fundamentos físicos de los procesos biológicos vol. 3 ... · iii introducción este libro,...
TRANSCRIPT

Fundamentos físicos de los procesos biológicos
Vol. 3 - Bioelectromagnetismo, ondas y radiación
Raúl Villar LázaroCayetano López Martínez
Fernando Cussó Pérez

Fundamentos físicos de los procesos biológicos
© Fernando Cussó Pérez Cayetano López Martínez Raúl Villar Lázaro
Fig. Cubierta: Células HeLa teñidas con Hoechst, observándose núcleos en interfase y una metafase. (Cortesía de Magdalena Cañete, Departamento de Biología, UAM)
ISBN: 978-84-15787-95-2Depósito legal: A 477-2013
Edita: Editorial Club Universitario. Telf.: 96 567 61 33C/ Decano, 4 – 03690 San Vicente (Alicante)[email protected]
Printed in SpainImprime: Imprenta Gamma. Telf.: 96 567 19 87C/ Cottolengo, 25 – 03690 San Vicente (Alicante)[email protected]
Reservados todos los derechos. Ni la totalidad ni parte de este libro puede reproducirse o transmitirse por ningún procedimiento electrónico o mecánico, incluyendo fotocopia, grabación magnética o cualquier almacenamiento de información o sistema de reproducción, sin permiso previo y por escrito de los titulares del Copyright.

iii
IntroducciónEste libro, Fundamentos físicos de los procesos biológicos, se ha conce-
bido como una ayuda para los estudiantes de los primeros cursos de titula-ciones cuyo objeto de estudio principal sean las Ciencias de la Vida, como la Biología, la Medicina, la Farmacia, la Veterinaria y otras, en las que sea necesario manejar algunas nociones básicas de la Física. No se supondrá, por lo tanto, la existencia de ningún conocimiento previo en esta materia, circuns-tancia frecuente en quienes inician estudios en Ciencias de la Vida. Lo único que se presupone es el conocimiento de las matemáticas elementales corres-pondientes al nivel de bachillerato. De todas formas, se recuerdan algunos conceptos y fórmulas importantes en el texto y en los apéndices.
El contenido del libro resulta de una modificación de un libro anterior, escrito por los mismos autores, llamado Física de los procesos biológicos y publicado en 2004 por Ariel. La modificación principal consiste en la elimi-nación de las partes más generales de la Física y un enfoque más centrado en las aplicaciones al mundo de la Biología, lo que se ha traducido en una reduc-ción neta de contenidos y una más estricta selección de estos en lo que afecta a la explicación de ciertos fenómenos biológicos. Una gran parte del texto que ahora presentamos es similar al anterior, aunque se ha cambiado el contexto en que se introducen algunas nociones de Física con objeto de hacerlas más directamente aplicables a las Ciencias de la Vida. Al mismo tiempo, se han incorporado más ejercicios para mejor adaptarse a los cambios que se están produciendo en el perfil y desarrollo de los estudios universitarios.
Con todo, sigue siendo y se ha construido como un libro de Física en el que se introducen los conceptos y la metodología propios de esta ciencia, aunque se busca, en todo momento, aplicar los resultados obtenidos al ámbito de los seres vivos. El hecho, además, de que no se requieran conocimientos previos en esta materia por parte del lector implica que cada tema sea presen-tado desde su inicio, indicando lo que significa en el mundo físico en general, con aplicaciones y ejemplos, y con un énfasis especial en el ámbito de las Ciencias de la Vida, a fin de mejor entender sus leyes y familiarizar al lector con el modo de tratar y formular los problemas físicos.

