt (Abb. 7). In dieser Zeit entstehen diehöchsten Kamesterrassen an der Barleit südlichvon Kaltern (Abb. 6) und die Lateralmoräne vonFuschgalai am Westhang des Unter berges gegenü-ber von Kaltern (Fuschgalai-Stadium, Abb. 6).
16 Geo.Alp, Vol. 2, 2005
2. Der Etschgletscher schmilzt weiter zurück, der Eis-spiegel der Kalterer Zunge sinkt etwas ab. Dergrößte Teil der Kamesterrassen zwischen Kalternund dem Kalterer See entsteht, außer dem Kame-sterrassen unterhalb der Lateralmoräne vonFuschgalai und im Leuchtenburger Wald (Stadiumvon Kaltern, Abb. 6, 7). Beim Absinken des Eisspie-gels werden durch Schmelzwässer parallel zur La-teralmoräne bzw. parallel zur Kamesterrasse süd-lich von Kaltern die Erosions täler des Fuschgalaibzw. am Barleiter Weg eingetieft.
3. Die Kalterer Zunge schmilzt zurück. Durch das Ab-sinken des Eisspiegels dünnt das Eis bei Kaltern soweit aus, dass sich von der Kalterer Zunge einegroße Toteismasse im Kalterer See abtrennt. Zwi-schen der Toteismasse und dem aktiven Eisrandbei Unterplanitzing akkumu lieren flächenhaftKiese, die bei einem kurzen Vorstoß dieser Zungenochmals überfahren werden. Die Schmelzwässerfließen um die Toteismasse herum und mündensüdlich des Kalterer Sees ins Etschtal. Hier entste-hen im Niveau des Etschtales vermutlich Deltakie-se.
4. Mit dem Rückschmelzen der Kalterer Zunge, demallmählichen Kleinerwerden der Toteis masse und
dem dadurch bedingten Tieferlegen des Vorflutersschneiden sich die Schmelz wässer in die zuerst ge-bildeten Kiesflächen ein. In den Rinnen des Lava-sontales, Val Eusca, Frühlingstalele etc. werdenSchmelzwassersedimente erodiert und nördlichder Toteismasse auf tieferen Niveaus erneut abge-lagert. Zunehmend sind weniger Schmelzwasser -rinnen aktiv, am längsten die des Lavasontales unddie tief eingeschnittene Rinne von Oberpla nitzing.Die Schmelzwässer fließen immer noch um dieToteismasse herum und münden südlich des Kalte-rer Sees ins Etschtal.
5. Während die Kalterer Zunge langsam nach Eppanzurückschmilzt, ist zuletzt nur noch die Schmelz-
17Geo.Alp, Vol. 2, 2005
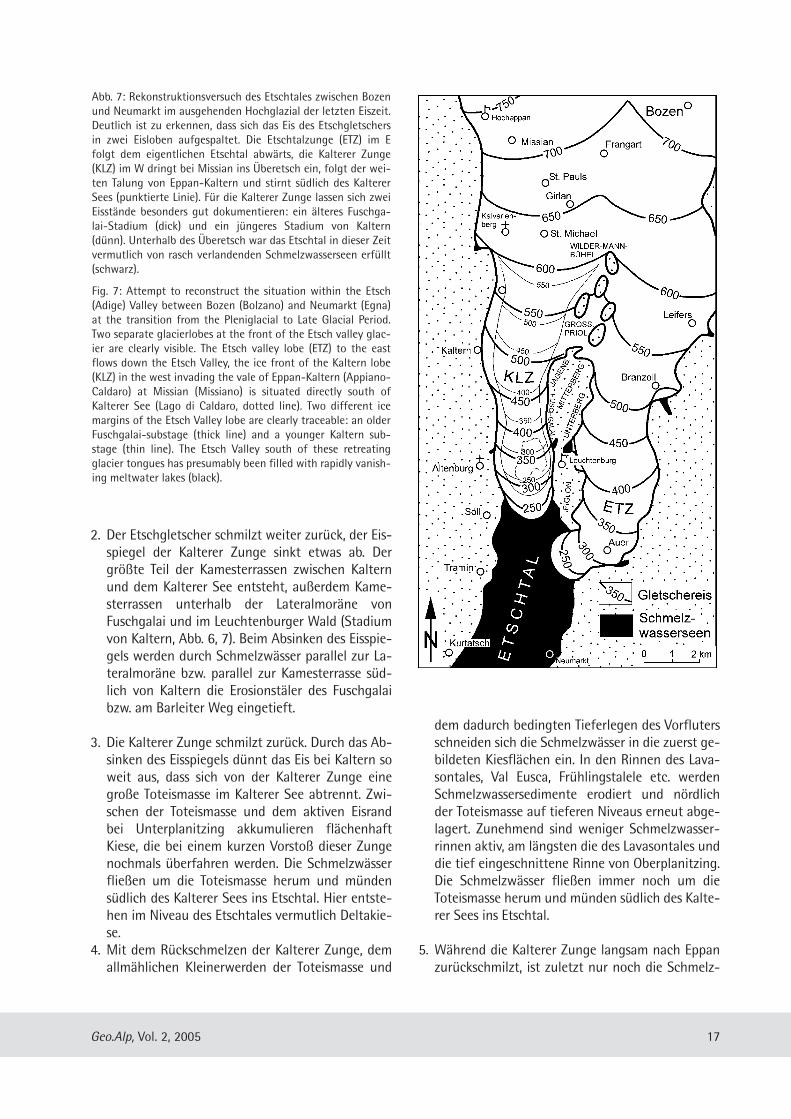
Abb. 7: Rekonstruktionsversuch des Etschtales zwischen Bozenund Neumarkt im ausgehenden Hochglazial der letzten Eiszeit.Deutlich ist zu erkennen, dass sich das Eis des Etschgletschersin zwei Eisloben aufgespaltet. Die Etschtalzunge (ETZ) im Efolgt dem eigentlichen Etschtal abwärts, die Kalterer Zunge(KLZ) im W dringt bei Mis sian ins Überetsch ein, folgt der wei-ten Talung von Eppan-Kaltern und stirnt südlich des KaltererSees (punktierte Linie). Für die Kalterer Zunge lassen sich zweiEisstände besonders gut dokumentieren: ein älteres Fuschga-lai-Stadium (dick) und ein jüngeres Stadium von Kaltern(dünn). Unterhalb des Überetsch war das Etschtal in dieser Zeitvermutlich von rasch verlandenden Schmelzwasserseen erfüllt(schwarz).
Fig. 7: Attempt to reconstruct the situation within the Etsch(Adige) Valley between Bozen (Bolzano) and Neumarkt (Egna)at the transition from the Pleniglacial to Late Glacial Period.Two separate glacierlobes at the front of the Etsch valley glac-ier are clearly visible. The Etsch valley lobe (ETZ) to the eastflows down the Etsch Valley, the ice front of the Kaltern lobe(KLZ) in the west invading the vale of Eppan-Kaltern (Appiano-Caldaro) at Missian (Missiano) is situated directly south ofKalterer See (Lago di Caldaro, dotted line). Two different icemargins of the Etsch Valley lobe are clearly traceable: an olderFuschgalai-substage (thick line) and a younger Kaltern sub-stage (thin line). The Etsch Valley south of these retreatingglacier tongues has presumably been filled with rapidly vanish-ing meltwater lakes (black).

wasserrinne des Lavasontales aktiv. In dem Maßewie sich die Toteismasse im Becken des KaltererSees verkleinert, vergrößern sich die Kiesflächennördlich und südlich davon. Das Eis im Becken desKalterer Sees verschwindet erst, als kein Schmelz-wasser mehr von N her zufließt. Dadurch bleibtein Teil der Hohlform bis heute als See erhalten.Die eis zeitlichen Ablagerungen werden stellenwei-se erodiert, teilweise auch durch junge Schwemm-und Murfächer überdeckt.
44.. SScchhlluussssffoollggeerruunnggeenn
In der weiten Talung von Eppan-Kaltern ist einganzes System von kiesigen Lateralmoränen, breitenKamesterrassen (Taf. 1, 2) und peripheren Rinnenerkennbar (Abb. 1, 6), mit dessen Hilfe sich unter-schiedliche Rückschmelzstadien einer „KaltererZunge“ rekonstruieren lassen. Sie muss während desEisstandes von Auer im ausgehenden Hochglazial inder Talung von Eppan-Kaltern gelegen und knappsüdlich des heutigen Kalterer Sees gestirnt haben(Abb. 7). Die Kamesterrassen bestehen vor allem aussehr kristallinreichen Schmelzwasserkiesen und -sanden, Stillwassersedimenten und zu einem klei-nen Teil auch aus einer Vielzahl von diamiktischenSedimenten, darunter Geschiebelehme (Tille) undMursedimente (Abb. 3). Bergwärts gehen die den Eisrand begleitenden,
leicht nach Süden hin einfallenden Terrassen stel-lenweise tatsächlich in stärker geneigte alluvialeFächer aus karbonatreichem Murschutt über, dervon den Hängen unterhalb des Mendelzugesstammt. Neben Murfächern, die mit diesen Eis -randterrassen direkt verbunden sind (Abb. 2), tretenauch Strukturen auf, die hier „Murkames“ genanntwerden. Es handelt sich um Erosionsreste von starkgeneigten Murfächern, die offen sichtlich direktgegen den absinkenden Eisrand des Etschgletschersgeschüttet wurden. Diese „Murkames“ besitzen aufihrer talwärtigen Seite einen deutlichen Gefälle-knick (Taf. 2), eine Sackungskante, die ihre Entste-hung dem Eisrand verdankt, gegen den die Sedi-mente ursprünglich geschüttet worden waren(Abb. 4). Daneben gibt es auch jüngere, aktive undinaktive Murfächer, aber ebenso Erosionsreste vondeutlich älteren, die offensichtlich vom Eis überfah-ren und dadurch überkonsolidiert sind (Abb. 6).Diese müssen aus der Zeit vor dem Eishöchststandder Würmeiszeit stammen. Stellenweise tritt extrem
matrixarmer Karbonatschutt auf, der bei GraunOberflächenstrukturen zeigt, wie sie für einen (si-cher nicht mehr aktiven) Blockgletscher typischsind (Abb. 1). Ähnliche Ablagerungen sind im Höl-lental und oberhalb Kaltern bei St. Anton durchkarbonatische Zemente zu festen Brekzien verfe-stigt worden. Mursedimente unterschiedlichen Al-ters bedecken in überraschend großer Mächtigkeitweite Flächen an der Ostflanke des Mendelzuges bishinunter ins Tal, besonders in der Umge bung vonKurtatsch. Tille, diamiktische Sedimente (Taf. 4) undBrekzien unterschiedlicher Zusammensetzung undGenese werden im Rahmen dieser Arbeit ausführ-lich beschrieben (Abb. 3). Die gesamte Talung von Eppan-Kaltern wird von
tief eingeschnittenen, breiten, kastenförmigenTrockentälern durchzogen (Abb. 1, 6). Abschnitts-weise werden diese Trockentäler auch von heu tigenGewässern benutzt, die die alten Täler teilweiseanerodiert und zerstört, in einigen Fällen auch mitihren Ablagerungen aufgefüllt haben. DieTrockentäler bilden ein verzweigtes Tal system, dasin südlicher Richtung zum Kalterer See hin entwäs-sert. Das größte und am wenigsten von jüngerenSchwemmfächern zugeschüttete Trockental, das La-vasontal, lässt sich von den Reitwiesen am KaltererSee über 6 km Richtung N bis nach Eppan (St.Michael) verfolgen. Die Talböden der hiervon ab-zweigenden Trockentäler werden teilweise vomHaupttal deutlich unterschnitten. Bei allen diesenTrockentälern handelt es sich um Schmelzwasser-rinnen, die zu einem Zeitpunkt entstanden, als die„Kalterer Zunge“ nach Norden in Richtung Eppanzurückschmolz (Abb. 8). Das komplexe System aus mächtigen Schmelz-
wassersedimenten, erosiven Schmelzwasserrinnen,Kamesterrassen und Lateralmoränen in der Talungvon Eppan-Kaltern lässt sich nur dann zwanglosdeuten, wenn man eine große, langsam abschmel-zende Toteismasse im Gebiet des Kalterer See-beckens annimmt (Abb. 8/ 3-5). Diese Toteismassemuss während des Rückschmelzens des Etschglet-schers dafür gesorgt haben, dass sich im Nordendavon zunächst mächtige Schmelzwassersedimenteakkumulieren konnten (Abb. 8/ 3), die mit demlangsamen Zurückschmelzen des Toteises und demdadurch bedingten Absinken des Vorfluters allmäh-lich wieder ausgeräumt wurden (Abb. 8/ 4). Zudemsorgte sie offensichtlich dafür, dass das Becken desKalterer Sees, trotz erheblichen Sedimenteintrages,nicht restlos aufgefüllt werden konnte.
18 Geo.Alp, Vol. 2, 2005
55.. CCoonncclluussiioonnss
Within the wide vale of Eppan-Kaltern (Appiano-Caldaro) at Überetsch (Oltradige, Sella di Appiano-Caldaro) close to Bozen (Bolzano) in South Tyrol(Alto Adige) a complex system of gravelly lateralmoraines, large kame terraces (plate 1, 2) as well aserosive peripheral meltwater valleys can be identi-fied (fig. 1, 6). With the help of these structures it ispossible to reconstruct different substages of the„Kaltern lobe“, a late Pleniglacial tongue of theEtsch (Adige) valley glacier. Originally the vale ofEppan-Kaltern was filled with the glacier ice of thislobe. At the transition from the Pleniglacial to theLate Glacial Period, in a time roughly correspondingto the stage of Auer (Ora), the front of this glacierwas situated directly south of present Kalterer See(Lago di Caldaro, fig. 7). The kame terraces are builtup of meltwater sands and gravel extremely rich incrystalline material, lake sediments and a variety ofdifferent diamictons, for example tills and debrisflow deposits (fig. 3). The kame terraces which dip gently downvalley,
gradually change into steeper inclined fossil alluvial
fans to the west (fig. 2), built up entirely by angularfragments of limestone and dolomite, deriving fromthe steep slopes below the Mendola-Roèn-Ridgeabove the terraces. Apart from these structuresconnected with kame terraces isolated erosionalremnants of steeply inklined debris fans can beidentified, obviously deposited in the gap betweenthe mountain slope and the western rim of theshrinking glacier. These „fankame“ expose a typicalsteep edge at their lower parts (plate 2), generatedby the glacier which formerly served as an abut-ment for these sedi ments (fig. 4). They have beendeeply cut by erosional valleys since the glacier icehas disap peared. Apart from these fossil alluvial fans younger ac-
tive and inactive fans can be identified. Beyondthat there are erosional remnants of debris fans,which are overconsolidated (fig. 6), because theyhave been overridden by the glacier ice and there-fore are clearly older. Structures of this type as forexample the hill of St. Jakob in Kastelaz at Tramin(Termeno) should have formed in the time beforethe LGM. Thick diamictons of this type which areobviously no tills at all cover great areas around
19Geo.Alp, Vol. 2, 2005
Abb. 8: Das Rückschmelzen der Kalterer Zunge des Etschgletschers an der Wende vom Hoch- zum Spätglazial der Würmeiszeit, dar-gestellt in 5 Etappen. Rekonstruktionsversuch auf der Grundlage von geologischen und geomor phologischen Detailkartierungen imMaßstab 1:10.000. Stand 1 entspricht dem Fuschgalai-Stadium, Stand 2 dem Stadium von Kaltern. Nähere Erläuterungen zu denRückzugsetappen im Abschnitt 3.
Fig. 8: An attempt to reconstruct 5 substages of the ice recession at the transition from the Pleniglacial to Late Glacial Period: theKaltern lobe (ETZ), part of the Etsch (Adige) svalley glacier. Based on detailed geological and geomor pho logical mapping in the re-gion between Eppan (Appiano) and Margreid (Magrè) at a scale of 1:10.000. The sketch on the left (1) corresponds to the Fuschgalaisubstage, the next one (2) to the Kaltern substage. For more information concerning the different substages of ice recession seechapter 2.

Kurtatsch (Cortaccia) and Tramin (Termeno). Manyvineyards are situated on these rigid stony deposits,called „Kampferde“ (which means „soil to fightwith“) by locals. Most of these sediments are pre-sumably debris flow deposits of different ages. In places coarse grained sediments with an ex-
tremely low content of silt and sand occur, consist-ing mostly of angular fragments of carbonate rocks.Close to Graun (Corona) sediments of this typeshow morphological surface structures characteris-tic for rock glaciers (fig. 1). Due to their comparablylow altitude of only 1000 m above sea level thisrock glacier is probably fossil and not active at pre-sent. Similar sediments within the Höllental (Valledel Inferno) and close to St. Anton near Kaltern(Caldaro) were transformed to breccias by carbon-ate cementation. Tills, diamictons and breccias ofdifferent composition, origin and age are men-tioned above in detail (fig. 3).Within the large vale of Eppan-Kaltern a variety
of erosive meltwater valleys can be identified,deeply incised into a thick and complex sequence ofPleistocene sediments, forming a branched fossildrainage system (fig. 1, 6). In places modern creeksuse parts of these valleys and have destroyed themboth, by erosion and infill of sediments. The Pleis-tocene dry valleys mostly show flat bottoms anddrain roughly to the south into the basin of LakeKalterer See. The Lavason Valley is the largest andthe best preserved of these meltwater valleys,tracable from Reitwiesen just north of Lake KaltererSee to St. Michael at Eppan (Appiano) over a dis-tance of 6 km. The bottoms of its tributary valleysare clearly cut by the main valley, and thereforeseem not to have been active for such a long timeas the Lavason Valley itself. All these valleys wereformed by meltwater streams of the „Kaltern lobe“in a later substage (fig. 8), when this glacier tonguemelted slowly back to Planitzing (Pianizza) andEppan (Appiano). The formation of the whole complicated system
of lateral moraines, kame terraces as well as ero sivefossil meltwater valleys within the vale of Eppan-Kaltern, can only be interpretated in a simple andsatisfying way, if a large and slowly vanishing massof stagnant ice is postulated to have existed withinthe basin of Kalterer See (fig. 8/ 3-5). This stagnantglacier ice may have been an obstacle for the melt-waters, streaming from the retreating glaciertongue in the north towards the lake basin in thesouth. North of this hypothetical abutment of stag-
nant ice a thick sequence of glaciofluviatile andglaciolacustrine sediments was accumulated for awhile (fig. 8/ 3). With the slow downmelting of thestagnant ice the meltwater rivers rather began toerode and several generations of erosional drainagesystems were formed here (fig. 8/ 4). The mass ofstagnant ice may also have prevented the lake basinfrom infill of meltwater sediments. OtherwiseKalterer See would not have survived.
LLiitteerraattuurr
Bestle, K.-H. (2005): Geologie der Höhen westlich desEtschtales zwischen Kurtatsch und Fennberg, mit derErstellung eines Gefahrenzonenplanes für diesen TeilSüdtirols.– unveröffentlicht, kombinierte Diplomkar-tierung und Diplomarbeit am Lehrst. f. Ingenieurge-ologie der TU München, 1 geol. Kt. 1 : 10.000; Mün-chen.
Blaas, J. (1892): Beiträge zur Geologie von Tirol. GlazialeAblagerungen bei Meran und Bozen. – Bd. 1, Verh. Geol.Reichsanst., 1892 (8), S. 217-222.
Bosselini, A. (1998): Geologie der Dolomiten.– 191 S.;Athesia-Verlagsanst., Bozen.
Brandner,R. & Mostler, H. (1982): Der geologische Aufbaudes Schlerngebietes und seiner weiteren Umge bung. –Exkursionsführer, Jahrestagung der ÖsterreichischenGeologischen Gesellschaft in Seis am Schlern, 108 S.;Österr. Geol. Ges., Seis.
Castiglioni, G.B., Trevisan, L. (1973): La Sella di Appiano-Caldaro presso Bolzano nel Quaternario.– Mem. I.G.Min. Univ. Padova, 29, S. 2–34.
Costa, J.E. (1984): Physical geomorphology of debrisflows.– In: Costa, J.E. & Fleisher, P.J. [Hrsg.] (1984): De-velopments and applications of geomorphology,S. 268–317; Springer-Verl., Berlin.
Costa, J.E. (1988): Rheologic, geomorphic and sedimento-logic differentiation of water floods, hyperconcentra-ted flows, and debris flows.– In: Baker, V.R., Kochel, R.C..& Patton, P.C. [Hrsg.] (1988): Flood geomorphology,S. 113–122; Springer-Verl., Berlin.
Coussot, P., Meunier, M. (1996): Recognition, classificationand mechanical description of debris flows, Earth-Science Reviews, 40, S. 209–227.
Davies, T.R.H. (1988): Debris flows – a laboratory investiga-tion.– Mitteilungen der Versuchsanstalt für Wasserbau,Hydrologie und Glaziologie an der ETH Zürich, 96,122 S.
Ebers, E. (1972): Das Quartär des Überetsch.– Schlern, 46(3), S. 111–119.
20 Geo.Alp, Vol. 2, 2005

Fuchs, F. (1969): Eine erste 14C-Datierung für das Paudorf-Interstadial am Alpensüdrand.– Eiszeitalter und Gegen -wart, 20, S. 68–71.
Geyer, O.F. (1993): Die Südalpen zwischen Gardasee undFriaul.– Sammlung Geol. Führer, 86, 576 S.; Gebr. Born-traeger-Verl., Stuttgart.
Gwinner, M.P. (1971): Geologie der Alpen.– 477 S.; Stutt-gart (Schweizerbart-Verl.).
Habbe, K.-A. (1969): Die würmeiszeitliche Vergletscherungdes Gardasee-Gebietes. – Freiburger geogr. Arb., 3,S. 1-254.
Haeberli, W. (1996): Gletscherschwund, Permafrostdegra-dation und periglaziale Murgänge im hochalpinen Be-reich.– In: Odsson, B. [Hrsg.] (1996): Instabile Hängeund andere risikorelevante Prozesse, Monte Verità,S. 163–181; Birkhäuser-Verl., Basel.
Hantke, R. (1983): Eiszeitalter. Die jüngste Erdgeschichteder Schweiz und ihre Nachbargebiete.– Bd. 3, 730 S.;Ott-Verl., Thun.
Heissel, W. (1982): Südtiroler Dolomiten.– Sammlg. Geol.Führer, 71, 172 S.; Gebr. Borntraeger-Verl., Stuttgart.
Husen, D. van (1982): Die Ostalpen in der Eiszeit.– 24 S.,1 Kt.; Geol. B.-Anst., Wien.
Jerz, H. (1993): Das Eiszeitalter in Bayern.– In: Geologievon Bayern, Bd. 1, 256 S.; E. Schweizer bart´ scheVerlags buchhandl., Stuttgart.
Johnson, A.M., Rodine, J.R. (1984): Debris flow.– In:Brunsden, D. & Prior, D.B. [Hrsg.] (1984): Slope Instabili-ty, S. 257–361; John Wiley & Sons, Chichester, NewYork etc.
Klebelsberg, R. von (1926): Über die Verbreitung intergla-zialer Schotter in Südtirol.– Zeitschrift für Gletscher -kunde, 14, S. 266–285.
Klebelsberg, R. von (1935): Geologie von Tirol.– 872 S.;Gebr. Borntraeger-Verl , Berlin.
Klebelsberg, R. von (1949): Handbuch der Gletscherkundeund Glazialgeologie.– Bd. 2, Historisch-regionaler Teil,1028 S.; Springer-Verl., Wien.
Moser, H. (1996): Blätterbach in Aldein.– 104 S.; Athesia-Verlagsanst., Bozen.
Penck, A. (1907): Interglaziale Ablagerungen im Etschthal-gebiet.– Z. dt. Geol. Ges., 59, Monatsber., S. 4–5.f.
Penck, A., Brückner, E. (1909): Die Alpen im Eiszeitalter, Bd.3.– 1197 S.; Tauchnitz-Verl., Leipzig.
Scholz, H. (1984): Westgrönland - ein lebendiges Modellfür die Eiszeit im Alpenvorland.– Natur u. Museum, 114(4), S. 89–103.
Scholz, H. (1986): Das Allgäu im Hochglazial – Westgrön-land heute: ein Vergleich. – Ber. Naturwiss. Ver. Schwa-ben, 90 (1), S. 1–26.
Stacul, P. (1980): Eine alte Gehängebrekzie am Mendel-hang oberhalb von St. Nikolaus bei Kaltern. – Schlern,54 (6), S. 289–291.
Werth, K. (2003): Geschichte der Etsch.– 341 S.; TappeinerVerl., Lana.
Willerich, S. (2005): Geologie der Höhen westlich desEtschtales zwischen Tramin und Kurtatsch (Penon),mit der Erstellung eines Gefahrenzonenplanes für die-sen Teil Südtirols. – unveröffentlicht, kombinierte Di-plomkartierung und Diplomarbeit am Lehrst. f. Inge-nieurgeologie der TU München, 1 geol. Kt. 1:10 000;München.
21Geo.Alp, Vol. 2, 2005
Manuscript submitted: August 25, 2004Manuscript accepted: February 2, 2005

TTaaffeelleerrllääuutteerruunnggeenn // EExxppllaannaattiioonn ooff ppllaatteess
1: Blick über die Talung von Eppan-Kaltern nach SE, von der Barleite zum Unterberg. Die begrünte Vereb nungsfläche istdie Kamesterrasse von Kaltern, am Hang des bewaldeten Berges ist die Lateralmoräne von Fusch galai als leicht nachrechts geneigte gerade Linie zu erkennen. Das Tal dahinter ist das Etschtal.
1: View to Unterberg from Barleite in the northwest, across the vale of Eppan-Kaltern (Appiano-Caldaro). The green plainis the kame terrace of Kaltern, the slightly inclined line at the slope of the wooded mountain is the lateral moraine ofFuschgalai. The valley behind that is the Etsch (Adige) Valley.
2: Blick auf Kaltern von S her. Bei der breiten Verebnungsfäche handelt es sich um die Kamesterrasse von Kaltern (vgl. Abb.2).
2: View to Kaltern from the south. The large green plain is the kame terrace of Kaltern (see fig. 2).
3: Blick von der Mendelpassstraße nach S in Richtung St. Nikolaus. Der bewaldete, nach links (E) gleichmäßig geneigteRücken ist der Erosionsrest des „Murkames“ oberhalb von Pfuss, das von rechts (W) vom Hang her gegen den Rand desGletschers geschüttet wurde. Deutlich ist der Gefälleknick an seinem unteren Ende zu erkennen, die Kante, an der dasMurkames ursprünglich ans Eis grenzte (vgl. Abb. 4).
3: View to St. Nikolaus (S. Nicolo) to the south from the road from Eppan (Appiano) to Mendelpass (Passo della Mendola).The wooded hill gently dipping from rigt (W) to left (E) belongs to the „fankame“ above Pfuss, originally supplied withdebris from the slope on the right hand side (W). There is a typical steep edge at its lower end generated by the glacierwhich formerly served as an abutment for these sediments (see fig. 4).
4: Straßenaufschlüsse in karbonatreichen, diamiktischen und nahezu ungeschichteten Mursedimenten an der Straße vonKurtatsch nach Penon. Es sind zahlreiche, größere, eckige Dolomitblöcke zu erkennen.
4: Roadside exposures of diamictic and nearly not stratified debris flow sediments ritch in carbonate fragments, at theroad from Kurtatsch (Cortaccia) to Penon (Penone). Some of the angular dolomite bolders can be identified.
22 Geo.Alp, Vol. 2, 2005

23Geo.Alp, Vol. 2, 2005
1 2
3 4

Geo.Alp, Vol. 2, S. 25–29, 2005
TTHHEE LLAATTEE PPLLEEIISSTTOOCCEENNEE VVEERRTTEEBBRRAATTEE FFAAUUNNAAFFRROOMM AAVVEETTRRAANNAA ((TTAARRAANNTTOO,, AAPPUULLIIAA,, SSOOUUTTHHEERRNN IITTAALLYY)):: PPRREELLIIMMIINNAARRYY RREEPPOORRTT
Raffaele Sardella1,2, Claudia Bedetti1, Luca Bellucci1, Nicoletta Conti1, Danilo Coppola3,Emmanuele Di Canzio2, Marco Pavia4, Carmelo Petronio1,2, Mauro Petrucci1 & Leonardo Salari1
With 3 figures
RRiiaassssuunnttooIn una cava inattiva, nei pressi della cittadina di Avetrana, a Est di Taranto (Puglia, Italia meridionale),
è stata rinvenuta un ricca associazione faunistica a vertebrati contenuta nel riempimento di una cavitàdi origine carsica. Nella cava affiora la formazione delle “Calcareniti di Gravina”, compatta di colore gial-lastro, con ricca malacofauna ed echinidi, ascrivibile al Pleistocene inferiore. A seguito di un saggio discavo condotto nell’ottobre 2003 è stato possibile condurre una prima analisi stratigrafica del riempi-mento carsico, con campionamento dei diversi livelli riconosciuti e recupero di alcuni degli abbondantireperti fossili affioranti. Attualmente i resti di vertebrati recuperati sono conservati presso laSoprintendenza di Taranto. Nel riempimento della fessura carsica indagata sono stati riconosciuti novelivelli fossiliferi e due tasche. Le specie presenti nel deposito sono riferibili al Pleistocene superiore.
AAbbssttrraaccttIn an abandoned quarry near Avetrana (Taranto, Apulia, Southern Italy) a fossiliferous karst filling deposit
rich in vertebrate remains has been discovered. This deposit fills a wide karst fracture crossing a massive, yel-low marine bio-calcarenite termed “Calcareniti di Gravina” Formation. This formation contains a rich macro-fauna and echinids, referable to the Early Pleistocene. In October 2003, field activities including sampling ofthe sediments and a preliminary excavation of the fossiliferous levels started. Within the karst filling depositnine levels and two pockets could be determined. The collected fossils are stored at the “Soprintendenza peri Beni Archeologici per la Puglia” (Taranto, Apulia). This fossiliferous karst deposit can be referred to the Late Pleistocene.
25
1 Dipartimento di Scienze della Terra, Università di Roma “La Sapienza”; e-mail: [email protected] CNR-IGAG Istituto di Geologia Ambientale e Geoingegneria3 Dipartimento di Storia, Università “Tor Vergata” di Roma4 Dipartimento di Scienze della Terra, Università di Torino
IInnttrroodduuccttiioonn
The Salento Peninsula is well known in thepalaeontological literature for its Late Pleistocenevertebrate faunas, mainly in its southern part(Blanc 1920, De Giuli 1983, Corridi 1987, Di Stefanoet alii 1992, Bologna et alii 1994, Rustioni et alii1994 among others). Data available for the Ioniannorthern part of Salento are quite rare. Here we
report the discovery of a new fossiliferous localityin the area of Taranto. It is a karst filling depositrich in fossil bones located in an abandoned cal-carenite quarry, in the area of Avetrana, not farfrom Manduria (Fig. 1).After a preliminary survey during May 2003, car-
ried on by some of the authors (D. Coppola and C.Petronio, in particular), a team of palaeontologistsof the “La Sapienza” University, leaded by Prof.

C. Petronio, started the field cam-paign, in accordance with “Soprin -ten denza per i Beni Archeologiciper la Puglia”. Field work was con-tinued in October 2003, withactivities of sampling sedimentsand a partial excavation of thefossiliferous levels. The collectedfossils are stored at “Soprinten -denza per i Beni Archeologici perla Puglia” (Taranto, Apulia).In the present paper we present
a preliminary analysis of the col-lected material and some generaloutlines of the fossiliferous karstdeposit.
SSttrraattiiggrraapphhyy
The “Calcareniti di Gravina” Formation is a mas-sive bio-calc arenite, rich in molluscs and echinids,that widely outcrops in the central-southernApulian Peninsula; its age spans the Late Pliocene(Adriatic side) and Early Pleistocene (Ionian side)(Ciaranfi et alii 1988). In the considered quarry nearAvetrana this formation is exposed in a sectionwhich is approximately 10 m thick. The vertebratefossil bones occur in a karst fissure filling (Fig. 3).The sediments containing fossil vertebrates aredivided into two parts: the upper part fills the maincavity (layers 1 to 9), the lower part fills a networkof small fissures which opened under the main one.The small fissures (layer 0 in Fig. 3) are filled withorange-yellow nonlaminated sandy clays rich insmall and medium-sized vertebrate remains. Themain cavity is filled with laminated sediments 4,5 to5,5 m thick. From the bottom to the top the follow-ing levels have been determined (Fig 3):
1) 30 cm of clayey sand with rare altered calcare-ous pebbles and bones. A continuous level ofcalcareous pebbles constitutes the basal part ofthe layer.
2) 20 cm of sandy clay very rich in fossil remains.3) 20 cm of clayey sand with some bones and rarecalcareous pebbles.
4) 20 cm of sandy clay rich in fossil bones and cal-careous pebbles. This layer is separated fromlayer 3 by an erosional surface which is markedby a level of calcareous pebbles and bones.
5) 140 cm of clayey sand with very abundant bonesand rare calcareous pebbles. This layers is char-acterised by a level of bones and pebbles at itsbase and also by a sandy lens with rare bones 20cm above the base of this layer.
6) 20 cm of sand with abundant bones and cal-careous pebbles.
7) 40 cm of clay with abundant bones and largecalcareous boulders, especially at the base of thelayer (Fig. 2).
8) 75 cm of clay and bones, the bones are also con-centrated at the base of the layer, separating itfrom the underlying layer.
9) 70 cm of clay with sparse bones, most of themdecalcified.
PPaallaaeeoonnttoollooggyy
Most of the fossil remains found at different lev-els of the main cavity are of medium to large size,mainly referable to Bos primigenius (Fig. 3). The pre-liminary analysis of the fossil material enables us topresent the following faunal list:Layer 0: AVES: Perdix perdix, Columba livia, Athenenoctua, Pyrrhocorax graculus; MAMMALIA:Erinaceus europaeus, Lepus europaeus,Oryctolagus cuniculus, Hystrix cf. H. cristata,Terricola savi, Felis silvestris.
26 Geo.Alp, Vol. 2, 2005
Fig. 1: Location of Avetrana

Layer 1: Bos primigenius.Layer 2: Vulpes vulpes, Canis lupus, Crocuta crocu-ta, Lynx lynx, Stephanorhinus sp., Bos primige-nius, Bovidae indet., Dama dama, Cervus elaphus.
Layer 3: Lepus europaeus, Vulpes vulpes, Canislupus, Bos primigenius, Dama dama, Cervus ela-phus.
Layer 4: Lepus europaeus, Vulpes vulpes, Canislupus, Bos primigenius, Dama dama, Cervus ela-phus.
Layer 5: Vulpes vulpes, Canis lupus, Bos primigenius,Cervus elaphus, Dama dama.
Layer 6: Vulpes vulpes, Canis lupus, Bos primigenius,Cervus elaphus, Dama dama.
Layer 7: Vulpes vulpes, Canis lupus, Crocuta crocu-ta, Stephanorhinus sp., Bos primigenius, Cervuselaphus, Dama dama, ?Megaloceros giganteus.
Layer 8: Vulpes vulpes, Canis lupus, Lynx lynx,Panthera leo, Bos primigenius, Cervus elaphus,Dama dama.
Layer 9: fossil bones are absent.
Layer 0 is characterised by the occurrence ofsmall vertebrate remains. Bird remains are repre-sented by some limb bones, while mammals aremainly represented by teeth. In particular, twolower molar teeth (M2 and M3) of Erinaceuseuropaeus, 7 well preserved mandibles and some M1of Terricola savi, M1 and two M2 of a porcupine,slightly smaller than the living Hystrix cristata, havebeen recorded. Among lagomorphs the hare and therabbit occur with some fragments of skull andmandible and some limb bones. Such taxa are alsorecorded from the main cavity deposit (from layers2 to 8) but are poorly represented. The wild cat isrepresented by fragmentary limb bones of peculiarsize.In the main cavity filling, Bos primigenius is the
best-represented taxon in each fossiliferous level(1–8); layers 5 and 6 are very rich in limb bones, insome cases in anatomical connection. Layer 8 is alsocharacterised by the occurrence of skull fragmentsand mandibles with jugal teeth.Cervids are represented by some isolated teeth
and limb bones (layers 2–8); two large-sized firstphalanxes recorded in level 7 can probably beascribed to Megaloceros giganteus. Among carni-vores, the occurrence of the wolf and the red fox istestified by some isolated teeth, occurring from lay-ers 2 to 8, the lynx and the cave lion are recordedfrom layer 8 (some teeth and a talus respectively).
The occurrence of a rhino is testified in layers 2 and7. This taxon is represented by one fragmentarymolar tooth and one pisiform in every layer.The faunal assemblage on the whole can be
referred to the Late Pleistocene. The occurrence ofthe fallow deer (in particular of the modern sub-species Dama dama dama) and of a rhino, generallyreferable to Stephanorhinus sp., recorded from lay-ers 2 and 7, gives important biochronological con-straints. In fact, the modern fallow deer was wide-spread in Italy at the beginning of the LatePleistocene, while rhinos referable to the genusStephanorhinus survived until the beginning of thePleniglacial (MIS 3) (Gliozzi et alii 1997).At the moment, only general considerations on the
palaeoenvironmental conditions can be pointed out.In layer 0, the occurrence of Terricola savii andHystrix cf. H. cristata suggests the presence of tem-perate climatic conditions with dry and openpalaeoenvironments. Moreover, such a generalframework is supported also by the occurrence of theavifauna including Perdix perdix and Athene noctua,while Columba livia and Pyrrhocorax graculus sug-gest the presence of rocky cliffs. In the sequence fill-ing the main cavity (layer 1 to 8), large mammalspecies of a wider ecological significance occur.Preliminary taphonomical observations indicate
that the fossil bones seem to be not oriented. This
27Geo.Alp, Vol. 2, 2005
Fig. 2: Fossil bones of layer 7 (scale bar: 20 cm).

fact suggests quick deposition of the fossil-bearingsediments, which is also supported by sedimento-logical observations. In fact, the different layers arecharacterised by a normally graded distribution ofthe sediment, with the heaviest material like bigbones and calcareous pebbles concentrated in thelowest part of the layer. Frequently such bones andpebbles constitute a well defined level at the baseof the layer to separate one layer from the underly-ing one.A detailed analysis of the fossil remains has just
begun, in accordance with the “Soprintendenza aiBeni Archeologici per la Puglia”, with the aim ofproviding further palaeontological information anda framework of the palaeoenvironmental evolutionof the area during the Late Pleistocene.
AAcckknnoowwlleeddggmmeennttss
All phases of fieldwork were financially support-ed by CNR – IGAG and by the Municipality ofAvetrana. A special mention is due to Francesco
Nigro, responsible for the Cultural Heritage. Thesupport of Maria Antonietta Gorgoglione, responsi-ble for “Soprintendenza per i Beni Archeologici perla Puglia”, is warmly acknowledged. Moreover, theEarth Science Department of the University of Turinand the “Museo delle Civiltà Preclassiche delleMurge Meridionali” provided facilities for research.We wish to thank Giuseppe “Pippo” Arcidiacono,Francesco Ciminelli, Vincenza Montenegro andMichael Giagnoni for participating the field workand, finally, Karl Krainer and Marzia Breda for theircomments and suggestions on the manuscript.
RReeffeerreenncceess
Blanc, G. A.(1920): Grotta Romanelli. – Arch. Antrop. Etn.,50(1-4): 1-39.
Bologna, P., Di Stefano, G., Manzi, G., Petronio, C.,Sardella, R., Squazzini, E. (1994): Late Pleistocenemammals from the Melpignano (Le) „Ventarole“: pre-liminary analysis and correlations. - Boll. Soc. Paleont.It., 33 (2): 265-274.
28 Geo.Alp, Vol. 2, 2005
Fig. 3: Stratigraphy of the fossiliferous deposit

Ciaranfi, N., Pieri, P., Ricchetti, G. (1988): Note alla CartaGeologica delle Murge e del Salento (Puglia cen-tromeridionale). - Mem. Soc. Geol. It., 41(1): 449-460.
Corridi, C. (1987): Faune pleistoceniche del Salento: 2. Lafauna di fondo Cattìe, Maglie, Lecce. - Quaderni delMuseo Comunale di Paleontologia, 3: 5-74.
De Giuli, C. (1983): Le faune pleistoceniche del Salento: 1.La fauna di S. Sidero 3. - Quaderni del MuseoComunale di Paleontologia, 1: 45-84.
Di Stefano, G., Petronio, C., Sardella, R., Savelloni, V.,Squazzini, E. (1992): Nuove segnalazioni di brecceossifere nella costa fra Castro Marina e Otranto(Lecce). - Il Quaternario, 5 (1): 3-10.
Gliozzi, E., Abbazzi, L., Argenti, P., Azzaroli, A., Caloi, L.,Capasso Barbato, L., Di Stefano, G., Esu, D., Ficcarelli,G., Girotti, O., Kotsakis, T., Masini, F., Mazza, P.,
Mezzabotta C., Palombo, M. R., Petronio, C., Rook, L.,Sala, B., Sardella, R., Zanalda, E., Torre, D. (1997):Biochronology of selected Mammals, Molluscs andOstracods from the Middle Pliocene to the LatePleistocene in Italy. The state of the art. – Riv. Ital.Paleont. Strat., 103(3): 369-388.
Rustioni, M., Mazza, P., Abbazzi, L., Delfino, M., Rook, L.,Petrucci, S., Vianello, F. (1994): The Würmian faunafrom Sternatia (Lecce, Apulia, Italy). - Boll. Soc.Paleont. It., 33 (2): 279-288.
29Geo.Alp, Vol. 2, 2005
Manuscript submitted: December 17, 2004Revised manuscript accepted: February 22, 2005

Geo.Alp, Vol. 2, S. 31–51, 2005
TTHHEE LLAADDIINNIIAANN FFLLOORRAA ((MMIIDDDDLLEE TTRRIIAASSSSIICC)) OOFF TTHHEE DDOOLLOOMMIITTEESS::PPAALLAAEEOOEENNVVIIRROONNMMEENNTTAALL RREECCOONNSSTTRRUUCCTTIIOONNSS AANNDD PPAALLAAEEOOCCLLIIMMAATTIICC CCOONNSSIIDDEERRAATTIIOONNSS
Evelyn Kustatscher1 & Johanna H.A. van Konijnenburg-van Cittert2
With 7 figures and 5 tables
AAbbssttrraaccttThe study of several plant fossils from the Ladinian of the Dolomites, that either had been described a long
time ago or had never been described at all, has led to a revision of this flora. The Ladinian flora now con-sists of the following taxa: Annalepis zeilleri (Lycophyta), Equisetites arenaceus (Sphenophyta), Cladophlebisleuthardtii, C. ruetimeyeri, Neuropteridium elegans, Scolopendrites sp., Gordonopteris lorigae (Pteridophyta),Ptilozamites heeri (Pteridospermae), Bjuvia dolomitica, Dioonitocarpidium moroderi, Pterophyllum jaegeri,?Pterophyllum sp., Sphenozamites wengensis, Sphenozamites cf. bronnii, Taeniopteris sp. (Cycadophyta),Voltzia dolomitica, V. ladinica, V. pragsensis, V. zoldana, Voltzia sp., Pelourdea vogesiaca und Elatocladus sp.(Coniferophyta).The flora, and especially the large number of specimens housed in the Natural History Museum at Bolzano
(I), indicates a dominance of conifers over (in this sequence) seedferns, cycads, ferns and horsetails. Severalfactors may have caused this: climatic (an arid climate on the mainland), edaphic (immature soil) or tapho-nomic (caused by selection during transport). Quantitative palynological analyses of three localities (Ritbergnear Wengen, and Seewald and Innerkohlbach near Prags, indicate a generally warm and humid climate. Thedominance of the conifers and seedferns may, therefore, have been caused by their larger resistance duringtransport rather than by climatic factors.Ladinian palaeoclimatic reconstructions and the plant fossils studies indicate that during the late Ladinian
the Dolomites consisted of carbonate or volcanic islands of various sizes, which were covered with severalbiotopes: coastal and ‚hinterland’; the latter divided into a more humid and a more arid zone.
ZZuussaammmmeennffaassssuunnggDas Studium verschiedener historischer, ebenso wie bisher unbeschriebener Pflanzenfossilien aus dem
Ladin der Dolomiten, die in italienischen und ausländischen Museen aufbewahrt werden, führte zu einigenErstbeschreibungen und systematischen Revisionen. Die Ladinflora setzt sich nunmehr aus folgenden Artenzusammen: Annalepis zeilleri (Lycophyta), Equisetites arenaceus (Sphenophyta), Cladophlebis leuthardtii, C.ruetimeyeri, Neuropteridium elegans, Scolopendrites sp., Gordonopteris lorigae (Pteridophyta), Ptilozamitesheeri (Pteridospermae), Bjuvia dolomitica, Dioonitocarpidium moroderi, Pterophyllum jaegeri, ?Pterophyllumsp., Sphenozamites wengensis, Sphenozamites cf. bronnii, Taeniopteris sp. (Cycadophyta), Voltzia dolomitica,V. ladinica, V. pragsensis, V. zoldana, Voltzia sp., Pelourdea vogesiaca und Elatocladus sp. (Coniferophyta).Die Flora, insbesondere die, die sich im Naturmuseum Südtirol (BZ) befindet, weist eine Dominanz der
Koniferen über Samenfarne, Cycadeen, Farne und Schachtelhalmen auf. Eine derartige Zusammensetzungkann auf verschiedenen Faktoren beruhen: (i) Klimatische (arides Klima auf dem Festland), (ii) edaphische
31
1 Dipartimento di Scienze della Terra, Università degli Studi di Ferrara, C.so Ercole I d’Este 32, 44100 Ferrara, Italy,e-mail [email protected] Laboratory of Palaeobotany and Palynology, Budapestlaan 4, 3584CD Utrecht, Netherlands,e-mail [email protected]

11 IInnttrroodduuccttiioonn
The first Ladinian plant remains from theDolomites have been figured by Wissmann andMünster (1841). Afterwards several authors men-tioned and figured plant fossils from the“Buchensteiner Schichten” and “Wengener Schich -ten” of various areas in the Dolomites (Mojsisovics,1879; Arthaber,1903; Ogilvie Gordon, 1927, 1934;Mutschlechner, 1932; P. Leonardi, 1953, 1968;Calligaris, 1983, 1986; Jung et al., 1992) and fromSappada (G. Leonardi, 1964) (for more detailedinformation see also, Wachtler & van Konijnenburg– van Cittert, 2000a, b; Kustatscher, 1999, 2001,2004). On this account, a high number of differentplant remains have been described from theDolomites at the end of the last century (Table 1).
However, an extended search and study of localand international plant collections with bothalready described and unpublished material fromthe Dolomites provided material for a first reportand some taxonomic revisions of the material(Kustatscher, 2004; Kustatscher et al., 2004).Also several palynological studies have been
applied during the last 25 years regarding succes-sions of Ladinian age from the Dolomites (Cros &Doubinger, 1982; van der Eem, 1982; Blendinger,1988; Roghi, 1995a, 1995b; Broglio Loriga et al.,1999). However, most of the articles use palynolog-ical data only on a biostratigraphic point of view(Blendinger, 1988, Roghi, 1995a, 1995b; BroglioLoriga et al., 1999). Only in one of them (van derEem, 1982) the palynomorphs are considered also asa source for paleoclimatic data.
32 Geo.Alp, Vol. 2, 2005
(unreife Böden) oder auch (iii) taphonomische (Selektion während des Transportes). Die quantitativenAnalysen der Palynofloren der drei Fundorte Ritberg (Wengen), Seewald und Innerkohlbach (Prags) weisenauf ein generell warmes und feuchtes Klima hin. Aus diesem Grund scheint die Dominanz der Koniferen undSamenfarne eher auf die größere Resistenz dieser Pflanzen gegen Zerstörung während des Transports, als aufklimatische Auslese zurückzuführen zu sein.Paläoklimatische Rekonstruktionen aus dem Ladin sowie die Analyse der Pflanzenfossilien weisen darauf
hin, dass die Dolomiten im oberen Ladin von karbonatischen oder vulkanischen Inseln verschiedener Größebedeckt waren, wo sich verschiedene Biotope gebildet hatten: das Küstengebiet und das Hinterland, das sichwiederum in feuchtere und trockener Zonen unterteilen lässt.
RRiiaassssuunnttooRecenti studi sistematici della flora ladinica delle Dolomiti, condotti su collezioni sia storiche che inedite
di musei italiani e stranieri hanno portato a nuove segnalazioni e ad alcune revisioni sistematiche. La floraladinica risulta essere composta dai seguenti taxa: Annalepis zeilleri (Lycophyta), Equisetites arenaceus(Sphenophyta), Cladophlebis leuthardtii, C. ruetimeyeri, Neuropteridium elegans, Scolopendrites sp.,Gordonopteris lorigae (Pteridophyta), Ptilozamites heeri (Pteridospermae), Bjuvia dolomitica, Dioonito -carpidium moroderi, Pterophyllum jaegeri, ?Pterophyllum sp., Sphenozamites wengensis, Spheno zamites cf.bronnii, Taeniopteris sp. (Cycadophyta), Voltzia dolomitica, V. ladinica, V. pragsensis, V. zoldana, Voltzia sp.,Pelourdea vogesiaca ed Elatocladus sp. (Coniferophyta).La flora, in particolare quella depositata nel Museo di Scienze Naturali dell’Alto Adige (BZ), presenta una
generale dominanza delle conifere e pteridosperme, sulle cicadee, felci e sfenofite. Una simile composizionepuò essere imputabile a vari fattori: climatici (aridità delle terre emerse), edafici (suoli immaturi) e tafono-mici (selezione tassonomica causata da un trasporto prolungato). Le analisi quantitative della palinoflora,effettuate nei tre affioramenti di Ritberg (La Valle), Seewald ed Innerkohlbach (Braies), indicano un climacomplessivamente caldo umido. Pertanto, la dominanza delle conifere e pteridosperme sembra essere dovu-ta ad una maggiore resistenza di queste piante ai processi putrefattivi, che avvengono durante il trasporto,piuttosto che a cause climatiche.Sulla base delle ricostruzioni paleogeografiche del Ladinico superiore e sui resti macrofloristici determi-
nati, le Dolomiti presentavano una serie di piccole piattaforme carbonatiche emerse e isole vulcaniche, sullequali si dovevano esistere biotopi differenti: aree costiere, e l’ambiente di entroterra, a sua volta suddivisain zone più umide e più aride.