iv Fundamentos físicos de los procesos biológicos
El programa, igual que el del texto anterior, cubre todos los contenidos mínimos u obligatorios de los programas de las carreras mencionadas en el primer párrafo, a desarrollar en uno o dos semestres. Pero va más allá, tanto en el desarrollo de cada tema como en el catálogo de temas, incorporando otros contenidos que serán de interés en estados más avanzados del estudio de la relación entre Física y Ciencias de la Vida. Intentamos que sirva de refe-rencia permanente para aclarar las nociones de Física que aparecen una y otra vez en determinadas ramas del estudio de los seres vivos.
Para el estudiante y el público en general, la Física y la Biología se presentan como disciplinas científicas muy distintas, incluso contradictorias o incompati-bles en sus métodos. Y es cierto que hay diferencias en su objeto de estudio, en su metodología y también en su lenguaje. La Física se ocupa normalmente de sis-temas simples que se pueden caracterizar con unos pocos parámetros, incidiendo siempre en los aspectos cuantitativos de las leyes que rigen su comportamiento, lo que implica un alto grado de formalización matemática y un método deduc-tivo a partir de unos principios generales, lo que le confiere una gran potencia predictiva. Por supuesto, se llega a esos principios generales desde la observa-ción y la experimentación de los fenómenos naturales, es decir, tienen una natu-raleza empírica, muy distinta de la de los postulados puramente matemáticos. La Biología, por su parte, se ocupa de los sistemas más complejos que existen en el mundo físico, los seres vivos, cuyo comportamiento no puede caracterizarse únicamente con ayuda de unos pocos parámetros y cuya estructura no puede entenderse fuera del marco evolutivo, de la historia concreta de la vida sobre este planeta, sujeta a contingencias impredecibles que cristalizan en las formas de vida actualmente existentes. La Biología es más descriptiva, menos deductiva, y su lenguaje no es el lenguaje altamente formalizado de las matemáticas.
Y, sin embargo, los seres vivos forman parte del mundo físico y, por lo tanto, se ven afectados por las leyes generales que rigen el comportamiento de cual-quier sistema físico, ya sea la gravedad, la tensión superficial, los intercambios de energía, la dinámica de los fluidos o las interacciones electromagnéticas. El funcionamiento de los seres vivos y de sus distintos órganos y tejidos, su estruc-tura y su relación con el entorno, están condicionados por las leyes generales de la Física, lo que constituye el nivel más básico de relación entre Física y Ciencias de la Vida. Existe otro nivel, más instrumental, que se refiere a la utilización, en los laboratorios de prácticas y de investigación, así como en otros ámbitos de la actividad profesional relacionados con las Ciencias de la Vida, de equipos e ins-trumentos basados en fenómenos físicos, o surgidos de la actividad de investi-gación en Física, que permiten observar ciertas características de los seres vivos. Ambos aspectos, básico e instrumental, están presentes en el libro.

Introducción v
Desde luego, sería absurdo pretender que entenderemos a los seres vivos si conocemos bien los fenómenos físicos en los que se basa su actividad vital. Se trata de sistemas tan complejos y con una componente “histórica” tan fuerte que, en nuestro estado actual de conocimientos, solo es posible formular en términos físicos algunos aspectos de su actividad y estructura. Pero también sería absurdo pensar que podemos entenderlos haciendo abs-tracción del mundo físico en el que viven y evolucionan y cuyas leyes les afectan. Es de gran importancia comprender que se trata de sistemas que deben ajustarse a lo estipulado por las leyes de la Física, desde las que se refieren a los átomos y moléculas que los constituyen, hasta las que regulan el comportamiento térmico y dinámico de conjuntos de muchas partículas y extrema complejidad. Las leyes de la Física plantean límites a la actividad vital que deben ser tenidos en cuenta, de forma que el juego de la evolución biológica debe discurrir dentro de esos límites. Como veremos en muchos de los casos tratados en el texto, lo difícil es plantear de forma correcta la aplicación de una ley física general al caso particular de un organismo o una función biológica.
En todo caso, la finalidad de las dos ciencias es idéntica: entender e inter-pretar los fenómenos naturales en términos de hipótesis que puedan ser con-frontadas con la observación o el experimento. Aunque, teniendo en cuenta que la complejidad de los seres vivos es superior a la de cualquier sistema inanimado, la aproximación usual en la Física se enfrenta con numerosas limitaciones y requiere adaptaciones a su nuevo objeto de estudio. Lo que no obsta para que, cuando se dilucida el fenómeno físico que subyace a una deter-minada función vital, se entienda esta de forma clara, así como el espectro de alternativas posibles permitidas por las leyes de la Física y la lógica del proceso evolutivo que ha conducido a esa solución específica.
A los autores, científicos cuya disciplina de origen es la Física, nos resulta especialmente grato dejar constancia de que, durante nuestra experiencia docente en la materia y la preparación de este texto, hemos adquirido con-ciencia de las contribuciones que la Física, a lo largo de su historia, debe a numerosos biólogos, fisiólogos y médicos, especialmente en el desarrollo de las teorías termodinámicas y la física de fluidos.
Como se ha indicado anteriormente, el presente texto resulta de la modi-ficación de un libro anterior. Afrontar una revisión y reelaboración de una obra con objetivos similares sólo es concebible si se ha recibido la justa com-binación de elogios y críticas relativas al primer texto que nos ha permitido concluir que el esfuerzo merece la pena. Así que, además de los agradeci-mientos que figuran en esa primera obra a las personas que nos ayudaron en

vi Fundamentos físicos de los procesos biológicos
la tarea de componerla, los autores nos sentimos en la obligación de añadir aquí nuestro agradecimiento a los muchos colegas y amigos que han tenido la generosidad de hacernos partícipes de sus opiniones y sugerencias de modi-ficación, lo que nos ha ayudado a identificar tanto aquellos contenidos que debían cambiarse como aquellos merecedores de mantenerse en este nuevo empeño. En particular, José Manuel Calleja, Juan Rojo y Elena Escudero, quienes han realizado una detallada lectura de la obra y cuyos comentarios han resultado especialmente esclarecedores. Sin olvidar a Carmen Gilabert y Álvaro Moreno, que han corrido con la siempre peliaguda tarea de crear las ilustraciones que ayudan a mejor entender el texto. Huelga decir que los res-ponsables de posibles deficiencias somos únicamente los autores. Por último, deseamos expresar nuestro agradecimiento a los estudiantes de Física del grado en Biología de la Universidad Autónoma de Madrid, que son respon-sables de muchos de los elogios y críticas a los que nos hemos referido, y que han constituido la principal referencia para juzgar la adecuación de nuestra descripción de los fundamentos físicos de los procesos biológicos.

vii
AgradecimientosMuchas son las personas que nos han ayudado en la tarea de componer
este libro. Proponiéndonos temas o enfoques o leyendo capítulos o fragmentos y mejorándolos con sus sugerencias, como Magdalena Cañete, Ángeles Villanueva, Elena Escudero, José Manuel Calleja, Arkadi Levanyuk, Sebastián Vieira, Francisco Jaque, Gines Lifante y Cristina Bernis. O proporcionándonos fotografías, ilustraciones o referencias con las que completar el texto, como M. C. y A. V., ya citadas, Estrella Rausell, Rodolfo Miranda, Jorgina Satrustegui, Eduardo Elizalde, Tomás López, Isabel Jimeno, Nicolás Fernández Baillo, Luis Munuera, José María Segura y José Manuel Suárez de Parga, así como el Servicio Interdepartamental de Investigación de la Universidad Autónoma de Madrid. Mención aparte merecen las personas que se han responsabilizado de las ilustraciones, una notable aportación sin la que el libro perdería una gran parte de su interés, sobre todo Carmen Gilabert, que ha debido plasmar grá-ficamente muchas de las ideas, y hasta los caprichos, de los autores, y Álvaro Moreno, así como E. E., ya citado, Carmen Morant y Eugenio Cantelar. Por último, es necesario agradecer a las numerosas promociones de alumnos de Biología que han debido cursar con nosotros la asignatura Física de los Procesos Biológicos en la UAM, por su paciencia, por sus sugerencias y por permitirnos hacernos una idea de cómo abordar esta disciplina. En realidad, hemos aprendido al mismo tiempo que ellos.


ix
Índice general abreviadoVolumen I. Biomecánica y leyes de escala
Capítulo 1. Cinemática. La percepción de la aceleraciónCapítulo 2. Dinámica. La fuerza en los animalesCapítulo 3. Fuerzas elementales y derivadas. Entornos con fricciónCapítulo 4. Bioestática y locomociónCapítulo 5. Trabajo y energía. La tasa metabólicaCapítulo 6. Leyes de escala en los seres vivosCapítulo 7. El metabolismo y las leyes de escalaApéndices A-F
Volumen II. Calor y dinámica de fluidos en los seres vivosCapítulo 8. Temperatura, calor y trabajoCapítulo 9. El primer principio: balance energético de los seres vivosCapítulo 10. Procesos espontáneos: segundo principio de la termodinámicaCapítulo 11. Transiciones de fase: la transpiración en los seres vivosCapítulo 12. Teoría cinética molecular de los gases: la difusiónCapítulo 13. Fluidos ideales: la flotaciónCapítulo 14. El flujo viscoso: la circulación sanguínea y el vueloCapítulo 15. Fenómenos de superficie y disolucionesCapítulo 16. Propiedades mecánicas de los biomateriales
Volumen III. Bioelectromagnetismo, ondas y radiaciónCapítulo 17. Fuerzas, campos y potenciales eléctricosCapítulo 18. Corriente eléctricaCapítulo 19. La membrana celular y el impulso nerviosoCapítulo 20. Magnetismo y campos electromagnéticosCapítulo 21. Ondas: luz y sonidoCapítulo 22. La visión y los instrumentos ópticosCapítulo 23. Átomos, núcleos y radiactividad


xi
Índice Volumen III. Bioelectromagnetismo, ondas y
radiaciónIntroducción ................................................................................................ iiiAgradecimientos ......................................................................................... viiÍndices .......................................................................................................... ix
Capítulo 17. Fuerzas, campos y potenciales eléctricos ............................. 117.1 Carga eléctrica y estructura de la materia ........................................ 117.2 Fuerzas eléctricas: ley de Coulomb .................................................. 317.3 Campo eléctrico ................................................................................ 517.4 Campos eléctricos en la materia. Polarización ............................... 1217.5 Interacción electrostática y energía ................................................ 1617.6 Propiedades eléctricas de la membrana celular .............................. 2517.7 Ejercicios propuestos ...................................................................... 3017.8 Referencias ..................................................................................... 37
Capítulo 18. Corriente eléctrica ................................................................ 3918.1 Movimiento de cargas: cargas libres, conductores y aislantes ....... 3918.2 Intensidad de corriente eléctrica ..................................................... 4218.3 Resistencia. Ley de Ohm ................................................................ 4318.4 Aspectos energéticos. FEM y circuitos .......................................... 4718.5 Circuitos simples ............................................................................ 5018.6 Circuitos con resistencia y capacidad (RC) .................................... 5418.7 Apéndice: pilas y baterías. Volta y Galvani .................................... 6418.8 Ejercicios propuestos ...................................................................... 6618.9 Referencias ..................................................................................... 70
Capítulo 19. La membrana celular y el impulso nervioso ...................... 7119.1 Biopotenciales ................................................................................ 71