22 MMaatteerriiaall aanndd mmeetthhooddss
The historical and often unpublished plant fossilcollections are stored in several local and interna-tional museum and universities. In detail, the plantremains figured by Ogilvie Gordon (1927) are keptin the “Paläontologisches Museum” (Munich, D),Mutschlechner’s (1932) material in the “Geologisch-Paläontologisches Institut” of the University ofInnsbruck. The plant fossils discussed in Leonardi(1953) are treasured at the “Museum de Gherdëina”(Ortisei, I) and at the “Museo di Geologia ePaleontologia” of the University of Padova (I). Somespecimens are kept at the “Museo di Paleontologia ePreistoria P. Leonardi” of the University of Ferrara (I)as also the fossil plants from Sappada figured by G.Leonardi (1964) and the plants figured in Leonardi(1968) and Bosellini (1989, 1996). The neuropterid-ian leaf fragment, figured by Zardini (1980) isexposed in the “Museo Paleontologico RinaldoZardini” (Cortina, I). The material discussed and fig-ured by Calligari (1986) is stored in the Museo diScienze Naturali (Trieste, I). Finally, the material dis-cussed by Wachtler & van Konijnenburg – vanCittert (2000a, 2000b) and Kustatscher (1999, 2001,2004 p.p.) is stored in the “Museo di ScienzeNaturali Alto Adige / Naturmuseum Südtirol”(Bolzano / Bozen, I), in the Museum de Gherdëina(Ortisei) and in the “Museo Paleontologico RinaldoZardini” (Cortina). Unfortunately the material men-tioned by Mojsisovics (1879) seems to have beenlost. Additionally unpublished material is stored inthe Museums discussed above and also at theMuseo Ladino Fodom (Livinallongo del Col di Lana,I), the Naturhistorisches Museum, the GeologischeBundesanstalt (Vienna, I) and at the GeologischesLandesamt (Munich, D).
For paleoclimatic considerations palynomorphanalyses have been carried out for 6 samples col-lected at two plant localities near Braies / Prags(Seewald and Innerkohlbach) and one near La Valle(Ritberg), belonging respectively to the upper partof the Fernazza Formation (Ritberg and Seewald)and to the base of the Wengen / La Valle Formation(Innerkohlbach) (see Fig. 1). The samples have been crushed into small frag-
ments and treated with the standard palynologicaltechniques, including HCl (37%), HF (40%) and sat-urated ZnCl2 solution (D ≈ 2,3 g/ml). Afterwards,the slides have been mounted in Canadian balsam.
For the quantitative analyses at least 300 paly-nomorphs have been counted for each sample; thematerial has been divided into the main groups aspollen, spores, fungal remains, algal cysts, acritarchsand foraminiferous lignins. For the quantitativedata the palaeoclimate methods proposed byVisscher & van der Zwan (1981) and Abbink (1998)have been applied. The frequencies of each grouphas been plotted with the aid of a specialised pro-gram, named Graph4win.All the material (macrofossil and palynological)
from the plant localities of Ritberg, Seewald andInnerkohlbach is stored at the Museo di ScienzeNaturali dell’Alto Adige / Naturmuseum Südtirol(Bolzano / Bozen).
33 MMaaccrroofflloorraall ccoommppoossiittiioonn
The Ladinian flora from the Dolomites is composedof the following taxa. The synonymy includes only allreferences from the Ladinian of the Dolomites, notfrom other areas. The localities from which materialhas been recovered, are indicated as well.
33Geo.Alp, Vol. 2, 2005
Fig. 1: Geographic distribution of the studied sections and fos-sil plant localities cited in the article. 1. Prags / Braies, Seewald,Innerkohlbach; 2. Gadertal / Val Badia, Wengen / La Valle, Rit-berg; 3 St. Leonhard in Abtei / S. Leonardo in Badia, St. Kassian/San Cassiano, 4. Grödental / Val Gardena; 5. Pufels / Bulla, Puf-latsch / Bullaccia, Schgaguler Alm / Malga Scagul, Seiser Alm /Alpe di Siusi; 6. Grödner Joch / Passo Gardena, Corvara; 7.Monte Sief, Arabba; 8. Forcella Giau, Corvo Alto, Mondeval; 8.Laste (Livinallongo); 9. Cercenà, Spiz Agnelessa; 10. Sappada.

DDIIVVIISSIIOONN LLYYCCOOPPHHYYTTAAOOrrddeerr IIssooeettaalleessAAnnnnaalleeppiiss zzeeiilllleerrii FFlliicchhee,, 11991100
2004 Annalepis zeilleri Fliche – Kustatscher, p. 157,pl. 10, fig. 1.
2004 Annalepis zeilleri Fliche – Kustatscher et al.,p. 58, pl. 1, fig. 1.
Localities: Wengen / La Valle.
DDIIVVIISSIIOONN SSPPHHEENNOOPPHHYYTTAAOOrrddeerr EEqquuiisseettaalleessFFaammiillyy EEqquuiisseettaacceeaaeeEEqquuiisseettiitteess aarreennaacceeuuss ((JJaaeeggeerr,, 11882277)) SScchheennkk,, 11886644
1999 Equisetites arenaceus - Avanzini & Wachtler,p. 118.
2000aEquisetites arenaceus (Jaeger) Schenk -Wachtler & van Konijnenburg - van Cittert, p.107, pl. 1, fig. 1, 2.
2000bEquisetites arenaceus (Jaeger) Schenk -Wachtler & van Konijnenburg - van Cittert, p.116, pl. 1, fig. 1, 2.
2004 Equisetites arenaceus (Jaeger) Schenk –Kustatscher, p. 158, pl. 10, fig. 2.
Localities: Wengen / La Valle, Sappada.
ccff.. EEqquuiisseettiitteess
1953 Equisetites vel Calamites? – Leonardi, pl. 4,figs. 4–5.
1964 impronta riferibile probabilmente adEquisetale - Leonardi, pl. 5, fig. 10.
1964 frammento di fusto di Equisetale, forseNeocalamites sp. - Leonardi, pl. 5, fig. 11.
2004 cf. Equisetites – Kustatscher, p. 159, pl. 10,fig. 3.
Localities: Pufels / Bulla, Wengen / La Valle,Seiser Alm / Alpe di Siusi, Arabba, Cercenà,Sappada.
DDIIVVIISSIIOONN PPTTEERRIIDDOOPPHHYYTTAAOOrrddeerr FFiilliiccaalleess FFaammiillyy OOssmmuunnddaacceeaaee oorr iinnddeett..CCllaaddoopphhlleebbiiss lleeuutthhaarrddttiiii LLeeoonnaarrddii,, 11995533
1841 Fahrenwedel – Wissmann & Münster, p. 22,pl. 16, fig. 10.
1953 Cladophlebis leuthardti Leonardi, p. 11, pl. 2,figs. 1-5.
1953 Cladophlebis rütimeyeri Heer n.var. heeri –Leonardi, p. 11, pl. 1, fig. 1.
1964 Cladophlebis sp. - Leonardi, p. 201 pl. 5, fig. 7.1968 Cladophlebis cfr. denticulata Brongniart –
Leonardi p. 179, pl. 28, fig. 7.1986 Cladophlebis leuthardti – Calligaris, p. 9, fig.
B29.1993 Cladophlebis leuthardti – Pozzi, p. 82, fig. 103.1998 cf. Pecopteris reticulata (Leuthardt) - Stingl &
Wachtler, p. 82.1999 ?Anomopteris mougeotii Brongniart, 1828 -
Kustatscher, p. 43, pl. 1, fig. B; pl. 2, fig. A.2000a Cladophlebis leuthardtii Leonardi - Wachtler
& van Konijnenburg – van Cittert, p. 109, pl.1, fig. 3.
2000b Cladophlebis leuthardtii Leonardi - Wachtler& van Konijnenburg - van Cittert, p. 117-8, pl.1, fig. 3.
2004 Cladophlebis leuthardtii Leonardi –Kustatscher, p. 160, pl. 10, fig. 5; pl. 11, fig. 1.
Localities: Prags / Braies, Wengen / La Valle, SeiserAlm / Alpe di Siusi, Pufels / Bulla, GrödnerJoch / Passo Gardena, Corvo Alto, Corvara,Monte Sief, Laste (Livinallongo), Cercenà,Sappada.
CCllaaddoopphhlleebbiiss rruueettiimmeeyyeerrii ((HHeeeerr,, 11887777))LLeeoonnaarrddii,, 11995533
1953 Cladophlebis rütimeyeri Heer - Leonardi, p.10, pl. 1, fig. 15, pl. 3 figs. 6.
1953 Cladophlebis sp. - Leonardi, pl. 1 figs. 3-4.1994 Pecopteris – Costamoling & Costamoling, p.
47, fig. 19.2004 Cladophlebis ruetimeyeri (Heer) Leonardi –
Kustatscher, p. 161, pl. 11, fig. 2.Localities: Seiser Alm / Alpe di Siusi, Col Alto,
Cercenà.
NNeeuurroopptteerriiddiiuumm eelleeggaannss ((BBrroonnggnniiaarrtt,, 11882288))SScchhiimmppeerr,, 11886699
1993 Cladophlebis sp. – Pozzi, p. 85, fig. 107.1998 Neuropteridium sp. - Stingl & Wachtler, p. 82.1999 Neuropteridium grandifolium (Schimper et
Mougeot) Schimper - Kustatscher, p. 44, pl. 2,fig. B.
34 Geo.Alp, Vol. 2, 2005
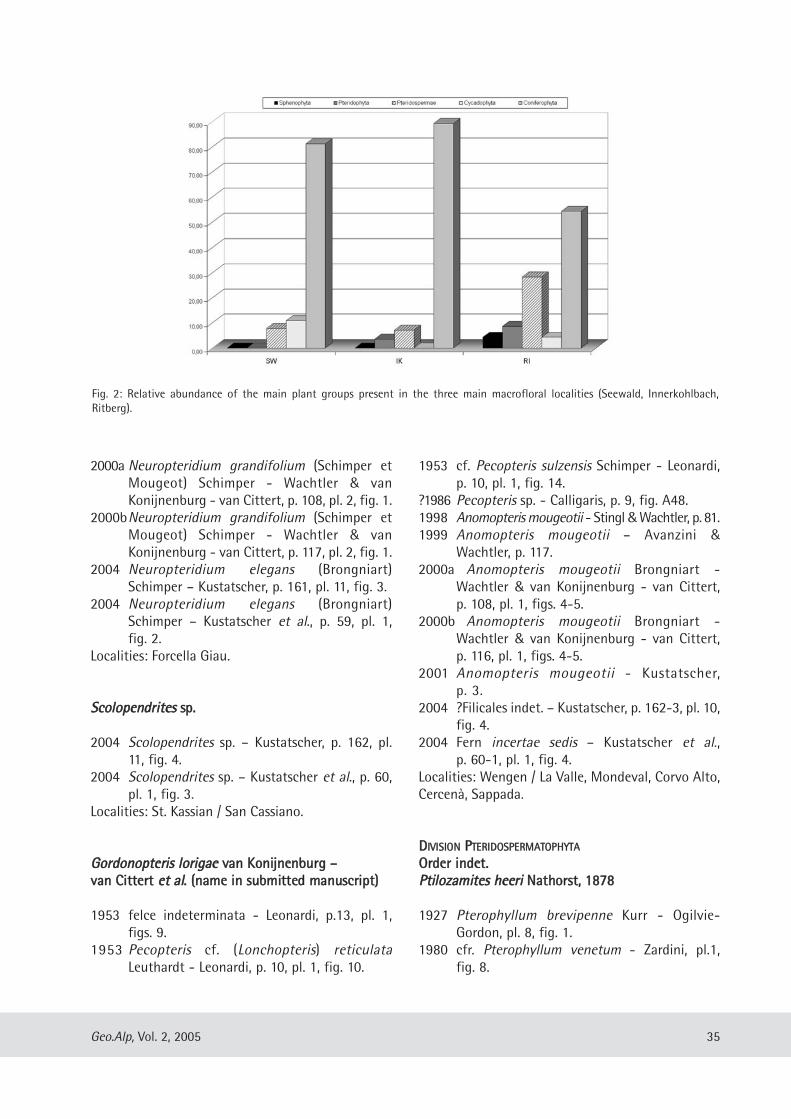
2000a Neuropteridium grandifolium (Schimper etMougeot) Schimper - Wachtler & vanKonijnenburg - van Cittert, p. 108, pl. 2, fig. 1.
2000bNeuropteridium grandifolium (Schimper etMougeot) Schimper - Wachtler & vanKonijnenburg - van Cittert, p. 117, pl. 2, fig. 1.
2004 Neuropteridium elegans (Brongniart)Schimper – Kustatscher, p. 161, pl. 11, fig. 3.
2004 Neuropteridium elegans (Brongniart)Schimper – Kustatscher et al., p. 59, pl. 1,fig. 2.
Localities: Forcella Giau.
SSccoollooppeennddrriitteess sspp..
2004 Scolopendrites sp. – Kustatscher, p. 162, pl.11, fig. 4.
2004 Scolopendrites sp. – Kustatscher et al., p. 60,pl. 1, fig. 3.
Localities: St. Kassian / San Cassiano.
GGoorrddoonnoopptteerriiss lloorriiggaaee vvaann KKoonniijjnneennbbuurrgg ––vvaann CCiitttteerrtt eett aall.. ((nnaammee iinn ssuubbmmiitttteedd mmaannuussccrriipptt))
1953 felce indeterminata - Leonardi, p.13, pl. 1,figs. 9.
1953 Pecopteris cf. (Lonchopteris) reticulataLeuthardt - Leonardi, p. 10, pl. 1, fig. 10.
1953 cf. Pecopteris sulzensis Schimper - Leonardi,p. 10, pl. 1, fig. 14.
?1986 Pecopteris sp. - Calligaris, p. 9, fig. A48.1998 Anomopteris mougeotii - Stingl &Wachtler, p. 81.1999 Anomopteris mougeotii – Avanzini &
Wachtler, p. 117. 2000a Anomopteris mougeotii Brongniart -
Wachtler & van Konijnenburg - van Cittert,p. 108, pl. 1, figs. 4-5.
2000b Anomopteris mougeotii Brongniart -Wachtler & van Konijnenburg - van Cittert,p. 116, pl. 1, figs. 4-5.
2001 Anomopteris mougeotii - Kustatscher,p. 3.
2004 ?Filicales indet. – Kustatscher, p. 162-3, pl. 10,fig. 4.
2004 Fern incertae sedis – Kustatscher et al.,p. 60-1, pl. 1, fig. 4.
Localities: Wengen / La Valle, Mondeval, Corvo Alto,Cercenà, Sappada.
DDIIVVIISSIIOONN PPTTEERRIIDDOOSSPPEERRMMAATTOOPPHHYYTTAAOOrrddeerr iinnddeett..PPttiilloozzaammiitteess hheeeerrii NNaatthhoorrsstt,, 11887788
1927 Pterophyllum brevipenne Kurr - Ogilvie-Gordon, pl. 8, fig. 1.
1980 cfr. Pterophyllum venetum - Zardini, pl.1,fig. 8.
35Geo.Alp, Vol. 2, 2005
Fig. 2: Relative abundance of the main plant groups present in the three main macrofloral localities (Seewald, Innerkohlbach, Ritberg).

1985 Cladophlebis cf. denticulata Brongniart -Moroder, p. 27, fig. 21.
1993 Cladophlebis cfr. denticulata – Pozzi, p. 83,fig. 105.
1999 Ptilozamites heeri - Avanzini & Wachtler, p.118.
2000a Ptilozamites heeri Nathorst - Wachtler & vanKonijnenburg - van Cittert, p. 108, pl. 2, figs.2-9.
2000b Ptilozamites heeri Nathorst - Wachtler & vanKonijnenburg - van Cittert, p. 118, pl. 2, figs.2-9.
2004 Ptilozamites heeri Nathorst – Kustatscher, p.163, pl. 11, fig. 5; pl. 12, fig. 1.
Localities: Prags / Braies, Wengen / La Valle, Gader-tal / Val Badia, Grödental / Val Gardena, CorvoAlto.
DDIIVVIISSIIOONN CCYYCCAADDOOPPHHYYTTAAOOrrddeerr CCyyccaaddaalleessBBjjuuvviiaa FFlloorriinn,, 11993333BBjjuuvviiaa ddoolloommiittiiccaa WWaacchhttlleerr eett vvaann KKoonniijjnneennbbuurrgg --vvaann CCiitttteerrtt,, 22000000
1927 Zamites sp. - Ogilvie-Gordon, p. 68, pl. 8,fig. 4.
1953 Pterophyllum sp. - Leonardi, p. 13, pl. 3, fig. 2.1999 Bjuvia dolomitica Wachtler et van
Konijnenburg - van Cittert (in stampa) -Kustatscher, p. 45, pl. 1, fig. C; p. 49, pl. 4, fig. A.
1999 Bjuvia dolomitica - Avanzini & Wachtler, p.113.
2000a Bjuvia dolomitica Wachtler et vanKonijnenburg - van Cittert, p. 110-111, pl. 4,fig. 1-3; pl. 5, fig. 1-5.
2000bBjuvia dolomitica Wachtler et vanKonijnenburg - van Cittert, p. 120-1, pl. 4, fig.1-3; pl. 5, fig. 1-5.
2004 Bjuvia dolomitica Wachtler et vanKonijnenburg - van Cittert – Kustatscher, p.165, pl. 12, fig. 3.
Localities: Wengen / La Valle, Grödental / ValGardena, Schgaguler Alm / Malga Scagul, Mon-deval.
ccff.. BBjjuuvviiaa
1927 “Zamites sp.“ - Ogilvie-Gordon, p. 68, pl. 8,fig. 4.
1927 Nilssonia sp. - Ogilvie-Gordon, p. 68, pl. 8,fig. 6.
2004 cf. Bjuvia – Kustatscher, p. 165.Localities: Schgaguler Alm / Malga Scagul, GrödnerJoch / Passo Gardena, Corvara, Sappada.
SSpphheennoozzaammiitteess wweennggeennssiiss WWaacchhttlleerr eettvvaann KKoonniijjnneennbbuurrgg -- vvaann CCiitttteerrtt,, 22000000
1999 Sphenozamites - Avanzini & Wachtler, p. 118. 2000a Sphenozamites wengensis Wachtler et van
Konijnenburg - van Cittert, p. 109, pl. 3,figs. 1-2.
2000b Sphenozamites wengensis Wachtler et vanKonijnenburg - van Cittert - Wachtler & vanKonijnenburg - van Cittert, p. 119, pl. 3,figs. 1-2.
2004 Sphenozamites wengensis Wachtler et vanKonijnenburg - van Cittert – Kustatscher,p. 166, pl. 12, fig. 4.
Localities: Prags / Braies, Wengen / La Valle.
SSpphheennoozzaammiitteess sspp.. ccff.. SS.. bbrroonnnniiii ((SScchheennkk))PPaassssoonnii && vvaann KKoonniijjnneennbbuurrgg -- vvaann CCiitttteerrtt,, 22000033
2004 Sphenozamites cf. bronnii (Schenk) Passoni &van Konijnenburg - van Cittert – Kustatscher,p. 166, pl. 13, fig. 2.
2004 Sphenozamites sp. cf. S. bronnii (Schenk)Passoni & van Konijnenburg - van Cittert –Kustatscher et al., p. 62, pl. 2, fig. 2-6.
Localities: St. Leonhard in Abtei / S. Leonardo inBadia, Laste (Livinallongo).
DDiioooonniittooccaarrppiiddiiuumm mmoorrooddeerrii ((LLeeoonnaarrddii))KKuussttaattsscchheerr eett aall..,, 22000044
1953 Cycadeoidea (?) moroderi Leonardi -Leonardi, p. 14, pl. 2, figs. 6-8.
1968 Cycadeoidea (?) moroderi Leonardi -Leonardi, p. 176, pl. 28, fig. 5.
1999 Dioonitocarpidium sp. - Kustatscher, p. 49,58, pl. 3, fig. A-B.
2000a Dioonitocarpidium sp. - Wachtler & vanKonijnenburg - van Cittert, p. 112, pl. 6, fig. 2.
2000bDioonitocarpidium sp. - Wachtler & vanKonijnenburg - van Cittert, p. 123, pl. 6,fig. 2.
36 Geo.Alp, Vol. 2, 2005

2004 Dioonitocarpidium moroderi (Leonardi) novcomb. – Kustatscher, p. 168, pl. 13, fig. 5.
2004 Dioonitocarpidium moroderi (Leonardi) novcomb. – Kustatscher et al., p. 61-2, pl. 2, fig. 1.
Localities: Schgaguler Alm / Malga Scagul.
OOrrddeerr BBeennnneettttiittaalleessPPtteerroopphhyylllluumm jjaaeeggeerrii BBrroonnggnniiaarrtt,, 11882288
1953 Pterophyllum jaegeri Brongniart - Leonardi,p. 13, pl. 2, fig. 12.
1968 Pterophyllum jaegeri Brongniart - Leonardi,p. 176, pl. 28, fig. 4.
1989 Pterophyllum – Bosellini, p. 19, fig. 2.1.1999 Pterophyllum jaegeri - Kustatscher, p. 57, pl.
4, fig. B.1999 Pterophylliium jaegeri - Avanzini & Wachtler,
p. 118.2000a Pterophyllum jaegeri Brongniart - Wachtler
& van Konijnenburg - van Cittert, p. 112, pl.3, figs. 3-4.
2000b Pterophyllum jaegeri Brongniart - Wachtler& van Konijnenburg - van Cittert, p. 122-3,pl. 3, figs. 3-4.
2001 Pterophyllum jaegeri - Kustatscher, p. 6.2004 Pterophyllum jaegeri Brongniart –
Kustatscher, p. 168, pl. 12, fig. 2.2004 Pterophyllum sp. – Kustatscher, p. 169.Localities: Prags / Braies, Wengen / La Valle, St. Kas-sian / San Cassiano, Corvara, Cercenà.
??PPtteerroopphhyylllluumm sspp..
2004 ?Pterophyllum sp. – Kustatscher, p. 170,pl. 13, fig. 3.
Localities: Laste (Livinallongo).
OOrrddeerr iinnddeett..TTaaeenniioopptteerriiss sspp..
1927 Taeniopteris angustifolia Schenk - Ogilvie-Gordon, p.67, pl. 8, fig. 2.
1953 cfr. Taeniopteris sp. - Leonardi, p. 12, pl. I,fig. 18.
1964 Taeniopteris (Nilssonia ?) - Leonardi, pl. 4,fig. 3.
1999 Taeniopteris sp. - Kustatscher, p. 57, pl. 2,fig. C; pl. 3, fig. C.
2000a Taeniopteris sp. - Wachtler & vanKonijnenburg - van Cittert, p. 112, pl. 6, fig. 1.
2000b Taeniopteris sp. - Wachtler & vanKonijnenburg - van Cittert, p. 122, pl. 6, fig.1.
2004 Taeniopteris sp. – Kustatscher, p. 171, pl. 13,fig. 1.
Localities: Prags / Braies, Grödental / Val Gardena,Gadertal / Val Badia, Corvara, Cercená, Sappada.
DDIIVVIISSIIOONN CCOONNIIFFEERROOPPHHYYTTAAOOrrddeerr CCoonniiffeerraalleessEEllaattooccllaadduuss sspp..
1968 Pterophyllum sp. - Leonardi, p. 176, pl. 28, fig.2.
1985 Pterophyllum - Moroder, p. 31, fig. 26.1989 Pterophyllum sp. - Bosellini, p. 89, fig. 12.9.1993 Pterophyllum sp. – Pozzi, p. 85, fig. 108.1996 Pterophyllum - Bosellini, p. 121, fig. 13.8.1999 Elatocladus sp. - Avanzini & Wachtler, p. 119. 1999 Elatocladus sp. - Kustatscher, p. 51, pl. 5,
fig. A.2000a Elatocladus sp. - Wachtler & van
Konijnenburg - van Cittert, p. 113, pl. 6, fig. 3.2000b Elatocladus sp. - Wachtler & van
Konijnenburg - van Cittert, p. 121, pl. 6, fig. 3.2004 Elatocladus sp. – Kustatscher, p. 172, pl. 14,
fig. 2.Localities: Puflatsch / Bullaccia.
PPeelloouurrddeeaa vvooggeessiiaaccaa ((SScchhiimmppeerr eett MMoouuggeeoott,, 11884444))SSeewwaarrdd 11991177
1953 Yuccites vogesiacus Schimper et Mougeot -Leonardi, p.15, pl. 2, fig. 9, 11; pl. 3, figs. 3-4.
1986 Yuccites sp. - Calligaris, p. 15, figs. B21, 42.1999 Yuccites vogesiacus - Avanzini & Wachtler,
p. 119. 2000a Yuccites vogesiacus Schimper et Mougeot -
Wachtler & van Konijnenburg – van Cittert,p. 113, pl. 6, figs. 4, 5.
2000b Yuccites vogesiacus Schimper et Mougeot -Wachtler & van Konijnenburg – van Cittert,p. 121-2, pl. 6, figs. 4, 5.
2004 Pelourdea vogesiaca (Schimper et Mougeot)Seward – Kustatscher, p. 172-4, pl. 13, fig. 4.
2004 Pelourdea vogesiaca (Schimper et Mougeot)Seward – Kustatscher et al., p. 63, pl. 1, fig. 5.
37Geo.Alp, Vol. 2, 2005

Localities: Prags / Braies, Wengen / La Valle, Schga-guler Alm / Malga Scagul.
??PPeelloouurrddeeaa sspp..
1953 Yuccites sp. – Leonardi, pl. 3, fig. 5.2004 ?Pelourdea sp. – Kustatscher, p. 174.Localities: Seiser Alm / Alpe di Siusi, Cercenà.
OOrrddeerr VVoollttzziiaalleessFFaammiillyy VVoollttzziiaacceeaaeeVVoollttzziiaa ddoolloommiittiiccaa WWaacchhttlleerr eett vvaann KKoonniijjnneennbbuurrgg --vvaann CCiitttteerrtt,, 22000000
1927 Voltzia recubariensis Schenk - Ogilvie-Gordon, p. 67, pl. 8, fig. 7.
1932 Voltzia sp. - Mutschlechner, p. 31.1953 Pagiophyllum (?) massalongi Zigno -
Leonardi, p. 18, pl. 3, figs. 8, 10; pl. 4, fig. 2.1968 Brachyphyllum sp. - Leonardi, p. 176, pl. 28, fig. 1.1986 Pagiophyllum cf. massalongi Zigno -
Calligaris, p. 16, figs. A64, B6, B7, B11, B19,B27, B31.
1995 Voltzia recubariensis Schenk - Jung et al., p.171, fig. 8.3.
1999 Voltzia dolomitica - Avanzini & Wachtler,p. 117, 119.
2000a Voltzia dolomitica Wachtler et vanKonijnenburg - van Cittert 2000, p. 113-14,pl. 7, fig. 1-4; pl. 5, fig.1-6.
2000bVoltzia dolomitica Wachtler et vanKonijnenburg - van Cittert - Wachtler & vanKonijnenburg - van Cittert, p. 123-4, pl. 7,fig. 1-4; pl. 5, fig.1-6.
2001 Voltzia dolomitica - Kustatscher, p. 4.2004 Voltzia dolomitica Wachtler et van
Konijnenburg - van Cittert – Kustatscher,p. 175, pl. 14, fig. 1.
Localities: Prags / Braies, Wengen / La Valle, Schga-guler Alm / Malga Scagul, Puflatsch / Bullaccia,Sappada.
VVoollttzziiaa llaaddiinniiccaa WWaacchhttlleerr eett vvaann KKoonniijjnneennbbuurrgg --vvaann CCiitttteerrtt,, 22000000
1999 Voltzia ladinica Wachtler et vanKonijnenburg - van Cittert (in stampa)-Kustatscher, p. 52, pl. 4, fig. C.
2000a Voltzia ladinica Wachtler et vanKonijnenburg - van Cittert, p. 115, pl. 10,figs. 1-5; pl. 11, figs. 1-4
2000bVoltzia ladinica Wachtler et vanKonijnenburg - van Cittert - Wachtler & vanKonijnenburg - van Cittert, p. 125-6, pl. 10,figs. 1-5; pl. 11, figs. 1-4
2004 Voltzia ladinica Wachtler et vanKonijnenburg - van Cittert – Kustatscher,p. 176-7, pl. 14, fig. 3.
Localities: Prags / Braies, Wengen / La Valle, Gröden-tal / Val Gardena.
VVoollttzziiaa pprraaggsseennssiiss WWaacchhttlleerr eett vvaann KKoonniijjnneennbbuurrgg --vvaann CCiitttteerrtt,, 22000000
1953 Pagiophyllum cfr. foetterlei Stur - Leonardi,p.19, pl. 4, fig. 6, 7, 9.
1986 Pagiophyllum cf. foetterlei Stur - Calligaris, p.17, figs. A58.
1998 Voltzia sp. - Stingl & Wachtler, p. 79.1999 Voltzia - Avanzini & Wachtler, p. 119. 2000a Voltzia pragsensis Wachtler et van
Konijnenburg - van Cittert, p. 115, pl. 9, fig.1-2.
2000bVoltzia pragsensis Wachtler et vanKonijnenburg - van Cittert - Wachtler & vanKonijnenburg - van Cittert, p. 125, pl. 9, fig.1-2.
2004 Voltzia pragsensis Wachtler et vanKonijnenburg - van Cittert – Kustatscher, p.177-8, pl. 14, fig. 4.
Localities: Prags / Braies, Wengen / La Valle, Schga-guler Alm / Malga Scagul.
VVoollttzziiaa zzoollddaannaa LLeeoonnaarrddii 11995533
1953 Voltzia zoldana - Leonardi, p. 19, pl. 4, fig. 11968 Voltzia zoldana Leonardi - Leonardi, p. 176,
pl. 28, fig. 3.2004 Voltzia zoldana Leonardi – Kustatscher, p.
178, pl. 14, fig. 5.Localities: Spiz Agnelessa.
VVoollttzziiaa sspp..
1927 Voltzia sp. - Ogilvie Gordon, p. 69, pl. 8, fig. 8.1953 Voltzia sp. - Leonardi, pl. 4, figs. 3, 8.
38 Geo.Alp, Vol. 2, 2005

1953 Pagiophyllum (?) massalongi Zigno -Leonardi, p. 18, pl. 4, fig. 2.
1964 Ramoscello di Brachyphyllum o Pagiophyllumsp. - Leonardi, pl. 4, fig. 4.
1994 Ullmannia Broni – Costamoling &Costamoling, p. 47, fig. 20.
2004 Voltzia sp. – Kustatscher, p. 178.Localities: Prags / Braies, Wengen / La Valle, SeiserAlm / Alpe di Siusi, Pufels / Bulla, Cercenà, Sappada.
44 PPaallaaeeoocclliimmaattiicc ccoonnssiiddeerraattiioonnss
MMaaccrroofflloorraa
Most of the studied plant fossil collections arecomposed of a few specimens only, collected in var-ious and often not well-defined localities. However,the main composition shows a dominance ofconifers, whereas cycads, pteridosperms, ferns andhorsetails occur only occasionally. Only one collec-tion (in Bolzano) is composed of a higher number ofspecimens (more than 150 specimens). Those plantremains have been collected at two plant localitiesnear Braies / Prags (Seewald and Innerkohlbach) andone near La Valle (Ritberg), belonging respectivelyto the upper part of the Fernazza Formation(Ritberg and Seewald) and to the base of theWengen / La Valle Formation (Innerkohlbach). The pollen samples collected at those fossil-bear-
ing horizons, attribute them to the secatus – vigensphase sensu Van der Eem (1982), or to thepseudoalatus-baculatus phase sensu Roghi (1995a,b). Moreover, the plant deposits of Ritberg andInnerkohlbach (Fig. 1) belong to theConbaculatisporites mesozoicus zone sensu Roghi(1995), referred to the upper part of NeumayriSubzone and to the base of Regoledanus Subzone(Protrachyceras Zone, uppermost Longobardian).The ammonoids (Lecanites glaucus, Protrachycerascf. ladinum, cf. Protrachyceras, “Eoprotrachyceras”neumayri, cf. Joannites, cf. Mepinoceras andMegaphyllites sp., det. P. Mietto) collocate thelocalities to the Neumayri Subzone of theProtrachyceras Zone (sensu Mietto & Manfrin,1995). On the other hand, at Seewald no paly-nomorph zonal marker of Roghi’s scale has beenfound. Also the collected ammonoid(Macleanoceras sp., det. P. Mietto) permits to referthe locality only to the Protrachyceras Zone(Longobardian). However, the lithostratigraphic
attribution of the deposit to the FernazzaFormation, narrows its age down to the upperLongobardian (De Zanche et al., 1993) (for moreinformation see also Kustatscher, 2004). The macrofossil collections, discussed already
partly in Kustatscher (1999, 2001, 2004), Wachtler &van Konijnenburg – van Cittert (2002a, 2002b) andKustatscher et al. (2004), permit us to take a closerlook at the quantitative composition of the UpperLadinian macroflora (Fig. 2). All three plant localitiesshow a distinct dominance of the conifers (Voltzia,Pelourdea). Also the pteridosperms (Ptilozamites) arewell represented in all three floras, whereas horse-tails (Equisetites), ferns (Cladophlebis,Gordonopteris) and cycadophytes (Pterophyllum,Sphenozamites and Taeniopteris) are rare and occuroften only in one or two of the plant deposits.
This composition may be due to various factorssuch as climate (aridity), edaphic (immature soils)and taphonomy (i.e. selection due to transport). Conifers are generally referred to arid environ-
ments due to their reduced leaf-surface, the thick-ness of their cuticles and the protection of theirstomata by papillae. On the base of these consider-ations, the composition of the Ladinian Flora fromthe Dolomites might be referred to an arid climatewhich the slightly imbricate pinnules ofCladophlebis might indicate as well. On the other hand, the fossil material is pre-
served within basinal sediments, and therefore, hasbeen subject to selection due to transport previous-ly to its deposition. The high abundance of coniferscompared with the other groups (Innerkohlbachand Seewald above 80%, Ritberg ca. 50%) could bereferred to selection caused by transport, as onlythe more woody and resistant plants preserved afterthe biostratinomic processes. However, the floralcomposition cannot be explained exclusively bymeans of taphonomy. The thickness of the cuticlessuggests also a certain degree of environmentalstress, related to adverse palaeoenvironment. Thiscould correspond to climatic or edaphic conditions.The latter would suggest immature soils and shallowwater level. In this case the papillae on the stomatamight protect the stomata from salted sprays. Onthe other hand, the presence of rare specimens offerns (Cladophlebis, Gordonopteris) and horsetails(Equisetites), suggests the presence of restrictedhumid microenvironments in the terrestrial habitatsas understorey and small ponds.
39Geo.Alp, Vol. 2, 2005

MMiiccrroofflloorraa
The hypothesis of an arid climate during theupper Ladinian is also in conflict with palynologicaldata available from literature. Van der Eem (1982)suggests a progressive increase in humidity duringthe Ladinian, opposed to the arid environmentalconditions at the end of the Anisian. These environ-ments are however considered to be local, due tothe considerable amount of elements derived fromxerophytic plant-communities often present as well(van der Eem, 1982, p. 72).Additionally palynological data are known also
from the plant deposits (Kustatscher, 2004); in thesmall outcrops of Seewald (SW) and Innerkohlbach(IK) one pollen sample each has been studied, whilefrom the more extensive outcrop of Ritberg (RI) foursamples have been analysed.Observing the main groups (spores, pollen grains,
algal cysts, acritarchs), Seewald is clearly dominatedby pollen grains, Innerkohlbach by spores whereasin the Ritberg section an upwards increase of thepollen fraction is observed (Table 2). These quantita-tive palynomorph fluctuations could be interpretedboth as climatic oscillations, and as variations in thedistance between the coast and the marine sedi-mentary environment, caused by sea level changes.
Applying the proposal of Visscher & Van derZwan (1981) for palaeoclimatic analysis, the paly-nomorphs have been divided into 15 groups (Table3). Some of the groups such as A - monolete aca-vate spores, F – Porcellispora complex and J –Samaropollenites complex are absent. Taxa, such asVallasporites ignacii and Enzonalasporites vigens,referred by Visscher & Van der Zwan (1981) and vander Eem (1982) to the vesicate pollen grains (M) arenow attributed to the (proto)monosaccate pollengrains (N). The pollen sample from Seewald (SW) is domi-
nated by the Triadispora complex (L), trilete acavatelaevigate or apiculate spores (B) and alete(proto)bisaccate pollen grains (I). Trilete laevigate orapiculate spores (B), on the other hand, dominatethe Innerkohlbach (IK) sample. This would suggest amore arid climate during deposition of the sedi-ments corresponding with the Seewald plantdeposit, and a more humid climate when theInnerkohlbach flora has been deposited.Trilete laevigate or apiculate spores (B) dominate
also in the Ritberg outcrop. Furthermore, from
the bottom to the top of this section, the Bgroup, while still dominating, decreases in abun-dance. A concomitant increase of the Ovalipolliscomplex (H, especially in RI 3), the Triadisporacomplex (L) and alete (proto)bisaccate pollengrains (I) can be observed. This would suggest anincrease of the aridity from the bottom to the topof the section.
Also Abbink’ s palynomorph quantitative analysis(1998) has been applied to the plant fossil deposits(Table 5). Seewald shows a dominance of theCoastal SEG, whereas Upland, Lowland, River andTidal SEGs are less abundant. At Innerkohlbach, onthe other hand, the more hygrophytic SEGs, such asRiver and Lowland, dominate. However, as there isonly one sample per outcrop, no extended consider-ations can be deduced.More information can be obtained from the
Ritberg section. This outcrop shows an upwardsincrease of the Coastal and Tidal SEGs, while theLowland and Upland SEGs decrease. This trend canbe interpreted as an increase of the distancebetween the coastal line and the area of plantdeposition (a transgression event) and thus it seemsto support that the palynomorph fluctuations maybe mostly due to sea level changes. Observing in detail the Lowland SEG, the most
sensible one to climatic changes (Abbink 1998),almost only taxa considered to be “more humid” canbe distinguished (Table 5). This suggests a prevailinghumid climate during the late Ladinian.
The hypothesis of sea level changes seems to beconfirmed also by the marine palynomorphs.Although acritarchs and algal cysts are only addi-tional elements (less than 20%), the acritarchs, con-sidered as elements of open marine environments,increase from the bottom to the top of the Ritbergsection, while algal cysts decrease (Table 2).
The hypothesis of taphonomic selection interact-ing with the Ladinian macrofloral deposition is sup-ported also by the comparison between the abun-dance of the main groups (divisions) on macrofloris-tic and microfloristic levels (Table 4). The conifers,represented by 50 to more than 80% in themacroflora, never exceed 45% in the microflora(max. 42,3 % at Seewald). Also pollen attributed tothe pteridosperms (2,6-17,9%) and cycads(microflora 0-1,3%) are less abundant than the
40 Geo.Alp, Vol. 2, 2005

macrofloral remains of these groups (respectively7,8-28,3% and 0-10,9%).On the other hand, ferns are much more impor-
tant in the microflora (20,9 – 50,8%) than in themacroflora (0-8,7%), becoming the most importantsporomorph group. This may be due to the highfragility of the pinnate fern leaves, which are easilydestroyed during transport. Considering on theother hand, that spores are generally underestimat-ed in basinal sediments (Neves effect, Chaloner &Muir, 1968) this dominance is even more important.Additionally, the lycophytes are quite abundant
in the microflora with 3,3 to 17,2 %, while only onemacrofloral species attributed to the lycophytes(Annalepis zeilleri) is known from the Ladinian ofthe Dolomites. Spores (especially Uvaesporites),however, are often preserved in tetrads probablydue to environmental stress of the mother-plants(Looy et al., 2001). In any case, this abundance sug-gests that the lycophytes were better represented inthe Ladinian of the Dolomites than suggested bythe macrofloral remains alone. Very abundant is also the genus Ovalipollis,
which botanical attribution is still unknown, as ithas been never found in situ.Observing the separate plant localities in detail
(Table 4), Seewald is dominated by conifers (42,3%),followed closely by ferns (20,9%) and pteridosperms(17,9%). At Innerkohlbach, on the other hand, ferns(50,8%) dominate among the lycophytes (17,1%)and conifers (13,8%). At Ritberg, from bottom totop lycophytes and ferns decrease in number(respectively12,2 - 7,1% and 35,5 – 21,7%), where-as pteridosperms (5 – 13,2%) and conifers increase(19,7 – 31,%).
Concluding, it can be suggested that the plantsgrew in a general warm and humid local climate.The high abundance of conifers and pteridospermsand respectively low abundance of horsetails, fernsand lycophytes in the macroflora seem to be moredue to local edaphic conditions and taphonomicselection than to climate.
55 PPaallaaeeooeennvviirroonnmmeennttaall rreeccoonnssttrruuccttiioonnss
During the late Ladinian, the Southern Alps werecharacterized by wide carbonate platforms bound-ed by more or less extended basins and were locat-ed north of an emerged land now buried under the
Po Plain („Southern Mobile Belt“ of Brusca et al.1981).Following the palaeogeographic reconstructions
of the uppermost Ladinian known from the litera-ture (Assereto et al., 1977; Brusca et al., 1981;Gianolla, 1993; Bosellini, 1996), Ritberg is situatedin a basin surrounded to the west by the carbonateplatforms of Putia / Peitler and Odle / Geißler and tothe northeast by the carbonate platform whichforms today the Piz da Peres. Southwards this basinwas bounded by the carbonate platforms ofSassolungo / Langkofel, Sella, Tofane andMarmolada. Additionally the volcanic complex ofMonzoni and Predazzo were exposed southwards aswell (Fig. 3). Some of these carbonate platforms andthe volcanic complex were subaerically exposedduring the time of deposition of the FernazzaFormation and, therefore, subject to erosion (i.e.Gianolla, 1993). The plant remains could have beentransported from the carbonate islands in thenortheast or west, or together with the volcano -clastic turbidites from the south. Seewald and Innerkohlbach, on the other hand,
are positioned in a basinal environment west of theTre Cime di Lavaredo / Drei Zinnen and east of thePiz da Peres platform. These platforms producedcarbonate sediments, whereas the terrigenousmaterial came from the south, from the volcaniccomplex of Predazzo/Monzoni and perhaps alsofrom source areas more southwards than theValsugana line.
Considering the palaeogeographic reconstruc-tions known from the literature and the paleocli-mate discussed also in this article, the Ladinianplants grew probably on more or less expanded car-bonate or volcanic islands. On these islands variousenvironments developed: the coastal belt and theso-called ‘hinterland’. The latter can be distin-guished in more humid and more arid areas (Fig. 4). The coastal environment (Fig. 5) was occupied
mainly by lycophytes (Annalepis) and pteridospermswith thick cuticles (Ptilozamites). The Annalepisscales were probably inserted on the top of somecentimetres high and thick stems with robust roots(Grauvogel-Stamm & Lugardon, 2001), whereasPtilozamites was likely a shrubby plant, although noreconstruction is so far known for this genus.The hinterland, on the other hand, might have
been composed of ferns (Neuropteridium, Gor don -opteris, Cladophlebis), cycads (Bjuvia, Spheno -
41Geo.Alp, Vol. 2, 2005
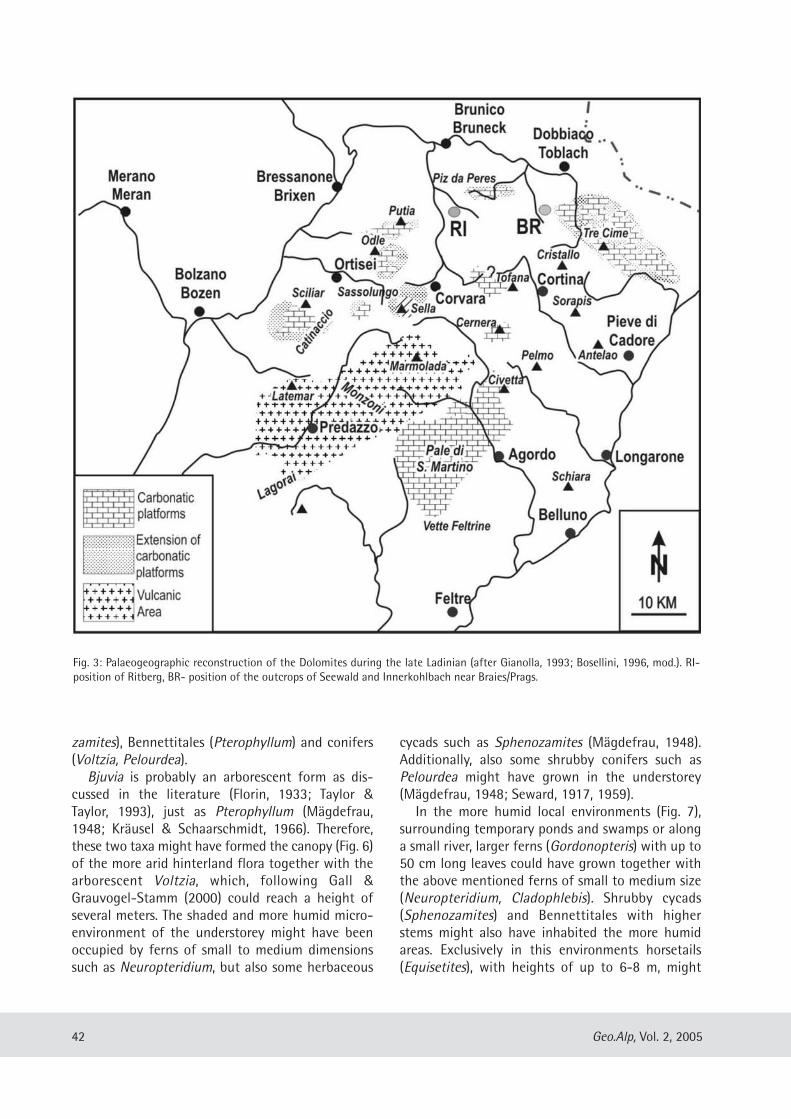
zamites), Bennettitales (Pterophyllum) and conifers(Voltzia, Pelourdea). Bjuvia is probably an arborescent form as dis-
cussed in the literature (Florin, 1933; Taylor &Taylor, 1993), just as Pterophyllum (Mägdefrau,1948; Kräusel & Schaarschmidt, 1966). Therefore,these two taxa might have formed the canopy (Fig. 6)of the more arid hinterland flora together with thearborescent Voltzia, which, following Gall &Grauvogel-Stamm (2000) could reach a height ofseveral meters. The shaded and more humid micro-environment of the understorey might have beenoccupied by ferns of small to medium dimensionssuch as Neuropteridium, but also some herbaceous
cycads such as Sphenozamites (Mägdefrau, 1948).Additionally, also some shrubby conifers such asPelourdea might have grown in the understorey(Mägdefrau, 1948; Seward, 1917, 1959).In the more humid local environments (Fig. 7),
surrounding temporary ponds and swamps or alonga small river, larger ferns (Gordonopteris) with up to50 cm long leaves could have grown together withthe above mentioned ferns of small to medium size(Neuropteridium, Cladophlebis). Shrubby cycads(Sphenozamites) and Bennettitales with higherstems might also have inhabited the more humidareas. Exclusively in this environments horsetails(Equisetites), with heights of up to 6-8 m, might
42 Geo.Alp, Vol. 2, 2005
Fig. 3: Palaeogeographic reconstruction of the Dolomites during the late Ladinian (after Gianolla, 1993; Bosellini, 1996, mod.). RI-position of Ritberg, BR- position of the outcrops of Seewald and Innerkohlbach near Braies/Prags.
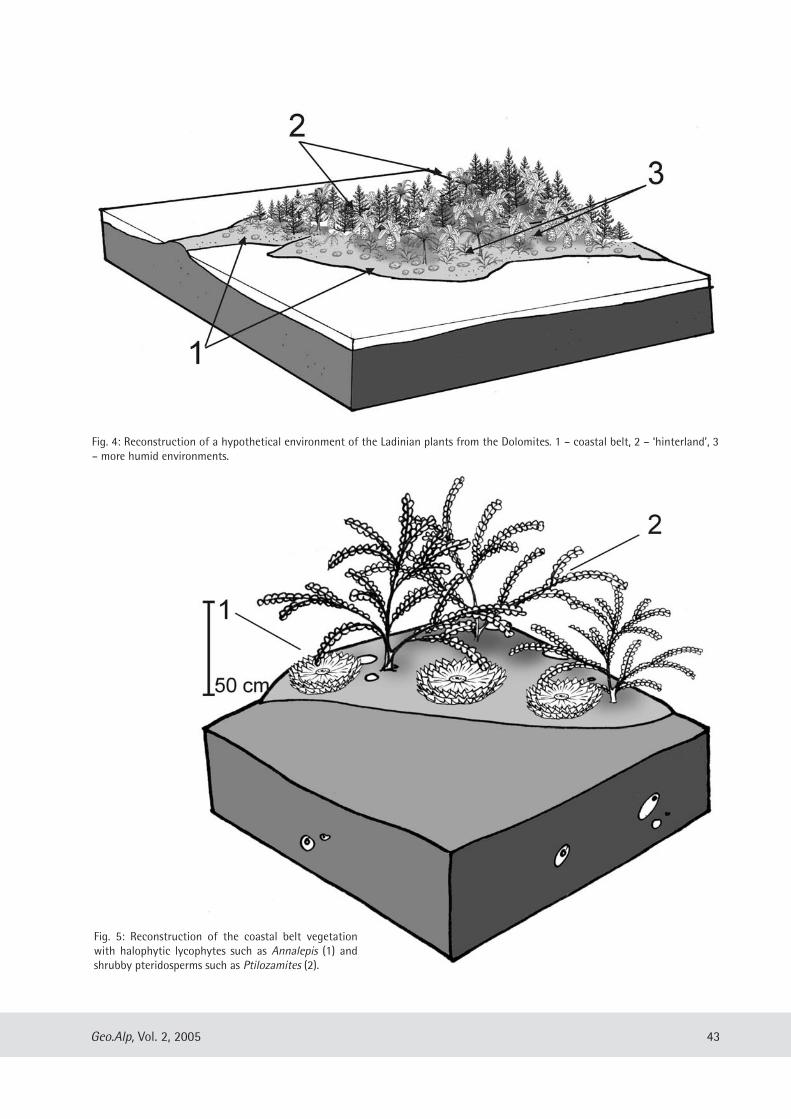
43Geo.Alp, Vol. 2, 2005
Fig. 4: Reconstruction of a hypothetical environment of the Ladinian plants from the Dolomites. 1 – coastal belt, 2 – ‘hinterland’, 3– more humid environments.
Fig. 5: Reconstruction of the coastal belt vegetationwith halophytic lycophytes such as Annalepis (1) andshrubby pteridosperms such as Ptilozamites (2).

have grown as well (Frentzen, 1933; Mägdefrau,1948, 1953; Kelber & Hansch, 1995; Kelber, 1999;Gall & Grauvogel-Stamm, 2000).
66.. DDiissccuussssiioonn
Quantitative variations of organic material (bothplant fossils and palynomorphs) within an outcropdepend on various factors. For those observedbetween the three studied plant deposits two dif-ferent hypotheses have been proposed; climaticoscillations of reduced time extension, or oscilla-tions of the sea level and, therefore, of the relativedistance between the coast line and the point ofdeposition.Throughout the Ritberg section and between the
Seewald and the Innerkohlbach sections, composi-tional variations have been observed (Tables 2-5).
However, the outcrops of Seewaldand Innerkohlbach, since they con-sist of one horizon only, do not per-mit to extrapolate any climatic con-siderations. It is possible that theincrease of the spores and algal cystsand decrease of pollen andacritarchs at Innerkohlbach com-pared to Seewald is due to anincrease of humidity, or an approachof the coastal line to the deposition-al area. The reduction of theacritarchs in favour of the algalcysts, however favours more the sec-ond hypothesis, variations of the sealevel, as would a comparison withthe sequence stratigraphy. TheSeewald outcrop is positioned at thetop of the Fernazza Formation, cor-responding to the HST (HighstandSystems Tract) of the depositionalsequence La3, composed of the basi-nal Zoppè Sandstone, the Acquatonaand the Fernazza Formation and theSciliar 3 platform (De Zanche et al.,1993; Gianolla, 1993). Innerkohl -bach, on the other hand, belongs tothe base of the La Valle / WengenFormation, and is, therefore, corre-sponding to the LST (LowstandSystems Tract) and TST (Transgressive
Systems Tract) of the following depositionalsequence (Car1, sensu De Zanche et al., 1993;Gianolla, 1993), to which also the base of the S.Cassian Formation and the Cassian Dolomite 1 plat-form belong. The lowering of the sea level betweenthese two depositional events could be, therefore,the principal factor of the observed quantitativevariation between these two outcrops.At the outcrop of Ritberg, on the other hand,
the four samples indicate an increase of pollengrains throughout the section (Table 2), and alsoan increase of the Coastal SEG, corresponding to adecrease of the Lowland and River SEGs (Table 5).Also in this case the most accredited hypothesis isa transgression. This hypothesis is confirmed bythe increase of the acritarchs, especially at Rit -berg 4 (Table 2). These (para)autochtonous marinepalynomorphs seem quite sensible to bathy metricand salinity variations, but not to climatic varia-tions.
44 Geo.Alp, Vol. 2, 2005
Fig. 6: Reconstruction of the more arid ‘hinterland’ vegetation with herbaceous(Sphenozamites, 1) and arboreous cycads (Bjuvia, 4), high stemmed Bennettitales(Pterophyllum, 3), shrubby (Pelourdea, 5) and arborescent conifers (Voltzia, 2).