xii
19.2 Conducción nerviosa ...................................................................... 9219.3 Apéndice 1: modelo de Hodgkin-Huxley del potencial de acción .. 13019.4 Apéndice 2: bases físicas de la electrocardiografía ...................... 13419.5 Ejercicios propuestos .................................................................... 14919.6 Referencias ................................................................................... 152
Capítulo 20. Magnetismo y campos electromagnéticos ........................ 15320.1 Introducción ................................................................................. 15320.2 Fuerza magnética sobre cargas en movimiento ............................ 15420.3 Campos magnéticos creados por corrientes ................................. 16720.4 Fuerzas entre corrientes: definición de amperio ........................... 17220.5 El magnetismo en la materia ........................................................ 17420.6 Biomagnetismo............................................................................. 17720.7 Campos electromagnéticos dependientes del tiempo ................... 18620.8 Ondas electromagnéticas .............................................................. 19320.9 Ejercicios propuestos .................................................................... 19520.10 Referencias ................................................................................. 203
Capítulo 21. Ondas: luz y sonido ............................................................ 20521.1 Introducción: propagación ............................................................ 20521.2 Tipos de ondas .............................................................................. 20621.3 Ondas: características generales ................................................... 20821.4 Luz y sonido ................................................................................. 22521.5 Fronteras: reflexión y refracción .................................................. 24921.6 Interferencia ................................................................................. 26421.7 Difracción ..................................................................................... 27021.8 Efecto Doppler ............................................................................. 28421.9 Apéndice: el oído humano ............................................................ 28721.10 Ejercicios propuestos .................................................................. 29321.11 Referencias ................................................................................. 298
Capítulo 22. La visión y los instrumentos ópticos ................................. 30122.1 Introducción: el ojo y la visión en los animales ........................... 30122.2 Imágenes sin lentes: la cámara oscura y el ojo del Nautilius ....... 30322.3 Uso de la refracción. El ojo con lente .......................................... 30422.4 Instrumentos ópticos .................................................................... 32022.5 Ejercicios propuestos .................................................................... 33922.6 Referencias ................................................................................... 342

xiii
Capítulo 23. Átomos, núcleos y radiactividad ....................................... 34523.1 Los fundamentos de la Física Cuántica ........................................ 34523.2 El núcleo atómico ......................................................................... 36923.3 Radiactividad ................................................................................ 37923.4 Efectos biológicos de la radiación ionizante ................................ 38723.5 Apéndice. La cadena de desintegración del U238 .......................... 40623.6 Ejercicios propuestos .................................................................... 40723.7 Referencias ....................................................................................411
Glosario ..................................................................................................... 413


1
Capítulo 17. Fuerzas, campos y potenciales eléctricos
17.1 Carga eléctrica y estructura de la materiaHoy día estamos familiarizados con el hecho de que la materia está for-
mada por átomos, que a su vez están formados por partículas cargadas. Cada átomo posee un núcleo que contiene protones con carga positiva y neutrones de carga nula. El número de protones en el núcleo es el número atómico Z y es ca racterístico del elemento de que se trate. Rodeando al núcleo existe un número igual de electrones, con carga negativa. La carga de los protones y electrones son exactamente iguales pero opuestas en signo (+e y –e), por lo que los átomos son eléctricamente neutros. La carga e constituye la unidad fundamental de carga, de forma que todas las cargas de la naturaleza se pre-sentan como múltiplos de dicha cantidad.1
En la casi totalidad de las manifestaciones de fenómenos eléctricos a escala macroscópica, incluyendo la mayoría de los fenómenos celulares, intervie nen siempre un número elevado de cargas (Q = Ne, con N un número entero muy grande), por lo que la cuantificación de la carga eléctrica no se observa normalmente.
Aunque la descripción rigurosa del átomo requiere del uso de la Mecánica Cuántica, como se verá en el capítulo 23, resulta útil en muchos casos acudir a un modelo atómico simplificado, de tipo “planetario”, donde los electrones de carga negativa dan vueltas en torno al núcleo de carga positiva, constituido por los protones y neutrones (fig. 17.1).
1 A escala subatómica se supone que los “hadrones” (partículas que interaccionan por medio de la fuerza nuclear fuerte, entre ellos los protones y neutrones) están formadas por “quarks”, con carga fraccionaria ± 1/3e o ± 2/3e. Sin embargo, no se ha detectado ningún quark libre y, en el marco de las teorías en vigor, sólo pueden existir combinados formando partículas de carga múltiplo de la fundamental e.