77.. CCoonncclluussiioonnss
The study of historical and ineditedmaterial stored in various collectionsof Italian and international Museumsand Institutions gives new insightsinto the composition of the Ladinianmacroflora of the Dolomites. The palaeoenvironmental recon-
struction based on both macro- andmicrofloral data shows more or lessexpanded carbonate or volcanicislands divided into various environ-ments: the coastal belt and the so-called ‘hinterland’; the latter subdivid-ed into more humid and more aridareas.Additionally, the integrated quanti-
tative analyses (macro- and microflo-ral) suggest that the dominance of theconifers results mostly from tapho-nomic selection. The flora probablygrew under environmental stress dueto salted spray, immature soils andshallow water level, but in a locallyhumid climate. Quantitative palynological analysis
suggests also that the variations infrequency between spores and pollenor algal cysts and acritarchs are prob-ably closer related to sea level changesthan to climatic changes. At presentthe limited extensions of the fossilhorizons do not permit to ex clude thepossibility of climate changes.
AAcckknnoowwlleeddggmmeennttss
The systematic revision would not have been pos-sible withouth the assistence of the various muse-ums and institutions visited by one of the authors,particularily by B. Baumgarten from theNaturmuseum Bozen / Museo di Scienze NaturaliAlto Adige (Bolzano), the family Moroder from theMuseum de Gherdëina (Ortisei), P. Fedele and A.Menardi from the Museo Paleontologico “R. Zardini”(Cortina), F. Deltedesco from the Museo LadinoFodom (Livinallongo del Col di Lana), R. Pancaldifrom the Museo di Paleontologia e Preistoria P.Leonardi (University of Ferrara), M. Fornasiero from
the Museo di Geologia e Paleontologia (Universityof Padova), W. Resch and R. Brandner from theGeologisch-Paläontologisches Institut (University ofInnsbruck), H.A. Kollmann from the Natur -historisches Museum, F. Stojaspal from theGeologische Bundesanstalt (both Vienna), H. Mayrfrom the Paläontologisches Museum and T. Sperlingfrom the Geologisches Landesamt (both Munich).Alberto Riva and Stefano Furin assisted during the
field work, Paolo Mietto determinated theammonoids found in the plant localities. We are par-ticulary thankful to Renato Posenato and GuidoRoghi for ample discussions which permitted toimprove noticable the PhD-thesis on which this workis based. The paleoenvironmental reconstructionshave been drawn by Mattia Guberti.
45Geo.Alp, Vol. 2, 2005
Fig. 7: Reconstruction of the more humid flora of the ‘hinterland’ environmentwith high stemmed Bennettitales (Pterophyllum, 3), arboreous horsetails (Equise-tites, 4) and herbaceous ferns (Neuropteridium, 1; Gordonopteris, 2) and cycads(Sphenozamites, 5).

This work was supported by the “Progetto GiovaniRicercatori 2001” with the titel “The terrestrial florafrom the Middle Triassic of the Dolomites: system-atic, biostratigraphy and palaeoclimate”.
RReeffeerreenncceess
Abbink, O.A. (1998): Palynological identification in theJurassic of the North sea region. – PhD-tesis, 191 pp.,Univ. Utrecht.
Arthaber von, G. (1903): Die alpine Trias des Meditterran-Gebietes. – In: Frech, F. (ed.): Lethaea geognostica.Handbuch der Erdgeschichte, pp. 223-472, Verlag derE. Schweizerbart’schen Verlagsbuchhandlung, Stutt-gart.
Assereto R., Brusca A., Gaetani M., Jadoul F. (1977): Lemineralizzazioni Pb-Zn nel Triassico delle Dolomiti:quadro geologico ed interpretazione genetica. – L’In-dustria Mineraria, 28: 367-402.
Avanzini, M., Wachtler, M. (1999): Dolomiten, Reisen indie Urzeit - 150 pp., Verlagsanstalt Athesia Ges.m.b.H., Bolzano.
Blendinger, E. (1988): Palynostratigraphy of the Late Ladi-nian and Carnian in the south-eastern Dolomites, Italy.– Rev. Paleobot. Palynol., 53: 329-348.
Bosellini, A. (1989): La storia geologica delle Dolomiti. –148 pp., Ed. Dolomiti, Pordenone.
Bosellini, A. (1996): Geologia delle Dolomiti. – 192 pp., Ver-lagsanstalt Athesia Ges. m.b.H., Bozen/Bolzano.
Broglio Loriga C., Cirilli S., De Zanche V., Di Bari D., Gia-nolla P., Laghi G. F., Lowrie W., Manfrin S., MastradreaA., Mietto P., Muttoni G., Neri C., Posenato R., Reichi-chi M., Rettori R., Roghi G. (1999): The Prati di Stuo-res/Wiesen Section (Dolomites, Italy): a candidate glo-bal Stratotype section and Point for the base of theCarnian stage. - Riv. It. Paleont. Strat., 105/1: 37-78.
Brusca C., Gaetani M., Jadoul F., Viel, G. (1981): Paleogeo-grafia ladino-carnica e metallogenesi del Sudalpino. -Mem. Soc. Geol. It. 22: 65-82.
Calligaris, R. (1983): Resti vegetali in terreni ladinicodella Val di Braies. – Tesina in Paleobotanica, 6 pp.,Univ. Trieste.
Calligaris, R. (1986): Geologia della Val di Braies e seg-nalazione di nuovo località fossilifere a vegetali nelLadinico superiore. – Atti Museo Civ. St. Nat., 24/1: 1-64.
Chaloner, W.G., Muir, M. (1968): Spores and flora.– In:Murchison, Westoll (eds.): Coal and Coal-bearingstrata, p. 127-146, Edinburgh.
Costamoling, H., Costamoling, H. (1994): Fossilien desGadertales. – 111 pp., Verlagsanstalt Athesia Ges.m.b.H., Bolzano.
Cros, P., Doubinger, J. (1982): Etudes palynologiques desediments terrigénes et pelagiques du Trias Moyen desDolomites italiennes, relation avec le paléoenvironne-ment. – Sci. Geol. Bull., 35: 157-182.
De Zanche V., Gianolla P., Mietto P., Siorpaes C., Vail P. R.(1993): Triassic sequence stratigraphy in the Dolomi-tes (Italy). - Mem. Sci. Geol., 45: 1-27.
Florin, C. (1933): Studien über die Cycadales des Meso-zoicums nebst Erörterungen über die Spaltöffnungs-apparate der Bennettitales. – Kungl. Svensk. Akad.Handl., ser. 3, 12(5): 1-119.
Frentzen, K. (1933): Equisetaceen des germanischen Keu-per. – Palaeont. Z., 15/1-4: 1-355.
Gall, J.C., Grauvogel-Stamm, L. (2000): L’Arenaria(«GRES») a Voltzia. – In: Pinna, G. (ed.): Alle radici dellastoria naturale d’Europa, seicento milioni di anni at-traverso i grandi giacimenti paleontologici, pp. 71-77,Jaca Book, Milano.
Gianolla, P. (1993): Le successioni stratigrafiche ladinico-carniche nel Sudalpino orientale. – Unpubl. PhD. the-sis, 199 pp., Univ. Padova.
Grauvogel-Stamm, L., Lugardon, B. (2001): The TriassicLycopsids Pleuromeia and Annalepis: Relationships,Evolution, and Origin. – Amer. Fern J., 91(3): 115-149.
Jung W., Selmeier A., Schaarschmidt F., Velitzelos E.,Dernbach U. (1995): Versteinerte Wälder, Ed. D’Oro,188 pp., Heppenheim.
Kelber, K.-P. (1999): Neue Befunde über die Schachtel-halme des Keupers. – In: Hauschke, N., Wilde, V. (eds.):Trias – eine ganze neue Welt, pp. 355-370, Verlag Dr.Friedrich Pfeil, München.
Kelber, K.-P., Hansch, W. (1995): Keuperpflanzen. Die Ent-rätselung einer über 200 Millionen Jahre alten Flora. –Museo, 11: 1-157.
Kräusel, R., Schaarschmidt, F. (1966): Die Keuperflora vonNeuewelt bei Basel. IV. Pterophyllen und Taeniopteri-den. – Schweiz. Paläontol. Abh., 84: 1-64.
Kustatscher, E. (1999): La Flora Ladinica (Triassico Medio)delle Dolomiti. – tesi di laurea AA 1998-1999, 95 pp.,Univ. Ferrara.
Kustatscher, E. (2001): Flora continentale del Ladinicodelle Dolomiti. – PaleoItalia, 4: 2-7.
Kustatscher, E. (2004): Macroflore terrestri del TriassicoMedio delle Dolomiti e loro inquadramento biocrono-stratigrafico e paleoclimatico mediante palinmorfi. –unpubl. PhD-Thesis, 220 pp., Univ. Ferrara.
Kustatscher, E., Wachtler, M., Van Konijnenburg-Van Cit-tert, J.H.A. (2004): Some additional and revised taxa
46 Geo.Alp, Vol. 2, 2005

from the Ladinian Flora of the Dolomites, NorthernItaly. – Geo.Alp 1: 57-70, Bolzano/Bozen.
Leonardi, G. (1964): Note stratigrafico-sedimentologichesul Ladinico della conca di Sappada (Belluno). – Ann.Univ. Ferrara (n. s.), sez. 9, Sci. Geol. Mine., 3 (10): 187-209.
Leonardi, P. (1953): Flora continentale Ladinica delle Do-lomiti. - Mem. Ist. Geol. Min., 18: 1-20.
Leonardi, P. (1968): Le Dolomiti. Geologia dei monti traIsarco e Piave. – 1019 pp., Ed. Manfrini, Rovereto.
Looy C.V., Twitchett R.J., Dilcher D.L., van Konjinenburg -van Cittert J.H.A., Visscher H. (2001). Life in the end-Permian dead zone. - Proc. Nat. Acad. Sci. U.S., 98(14):7879-7883.
Mägdefrau, K. (1948): Vegetationsbilder der Vorzeit. –18 Tafeln, Gustav Fischer Verlag, Jena.
Mägdefrau, K. (1953): Paläobiologie der Pflanzen. – 549pp., Gustav Fischer Verlag, Jena.
Mietto, P., Manfrin, S. (1995): A high resolution MiddleTriassic ammonoid standard scale in the Tethys Realm.A preliminary report. – Boll. Soc. Géol. Fr., 166/5: 539-563.
Mojsisovics, E. (1879): Die Dolomit-Riffe von Südtirolund Venetien: Beiträge zur Bildungsgeschichte derAlpen. - 551 pp., A. Hölder Verlag, Wien.
Moroder, R. et al. (1985). L’Museo de Gherdëina – IlMuseo della Val Gardena, Panoramica dell’ambientenaturale, della preistoria e dell’arte. - Ed. Museo dellaVal Gardena.
Mutschlechner, G. (1932). Geologie der St. Vigiler Dolo-miten. - Jb. Geol. Bundesanst., 82: 163-273.
Ogilvie Gordon, M. M. (1927): Das Grödner-, Fassa- undEnneberggebiet in den Südtiroler Dolomiten. – Abh.Geol. Bundesanst., 24(2): 1-376.
Ogilvie Gordon, M. M. (1934): Geologie von Cortinad’Ampezzo und Cadore. – Jahrb. Geol. Bundesanst., 84:60-215.
Pozzi, E. (1993): Die Fossilien der Dolomiten - 175 pp.,Tappeiner Verlag, Lana.
Roghi, G. (1995a): Analisi palinologica del Trias medio delSudalpino. – PhD thesis, pp. 121, Univ. Padova.
Roghi, G. (1995b): Analisi palinologica della sezione diStuores Wiesen (Prati di Stuores, Dolomiti): dati preli-minari. – In: Loriga Broglio, C., Neri, C. (eds.): Versouna definizione del limite Ladinico-Carnico nel Sudal-pino orientale. - Ann. Univ. Ferrara, 5 (suppl.): 37-40.
Seward, A.C. (1917): Fossil plants, III Pteridospermae, Cy-cadofilices, Cordaitales, Cycadophyta. – 559 pp., Uni-versity Press, Cambridge.
Seward, A.C. (1959): Plant life through the ages – A geo-logical and botanical retrospect – 603 pp., Hafner Pu-blishing Co., New York.
Stingl, V., Wachtler, M. (1998): Dolomiten, Das Werdeneiner Landschaft. - 147 pp., Verlagsanstalt ArthesiaGes. m.b.H., Bolzano.
Taylor, T.N., Taylor, E.L. (1993): The Biology and Evolutionof fossil Plants. – 982 pp., Prentice Hall, New York.
Van der Eem, J.G.L.A. (1982): Palynological investigationsin the Ladinian and Lower Karnian of the Western Do-lomites, Italy. – PhD thesis, 105 pp., Univ. Utrecht.
Visscher, H., Van der Zwan, C.J. (1981): Palynology of thecircum-Mediterranean Triassic: phytogeographicaland palaeoclimatological implications. – Geol.Rundsch., 70(1-2): 625-634.
Wachtler, M., van Konjinenburg van Cittert, J.H.A.(2000a): The fossil flora of the Wengen Formation (La-dinian) in the Dolomites (Italy). – Beitr. Paläont., 25:105-141.
Wachtler, M. & van Konjinenburg van Cittert, J.H.A.(2000b): La flora fossile della Formazione di La Valle –Wengen (Ladinico) nelle Dolomiti (Italia). – StudiTrent. Sci. Nat., 75 (1998): 113-146.
Wissmann, H.L., Münster, G. Graf von (1841): Beiträge zurGeognosie und Petrefactenkunde des südöstlichen Ti-rol’s vorzüglich der Schichten von St. Cassian. – pp.152, Buchner’sche Buchhandlung, Bayreuth.
Zardini, R. (1980): Geologia e fossili attorno a Cortinad’Ampezzo. – 26 pp., Ed. Ghedina, Cortina d’Ampezzo.
47Geo.Alp, Vol. 2, 2005
Manuscript submitted: August 25, 2004Manuscript accepted: March 11, 2005
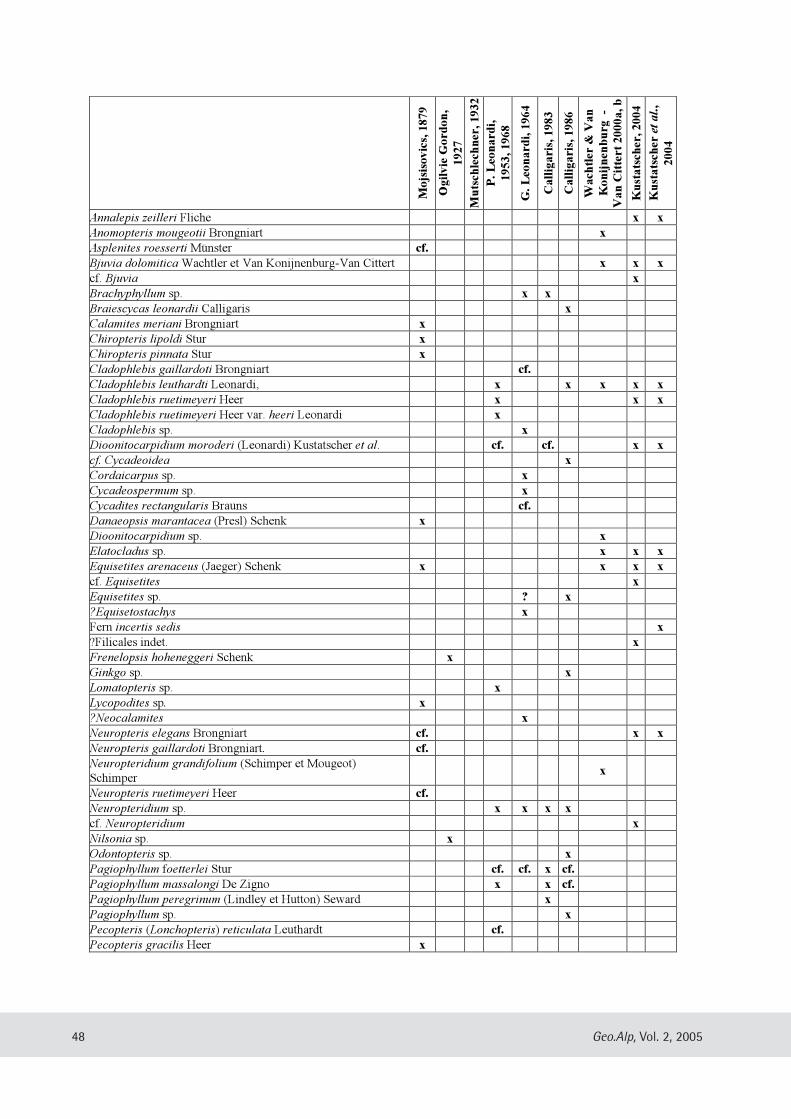
48 Geo.Alp, Vol. 2, 2005
Mo
jsis
ov
ics,
18
79
Og
ilv
ie G
ord
on
,
19
27
Mu
tsch
lech
ner
, 1
93
2
P. L
eon
ard
i,
19
53
, 1
96
8
G. L
eon
ard
i, 1
96
4
Ca
llig
ari
s, 1
98
3
Ca
llig
ari
s, 1
98
6
Wa
chtl
er &
Va
n
Ko
nij
nen
bu
rg -
Va
n C
itte
rt 2
00
0a
, b
Ku
sta
tsch
er, 2
00
4
Ku
sta
tsch
er e
t a
l.,
20
04
Annalepis zeilleri Fliche x x
Anomopteris mougeotii Brongniart x
Asplenites roesserti Münster cf.
Bjuvia dolomitica Wachtler et Van Konijnenburg-Van Cittert x x x
cf. Bjuvia x
Brachyphyllum sp. x x
Braiescycas leonardii Calligaris x
Calamites meriani Brongniart x
Chiropteris lipoldi Stur x
Chiropteris pinnata Stur x
Cladophlebis gaillardoti Brongniart cf.
Cladophlebis leuthardti Leonardi, x x x x x
Cladophlebis ruetimeyeri Heer x x x
Cladophlebis ruetimeyeri Heer var. heeri Leonardi x
Cladophlebis sp. x
Dioonitocarpidium moroderi (Leonardi) Kustatscher et al. cf. cf. x x
cf. Cycadeoidea x
Cordaicarpus sp. x
Cycadeospermum sp. x
Cycadites rectangularis Brauns cf.
Danaeopsis marantacea (Presl) Schenk x
Dioonitocarpidium sp. x
Elatocladus sp. x x x
Equisetites arenaceus (Jaeger) Schenk x x x x
cf. Equisetites x
Equisetites sp. ? x
?Equisetostachys x
Fern incertis sedis x
?Filicales indet. x
Frenelopsis hoheneggeri Schenk x
Ginkgo sp. x
Lomatopteris sp. x
Lycopodites sp. x
?Neocalamites x
Neuropteris elegans Brongniart cf. x x
Neuropteris gaillardoti Brongniart. cf.
Neuropteridium grandifolium (Schimper et Mougeot)
Schimperx
Neuropteris ruetimeyeri Heer cf.
Neuropteridium sp. x x x x
cf. Neuropteridium x
Nilsonia sp. x
Odontopteris sp. x
Pagiophyllum foetterlei Stur cf. cf. x cf.
Pagiophyllum massalongi De Zigno x x cf.
Pagiophyllum peregrinum (Lindley et Hutton) Seward x
Pagiophyllum sp. x
Pecopteris (Lonchopteris) reticulata Leuthardt cf.
Pecopteris gracilis Heer x
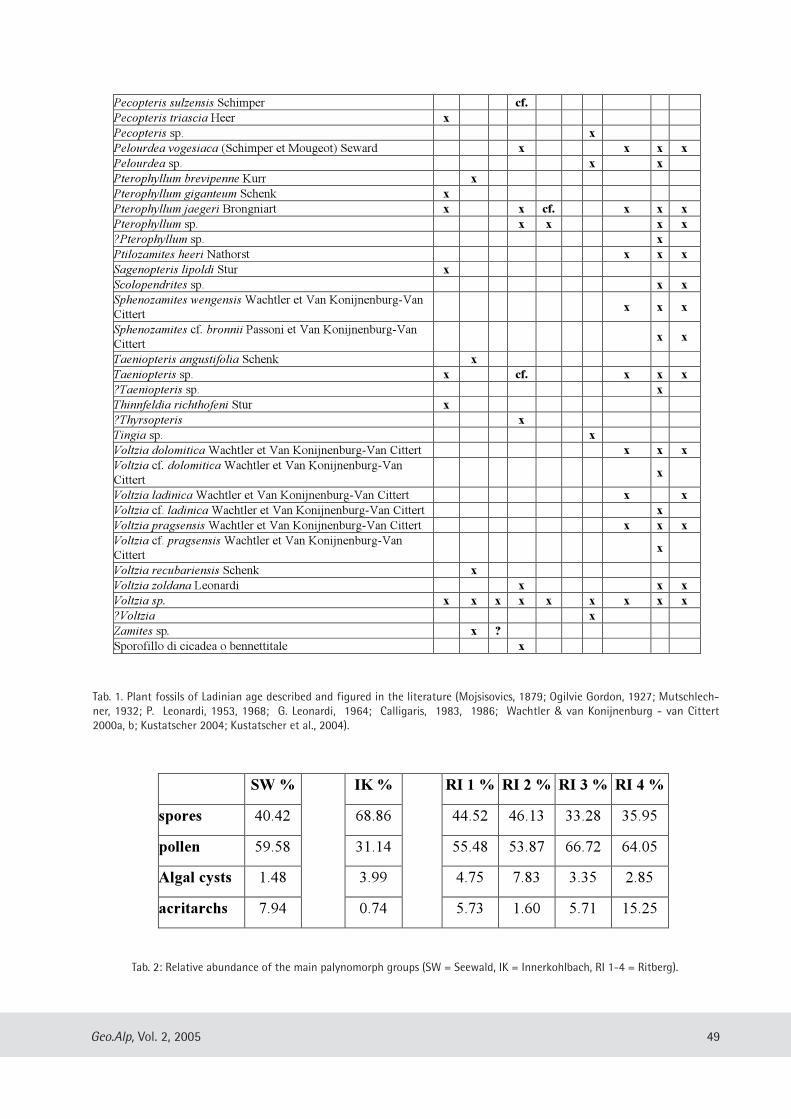
Pecopteris sulzensis Schimper cf.
Pecopteris triascia Heer x
Pecopteris sp. x
Pelourdea vogesiaca (Schimper et Mougeot) Seward x x x x
Pelourdea sp. x x
Pterophyllum brevipenne Kurr x
Pterophyllum giganteum Schenk x
Pterophyllum jaegeri Brongniart x x cf. x x x
Pterophyllum sp. x x x x
?Pterophyllum sp. x
Ptilozamites heeri Nathorst x x x
Sagenopteris lipoldi Stur x
Scolopendrites sp. x x
Sphenozamites wengensis Wachtler et Van Konijnenburg-Van
Cittertx x x
Sphenozamites cf. bronnii Passoni et Van Konijnenburg-Van
Cittertx x
Taeniopteris angustifolia Schenk x
Taeniopteris sp. x cf. x x x
?Taeniopteris sp. x
Thinnfeldia richthofeni Stur x
?Thyrsopteris x
Tingia sp. x
Voltzia dolomitica Wachtler et Van Konijnenburg-Van Cittert x x x
Voltzia cf. dolomitica Wachtler et Van Konijnenburg-Van
Cittertx
Voltzia ladinica Wachtler et Van Konijnenburg-Van Cittert x x
Voltzia cf. ladinica Wachtler et Van Konijnenburg-Van Cittert x
Voltzia pragsensis Wachtler et Van Konijnenburg-Van Cittert x x x
Voltzia cf. pragsensis Wachtler et Van Konijnenburg-Van
Cittertx
Voltzia recubariensis Schenk x
Voltzia zoldana Leonardi x x x
Voltzia sp. x x x x x x x x x
?Voltzia x
Zamites sp. x ?
Sporofillo di cicadea o bennettitale x
49Geo.Alp, Vol. 2, 2005
Tab. 1. Plant fossils of Ladinian age described and figured in the literature (Mojsisovics, 1879; Ogilvie Gordon, 1927; Mutschlech-ner, 1932; P. Leonardi, 1953, 1968; G. Leonardi, 1964; Calligaris, 1983, 1986; Wachtler & van Konijnenburg - van Cittert2000a, b; Kustatscher 2004; Kustatscher et al., 2004).
Tab. 2: Relative abundance of the main palynomorph groups (SW = Seewald, IK = Innerkohlbach, RI 1-4 = Ritberg).
SW % IK % RI 1 % RI 2 % RI 3 % RI 4 %
spores 40.42 68.86 44.52 46.13 33.28 35.95
pollen 59.58 31.14 55.48 53.87 66.72 64.05
Algal cysts 1.48 3.99 4.75 7.83 3.35 2.85
acritarchs 7.94 0.74 5.73 1.60 5.71 15.25
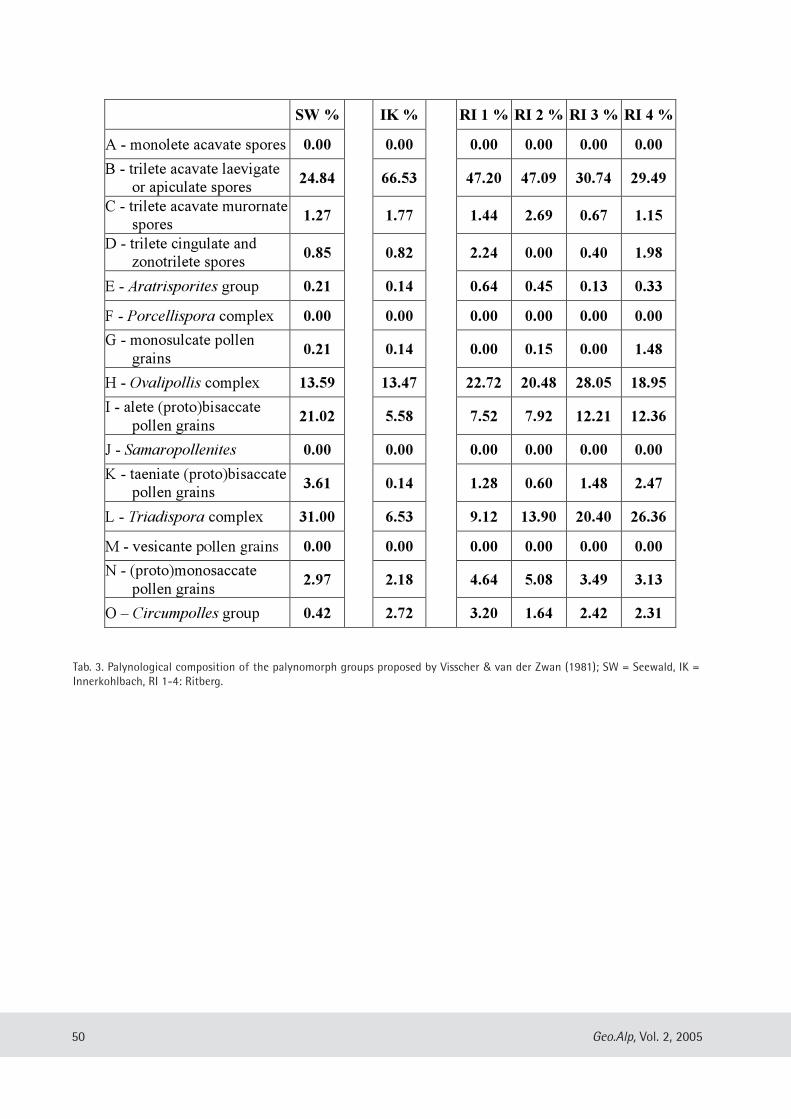
SW % IK % RI 1 % RI 2 % RI 3 % RI 4 %
A - monolete acavate spores 0.00 0.00 0.00 0.00 0.00 0.00
B - trilete acavate laevigate
or apiculate spores24.84 66.53 47.20 47.09 30.74 29.49
C - trilete acavate murornate
spores1.27 1.77 1.44 2.69 0.67 1.15
D - trilete cingulate and
zonotrilete spores0.85 0.82 2.24 0.00 0.40 1.98
E - Aratrisporites group 0.21 0.14 0.64 0.45 0.13 0.33
F - Porcellispora complex 0.00 0.00 0.00 0.00 0.00 0.00
G - monosulcate pollen
grains0.21 0.14 0.00 0.15 0.00 1.48
H - Ovalipollis complex 13.59 13.47 22.72 20.48 28.05 18.95
I - alete (proto)bisaccate
pollen grains21.02 5.58 7.52 7.92 12.21 12.36
J - Samaropollenites 0.00 0.00 0.00 0.00 0.00 0.00
K - taeniate (proto)bisaccate
pollen grains3.61 0.14 1.28 0.60 1.48 2.47
L - Triadispora complex 31.00 6.53 9.12 13.90 20.40 26.36
M - vesicante pollen grains 0.00 0.00 0.00 0.00 0.00 0.00
N - (proto)monosaccate
pollen grains2.97 2.18 4.64 5.08 3.49 3.13
O – Circumpolles group 0.42 2.72 3.20 1.64 2.42 2.31
50 Geo.Alp, Vol. 2, 2005
Tab. 3. Palynological composition of the palynomorph groups proposed by Visscher & van der Zwan (1981); SW = Seewald, IK =Innerkohlbach, RI 1-4: Ritberg.
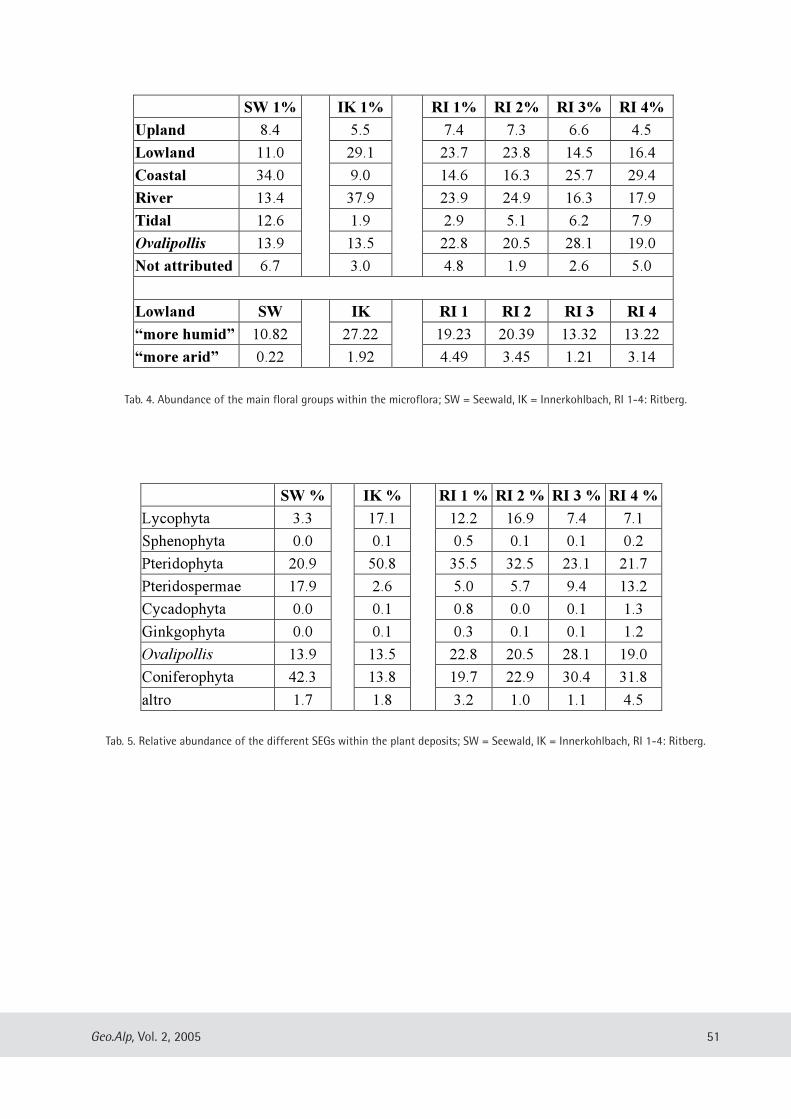
SW % IK % RI 1 % RI 2 % RI 3 % RI 4 %
Lycophyta 3.3 17.1 12.2 16.9 7.4 7.1
Sphenophyta 0.0 0.1 0.5 0.1 0.1 0.2
Pteridophyta 20.9 50.8 35.5 32.5 23.1 21.7
Pteridospermae 17.9 2.6 5.0 5.7 9.4 13.2
Cycadophyta 0.0 0.1 0.8 0.0 0.1 1.3
Ginkgophyta 0.0 0.1 0.3 0.1 0.1 1.2
Ovalipollis 13.9 13.5 22.8 20.5 28.1 19.0
Coniferophyta 42.3 13.8 19.7 22.9 30.4 31.8
altro 1.7 1.8 3.2 1.0 1.1 4.5
51Geo.Alp, Vol. 2, 2005
SW 1% IK 1% RI 1% RI 2% RI 3% RI 4%
Upland 8.4 5.5 7.4 7.3 6.6 4.5
Lowland 11.0 29.1 23.7 23.8 14.5 16.4
Coastal 34.0 9.0 14.6 16.3 25.7 29.4
River 13.4 37.9 23.9 24.9 16.3 17.9
Tidal 12.6 1.9 2.9 5.1 6.2 7.9
Ovalipollis 13.9 13.5 22.8 20.5 28.1 19.0
Not attributed 6.7 3.0 4.8 1.9 2.6 5.0
Lowland SW IK RI 1 RI 2 RI 3 RI 4
“more humid” 10.82 27.22 19.23 20.39 13.32 13.22
“more arid” 0.22 1.92 4.49 3.45 1.21 3.14
Tab. 4. Abundance of the main floral groups within the microflora; SW = Seewald, IK = Innerkohlbach, RI 1-4: Ritberg.
Tab. 5. Relative abundance of the different SEGs within the plant deposits; SW = Seewald, IK = Innerkohlbach, RI 1-4: Ritberg.

Geo.Alp, Vol. 2, S. 53–60, 2005
IITTAALLIIAANN FFOOSSSSIILL CCHHIIRROOPPTTEERRAANN AASSSSEEMMBBLLAAGGEESS:: AA PPRREELLIIMMIINNAARRYY RREEPPOORRTT
Cristiana Tata & Tassos Kotsakis
With 2 figures and 1 table
AAbbssttrraaccttThis work is a preliminary report on Italian fossil chiropteran faunas. During the Paleogene just one sam-
ple of Early Oligocene age, pertaining to an extinct species, has been reported. A few findings have beenreported from the Neogene. Just one complete assemblage from the Late Miocene site of Brisighella has beenexamined and has allowed palaeoecological inferences, whilst specimens from Late Miocene localities ofBaccinello V0 (Tuscany) and Gargano peninsula (Apulia) need a revision. A Late Pliocene assemblage has beencollected in Montagnola Senese (Tuscany) but it still needs a systematic revision.During the Quaternary and most of all since the Middle Pleistocene the fossil record becomes richer. Some
assemblages testify a Mediterranean climate analogous to the present one. The most significant are: the EarlyPleistocene ones from Pirro Nord (Apulia) and Ghar Dalam Cave (Malta), the early Middle Pleistocene onesfrom Slivia (Venezia Giulia) and Spinagallo Cave (Sicily) and the Late Pleistocene ones from Punta Padre Bellu(Sardinia) and Breuil Cave (Latium). In other cases the species represented in the assemblages are typical ofcolder climate and then they make it possible to infer cooler conditions in Italy during some periods. Goodexamples in this sense are the Middle Pleistocene assemblage from Vento Cave (Marche) and the LatePleistocene one from Cittareale Cave (Latium). A distributuion chart of all fossil bats from Italy and Malta isalso presented.
53
Dipartimento di Scienze Geologiche, Università Roma Tre, L.go S. L. Murialdo 1, 00146 Roma Italy; e-mail: [email protected], [email protected]
IInnttrroodduuccttiioonn
Nowadays it is quite common to supportpalaeoenvironmental reconstructions using samplesfrom fossil mammal (especially micromammal)assemblages as palaeoecological and/or palaeocli-matic markers. Among micromammals bats are really meaning-
ful in this respect but, especially in Italy, althoughwhen they are found they are really abundant(especially from Pleistocene sites), they are oftenlacking. The lack of interest in this group is causedby the bradytelic evolution of these animals thatmakes them useless for biochronological studiesthat, in the past decades, have been attractingpalaeontologists attention. Anyway it has to be underlined that bats,
because of their peculiar ecological habits arestrongly influenced in their distribution by climat-ic and ecological parameters and this is why they
can be considered as good environmental markers.In addition, just because of the low rates of evolu-tion, living species are mostly analogous to fossilones. Since the present distribution and the cli-matic context of their life are known it is reason-able to make palaeoclimatic and palaeoecologicalinferences from studying species pertaining to fos-sil assemblages. However first of all it is necessaryto review chiropteran assemblages and this workrepresents a preliminary approach to this researchproject.
TTeerrttiiaarryy cchhiirroopptteerraann aasssseemmbbllaaggeessaanndd tthheeiirr ppaallaaeeooeeccoollooggiiccaall mmeeaanniinngg
Just one species of bat is known from Paleogenesediments in Italy: Archaeopteropus transiensMeschinelli, 1903. It has been collected in the earlyOligocene (MP 21) lignites of Monteviale (Veneto)

(Meschinelli, 1903; Kotsakis et al., 1997) (Figs. 1, 2).It is a large chiropteran classified in its own sub-family, Archaeopteropodinae, and considered bysome authors (Russel & Sigé, 1970) to belong to thesuborder Microchiroptera and by others (Smith &Storch, 1981) to the suborder Megachiroptera.Unfortunately the original sample has been lostduring the Second World War and only some rathergood casts are available. Archaeopteropus was partof an assemblage that, if considered as a whole,shows a tropical character.In the Italian Miocene the presence of Chiroptera
indet. has been signalled from clays of Baccinello V0(Tuscany - MN11) (Kosakis et al., 1997). Just oneMiocene assemblage is known, coming fromMonticino Quarry (Brisighella, Romagna) karst fis-sures. The assemblage is of Late Turolian age (MN13)and it is composed of six species: Megaderma cf. M.mediterraneum Sigé, 1974, Rhinolophus cf. R.kowalskii Topál, 1979, Rhinolophus sp., Hipposideros(Syndesmotis) cf. H. (S.) vetus (Lavocat, 1961),Asellia cf. A. mariaetheresae Mein, 1958 and Myotiscf. M. boyeri Mein, 1964 (Kotsakis & Masini, 1989).Three species, the two rhinolophids and the vesper-tilionid, are similar to living forms now inhabiting
this same site. On the other hand the remainingthree genera, Megaderma, Hipposideros and Asellianow live in tropical and subtropical areas. In partic-ular the presence of Megaderma is indicative ofminimum temperatures higher than 14-15°C allaround the year, while the presence of Asellia isindicative of subdesertic conditions (Sigé, 1974).From the species represented here a littoral sandyhabitat has been inferred (Kotsakis & Masini, 1989).Another finding from the Italian Miocene is from
Gargano Peninsula (Apulia) from karst fissure fill-ings characterized by the Hoplitomeryx andMicrotia assemblage and ascribed to the LateMiocene – ?Early Pliocene. Here a single species, notdefinitely studied yet, has been collected and previ-ously ascribed to the genus Megaderma but an attri-bution to other megadermatid genera is possible(Kotsakis et al., 1997). As in the preceding case thepresence of this genus, now inhabiting hot regions,has suggested tropical temperatures in this area.Another Neogene assemblage is from the Late
Pliocene (MN17, Middle Villafranchian Mammal Ageor Late Villanyian Micromammal Age, Costa SanGiacomo Faunal Unit/Olivola Faunal Unit; Kotsakiset al., 2003) fissure fillings of Montagnola Senese(Tuscany) (Fondi, 1972). It includes four species:Myotis blythii (Tomes, 1857), M. gr. schaubi Kormos,1934 – rapax Heller, 1936, Myotis sp. and ? Tadaridasp. The fauna needs a systematic revision and it isimpossible to infer palaeoecological informations.
QQuuaatteerrnnaarryy cchhiirroopptteerraann aasssseemmbbllaaggeessaanndd tthheeiirr ppaallaaeeooeeccoollooggiiccaall mmeeaanniinngg
During the Pleistocene, particularly in the LatePleistocene, an increase in the Italian fossil record isobserved. Among various sites under study at pre-sent the most meaningful are Pirro Nord (Gargano,Apulia), Spinagallo Cave (Sicily), Punta Padre Bellu(Alghero, Sardinia), Breuil Cave (Monte Circeo,Latium), Monte Cucco Cave (Perugia, Umbria) andCittareale Cave (Rieti, Latium). One assemblage fromthe late Early Pleistocene/earliest Middle Pleisto -cene from the Ghar Dalam Cave (Malta) is stronglyrelated to Italian faunas.The oldest chiropteran assemblage is Pirro Nord,
ascribed to the early Pleistocene (Late VillafranchianM.A. or Early Biharian Micromammal Age, Pirro F.U.)(Gliozzi et al., 1997). A rich assemblage has been col-lected from one of the karst fissures in the area. It is
54 Geo.Alp, Vol. 2, 2005
Fig.1: Map of main fossiliferous localities of Italy and Malta.1- Monteviale (Veneto); 2- Baccinello V0 (Tuscany); 3- Brisig-hella (Romagna); 4- Gargano (Apulia); 5- Montagnola Senese(Tuscany); 6- Pirro Nord (Apulia); 7- Ghar Dalam Cave (Malta);8- Spinagallo Cave (Sicily); 9- Slivia (Venezia Giulia); 10- VentoCave (Marche); 11- Punta Padre Bellu (Sardinia); 12- BreuilCave (Latium); 13- Monte Cucco Cave (Marche); 14- CittarealeCave (Latium).

55Geo.Alp, Vol. 2, 2005
Fig. 2: Biochronological scheme of localities bearing fossil bats of Italy and Malta.

composed of six species: Rhinolophus ferrume-quinum (Schreber, 1774), R. birzebbugensis Storch,1974, Myotis blythii, M. capaccinii (Bonaparte,1837), Miniopterus schreibersi (Kuhl, 1819) andMiniopterus n. sp. (Masini et al., 1996; Tata, 2003).Among them the living species R. ferrumequinum,M. blythii, M. capaccinii and M. schreibersi are pre-sent in Europe in the central and southern part ofthe continent (except for R. ferrumequinum thatextends to more northern latitudes). The remainingtwo species R. birzebbugensis and Miniopterus n. sp.cannot be considered as strong palaeoclimatic mark-ers since the rhinolophid is known as a fossil onlyfrom a few localities (Malta, Bulgaria and probablySpain) (Storch, 1974; Popov, 2004; Tata & Kotsakis,in prep.) while the miniopterid has been collectedhere for the first time. Considered as a whole theassemblage has a strong Mediterranean character. Adifferent assemblage including three species hasbeen collected from another fissure filling in thesame area (De Giuli & Torre, 1984): Rhinolophus gr. R.euryale Blasius, 1853, Myotis cf. M. blythii and Myotissp. (small size). From a climatic point of view theassemblage does not differ from the previous one.Close in age to the previous assemblage is that
collected in the Ghar Dalam Cave (strata with Leithiacartei; Storch, 1974) including ten species:Rhinolophus hipposideros (Bechstein, 1800), R.birzebbugensis, R. blasii Peters, 1866, Myotis exilisHeller, 1936, M. bechsteini robustus Topál, 1963, M.ghardalamensis Storch, 1974, M. capaccinii,Eptesicus praeglacialis Kormos, 1930, Pipistrelluspipistrellus (Schreber, 1774) and Miniopterusschreibersi. The assemblage shows a Mediterraneancharacter with forested and open habitats and thepresence of fresh water.Also the assemblage from the Spinagallo Cave,
ascribed to the early Middle Pleistocene (Elephas fal-coneri Faunal Complex) (Bonfiglio et al., 2003) isquite rich including ten species: R. ferrumequinum,R. hipposideros, R. mehelyi Matschie, 1901, R. cf. R.blasii, Myotis mystacinus (Leisler in Kuhl, 1819), M.bechsteini (Leisler in Kuhl, 1819), M. capaccinii,Eptesicus serotinus (Schreber, 1774), Barbastella bar-bastellus (Schreber, 1774) and M. schreibersi (cfr.Kotsakis & Petronio, 1980). Very probably the newspecies of Miniopterus collected in Pirro Nord is alsopresent in the Spinagallo Cave assemblage.The species are derived from two different strata;
in the lower one only three species are represented:M. schreibersi, R. ferrumequinum and M. capaccinii,
the first being decisely dominant. In the higher stra-tum all the species, except M. capaccinii, are repre-sented. M. schreibersi is always dominant althoughless numerous than in the lower stratum. On thewhole the assemblage has a Mediterranean charac-ter although forms such as M. mystacinus and B.barbastellus usually have a more northern distribu-tion. All but one species are still living in Sicily, M.bechsteini and B. barbastellus are less widespreadthen the others (Kotsakis & Petronio, 1980). R. blasiihas a recent eastern Mediterranean distribution andis present in the easternmost province of Italy nearthe Italian-Slovenian boundary (Lanza & Agnelli,1999). Palaeoecological conditions similar to thoseof Ghar Dalam can be inferred. At several fossil localities a small number of bat
species has been collected; usually the specimensrepresented there belong to recent species now liv-ing in the same areas. A good example is the earlyMiddle Pleistocene (Early Galerian M.A., Slivia F.U.)assemblage collected in Slivia karst fissure (Trieste,Venezia Giulia) where two species have been recog-nized: Rhinolophus ferrumequinum and Mini -opterus schreibersi (cfr. Ambrosetti et al., 1979). Inother cases some elements indicating colder condi-tions have been recognized in mammal assemblagesas for example in the late Middle Pleistocene (EarlyAurelian M.A.) deposit from Vento Cave (Ancona,Marche), where two species are found: Rhinolophusferrumequinum and Myotis dasycneme (Boie, 1825)(Esu et al., 1990). Among them particularly signifi-cant in this respect is the presence of M. dasycneme,typical of cold conditions, known at present by justone erratic individual in the north-eastern part ofthe Italian peninsula . In the Late Pleistocene the findings are abun-
dant, isolated remains are often reported(Lombardy, Santi, 2000; Sardinia, Abbazzi et al.,2004), but systematic analysis has been conductedonly on a few cave deposits.The assemblage from Punta Padre Bellu, collect-
ed in a destroyed cave near Alghero, has beenascribed to the Late Pleistocene and is composed ofsix species: R. ferrumequinum, R. hipposideros,Myotis myotis (Borkhausen, 1797), M. capaccinii,Nyctalus cf. N. lasiopterus (Schreber, 1780) and M.schreibersi (cfr. Kotsakis, 1987). All the species withthe exeption of N. lasiopterus (that however is quiterare in the peninsula today) are still living inSardinia suggesting a climatic context similar to thepresent one.
56 Geo.Alp, Vol. 2, 2005
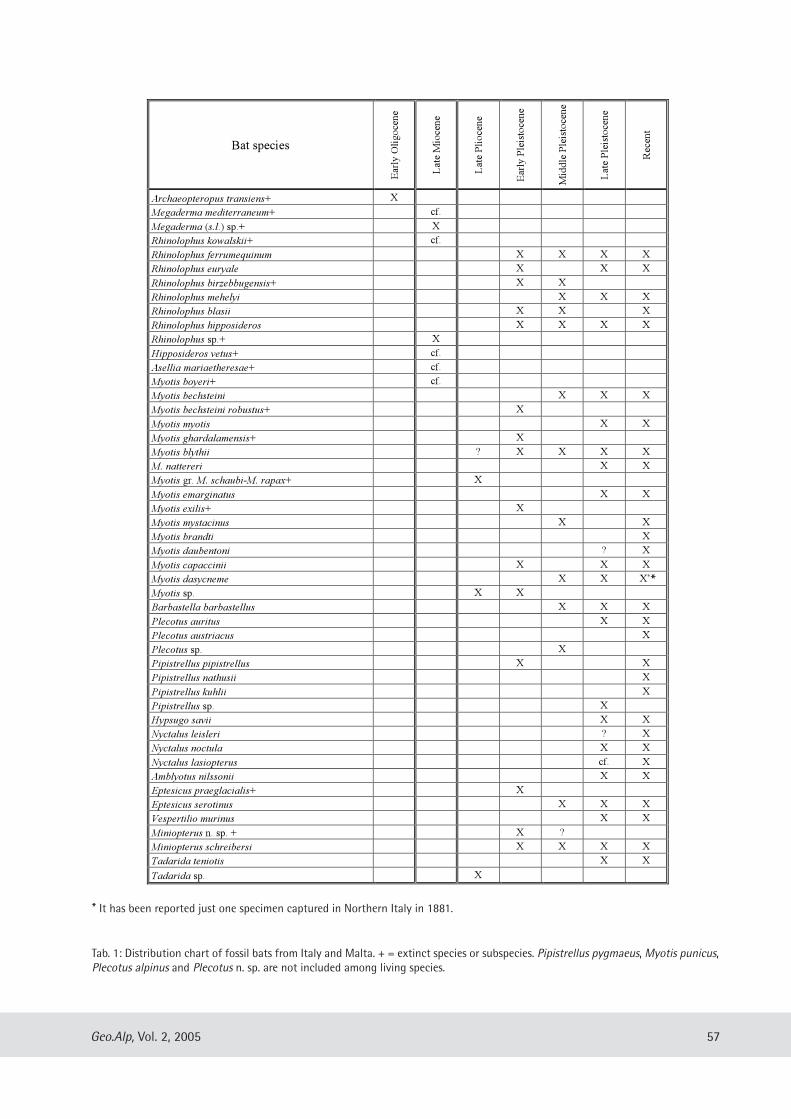
57Geo.Alp, Vol. 2, 2005
14
Bat species
Ear
ly O
ligoce
ne
Lat
e M
ioce
ne
Lat
e P
lioce
ne
Ear
ly P
leis
toce
ne
Mid
dle
Ple
isto
cene
Lat
e P
leis
toce
ne
Rec
ent
Archaeopteropus transiens+ X
Megaderma mediterraneum+ cf.
Megaderma (s.l.) sp.+ X
Rhinolophus kowalskii+ cf.
Rhinolophus ferrumequinum X X X X
Rhinolophus euryale X X X
Rhinolophus birzebbugensis+ X X
Rhinolophus mehelyi X X X
Rhinolophus blasii X X X
Rhinolophus hipposideros X X X X
Rhinolophus sp.+ X
Hipposideros vetus+ cf.
Asellia mariaetheresae+ cf.
Myotis boyeri+ cf.
Myotis bechsteini X X X
Myotis bechsteini robustus+ X
Myotis myotis X X
Myotis ghardalamensis+ X
Myotis blythii ? X X X X
M. nattereri X X
Myotis gr. M. schaubi-M. rapax+ X
Myotis emarginatus X X
Myotis exilis+ X
Myotis mystacinus X X
Myotis brandti X
Myotis daubentoni ? X
Myotis capaccinii X X X
Myotis dasycneme X X X!
Myotis sp. X X
Barbastella barbastellus X X X
Plecotus auritus X X
Plecotus austriacus X
Plecotus sp. X
Pipistrellus pipistrellus X X
Pipistrellus nathusii X
Pipistrellus kuhlii X
Pipistrellus sp. X
Hypsugo savii X X
Nyctalus leisleri ? X
Nyctalus noctula X X
Nyctalus lasiopterus cf. X
Amblyotus nilssonii X X
Eptesicus praeglacialis+ X
Eptesicus serotinus X X X
Vespertilio murinus X X
Miniopterus n. sp. + X ?
Miniopterus schreibersi X X X X
Tadarida teniotis X X
Tadarida sp. X
Tab. 1: Distribution chart of fossil bats from Italy and Malta. + = extinct species or subspecies. Pipistrellus pygmaeus, Myotis punicus,Plecotus alpinus and Plecotus n. sp. are not included among living species.
* It has been reported just one specimen captured in Northern Italy in 1881.
*

The rich assemblage from Breuil Cave (MonteCirceo, Latium) collected in two strata (stratum “e”and stratum “d”) must be referred to the LatePleistocene (OIS 3). Among micromammals bats arewell represented with five species: R. ferrume-quinum, M. myotis, Nyctalus noctula (Schreber,1774), M. schreibersi and Tadarida teniotis(Rafinesque, 1814). All these species are present inthe lower part of the stratum “e”, while only R. fer-rumequinum is represented in the upper part of thestratum “d” (Kotsakis, 1989). The assemblagederived from stratum “e” is constituted partly bytypical Mediterranean species such as M. schreiber-si and T. teniotis (that are more aboundantly repre-sented) and partly by species having a more north-ern distribution such as N. noctula. If this assem-blage is considered in the general faunal context itbecomes quite clear that its interstadial characterdenotes a woodland environment with moist areasin the neighbourhood of the cave.Another Late Pleistocene (OIS 2) chiropteran
assemblage is that from the Monte Cucco Cave(Perugia, Umbria) (Capasso Barbato & Kotsakis,1986), including five species: R. ferrumequinum, M.myotis, M. blythii, M. bechsteini and M. emargina-tus (E. Geoffroy, 1806). The absence of Miniopterusschreibersi is interesting because it is a usual com-ponent of Italian cave- dwelling faunas. The assem-blage does not show any peculiar characteristicsthat allow palaeoclimatic inferences. The chiropteran assemblage from Cittareale Cave
(Rieti, Latium) is clearly colder and ascribed to theLate Pleistocene (OIS 2, Younger Dryas?); fivespecies are present: R. ferrumequinum, R. hip-posideros, M. myotis, M. bechsteini and M. dasyc-neme (cfr. Argenti et al., in press). Particularlymeaningful in a climatic sense is the presence of M.dasycneme that suggests the attribution of theassemblage to a cold interval, presumably to theYounger Dryas period. In addition all the species,with the exeption of M. myotis that usually prefersopen and slightly wooded terrain, are common inwooded areas suggesting then, for the assemblage,a forested environment with open space and ponds.
CCoonncclluussiioonnss
Among the recent mammalian faunas of Italyand Malta, Mitchell-Jones et al. (1999) indicate thepresence of 28 species of bats. This number has
been increased in the last years by new researches:Pipistrellus pygmaeus (Leach, 1825) (Russo & Jones,2000); Myotis punicus Felten, 1977 (Castella et al.,2000; Beuneux, 2004); Plecotus alpinus Kiefer &Veith, 2001 (Trizio et al., 2003); Plecotus n. sp. fromSardinia (Mucedda et al., 2002) have been added inthe list of bats of Italy. However for an attributionof Italian fossil material to these species a completesystematic revision is necessary.The number of Italian Tertiary fossil species is
much less; it has been calculated to include 12species, among them 11 are surely extinct, but alsothe twelwth, which has been attributed to a livingspecies, needs a systematic revision. During the Quaternary an increase in the number
of species is observed with at least 31 represented;5 of this number are extinct (R. birzebbugensis, M.ghardalamensis, M. exilis, E. praeglacialis andMiniopterus n. sp.). A fossil subspecies has also beenreported M. bechsteini robustus. Another specieshas to be mentioned pertaining to the genusRhinolophus, R. botegoi Regàlia, 1893 described byRegàlia (1893), from fossil remains collected inColombi Cave (Palmaria Island, Liguria). Its validityseems to be improbable, but in any case the mater-ial needs to be revised. In the fossil record of Italianbats the presence of troglophilous species is domi-nant, whilst non-cave dwelling species are not wellrepresented (see Table 1).The analysis of the chiropteran assemblages con-
firms that during the time span between theMiocene and Pleistocene the Italian peninsula hasbeen subjected to a general decrease of tempera-ture. This inference comes from the observationthat species typical of tropical and subtropical envi-ronments present in the Neogene assemblages arecompletely lacking from more recent assemblages.It has to be emphasised that in some cases the pres-ence of a single species with peculiar ecologicalrequirements gives clear palaeoecological informa-tion whilst in other cases it is the assemblage as awhole (considering the percentage composition ofeach single species) that allows palaeoecolgicalinferences.
AAcckknnoowwlleeddggmmeennttss
We wish to thank Prof. G. Tichy of SalzburgUniversity for reviewing the manuscript and Dr.D. Harrison of Harrison Institute for the helpful dis-
58 Geo.Alp, Vol. 2, 2005

cussions about bat systematics with the first authorand the correction of the English.
RReeffeerreenncceess
Abbazzi, L., Angelone, C., Arca, M., Barisone, G., Bedetti,C., Delfino, M., Kotsakis, T., Marcolini, F., Palombo,M.R., Pavia, M., Piras, P., Rook, L., Torre, D., Tuveri, C.,Valli, A., Wilkens, B. (2004): Plio-Pleistocene fossilvertebrates of Monte Tuttavista (Orosei, E. Sardinia,Italy), an overview. — Riv. Ital. Paleont. Strat., 110:613-638.
Ambrosetti, P., Bartolomei, G., De Giuli, C., Ficcarelli, G.,Torre, D. (1979): La breccia ossifera di Slivia (Aurisina– Sistiana) nel carso di Trieste. – Boll. Soc. Paleont.Ital., 18: 207-220.
Argenti, P., Kotsakis, T., Sabatini, F. (in press): The latestPleistocene bats of Cittareale Cave (Rieti, Latium,Central Italy).
Beuneux, G. (2004): Morphometrics and ecology ofMyotis cf. punicus (Chiroptera, Vespertilionidae) inCorsica. – Mammalia, 68: 269-273.
Bonfiglio, L., Di Maggio, C., Marra, A. C., Masini, F.,Petruso, D. (2003): Biochronology of Pleistocene ver-tebrate faunas of Sicily and correlation of vertebratebearing deposits with marine deposits. – IlQuaternario, 16 (1bis): 102-114.
Capasso Barbato, L., Kotsakis, T. (1986): Les chiroptèresdu Pléistocène supérieur de la Grotte de Monte Cucco(Italie Centrale). – Geol. Romana, 25: 309-316.
Castella, V., Ruedi, M., Excoffier, L., Ibáñez, C., Arlettaz, R.,Hauser, J. (2000): Is the Gibraltar Strait a barrier togene flow for the bat Myotis myotis (Chiroptera :Vespertilionidae)? – Mol. Ecol., 9: 1761-1772.
De Giuli, C., Torre, D. (1984): A microfauna withAllophaionys pliocaenicus from Gargano (SouthernItaly). – Palaeontogr. Ital., 73: 115-128.
Esu, D., Galdenzi, S., Kotsakis, T. (1990): Molluschi emicrovertebrati del deposito pleistocenico della Grottadel Vento (Gola della Rossa, Ancona). – Nota prelimi-nare. Mem. Ist. Ital. Spel., 4: 193-198.
Fondi, R. (1972): Fauna cromeriana della Montagnolasenese. Palaeontogr. Ital., 68: 1-27.
Gliozzi, E., Abbazzi, L., Argenti, P., Azzaroli, A., Caloi, L.,Capasso Barbato, L., Di Stefano, G., Esu, D., Ficcarelli,G., Girotti, O., Kotsakis, T., Masini, F., Mazza, P.,Mezzabotta, C., Palombo, M.R., Petronio, C., Rook, L.,Sala, B., Sardella, R., Zanalda, E., Torre, D. (1997):Biochronology of selected mammals, molluscs andostracods from the Middle Pliocene to the Late
Pleistocene in Italy. The state of the art. – Riv. Ital.Paleont. Strat., 103: 369-388.
Kotsakis, T. (1987): Les chiroptères du Pléistocène supé-rieur des environs de Alghero (Sardaigne, Italie). –Geol. Romana, 26: 103-108.
Kotsakis, T. (1989): Late Pleistocene fossil microverte-brates of Grotta Breuil (Monte Circeo, Central Italy). –Quaternaria Nova, 1: 325-332.
Kotsakis, T., Abbazzi, L., Angelone, C., Argenti, P., Barisone,G., Fanfani, F., Marcolini, F., Masini, F. (2003): Plio-Pleistocene biogeography of Italian mainland micro-mammals. – In Reumer J.W.F. & Wessels W. (Eds.):Distribution and migration of Tertiary mammals inEurasia. A volume in honour of Hans de Brujin.Deinsea, 10: 313-342.
Kotsakis, T., Barisone, G., Rook, L. (1997): Mammalianbiochronology in an insular domain: the ItalianTertiary faunas. – Mém. Trav. E. P. H. E. Inst.Montpellier, 21: 431-441.
Kotsakis, T., Masini, F. (1989): Late Turolian bats fromBrisighella (Northern Italy). Boll. Soc. Paleont. Ital., 28:281-285.
Kotsakis, T., Petronio, C. (1980): I chirotteri del Pleistocenesuperiore della grotta di Spinagallo (Siracusa, Sicilia).Boll. Serv. Geol. Ital., 101: 49-76.
Lanza, B., Agnelli, P. (1999): Chirotteri. – In Spagnesi, M.,Toso, S. (Eds.): Iconografia dei mammiferi d’Italia.Istituto Nazionale per la Fauna Selvatica “AlessandroChigi”, 44-142.
Masini, F., Rook, L., Abbazzi, L., Ambrosetti, P., Azzaroli, A.,Ficcarelli, G., Gentili, S., Kotsakis, T., Sala, B., Torre, D.(1996): Mammalian faunas of selected Villafranchianlocalities of Italy. – In Carraro F. (Ed.): Revisione delVillafranchiano nell’area tipo di Villafranca d’Asti. IlQuaternario, 9, tab. 2.
Meschinelli, L. (1903): Un nuovo chirottero fossile(Archaeopteropus transiens Mesch.) delle ligniti diMonteviale. – Atti R. Ist. Veneto Sci. Lett. Arti, 62:1329-1344.
Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W.,Kryštufek, B., Reijnders, P.J.H., Spitzenberger, F.,Stubbe, M., Thissen, J.B.M., Vohralík, V., Zima, J. (1999):The atlas of European mammals. Academic Press, 484pp..
Mucedda, M., Kiefer, A., Pidinchedda, E., Veith, M. (2002):A new species of long-eared bat (Chiroptera,Vespertilionidae) from Sardinia (Italy). – ActaChiropterol., 4: 121-135.
Popov, V. (2004): Pliocene small mammals (Mammalia,Lipotyphla, Chiroptera, Lagomorpha, Rodentia) fromMuselievo (North Bulgaria). – Geodiversitas, 26: 403-491.
59Geo.Alp, Vol. 2, 2005

Regàlia, E. (1893): Sulla fauna della «Grotta dei Colombi»(Is. Palmaria, Spezia). – Nota paleontologica. Arch.Antropol. Etnol., 23: 257-366.
Russel, D. E., Sigé, B. (1970): Révision des chiroptères luté-tiens de Messel (Hesse, Allemagne). – Palaeo verte -brata, 3: 83-182.
Russo, D., Jones, G. (2000): The two cryptic species ofPipistrellus pipistrellus (Chiroptera: Vespertilionidae)occur in Italy: evidence from echolocation and socialcalls. – Mammalia, 64: 187-197.
Santi, G. (2000): Remains of Würmian Myotis into Ursusspelaeus skull from Buco dell’Orso Cave (Laglio: Como– Lombardy, Italy). – Atti Ticin. Sci. Terra, 41: 41-47.
Sigé, B. (1974): Presence d’un Megaderma (Mammalia,Chiroptera) dans le Pléistocène inférieur à Sète(Hérault). – Géol. Méditerr., 1: 97-104.
Smith, J.D., Storch, G. (1981): New Middle Eocene batsfrom “Grube Messel” near Darmstadt, W-Germany.(Mammalia: Chiroptera). – Senckenberg. Biol., 61:153-167.
Storch, G. (1974): Quartäre Fledermaus-Faunen von derInsel Malta. – Senckenberg. Lethaea, 55:: 407-434.
Tata, C. (2003): Fauna a chirotteri del Villa franchianosuperiore del Gargano (Puglia, Italia meridionale). –Unpublished Laurea Thesis, University Roma Tre.
Tata, C., Kotsakis, T. (in prep.): A chiropteran fauna fromEarly Pleistocene of Gargano Peninsula (Apulia,Southern Italy).
Trizio, I., Patriarca, E., Debernardi, P., Preatoni, A., Tosi, G.,Martinoli, A. (2003): The alpine long-eared bat(Plecotus alpinius Kiefer and Veith, 2001) is presentalso in Piedmont region: first record revealed by DNAanalysis. – Hystrix, n. s., 14: 113-115.
60 Geo.Alp, Vol. 2, 2005
Manuscript submitted: December 12, 2004Revised manuscript accepted: March 30, 2005

Geo.Alp, Vol. 2, S. 61–70, 2005
CCEERRVVUUSS EELLAAPPHHUUSS SSIICCIILLIIAAEE FFRROOMM PPLLEEIISSTTOOCCEENNEE LLAACCUUSSTTRRIINNEE DDEEPPOOSSIITTSSOOFF AACCQQUUEEDDOOLLCCII ((NNOORRTTHH--EEAASSTTEERRNN SSIICCIILLYY,, IITTAALLYY)) AANNDD IITTSS TTAAPPHHOONNOOMMIICC SSIIGGNNIIFFIICCAANNCCEE
Gabriella Mangano
With 6 figures and 12 tables
AAbbssttrraaccttSystematic excavations carried out on the Pleistocene lacustrine deposits of Acquedolci (North-Eastern
Sicily, Italy) yelded a very rich fossil vertebrate assemblage, containing thousands of remains ofHippopotamus pentlandi, the endemic hippo of the Siculo-Maltese archipelago, associated with remains ofCervus elaphus siciliae and scarce remains of Ursus cf. arctos, Canis lupus, Testudo cf. hermanni, Elephasmnaidriensis and Aves. This paper presents a morphological, biometrical and taphonomical analysis of Cervuselaphus siciliae remains. Morphological and biometrical features are in the range of the variability of Cervuselaphus siciliae Pohlig, the endemic deer of Sicily which is characterized by a slightly smaller size comparedto the populations of the Italian peninsula. Taphonomical features, such as spatial distribution and orienta-tion of the remains, composition of the skeletal part, age distribution, degree of skeletal articulation, frag-mentation and bone modification, indicate that Cervus elaphus siciliae remains did not accumulate “in situ”,unlike the autochthonous remains of Hippopotamus pentlandi, but probably they were occasionally deposit-ed in the lacustrine basin as fragments of carcasses belonging to the animals inhabiting the surroundingarea.
61
Dipartimento di Scienze della Terra, Università degli Studi di Messina; e-mail: [email protected]
IInnttrroodduuccttiioonn
The lacustrine sediments of Acquedolci arelocated on the northern flank of the Nebrodi range(North-Eastern Sicily), at the base of the high ver-tical cliff of the Pizzo Castellaro carbonatic massif,on which the well-known S. Teodoro Cave opens.The deposit is composed of silt, gravel and pebblesof variable size, probably fallen from the adjacentcliff. It is superimposed on a Late Pleistocenemarine terrace located 131 m a.s.l. and representsthe remains of a late Middle Pleistocene lacustrinebasin (Bonfiglio, 1985; 1987; 1989; 1992). Duringthe years 1982-1987 systematic excavations werecarried out and seven trenches of different widthand depth have been excavated over an area of104 m2 (Fig. 1). About 130 m3 of sediments wereremoved and the entire succession of the deposit,which was originally about 14 m thick, was investi-gated. In trench G the lacustrine sediments con-taining fossil remains are absent.
Most of the collected remains come from thetrench F, which was deepened for 6 m. In the othertrenches, which have a maximum depth of about 2m, the fossil bones have been partially preserved insitu because of their spectacular abundance, inorder to establish a field Museum.
A total number of 3.016 remains of Hippo -potamus pentlandi, the endemic hippo of theSiculo-Maltese archipelago, together with 104remains of the endemic deer of Sicily, Cervus ela-phus siciliae, and very scarce remains of Ursus cf.arctos (15), Canis lupus (7), Testudo cf. hermanni(6), Elephas sp. (1) and Aves (2) were collected(Bonfiglio, 1995). One of the two remains of Avesbelongs to Gyps melitensis Lydekker, an extinct vul-ture (griffon) species (Pavia, 2001).
This faunal assemblage belongs to the “Elephasmnaidriensis Faunal Complex”, one of the fivePleistocene faunal complexes recognized in Sicily,referred to the late Middle Pleistocene-early LatePleistocene (Bonfiglio et al., 2001; 2002). Amino-
acid racemization dating yielded an age of 200 + 40Ky for the Hippopotamus pentlandi remains ofAcquedolci (Bada et al., 1991).
MMoorrpphhoollooggiiccaall aanndd bbiioommeettrriiccaall ddeessccrriippttiioonnss
A total number of 104 strongly fragmentedremains of Cervus elaphus siciliae were collected.The only complete and well preserved bones arerepresented by two metacarpals. Antlers andmetapodials are the most frequent skeletal elements(Tab. 1). A morphological and biometrical compari-son with the remains of Cervus elaphus siciliaePohlig from different Pleistocene deposits of Sicily,described by Gliozzi et al. (1993), is presented. Atpresent, the data published by Gliozzi et al. (1993)about the remains of Cervus elaphus siciliae fromSicily are the only available ones. The remains fromAcquedolci do not have a catalogue number.
62 Geo.Alp, Vol. 2, 2005
Fig. 1: Topography of the Acquedolci area and location of theexcavation trenches (A-G) (modified from Bonfiglio, 1987).
Tab. 2: Measurements (mm) of the antler of Cervus elaphus siciliae from Acquedolci compared with the dimensions of antlersdescribed by Gliozzi et al., 1993.
Fig. 2: Right shed antler of Cervus elaphus siciliae, internalview.
Tab. 1: Composition of the skeletal part of Cervus elaphus siciliae remains from Acquedolci.
ANTLER right (Gliozzi et al., 1993)
transverse diameter of the burr 47 –
antero-posterior diameter of the burr 63 min 51 – max 67
transverse diameter of the beam above the bez-tine 37 –
antero-posterior diameter of the beam above the bez-tine 42 min 34 – max 44
Skeletal element N.R.
skull 3
antler 23
vertebrae 7
ribs 15
scapula 1
humerus 4
radius 11
femur 2
tibia 4
podials 5
metapodials 22
phalanges 7
Total 104

Skull. The skull remains are represented by 3pedicle fragments only. The most complete of thesebones is a left pedicle, which is rather short andstrong. The antero-posterior diameter is 44 mm,while the transverse diameter is 41 mm. The skullsof Cervus elaphus siciliae collected in the PuntaliCave (Palermo) have antero-posterior diameters ofthe pedicles varying between 34.8 and 40.6 mm,and the transverse diameter of the pedicles ranging
between 33.4 and 41 mm (Gliozzi et al., 1993).Another fragment of skull from the VillafrancaTirrena deposit (Messina) has an antero-posteriordiameter of the pedicle measuring 43 mm(Mangano, 2000).
Antlers. A total number of 23 antler fragmentswere recovered: 7 fragments of tines, 9 fragmentsof beams and 7 shed antler fragments with burr. Theonly measurable remain is a right shed antler frag-ment, which was strongly fractured and recon-structed by restoration (Fig. 2). The burr and thefirst portion of the beam are preserved, the brow-tine and bez-tine are broken. The burr is moderate-ly developed and formed by little pearls. Theapproximate measurements of this specimen arelisted in Tab. 2. The dimensions of the antero-poste-rior diameter of the burr and of the beam above thebez-tine are in the range of the variability of Cervuselaphus siciliae (Gliozzi et al., 1993).
Vertebrae. 6 vertebrae are present. They are frac-tured and incomplete. Two fragments belong toyoung individuals.
Ribs. 15 fragments of ribs lacking the articulationsurface were recovered.
Scapula. The scapula is represented only by oneproximal right fragment (Tab. 3). The glenoid cavityis slightly ovoidal in shape with a well developedconcave surface. The glenoid tubercle is very strong.The neck is rather short and slender. The remains ofscapula of Cervus elaphus siciliae recovered in theFata Donnavilla Cave (Messina) display the samemorphological features (Gliozzi et al., 1993).
Humerus. The humerus is poorly represented by 4fragmentary specimens: 2 distal fragments preserv-
63Geo.Alp, Vol. 2, 2005
Tab. 3: Measurements (mm) of the scapula of Cervus elaphus siciliae from Acquedolci.
Tab. 4: Measurements (mm) of the humerus of Cervus elaphus siciliae from Acquedolci compared with the dimensions of the remainsdescribed by Gliozzi et al., 1993.
Fig. 3: Right distal humerus of Cervus elaphus siciliae; a) ante-rior view, b) posterior view.
SCAPULA right
transverse diameter of the glenoid cavity 33
antero-posterior diameter of the glenoid cavity 37
antero-posterior diameter of the neck 31
antero-posterior diameter of the articulation surface 49
HUMERUS right left (Gliozzi et al., 1993)
transverse diameter of the distal end 48 42 min 40 – max 49.2
antero-posterior diameter of the distal end 49 43 min 37.5 – max 46
transverse diameter of the trochlea 45 40 –
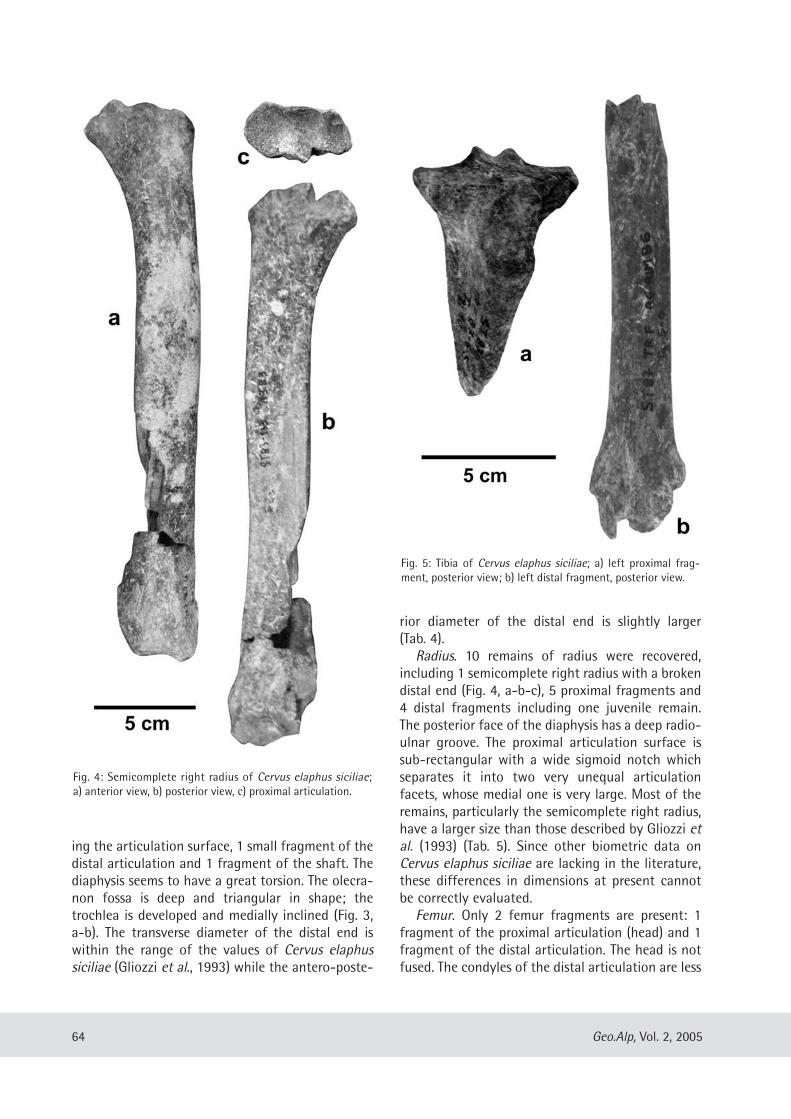
ing the articulation surface, 1 small fragment of thedistal articulation and 1 fragment of the shaft. Thediaphysis seems to have a great torsion. The olecra-non fossa is deep and triangular in shape; thetrochlea is developed and medially inclined (Fig. 3,a-b). The transverse diameter of the distal end iswithin the range of the values of Cervus elaphussiciliae (Gliozzi et al., 1993) while the antero-poste-
rior diameter of the distal end is slightly larger(Tab. 4).
Radius. 10 remains of radius were recovered,including 1 semicomplete right radius with a brokendistal end (Fig. 4, a-b-c), 5 proximal fragments and4 distal fragments including one juvenile remain.The posterior face of the diaphysis has a deep radio-ulnar groove. The proximal articulation surface issub-rectangular with a wide sigmoid notch whichseparates it into two very unequal articulationfacets, whose medial one is very large. Most of theremains, particularly the semicomplete right radius,have a larger size than those described by Gliozzi etal. (1993) (Tab. 5). Since other biometric data onCervus elaphus siciliae are lacking in the literature,these differences in dimensions at present cannotbe correctly evaluated.
Femur. Only 2 femur fragments are present: 1fragment of the proximal articulation (head) and 1fragment of the distal articulation. The head is notfused. The condyles of the distal articulation are less
64 Geo.Alp, Vol. 2, 2005
Fig. 4: Semicomplete right radius of Cervus elaphus siciliae;a) anterior view, b) posterior view, c) proximal articulation.
Fig. 5: Tibia of Cervus elaphus siciliae; a) left proximal frag-ment, posterior view; b) left distal fragment, posterior view.
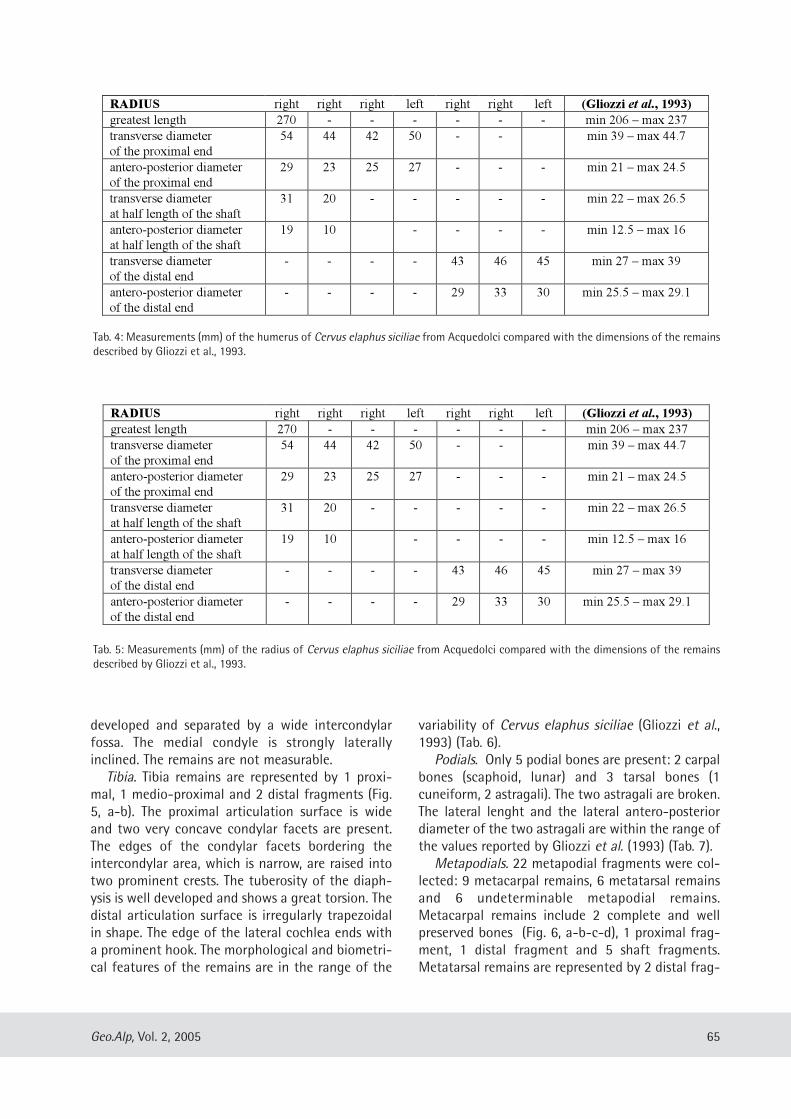
developed and separated by a wide intercondylarfossa. The medial condyle is strongly laterallyinclined. The remains are not measurable.
Tibia. Tibia remains are represented by 1 proxi-mal, 1 medio-proximal and 2 distal fragments (Fig.5, a-b). The proximal articulation surface is wideand two very concave condylar facets are present.The edges of the condylar facets bordering theintercondylar area, which is narrow, are raised intotwo prominent crests. The tuberosity of the diaph-ysis is well developed and shows a great torsion. Thedistal articulation surface is irregularly trapezoidalin shape. The edge of the lateral cochlea ends witha prominent hook. The morphological and biometri-cal features of the remains are in the range of the
variability of Cervus elaphus siciliae (Gliozzi et al.,1993) (Tab. 6).
Podials. Only 5 podial bones are present: 2 carpalbones (scaphoid, lunar) and 3 tarsal bones (1cuneiform, 2 astragali). The two astragali are broken.The lateral lenght and the lateral antero-posteriordiameter of the two astragali are within the range ofthe values reported by Gliozzi et al. (1993) (Tab. 7).
Metapodials. 22 metapodial fragments were col-lected: 9 metacarpal remains, 6 metatarsal remainsand 6 undeterminable metapodial remains.Metacarpal remains include 2 complete and wellpreserved bones (Fig. 6, a-b-c-d), 1 proximal frag-ment, 1 distal fragment and 5 shaft fragments.Metatarsal remains are represented by 2 distal frag-
65Geo.Alp, Vol. 2, 2005
RADIUS right right right left right right left (Gliozzi et al., 1993)
greatest length 270 - - - - - - min 206 – max 237
transverse diameter
of the proximal end
54 44 42 50 - - min 39 – max 44.7
antero-posterior diameter
of the proximal end
29 23 25 27 - - - min 21 – max 24.5
transverse diameter
at half length of the shaft
31 20 - - - - - min 22 – max 26.5
antero-posterior diameter
at half length of the shaft
19 10 - - - - min 12.5 – max 16
transverse diameter
of the distal end
- - - - 43 46 45 min 27 – max 39
antero-posterior diameter
of the distal end
- - - - 29 33 30 min 25.5 – max 29.1
Tab. 4: Measurements (mm) of the humerus of Cervus elaphus siciliae from Acquedolci compared with the dimensions of the remainsdescribed by Gliozzi et al., 1993.
RADIUS right right right left right right left (Gliozzi et al., 1993)
greatest length 270 - - - - - - min 206 – max 237
transverse diameter
of the proximal end
54 44 42 50 - - min 39 – max 44.7
antero-posterior diameter
of the proximal end
29 23 25 27 - - - min 21 – max 24.5
transverse diameter
at half length of the shaft
31 20 - - - - - min 22 – max 26.5
antero-posterior diameter
at half length of the shaft
19 10 - - - - min 12.5 – max 16
transverse diameter
of the distal end
- - - - 43 46 45 min 27 – max 39
antero-posterior diameter
of the distal end
- - - - 29 33 30 min 25.5 – max 29.1
Tab. 5: Measurements (mm) of the radius of Cervus elaphus siciliae from Acquedolci compared with the dimensions of the remainsdescribed by Gliozzi et al., 1993.
66 Geo.Alp, Vol. 2, 2005
TIBIA left left left right (Gliozzi et al., 1993)
transverse diameter of the proximal end 63 59 - - min 48.5 – max 63.5
antero-posterior diameter of the proximal end - 57 - - min 50 – max 64
transverse diameter at half length of the shaft 26 - 23 - min 21 – max 28.6
antero-posterior diameter at half length of the shaft 24 - 20 - min 19 – max 27.5
transverse diameter of the distal end - - 37 46 min 33 – max 49
antero-posterior diameter of the distal end - - 30 32 min 24.5 – max 35
Tab. 6: Measurements (mm) of the tibia of Cervus elaphus siciliae from Acquedolci compared with the dimensions of the remains de-scribed by Gliozzi et al., 1993.
METATARSAL right left (Gliozzi et al., 1993)
transverse diameter
of the distal end
35 34 min 29 – max 35
antero-posterior diameter
of the distal end
24 22 min 18 – max 23.4
Tab. 9: Measurements (mm) of the metatarsal of Cervus elaphus siciliae from Acquedolci compared with the dimensions of the re-mains described by Gliozzi et al., 1993.
METACARPAL right left left left (Gliozzi et al., 1993)
greatest lenght 221 222 - - min 195 – max 226
transverse diameter
of the proximal end
33 34 33 - min 29 – max 35.6
antero-posterior diameter
of the proximal end
23 24 24 - min 19 – max 26
transverse diameter
at half length of the shaft
18 20 - - min 16 – max 23.7
antero-posterior diameter
at half length of the shaft
21 22 - - min 18 – max 22.4
transverse diameter
of the distal end
34 35 - 34 min 27 – max 38.8
antero-posterior diameter
of the distal end
22 22 - 22 min 19 – max 24.4
Tab. 8: Measurements (mm) of the metacarpal of Cervus elaphus siciliae from Acquedolci compared with the dimensions of the re-mains described by Gliozzi et al., 1993.
PODIAL BONES SCAPH.
rightLUNAR
rightCUNEIF.
leftASTR.
rightASTR.
left(Gliozzi et al., 1993)
transverse diameter 27 23 30 - - –
lateral lenght - - - 47 - min 41.3 – max 47.8
medial lenght - - - 44 42 –
transverse diameter of the distal end - - - 28 27 –
lateral antero-posterior diameter - - - 25 - min 24 – max 32
medial antero-posterior diameter - - - 25 21 –
Tab. 7: Measurements (mm) of the podial bones of Cervus elaphus siciliae from Acquedolci compared with the dimensions of the re-mains described by Gliozzi et al., 1993.
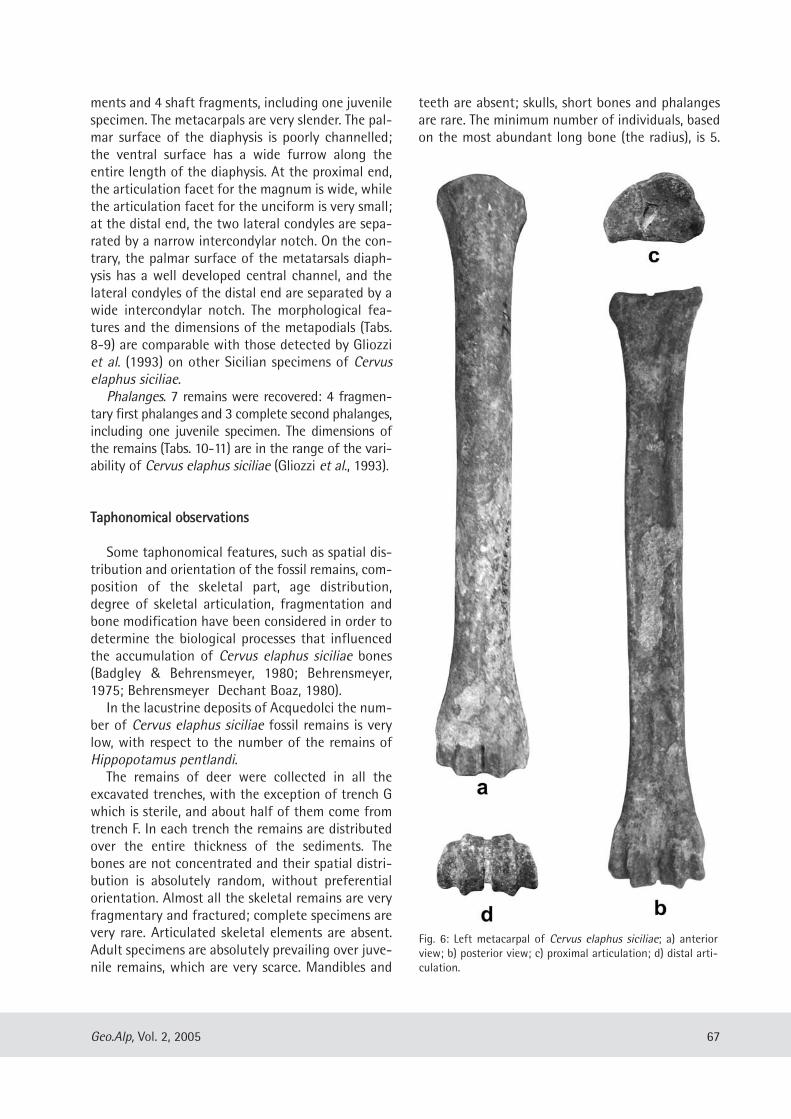
ments and 4 shaft fragments, including one juvenilespecimen. The metacarpals are very slender. The pal-mar surface of the diaphysis is poorly channelled;the ventral surface has a wide furrow along theentire length of the diaphysis. At the proximal end,the articulation facet for the magnum is wide, whilethe articulation facet for the unciform is very small;at the distal end, the two lateral condyles are sepa-rated by a narrow intercondylar notch. On the con-trary, the palmar surface of the metatarsals diaph-ysis has a well developed central channel, and thelateral condyles of the distal end are separated by awide intercondylar notch. The morphological fea-tures and the dimensions of the metapodials (Tabs.8-9) are comparable with those detected by Gliozziet al. (1993) on other Sicilian specimens of Cervuselaphus siciliae.
Phalanges. 7 remains were recovered: 4 fragmen-tary first phalanges and 3 complete second phalanges,including one juvenile specimen. The dimensions ofthe remains (Tabs. 10-11) are in the range of the vari-ability of Cervus elaphus siciliae (Gliozzi et al., 1993).
TTaapphhoonnoommiiccaall oobbsseerrvvaattiioonnss
Some taphonomical features, such as spatial dis-tribution and orientation of the fossil remains, com-position of the skeletal part, age distribution,degree of skeletal articulation, fragmentation andbone modification have been considered in order todetermine the biological processes that influencedthe accumulation of Cervus elaphus siciliae bones(Badgley & Behrensmeyer, 1980; Behrensmeyer,1975; Behrensmeyer Dechant Boaz, 1980).
In the lacustrine deposits of Acquedolci the num-ber of Cervus elaphus siciliae fossil remains is verylow, with respect to the number of the remains ofHippopotamus pentlandi.
The remains of deer were collected in all theexcavated trenches, with the exception of trench Gwhich is sterile, and about half of them come fromtrench F. In each trench the remains are distributedover the entire thickness of the sediments. Thebones are not concentrated and their spatial distri-bution is absolutely random, without preferentialorientation. Almost all the skeletal remains are veryfragmentary and fractured; complete specimens arevery rare. Articulated skeletal elements are absent.Adult specimens are absolutely prevailing over juve-nile remains, which are very scarce. Mandibles and
teeth are absent; skulls, short bones and phalangesare rare. The minimum number of individuals, basedon the most abundant long bone (the radius), is 5.
67Geo.Alp, Vol. 2, 2005
Fig. 6: Left metacarpal of Cervus elaphus siciliae; a) anteriorview; b) posterior view; c) proximal articulation; d) distal arti-culation.
68 Geo.Alp, Vol. 2, 2005
FIRST PHALANX (Gliozzi et al., 1993)
transverse diameter
of the proximal end
18 - - 17 –
antero-posterior diameter
of the proximal end
22 - - 22 –
transverse diameter
at half length of the shaft
13 15 12 - min 11 – max 15.7
antero-posterior diameter
at half length of the shaft
17 18 - - –
transverse diameter
of the distal end
- 17 15 - –
antero-posterior diameter
of the distal end
- 10 13 - –
Tab. 10: Measurements (mm) of the first phalanx of Cervus elaphus siciliae from Acquedolci compared with the dimensions of theremains described by Gliozzi et al., 1993.
Cervus elaphus siciliae Hippopotamus pentlandi
Number of Remains 104 3.016
Spatial distribution random
absence of orientation
absence of concentration
random
absence of orientation
extreme concentration
Minimum Number of Individuals 5 33
Age distribution predominantly adult, rare juvenile adult, juvenile, infantile
Skeletal part composition absence of mandibles and teeth
rare skull, short bones and phalanges
all skeletal parts represented
Skeletal articulation disarticulated bones anatomical connection
Degree of fragmentation very high very low
Bone modification cracking, abrasion (not frequently) no
Tab. 12: Comparison between taphonomical features of Cervus elaphus siciliae and Hippopotamus pentlandi remains from Acque-dolci deposit (taphonomic data about Hippopotamus pentlandi from Bonfiglio, 1995).
SECOND PHALANX (Gliozzi et al., 1993)
greatest lenght 37 34 min 33 – max 38
transverse diameter
of the proximal end
18 16 –
antero-posterior diameter
of the proximal end
23 21 –
transverse diameter
at half length of the shaft
13 13 min 10 – max 15
antero-posterior diameter
at half length of the shaft
15 16 –
transverse diameter
of the distal end
14 14 –
antero-posterior diameter
of the distal end
19 20 –
Tab. 11: Measurements (mm) of the second phalanx of Cervus elaphus siciliae from Acquedolci compared with the dimensions of theremains described by Gliozzi et al., 1993.

Bone modifications are observed at about 20 % ofthe remains, showing traces of cracking (stage 1,according to Behrensmeyer, 1978) and/or abrasion.
A comparison between taphonomical features ofCervus elaphus siciliae and Hippopotamus pentlan-di remains from the Acquedolci deposit is shown inTab. 12.
From a taphonomical point of view, the smallnumber of recovered remains of Cervus elaphussiciliae, with respect to the extension of the depositand to the number of the hippo remains, their ran-dom distribution over the entire thickness of thedeposit, the lack of skeletal articulation, the pres-ence of selected skeletal elements and the degree offragmentation, indicate an allochthonous fossiliza-tion, although the slight traces of abrasion andcracking suggest a minimal transportation and/or ashort period of subaerial exposure.
The taphonomical analysis indicates that theremains are allochthonous and probably weredeposited in the lacustrine basin as fragments ofcarcasses from animals living in the area,
testifying, therefore, a different accumulationprocess in comparison with the remains ofHippopotamus pentlandi, which accumulated andfossilized “in situ”, in the lacustrine basin where thehippos have lived (Bonfiglio, 1995).
CCoonncclluussiioonn
The morphological and biometrical features ofthe remains are in the range of the variability ofCervus elaphus siciliae POHLIG, the Pleistoceneendemic deer of Sicily which is characterized by amoderately reduced size compared to the popula-tions of the Italian peninsula.
The small number of specimens belonging todeer, as well as those belonging to the other associ-ated species, if compared with the very large num-ber of the recovered hippo remains, probably is tocorrelate with the different accumulation processesof the remains and it does not reflect the real com-position of the faunal populations living in the area.
AAcckknnoowwlleeddggmmeennttss
Work supported by grants CoFin MURST 2003“Faunal turnover in Sicily during the two lastGlacial cycles”. Thanks to Dr. R. Sardella, for the crit-
ical reading of the manuscript and the precious ad-vises, and to Prof. K. Krainer, for the helpful sugges-tions in the revision of the English version.
RReeffeerreenncceess
Bada, J. L., Belluomini, G., Bonfiglio, L., Branca, M., Burgio,E., Delitala, L. (1991): Isoleucine epimerization ages ofquaternary mammals of Sicily. – Il Quaternario, vol. 4(1a): 5-11.
Badgley, C., Behrensmeyer, A. K. (1980): Paleoecology ofMiddle Siwalik sediments and faunas. – Palaeo geo -graphy, Palaeoclimatology, Palaeoecology, vol. 30:133-155.
Behrensmeyer, A. K. (1975): The taphonomy and paleoe-cology of Plio-Pleistocene vertebrate assemblages ofLake Rudolf, Kenya. – Museum of ComparativeZoology Bulletin Harvard, vol. 146: 473-578.
Behrensmeyer, A. K. (1978): Taphonomic and ecologicinformation from bone weathering. – Paleobiology,vol. 4(2): 150-162.
Behrensmeyer, A. K., Dechant Boaz, D. E. (1980): Therecent bones of Amboseli National Park, Kenya, inrelation to East African paleoecology. – In:Behrensmeyer A. K., Hill A. P. (eds.): Fossils in themaking, 72-93. University of Chicago Press,Chicago.
Bonfiglio, L. (1985): Prima campagna di scavo dei depo-siti a mammiferi pleistocenici dell’area della grotta diS. Teodoro (Acquedolci, Messina, Sicilia). – GeologicaRomana, vol. 22: 271-285.
Bonfiglio, L. (1987): Primi elementi di stratigrafia deltalus della grotta di S. Teodoro (Acquedolci, Messina,Sicilia). – Il Naturalista Siciliano, s. 4, vol. 10 (1-4):43-57.
Bonfiglio, L. (1989): Distribuzione quantitativa dei resti diHippopotamus sp. del deposito di bacino del talusdella grotta di S. Teodoro (Acquedolci, Messina,Sicilia). – Atti 3° Simposio di Ecologia e Paleoecologiadelle Comunità bentoniche: 299-317.
Bonfiglio, L. (1992): Campagna di scavo 1987 nel deposi-to pleistocenico a Hippopotamus pentlandi diAcquedolci (Sicilia nord-orientale). – Bollettino dellaSocietà Paleontologica Italiana, vol. 30 (2): 157-173.
Bonfiglio, L. (1995): Taphonomy and depositional settingof Pleistocene mammal-bearing deposits fromAcquedolci (North-Eastern Sicily). – Geobios, M. S.,vol. 18: 57-68.
Bonfiglio, L., Mangano, G., Marra, A. C., Masini, F. (2001):A new late Pleistocene vertebrate faunal complex
69Geo.Alp, Vol. 2, 2005

from Sicily (S. Teodoro Cave, North-Eastern Sicily,Italy). – Bollettino della Società PaleontologicaItaliana, vol. 40 (2): 149-158.
Bonfiglio, L., Marra, A. C., Masini, F., Pavia, M., Petruso, D.(2002): Pleistocene faunas of Sicily: a review. – In:Waldren W. H., Ensenyat J. A. (eds.): World Islands inPrehistory, International Insular Insular Investigations,428-436. BAR International Series, 1095.Archaeopress, Oxford.
Gliozzi, E., Malatesta, A, Scalone, E. (1993): Revision ofCervus elaphus siciliae Pohlig, 1893, Late Pleistoceneendemic deer of the Siculo-Maltese district. –Geologica Romana, vol. 29: 307-354.
Mangano G. (2000): Nuovi resti di elefante e revisione dialcuni resti di mammiferi del Pleistocene superioredella Sicilia nord-orientale. – Giornale di Geologia,Supplemento, serie 3a, vol. 62: 103-109.
Pavia, M. (2001): The Middle Pleistocene fossil avifaunafrom the “Elephas mnaidriensis Faunal Complex” ofSicily (Italy): preliminary results. – In: Cavarretta G.,Gioia P., Mussi M., Palombo M. R. (eds.): La Terra degliElefanti, 497-501. Consiglio Nazionale delle Ricerche,Roma.
70 Geo.Alp, Vol. 2, 2005
Manuscript submitted: December 14, 2004Revised manuscript accepted: April 8, 2005

Geo.Alp, Vol. 2, S. 71–76, 2005
EEXXCCAAVVAATTIIOONNSS OOFF 22000033 AATT TTHHEE SS.. TTEEOODDOORROO CCAAVVEE ((NNOORRTTHH--EEAASSTTEERRNN SSIICCIILLYY,, IITTAALLYY))::PPRREELLIIMMIINNAARRYY FFAAUUNNIISSTTIICC AANNDD SSTTRRAATTIIGGRRAAPPHHIICC DDAATTAA
Gabriella Mangano1, Laura Bonfiglio1 & Daria Petruso2
With 2 figures and 2 tables
AAbbssttrraaccttSystematic excavations have been carried out at the S. Teodoro Cave since 1998. Two trenches have been
excavated on the eastern side of the cave. The “1998 trench”, located between 9 and 13 meters from theentrance, covers an area of 25 m2. The “2002 trench” was located between 30 and 32 meters from the ent-rance and covers an area of 9 m2.
The 2003 excavations included the 2002 trench, which has been deepened for 1 m and enlarged by ad-ding new squares. Remains of Cervus elaphus siciliae, Bos primigenius siciliae, Elephas mnaidriensis, Crocutacrocuta spelaea, Equus hydruntinus, small mammals, birds, reptiles, invertebrates and plant remains havebeen recovered. The composition of the faunal assemblage and the lithologic features of the deposit, whichis composed of clayey sands and gravels, are similar to those recognized in the 1998 trench. Fragmentationof remains, damages on almost all large mammal bones and abundant hyena coprolites testify an intensehyena activity. Complete and not damaged remains of elephant and deer have also been collected and ac-tually represent a taphonomic novelty. A new sedimentary unit of clayey sands and gravels which does notcontain fossil remains has been detected in the southern part of the trench. The age and the environmentalsignificance of this new sedimentary unit are to be clarified.
71
1 Dipartimento di Scienze della Terra, Università degli Studi di Messina; e-mail: [email protected]; [email protected] Dipartimento di Geologia e Geodesia, Università degli Studi di Palermoe-mail: [email protected]
IInnttrroodduuccttiioonn
The San Teodoro Cave opens in Jurassic lime-stone at an altitude of 150 m a.s.l.; it has hugedimensions (about 60 m long, 20 m wide and up to20 m high) and a total surface of more than 1.000m2. In previous excavations the authors (Anca,1860; Vaufrey, 1928, 1929; Tricomi, 1938; Maviglia,1941; Graziosi, 1943, 1947; Graziosi and Maviglia,1946) distinguished an upper sedimentary unit,Late Glacial in age, containing human feedingremains (mammal bones) associated with lateUpper Palaeolithic (Epigravettian) stone artifacts(unit A in Bonfiglio et al., 2001), and a lower sedi-mentary unit (unit B in Bonfiglio et al., 2001) con-taining late Pleistocene endemic mammals.
The 1998 excavations were devoted to thereconstruction of the stratigraphy of the cave
deposits and to a better knowledge of the faunalassemblages, especially the older one. The 1998trench has been located on the eastern side of thecave at a square surface of 25 m2, between 9 and13 meters from the entrance (coordinates 9-13/E-I)(Fig. 1) and it has been deepened for 1.40 m. Theinvestigated unit B is composed of clayey sands andgravels containing a highly diverse assemblage ofvertebrates, invertebrates (molluscs) and plantremains. The large mammal assemblage which con-tains elephant (Elephas mnaidriensis), wild ox (Bosprimigenius siciliae), deer (Cervus elaphus siciliae),wild boar (Sus scrofa), wolf (Canis lupus), hyena(Crocuta crocuta spelaea), fox (Vulpes vulpes), asso-ciated with the equid Equus hydruntinus and thesmall mammal taxa Microtus (Terricola) ex gr. savii,Apodemus cf. sylvaticus, Erinaceus cf. europaeusand Crocidura cf. sicula, has been attributed to a

new faunal complex in the Pleistocene of Sicily,named “S. Teodoro Cave-Pianetti” faunal complex,which contain some endemic taxa surviving fromthe previous faunal complex (“Elephas mnaidriensisF.C.”) associated with non-endemic taxa (Equushydruntinus, Microtus (Terricola) ex gr. savii,Erinaceus cf. europaeus) (Bonfiglio et al., 2001).
The various evidences of cave frequentation byspotted hyena populations are the most prominenttaphonomic feature of this deposit. Evidence comesfrom the occurrence of several Crocuta skeletal ele-ments (skull, teeth, limb bones), an impressive quan-tity of coprolites, and from ubiquitous traces ofcrushing, gnawing, chewing and digestion that havebeen detected on almost all the large mammalremains (Bonfiglio et al., 1999, 2001). These tapho-nomic characters have been found so far only in afew cave deposits of the Italian peninsula (“Grottadei Moscerini”, Stiner, 1990-91; “Buca della Iena”,Pitti and Tozzi, 1971; Stiner, 1990-91; “GrottaGuattari”, Piperno and Giacobini, 1990-1991; Stiner,1990-91; “Tana delle iene”, Giaccio and Coppola,2000) and are actually a novelty for insular envi-ronments.
Geochemical and radiometric data are not avail-able for the deposits of the S. Teodoro Cave; the dis-persal to Sicily of the ground vole, which has a fos-sorial habit, and of horses, that prefer open land-scapes, might imply that a fully exposed connection(a temporary land bridge related to an eustatic sea-
level lowstand) existed, perhaps more than onceduring the last glaciation (Bonfiglio et al., 2002).
Pollen spectra from samples of coprolites fromunit B show the existence of a vegetation whichwas mainly dominated by grass with moderatearboreal taxa (Artemisia, Ephedra) and low percent-ages of mesophilous pollen taxa (Quercus, Betula,Abies, Alnus, Pistacia, among others) which depict aglacial landscape (Yll et al., in press).
During the 2002 excavations a new trench hasbeen located on the inner eastern side of the caveat a square surface of 9 m2, between 30 and 32 mfrom the entrance (coordinates 30-32/B-D) (Fig. 1),in order to verify the extension of the evidences ofthe frequentation by spotted hyenas in the innerpart of the cave. The 2002 trench has been deep-ened for about 40 cm. The sediments of unit B areagain composed of clayey sands and gravels andcontain several carbonatic concretion levels oftenincorporating fossil remains. Remains of the samelarge mammals collected during the 1998 excava-tions have been found together with small mam-mals (Microtus (Terricola) ex gr. savii, Crocidura cf.sicula, Myotis sp.), birds, reptiles and hyena copro-lites. The taphonomic features are very similar tothose detected in the 1998 trench and confirm theextension of the deposit as far as 32 m from theentrance of the cave, as well as the intense andextensive frequentation by hyenas (Mangano andBonfiglio, 2003).
72 Geo.Alp, Vol. 2, 2005
Fig. 1: Plan of the S. Teodoro Cave with the location of the excavation areas of 1998 and 2002-2003. The black arrow indicates thecave entrance.