2 Fundamentos físicos de los procesos biológicos
Fig. 17.1. Modelo atómico planetario. Los electrones (carga –e) giran en torno a un núcleo cargado positivamente de unos 10-15 m de radio. El radio de las órbitas electrónicas es
100.000 veces mayor.
El radio atómico es del orden del angstrom (1 Å = 10-10 m) mientras el radio del núcleo es del orden de tan solo 10–15 m (100.000 veces más pequeño). Por su parte, la masa del átomo se concentra en el núcleo, ya que la masa del electrón es aproximadamente 1.840 veces menor que la del protón o neutrón.
Se conocen cerca de 90 átomos estables, con número atómico desde Z = 1 (hidrógeno) hasta Z ≈ 90, estando sus propiedades sistematizadas en la tabla periódica de los elementos. En los átomos multielectrónicos, los electrones internos apantallan en gran medida la interacción eléctrica entre el núcleo y los electrones externos, que pueden ser transferidos con más o menos faci-lidad, constituyendo los electrones de valencia, responsables de la mayoría de las propiedades químicas de estos átomos.
También es posible que algún átomo (o molécula), en principio eléctrica-mente neutro, pueda captar o perder algún electrón, dando lugar a iones de signo negativo (aniones) o positivo (cationes). La facilidad para este proceso está determinada por la energía de ligadura de los electrones y depende de la estructura de la “corteza electrónica” de cada átomo. En particular, muchas

Fuerzas, campos y potenciales eléctricos 3
moléculas eléctricamente neutras se disocian formando iones al incorpo-rarse a una disolución, propiedad de gran importancia para los fenómenos biológicos, que se desarrollan básicamente en medio acuoso. Por ejemplo, el cloruro sódico (NaCl) en disolución acuosa se disociaría dando lugar al catión Na+ y al anión Cl‒:
NaCl → Na+ + Cl‒
Mientras en la mayoría de las aplicaciones macroscópicas de la electri-cidad (metales, conductores) lo que se mueve son electrones, en los seres vivos los fenómenos de tipo eléctrico están ligados a la presencia de dife-rentes iones (Na+, K+, Ca2+, Cl‒ …) que se encuentran presentes en los fluidos orgánicos.
17.2 Fuerzas eléctricas: ley de CoulombLa interacción entre cuerpos cargados fue estudiada en el siglo XVIII por
Charles Augustin de Coulomb (1736-1808). La ley de Coulomb expresa el valor de las fuerzas ejercidas entre sí por dos cargas puntuales.
Sean dos cargas puntuales, q1 y q2, separadas una cierta distancia r (fig. 17.2). Las fuerzas que las cargas ejercen entre sí son proporcionales al pro-ducto de las cargas e inversamente proporcionales a la distancia entre ellas, estando dirigidas según la línea que une ambas cargas. Si las dos cargas tienen el mismo signo la fuerza es repulsiva, y si tienen signos distintos la fuerza es atractiva. En forma vectorial, la fuerza (
F12 ) que la carga q1 ejerce sobre la carga q2 puede expresarse de la forma:
F K q qr
ue r121 2
2= (17.1)
donde u r rr = es un vector unitario en la dirección de q1 a q2 y Ke es una cierta constante que depende del sistema de unidades. A su vez, de acuerdo con la tercera ley de Newton, la fuerza que la carga q2 ejerce sobre la carga q1 es: .

4 Fundamentos físicos de los procesos biológicos
Fig. 17.2. La fuerza entre dos cargas puntuales tiene un módulo Keq1q2/r2 y está dirigida
según la línea de unión entre las cargas. La fuerza es repulsiva cuando las cargas son de igual signo y atractiva si son de signo opuesto (ley de Coulomb).
En el SI de unidades, la unidad de carga es el culombio (C) y la cons-tante Ke, determinada experimentalmente, es muy aproximadamente:
K N m Ce ≈ × −9 109 2 2
En ocasiones la constante de Coulomb, Ke, se expresa en función de otra constante denominada permitividad dieléctrica del vacío (ε0).
Ke =1
4 0πε
El valor de la permitividad dieléctrica del vacío en unidades SI es, por tanto:
επ π0 9
12 2 1 214
14 9 10
8 85 10= ≈× ×
= × − − −
KC N m
e
,
La ley de Coulomb y la ley de Newton de atracción gravitatoria son nota-blemente semejantes. La magnitud de las fuerzas eléctricas o gravitatorias es proporcional al producto de los valores de las cargas o masas respectivamente, y ambas son inversamente proporcionales al cuadrado de la distancia entre las cargas o masas. Se diferencian, claro está, en que la interacción gravitatoria es siempre atractiva mientras la interacción eléctrica puede dar lugar a atracción o repulsión dependiendo del signo de las cargas.
Observación. Sistema Internacional de Unidades. En realidad, es la unidad de corriente eléctrica (el amperio, A) la que se adopta como unidad fundamental en el SI. El culombio se define a partir de ella como la cantidad de carga que fluye por dicha corriente en cada segundo 1 1 1C A s= ×( ) .

Fuerzas, campos y potenciales eléctricos 5
El valor de la carga fundamental es:
e C= × −1 60 10 19,
por lo que hacen falta del orden de 1019 electrones para conseguir un culombio de carga.
Esto significa también que el culombio es una carga muy grande. En efecto, si calculamos cuál sería la fuerza entre dos cargas de 1 C separadas una distancia de 1 m, aplicando directamente la ley de Coulomb, ec. (17.1), se obtiene:
F N= × × = ×9 10 11
9 1092
9
Esta fuerza es aproximadamente igual al peso de 1 millón de toneladas (109 kg), lo que equivale aproximadamente a diez petroleros cargados o unas tres veces el Empire State Building o unas cinco veces el iceberg con que chocó el Titanic o la sexta parte de la pirámide de Keops.
17.3 Campo eléctrico
17.3.1 Campo eléctrico creado por una carga puntualLa descripción de las fuerzas que actúan sobre una partícula cargada puede
hacerse también utilizando el concepto de campo eléctrico.
El lenguaje es análogo al utilizado al describir la interacción gravitatoria. Cuando un satélite se mueve en el espacio bajo la atracción terrestre decimos que se encuentra en el campo gravitatorio terrestre. Análogamente, cuando una carga se encuentra en una región del espacio donde le afectan otras cargas, se dice que se encuentra en el campo eléctrico que han creado dichas cargas. Es decir, se adopta el punto de vista de considerar que las cargas crean en su entorno una perturbación (un campo eléctrico) y este campo actúa sobre cualquier carga que se encuentre allí, dando lugar a la fuerza descrita por la ley de Coulomb (fig. 17.3).

6 Fundamentos físicos de los procesos biológicos
Fig. 17.3. La carga Q crea en su entorno un campo eléctrico (r), y la fuerza eléctrica sobre una carga q es igual al producto de dicha carga por el campo eléctrico.
La intensidad de campo eléctrico en un punto se define como la fuerza por unidad de carga colocada en ese punto.
E Fq
= (17.2)
La unidad de campo eléctrico en el SI es la unidad de fuerza dividida por la unidad de carga, es decir, el N C‒1.
El campo eléctrico creado por una carga puntual se calcula inmediata-mente a partir de la ley de Coulomb:
E Fq
K Qr
ue r= = 2 (17.3)
donde Q es la carga que crea el campo eléctrico en un punto situado a una distancia r (fig. 17.3). La dirección del campo viene dada por el vector unitario u r rr = y el sentido del campo depende del signo de la carga: hacia fuera si ésta es positiva y hacia dentro si es negativa.
La existencia del campo eléctrico se pondría de manifiesto al situar una carga de prueba, q, en dicho punto, ya que la carga experimentará, de acuerdo con la ecuación (17.2), una fuerza:
F q E=

Fuerzas, campos y potenciales eléctricos 7
Cuando el campo eléctrico es debido a la presencia de varias cargas Q1, Q2, ..., QN (fig. 17.4), su valor es el resultante de la suma vectorial del campo creado por cada una de ellas:
E Etot i
N
= ∑1
y la fuerza total sobre una carga q puede calcularse a partir del campo total:
F qE q E qE Ftot tot i
N
i
N
i
N
= =
= ( ) =∑ ∑ ∑
1 1 1
Fig. 17.4. Cuando una carga q interacciona con varias cargas Qi, la fuerza resultante es la suma de las fuerzas de cada una de las cargas (a) o, alternativamente, se puede calcular como el producto de la carga por el campo resultante, igual a la suma de todos los campos
eléctricos (b).
Para visualizar el campo eléctrico creado por una distribución cualquiera de cargas, resulta conveniente utilizar una representación basada en la uti-lización de las líneas de campo eléctrico. Dicha representación consiste en dibujar mediante líneas continuas la dirección del campo eléctrico resultante que, a veces, reciben también el nombre de líneas de fuerza, ya que indican la dirección de la fuerza que el campo ejercería sobre una carga positiva.
La figura 17.5 representa las líneas de campo eléctrico en presencia de cargas puntuales positiva (a) y negativa (b).