EExxccaavvaattiioonnss ooff 22000033 ((GG.. MMaannggaannoo))
During the 2003 excavation the “2002 trench”has been deepened for 1 m and enlarged by addingtwo new squares on the southern side (coordinates33/E-F) (Fig. 1).
Stratigraphic dataBesides unit B containing the fossil remains, in
the southern area of the trench (squares 32B/C/D,and part of the squares 31B and 31C) a new unit ofclayey sands and gravels lacking fossil remains hasbeen detected. In this unit numerous white-yellow-ish pisolith-like elements with phosphatic composi-tion, diameters between 1 and 5 cm and lackingcrystalline structure, are scattered. A subvertical,quite irregular surface separates the fossiliferousunit B from the sterile deposit and suggests that anerosional phase cut the sterile deposit unit beforedeposition of unit B. Age and precise environmentalsignificance of this new sedimentary unit are to beclarified by deepening the trench.
Faunistic dataA total number of 543 large mammal bones and
1064 coprolites have been recovered (Tab.1). Almostall the skeletal remains are strongly fragmented, notarticulated and horizontally and vertically scatteredwithout preferential orientation. A very large num-ber of them (437) is represented by unidentifiablebone splinters. The composition of the skeletal partis characterized by the abundance of isolated teethand antlers (Tab. 2).
Cervus elaphus siciliae Pohlig, 1893. The endemicred deer of Sicily is the most aundant species: 8shed antlers, 13 antlers, 2 skull fragments, 3 hemi-mandibles, 18 teeth, 3 scapulae, 8 anterior limbbones, 9 metapodials, 5 podials and 6 phalangeshave been recovered. Morphological and biometri-cal features ascribe them to Cervus elaphus siciliae(Gliozzi et al., 1994). Particularly, two almost com-plete right shed antlers, different in size, have beenrecovered arranged side by side. They were totallycovered by carbonatic concretions. The largest oneis 1.20 m long and actually is the largest antler frag-ment belonging to this species so far recovered (Fig.2, a). Teeth grooves which cannot be ascribed cer-tainly to hyenas are present on the surface of theseantlers.
Elephas mnaidriensis A.L. Adams, 1870. The ele-phant is represented by a small fragment of amandible, 3 teeth, 2 vertebrae, 1 rib, 1 pelvis, 1anterior limb bone, 1 posterior limb bone and 1metapodial. Teeth include one large fragment ofincisor and two very worn molar fragments belong-ing to an adult specimen. A complete and not dam-aged right tibia, absolutely lacking typical damagesproduced by hyenas, is also preserved (Fig. 2, b).Morphological features and biometrical data allowto identify these specimens as Elephas mnaidriensis(Ambrosetti, 1968; Bonfiglio and Berdar, 1979).
Bos primigenius siciliae Pohlig, 1911. The endem-ic wild ox of Sicily is represented by 2 hemi-mandibles, 1 femur shaft, 1 tibia, 1 metatarsal bone
73Geo.Alp, Vol. 2, 2005
2002 trench 1998 trench
2003 excavations 2002 excavations 1998 excavations
Coprolites 1064 291 4271
Bones 543 132 2228
Unidentifiable bones 437 94 1686
Identifiable bones 106 38 542
Cervus elaphus siciliae 75 24 392
Bos primigenius siciliae 6 1 21
Equus hydruntinus 5 5 41
Elephas mnaidriensis 11 1 26
Crocuta crocuta spelaea 9 3 38
Sus scrofa 2 14
Vulpes vulpes 1 7
Canis lupus 1 3
Tab. 1: Number of recovered remains during the three excavation surveys at S. Teodoro Cave.

and 1 scaphoid bone. M/1, M/2 and M/3 are pre-served on mandibular fragments. M/3 has a slightlyinclined hypoconulid. The femur shaft belongs to ajuvenile specimen. The proximal end of the left tibiawas totally removed by crunching of the hyenas(Fig. 2, c). The dimensions of remains are within therange of the variation of Bos primigenius siciliae(Brugal, 1987).
Equus hydruntinus Regàlia, 1904. The small equidis represented by 1 right mandible fragment includ-ing the tooth row from M/2 to P/2 (Fig. 2, d), 2upper molars and 2 deciduous premolars. The uppermolars have a short protocone and a well marked plicaballin. In the lower cheek teeth the pli caballin isless evident.
Crocuta crocuta spelaea (Goldfuss, 1832). Thespotted hyena is the only carnivore recovered dur-ing the 2003 excavations. One small maxillar bonefragment, 4 heminandibles, 3 isolated teeth(canines) and 1 metapodial small fragment are pre-sent. Two right hemimandibles include the toothrow from M/1 to C (Fig. 2, e). Lower premolars aresturdy and oval in section.
The preliminary study of mammal remains indi-cates the predominance of the non-endemic speciesMicrotus (Terricola) ex gr. savii.
The recovered taxa belong to the “S. TeodoroCave-Pianetti” faunal complex, late Pleistocene inage, just recognized for the first time at the S.Teodoro Cave (Bonfiglio et al., 2001).
Almost all large mammal bones are fragmentaryand show typical damages produced by the activityof hyenas, such as strong fragmentation, raggededges, tooth grooves, tooth pits, digestion traces,
scooping out of cancellous bone (Sutcliffe, 1970;Brain, 1981; Bunn, 1983). Nevertheless, some com-plete and undamaged bones of elephant (tibia) anddeer (antlers) have also been recovered and actual-ly represent a taphonomic novelty.
CCoonncclluussiioonn
Fossil remains collected during the 2003 excava-tions at the S. Teodoro Cave belong to the sametaxa previously recovered.
Most of the remains are fragmentary andunequivocally damaged by hyenas, but some com-plete and undamaged bones are also present.
The cave is confirmed as a very large hyena denand the spotted hyena is assumed to be the maincollecting agent of the skeletal elements of unit B,although some new recognized features could indi-cate the existence of a different accumulationprocess of the faunal remains.
A new sterile sedimentary unit has been discov-ered, but its age and environmental significance areto be clarified.
AAcckknnoowwlleeddggmmeennttss
Work supported by grants CoFin MURST 2003“Faunal turnover in Sicily during the two last Glacialcycles” . The excavations have been supported byUniversity of Messina (2003, extraordinary con-tribute to L. Bonfiglio) and by Acquedolci
74 Geo.Alp, Vol. 2, 2005
Cervus elaphus
siciliae
Bos
primigenius
siciliae
Equus
hydruntinus
Elephas
mnaidriensis
Crocuta
crocuta
spelaea
shed antlers 8
antlers 13
skull 2 1
mandible 3 2 1 1 4
teeth 18 4 3 3
axis 3
girdles 3 1
anterior limb 8 1
posterior limb 2 1
metapodials 9 1 1 1
podials 5 1
phalanges 6
Tab. 2: Skeletal element distribution of large mammals recovered in the excavations of 2003 at S. Teodoro Cave.
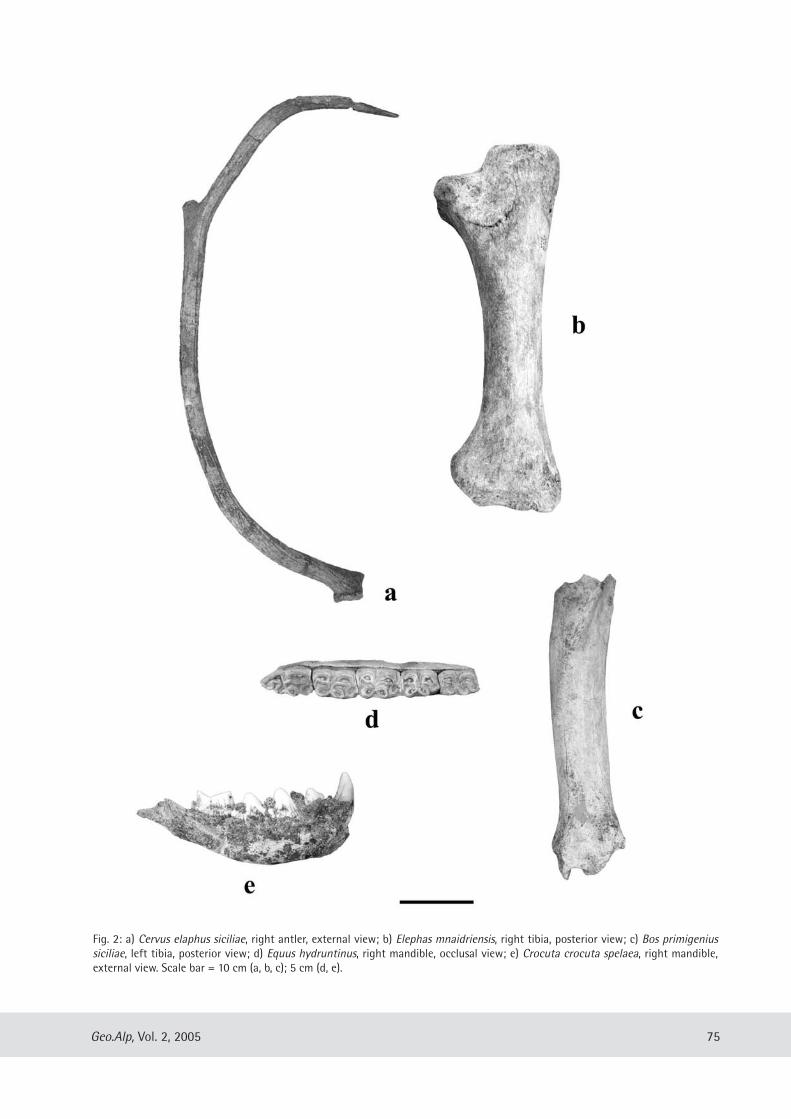
75Geo.Alp, Vol. 2, 2005
Fig. 2: a) Cervus elaphus siciliae, right antler, external view; b) Elephas mnaidriensis, right tibia, posterior view; c) Bos primigenius siciliae, left tibia, posterior view; d) Equus hydruntinus, right mandible, occlusal view; e) Crocuta crocuta spelaea, right mandible,external view. Scale bar = 10 cm (a, b, c); 5 cm (d, e).

Commune. Thanks are due to Dr. G.F. Villari,Superintendent to Archaeological and CulturalHeritage of Messina and to Dr. U. Spigo, responsibleof the Archaeological Service. A particular acknowl-edgment to Prof. A. Kotsakis for the critical readingof the text and for the revision of English version,and to Prof. K. Krainer for the helpful suggestions inthe revision of the final text.
RReeffeerreenncceess
Ambrosetti, P. (1968): The Pleistocene dwarf elephants ofSpinagallo (Siracusa, south eastern Sicily). – GeologicaRomana, vol. 7: 277-398.
Anca, F. (1860): Note sur deux nouvelles grottes ossifèresdécouvertes en Sicile en 1859. - Bulletin de la SociétéGéologique de France, vol. 17: 684-695.
Bonfiglio, L., Berdar, A. (1979): Gli elefanti delle ghiaiepleistoceniche di Messina. - Quaternaria, vol. 21:139-177.
Bonfiglio, L., Mangano, G., Marra, A.C. (1999): LatePleistocene hyaena den from a large cave deposits ofSicily (Italy). - INQUA XV International Congress,Durban 3-11 August 1999, abstract book: 27-28.
Bonfiglio, L., Mangano, G., Marra, A.C., Masini, F. (2001):A new Late Pleistocene vertebrate faunal complexfrom Sicily (S. Teodoro cave, north-eastern Sicily,Italy). - Bollettino Società Paleontologica Italiana,vol. 40 (2): 149-158.
Bonfiglio, L., Mangano, G., Marra, A.C., Masini, F., Pavia,M., Petruso, D. (2002): Pleistocene Calabrian andSicilian bioprovinces. – Geobios, M. S., vol. 24: 29-39.
Brain, C. K. (1981): The hunters or the hunted? An intro-duction to African cave taphonomy. - pp. 1-365,Chicago University Press, Chicago.
Brugal, J.P. (1987): Cas de „nanisme“ insulaire chez l’au-rochs. - In: 112th Congrès National des Sociétéssavants, Lyon, vol. 2: 53-66.
Bunn, H.T. (1983): Comparative analysis of modernbone assemblages from a San hunter-gatherer campin the Kalahari Desert, Botswana, and from a spot-ted hyena den near Nairobi, Kenya. - Animal andArchaeology, BAR International Series, vol. 163:143-148.
Giaccio, B., Coppola, D. (2000): Note preliminari sul con-testo stratigrafico e paleoecologico del sito “Tana delleiene” (Ceglie Messapica, Brindisi, SE Italia). - IlQuaternario, vol. 13 (1/2): 5-20.
Ghiozzi, E., Malatesta, A., Scalone E. (1994): Revision ofCervus elaphus siciliae Pohlig, 1893, Late Pleistocene
endemic deer of the Siculo-Maltese district. –Geologica Romana, vol. 29: 307-353.
Graziosi, P. (1943): Gli scavi dell’Istituto Italiano diPaleontologia Umana nella grotta di S. Teodoro(Messina): nota preliminare. - Atti Società ToscanaScienze Naturali, Memorie, vol. 52: 82-99.
Graziosi, P. (1947): Gli uomini paleolitici della grotta di S.Teodoro (Messina). - Rivista di Scienze Preistoriche,vol. 2 (2-3): 123-224.
Graziosi, P., Maviglia, C. (1946): La grotta di S. Teodoro(Messina). - Rivista di Scienze Preistoriche, vol. 1 (4):227-283.
Mangano, G., Bonfiglio, L. (2003): Campagna di scavo2002 nei depositi pleistocenici della Grotta diS. Teodoro (Acquedolci, Messina – Sicilia nord-orien-tale). Giornate di Paleontologia 2003, Alessandria22-25 maggio 2003, abstract book: 31.
Maviglia, C. (1941): Scheletri umani del Paleolitico supe-riore rinvenuti nella grotta di S.Teodoro. - Archivio perl’Antropologia e l’Etnologia, vol. 70: 94-104.
Piperno, M., Giacobini, G. (1990-1991): A taphonomicstudy of the paleosurface of Guattari Cave (MonteCirceo, Latina, Italy). - Quaternaria Nova, vol. 1:143-161.
Pitti, C., Tozzi, C. (1971): La Grotta del Capriolo e la Bucadella Iena presso Mommio (Camaiore, Lucca). - Rivistadi Scienze Preistoriche, vol. 26 (2): 213-258.
Stiner, M. (1990-1991): The Guattari faunas then andnow. - Quaternaria Nova, vol. 1: 163-192.
Sutcliff, A. J. (1970): Spotted hyena: crusher, gnawer,digestor and collector of bones. – Nature, vol. 227:1110-1113.
Tricomi, G. (1938): Cenni su un recente assaggio nellagrotta di S. Teodoro in provincia di Messina. -Bollettino Società Scienze Naturali ed Economiche,vol. 20: 1-4.
Vaufrey, R. (1928): Le Paléolithique Italien. - Archives del’Institute de Paléontologie Humaine, Mémoire, vol. 3:1-196.
Vaufrey, R. (1929): Les éléphants nains des iles mediter-ranèennes et la question des isthmes pléistocènes. -Archives de l’Institute de Paléontologie Humaine,Mémoire, vol. 6: 1-220.
Yll, R., Carrion, J. S., Marra, A.C., Bonfiglio, L. (in press):Pollen in Late Pleistocene hyena coprolites from SanTeodoro Cave (Sicily, Italy). - Palaeogeography,Palaeoclimatology, Palaeoecology, Special volume,Elsevier.
76 Geo.Alp, Vol. 2, 2005
Manuscript submitted: December 14, 2004Revised manuscript accepted: April 8, 2005

Geo.Alp, Vol. 2, S. 77–90, 2005
LLOOWWEERR PPEERRMMIIAANN PPAALLAAEEOOIICCHHNNOOLLOOGGYY FFRROOMM TTHHEE OORROOBBIICC BBAASSIINN ((NNOORRTTHHEERRNN IITTAALLYY))
Giuseppe Santi
With 4 figures and 2 plates
AAbbssttrraaccttThe Lower Permian palaeoichnofauna of the South-Alpine region comes from the Collio Formation only,
and mainly crops out in the Orobic and Trompia basins. It consists of traces of vertebrates (amphibians andreptiles) and invertebrates (insects, arthropods, burrowing, probable myriapods, gastropods, freshwater jelly-fishes, conchostraceans and freshwater bivalves). It is poor in taxa and similar to the coeval ichnoassociationof Central Europe, N. America and Argentina. Inside the trophic pyramid relevant to the biodiversity of thelower unit of the Collio Fm., the top carnivore is absent. This role is partially occupied by reptiles (e.g. araeos-celids) having features similar to true lizards. Shifting from the sediments of the lower part of the Collio For-mation to the upper part, impoverishment of the ichnocoenosis composition is linked either to a climaticshift (from more humid towards drier), or to tectonic activity that prevented the persistence of biotope for-mation. The Upper Permian uplift marks a strong ichnofaunistic change with the introduction of Triassiccomponents.
77
Dipartimento di Scienze della Terra, Via Ferrata 1, 27100 Pavia (Italia). e-mail: [email protected]
IInnttrroodduuccttiioonn
In the South-Alpine region the continentalLower Permian is characterized by vertebrate andinvertebrate ichnofossils and by rare floral remains(macroplants, spores and pollen). They comealmost exclusively from the Collio Fm. croppingout in the Orobic and Trompia (= Collio) basins,with the exception of rare fossiliferous remainsfrom the Tregiovo Basin. The features, problemsand hypotheses relating to the composition, distri-bution and behavioural features of the trackmak-ers and the ichnocoenosis variation are also dif-ferent, moving from the lower “Collio” to theupper unit of the same formation, and from thisto the Upper Permian when the faunistic changeis profound. These aspects are very clear fromanalysing the fossil beds of the Collio Fm. in theOrobic Basin; this article will review the ichnofau-nistic assemblages from this basin and discusstheir significance and the problems inherent tothem.
BBrriieeff ssttrraattiiggrraapphhiiccaall ffrraammeewwoorrkk
For a long time it has been known that thePalaeozoic of the Alps is very poor in vertebrateremains, with the only exception being the ich-nites, which have recently become a great strati-graphical tool (Avanzini et al., 2001). Recentdetailed research on the Permian of Europe (main-ly in France and Germany) has enlarged ourknowledge, particularly of the invertebrates, andof the stratigraphical-chronological role played bythem (Gand et al., 2001 with references therein).The South-Alpine region is characterized by aseries of basins created from W to E, as inheritedHercynian structures have produced structuralhighs of a metamorphic or igneous nature(Cassinis & Perotti, 1994; Cassinis et al., 1999 withreferences therein; Perotti, 1999). The main basinsare the Orobic Basin and, to the east, the TrompiaBasin (Fig. 1), but other smaller basins are impor-tant for their ichnofossil content (Tregiovo Basin,Tione Basin) (Conti et al., 1997).

Permian sediments occur in two tectono-sedi-mentary cycles separated by an uncertain age gap(between 14 and 25 My, according to the mostrecent data in Cassinis et al., 2002a). The first cycle,of ?Upper Carboniferous–Lower Permian age, iscomposed of a continental succession of volcanicdeposits (from intermediate to acid chemistry) andby alluvial-to-lacustrine sediments that comprisethe Basal Conglomerate, the Collio Fm, theTregiovo Fm, the Ponteranica Conglomerate, theDosso dei Galli Conglomerate and the AucciaVolcanics. The second cycle is assigned to theUpper Permian and is composed of the reddishclastic deposits of the Verrucano Lombardo-Val
Gardena Sandstone complex (Fig. 2). With theirdeposition the Palaeozoic ended.
The question of the use of a two- or three-foldsubdivision of the Permian System for dating ofcontinental successions has been debated for along time. A detailed discussion relating to datingof the Permian continental beds in the South-Alpine region was recently carried out by Cassinis(2003), Cassinis and Ronchi (2001) and Cassinis etal. (2002b). The traditionally adopted Permian sub-division for research in the South-Alpine area is“Lower Permian” (from about the Asselian toKungurian) and “Upper Permian” (from the
78 Geo.Alp, Vol. 2, 2005
Fig. 1: Schematic non-palinspastic section of the main Permian basin distribution of the South-Alpine region (Conti et al., 1997,mod.). 1 – Pre-Permian basement, 2 – Volcanic deposits, 3 – Clastic units of the first cycle of sedimentation (Basal Conglomerate,Collio Fm, Ponteranica Conglomerate), 4 – Verrucano Lombardo-Val Gardena Sandstone complex, 5 – Bellerophon Fm.
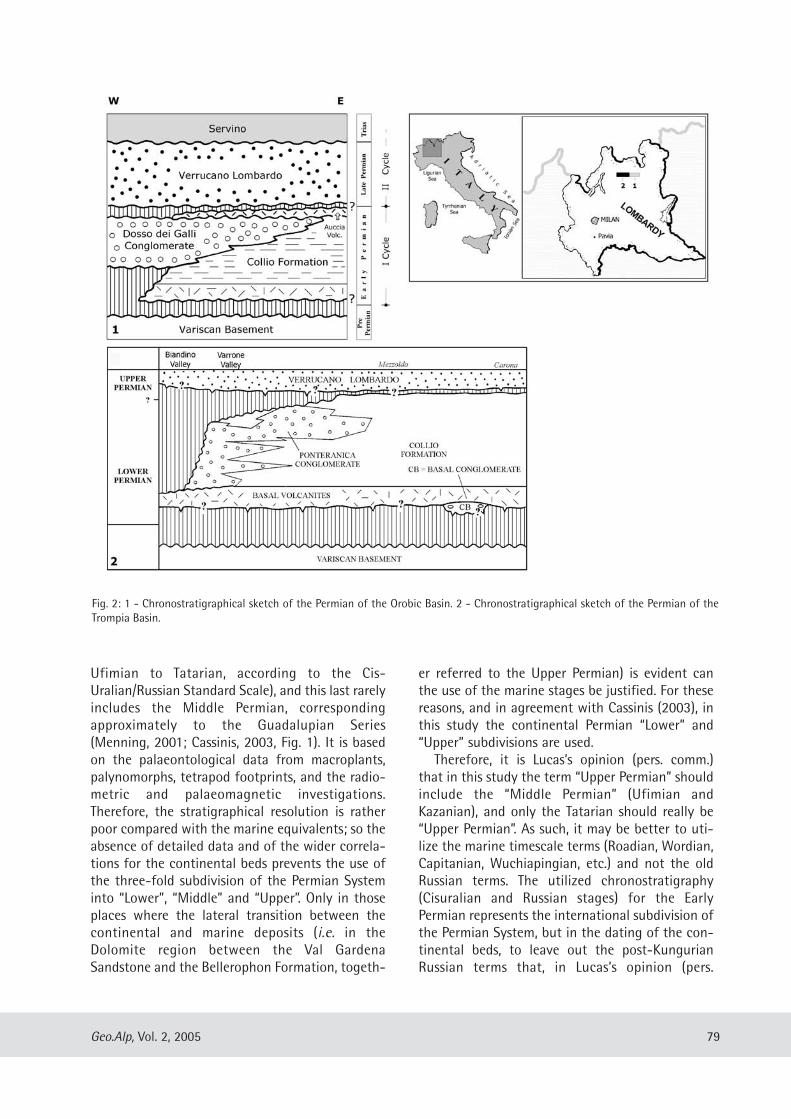
Ufimian to Tatarian, according to the Cis-Uralian/Russian Standard Scale), and this last rarelyincludes the Middle Permian, correspondingapproximately to the Guadalupian Series(Menning, 2001; Cassinis, 2003, Fig. 1). It is basedon the palaeontological data from macroplants,palynomorphs, tetrapod footprints, and the radio-metric and palaeomagnetic investigations.Therefore, the stratigraphical resolution is ratherpoor compared with the marine equivalents; so theabsence of detailed data and of the wider correla-tions for the continental beds prevents the use ofthe three-fold subdivision of the Permian Systeminto “Lower”, “Middle” and “Upper”. Only in thoseplaces where the lateral transition between thecontinental and marine deposits (i.e. in theDolomite region between the Val GardenaSandstone and the Bellerophon Formation, togeth-
er referred to the Upper Permian) is evident canthe use of the marine stages be justified. For thesereasons, and in agreement with Cassinis (2003), inthis study the continental Permian “Lower” and“Upper” subdivisions are used.Therefore, it is Lucas’s opinion (pers. comm.)
that in this study the term “Upper Permian” shouldinclude the “Middle Permian” (Ufimian andKazanian), and only the Tatarian should really be“Upper Permian”. As such, it may be better to uti-lize the marine timescale terms (Roadian, Wordian,Capitanian, Wuchiapingian, etc.) and not the oldRussian terms. The utilized chronostratigraphy(Cisuralian and Russian stages) for the EarlyPermian represents the international subdivision ofthe Permian System, but in the dating of the con-tinental beds, to leave out the post-KungurianRussian terms that, in Lucas’s opinion (pers.
79Geo.Alp, Vol. 2, 2005
Fig. 2: 1 - Chronostratigraphical sketch of the Permian of the Orobic Basin. 2 - Chronostratigraphical sketch of the Permian of theTrompia Basin.

comm.), are only the regional stages for the marinetimescale, is more difficult for the reasonsadvanced above. Fig. 3 shows the different scalesof the Permian stratigraphy.
In the classic succession of the Trompia Valley(Collio Basin) the COLLIO FORMATION was deposited onvolcaniclastic rocks (ignimbrites) which do notcrop out with continuity within the Orobic Basin,but are abundant in other areas (e.g. in theAcquaduro Valley –Introbio- and in the CedrinoPass) (Sciunnach, 2001) and in the mainly “berga-mask” sector of the same basin (Jadoul et al.,2000). Other subdivisions of lithofacies have beenproposed on a petrographical basis by Cassinis etal. (1988), Cadel et al. (1996), Forcella et al. (2001)and Sciunnach (2001). The Collio Fm. can be infor-mally subdivided into two units: the lower one iscomposed of grey-green and black sandstones andsiltstones, while the upper unit is defined by main-ly reddish sandstones and pelites of volcanic ele-ments with quartz, plagioclase and muscovite. It iswell stratified and locally contains some conglom-eratic beds. The typical arenaceous zones frequent-ly contain fragments of black clay (clay chips) anddisplay planar lamination, while in the peliticintervals there are different structures such as mud
cracks, raindrop imprints, ripple marks and fossilplant remains, as well as vertebrate and inverte-brate ichnites.This formation is interfingered with the
Ponteranica Conglomerate (Casati & Gnaccolini,1965, 1967). Utilising the fossils collected in theTrompia Basin, the Collio Fm. is referred to theLower Permian based on chronological data pro-vided by macroflora (Geinitz, 1869; Jongmans,1960; Remy & Remy, 1978; Kozur, 1981; Visscher etal., 1999), pollen (Clement-Westerholf et al., 1974;Cassinis & Doubinger, 1991, 1992) and tetrapodfootprints (Ceoloni et al., 1987; Conti et al., 1991,1997), and also for its position below the angularunconformity ascribed to the main post-Saalianphase (Palatine) of the Hercynian orogenesis.
VVeerrtteebbrraattee aanndd iinnvveerrtteebbrraattee iicchhnnooccooeennoosseess ooff tthheeOOrroobbiicc BBaassiinn
In Italy, early knowledge of vertebrate foot-prints from the Collio Fm. in the Trompia Valleywas advanced by Geinitz (1869) and Curioni(1870). Later, these fossils were studied by Gümbel(1880); the same ichnofauna from the Orobic Basinwas analysed by Dozy (1935) and later re-exam-
80 Geo.Alp, Vol. 2, 2005
Fig. 3: Permian stratigraphy (SCPS = Sub-Commission of Permian stratigraphy) (Vachard & Argyriadis, 2002. mod.)

ined by Haubold (1971). The studies of Berruti(1968), Haubold (1996, 2000), Haubold & Stapf(1998), Casati & Gnaccolini (1967), Ceoloni et al.(1987), Conti et al. (1991, 1997, 1999), Nicosia etal. (2000) and Santi & Krieger (2001) haveadvanced our knowledge of the vertebrate ichno-fauna of the Lower Permian. Footprints from boththe Orobic Basin and the Trompia Valley are ofamphibians and reptiles, and they come from dif-ferent parts of the volcano-sedimentary depositsof the Collio Formation (Conti et al., 1991; Santi,2003) relating to main vegetated areas, to otheralluvial zones, to more emergent humid areas, andothers with shallow water.Together with small- to medium-sized verte-
brates, lived insects and arthropods (BifurculapesHitchcock, 1858, Dendroidichnites elegansDemathieu, Gand & Toutin-Morin, 1992, cfr.Heteropodichnus variabilis Walter, 1983,Eisenachichnus sp. (= Secundumichnus), Tambiaspiralis Müller, 1956, Permichnium Guthörl, 1934,burrowing invertebrates (?Scoyenia White, 1929),gastropods (Paleobullia sp. vel. ?Cochlea sp.), prob-ably myriapods and some unidentified trails,bivalves (Anthracosiidae), small crustaceans(“Estheria”) and freshwater jellyfish (Medusinalimnica Müller, 1978 and Medusina atava (Pohlig,1892, Walcott, 1898) (Ronchi & Santi, 2003) (Pl. 1).Up to now, from these former data the compo-
sition of the invertebrate ichnocoenosis shows: (a)imprints are typically of freshwater animals, (b) adominance of surface traces and not infaunal bur-rows, (c) low biodiversity, (d) a lack of monospeci-fity, and (e) the ichnodiversity and the taxonomiccomposition suggest a terrestrial-freshwater ori-gin.
The tetrapod ichnofauna of the Collio Basinconsists of: Batrachichnus sp., Camunipes cassinisiCeoloni et al., 1987, Amphisauropus imminutusHaubold, 1970, Amphisauropus latus Haubold,1970, Varanopus curvidactylus Moodie, 1929,Dromopus lacertoides (Geinitz, 1861), Dromopusdidactylus Moodie, 1930 and Ichniotherium cottae(Pohlig, 1885). That of the Orobic Basin is com-posed of: “Batrachichnus” salamandroides (Geinitz,1861), Camunipes cassinisi Ceoloni et al., 1987,Amphisauropus imminutus Haubold, 1970,Amphisauropus latus Haubold, 1970, Varanopuscurvidactylus Moodie, 1929 and Dromopus lacer-toides (Geinitz, 1861) (Pl. 2). The ichnocoenoses re-
enter in the so-called “red-bed ichnofacies”(defined as a variety of fluvial, deltaic, lacustrineand marginal marine environments; Haubold &Lucas, 1999), typically different from the“Chelichnus ichnofacies” related to the desertenvironment and aeolian facies (Lockley et al.,1994; Lockley & Meyer, 2000; Lucas, 2002). A great affinity between the ichnocoenoses of
the two basins is evident, with the only exceptionbeing Ichniotherium cottae and Dromopus didac -tylus presenting together inside the Collio Basin,but lacking in the Orobic Basin. This last ich-nospecies is present not only in the highest strataof the Collio Fm. in the Trompia Valley, but it is alsoa monotypic taxon of the Tregiovo Basin (Conti etal., 1997; Nicosia et al., 2000). At present I. cottaeshould be a local taxon of the Trompia Basin.Besides, there is the problem linked to the validityof the ichnogenus Camunipes, namely if it effec-tively should be a true ichnogenus, or should beconsidered a synonym of Erpetopus. A discussionof this taxonomic problem is advanced by Haubold& Lucas (2001, 2003) and Santi (2004). On thewhole, the Lower Permian ichnocoenosis actuallyconsists of mostly reptiles and one amphibian(Batrachichnus); among the former we have a rel-evant “large” herbivore component, while the oth-ers are of smaller size.
The time interval into which the tetrapod ich-nofauna is limited is between 286/283 Ma at thebase and 278/273 Ma at the top (Avanzini et al.,2001). In agreement with the Permian subdivisioneffected by Menning (2001), this ichnoassociationmay belong to the Artinskian and Kungurian, butother scales (i.e. Harland et al., 1990; Odin, 1994;Gradstein & Ogg, 1996) consider these values to beSakmarian and upper Asselian. The South-Alpineichnoassociation has a similarity to that of NorthAmerica, with strong Wolfcampian affinities show-ing a great interaction between W-Central Europeand this continent.It is a mostly homogeneous association, but
also very poor in taxa, and even more reduced inthe highest strata of the Collio Fm. In the OrobicBasin, the passage between the lower unit of thisformation and the upper is marked among thetetrapod palaeoichnofauna by the absence ofBatrach ichnus, Camunipes (Erpetopus) and A.imminutus, and by the presence of only A. latus, D.lacertoides and V. curvidactylus, and among the
81Geo.Alp, Vol. 2, 2005

invertebrates, Dendroidichnites and Medusinaatava are present. In agreement with the “GlobalPermian series of the marine Permian System”, theabove-mentioned ichnoassociation is consideredcoeval with the “Lower Permian Cisuralian”(Cassinis et al., 2002).
On the whole, factors producing the taxonomiccompression of the Lower Permian palaeoichnofau-na are different (Lucas, 1998), but regionally, the“deposition time compression” hypothesis (Nicosiaet al., 2000) can be advanced on the basis of radio-metric data presented by Schaltegger & Brack(1999) in the volcanic beds at the base and at thetop of the Collio Fm. s.s. (= sedimentary “Collio”) inthe Trompia Valley. According to these authors,about 700 m of sediments were laid down in 4–5My: a very high rate linked to strong tectonic activ-ity. In my opinion this would prevent the establish-ment of useful biotopes for the survival of animals.A clear example is shown near to the Pizzo delDiavolo (Brembana Valley) neighbouring theBocchetta di Poddavista (“Podavit”) where the lowerunit of the Collio Fm. (600 m up) is well exposed. Inits lower portion abundant “signatures” of the tec-tonic activity are well evident. Repeated pyroclasticfall intercalations and the soft sediment deforma-tions (seismites), sedimentary dykes, “ball & pillow”and slumping structures, were probably triggered bysynsedimentary tectonics and frequent volcano-seismic activity. Only in the homogeneous silty-muddy part (last ten of metres) did the tectonic“peace” allow the development of more firmbiotopes. Only in this position were the taxa of the“orobic” ichnoassociation identified. Furthermore, the orogenic activity is not tthhee
cause, but oonnee cause of the taxonomic paucity,together with climatic change (Santi, 2004). Partially in agree with the opinion of Lucas
(pers. comm.) that the global paucity in Permianichnotaxa reflects the conservative nature of thefootprint structure (Santi, 2004), the ichnoassocia-tion of the South-Alpine region is very similar tothe other European and extra-European countries(see later): then a priori it is not possible to excludethe hypothesis that it could accurately reflect theoriginal vertebrate biodiversity. Overall, local geo-logical events could have played a crucial role forthe original biodiversity composition in this sectorof Palaeoeurope (“deposition time compression”hypothesis).
Paucity in taxa could depend on internal prop-erties and external conditions:aa)) linked to niche dimensions for vertebrates andinvertebrates. In fact, the species with the nar-rowest niches have high probabilities of specia-tion either because species are unstable andhave patchy populations, or because there arepotential new niches to invade through evolu-tionary divergences. The “Collio” area wasundoubtedly large and less ecologically diversi-fied, and this should favour extinction ratherthan speciation.
bb)) Species with small and patchy populations tendto isolate frequently; consequently this patternof species has a greater probability of extinction(Stanley, 2001). The orogenic forces and climat-ic changes probably operated above a very brit-tle biodiversity with low numbers, and deter-mined their extinction. Only the ability of sometaxa to disperse and to colonize differentbiotopes might have allowed them to survive(Amphisauropus, Dromopus, Varanopus), butprobably the attempt did not occur completelywithin an unstable framework (coeval orogene-sis + climatic changes).
In the palaeo-European domain, documentedexamples of terrestrial environments with fossilif-erous assemblages have been described (e.g.Debriette & Gand, 1990; Schneider, 1994; Gand etal., 1997 a, b, c; Eberth et al., 2000). It is notewor-thy that in many European Lower Permian basins,which can represent excellent analogues to thoseof the central Southern Alps, the facies distribu-tions and environmental settings record, from baseto top, an evolution from grey-black alluvial-to-lacustrine deposits to reddish flood-plain andplaya sediments. Over a large part of WesternEurope, Early Permian times were characterised bya climatic shift from warm, with alternating wetand dry seasons, to semi-arid, up to the very warmand hot conditions of the Late Permian (Ori, 1988;Dickins, 1993; Parrish, 1993; Golonka & Ford,2000). Thus, during the mid to late Early Permian(Artinskian–Kungurian?), a regional and geologi-cally rapid decrease in the rate of precipitation andthe onset of oxidising climatic conditions weresuggested by both lithofacies and biofacieschanges. In the Orobic Basin (at least in its westernsectors), the dominant alluvial-to-lacustrine dark-coloured facies pass quite abruptly, towards the
82 Geo.Alp, Vol. 2, 2005

stratigraphic top of the succession, to reddish finesediments. The former dark deposits suggest that ahigher groundwater level produced reducing con-ditions, while the red fines indicate muddy playaconditions with high evaporation rates and an oxicenvironment. A similar environmental–climatictransition could also be envisaged in the westernVal Trompia Basin, where the Collio Fm. fluvial andlacustrine scenario evolves from the proximal todistal alluvial-fan facies (Dosso dei GalliConglomerate) and up-section to the lateral andbioturbated, purple-red, fine sandstones and silt-stones (Pietra Simona Mb.). The consequenceswere, at the beginning of the Upper Permian, aclear change in fauna with more modern features(Conti et al., 1999); its origin is contained in theregional temporal gap which divides the first cyclefrom the second.
BBeehhaavviioouurraall ffeeaattuurreess oofftthhee EEaarrllyy PPeerrmmiiaann tteettrraappooddss
It seems opportune to talk about the problem ofthe behavioural features of the trackmakers. Therarity of fossil remains of vertebrates in the conti-nental deposits of the Permian of Central and SouthEurope makes a discussion about their behaviouralfeatures rather difficult, but the ichnoassociationcan be considered as a good starting point for thisgoal. The Lower Permian ichnoassociation of theSouth-Alpine zone reflects the vertebrate associa-tion living in this area of Palaeoeurope at the time,like those of France, Germany and also NorthAmerica and Argentina, with only rare exceptions ofelements considered as “local form” (i.e.Ichniotherium for the South-Alpine region) (Contiet al., 1999). Within the ichnoassociation of theSouth-Alpine region (Orobic and Trompia Basins),until now typical prints attributed to a top carni-vore are absent; either the trackmaker belonged toa population effectively reduced in number com-pared with the herbivores, or it was totally absent.Maybe during the Lower Permian of southernEurope, its specific role was partially occupied byother vertebrates. The low number of taxa (commonalso in the Lower Permian ichnoassociations fromother countries) suggests that the ichnodiversitycould be, if not real, then the almost complete com-position of the vertebrate biodiversity. Then theprints can be, if not an exact mirror, then at least a
significant indicator of the original vertebrate andinvertebrate biodiversity. This would not explainwhy the trophic pyramid should effectively be thathere carried out, but until now the ichnocoenosiscomposition and the frequency with which somefootprints are discovered (i.e. Batrachichnus is veryrare compared with the reptiles, and among theseAmphisauropus latus and Dromopus lacertoides areclearly much frequent in comparison withVaranopus) allows us to propose the hypothesesadvanced here. This is rather different to Lucas’sopinion (pers. comm.) referring to the Moenkopiichnoassociation from the Triassic of the USA:“…The tracks are almost all of archosaurs (chi-rotheres), but the bones from the same formationare almost all of amphibians…”.Not withstanding the paucity of taxa of the
tetrapod ichnofauna, the ichnocoenoses have notbeen utilised to examine the behavioural features ofthe trackmakers. A similar gap is also underlined byKramer et al. (1995) referring to the ichnites fromthe Coconino Sandstone (North America): “…behav-ioural aspects of extinct animals cannot be tested “(Brand, 1978 p. 81) (Kramer et al., 1995 p. 245).Furthermore, behavioural evidence from trackmak-ers can be discussed when studying “terminatedtrackways” sensu Kramer et al. (1995), or those thatsuddenly change direction. From the “orobic” LowerPermian beds come some data on the reptilian diet.Among the components of the ichnocoenosis, theDromopus trackmaker is commonly ascribed to thearaeoscelid, considered a consumer of small inverte-brates with exoskeletons. Figure 4A suggests thefollowing event sequence, pointing to a lack ofsuperimposition of walking-trail and footprints. Atrackmaker arthropod (Dendroidichnites elegans)is moving on a firm silty bed (point A). On its leftside a probable adult araeoscelid reptile, trackmak-er of Dromopus, is approaching. At point B thearthropod abruptly deviates towards its right side,probably trying an evasive manoeuvre; by thispoint the trail impression is not very clear, proba-bly because the trackmaker was alarmed andprogress was disordered. The final trackway-tractwas not preserved by the sediments, but we realisethat the araeoscelid preyed upon the arthropodwithout pursuing it. Figure 4B shows a clear “ter-minated trackway” sensu Kramer et al. (1995) of anarthropod (Heteropodichnus trackmaker) pursuedby a Dromopus one; traces of its trail abruptly dis-appear.
83Geo.Alp, Vol. 2, 2005

As witnessed by the prints upon the slabs in Fig.4, it is possible that the predator role in the LowerPermian of the South-Alpine region was playedpartially by these reptiles. Rare amphibians andmainly reptiles compose the tetrapod ichnocoeno-sis; it is an association with a paucity in taxa andcomprises herbivores from small size(Amphisauropus imminutus) to medium-large size(Amphisauropus latus). At present, large footprintsreferred to large vertebrates (i.e. such as theMiddle Permian pareiasaur Pachypes) have notbeen found. A top carnivore seems lacking. Thus, inthe Lower Permian of the South-Alpine region thetrophic pyramid was probably like this:PPrriimmaarryy ccoonnssuummeerr.. MMeeddiiuumm--ssiizzeedd hheerrbbiivvoorree:
cotylosaurs identified as the trackmakerAmphisauropus latus, a tetrapod of relatively largedimensions (the true “giant” of the association incomparison with the sizes of other trackmakers),with short and stumpy legs, probably strong and
adapted to support a relatively great weight. Thefrequency with which the A. latus footprints arefound is highest, so it represented the dominantanimal of “Collio” lands. Similar in size or possiblylarger was the Ichniotherium trackmaker (anedaphosaur pelycosaur), but as seen above, itspresence is very rare, and thus its role inside thetrophic pyramid is much diminished. SSeeccoonnddaarryy ccoonnssuummeerr.. CCaarrnniivvoorreess: the ichnologi-
cal association seems to lack typical footprintsattributed to this consumer. MMiixxeedd ddiieett.. OOppppoorrttuunniissttiicc ccoonnssuummeerrss: on the
whole these are small reptiles, morphologically andin their general structure similar to small lizards,also with autopodia features and with more or lesssharp teeth (Camunipes trackmakers). Their dietcould be similar to that of true lizards of smalldimensions, swallowing and biting anything eitherliving or dead. In this category should re-enter theDromopus trackmaker which, together with the
84 Geo.Alp, Vol. 2, 2005
Fig. 4: A - Interaction between the Dendroidichnites elegans Demathieu, Gand and Toutin-Morin, 1992 trail and Dromopus sp. foot-prints. Black arrows indicate the arthropod trail directions. B - “Terminated trackway” of cfr. Heteropodichnus variabilis Walter, 1983with Dromopus sp

Amphisauropus, is a common form, and less fre-quently that of Varanopus. A novel feature of anaraeoscelid trackmaker (Araeoscelis) is the lateraltemporal opening, which could have been closed inrelation to the skull extension as the consequenceof a more massive dentition (Carroll, 1988). Sucharaeoscelids could prey upon protein-bearingorganisms and consume some strong parts such astheir exoskeleton (arthropods), or small vertebrates(amphibians?) also.Thus, it does not seem that the Lower Permian
association of the South-Alpine area needs to bebalanced. It is possible that the araeoscelids andthe Dromopus trackmaker could have partiallyoccupied the small predator role.
CCoonncclluussiioonnss
Lower Permian palaeoichnoassociations of theOrobic Basin (the Collio Fm, the only fossiliferousunit), although poor in ichnotaxa, perfectly reflectthe ichnofaunistic panorama of Central Europe,North America and Argentina. On the basis ofactual knowledge, different conclusions can beadvanced.
11)) The ichnocoenosis has a similarity to thosefrom Central Europe, North America andSouth America (Argentina; Melchor &Sarjeant, 2004) (Lucas et al., 2004 and refer-ences therein).
22)) An impoverishment of the ichnofaunistic com-position, shifting from the lower unit to theupper unit of the Collio Fm., is recognized andprobably linked to the mutual action of tec-tonic activity and climatic change (from morehumid to drier). It is also probable that theoriginally low number of ichnotaxa might bedue to both internal properties of the biodi-versity and external conditions.
33)) The presence of Camunipes in the lower“Collio” may have created some problems,either evolutive or systematic, for its similarityto Varanopus and mainly with Erpetopus(Haubold & Lucas, 2003). If it should be con-sidered a separate ichnogenus, its presence inthe South-Alpine region could be interpretedas a local form (more frequent in the OrobicBasin, less so in Trompia Valley) as isIchniotherium cottae.
44)) The trophic pyramid relevant to the LowerPermian of the South-Alpine region does notseem balanced because of the lack of a topcarnivore. This role might have been occupiedby reptiles, some araeoscelids having featuressimilar to true lizards.
AAcckknnoowwlleeddggeemmeennttss
The author is deeply indebted to S.G. Lucas(Albuquerque, New Mexico) for his useful adviceand critical review of the text and S. Jones (Cardiff)for revision to English. This study was carried outwith a grant from FAR.
RReeffeerreenncceess
Avanzini, M., Ceoloni, P., Conti, M.A., Leonardi, G.,Manni, R., Mariotti, N., Mietto, P., Muraro, C., Nicosia,U., Sacchi, E., Santi, G., Spezzamonte, M. (2001):Permian and Triassic tetrapod ichnofaunal units ofNorthern Italy, potential contribution to continentalbiochronology. – In: Cassinis G. (ed.): Permian conti-nental deposits of Europe and other areas. Regionalreports and correlations. – Natura Bresciana,Monografia n. 25, 89-107.
Berruti, G. (1968): Osservazioni biostratigrafiche sulleformazioni continentali pre-quaternarie delle ValliTrompia e Sabbia. II. Sulla fauna fossile dellaFormazione di Collio (alta Val Trompia). – NaturaBresciana, Ann. Mus. Civ. St. Nat. Brescia, 6: 3-32.
Brand, L. R. (1978): Footprints in the Grand Canyon. –Origins, 5: 64-82.
Cadel, G., Cosi, M., Pennacchioni, G., Spalla, M.J. (1996): Anew map of the Permo-Carboniferous cover andvariscan metamorphic basement in the Central OrobicAlps, Southern Alps-Italy: structural and stratigraphicdata. – Mem. Sci. Geol. Univ. Padova, 48: 1-53.
Carroll, R.T. (1988): Vertebrate Paleontology andEvolution. – Freeman and Company, 698 pp.
Casati, P., Gnaccolini, M. (1965): Una nuova formazionepaleozoica dell’alta Val Brembana: la Formazione diPonteranica. – Riv. It. Paleont. Strat., 70: 791-804.
Casati, P., Gnaccolini, M. (1967): Geologia delle AlpiOrobie occidentali. – Riv. It. Paleont. Strat., 73:25-162.
Cassinis, G. (2003): Towards a stratigraphic referencetable for the continental Permian of Italy. – Boll. Soc.Geol. It., Vol. Spec. 2: 153-168.
85Geo.Alp, Vol. 2, 2005

Cassinis, G., Doubinger, J. (1991) : On the geologicaltime of the typical Collio and Tregiovo continentalbeds in the Southalpine Permian (Italy), and someadditional observations. – Atti Ticinensi di Scienzedella Terra, Pavia, 34: 1-20.
Cassinis, G., Doubinger, J. (1992) : Artinskian to Ufimianpalynomorph assemblages from the Central SouthernAlps, Italy and their regional stratigraphic implica-tions. – In Nairn A.E.M. & Koroteev, V. (Chief Eds):Contributions to Eurasian Geology, Int. Congr. PermianSystem of the World, Perm, Russia, 1991, OccasionalPubblication ESRI New Series 8b, Columbia Universityof South Carolina, Part I: 9-18.
Cassinis, G., Perotti, C.R. (1994). Iterazione strutturalepermiana tra la Linea delle Giudicarie ed i Bacini diCollio, Tione e Tregiovo (Sudalpino centrale, N. Italia).– Boll. Soc. Geol. It., 112 (1993): 1021-1036.
Cassinis, G., Ronchi, A. (2001) : Permian chronostratigra-phy of the Southern Alps (Italy) – an update. –Contribution to Geology and Palaeontology ofGondwana in honour of Helmut Wopfner, Cologne2001: 73-87.
Cassinis, G., Massari, F., Neri, C., Venturini, C. (1988): Thecontinental Permian in the Southern Alps (Italy). Areview. – Zt. Geol. Wiss., 16: 1117-1126.
Cassinis, G., Nicosia, U., Pittau, P., Ronchi, A. (2002 a):Palaeontological and radiometric data from thePermian continental deposits of the Central-EasternSouthern Alps (Italy), and their stratigraphic implica-tions. Mém. de l’Association des Géologues duPermien, 2: 53-74.
Cassinis, G., Nicosia, U., Lozovsky, V.R., Gubin, Y.M.(2002 b) : A view on the Permian continentalstratigraphy of the Southern Alps, Italy, and generalcorrelation with the Permian of Russia. –Permophiles, 40: 4-16.
Cassinis, G., Cortesogno, L., Gaggero, L., Massari, F., Neri,C., Nicosia, U., Pittau, P. (coord.) (1999): Stratigraphyand facies of the Permian deposits between easternLombardy and the western Dolomites. – Field tripguide-book Inter. Congr. on: “The continentalPermian of the Southern Alps and Sardinia (Italy).Regional reports and general correlations”. Brescia15-25 September 1999, 19-20.
Ceoloni, P., Conti, M.A., Mariotti, N., Mietto, P., Nicosia,U. (1987): Tetrapod footprints from Collio Formation(Lombardy, Northern Italy). – Mem. Sci. Geol. Univ.Padova, 39: 213-233.
Clement-Westerholf, J.A., Van der Eem, J.G.L.A., Van Erve.,A.W, Klasen, J.J., Schuurman, W.M.L., Visscher, H.(1974): Aspects of Permian, Triassic and Early Jurassic
palynology of Western Europe-A research project. –Geologie en Mijnbouw, 53: 329-341.
Conti, M.A., Mariotti, N., Mietto, P., Nicosia, U. (1991):Nuove ricerche sugli icnofossili della Formazione diCollio in Val Trompia (Brescia). – Natura Bresciana.Ann. Mus. Civ. St. Nat. Brescia, 26 (1989): 109-119.
Conti, M.A., Mariotti, N., Nicosia, U., Pittau, P. (1997):Selected bioevents succession in the continentalPermian of the Southern Alps (Italy): improvementsof intrabasinal and interregional correlations. – In:Dickins J.M., Yang Zunyi, Yin Hongfu, Lucas S.G. &Acharyya S.K. (eds): Late Palaeozoic and EarlyMesozoic Circum-Pacific events and their global cor-relation. – Cambridge University Press, 51-65.
Conti, M.A., Mariotti, N., Manni, R., Nicosia, U. (1999):Tetrapod footprints in the Southern Alps: anoverview. In: Cassinis G., Cortesogno L., Gaggero L.,Massari F., Neri C., Nicosia U., Pittau P. (coord.):Stratigraphy and facies of the Permian depositsbetween eastern Lombardy and the westernDolomites. – Field trip guide-book Inter. Congr. on:“The continental Permian of the Southern Alps andSardinia (Italy). Regional reports and general correla-tions”. Brescia 15-25 September 1999, 137-138.
Curioni, G. (1870): Osservazioni geologiche sulla ValTrompia. – R. Ist. Lomb. Sc. Lett. Arti – Mem. Ser. 3(2): 1-60.
Debriette, P., Gand, G. (1990) : Conséquences strati-graphiques et paléoenvironmentales de nouvellesobservations paléontologiques dans le Permien de lapartie occidentale du Bassin de Lodève (Sud duMassif central). – Géologie de la France, 1 : 19-32.
Dickins, J. M. (1993): Climate of the Late Devonian toTriassic. – Palaeog. Palaeocl. Palaeoec.,100: 89-94.
Dozy, J.J. (1935): Einige Tierfärhten aus dem UnterenPerm der Bergamasker Alpen. – Paläont. Z., 17 (1/2):45-55.
Eberth, D. A., Berman, D. V., Sumida, S. S., Hopf, H.(2000): Lower Permian terrestrial paleonvironmentsand vertebrate paleoecology of the Tambach Basin(Thuringia, Central Germany): the upland holy Grail.– Palaios, 15: 293-313.
Forcella, F., Sciunnach, D., Siletto, G. B. (2001): The LowerPermian in the Orobic Anticlines (Lombardy, SouthernAlps): criteria for field mapping towards a strati-graphic revision of the Collio Formation. In: CassinisG. (ed.): Permian continental deposits of Europe andother areas. – Regional reports and correlations -Natura Bresciana, Monografia n. 25: 51-57.
Gand, G., Garric, J., Schneider, J., Sciau, J., Walter, H.(1997a): Biocoenoses à méduses du Permien Français
86 Geo.Alp, Vol. 2, 2005