8 Fundamentos físicos de los procesos biológicos
Fig. 17.5. Líneas de campo eléctrico en presencia de una carga puntual: (a) positiva y (b) negativa.
Como indica la ecuación (17.3), las líneas de campo son radiales y se dirigen (a) hacia fuera en el caso de campo creado por una carga positiva y (b) hacia la carga si el campo es creado por una carga negativa. Al alejarnos de la carga el campo disminuye y, como vemos en la figura, las líneas de campo están más separadas. Análogamente, en distribuciones de carga más complejas, el número de líneas representado en cada región del espacio indica la intensidad del campo.
Las líneas de campo pueden visualizarse experimentalmente utilizando pequeños objetos alargados (semillas, trocitos de hilo...) que tienden a orien-tarse según la dirección del campo.
Los diagramas de líneas de campo eléctrico nos indican también la direc-ción de las fuerzas que actúan sobre las cargas, si bien hay que recordar que el sentido de la fuerza viene determinado por el signo de la carga (
F q E= ): las cargas positivas se mueven en la dirección del campo, mientras las cargas negativas lo hacen en dirección opuesta al campo.
17.3.2 Algunas distribuciones de carga i) Dipolo eléctrico
Aunque la materia es neutra, en general, ya que tiene el mismo número de cargas positivas que negativas, la distribución de las mismas no es siempre simétrica, por lo que en muchas moléculas el centro de las cargas positivas y negativas está desplazado. Estas moléculas constituyen dipolos eléctricos y este carácter polar tiene importantes consecuencias en sus propiedades fisi-coquímicas. La forma más simple de dipolo eléctrico lo constituye un par de cargas iguales, pero de signo opuesto, +q y –q, separadas una pequeña distancia, d.
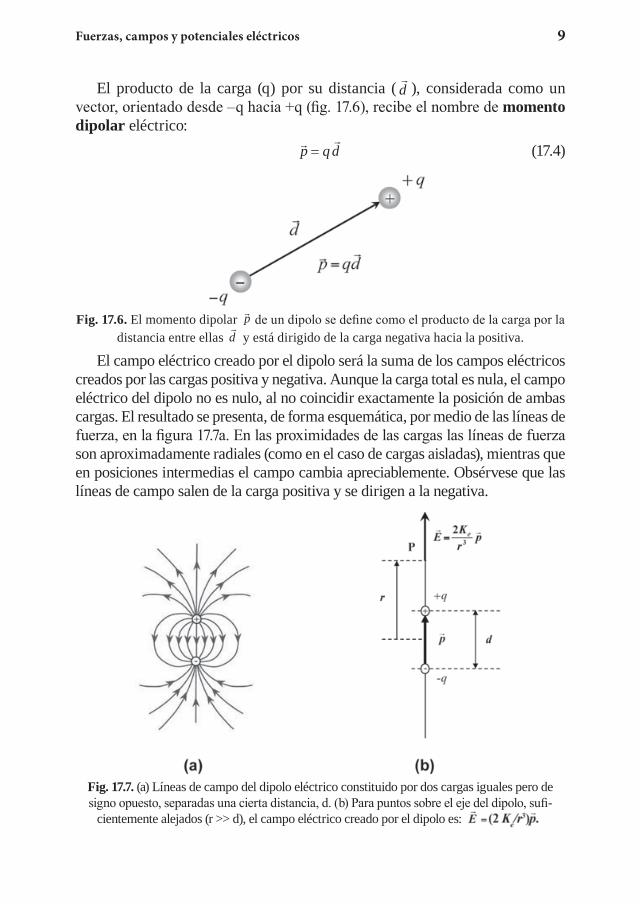
Fuerzas, campos y potenciales eléctricos 9
El producto de la carga (q) por su distancia (
d ), considerada como un vector, orientado desde –q hacia +q (fig. 17.6), recibe el nombre de momento dipolar eléctrico:
p q d= (17.4)
Fig. 17.6. El momento dipolar
p q d= de un dipolo se define como el producto de la carga por la distancia entre ellas
d y está dirigido de la carga negativa hacia la positiva.
El campo eléctrico creado por el dipolo será la suma de los campos eléctricos creados por las cargas positiva y negativa. Aunque la carga total es nula, el campo eléctrico del dipolo no es nulo, al no coincidir exactamente la posición de ambas cargas. El resultado se presenta, de forma esquemática, por medio de las líneas de fuerza, en la figura 17.7a. En las proximidades de las cargas las líneas de fuerza son aproximadamente radiales (como en el caso de cargas aisladas), mientras que en posiciones intermedias el campo cambia apreciablemente. Obsérvese que las líneas de campo salen de la carga positiva y se dirigen a la negativa.
Fig. 17.7. (a) Líneas de campo del dipolo eléctrico constituido por dos cargas iguales pero de signo opuesto, separadas una cierta distancia, d. (b) Para puntos sobre el eje del dipolo, sufi-
cientemente alejados (r >> d), el campo eléctrico creado por el dipolo es:

10 Fundamentos físicos de los procesos biológicos
Como puede verse en la figura, el campo eléctrico en un punto del eje del dipolo es paralelo a dicho eje. Es fácil calcular su valor, sumando las contribu-ciones debidas a ambas cargas. El valor del campo en un punto P, situado a una distancia r del centro del dipolo, suficientemente alejado (r >> d), puede escribirse como:
E Kr
pe=2
3 (17.5)
donde p es el momento dipolar, tal como se indica en la figura 17.7b. Nótese que el campo eléctrico creado por el dipolo decrece como el cubo de la dis-tancia (E ~ 1/r3), es decir, más rápidamente que en el caso del campo creado por una carga aislada (E ~ 1/r2).
La mayoría de las moléculas formadas por átomos distintos, aun siendo eléctricamente neutras, presentan una distribución de carga asimétrica, teniendo alguno de sus átomos un exceso de carga positiva y otros exceso de carga negativa, por lo que se comportan como dipolos eléctricos. Un ejemplo, particularmente importante, lo constituye la molécula de agua, donde el átomo de oxígeno está cargado negativamente y los átomos de hidrógeno positiva-mente, lo que dota a la molécula de agua de un marcado carácter dipolar, de gran importancia para sus propiedades fisicoquímicas.
ii) Distribuciones continuas de carga
Con frecuencia, el número de cargas que intervienen en las situaciones de interés es muy elevado, por lo que en lugar de describir individualmente cada carga, se puede considerar que ésta se distribuye continuamente en el espacio o sobre una superficie. En esos casos la carga se define a partir de la densidad de carga, bien de volumen (ρ = dQ/dV), o bien de superficie (σ = dQ/dS).
Distribución de carga con simetría esférica
El campo eléctrico creado por una distribución de carga con simetría esférica, en un punto exterior a dicha distribución, es el mismo que si toda la carga estuviese concentrada en el centro de la esfera. Este resultado es similar al que se obtiene para el campo gravitatorio, debido a la similitud entre la ley de la gravitación de Newton y la ley de Coulomb.
Planos cargados uniformemente
Es fácil comprender que el campo eléctrico creado por un plano cargado uniformemente debe ser perpendicular al plano (fig. 17.8). Al calcular el valor del campo en un punto cualquiera P, y sumar la contribución de todas las cargas del plano, por cada carga situada en un punto A existe otra igual