(Bassin de Saint-Affrique, Massif Central). Geobios,29 (4): 370-400.
Gand, G., Lapeyrie, J., Garric, J., Nel, A., Schneider, J.,Walter, H. (1997b): Découverte d’Arthropodes et debivalves inédits dans le Permien continental(Lodévois, France). Compte Rendu de l’Acad. desSciences de Paris, 325 : 891-898.
Gand, G., Kerp, H., Parsons, C. A., Martinez-Garcia E.(1997c): Palaeoenvironmental and stratigraphicaspects of animal traces and plant remains inSpanish Permian red beds (Pena Sagra, CantabrianMountains, Spain). Geobios, 30: 295-318.
Gand, G., Galtier, J., Garric, J., Schneider, J., Körner, F.,Nel, A., Béthoux, O., Demathieu, G., Broutin, J.,Hoernes, S., Kleeberg, R., Lapeyrie, J. (2001):Excursion n. 3: The Graissessac Carboniferous andLodève Permian Basins (Languedoc-France). FieldTrip Guidebook, Inter. Field Conf. on: “The strati-graphic and structural evolution of the LateCarboniferous to Triassic continental and marinesuccessions in Tuscany (Italy). Regional reports andgeneral correlations”. Siena 30 April-7 May 2001,1-21.
Geinitz, H.B. (1869): Über Fossile Pflanzenreste aus derDyas von Val Trompia. N. Jb. Mineral. Geol. Paläont.:456-461.
Golonka, J., Ford, D. (2000): Pangean (LateCarboniferous-Middle Jurassic) palaeoenvironmentsand lithofacies. Palaeog. Palaeocli. Palaeoec., 161: 1-34.
Gradstein, F.M., Ogg, J. (1986): A Phanerozoic time scale.Episodes, insert 19: 3-4, Nottingham.
Gümbel von, C.W. (1880): Geognostiche Mitteilungenaus den Alpen. VI. Ein geognosticher Streifzug durchdie Bergamasker Alpen. Sitzberg. K. Ak. Wiss.München, Math.-Nat. Kl., 10 (2): 164-240.
Harland, W. B., Armstrong, R. I., Cox, A. V., Craig, L. E.,Smith, A. G., Smith, D. G. (1990): A geological timescale. (1989). Cambridge University Press.
Haubold, H. (1971): Ichnia Amphibiorum et Reptiliorumfossilium. In : Kuhn O. (ed.), Handbuch derPalaeoherpetologie. G. Fisher-Verlag, 18, 1-124.Stuttgart-Portland, USA.
Haubold, H. (1996): Ichnotaxonomie und Klassificationvon Tetrapodenfährten aus dem Perm. Hallesches Jb.Geowiss., B 18: 23-88.
Haubold, H. (2000): Tetrapodenfährten aus dem Perm-Kenntnisstand und Progress 2000. Hallesches Jb.Geowiss., B 22: 1-16.
Haubold, H., Lucas, S.G. (1999): Early Permian tetrapodtracks-Preservation, taxonomy, and Euramerican dis-
tribution. – J. Vert. Paleont., 19, supplement to n. 3:49A-50A.
Haubold, H., Lucas, S.G. (2001): Die Tetrapodenfährtender Choza Formation (Texas) und das Artinsk-Alterder Redbed-Ichnofaunen des Unteren Perm. –Hallesches Jb. Geowiss., B 23: 79-108.
Haubold, H., Lucas, S.G. (2003): Tetrapod footprints ofthe Lower Permian Choza Formation at Castle Peak,Texas. – Paläont. Zt, 77 (2): 247-261.
Haubold, H., Stapf, H. (1998): The Early Permian tetra-pod track assemblage of Nierstein, Standen-bühlBeds, Rotliegend, Saar-Nahe Basin, SW Germany. –Hallesches Jb. Geowiss., B 20: 17-32.
Jadoul, F., Forcella, F., Bini, A., Ferliga, C. (coord.) (2000):Carta geologica della Provincia di Bergamo. – A curadi: Servizio Territorio della Provincia di Bergamo,Dipartimento di Scienze della Terra dell’Università diMilano-Centro di Studio per la geodinamica alpina edel Quaternario del C.N.R.-.
Jongmans, W. (1960): Die Karbonflora der Schweiz. mitEinem Beitrag von Ritter, E: Die Karbon-Vorkommender Schweiz. – Btg. Geol. Karte Schweiz, N.F. 108: 1-95.
Kozur, H. (1981): Weitere Beiträge zur Paläontologie undStratigraphie des Perms. – Geol. Paläont. Mitt.Innsbruck, 11(6): 243-257.
Kramer, J.M., Erickson, B.R., Lockley, M.G., Hunt, A.P.,Braddy, S.J. (1995): Pelycosaur predation in thePermian: evidence from Laoporus trackways from theCoconino Sandstone with description of a newspecies of Permichnium. – In: Lucas S.G., Heckert A.B.(eds.): Early Permian Footprints and Facies. NewMexico Museum of Natural History and ScienceBulletin N. 6, 246-249.
Lockley, M.G., Meyer, C.A. (2000): Dinosaur tracks andother fossil footprints of Europe. 323 pp., ColumbiaUniversity Press, New York.
Lockley, M.G., Hunt A.P., Meyer C.A. (1994): Vertebratetracks and the ichnofacies concept: implications forpalaeoecology and palichnostratigraphy. – InDonovan S. K. (ed.): The Paleobiology of trace fossils,241-268, Wiley and Sons, New York.
Lucas, S.G. (1998): Permian tetrapod biochronology. –Permophiles, 32: 17-24.
Lucas, S.G. (2002): Global Permian tetrapod footprintbiostratigraphy and biochronology. – Permophiles,41: 30-34.
Lucas, S.G., Lerner, A.J., Hunt A.P. (2004): Permian tetra-pod footprints from the Lucero uplift, Central NewMexico, and Permian footprint biostratigraphy. – In:Lucas S.G., Ziegler K.E (eds.): Carboniferous-Permiantransition at Carrizo Arroyo, Central New Mexico.
87Geo.Alp, Vol. 2, 2005

New Mexico Museum of Natural History & ScienceBulletin, 25: 291-300.
Melchor, R.N., Sarjeant, W.A.S. (2004): Small Amphibianand Reptile footprints from the Permian CarapachaBasin, Argentina. – Ichnos, 11 (1-2): 55-78.
Menning, M. (2001): A Permian time-scale 2000 andcorrelation of marine and continental sequencesusing the IIlawarra Reversal (265 Ma). In: Cassinis G.(ed.): Permian continental deposits of Europe andother areas. Regional reports and correlations. –Natura Bresciana, Monografia n. 25, 355-362.
Nicosia, U., Ronchi, A., Santi, G. (2000): Permian tetra-pod footprints from W Orobic Basin (Northern Italy).Biochronological and evolutionary remarks. –Geobios, 33 (6): 753-768.
Odin, G.S. (1994): Geological time scale (1994). –Compte Rendue de l’ Acad. de Sciences de Paris sérieII, 318: 59-71.
Ori, G.G. (1988): The nature of Permian rivers. – In:Cassinis, G., (Ed.), Permian and Permian-Triassicboundary in the South-Alpine segment of theWestern Tethys, and additional regional reports. –Mem. Soc. Geol. It., 34 (1986): 155-160.
Parrish, J. T. (1993): Climate of the SupercontinentPangaea. – Journal of Geology, 101: 215-233.
Perotti, C.R. (1999): Permian tectonic in the CentralSouthern Alps. – In: Cassinis G., Cortesogno L.,Gaggero L., Massari F., Neri C., Nicosia U., Pittau P.(coord.): Stratigraphy and facies of the Permiandeposits between eastern Lombardy and the westernDolomites. Field trip guide-book Inter. Congr. on:“The continental Permian of the Southern Alps andSardinia (Italy). Regional reports and general correla-tions”. Brescia 15-25 September 1999, 19-20.
Remy, W., Remy, R. (1978): Die Flora des Perms imTrompia-Tal und die Grenze Saxon/Thuring in denAlpen. – Argumenta Palaeobotanica, 5: 57-90.
Ronchi, A., Santi, G. (2003): Non marine biota from theLower Permian of the Central Southern Alps (Orobicand Collio Basins, N. Italy): a key for paleoenviron-ment. – Geobios, 36 (6): 749-760.
Santi, G. (2003): Early Permian tetrapod ichnology fromthe Orobic Basin (Southern Alps-Northern Italy).Data, problems, hypotheses. – Boll. Soc. Geol. It., Vol.Spec. 2: 59-66.
Santi, G. (2004): Tetrapod footprints in the LowerPermian of Southalpine (Northern Italy): is it differ-
ent Camunipes from Erpetopus and Varanopus? ––32nd Int. Geol. Congr., 2004, Abstract vol., pt. 1, Abs.128-16: p. 579.
Santi, G., Krieger, C. (2001): Lower Permian tetrapodfootprints from Brembana Valley – Orobic Basin-(Lombardy, Northern Italy). – Revue de Paléobiologie,20 (1): 45-68.
Schaltegger, U., Brack, P. (1999): Radiometric age con-straints on the formation of the Collio Basin(Brescian Prealps). – In: Cassinis G., Cortesogno L.,Gaggero L., Massari F., Neri C., Nicosia U., Pittau P.(coord.): Stratigraphy and facies of the Permiandeposits between eastern Lombardy and the westernDolomites. – Field trip guide-book Inter. Congr. on:“The continental Permian of the Southern Alps andSardinia (Italy). Regional reports and general correla-tions”. Brescia 15-25 September 1999, 71.
Sciunnach, D. (2001): The Lower Permian in the OrobicAnticline (Southern Alps, Lombardy): a review basedon new stratigraphic and petrographic data. – Riv. It.Paleont. Strat., 107 (1): 47-68.
Schneider, J. (1994): Environments, biotas and taphono-my of the Lower Permian lacustrine Niederhäslichlimestone, Döhlen Basin, Germany. – Trans. RoyalSoc. Edinburgh-Earth Sciences, 84: 453-464.
Stanley, S.M. (2001): Controls on rates of evolution. – In:Briggs D.E.G., Crowther P.R. (eds.), Palaeobiology II,166-171, Blackwell Publishing, Oxford.
Vachard, D., Argyriadis J. (2002): Quelques problèmes debiostratigraphie dans le Permien mèsogéen, des AlpesCarnique à la Turquie. – Mém. de l’Association desGéologues du Permien, 2: 75-93.
Visscher, H., Kerp, H., Clement-Westerholf, J.A., Looy, C.V.(1999): Permian floras of the Southern Alps. – In:Cassinis G., Cortesogno L., Gaggero L., Massari F., NeriC., Nicosia U., Pittau P. (coord.): Stratigraphy andfacies of the Permian deposits between easternLombardy and the western Dolomites. Field trip guide-book Inter. Congr. on: “The continental Permian of theSouthern Alps and Sardinia (Italy). Regional reportsand general correlations”. Brescia 15-25 September1999, 139-146.
88 Geo.Alp, Vol. 2, 2005
Manuscript submitted: November 26, 2004Revised manuscript accepted: April 25, 2005

PPllaattee 11
AA – Bifurculapes sp. Bocchetta di Poddavista, Orobic Basin. BB – cfr. Heteropodichnus variabilis Walter, 1983. MincuccoMt. Orobic Basin. CC - Paleobullia Götzinger & Becker, 1932 vel. ?Cochlea Hitchcock, 1858. Brembana Valley, OrobicBasin. DD - Secundumichnus sp. Brembana Valley, Orobic Basin. EE – Undetermined traces. Brembana Valley, OrobicBasin. FF – Medusina atava (Pohlig, 1982) Walcott, 1898. Inferno Valley, Orobic Basin. GG – Anthracosiidae TrompiaValley Basin. HH – Dendroidichnites elegans Demathieu, Gand & Toutin-Morin, 1992, Mincucco Mt. Orobic Basin.II,, JJ – Medusina limnica Müller, 1978. Trompia Valley Basin .
PPllaattee 22 (continued on next page)
AA- Camunipes cassinisi Ceoloni et al., 1987, reverse print left couple manus-pes. Brembana Valley, Orobic Basin.BB – Amphisauropus latus Haubold, 1971, reverse print right pes. Inferno Valley, Orobic Basin. CC - Amphisauropus latusHaubold, 1971, reverse print left manus. Inferno Valley, Orobic Basin. DD – Varanopus curvidactylus Moodie, 1929,reverse print left pes. Inferno Valley, Orobic Basin. EE - Varanopus curvidactylus Moodie, 1929, reverse print left couple
89Geo.Alp, Vol. 2, 2005
PPllaattee 11
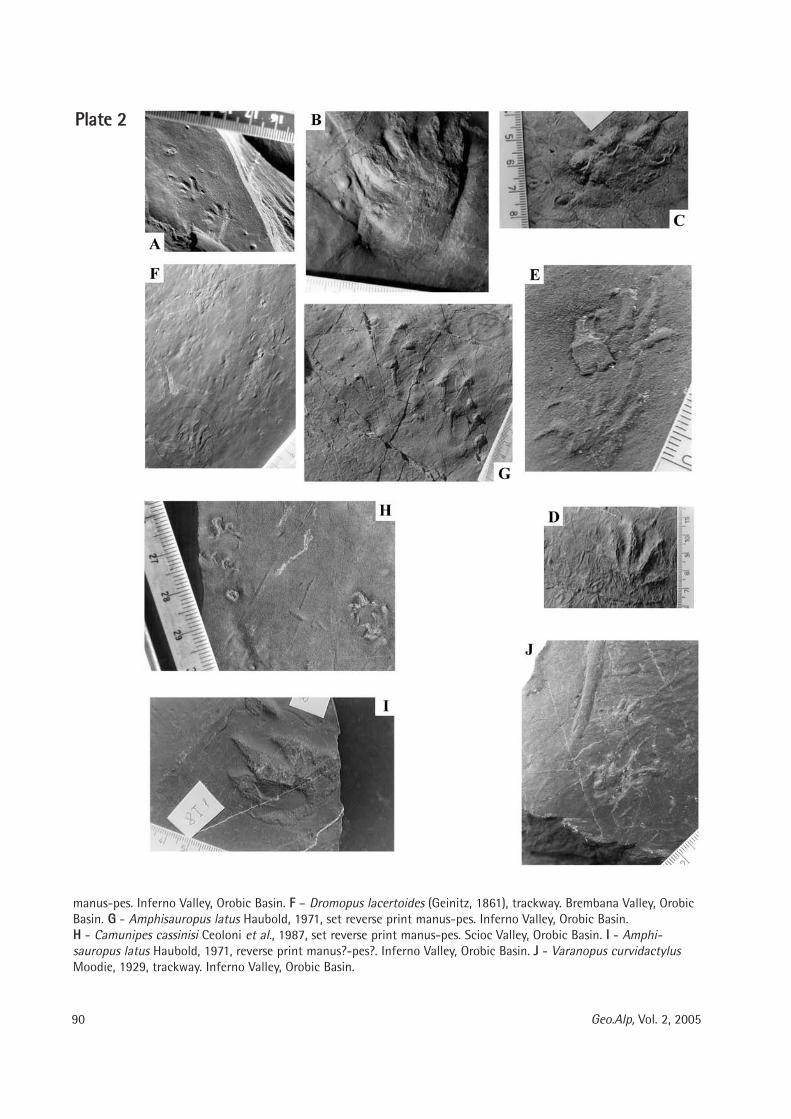
90 Geo.Alp, Vol. 2, 2005
manus-pes. Inferno Valley, Orobic Basin. FF – Dromopus lacertoides (Geinitz, 1861), trackway. Brembana Valley, OrobicBasin. GG - Amphisauropus latus Haubold, 1971, set reverse print manus-pes. Inferno Valley, Orobic Basin.HH - Camunipes cassinisi Ceoloni et al., 1987, set reverse print manus-pes. Scioc Valley, Orobic Basin. II - Amphi -sauropus latus Haubold, 1971, reverse print manus?-pes?. Inferno Valley, Orobic Basin. JJ - Varanopus curvidactylusMoodie, 1929, trackway. Inferno Valley, Orobic Basin.
PPllaattee 22

Geo.Alp, Vol. 2, S. 91–98, 2005
PPRREELLIIMMIINNAARRYY AANNAALLYYSSIISS OOFF TTHHEE FFIIRRSSTT LLOOWWEERR MMOOLLAARR VVAARRIIAABBIILLIITTYYIINN LLAATTEE PPLLEEIISSTTOOCCEENNEE AANNDD LLIIVVIINNGG PPOOPPUULLAATTIIOONNSS OOFF TTEERRRRIICCOOLLAA SSAAVVIIII ((AARRVVIICCOOLLIIDDAAEE,, RROODDEENNTTIIAA))
Maria Teresa Curcio1, Longino Contoli2, Emanuele Di Canzio3, Tassos Kotsakis1
With 7 figures and 3 tables
AAbbssttrraaccttThe main object of this research is the study of the variability degree of the first lower molar in Late
Pleistocene and living populations of Terricola savii in Italy (whose conspecificity has been proved by genet-ic analyses) and its comparison with that of fossil populations (assigned to T. savii on a morphological basis)in order to find a way to attribute isolated fossil remains to specific systematic groups. On this basis, weattempted to establish, through different analyses and direct observations on the occlusal dental surfacemorphology, the relationships that exist between fossil and living populations, and to verify the existence ofa temporal and/or geographic cline.
91
1 Dipartimento di Scienze Geologiche, Università Roma Tre, Largo San Leonardo Murialdo 1 - 00146 Roma, Italy2 Via Arno 38 - 00198 Roma, Italy3 Dipartimento di Scienze della Terra, Università di Roma “La Sapienza”, Piazzale Aldo Moro 1 - 00185 Roma, Italye-mail: [email protected]; [email protected]; [email protected]; [email protected]
IInnttrroodduuccttiioonn
Terricola savii (DE SELYS LONGCHAMPS, 1838)(Arvicolidae, Rodentia) is the most common livingvole on the Italian peninsula (south of theApennines) and it is common in northern Italy too(between the Alps and the Apennines). During theLate Pleistocene T. savii colonized Sicily (Petruso,2002). Voles classified as T. gr. T. savii are present inEarly Toringian mammal assemblages (San Giovannidi Duino, Venezia Giulia - Campani Quarry, Tuscany– Case Picconetto, Abruzzi) (Bartolomei, 1976;Marcolini, 2002; Marcolini et al., 2003) belongingto the Fontana Ranuccio Faunal Unit (Gliozzi et al.,1997). True Savi’s ground voles are reported sincethe beginning of Late Toringian (Bartolomei, 1980),corresponding to the latest phase of the MiddlePleistocene. During the Late Pleistocene the geo-graphic range of T. savi expanded or reduced due toclimatic changes. During the temperate-warmoscillations, this species reached the Alpine region,whilst during the cool or cold periods, it wasrestricted to the southern and central parts of theItalian peninsula (Kotsakis et al., 2003).
Studies on local populations of T. savii have beencarried out by several authors: De Giuli (1983),Corridi (1987), Rustioni et al. (1994), Abbassi &Brunet-Lecomte (1997), Masini & Abbazzi (1997),Ronchitelli et al. (1998). A general analysis of fossiland living populations of Italian ground voles hasbeen already published by Brunet-Lecomte et al.(1994a). The present study focuses on the compar-ison of living and fossil populations of T. savii only.Our target is to examine the relationships of recentpopulations of central and southern areas of theItalian peninsula with the fossil ones of the samearea. A similar work was performed by Brunet-Lecomte et al. (1994b) for Terricola gerbei (Gerbe,1879) (= Terricola pyrenaica (de Sélys Longchamps,1847)) of northern Spain and south-westernFrance.
MMaatteerriiaallss aanndd MMeetthhooddss
The studied samples are derived from 11 locali-ties. Five out of this number are fossil populations,while six belong to living samples. The latter are

from: Cervia (Ravenna, Emilia Romagna), Civitelladel Tronto (Teramo, Abruzzi), Torraccia di SanGennaro (Rome, Latium), Casarano (Lecce, Apulia),Sila National Park (Calabria), Noto (Syracuse, Sicily).The five fossil populations were collected inMelpignano (Lecce, Apulia), Ingarano (Foggia,Apulia), Praia a Mare (Cosenza, Calabria), Ostuni(Foggia, Apulia) and Riparo Salvini (Latina, Latium)(fig. 1).
As to the fossil localities, in the fossiliferous site ofMelpignano a fauna testyfing warm climatic condi-tions has been recognized. In particular the macro-fauna collected in sediments of karst cavities allowedits attribution to MIS 5a-5c (Bologna et al., 1994).Petronio et al. (1996) assigned the assemblage fromIngarano to the middle part of MIS 3. CapassoBarbato & Gliozzi (2001) assigned the small mammalassemblage from Praia a Mare to the final phase ofMIS 3. The Ostuni fossil assemblage is ascribed toMIS 2 by Angelone et al. (2004). The fauna from RiparoSalvini has been ascribed to the latest Pleistocene(Tardiglacial - final phase of MIS 2) (Cassoli &Guadagnoli, 1987; Alessio et al., 1993) (fig. 2).
The material studied in this research pertains topublic and private collections. In particular the fos-sil material from Melpignano and Ingarano is storedin the Dipartimento di Scienze della Terra of theUniversity of Rome „La Sapienza“, the fossils fromPraia a Mare, Ostuni and Riparo Salvini are stored inthe Laboratory of Palaeontology of the Diparti -mento di Scienze Geologiche of the UniversityRoma Tre. The recent material belongs to the„Contoli Col lection“ and it is stored in theDipartimento di Biologia Animale e dell’Uomo ofthe University of Rome „La Sapienza“.
The decision to take dental measurements, par-ticulary on the first lower molars (M1) (fig. 3b;tabs. 2, 3) is necessary because the systematics ofthe Arvicolidae is based on the morphology of thistooth, and because teeth are often the only com-mon fossil elements available. Quantitative andqualitative analyses have been carried out on thestudied material. The pictures of teeth were takenby using a digital camera Nikon Coolpix 995 con-nected to a stereoscopic microscope Nikon SMZ-U.The measurements were carried out with the graph-ic program CorelDraw 8. The statistic analyses werecarried out with the program KyPlot ver.2.0 beta 15.Some illustrations have been produced with the aidof a Leica L2 camera lucida and of a graphicaltablet.
92 Geo.Alp, Vol. 2, 2005
Fig. 1: Geographical location of the studied populations.
Fig. 2: Chronostratigraphy of the late Middle Pleistocene andHolocene.
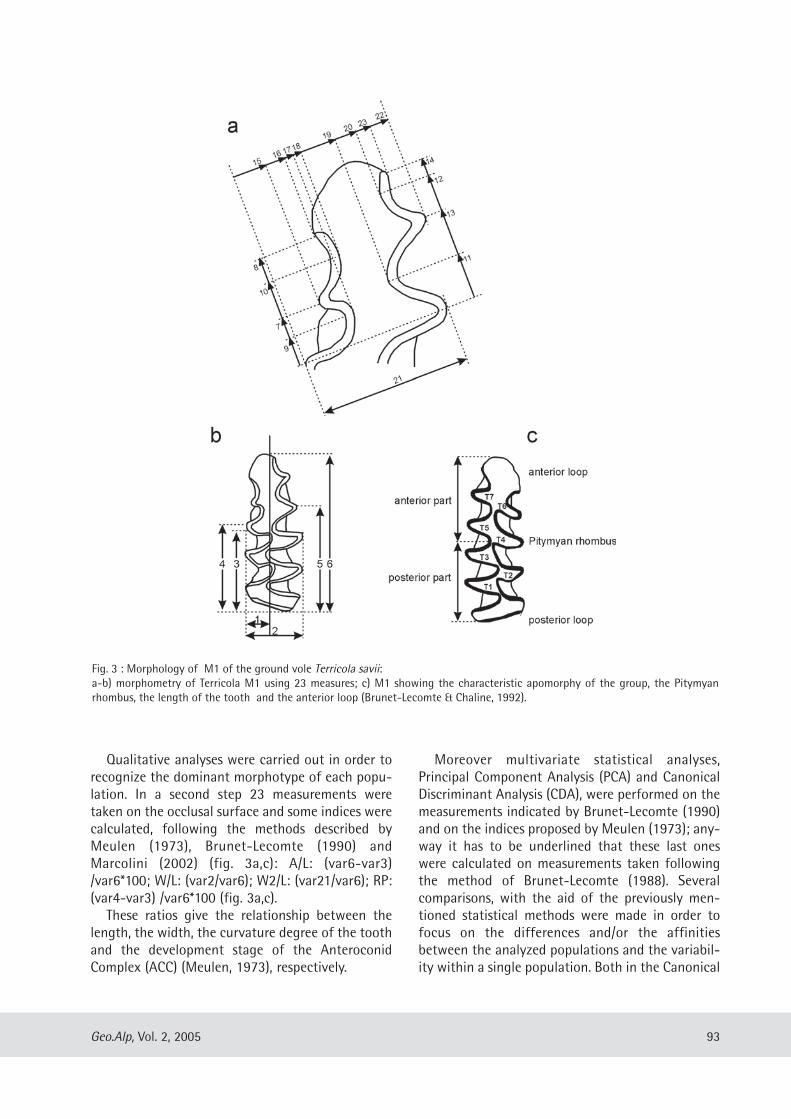
Qualitative analyses were carried out in order torecognize the dominant morphotype of each popu-lation. In a second step 23 measurements weretaken on the occlusal surface and some indices werecalculated, following the methods described byMeulen (1973), Brunet-Lecomte (1990) andMarcolini (2002) (fig. 3a,c): A/L: (var6-var3)/var6*100; W/L: (var2/var6); W2/L: (var21/var6); RP:(var4-var3) /var6*100 (fig. 3a,c).
These ratios give the relationship between thelength, the width, the curvature degree of the toothand the development stage of the AnteroconidComplex (ACC) (Meulen, 1973), respectively.
Moreover multivariate statistical analyses,Principal Component Analysis (PCA) and CanonicalDiscriminant Analysis (CDA), were performed on themeasurements indicated by Brunet-Lecomte (1990)and on the indices proposed by Meulen (1973); any-way it has to be underlined that these last oneswere calculated on measurements taken followingthe method of Brunet-Lecomte (1988). Severalcomparisons, with the aid of the previously men-tioned statistical methods were made in order tofocus on the differences and/or the affinitiesbetween the analyzed populations and the variabil-ity within a single population. Both in the Canonical
93Geo.Alp, Vol. 2, 2005
Fig. 3 : Morphology of M1 of the ground vole Terricola savii:a-b) morphometry of Terricola M1 using 23 measures; c) M1 showing the characteristic apomorphy of the group, the Pitymyanrhombus, the length of the tooth and the anterior loop (Brunet-Lecomte & Chaline, 1992).

Discriminant Analysis and in the PrincipalComponent Analysis the populations were analyzedin a first moment all together. Subsequently, thesesame analyses (PCA and CDA) have been repeateddividing the populations in fossil and recent onesand all populations have been compared pair bypair. In all tests an outgroup was present. The out-group population comes from the lower level ofGran Dolina (Atapuerca, Burgos, Spain) and it iscomposed by Terricola arvalidens (Cuenca - Bescoset al., 1995). This material has been found in a karstfilling sediment (approximately 18 meters thick)partly ascribed to Early Pleistocene and partly toMiddle Pleistocene. This population is not temporal-ly or geographically related to ours (both living andfossil), nevertheless shows similar characteristics tothose of the studied populations and for this reasonhas been included in the analyses.
The matrix used for PCA and CDA are available in thesite http://host.uniroma3.it/laboratori/paleontologia.
RReessuullttss
Three different morphotypes were identified, onthe basis of the number of salient and re-entrantangles, of the complication and development of the
Anterior Loop (AL) and of the greater or smallerconfluence of the triangles in the AnteroconidComplex.
MORPHOTYPE 1( morphotype savii s.s.) is character-ized by a simple and wide anterior loop, with a wideneck and widely confluent with the triangles T7 andT6. T5 and T4 are broadly confluent. The reentrantangles are quite marked and slightly more flattenedon the lingual side (fig. 4 a,b,c).
The anterior loop in MORPHOTYPE 2 is more com-plex than in morphotype 1. T7 and its reentrantangle are much more evident while T6 and its reen-trant angle are only outlined or even absent. T4 andT5 are not confluent and consequently thepitymyan rhombus is not clearly visible (fig. 4 d,e,f).The anterior loop of MORPHOTYPE 3 is as simple as inmorphotype 1 although the triangles are ratherirregular in shape (fig. 4 g,h).
The analysis of the morphotypes shows a cleardominance of morphotypes 1 and 2 in all the exam-ined populations, both fossil and recent, while mor-photype 3 is present only marginally in the recentpopulations (tab.1).
As to the variability of the M1’s within the ana-lyzed populations, as it is shown by the qualitativedata, it is clear that M1 follows a mosaic modelcomposed by the Anteroconid Complex (ACC),which is more variable and characterizing most ofthe morphotypes and by a more conservativeTalonid-Trigonid Complex (TTC). The observed vari-ability is both inter- and intra-populational.Moreover, it was possible to divide all the analyzedM1 into two different morphotypes of both thefossil populations and the living ones (the thirdmorphotype is present as we have seen only in theliving populations with low percentages) and, inboth cases, the percentages of the morphotypesare similar. Nearly none of the performed PCAhave brought statistically significant results. In theplots obtained by statistically significant analysesthere seem to be no differences within the fossilpopulations or the recent ones. And there seem tobe no differences between fossil and living popu-lations.
As to the living populations, differences havebeen recognized between the populations of Noto(Siracusa, Sicily) and Cervia (Ravenna, EmiliaRomagna), but this is a rather obvious result, beinggeographically the two farthest populations with-in those considered. Moreover, the population ofNoto, coming from the island of Sicily, introduces
94 Geo.Alp, Vol. 2, 2005
Fig. 4: Morphotypes of Terricola savii: a,b,c) morphotype 1(morphotype savii s.s.); d,e,f) morphotype 2; g,h) morpho -type 3.
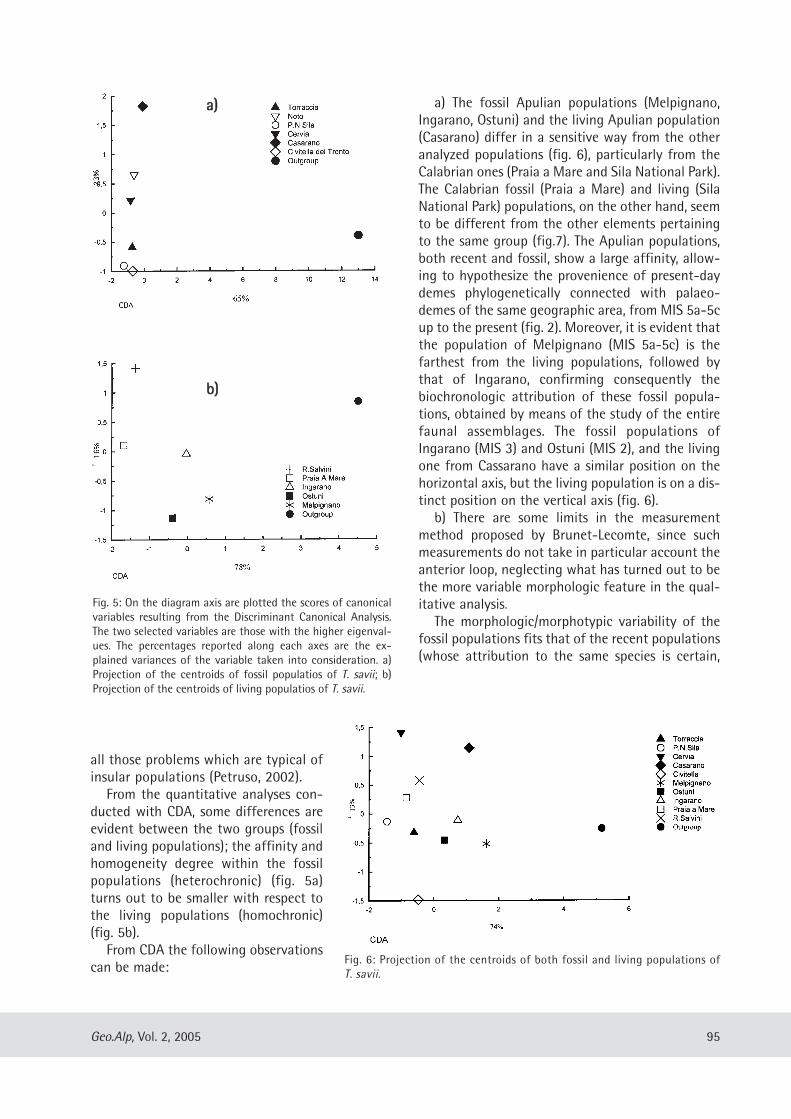
all those problems which are typical ofinsular populations (Petruso, 2002).
From the quantitative analyses con-ducted with CDA, some differences areevident between the two groups (fossiland living populations); the affinity andhomogeneity degree within the fossilpopulations (heterochronic) (fig. 5a)turns out to be smaller with respect tothe living populations (homochronic)(fig. 5b).
From CDA the following observationscan be made:
a) The fossil Apulian populations (Melpignano,Ingarano, Ostuni) and the living Apulian population(Casarano) differ in a sensitive way from the otheranalyzed populations (fig. 6), particularly from theCalabrian ones (Praia a Mare and Sila National Park).The Calabrian fossil (Praia a Mare) and living (SilaNational Park) populations, on the other hand, seemto be different from the other elements pertainingto the same group (fig.7). The Apulian populations,both recent and fossil, show a large affinity, allow-ing to hypothesize the provenience of present-daydemes phylogenetically connected with palaeo -demes of the same geographic area, from MIS 5a-5cup to the present (fig. 2). Moreover, it is evident thatthe population of Melpignano (MIS 5a-5c) is thefarthest from the living populations, followed bythat of Ingarano, confirming consequently thebiochronologic attribution of these fossil popula-tions, obtained by means of the study of the entirefaunal assemblages. The fossil populations ofIngarano (MIS 3) and Ostuni (MIS 2), and the livingone from Cassarano have a similar position on thehorizontal axis, but the living population is on a dis-tinct position on the vertical axis (fig. 6).
b) There are some limits in the measurementmethod proposed by Brunet-Lecomte, since suchmeasurements do not take in particular account theanterior loop, neglecting what has turned out to bethe more variable morphologic feature in the qual-itative analysis.
The morphologic/morphotypic variability of thefossil populations fits that of the recent populations(whose attribution to the same species is certain,
95Geo.Alp, Vol. 2, 2005
Fig. 5: On the diagram axis are plotted the scores of canonicalvariables resulting from the Discriminant Canonical Analysis.The two selected variables are those with the higher eigenval-ues. The percentages reported along each axes are the ex-plained variances of the variable taken into consideration. a)Projection of the centroids of fossil populatios of T. savii; b)Projection of the centroids of living populatios of T. savii.
Fig. 6: Projection of the centroids of both fossil and living populations ofT. savii.
aa))
bb))
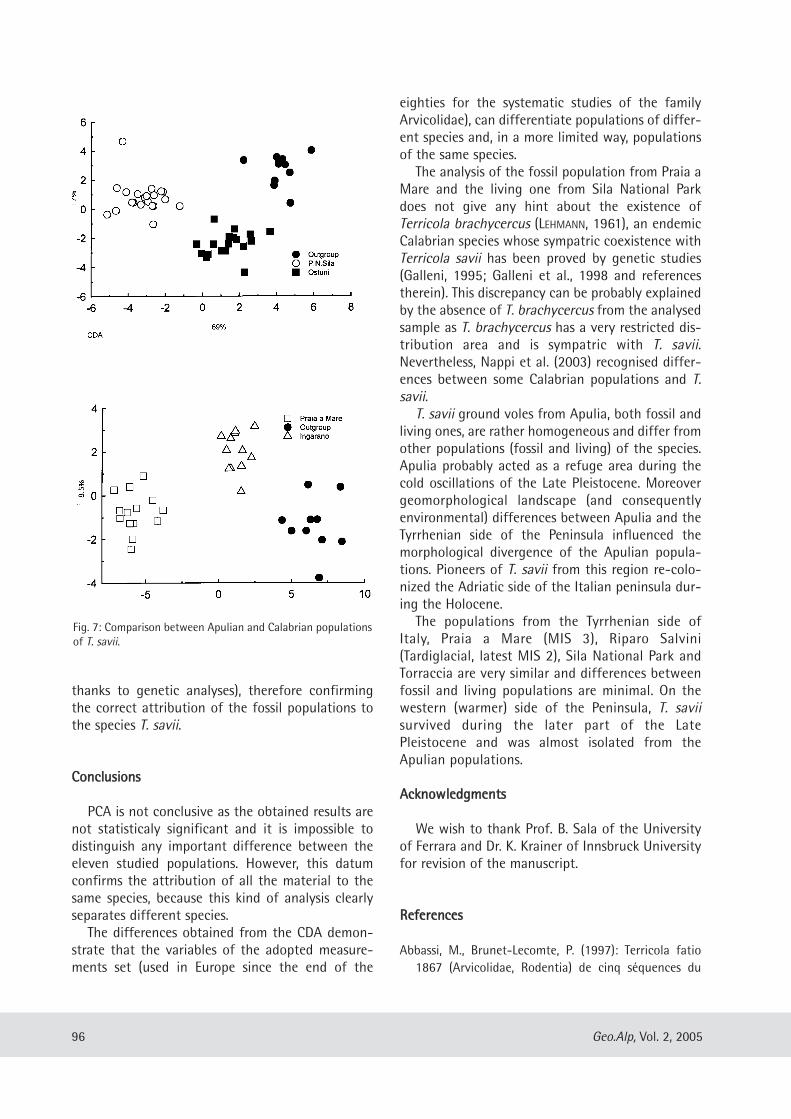
thanks to genetic analyses), therefore confirmingthe correct attribution of the fossil populations tothe species T. savii.
CCoonncclluussiioonnss
PCA is not conclusive as the obtained results arenot statisticaly significant and it is impossible todistinguish any important difference between theeleven studied populations. However, this datumconfirms the attribution of all the material to thesame species, because this kind of analysis clearlyseparates different species.
The differences obtained from the CDA demon-strate that the variables of the adopted measure-ments set (used in Europe since the end of the
eighties for the systematic studies of the familyArvicolidae), can differentiate populations of differ-ent species and, in a more limited way, populationsof the same species.
The analysis of the fossil population from Praia aMare and the living one from Sila National Parkdoes not give any hint about the existence ofTerricola brachycercus (LEHMANN, 1961), an endemicCalabrian species whose sympatric coexistence withTerricola savii has been proved by genetic studies(Galleni, 1995; Galleni et al., 1998 and referencestherein). This discrepancy can be probably explainedby the absence of T. brachycercus from the analysedsample as T. brachycercus has a very restricted dis-tribution area and is sympatric with T. savii.Nevertheless, Nappi et al. (2003) recognised differ-ences between some Calabrian populations and T.savii.
T. savii ground voles from Apulia, both fossil andliving ones, are rather homogeneous and differ fromother populations (fossil and living) of the species.Apulia probably acted as a refuge area during thecold oscillations of the Late Pleistocene. Moreovergeomorphological landscape (and consequentlyenvironmental) differences between Apulia and theTyrrhenian side of the Peninsula influenced themorphological divergence of the Apulian popula-tions. Pioneers of T. savii from this region re-colo-nized the Adriatic side of the Italian peninsula dur-ing the Holocene.
The populations from the Tyrrhenian side ofItaly, Praia a Mare (MIS 3), Riparo Salvini(Tardiglacial, latest MIS 2), Sila National Park andTorraccia are very similar and differences betweenfossil and living populations are minimal. On thewestern (warmer) side of the Peninsula, T. saviisurvived during the later part of the LatePleistocene and was almost isolated from theApulian populations.
AAcckknnoowwlleeddggmmeennttss
We wish to thank Prof. B. Sala of the Universityof Ferrara and Dr. K. Krainer of Innsbruck Universityfor revision of the manuscript.
RReeffeerreenncceess
Abbassi, M., Brunet-Lecomte, P. (1997): Terricola fatio1867 (Arvicolidae, Rodentia) de cinq séquences du
96 Geo.Alp, Vol. 2, 2005
Fig. 7: Comparison between Apulian and Calabrian populationsof T. savii.
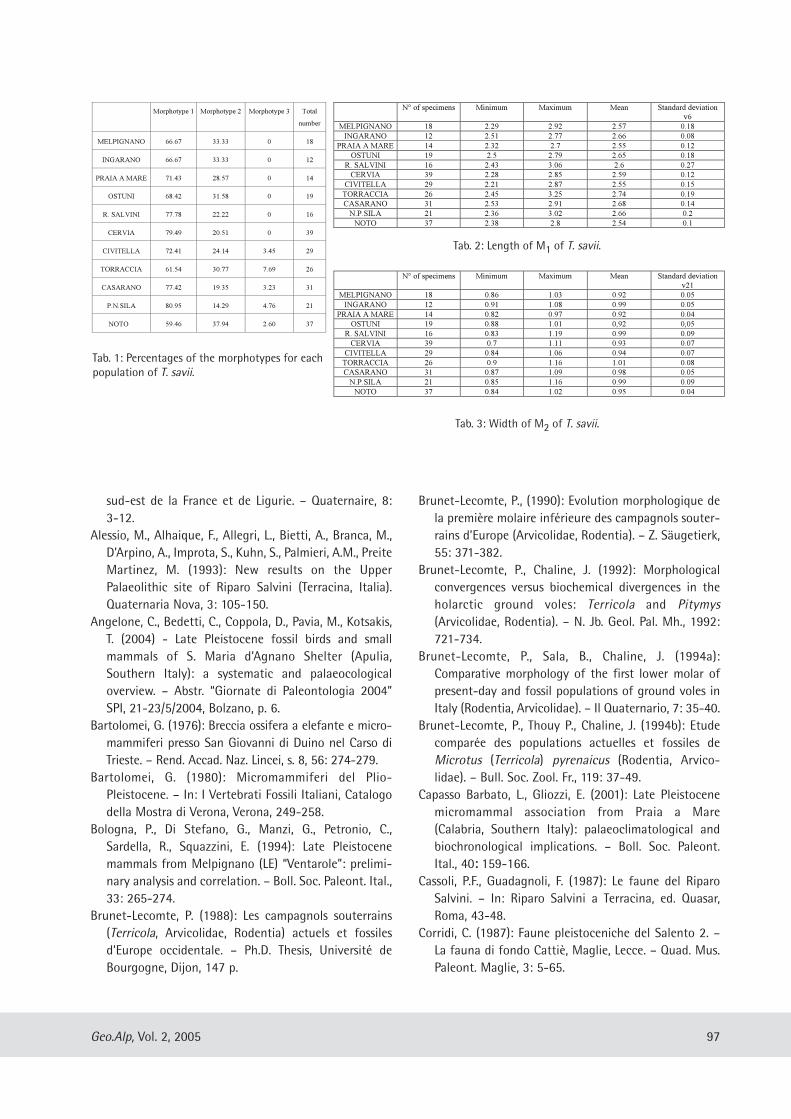
sud-est de la France et de Ligurie. – Quaternaire, 8:3-12.
Alessio, M., Alhaique, F., Allegri, L., Bietti, A., Branca, M.,D’Arpino, A., Improta, S., Kuhn, S., Palmieri, A.M., PreiteMartinez, M. (1993): New results on the UpperPalaeolithic site of Riparo Salvini (Terracina, Italia).Quaternaria Nova, 3: 105-150.
Angelone, C., Bedetti, C., Coppola, D., Pavia, M., Kotsakis,T. (2004) - Late Pleistocene fossil birds and smallmammals of S. Maria d’Agnano Shelter (Apulia,Southern Italy): a systematic and palaeocologicaloverview. – Abstr. “Giornate di Paleontologia 2004”SPI, 21-23/5/2004, Bolzano, p. 6.
Bartolomei, G. (1976): Breccia ossifera a elefante e micro-mammiferi presso San Giovanni di Duino nel Carso diTrieste. – Rend. Accad. Naz. Lincei, s. 8, 56: 274-279.
Bartolomei, G. (1980): Micromammiferi del Plio-Pleistocene. – In: I Vertebrati Fossili Italiani, Catalogodella Mostra di Verona, Verona, 249-258.
Bologna, P., Di Stefano, G., Manzi, G., Petronio, C.,Sardella, R., Squazzini, E. (1994): Late Pleistocenemammals from Melpignano (LE) “Ventarole”: prelimi-nary analysis and correlation. – Boll. Soc. Paleont. Ital.,33: 265-274.
Brunet-Lecomte, P. (1988): Les campagnols souterrains(Terricola, Arvicolidae, Rodentia) actuels et fossilesd’Europe occidentale. – Ph.D. Thesis, Université deBourgogne, Dijon, 147 p.
Brunet-Lecomte, P., (1990): Evolution morphologique dela première molaire inférieure des campagnols souter-rains d’Europe (Arvicolidae, Rodentia). – Z. Säugetierk,55: 371-382.
Brunet-Lecomte, P., Chaline, J. (1992): Morphologicalconvergences versus biochemical divergences in theholarctic ground voles: Terricola and Pitymys(Arvicolidae, Rodentia). – N. Jb. Geol. Pal. Mh., 1992:721-734.
Brunet-Lecomte, P., Sala, B., Chaline, J. (1994a):Comparative morphology of the first lower molar ofpresent-day and fossil populations of ground voles inItaly (Rodentia, Arvicolidae). – Il Quaternario, 7: 35-40.
Brunet-Lecomte, P., Thouy P., Chaline, J. (1994b): Etudecomparée des populations actuelles et fossiles deMicrotus (Terricola) pyrenaicus (Rodentia, Arvi co -lidae). – Bull. Soc. Zool. Fr., 119: 37-49.
Capasso Barbato, L., Gliozzi, E. (2001): Late Pleistocenemicromammal association from Praia a Mare(Calabria, Southern Italy): palaeoclimatological andbiochronological implications. – Boll. Soc. Paleont.Ital., 40:: 159-166.
Cassoli, P.F., Guadagnoli, F. (1987): Le faune del RiparoSalvini. – In: Riparo Salvini a Terracina, ed. Quasar,Roma, 43-48.
Corridi, C. (1987): Faune pleistoceniche del Salento 2. –La fauna di fondo Cattiè, Maglie, Lecce. – Quad. Mus.Paleont. Maglie, 3: 5-65.
97Geo.Alp, Vol. 2, 2005
N° of specimens Minimum Maximum Mean Standard deviation
v6
MELPIGNANO 18 2.29 2.92 2.57 0.18
INGARANO 12 2.51 2.77 2.66 0.08
PRAIA A MARE 14 2.32 2.7 2.55 0.12
OSTUNI 19 2.5 2.79 2.65 0.18
R. SALVINI 16 2.43 3.06 2.6 0.27
CERVIA 39 2.28 2.85 2.59 0.12
CIVITELLA 29 2.21 2.87 2.55 0.15
TORRACCIA 26 2.45 3.25 2.74 0.19
CASARANO 31 2.53 2.91 2.68 0.14
N.P.SILA 21 2.36 3.02 2.66 0.2
NOTO 37 2.38 2.8 2.54 0.1
N° of specimens Minimum Maximum Mean Standard deviation
v21
MELPIGNANO 18 0.86 1.03 0.92 0.05
INGARANO 12 0.91 1.08 0.99 0.05
PRAIA A MARE 14 0.82 0.97 0.92 0.04
OSTUNI 19 0.88 1.01 0,92 0,05
R. SALVINI 16 0.83 1.19 0.99 0.09
CERVIA 39 0.7 1.11 0.93 0.07
CIVITELLA 29 0.84 1.06 0.94 0.07
TORRACCIA 26 0.9 1.16 1.01 0.08
CASARANO 31 0.87 1.09 0.98 0.05
N.P.SILA 21 0.85 1.16 0.99 0.09
NOTO 37 0.84 1.02 0.95 0.04
Morphotype 1 Morphotype 2 Morphotype 3 Total
number
MELPIGNANO 66.67 33.33 0 18
INGARANO 66.67 33.33 0 12
PRAIA A MARE 71.43 28.57 0 14
OSTUNI 68.42 31.58 0 19
R. SALVINI 77.78 22.22 0 16
CERVIA 79.49 20.51 0 39
CIVITELLA 72.41 24.14 3.45 29
TORRACCIA 61.54 30.77 7.69 26
CASARANO 77.42 19.35 3.23 31
P.N.SILA 80.95 14.29 4.76 21
NOTO 59.46 37.94 2.60 37
Tab. 2: Length of M1 of T. savii.
Tab. 3: Width of M2 of T. savii.
Tab. 1: Percentages of the morphotypes for eachpopulation of T. savii.

Cuenca-Bescós, G., Canudo, J.I., Laplana, C. (1995): Losarvicólidos (Rodentia, Mammalia) de los niveles inferi-ores de Gran Dolina (Pleistoceno Inferior, Atapuerca,Burgos, España). – Rev. Esp. Paleont. 10:: 202-218.
De Giuli, C. (1983) – Faune pleistoceniche del Salento 1.La fauna di San Sidero 3. Quad. Mus. Paleont. Maglie,1: 45-84.
Galleni, L. (1995): Speciation in the Savi pine vole,Microtus savii (de Sélys-Longchamps) (Rodentia,Arvicolidae): a theoretical biology approach. – Boll.Zool., 62: 45-51.
Galleni, L., Stanyon, R., Contadini, L., Tellini, A. (1998):Biogeographical and karyological data of the Microtussavii group (Rodentia, Arvicolidae) in Italy. – Bonn.Zool. Beitr., 47: 277-282.
Gliozzi, E., Abbazzi, L., Argenti, P., Azzaroli, A., Caloi, L.,Capasso Barbato, L., Di Stefano, G., Esu, D., Ficcarelli,G., Girotti, O., Kotsakis, T., Masini, F., Mazza, P.,Mezzabotta, C., Palombo, M. R., Petronio, C., Rook, L.,Sala, B., Sardella, R., Zanalda, E., Torre, D. (1997):Biochronology of selected mammals, molluscs andostracods from the middle Pliocene to the latePleistocene in Italy. The state of the art. – Riv. Ital.Paleont. Strat., 103: 369-388.
Kotsakis, T., Abbazzi, L., Angelone, C., Argenti, P., Barisone,G., Fanfani, F., Marcolini, F., Masini, F. (2003): Plio-Pleistocene biogeography of Italian mainland micro-mammals. – DeinseA, 10: 313-342.
Marcolini, F. (2002) – Continental Lower Valdarno rodentbiochronology and two new methods for the system-atics of Mimomys (Arvicolidae, Rodentia). – Ph.D.Thesis, Università di Pisa, 165 p.
Marcolini, F., Bigazzi, G., Bonadonna, F.P., Cioni, R.,Zanchetta, G. (2003): Tephrochronolgy and tephros-tratigraphy of two Pleistocene continental fossilifer-ous successions of Central Italy. – J. Quat. Sci., 18:545-556.
Masini, F., Abbazzi, L. (1997) – L’associazione di mam-miferi della grotta di Castelcivita. – In Gambassini, P.(Ed.) – Il Paleolitico di Castelcivita, 33-59.
Meulen, A. van der (1973): Middle Pleistocene smallmammals from the Monte Peglia (Orvieto, Italy) withspecial reference to the phylogeny of Microtus(Arvocolidae, Rodentia). – Quaternaria, 17: 1-144.
Nappi, A., Montuire, S., Brunet-Lecomte, P. (2003) –Sintesi sulla morfometria del primo molare inferiorenel gruppo Microtus (Terricola) savii. – Hystrix, n.s.,suppl., Abstr. IV Congr. Ital. Teriologia, p. 125.
Petronio, C., Billia, E., Capasso Barbato, L., Di Stefano, G.,Mussi, M., Parry, S., Sardella, R., Voltaggio, M. (1996):The late Pleistocene fauna of Ingarano (Gargano,Italy): biocronological, palaeoecological, palaeoetho-logical and geocronological implications. – Boll. Soc.Paleont. Ital., 34(1995): 333-339.
Petruso, D. (2002): Il contributo dei micromammiferi allastratigrafia e paleogeografia del Quaternario conti-nentale siciliano. – Ph.D. Thesis, Università di Napoli,315 p.
Ronchitelli, A., Abbazzi, L., Accorsi, C.A., BandiniMazzanti, M., Bernardi, M., Masini, F., Mercuri, A.,Mezzabotta, C., Rook, L. (1998) – The Grotta Grande diScario (Salerno – southern Italy): stratigraphy, archae-ological finds, pollen and mammals. – Proc. 1st Intern.Congr. “Science and Technology for the Safeguard ofCultural Heritage in the Mediterranean Basin”, 1529-1535, Tip. Rustioni, M., Mazza, P., Abbazzi, L., Delfino,
M., Rook, L., Petrucci, S., Vinello, F. (1994): The würmianfauna from Sternatia (Lecce, Apulia, Italy).
98 Geo.Alp, Vol. 2, 2005
Manuscript submitted: December 14, 2004Manuscript accepted: May 25, 2005

Geo.Alp, Vol. 2, S. 99–106, 2005
AA TTEESSTT AAPPPPLLIICCAATTIIOONN OOFF TTHHEE SSHHEE MMEETTHHOODD AASS AA BBIIOOSSTTRRAATTIIGGRRAAPPHHIICCAALL PPAARRAAMMEETTEERR
Davide Mana
With 6 figures and 1 table
AAbbssttrraacctt::Biodiversity – the index “expressing the variety and variability of living organisms and of the ecological
systems comprising them” (Ferrari, 2001) – is essential in the characterization and study of past and presentbiological systems, and is generally expressed by a number (the “biodiversity index”), statistically derivedfrom empirical observations. The SHE indexing method (Buzas & Hayek, 1996, Hayek & Buzas 1997), is expressed by the Shannon Index,
H (a measure of the system’s entropy) as the composition of two factors representing respectively the num-ber of species in the sample (S) and the distribution uniformity (E).The SHE index does not only describe in a thorough way the system’s biodiversity, but, as a function of
abundance and evenness, can be used to identify biofacies (SHEBI – SHE for Biofaces Identification) or tocharacterize the whole structure of the analysed community (SHECSI – SHE for Community StructureIdentification). SHE analysis, independently of its application purposes, appears to be highly flexible, does not require the
adoption of specific computer packages beyond a common spreadsheet, and is based on a simple graphicalanalysis; widely adopted in botanics, SHEBI analysis in particular has been applied with satisfactory resultsto the study of benthic foraminiferal faunas from the Atlantic ocean (Buzas & Hayek,1998]. In this work, the SHEBI method has been applied to 87 samples from the Falconara section (Southern
Sicily) – the purpose of the study is to verify the possibility of applying SHE/SHEBI to Messinian planktonicforaminiferal assemblages. Our study has to face issues that are typical of planctonic faunas – such as the lower number of species
and the ample variability in single taxa abundances; a further factor to be taken into account in setting upand executing the analysis is the progressive deterioration of the ecosystem as the peak of the Messinian cri-sis approaches. Biofacies identification through SHEBI in less than ideal conditions, but on such a widelystudied and described section, offers an excellent opportunity to test the method and its limits, its applica-tion range and the reliability of its results.
99
C.so Traiano 24/8, 10135 – Torino, Italia
11.. IInnttrroodduuccttiioonn –– SSHHEE aanndd tthhee mmeeaassuurree ooff ddiivveerrssiittyy
This work aims at identifying and evaluating thelimits (if any) of the application of the SHE analysisto planktonic foraminiferal faunas, in order to sim-plify the application of this powerful diversity-based technique to the field of planktonicforaminiferal biostratigraphy.
Diversity is one of the defining factors in anystudy of an ecological system. A number of indiceswas developed through the years by differentresearchers, to quantitatively express diversity asobserved in the field or in laboratory; among themore widely used indices are Fisher’s α index (ameasure of species richness), Simpson’s λ index,Equitability (E, a measure of evenness) and

Shannon’s H index (a derivation of the informationfunction) (Smart, 2002).Species richness itself (expressed as S, total num-
ber of species) has been used in the past as a roughmeasure of diversity.The recognition of the mathematical relation
between Species Richness and Taxa Abundance, andits meaning in terms of Diversity and Dominance isthe basis of the recent SHE approach to the study ofbiodiversity (Hayek & Buzas, 1997).The mathematical expression summing up this
relationship is (1) HH == llnn((SS)) ++ llnn((EE))in whichHH is the Shannon Diversity IndexSS is the Species RichnessEE is the Dominance or Evenness of the distribution
Relation (1) is constant as long as species propor-tions are constant.As a change of the proportions of species to each
other is clearly a sign of change in diversity, the SHErelationship has to be interpreted as an expressionof diversity. This allows a simple graphical analysis of the
variations of biodiversity: each of the three vari-ables can in fact be plotted against the Abundance(N) of the sample; changes in proportions (andtherefore in diversity) will be signalled by a changein the graphic line slope (a “slope break” in the “hol-low curve” following Hayek & Buzas’ terminology). Operationally, the method is not as mathemati-
cally intensive as other well-established analysisprocedures (i.e., Cluster Analysis), requiring simply a
logarithmic transformation of the indices, and whilespecific software is easily available to calculate thevalues of H and E, given standard sample counts, thewhole analysis can be carried out on a simplespreadsheet software (i.e., Microsoft Excell orOpenCalc) with a minimum of fuss.Conceptually, the analysis can be carried out
through time (i.e., vertically, comparing levels alonga geological section) or through space (i.e. laterally,comparing sectors in a landscape).
Introduced in the late 1990s as a way to sidestepsome perceived limits in more popular diversityindices (Shannon-Weiner in particular), SHE’s fieldof application was later extended and redefined,with the introduction of SHEBI (SHE Analysis forBiofacies Identification) and SHECSI (SHE Analysisfor Community Structure Identification) (Buzas andHayek, 1996, 1998 ; Hayek and Buzas, 1997, 1998).Examples of applications of the SHE approach to
biodiversity have been published as part of botani-cal (Hayek and Buzas, 1996, 1997, Small andMcCarthy 2002) and zoological studies (Leponce etal. 2004); closer to the concerns of this paper, SHEhas been applied to the study of quaternary benth-ic foraminiferal faunas in what can be defined as anon-perturbed environmental setting (Buzas andHayek, 1998, Osterman et al, 2002).By all accounts, when applied to current or
recent environments and populations, SHE appearsto be a solid, easily applied method for describingdiversity; in particular, it allows a high-resolutionvisualization of changes in diversity through time orspace; the method allows researchers “to examineevenness separately from richness within a singlemultispecies system” (Buzas and Hayek, 1998) andit does not suffer from some of the limits signalledfor other diversity-based indices (Hayek and Buzas,1997).Some doubts might still remain when SHE is to
be applied to situations in which those factors themethod takes into account (population density, spe-cific richness, etc.) are subject to extreme or unpre-dictable variations – i.e. due to drastic changes inenvironmental conditions, or to other externalcauses.To verify the viability of SHE analysis in such crit-
ical conditions, this study has been carried out onplanktonic foraminiferal faunas from Messinianstrata of the Mediterranean, which are normallycharacterized by lower species richness (S) than ben-
100 Geo.Alp, Vol. 2, 2005
Fig. 1: Location map of the Falconara outcrop in Sicily.

thic faunas. Proximity to the peak of the MessinianSalinity Crisis further weakens the species richnesssignal, due to increased environmental stress.This paper briefly summarizes the study and its
results.
22.. TThhee FFaallccoonnaarraa SSeeccttiioonnaanndd tthhee ppllaannkkttoonniicc ssaammpplleess
The samples used for this study were collected inthe alternating clay/diatomite cycles of the Tripoliformation (Upper Tortonian-Messinian) with anexposed thickness of one hundred meters in theFalconara Section.Located on the southern face of Monte
Caltagirone, on the southern coast of Sicily betweenGela and Licata (see fig. 1), the Falconara Section(fig.2) was originally proposed as the type-sectionfor the Messinian (Colalongo et al., 1979), and hasbeen the object of continuing studies, criticism andrevisions, due to its paramount importance for thecomprehension of Mediterranean events; in morethan thirty years, studies have shifted from bio -stratigraphical and chronostratigraphical concernsand techniques to cyclostratigraphical and astro-chronological methods. (summarized in Hilgen et al.,2000).The abundance of previous studies and the
detailed description of the Falconara faunas(Colalongo et al., 1979, Hilgen & Krijgsman, 1999,Hilgen et al., 2000) by previous authors provides anexcellent background for our test-run of the SHEapproach to planktonic foraminifera biostratigra-phy. Our study does not mean to redefine in anyway the stratigraphy of the Falconara section, butto use a well-studied section and its wealth ofaccumulated paleontological and stratigraphicalknowledge as the consensus against which theresults of the SHE test will be compared for vali-dation.
The samples used in this study were collectedfrom the Falconara Section in 1994 (fig. 3), as partof a wide-ranging campaign of studies on theMessinian Salinity Crisis in the Mediterranean; inthe field, both clay and diatomite layers were sam-pled separately, and were later subjected to stan-dard micropaleontological analysis and quantitativestudies in the laboratories of the Università degliStudi, Torino.
The environmental information provided by thefaunas contained in the sediments was presentedand discussed in the author’s graduation paper(Mana, 2001) concerning the same samples used inthis study; in that work, a general biozonationbased on a traditional method (Cluster Analysis),was proposed, identifying seven distinct biofacies,each connected with the progressive environmentalcrisis of the Messinian sea. That work, and the excellent synthesis by Hilgen
and Krijgsman (1999) will be our two chief refer-ences for comparison.
33.. SSHHEE AAnnaallyyssiiss
For the purposes of this study, 87 samples wereobserved, and 300 individuals counted according tostandard statistical data-gathering practices; seven-teen planktonic taxa were recognized (see below)and counted; to these, a class labelled “others” wasadded to include the few non-planktonic individu-als (mostly Bulimina echinata). For the speciesNeogloboquadrina acostaensis, sinistral coilingindividuals were counted separately from dextralcoiling individuals.
101Geo.Alp, Vol. 2, 2005
Fig. 2: View of the Falconara outcrop.

The taxa used in this study are:
Globigerina angustiumbilicataGlobigerina spGlobigerinoides ruberGlobigerinoides spTurborotalita multilobaTurborotalita spGloborotalia conoideaGloborotalia praemenardiGloborotalia spNeogloboquadrina continuosaNeogloboquadrina acostaensis sin.Neogloboquadrina acostaensis dex.Orbulina universaGlobigerinella obesaGlobigerinella praesiphonipheraGlobigerinella spSphaeroidinellopsisOther
As described in Buzas and Hayek (1998), from thespecies counts, the cumulative values of N (numberof individuals in sample), S (number of species insample or Specific Richness), H (Shannon’s Index)and E (Evenness) were calculated using an Excellspreadsheet, and the natural logarithms extractedfor each value (Table 1).Cumulative values (a stepwise addition of values)
were used so that S will be steadily increasingthrough the sequence. Considering now equation
(1) HH == llnn((SS)) ++ llnn((EE))as we have already stated, this relation remains con-stant as long as species proportions remain con-stant. More to the point, if – as in the case of ouranalysis – the value of S increases steadily due tothe cumulative process, two possibilities canbecome apparent: if, as S increases, H remains con-stant, this will mean a progressive decrease in thevalue of the samples’ cumulative Evenness; shouldinstead the value of ln(E) remain constant, thiswould mean a progressive variation in the value ofH. Plotting linear graphics for
llnn((SS)) vvss llnn((NN))HH vvss llnn((NN))llnn((EE)) vvss llnn((NN))allows us to pinpoint biofacies changes, representedby slope breaks on the graphs (fig. 4).
102 Geo.Alp, Vol. 2, 2005
Fig. 3: Summary sketch of the Falconara Section, with samplenumbers.

In fig. 4 the three “hollow curves” (Hayek &Buzas, 1997, 1998) are plotted in a single graph inthe same order in which we introduced them above;as our fig. 4 shows, a number of breaks are evident,each of them potentially marking a change in asso-ciation, and therefore in biofacies.It is important at this point to notice that mat-
ters of scale, and the high number of individualsprojected, might distort the curve plot, causing aloss of definition and actually masking some signif-icant slope breaks. To avoid this distortion effect,the suggested practice consists in breaking thesequence into smaller intervals – which is achievedin practice by stepwise deleting the samples whosetrend has already been analysed, recalculating allthe values in the system. The stepwise deletion procedure also corrects
another important distortion which may presentthe single-plot SHE model in fig. 4 – the one causedby the disappearance of certain taxa as thesequence develops. Cumulative addition of SpecificRichness alone, does account for the appearance ofnew species, but not for the loss of those speciesthat, while present in the earlier levels of thesequence, disappear later. By stepwise deleting ear-lier data-points from the plot as the analysis pro-
gresses, and recalculating the values of S, N, E andH, disappearances are now computed into themodel.For the purposes of this work, the SHE analysis
procedure was applied six times (fig. 5) in order toheighten the definition of the hollow curve. The resulting graphs appear choppy and uneven,
especially when compared to similar plots for ben-thic faunas (Buzas and Hayek, 1998]; this is aneffect most likely caused by the characters of theplanktonic assemblage (low Specific Richness, sud-den disappearances) and the time interval consid-ered (wide and sudden variations in environmentalconditions as the situation evolves towards the cri-sis). The operator has also to take into account thevery low values of the indices, a product of the gen-erally low Species Richness and of the scarcity ofbiological remains in some samples (fifteen ofwhich lack fossils). Our biofacies analysis is based chiefly on the joint
observation of both ln(S) and ln(E) plots; the latteris considered to be most sensitive to specific assem-blage changes by most authors, but considering thescarcity of species represented in the samples, andthe low abundances, using the former as a controland as a support in the definition of biofacies
103Geo.Alp, Vol. 2, 2005
Table 1: List of Falconara samples showing calculated values of indexes for SHE analysis. Missing samples were found to be sterile.Abbreviations: N, counted individuals; (N), cumulated N; (S), cumulated species richness; H, Shannon’s Index; (E), Evenness.
SAMPLE N (N) ln(N) (S) ln(S) H (E) LN(E) SAMPLE N (N) ln(N) (S) ln(S) H (E) LN(E)
1 307 307 5,73 7 1,95 0,97 0,38 -0,97 45 316 10913 9,30 54 3,99 0,14 0,02 -3,85
2 288 595 6,39 11 2,40 1,00 0,25 -1,40 47 304 11217 9,33 54 3,99 0,11 0,02 -3,88
3 329 924 6,83 20 3,00 0,86 0,12 -2,14 48 341 11558 9,36 54 3,99 0,11 0,02 -3,87
4 302 1226 7,11 22 3,09 0,62 0,08 -2,47 49 343 11901 9,38 54 3,99 0,15 0,02 -3,84
5 315 1541 7,34 26 3,26 0,62 0,07 -2,64 50 337 12238 9,41 54 3,99 0,10 0,02 -3,89
6 303 1844 7,52 31 3,43 0,53 0,05 -2,91 51 396 12634 9,44 54 3,99 0,14 0,02 -3,85
7 300 2144 7,67 31 3,43 0,43 0,05 -3,01 52 347 12981 9,47 56 4,03 0,13 0,02 -3,90
8 327 2471 7,81 32 3,47 0,38 0,05 -3,09 53 317 13298 9,50 56 4,03 0,11 0,02 -3,91
9 329 2800 7,94 34 3,53 0,34 0,04 -3,18 54 354 13652 9,52 57 4,04 0,12 0,02 -3,93
10 396 3196 8,07 35 3,56 0,40 0,04 -3,16 55 326 13978 9,55 57 4,04 0,11 0,02 -3,93
11 285 3481 8,16 37 3,61 0,31 0,04 -3,30 56 300 14278 9,57 57 4,04 0,11 0,02 -3,93
12 319 3800 8,24 37 3,61 0,27 0,04 -3,34 57 307 14585 9,59 57 4,04 0,11 0,02 -3,93
13 286 4086 8,32 38 3,64 0,29 0,04 -3,34 58 198 14783 9,60 57 4,04 0,07 0,02 -3,98
14 296 4382 8,39 39 3,66 0,28 0,03 -3,39 59 327 15110 9,62 57 4,04 0,12 0,02 -3,93
15 323 4705 8,46 45 3,81 0,30 0,03 -3,51 61 331 15441 9,64 58 4,06 0,12 0,02 -3,94
16 312 5017 8,52 46 3,83 0,26 0,03 -3,57 62 350 15791 9,67 58 4,06 0,10 0,02 -3,97
17 328 5345 8,58 48 3,87 0,21 0,03 -3,66 64 349 16140 9,69 60 4,09 0,11 0,02 -3,98
20 303 5648 8,64 48 3,87 0,23 0,03 -3,64 65 287 16427 9,71 60 4,09 0,09 0,02 -4,00
22 305 5953 8,69 49 3,89 0,22 0,03 -3,67 67 344 16771 9,73 60 4,09 0,11 0,02 -3,99
24 172 6125 8,72 50 3,91 0,13 0,02 -3,78 68 311 17082 9,75 60 4,09 0,10 0,02 -3,99
25 313 6438 8,77 50 3,91 0,20 0,02 -3,72 70 314 17396 9,76 60 4,09 0,10 0,02 -4,00
26 293 6731 8,81 50 3,91 0,19 0,02 -3,72 72 320 17716 9,78 60 4,09 0,10 0,02 -3,99
27 209 6940 8,85 50 3,91 0,13 0,02 -3,78 73 72 17788 9,79 60 4,09 0,03 0,02 -4,06
29 303 7243 8,89 51 3,93 0,20 0,02 -3,73 74 347 18135 9,81 60 4,09 0,10 0,02 -4,00
31 289 7532 8,93 52 3,95 0,19 0,02 -3,77 75 332 18467 9,82 60 4,09 0,08 0,02 -4,01
33 300 7832 8,97 53 3,97 0,17 0,02 -3,80 76 350 18817 9,84 60 4,09 0,08 0,02 -4,01
34 309 8141 9,00 53 3,97 0,17 0,02 -3,80 77 304 19121 9,86 60 4,09 0,07 0,02 -4,03
35 304 8445 9,04 54 3,99 0,16 0,02 -3,83 78 197 19318 9,87 60 4,09 0,06 0,02 -4,03
37 321 8766 9,08 54 3,99 0,17 0,02 -3,82 79 300 19618 9,88 60 4,09 0,08 0,02 -4,01
38 304 9070 9,11 54 3,99 0,15 0,02 -3,84 80 370 19988 9,90 60 4,09 0,08 0,02 -4,01
39 291 9361 9,14 54 3,99 0,14 0,02 -3,85 81 338 20326 9,92 60 4,09 0,08 0,02 -4,02
40 316 9677 9,18 54 3,99 0,14 0,02 -3,85 82 345 20671 9,94 60 4,09 0,09 0,02 -4,01
42 302 9979 9,21 54 3,99 0,14 0,02 -3,85 84 201 20872 9,95 60 4,09 0,05 0,02 -4,04
43 300 10279 9,24 54 3,99 0,13 0,02 -3,85 85 267 21139 9,96 60 4,09 0,06 0,02 -4,04
44 318 10597 9,27 54 3,99 0,14 0,02 -3,85 87 332 21471 9,97 60 4,09 0,07 0,02 -4,02
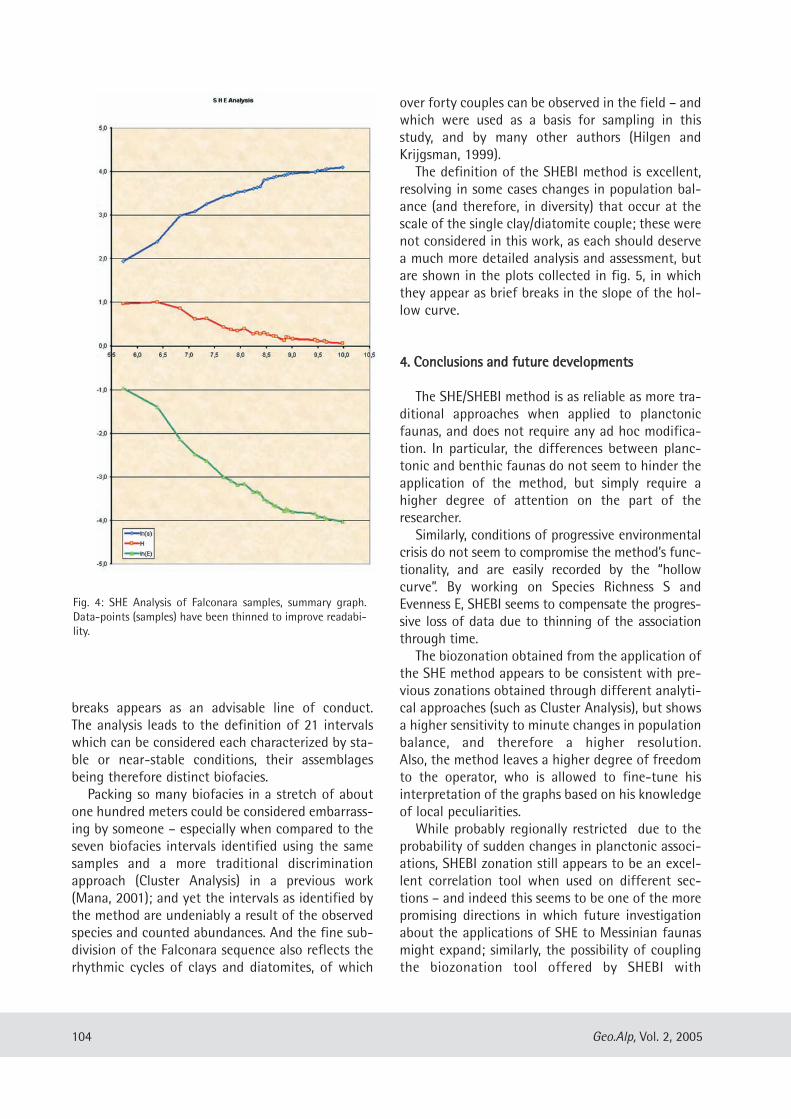
breaks appears as an advisable line of conduct.The analysis leads to the definition of 21 intervalswhich can be considered each characterized by sta-ble or near-stable conditions, their assemblagesbeing therefore distinct biofacies.Packing so many biofacies in a stretch of about
one hundred meters could be considered embarrass-ing by someone – especially when compared to theseven biofacies intervals identified using the samesamples and a more traditional discriminationapproach (Cluster Analysis) in a previous work(Mana, 2001); and yet the intervals as identified bythe method are undeniably a result of the observedspecies and counted abundances. And the fine sub-division of the Falconara sequence also reflects therhythmic cycles of clays and diatomites, of which
over forty couples can be observed in the field – andwhich were used as a basis for sampling in thisstudy, and by many other authors (Hilgen andKrijgsman, 1999). The definition of the SHEBI method is excellent,
resolving in some cases changes in population bal-ance (and therefore, in diversity) that occur at thescale of the single clay/diatomite couple; these werenot considered in this work, as each should deservea much more detailed analysis and assessment, butare shown in the plots collected in fig. 5, in whichthey appear as brief breaks in the slope of the hol-low curve.
44.. CCoonncclluussiioonnss aanndd ffuuttuurree ddeevveellooppmmeennttss
The SHE/SHEBI method is as reliable as more tra-ditional approaches when applied to planctonicfaunas, and does not require any ad hoc modifica-tion. In particular, the differences between planc-tonic and benthic faunas do not seem to hinder theapplication of the method, but simply require ahigher degree of attention on the part of theresearcher.Similarly, conditions of progressive environmental
crisis do not seem to compromise the method’s func-tionality, and are easily recorded by the “hollowcurve”. By working on Species Richness S andEvenness E, SHEBI seems to compensate the progres-sive loss of data due to thinning of the associationthrough time.The biozonation obtained from the application of
the SHE method appears to be consistent with pre-vious zonations obtained through different analyti-cal approaches (such as Cluster Analysis), but showsa higher sensitivity to minute changes in populationbalance, and therefore a higher resolution. Also, the method leaves a higher degree of freedomto the operator, who is allowed to fine-tune hisinterpretation of the graphs based on his knowledgeof local peculiarities.While probably regionally restricted due to the
probability of sudden changes in planctonic associ-ations, SHEBI zonation still appears to be an excel-lent correlation tool when used on different sec-tions – and indeed this seems to be one of the morepromising directions in which future investigationabout the applications of SHE to Messinian faunasmight expand; similarly, the possibility of couplingthe biozonation tool offered by SHEBI with
104 Geo.Alp, Vol. 2, 2005
Fig. 4: SHE Analysis of Falconara samples, summary graph.Data-points (samples) have been thinned to improve readabi-lity.
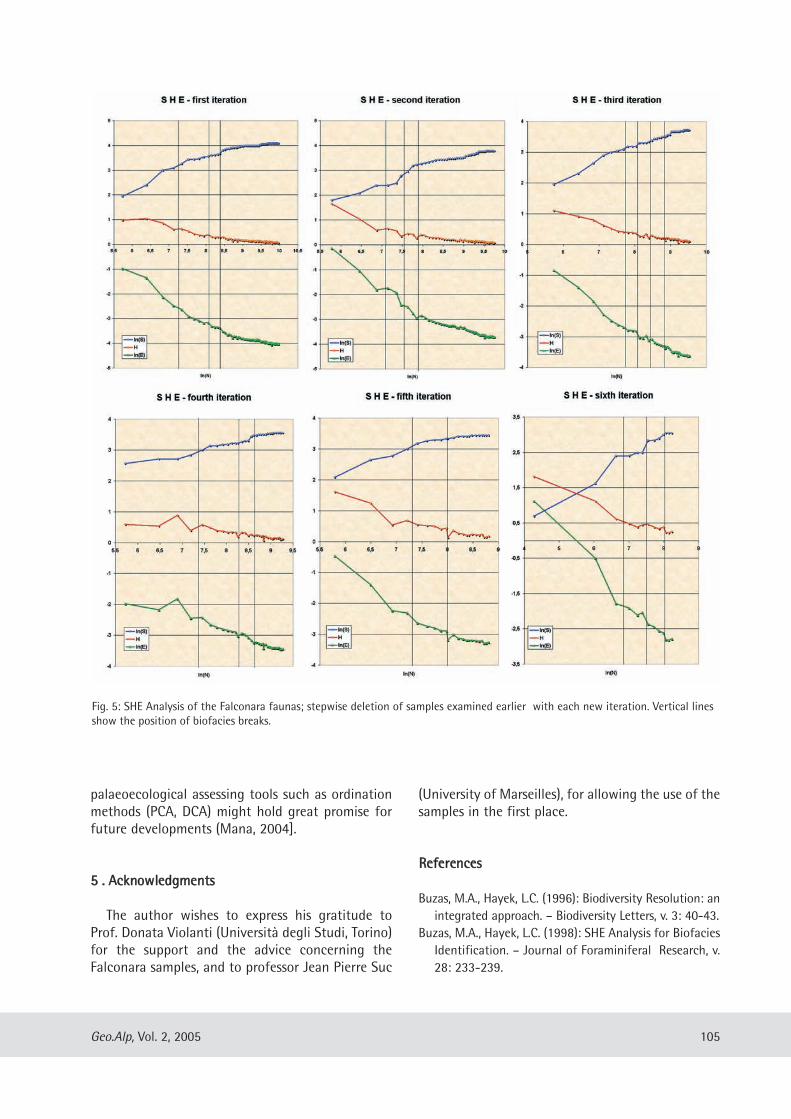
palaeoecological assessing tools such as ordinationmethods (PCA, DCA) might hold great promise forfuture developments (Mana, 2004].
55 .. AAcckknnoowwlleeddggmmeennttss
The author wishes to express his gratitude toProf. Donata Violanti (Università degli Studi, Torino)for the support and the advice concerning theFalconara samples, and to professor Jean Pierre Suc
(University of Marseilles), for allowing the use of thesamples in the first place.
RReeffeerreenncceess
Buzas, M.A., Hayek, L.C. (1996): Biodiversity Resolution: anintegrated approach. – Biodiversity Letters, v. 3: 40-43.
Buzas, M.A., Hayek, L.C. (1998): SHE Analysis for BiofaciesIdentification. – Journal of Foraminiferal Research, v.28: 233-239.
105Geo.Alp, Vol. 2, 2005
Fig. 5: SHE Analysis of the Falconara faunas; stepwise deletion of samples examined earlier with each new iteration. Vertical linesshow the position of biofacies breaks.

Colalongo, M.L., di Grande, A., D’Onofrio, S., Giannelli, L.,Iaccarino, S., Mazzei, R., Poppi Brigatti, M.F., Romeo,M., Rossi, A., Salvatorini, G., (1979): A proposal for theTortonian/Messinian boundary. – Ann. Géol. PaysHellén., Tome hors série, v. 1, pp. 285-294.
Ferrari, C (2001): Biodiversità. – 36 pp, Zanichelli,Bologna.
Hayek, L.C., Buzas, M.A. (1997): Surveying NaturalPopulations. – 563 pp., Columbia University Press, NewYork.
Hayek, L. C., Buzas, M. A. (1998): SHE analysis: an inte-grated approach to the analysis of forest biodiversity.– In Dallimeier, F., Comkey, J. (eds.) Forest BiodiversityResearch, Monitoring and Modeling, 311-321,UNESCO and Parthenon Publishing Group, Paris.
Hilgen, F.J., Krijgsman, W. (1999): Cyclostratigraphy adastrochronology of the Tripoli diatomite formation. –Terra Nova, vol. 11, No. 1: 16-22.
Hilgen F.J., Iaccarino S., Krijgsman,W., Villa, G., LangereisC.G., Zachariasse W.J. (2000): The Global BoundaryStratotype Section and Point (GSSP) of the MessinianStage (uppermost Miocene). – Episodes, Vol. 23,no. 3:172-178.
Leponce, M., Theunis, L., Delabie, J. H. C. and Roisin, Y.(2004): Scale dependence of diversity measures in aleaf-litter ant assemblage. – Ecography v. 27: 253-267.
Mana, D. (2001): Metodi Quantitativi applicati allo studiodei Foraminiferi del Messiniano di Falconara (SiciliaMeridionale). – Università degli Studi, Torino, gradua-tion research paper.
Mana, D. (2004): SHE Characterization of the PlanktonicForaminifera Assemblages from the Falconara andCapodarso Sections (Messinian), Sicily, Italy. – Oralpresentation to the 32nd International GeologicalCongress, Florence, August 20-28, 2004; abstract in32nd International Geological Congress, Abstracts,Part 1, IUGS, Florence.
Osterman, L.E., Buzas, M.A., Hayek, L.C. (2002): SHEAnalysis for Biozonation of Benthic ForaminiferalAssemblages from Western Arctic Ocean. – PALAIOS,v. 17: 297-303.
Small, C.J., McCarthy, B.C.(2002): Spatial and temporalvariability of herbaceous vegetation in an easterndeciduous forest. – Plant Ecology, v. 64: 37-48.
Smart, C.W. (2002): Environmental Applications of Deep-Sea Benthic Foraminifera. – In Haslett, K.S. (ed.):Quaternary Environmental Micropalaeontology,14-58, Arnold Publishers, London.
106 Geo.Alp, Vol. 2, 2005
Fig. 6: Schematic comparison of the biozonation based on Clu-ster Analysis [Mana, 2001], and the SHE biozonation (thiswork). Colors are purely indicative and have no stratigraphicalmeaning.
Manuscript submitted: November 29, 2004Revised manuscript accepted: June 2, 2005

Geo.Alp, Vol. 2, S. 107–113, 2005
UURRSSUUSS SSPPEELLAAEEUUSS RROOSSEENNMMÜÜLLLLEERR,, 11779944 FFRROOMM TTHHEE VVEENNEETTIIAANN RREEGGIIOONN OOFF NNOORRTTHHEERRNN IITTAALLYY::PPRREELLIIMMIINNAARRYY NNOOTTEESS OONN IITTSS EEVVOOLLUUTTIIOONNAARRYY PPAATTHH
Cinzia Galli1, Mario Rossi2, Giuseppe Santi3
With 6 figures and 1 plate
AAbbssttrraaccttAccording to morphometric data, population remains of bears ascribed to the deningeri-spelaeus group have been
identified in caves of the Venetian region of Northern Italy: Cerè Cave, Covoli di Velo, San Donà di Lamon and Veja.Some initial observations about the evolutionary path of these cave bears are presented.
107
1 Museo Civico di Storia Naturale, Parco del Vecchio Passeggio, I-26100 Cremona (Italy)2 Museo Civico di Storia Naturale, Lungadige Porta Vittoria 9, I-37129 Verona (Italy)3 Dipartimento di Scienze della Terra, Via Ferrata 1, I-27100 Pavia (Italy), e-mail [email protected] (corresponding author)
IInnttrroodduuccttiioonn aanndd pprreevviioouuss wwoorrkk
Numerous caves distributed over the Alpine and pre-Alpine areas including Cerè Cave, the Covoli di Velo andVeja caves (Province of Verona), and the San Donà diLamon cave (Province of Belluno) (Fig. 1) are of greatimportance, both historically and for theabundance of bear fossils. They provide alarge number of morphometric datastimulating several interesting considera-tions on the presence of these fossils inthe Venetian region. The studied depositsshow that these areas were inhabitedeither by Ursus spelaeus Rosenmüller,1794, or Ursus arctos Linnaeus, 1758albeit in different proportions; in fact,the cave bear – Ursus spelaeus - repre-sents the most abundant species. Untilnow, the presence of Ursus deningeri VonReichenau, 1906 has only been assumed(Zorzin et al., in press). The most recentstudies (i.e Rossi & Santi, 2005) on newlyfound fossils from the Cerè Cave, themost significant results of which will bepresented in this paper, have confirmedthe presence of this species. Currently,
radiometric data are available for only a few caves inNorthern Italy. The best known is the Grotta SopraFontana Marella (Varese Province, Lombardy). For othercaves (e.g. the Caverna Generosa, Varese Province) dataare still incomplete (Bona, 2004) and are lacking in theVenetian region. Radiometric data and aminoacid
Fig. 1: Geographic position of the main caves of the Venetian region.

racemization of the bear bones from Grotta SopraFontana Marella provide the following ages: FM4 over26000 years BP, FM2, 22310 ±200 and FM1 21810±200years BP (Perego et al., 2001).
For many years the Venetian caves have constitutedan important research target. A review of the inventoryof the Pleistocene-Holocene fauna from these caves wascompiled by Bon et al. (1991, cum bibl.) based on fossilsstored at different localities in Northern Italy. Morerecently, studies on populations of bears and other fossilgroups from the Cerè Cave, Covoli di Velo, San Donà diLamon and Veja caves have been published by Rossi &Santi (2001 a, b, 2002), Zorzin et. al. (2003, 2004 and inpress) and Rossi et al. (2004).
BBrriieeff bbaacckkggrroouunndd oonn tthhee ssttrraattiiggrraapphhyyooff tthhee CCeerrèè CCaavvee
The Cerè Cave, known also as the “Tana dell’Orso” orthe “Tanasela” (Fig. 2), is located at an altitude of about750 m a.s.l. and is 12 m deep; it opens at the hydro-graphically right side of the Vajo dell’Anguilla within theRosso Ammonitico Formation about 150 m east ofCeredo (S. Anna di Alfaedo) village. The entrance is neara distinct fracture of the slope that characterizes theright side of the Vajo dei Falconi. From bottom to top, the
7.50 m thick stratigraphic succession is composed (Zorzinet al., 2003) as follows:11. Ferrous-manganesiferrous clay containing concre-
tions (at the karstic bed rock contact).22. Calcareous concretions, locally very thick.33. Fine-grained, mixed carbonate-siliciclastic sand with
small amounts of clay filling the bottom of thedepressions and the karst fissures. Locally, a thin layerof yellow or reddish clay is present below concre-tion 4.
44. Concretion rich in siliceous and patinated detritalmaterial.
55. Plastic clay containing pebbles up to 1 cm in size.66. Horizon with concretions.77. Plastic red clay containing rare fossil remains and
siliceous detrital fragments with diameters up to 5cm.
88. Red clay with abundant pebbles of chert and slightlyaltered gravel.
99. Dark layer rich in bone remains mostly belonging toCanis lupus containing concretions and rich insiliceous and rare chert pebbles with diameters of 1 to3 cm.
1100. Dark layer rich in bone remains predominantlybelonging to Ursus, with calcareous pebbles about 2cm in size.
1111. Strongly cemented bone-breccia, with abundantremains of Ursus, Canis lupus and Marmota.
108 Geo.Alp, Vol. 2, 2005
Fig. 2: Geographic position of the Cerè Cave. A – Lateral wall of the cave in bone breccias. B – Entrance of the Cerè Cave.
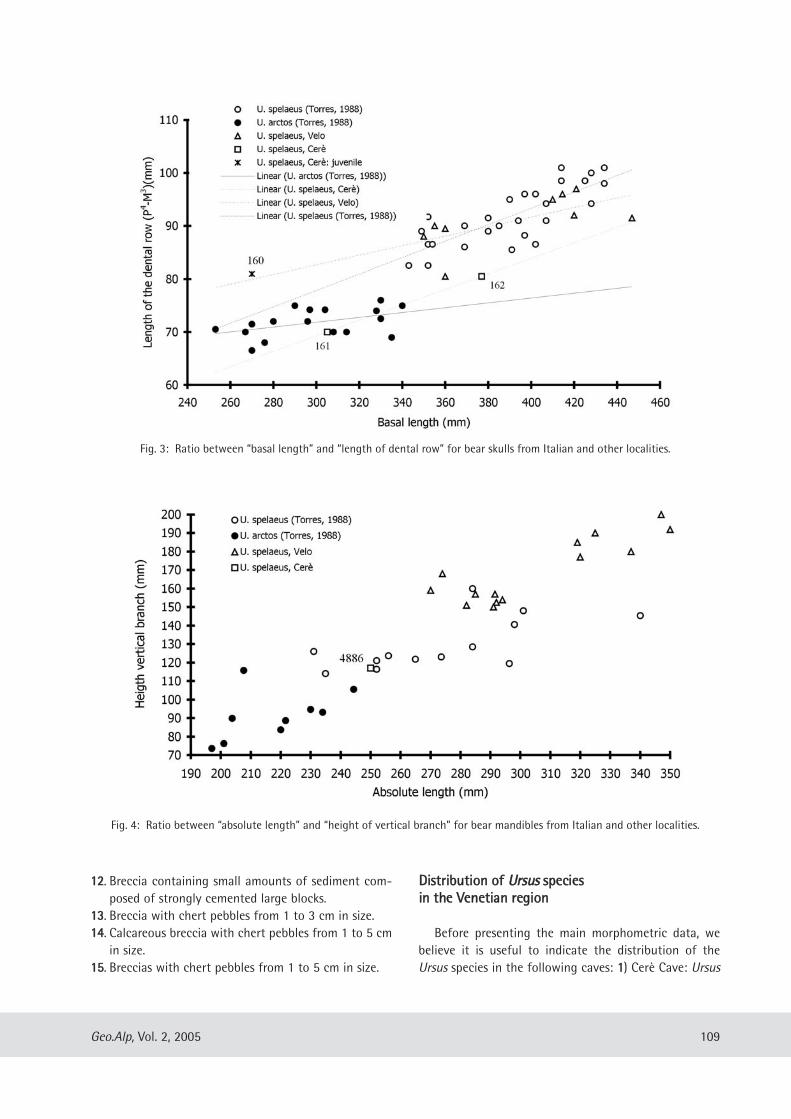
1122. Breccia containing small amounts of sediment com-posed of strongly cemented large blocks.
1133. Breccia with chert pebbles from 1 to 3 cm in size.1144. Calcareous breccia with chert pebbles from 1 to 5 cm
in size.1155. Breccias with chert pebbles from 1 to 5 cm in size.
DDiissttrriibbuuttiioonn ooff UUrrssuuss ssppeecciieessiinn tthhee VVeenneettiiaann rreeggiioonn
Before presenting the main morphometric data, webelieve it is useful to indicate the distribution of theUrsus species in the following caves: 11) Cerè Cave: Ursus
109Geo.Alp, Vol. 2, 2005
Fig. 3: Ratio between “basal length” and “length of dental row” for bear skulls from Italian and other localities.
Fig. 4: Ratio between “absolute length” and “height of vertical branch” for bear mandibles from Italian and other localities.
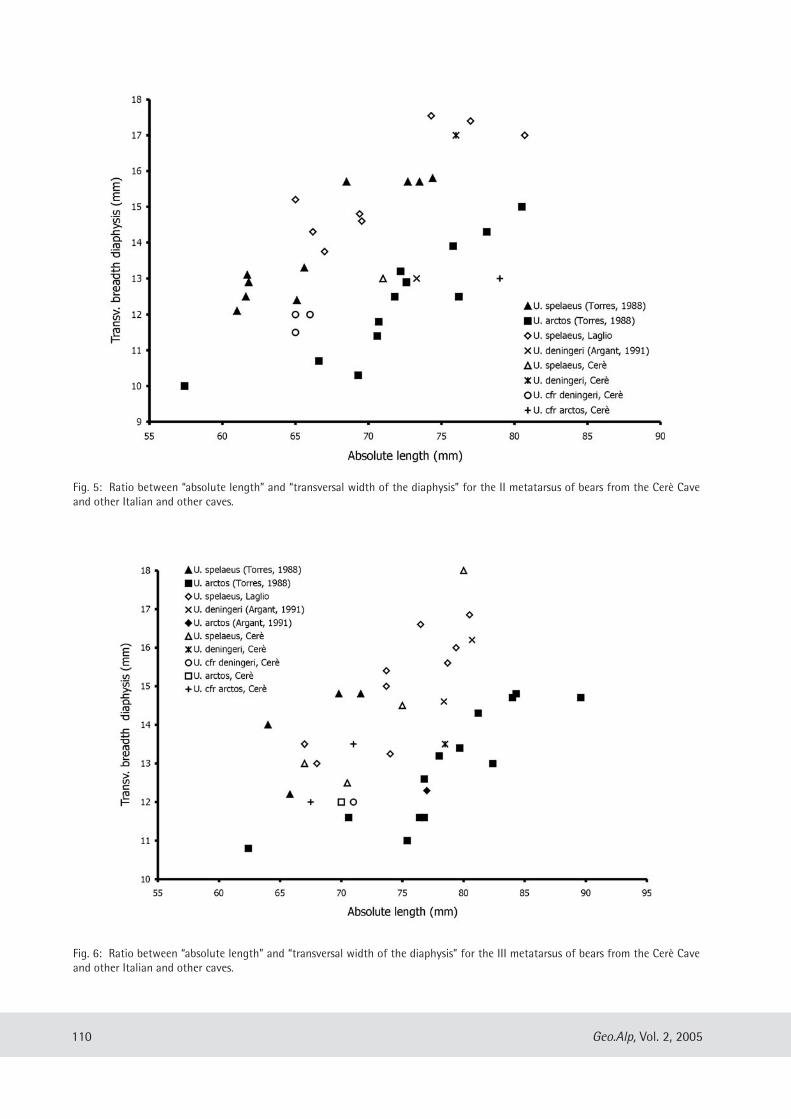
110 Geo.Alp, Vol. 2, 2005
Fig. 6: Ratio between “absolute length” and “transversal width of the diaphysis” for the III metatarsus of bears from the Cerè Caveand other Italian and other caves.
Fig. 5: Ratio between “absolute length” and “transversal width of the diaphysis” for the II metatarsus of bears from the Cerè Caveand other Italian and other caves.

deningeri, U. spelaeus, U. arctos; 22) Covoli di Velo: Ursusspelaeus; 33) Veja: Ursus spelaeus; 44) San Donà di Lamon:Ursus spelaeus (Pl. 1). Considering the rarity of fossilspertianing to U. deningeri not only in Northern Italy, butalso in the rest of the peninsula, their presence within theCerè Cave is of great importance.
MMoorrpphhoommeettrryy
Morphometric analysis was carried out on severalhundreds of fossils from a large portion of the skeleton(except for the vertebrae, ribs and other anatomic partswhose limited number of specimens prevented an indis-putable analysis) stored in the Museo Civico di StoriaNaturale of Verona and compared with other fossils fromNorthern Italy (Grotta del Buco dell’Orso – Laglio, ComoProvince; Grotta Sopra Fontana Marella - VareseProvince; Grotta delle Streghe – Sambughetto Valstrona,Verbania Province) including foreign examples, particu-larly from Spanish caves (Torres, 1988). The findings haveallowed us to advance a number of hypotheses (Figs. 3-6).aa)) CCeerrèè CCaavvee: The morphometric data show the presence
of populations from the deningeri-spelaeus groupand the large number of fossils, especially of themetapodial bones, have confirmed the above men-tioned observations.
bb)) CCoovvoollii ddii VVeelloo: Unlike the Cerè Cave, the fossils areexclusively from larger-sized bears while those inmedium- to small-size ranges ones appear to be verypoorly represented.
cc)) SSaann DDoonnàà ddii LLaammoonn aanndd VVeejjaa: The morphometricanalysis of the rather limited remains in these locali-ties confirms the presence of relatively medium- tolarge-sized populations similar to those that lived inthe Covoli di Velo region.
HHyyppootthheessiiss aabboouutt tthhee ppoossssiibblleeUUrrssuuss ddeenniinnggeerrii ““ttrraacckk ooff wwaayyss””
The presence of the deningerian remains in the CerèCave, rarely found in Central Italy and the Alpine andpre-Alpine sectors of the Western and Central Alps, mayindicate migration paths that initially followed a N-S-direction, possibly encouraged by the overall mildness ofthe climate in the more southern regions, and later alsoin an E-W-direction. The lack of the Ursus deningeriremains in other areas may be due to a gap in the fossilrecord linked to inadequate fossil preservation or unsuc-cessful discovery of the deposits. However, it may also be
a consequence of a limited expansion of these formergroups that were able to colonize only in this limited areain Northern Italy. The remaining zones could have beenfurther colonized starting from a supposed initial point,represented by the Venetian populations originally fromCentral Europe, which experienced a rapid and articulat-ed evolution.
PPrreelliimmiinnaarryy ccoonncclluuddiinngg rreemmaarrkkss
Based on the morphometric analysis that shows thebear fossils belong to the deningeri-spelaeus group, somepreliminary conclusions can be drawn:
aa)) The main caves of the Venetian region were inhab-ited by bears of the deningeri-spelaeus group, but inthe Cerè Cave the continuous presence of bothUrsus deningeri and Ursus spelaeus (medium- tolarge-sized) from their intermediate to final evolu-tionary stages is certain. In other regions only largesized cave bear populations are evident and linkedto the final phase of the evolutionary path of thisspecies.
bb)) The presence of the three species in the Cerè Caveindicates its prolonged inhabitation in ancient timescompared to the other caves. Hence, the Ursusdeningeri population may represent the originalnucleus from which subsequent forms may havedeveloped with their final examples being discoveredin the other caves examined. These populations aremorphometrically comparable to those from the morerecent beds of the Grotta Sopra Fontana Marelladated 21810±200 years BP (Perego et al., 2001). Some data indicate the presence of Ursus deningeri in
the Delle Ossa Cave near Zandobbio village (BergamoProvince, Lombardy), but further investigations arerequired to confirm its occurrence in this area. If futureresearch confirms the exclusiveness of the findings in theCerè Cave, its importance will increase. In fact, on thebasis of this data, this zone could represent an expansionnucleus for the Venetian region as well for the whole ofNorthern Italy.
AAcckknnoowwlleeddggmmeennttss
The authors thank Prof. D. Nagel (Vienna) for usefuladvice and critical reading of the manuscript and Dr. G.Papalia (Pavia) for revision of the English.
This study was supported by a FAR grant contribution.
111Geo.Alp, Vol. 2, 2005

RReeffeerreenncceess
Argant, A. (1991): Carnivores quaternaires de Bourgogne.D– ocuments des Laboratoire de Géologie Lyon, 115 :1-301.
Bon, M., Piccoli, G. & Sala B. (1991): I giacimenti quater-nari di vertebrati fossili nell’Italia nord-orientale. –Mem. Sci. Geol., Padova, 43: 185-231.
Bona, F. (2004): Preliminary analysis on Ursus spelaeusRosenmüller & Heinroth, 1794 populations from“Caverna Generosa” (Lombardy-Italy). – CahiersScientifiques, Hors série 2: 87-98.
Perego, R., Zanalda, E., Tintori, A. (2001): Ursus spelaeusfrom Grotta sopra Fontana Marella, Campo dei FioriMassif (Varese, Italy): morphometry e paleoecology. –Riv. It. Paleont. Strat., 107 (3): 451-462.
Rossi, M., Santi, G. (2001a): La fauna pleistocenica dellaGrotta del Cerè (Verona). 1 – Prime osservazioni suiresti craniali e mandibolari di ursidi. – Bollettino delMuseo Civico di Storia Naturale di Verona, sez. diGeologia, Paleontologia e Preistoria, 25: 59-72.
Rossi, M., Santi, G. (2001b): Archaic and recent Ursusspelaeus forms from Lombardy and Venetia region(North Italy). – Cadernos Lab. Xeológico de LaxeCoruña, 26: 317-323.
Rossi, M., Santi, G. (2002): Gli ursidi dei Covoli di Velo(Verona) e di S. Donà di Lamon (Belluno). I –Preliminare analisi morfologica e morfometrica deiresti craniali e mandibolari. – Bollettino del MuseoCivico di Storia Naturale di Verona, sez. di Geologia,Paleontologia e Preistoria, 26: 33-41.
Rossi, M., Santi, G. (2005): What differences betweenUrsus deningeri Von Reichenau and Ursus spelaeusRosenmüller-Heinroth? The bear mandibles fromVenetia Region caves (N. Italy). – “V Giornate diPaleontologia” Urbino 20-22 Maggio 2005, Abstractsvol., p. 61.
Rossi, M., Santi, G., Zorzin, R. (2004): Distribuzione diUrsus gr. deningeri-spelaeus nell’Italia Settentrionalenel Pleistocene medio-superiore ed implicazioni cli-matico-evolutive. – XXXV Congresso Società Italianadi Biogeografia “Biogeografia delle Alpi e PrealpiCentro-orientali” Rabbi (TN) 6-9 Settembre 2004,Abstracts vol., p. 67.
Torres Pérez Hidalgo T. (1988) - Osos (Mammalia,Carnivora, Ursidae) del Pleistocene Ibérico (U.deningeri Von Reichenau, U. spelaeus Rosemüller-Heinroth, U. arctos Linneo). – Boletín Geológico yMinero. I Filogenia, distribution stratigrafica ygeografica. Estudio anatomico y metrico del craneo:3-46. II Estudio anatomico y metrico de la mandibula,hioides, atlas y axis: 220-249. III Estudio anatomico ymetrico del miembro toracico, carpo y metacarpo:359-412. lV Estudio anatomico y metrico del miembropelviano, tarso, metatarso y dedos: 516-577. VDentiction decidual, formula dentaria y denticionsuperior: 660-714. VI Denticion inferior: 886-940.
Zorzin, R., Santi, G., Rossi, M. (2003): I principali mam-miferi quaternari della Grotta del Cere’ (Monti Lessini- VR) conservati presso il Museo Civico di StoriaNaturale di Verona. – Thalassia Salentina, 26 (2003)suppl.: 183-190.
Zorzin, R., Bona, F., Accordini M. (2004): Cave bearremains from “Covoli di Velo” (Verona-Italy): newfindings from recent stratigraphic excavations. –Cahiers Scientifiques, Hors série 2: 135-138.
Zorzin, R., Rossi, M., Santi, G. (in press): Metapodial bonesof Ursus from Cerè Cave (Venetia Region, North Italy).Cranium.
112 Geo.Alp, Vol. 2, 2005
Manuscript submitted: November 26, 2004Revised manuscript accepted: June 14, 2005
Plate 1: Ursus spelaeus Rosenmüller, 1794. A – Skull V160 (Cerè Cave), dorsal view, B – Skull V 162 (Cerè Cave), dorsal view, C – Man-dible V 4673 (Cerè Cave), internal lateral view, D – Mandible V 2886 (Veja), external lateral view, E – Mandible V 2887 (Veja), internallateral view, F – Mandible V 9889 B (Covoli di Velo), external lateral view, G – Skull V 161 (Cerè Cave), lateral view.
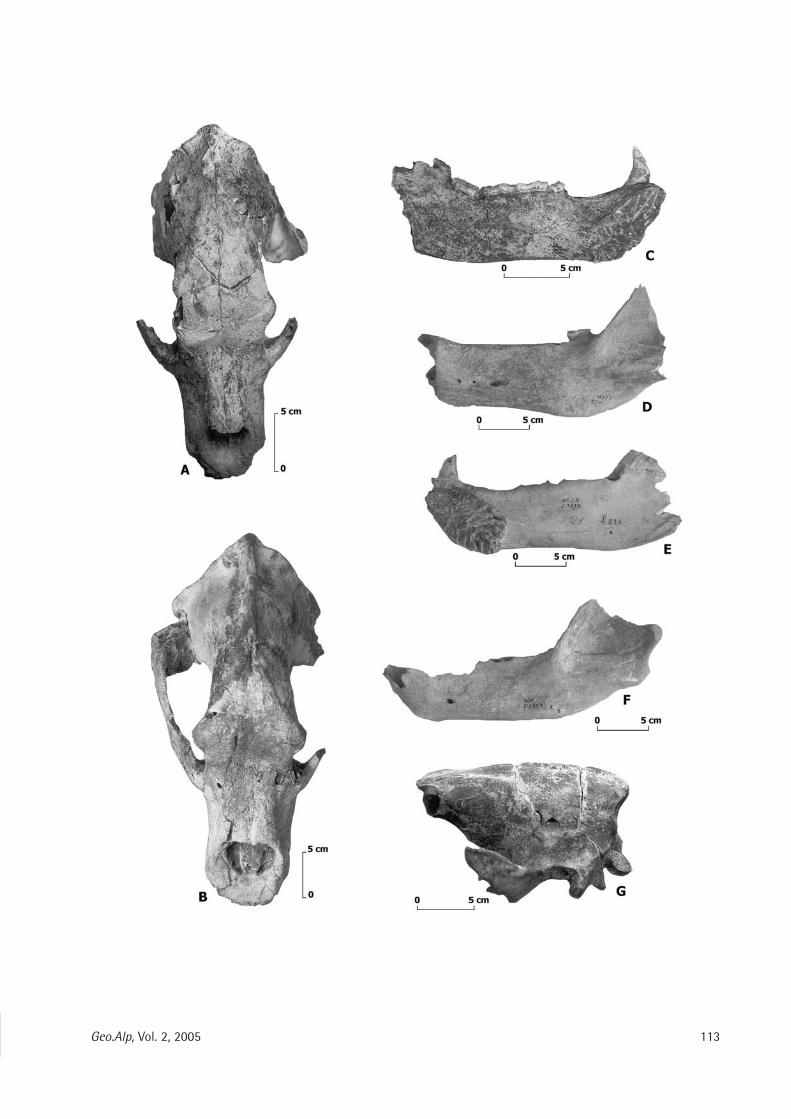
113Geo.Alp, Vol. 2, 2005

Geo.Alp, Vol. 2, S. 115–126, 2005
MMOORRPPHHOOMMEETTRRYY OOFF TTHHEE UURRSSUUSS SSPPEELLAAEEUUSS RREEMMAAIINNSS FFRROOMM VVAALLSSTTRROONNAA ((NNOORRTTHHEERRNN IITTAALLYY))
Alessandro de Carlis1, Enrico Alluvione1, Alessandro Fonte1, Mario Rossi2 & Giuseppe Santi1
With 14 figures and 2 plates
AAbbssttrraaccttMorphometric analyses on cave bear fossils of the Valstrona-Valsesia region (Piedmont, Northern Italy) (in
particular from the Delle Streghe Cave), allow the distinction of at least two or three populations of Ursusspelaeus Rosenmüller, 1794, of different sizes. Elements of smaller size are likely to be found in the Bucodell’Orso Cave (Laglio, Como province, Lombardy), and in the older strata of the Grotta Sopra Fontana Marella–GSFM- (Varese province). Differences in size can be linked to the chronological position of the fossils stud-ied: in fact, fossils of smaller dimensions should be chronologically older, but can also be linked to climaticand thermoregulation factors. The increase of size could represent a response to a cooler climate. An alter-native hypothesis associates this reduction of size to the cooler climatic shift. Observations on the DelleStreghe Cave fossils indicate that they are very similar to those inferred for the GSFM population, linkingthis trend to climatic variation. The reason for an increase in size could also be linked to the rapid evolutionof the cave bears and the Delle Streghe fossils should cover a chronological range approximately similar tothe fossils from GSFM.
115
1 Dipartimento di Scienze della Terra, Via Ferrata 1, I-27100 Pavia (Italy); e-mail: [email protected] (corresponding author)2 Museo Civico di Storia Naturale, Lungadige Porta Vittoria 9, I-37129 Verona (Italy)
IInnttrroodduuccttiioonn
During the Late Pleistocene Ursus spelaeusRosenmüller, 1794 (Rosendhal & Kempe, 2004) col-onized a large part of Europe, migrating to thesouth (central Italy), towards the west (Spain), andtowards the north-west (Great Britain) exhibitingthe most numerous morphological and morphome-trical varieties. Several studies concerning the cavebear group, Ursus spelaeus and its ancestor Ursusdeningeri Von Reichenau, 1906 and U. arctosspecies coeval to U. spelaeus (Rabeder, 1999;Weinstock, 1999; Perego et al., 2001; Rossi & Santi,2001 a, b;; Santi & Rossi, 2001; Santi et al., 2003 andothers) have allowed several hypotheses to beadvanced about the relationships between cave
bears, U. deningeri and coeval species, and aboutpossible phyletic lines indicated by the fossiliferousrecord and by recent mtDNA examination(Hofreiter et al., 2002, 2004). Currently, radiometricdata is only available for a few caves in NorthernItaly: the best known example is the Grotta SopraFontana Marella (Varese Province, Lombardy). Forthe other caves (i.e. the Caverna Generosa, VareseProvince) the data are still incomplete (Bona, 2004).Radiometric data and aminoacid racemization ofthe bear bones from Grotta Sopra Fontana Marellaprovide the following ages: sample FM4 over 26000years BP, FM2, 22310 ±200 years BP and FM121810±200 years BP (Perego et al., 2001). Thewealth of discoveries in various caves in NorthernItaly have shed new light on the distribution of the

vertebrates in this area. In some zones research hasonly just started; one such example is the Valsesia-Valstrona (Piedmont) area (Fig. 1). The aim of thispaper is to summarize previous results regardingthese cave bear populations.
GGeeooggrraapphhiiccaall--ggeeoollooggiiccaall ffrraammee ooff tthhee ssttuuddiieedd aarreeaa
Valstrona is a narrow valley with a V-shaped pro-file in its lower reaches while at its head, near Cimadi Capezzone-Punta del Pizzo (2240 m)-Puntad’Issola (2146 m), it enlarges into a wide cirque. Itwinds for 20 km to Omegna village where itdebouches onto the Orta Lake (Cusio). Near theSambughetto village some caves have formed viakarst processes within the lens of the “MarmoValstrona” formation; this lenticular body is interca-lated between gneisses and micaschists of the “SerieKinzigitico-sillimanitica”. Inside the caves the osteo-logical material, accompanied by yellow loessic clay,collects in the lower parts along the side lanes andcavities. This sediment is frequently covered by hardstalagmitic soil (about 15-20 cm thick), and by greymicaceous and sterile sands interspersed with small-er gravel of more recent age linked to the pluvialwashing away phase. To ensure good preservation
of bones, deposits were only slightly dis-turbed and analysed in situ.
Fossils from Valstrona have beenfound inside the caves known asComplesso dell’Intaglio and Cavernadelle Streghe, near the Cava Sam -bughetto village. The first of these cavesopens out in the upper part of the mar-ble quarry (“Sass Muiè”), it has fiveentrances and a subcircular small gallerycomplex correlated with an older level ofthe water-bearing stratum. The secondcave, called Caverna delle Streghe, is thewidest cave in Verbania Province. It iscomposed of a fossil branch presentlyfoliated by water and by a second activebranch in the marble eroded by the river(Fig. 2). The water source is from theChignolo stream that, after havingcrossed the cave and swelled water fromother tributaries, re-emerges in theStrona River.
The Valsesia fossils are derived from theMt. Fenera (Fig. 3) caves and mainly from the“Ciutarun” and the “Ciota Ciara” cave. The former issituated at 650 m asl, with a large ogival entrance,and it is 55 m long and up to 13 m high. The “CiotaCiara” is located at an altitude of 685 m asl, it is 57m long and the difference in levels internally is upto 18 m. There are two entrances: a southern, nat-ural and a northwestern entrance which wasformed by the collapse of a part of the vault. Thiscave rises upward from SE-NW and ends towardsthe N (Strobino, 1981).
MMaatteerriiaallss aanndd mmeetthhooddss
About one thousand Ursus remains currentlystored in the Museo Civico di Storia Naturale diMilano have been analysed. They have been labelled“MMCCSSNNMM VV”, (abbrevation of MMuseo CCivico di SStoriaNNaturale and VVertebrate), followed by a progressivenumber. A substantial portion of the skeletons ofcubs, juveniles and adult elements is represented(Pls. 1-2). The material is rarely complete, especiallythe skull remains, and in particular in the case ofcubs only skull-caps have been preserved.Preservation is generally good, although some
116 Geo.Alp, Vol. 2, 2005
Fig. 1: Geographic position of the Delle Streghe Cave (Valstrona, Piedmont,Northern Italy).
117Geo.Alp, Vol. 2, 2005
Fig. 2: A – Planimetric scheme and profiles; B – of the Delle Streghe Cave (Cella, 1993, mod.).

traces of erosion can be found in the proximal anddistal ends of limb bones. In addition, some speci-mens showed traces of pathologies (e.g. periarthri-tis and pesudoarthrosis) and generic malformations,traces of predator activities (Fig. 4). The presence ofpredators is indicated by the catlike remains insidethe Delle Streghe fauna with an incomplete rightradius fragment (MSNM V4329) belonging to
Panthera leo spelaea (Goldfuss, 1810) (Fig. 5). Mostof the fossils belong to Ursus spelaeus Rosenmüller,1794, while others with disputed morphologicalfeatures could be classified as Ursus deningeri VonReichenau 1906. However we have considered theseremains as U. spelaeus on the basis of the broadermorphological relationships within this species.Useful morphometric parameters were deducedfrom Hue (1908), Von den Driesch (1976) and Torres(1988).
MMoorrpphhoommeettrryy
SKULL – These fossils, although incomplete, havesome morphometric features that seem to be typi-cal of cave bears. They are generally similar in sizeto examples of U. spelaeus from caves in Spain andslightly larger than those from Caverna delle Ossa(Zandobbio, Bergamo Province, North of Milan).
MANDIBLE – The relationship between the trans-versal diameter of the condyle and the verticaldiameter (Fig. 6) confirms what has been inferredregarding skull morphometric analysis. TheSambughetto specimens are similar in size to thetypical spelaeus (in this paper represented by fossils
118 Geo.Alp, Vol. 2, 2005
Fig. 3: Distribution of the main caves in the Fenera Mt. (Valsesia, Piedmont, Northern Italy). Number 1 is the “Ciutarun”, 2 and 3refer to the “Ciota Ciara”. (Strobino, 1981, mod.).
Fig. 4: AA.. Pathological Ursus bone (specimen MSNM V 4362,Delle Streghe Cave). BB.. Predatory activity traces (specimenMSNM V 4097, Delle Streghe Cave).
from Covoli di Velo Veronese, Verona Province), butthey are larger than those from the Buco dell’Orsocave, whose small sizes can be linked to climaticfactors (Bergmann’s rule). The dimensions of themandibular condyle, but especially the height ofthe mandible below P4, provided additional evi-dence supporting what has been deduced fromskull analysis. Comparison between the fossils stud-ied and samples from some Venetia caves (Grottadel Cerè whose population appears to be older,Covoli di Velo Veronese and S. Donà di Lamon) andfrom Grotta Sopra Fontana Marella –GSFM- (VareseProvince, Lombardy), allows us to place the DelleStreghe bears in an intermediate position betweenancient and modern forms. These data are also sup-ported by dental surface features. Data referred tothe M1 and M2 show the greatest range comparedto those of the other specimens considered (Pocala,Equi, GSFM, Covoli di Velo, Buco dell’Orso, Cavernadelle Fate, Grotta delle Ossa) and a smallerlength/width ratio. This feature could be probablyrelated to local factors and particularly to foodpreferences. But we cannot exclude that this dif-ference in size may be related to sexual dimor-phism.
HUMERUS – As shown in diagram Fig. 7, the DelleStreghe specimens show similar features to thosefrom GSFM. In fact, the absolute dimensions aresimilar. The main difference is evident from thegreater deformation of the diaphyses of theanalysed remains, and particularly in the morerecent forms due to a smaller antero-posteriordiameter.
RADIUS – Data concern-ing the radius seem toconfirm what is shown bythe humeri. In particularsome morphometric rela-tionships (Fig. 8) allow usto affirm that: a) themorphometric character-istics of the specimensstudied are comparablewith those of the GSFM,b) generally, adult ele-ments can be comparedwith those from the olderand intermediate levelsof the GSFM, while the
119Geo.Alp, Vol. 2, 2005
Fig. 5: Panthera leo spelaea (Goldfuss, 1810). Specimen MSNMV 4329 (Delle Streghe Cave). Right radius. AA: External view,BB – Internal view.
Fig. 6: Relationship between Transversal Diameter of the condyle and Vertical diameter inmandibles of Ursus spelaeus from c
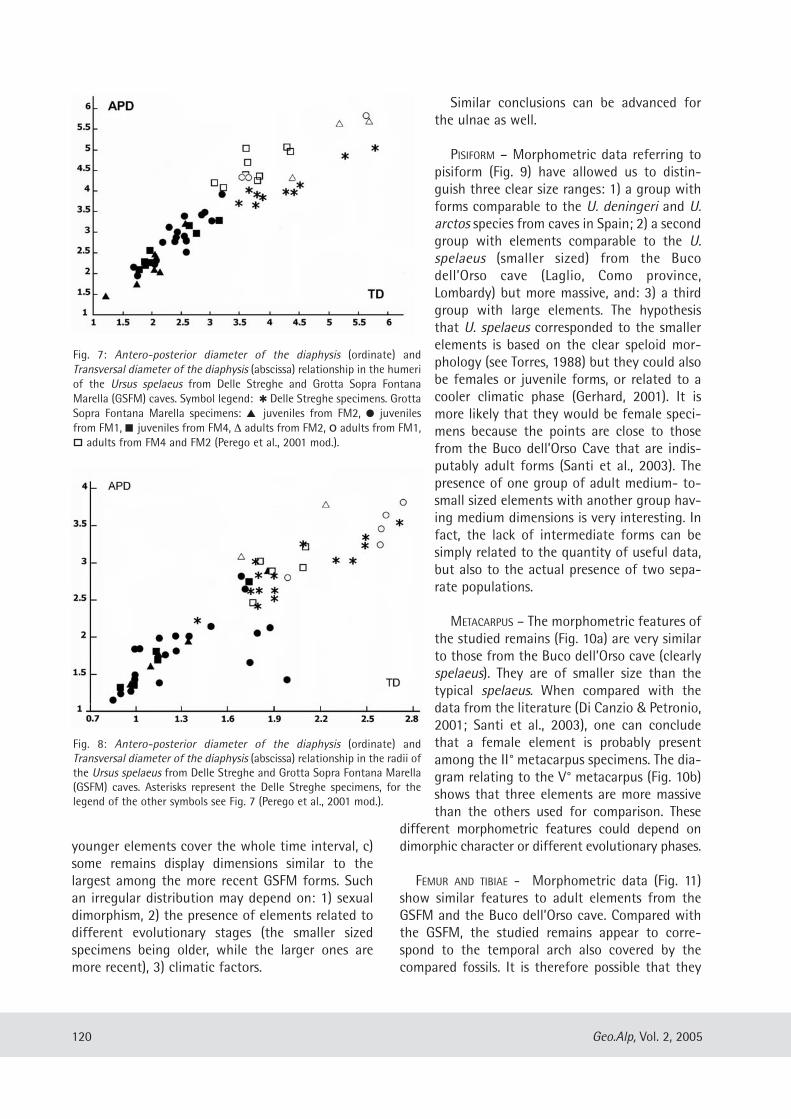
younger elements cover the whole time interval, c)some remains display dimensions similar to thelargest among the more recent GSFM forms. Suchan irregular distribution may depend on: 1) sexualdimorphism, 2) the presence of elements related todifferent evolutionary stages (the smaller sizedspecimens being older, while the larger ones aremore recent), 3) climatic factors.
Similar conclusions can be advanced forthe ulnae as well.
PISIFORM – Morphometric data referring topisiform (Fig. 9) have allowed us to distin-guish three clear size ranges: 1) a group withforms comparable to the U. deningeri and U.arctos species from caves in Spain; 2) a secondgroup with elements comparable to the U.spelaeus (smaller sized) from the Bucodell’Orso cave (Laglio, Como province,Lombardy) but more massive, and: 3) a thirdgroup with large elements. The hypothesisthat U. spelaeus corresponded to the smallerelements is based on the clear speloid mor-phology (see Torres, 1988) but they could alsobe females or juvenile forms, or related to acooler climatic phase (Gerhard, 2001). It ismore likely that they would be female speci-mens because the points are close to thosefrom the Buco dell’Orso Cave that are indis-putably adult forms (Santi et al., 2003). Thepresence of one group of adult medium- to-small sized elements with another group hav-ing medium dimensions is very interesting. Infact, the lack of intermediate forms can besimply related to the quantity of useful data,but also to the actual presence of two sepa-rate populations.
METACARPUS – The morphometric features ofthe studied remains (Fig. 10a) are very similarto those from the Buco dell’Orso cave (clearlyspelaeus). They are of smaller size than thetypical spelaeus. When compared with thedata from the literature (Di Canzio & Petronio,2001; Santi et al., 2003), one can concludethat a female element is probably presentamong the II° metacarpus specimens. The dia-gram relating to the V° metacarpus (Fig. 10b)shows that three elements are more massivethan the others used for comparison. These
different morphometric features could depend ondimorphic character or different evolutionary phases.
FEMUR AND TIBIAE - Morphometric data (Fig. 11)show similar features to adult elements from theGSFM and the Buco dell’Orso cave. Compared withthe GSFM, the studied remains appear to corre-spond to the temporal arch also covered by thecompared fossils. It is therefore possible that they
120 Geo.Alp, Vol. 2, 2005
Fig. 7: Antero-posterior diameter of the diaphysis (ordinate) andTransversal diameter of the diaphysis (abscissa) relationship in the humeriof the Ursus spelaeus from Delle Streghe and Grotta Sopra FontanaMarella (GSFM) caves. Symbol legend: � Delle Streghe specimens. GrottaSopra Fontana Marella specimens: � juveniles from FM2, � juvenilesfrom FM1, � juveniles from FM4, ∆ adults from FM2, o adults from FM1,
adults from FM4 and FM2 (Perego et al., 2001 mod.).
Fig. 8: Antero-posterior diameter of the diaphysis (ordinate) andTransversal diameter of the diaphysis (abscissa) relationship in the radii ofthe Ursus spelaeus from Delle Streghe and Grotta Sopra Fontana Marella(GSFM) caves. Asterisks represent the Delle Streghe specimens, for thelegend of the other symbols see Fig. 7 (Perego et al., 2001 mod.).
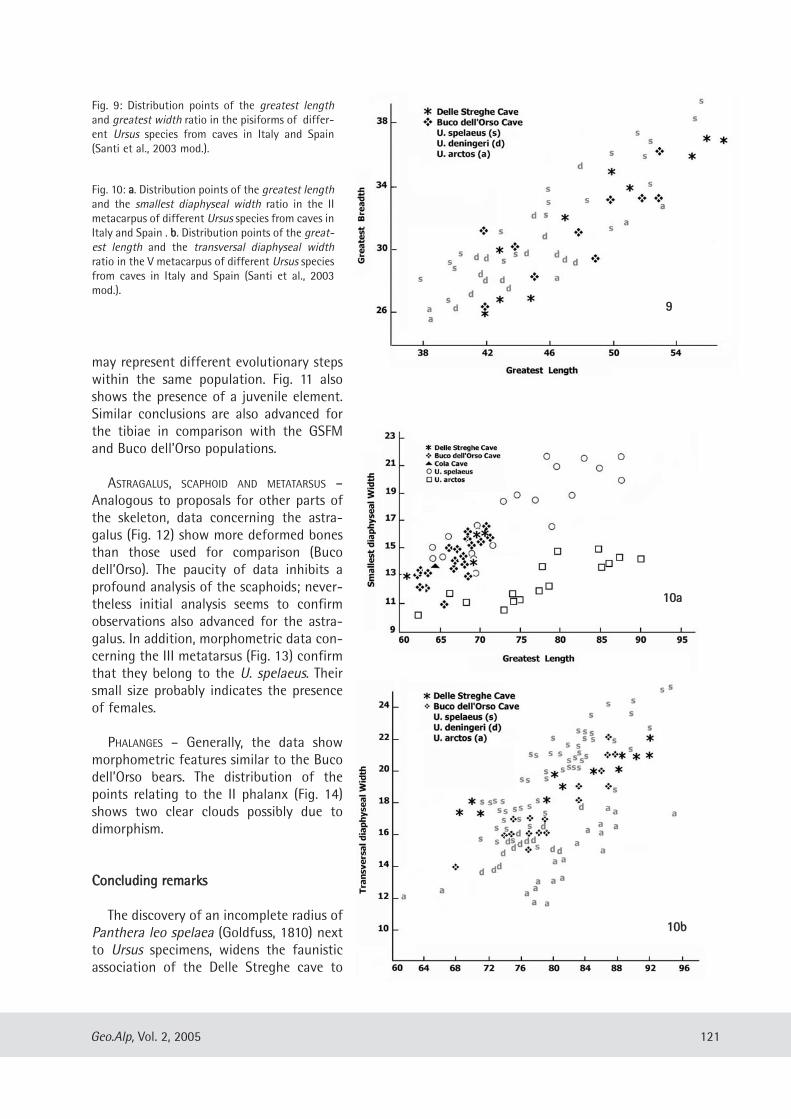
may represent different evolutionary stepswithin the same population. Fig. 11 alsoshows the presence of a juvenile element.Similar conclusions are also advanced forthe tibiae in comparison with the GSFMand Buco dell’Orso populations.
ASTRAGALUS, SCAPHOID AND METATARSUS –Analogous to proposals for other parts ofthe skeleton, data concerning the astra-galus (Fig. 12) show more deformed bonesthan those used for comparison (Bucodell’Orso). The paucity of data inhibits aprofound analysis of the scaphoids; never-theless initial analysis seems to confirmobservations also advanced for the astra-galus. In addition, morphometric data con-cerning the III metatarsus (Fig. 13) confirmthat they belong to the U. spelaeus. Theirsmall size probably indicates the presenceof females.
PHALANGES – Generally, the data showmorphometric features similar to the Bucodell’Orso bears. The distribution of thepoints relating to the II phalanx (Fig. 14)shows two clear clouds possibly due todimorphism.
CCoonncclluuddiinngg rreemmaarrkkss
The discovery of an incomplete radius ofPanthera leo spelaea (Goldfuss, 1810) nextto Ursus specimens, widens the faunisticassociation of the Delle Streghe cave to
121Geo.Alp, Vol. 2, 2005
Fig. 9: Distribution points of the greatest lengthand greatest width ratio in the pisiforms of differ-ent Ursus species from caves in Italy and Spain(Santi et al., 2003 mod.).
Fig. 10: aa. Distribution points of the greatest lengthand the smallest diaphyseal width ratio in the IImetacarpus of different Ursus species from caves inItaly and Spain . bb. Distribution points of the great-est length and the transversal diaphyseal widthratio in the V metacarpus of different Ursus speciesfrom caves in Italy and Spain (Santi et al., 2003mod.).
99
1100aa
1100bb
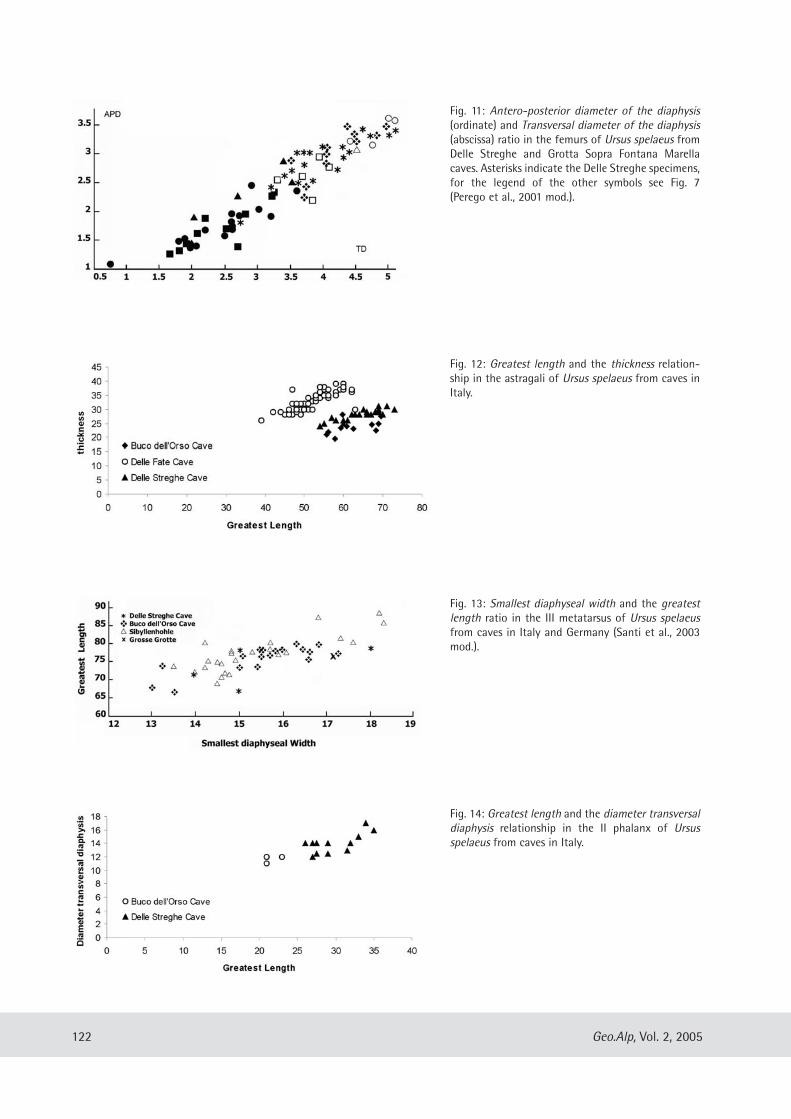
122 Geo.Alp, Vol. 2, 2005
Fig. 11: Antero-posterior diameter of the diaphysis(ordinate) and Transversal diameter of the diaphysis(abscissa) ratio in the femurs of Ursus spelaeus fromDelle Streghe and Grotta Sopra Fontana Marellacaves. Asterisks indicate the Delle Streghe specimens,for the legend of the other symbols see Fig. 7(Perego et al., 2001 mod.).
Fig. 13: Smallest diaphyseal width and the greatestlength ratio in the III metatarsus of Ursus spelaeusfrom caves in Italy and Germany (Santi et al., 2003mod.).
Fig. 14: Greatest length and the diameter transversaldiaphysis relationship in the II phalanx of Ursusspelaeus from caves in Italy.
Fig. 12: Greatest length and the thickness relation-ship in the astragali of Ursus spelaeus from caves inItaly.

other nearby caves (Buco dell’Orso Cave, Delle OssaCave – Zandobbio in Bergamo Province).Pathologies are rare, mainly confined to limbs, andrelated to the senescence of the bears.Morphometric data indicate the presence of at leasttwo populations of cave bears characterized by dif-ferent sizes: the small-size bears are comparable tothe Buco dell’Orso cave bears and those specimensfrom the older levels to the Grotta Sopra FontanaMarella. According to Perego et al. (2001), the dif-ference in size is related to a different evolutionarystep of the bear; small size could correspond tomore ancient forms, namely more primitive ones.The increase in size can be linked to a thermoregu-lation factor following Bergmann’s rule (1847): theincrease in body size yields an advantage in ther-moregulation. Loss of heat in bodies of large size islower, causing a smaller surface-to-volume ratio. Inthis manner large sized populations can colonizecool regions. Moreover, in the case of the studiedbears, an increase in dimensions could also repre-sent a response to a shift towards a cooler climate.In contrast to these authors, Gerhard (2001) andRabeder & Nagel (2001) associate a similar reduc-tion in size to the shift toward cooler conditionsalthough this should be observable in high Alpineregions. The similarity between the Grotta SopraFontana Marella and Delle Streghe Cave fossils leadsus to link this trend to a climatic change, ratherthan to rapid evolution by cave bears.
AAcckknnoowwlleeddggmmeennttss
The authors thank D. Nagel (Vienna) for usefuladvise and critical reading of the manuscript, and G.Papalia (Pavia) for revision of the English. This studywas supported by a FAR grant contribution.
RReeffeerreenncceess
Bergmann, C. (1847): Ueber die Verhaeltnisse derWaemeoekonomie der Thiere zu ihrer Groesse. –Goettinger Studien 3, Pt. 1: 595-708.
Bona, F. (2004): Preliminary analysis on Ursus spelaeusRosenmüller & Heinroth, 1794 populations from“Caverna Generosa” (Lombardy-Italy). – CahiersScientifiques, Hors série 2: 87-98.
Cella, D.G. (1993): Il patrimonio speleologico dellaValstrona. – Labirinti, 13: 2-4.
Di Canzio, E., Petronio, C. (2001): Osservazioni sulla faunaa vertebrati pleistocenici della Grotta Cola ( Abruzzo,Aquila). – Boll. Soc. Paleont. It., 40 (1): 105-114.
Gerhard, W. (2001): The evolution of metapodial bones inthe cave bear group and its biostratigraphical implica-tions. – Cadernos Lab. Xeolòxico de Laxe Coruña, 26 :365-371.
Hofreiter, M., Capelli, C., Krings, M., Waits, L., Conard, N.,Munzel, S., Rabeder, G., Nagel, D., Paunovic, M.,Jambresic, G., Meyer, S., Weiss, G., Pääbo, S. (2002):Ancient DNA analyses reveal high mitochondrial DNAsequence diversity and parallel morphological evolu-tion of late Pleistocene cave bears. – MolecularBiology and Evolution, 19 (8):1244-1250.
Hofreiter, M., Rabeder, G., Jaenicke-Deprés, V., Withalm,G., Nagel, D., Paunovic, M., Jambr?sic, G. & Pääbo, S.(2004): Evidence of reproductive isolation betweencave bear population. – Current Biology, 14: 40-43.
Hue, E. (1907): Musée ostéologique. Étude de la faunequaternaire. Ostéometrie des Mammifères. 2 vol. –Librairie C. Reinwold, Schleicher Frères Editeurs, Paris.
Perego, R., Zanalda, E., Tintori, A. (2001): Ursus spelaeusfrom Grotta sopra Fontana Marella, Campo dei FioriMassif (Varese, Italy): morfometry e paleoecology. –Riv. It. Paleont. Strat., 107 (3): 451-462.
Rabeder, G. (1999): Die Evolution des Höhlenbären gebis -ses. – Mitteilungen der Kommission für Quartär for -schung der Österreichischen Akademie der Wissen -schaften, Band II, 102 pp.
Rabeder, G., Nagel, D. (2001): Phylogenetic problems ofthe Alpine Cave Bears. – Cadernos Lab. Xeológico deLaxe Coruña, 26: 359-364.
Rosendhal, W., Kempe, S. (2004): Johann Christian Rosen -müller und der Höhlenbär (1794-2004). „Lebensbilder“aus 210 Jahren. – Natur und Mensch 2003: 145-159.
Rossi, M., Santi, G. (2001 a): Gli ursidi della Grottadell’Orso (Laglio, Como, Lombardia, Italia Setten -trionale). Analisi morfometrica degli arti: indaginepreliminare. – Atti Soc. it. Sc. Nat. Mus. Civ. St. Nat.Milano, 141/2000 (2): 329-336.
Rossi, M., Santi, G. (2001 b): Archaic and recent Ursusspelaeus forms from Lombardy and Venetia region(North Italy). – Cadernos Lab. Xeológico de LaxeCoruña, 26: 317-323.
Santi, G., Rossi, M. (2001): Bears from the Buco dell’OrsoCave (Laglio-Como, Lombardy-Northern Italy). I:Morphometric study of the cranial and mandibularfossil remains. – Atti Ticinensi di Scienze della Terra,Pavia, 42: 75-100.
Santi, G., Rossi, M., Pomodoro, S. (2003): Cave bearsremains from the Buco dell’Orso cave (Lombardy-
123Geo.Alp, Vol. 2, 2005

Italy). Part III – Morphometric analysis of metapodialbones. – Bull. Inst. Royal Sc. Nat. de Belgique, 73: 195-219.
Strobino, F. (1981): Preistoria in Valsesia. Studi sul MonteFenera. – Società Valsesiana di cultura. pp.89.
Torres Pérez Hidalgo, T. (1988). Osos (Mammalia,Carnivora, Ursidae) del pleistocene Ibérico (U. denin-geri Von Reichenau, U. spelaeus Rosenmüller-Heinroth, U. arctos Linneo). – Boll. Geol. y Min.- lFilogenia, distribution stratigrafica y geografica.Estudio anatomico y metrico del craneo: 3-46. llEstudio anatomico y metrico de la mandibula, hioides,atlas y axis: 220-249. lll Estudio anatomico y metricodel miembro toracico, carpo y metacarpo: 359-412. lVEstudio anatomico y metrico del miembro pelviano,tarso, metatarso y dedos: 516-577. V Dentiction deci-
dual,formula dentaria y denticion superior: 660-714.Vl Denticion inferior: 886-940.
Weinstock, J. (1999): The upper Pleistocene mammalianfauna from the Grosse Grotte near Blauberen(Southwestern Germany). – Stuttgarter Beitr. Naturk.,Serie B: 277, 1-49.
Von den Driesch, A. (1976): A guide to the measurementof animal bones from archaeological sites. – PeabodyMuseum Bullettin, 1: 1-137.
124 Geo.Alp, Vol. 2, 2005
Plate 1: Ursus spelaeus Rosenmüller, 1794. Delle Streghe Cave (Sambughetto Valstrona, Piedmont, North Italy). AA – Skull. SpecimenMSNM V 4486, dorsal view. BB – Skull. Specimen MSNM V 5043, dorsal view. CC – Skull. Specimen MSNM V 5041, dorsal view. DD –Skull-cap of cub. Specimen MSNM V 4614, dorsal view. EE – Skull-cap of cub. Specimen MSNM V 4736, dorsal view. FF – Skull-cap ofcub. Specimen MSNM V 4721, dorsal view. GG - III phalanx. Specimen MSNM V 5028, lateral view. HH – Mandible. Specimen MSNM V5059, internal view. II - Skull. Specimen MSNM V 5043, frontal view. JJ – I phalanx. Specimen MSNM V 4988, dorsal view. KK – Radius.Specimen MSNM V 4331, external view. LL – Scapholunar. Specimen MSNM 4781, lateral view. MM – Astragalus. Specimen MSNM 4874,dorsal view. NN – Femur. Specimen MSNM V 4393, caudal view. OO – Radius. Specimen MSNM V 4304, dorsal view.
Manuscript submitted: November 26, 2004Revised manuscript accepted: June 22, 2005
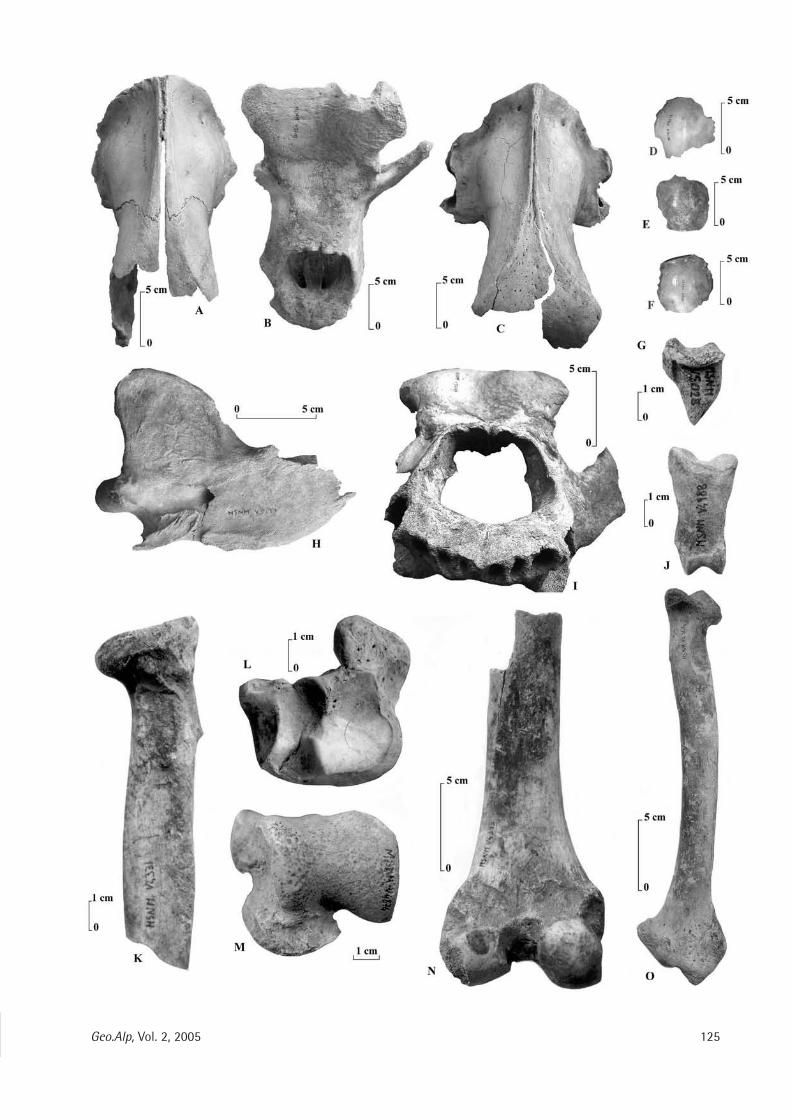
125Geo.Alp, Vol. 2, 2005

126 Geo.Alp, Vol. 2, 2005
Plate 2: Ursus spelaeus Rosenmüller, 1794. Delle Streghe Cave (Sambughetto Valstrona, Piedmont, North Italy). AA - Calcaneus.Specimen MSNM V 4904, dorsal view. BB - IV° metacarpus. Specimen MSNM V 4827, medial view. CC – III° metacarpus. SpecimenMSNM V 4824, medial view. DD – IV° metacarpus. Specimen MSNM V 4828, medial view. EE – II° metacarpus. – V° metacarpus.Specimen MSNM V 4845, lateral view.

Geo.Alp, Vol. 2, S. 127–129, 2005
TTHHEE AACCTTIIVVIITTIIEESS OOFF TTHHEE LLIIGGAABBUUEE SSTTUUDDYY RREESSEEAARRCCHH CCEENNTTRREEOONN TTHHEE TTHHIIRRTTIIEETTHH AANNNNIIVVEERRSSAARRYY OOFF IITTSS FFOOUUNNDDAATTIIOONN
Francesco Garofalo1, Fabrizio Bizzarini2, Federica Ferrieri3
With 4 figures
AAbbssttrraacctt
In 2003 the Ligabue Study Research Centre celebrated its first thirty years of activity with various projectsconcerning research and scientific promotion: the opening of a new exhibition area in the Venice Museumof Natural History and the creation of a multi-themed exhibition in the Palazzo delle Miniere at Fiera diPrimiero (Trento).The new room in the Venice Museum is dedicated to the scientific expedition which took place in the
Ténéré Desert between 1971 and 1973. It briefly examines the history of the expedition, which contributedtowards the foundation of the Ligabue Study Research Centre.The exhibition “From Meteorites to Dinosaurs … to Men” has been staged with the collaboration of the
Comprensorio del Primiero (Trento). Theories about biological evolution act as a bond throughout the exhibi-tion: gathering a wide range of exhibits, the exhibition links the evolutionary potentials which can be foundin the history of terrestrial organisms to the global evolution of the solar system and to human cultural evo-lution.
RRiiaassssuunnttooL’attività del CENTRO STUDI RICERCHE LIGABUE in occasione del trentennale della sua fondazione.
Nel 2003 il Centro Studi Ricerche Ligabue ha celebrato i suoi primi trent’anni di attività con numerose in-iziative nel campo della ricerca e della divulgazione scientifica. Questo secondo aspetto è stato caratterizza-to da due manifestazioni: l’apertura di un nuovo percorso espositivo al Museo di Storia Naturale di Venezia euna mostra politematica presso il Palazzo delle Miniere a Fiera di Primiero.La nuova sala del Museo Veneziano è dedicata alla spedizione scientifica nel deserto del Ténéré, svoltasi
negli anni tra il 1971 e il 1973. Riassume brevemente la storia di quella spedizione che stimolò la nascitastessa del Centro Studi Ricerche Ligabue. La mostra “dalle Meteoriti ai Dinosauri…all’Uomo” è stata realizzata in collaborazione con il Comprensorio
del Primiero. Le teorie dell’evoluzione biologica fanno da collante all’intero percorso espositivo, che nel riu-nire l’ampia varietà di reperti, collega le potenzialità evolutive riscontrabili nella storia degli organismi terre-stri, all’evoluzione complessiva del Sistema Solare e alla stessa evoluzione culturale umana.
127
1 Via Monte San Michele 20/A, 30171 Mestre – [email protected] Cannaregio 1269/A, 30121 Venezia 3 Università Ca’ Foscari di Venezia, Dottorato in Studi Iberici, Anglo-Americani e dell’Europa Orientale

In 2003 the Ligabue Study Research Centre cele-brated its first thirty years of scientific-cultural ac-tivities with various projects concerning both re-search and promotion. In particular, the Centre’sendeavours have been promoted by two exhibitions.On August 9, 2003, the permanent exhibition enti-tled “From Meteorites to Dinosaurs … to Men” wasopened in the 14th century Palazzo delle Miniere atFiera di Primiero, while on October 25, 2003 theVenice Museum of Natural History, including aroom called the “Dinosaur Fossil Deposit”, was offi-cially reopened to the public. This room is dedicatedto the scientific expedition conducted in 1973 bythe Ligabue Study Research Centre and the Nation-al Museum of Natural History of Paris, and led byGiancarlo Ligabue and Philippe Taquet. This expedi-tion enabled the study of the dinosaur fossil de-posits of Gadoufaoua, in the Ténéré Desert (Niger),whose sands yielded the skeleton of an Oura-nosaurus nigeriensis, now exhibited in Venice. Thesediments of this deposit belong to the Elrhaz for-mation, upper Aptian (lower Cretaceous), andformed in a marshy and deltaic environment, whichwas rich in vegetation and populated by dinosaurs,crocodiles, pterosaurs and fish. The exhibition areaof the Venice Museum enables the visitor to retracethe history of the expedition, its difficulties and thetechnologies which were used to save the palaeon-tologic material. The central part of the exhibitionis dominated by the skeleton of the Ouranosaurusnigeriensis as well as the sizeable skull and the restof the dermic part of the Sarcosuchus imperator,possibly the largest crocodile found to date. The in-teractive material and a big central screen for theprojection of footage relating to the expedition
permit the exhibitors to engage the public immer-sively in the history of the Gadoufaoua deposit andthe discovery of the remains of dinosaurs, croco-diles, turtles, fish and shellfish, as well as vegetablefinds which are now exhibited inside the showcasesthat complete the exhibition area. Therefore, notonly the public, particularly young visitors, can ad-mire the richness of the exhibited material, but theycan also experience the main moments of the firstexpedition with Italian participants dedicated tothe research and the study of dinosaurs.The exhibition “From Meteorites to Dinosaurs…
to Men” is the result of a collaboration between theLigabue Study Research Centre and the seven townsof Primiero. It is currently hosted in two rooms ofthe 14th century Palazzo delle Miniere of Fiera diPrimiero, already the venue of an ethnographic mu-seum. The exhibited findings represent a part ofthose which have been gathered during the activi-ties of the Research Centre. The materials comefrom different continents, in addition to various ge-ological eras and historical periods. Their acquisi-tion by the region of Fiera di Primiero representedthe origin of a small but active scientific museum,which is clearly separated from similar initiatives inthe area, mainly centred on materials of local ori-gin. Therefore, a private collection became a publicheritage and an instrument for the developmentand the promotion of scientific culture.The exhibition includes some fragments of mete-
orites, which document the origin and the firstphases of the solar system; various fossilised re-mains of different organisms; and two manufac-tured exhibits – a female statuette of Olmecan ori-gin and a fragment of cuneiform writing – which
128 Geo.Alp, Vol. 2, 2005
Fig.1: The ceremony of the new exhibition area at the VeniceMuseum of Natural History.
Fig. 2: An example of the interactive material in the Venetianshow room.

constitute evidence of ancient human civilizations.The main theme of the exhibition as a whole is thestate of transformation pervading Nature and thepossibility of reconstructing the subsequent phasesof Natural History through the analysis and the in-terpretation of documents. Man is, at the sametime, both the spectator and interested party ofNatural History; he is the result of biological evolu-tion as well as of the cultural evolution whichemerges in various terms and conditions.In order to organise the exhibition area, it was
necessary to start with the chronological sequenceof the finds, but we tried to avoid suggesting theidea of a “project” which – according to some peo-ple – could act as a background to the evolution ofliving organisms, a progression from initial simplici-ty towards the ultimate improvement of the organ-isms. On the contrary, we emphasised the synchron-ic aspects of evolution, classifying contemporaryevents on parallel levels of the exhibition. For ex-ample, in the showcase dedicated to invertebrates,we tried to show the evolutionary potential of “lifewithout vertebrates” and underline the structuralcomplexity which has been present since the veryfirst moments of the Cambrian explosion of life. Incontrast, the evolution of vertebrates is not seen asa progression towards the colonisation of the emer-gent lands, but as a contemporary development ofdifferent evolutionary lines, that allowed dinosaurs– widely present in the two exhibition rooms – toadapt to a great variety of habitats in the Mesozoicera. In that period there also appeared the firstmammals and birds: in the exhibition, the latter arerepresented by the rare specimen of Cathayornis.The same period ended with the decline of largereptiles and the subsequent ascendancy of mam-mals. However, the link between the skull of theCynodont, a small mammal of the Triassic period,and that of the Miocenic Machairodus giganteus,the extraordinary sabre-toothed tiger which is thesymbol of the current exhibition, is not at all linear.Similarly, the subsequent appearance of Man doesnot seem automatic. However, it is only in the lightof Darwin’s Theory of Evolution that such phenom-ena find their explanation, even though the newdiscoveries, which came consecutively in the pasthundred and fifty years that separate us from thepublication of The Origin of Species, led to an over-all revision of Darwin’s original idea. Unfortunately,the confirmation of the exact development of evo-lutionary processes will never come from a labora-
tory test, which is conventional for the empiricalsciences. On the contrary, evolutionist biology,which adopts the method of historical sciences, be-came the research of those biological traces thatmark the different phases of the history of livingorganisms. In this paradigm, palaeontology remainsa field full of potentials, which could provide solu-tions to some problems concerning the origin andthe extinction of the species. Therefore, fossils con-tain proof of the transformations that constitutethe history of life on this planet, petrified remainsof organisms that lived in a remote past, and whichnow, from a show-glass in a small mountain muse-um, continue to educate us in the “grandeur in thisview of life”.
129Geo.Alp, Vol. 2, 2005
Fig. 3: The Miocenic Machairodus giganteus, the sabre-toothedtiger, symbol of the exhibition “From Meteorites to Dinosaurus… to Men”.
Fig. 4: The characteristic skull of Psittacosaurus mongoliensis, aCretaceous dinosaur, the latest acquisition of the exhibition ofthe Primiero.

Geo.Alp, Vol. 2, S. 131, 2005
TTHHEE OORRIIGGIINN OOFF TTHHEE PPAALLAAEEOONNTTOOLLOOGGIICCAALL FFOOSSSSIILL CCOONNCCEEPPTT
Nicola Dall’Olio
131
Provincia di Parma, Piazzale Barezzi 3, 43100 Parma; e-mail: n.dall’[email protected]
In the history of science, the interpretation offossils as petrified remains of living organisms wasa first decisive step towards both the developmentof a dynamic and evolutional conception of geo-logical and biological forms, and the adoption of atemporal perspective on a scale of billions of years.In line with an underlying radicalism particularlywidespread within the scientific community, thecurrent definition of the fossil, and the relatedattribution of an organic origin to a particular classof stone objects, are usually seen as assumptionsthat arose almost automatically when, in the mod-ern age, natural scientists set aside their religiousdogma and metaphysical speculation and began tocarefully observe the world around them with anopen and objective mind, in an attempt to work out„how things really stood“. Today, the ease andimmediacy with which we recognise the vestiges ofwhat was once a living thing in a spiral object setin rock, lead us to conclude that a careful andobjective observation, free from prejudice or pre-conceived ideas based on mere speculation, isenough to determine the organic origin of fossils(or at least most of them) and to clearly distinguishthem from other mineral stone objects.
In light of a historical examination of fossil the-ories developed in Europe between 1500 and 1600,this intuitive and simplified conception of the originof palaeontology would appear to be incorrect andunfounded. Although the recognition of a researchmethod based on the careful observation of thenatural world was fundamental in achieving thesystem of classification shared today, this neverthe-less appears to be insufficient from a historical pointof view. That which is considered an almost logicalconsequence of the adoption of an objective pointof view, would appear rather as the result of assum-ing a vast combination of theories on nature andthe workings of the physical world which act as fil-ters and classifiers of the object being examined.When the problem of the classification of fossils is
limited to the recognition or denial of their organicorigin through observation, distinctions, classifica-tions and the dividing lines between natural worldsand beings (such as that between organic and inor-ganic) are taken for granted. These factors are theresult of a complex theoretical scheme, indeed onlya few centuries ago they neither existed nor couldthey even be outlined. In the absence of these divid-ing lines, the term fossil, coined by Georg Bauer,better known as Georgius Agricola in the 16th cen-tury, was simply used to describe any object in rockextracted from the subsoil.
The poster, with the help of some illustrationsfrom that era, aims to represent the decisive episte-mological change which, at the beginning of the17th century, enabled us to conceive the world ofmineral „things“ as distinct from that of organic„things“, thus providing the essential bases for theformulation of a more restrictive palaeontologicalconcept of fossils.
RReeffeerreenncceess::
Aldrovandi, U. (1648): Museum Metallicum; Ferroni eBernia ed., Bologna.
Dall’Olio, N. (2004): Vedere il Tempo. Fossili e strati nellaScienza tra 1600 e 1700. _ MUP ed., Parma.
Morello, N. (1979): La nascita della paleontologia nelSeicento: Colonna, Stenone, Scilla. – Franco Angeli,Milano
Rossi P. (1979): I segni deI Tempo. Storia della Terra e sto-ria delle nazioni da Hooke a Vico. – Feltrinelli, Milano
Rudwick, M.J.S. (1976): The meaning of fossils. Episodesin the History of Paleontology. – The University ofChicago Press, Chicago & London
Stenone, N. (1667) Canis Carchariae dissecturn caput[Trad. it. a cura di N. Morello in Morello, 1979, op. cit.]
Stenone, N. (1669) De solido intra solidum naturalitercontento dissertationis prodromus. – Trad. it. A cura diA. Mottana, Teknos Ed., Roma 1995.

Die neue Veröffentlichungsreihe „Gredleriana" des Naturmuseums Südtirol (Bozen) ist ein Forum für naturwissenschaftliche Forschung in und über Südtirol. Sie stellt eine Kommunikationsplattform dar für alle jene, die in Südtirol forschen oder in der FerneSüdtirol und den alpinen Raum als Ziel ihrer naturwissenschaftlichen Forschung haben.Band 4: 430 Seiten mit Schwerpunkt „Lebensraum Etsch“; 25 EuroAbonnement (1 Band jährlich): 20 Euro