Fuerzas, campos y potenciales eléctricos 11
ubicada simétricamente, B, con lo que la suma de los campos creados por ambas cancela las componentes paralelas al plano y la resultante es perpen-dicular al mismo. Este razonamiento es válido si el plano es infinito o, en tér-minos más realistas, si la distancia entre el punto P y el plano es mucho menor que las dimensiones de éste, es decir, cuando P está “próximo” al plano.
Se demuestra que el campo eléctrico creado por un plano infinito cargado es:
E uN=σε2 0
(17.6)
donde σ es la densidad superficial de carga y uN es un vector unitario per-pendicular al plano. Nótese que el campo resulta ser independiente de la dis-tancia al plano y, salvo constantes, su valor depende exclusivamente de la densidad de carga eléctrica. Si el plano está cargado positivamente (+σ) el campo se dirige hacia fuera (fig. 17.8), mientras que si la carga es negativa (–σ), el campo se dirige hacia el plano.
Fig. 17.8. El campo eléctrico en las proximidades de un plano cargado uniformemente, con densidad de carga σ = Q/S, es perpendicular al plano y su módulo (E = σ/2ε0) es inde-
pendiente de la distancia al plano.
Una configuración habitual es el conjunto de dos planos cargados, con densidades de carga iguales pero de signo contrario +σ y –σ, lo que consti-tuye un condensador plano. El campo eléctrico resultante será la suma de los campos creados por cada plano, que son iguales en módulo (e independientes de la distancia a los planos) cambiando tan solo el sentido respecto de cada plano debido al signo de la carga. En consecuencia, en la región exterior a los planos la dirección de los campos es opuesta y la suma se cancela (
E =
0 ), mientras en la región comprendida entre ambos planos las direcciones coin-ciden y el campo resultante se suma, resultando un campo eléctrico uniforme en el espacio comprendido entre los planos cargados, de valor:

12 Fundamentos físicos de los procesos biológicos
E u uN N= ×
=2
2 0 0
σε
σε (17.7)
estando dirigido del plano positivo hacia el negativo (fig. 17.9).
Fig. 17.9. Campo eléctrico creado por dos planos cargados (condensador plano). Al sumar el campo creado por el plano positivo y el negativo los campos se cancelan en el exterior y se suman en el interior, resultando un campo eléctrico uniforme dirigido del plano positivo
al negativo.
17.4 Campos eléctricos en la materia. Polarización
17.4.1 Polarización inducidaHemos dicho con anterioridad que la mayoría de las moléculas pre-
sentan un carácter dipolar, debido a la falta de simetría en la distribución de carga. Este carácter dipolar lo adquieren también las moléculas e incluso los átomos no polares cuando se encuentran en un campo eléctrico (polarización inducida).
Si consideramos un átomo como un núcleo positivo rodeado por una nube electrónica, con simetría esférica en torno al núcleo, el centro de ambas cargas coincide (fig. 17.10.a). Sin embargo, en presencia de un campo eléctrico externo, el centro de ambas cargas no coincidirá: el campo ejerce una fuerza sobre el núcleo cargado positivamente en la dirección del campo, mientras la nube electrónica, cargada negativamente, experimenta una fuerza en direc-ción opuesta. Las cargas positiva y negativa son separadas hasta que la fuerza atractiva entre ellas compensa la fuerza del campo eléctrico externo. Al sepa-rase las cargas, el átomo (o molécula) se ha convertido en un dipolo eléctrico inducido por el campo externo (fig. 17.10.b).

Fuerzas, campos y potenciales eléctricos 13
Fig. 17.10. (a) En un átomo, en ausencia de campo externo, el centro de las cargas positiva y negativa coincide. (b) Un campo externo separa ambas cargas, dando lugar a un dipolo
eléctrico inducido.
Orientación de dipolos en un campo eléctrico
La materia, en presencia de un campo eléctrico externo, se comporta siempre como un conjunto de dipolos, ya sea porque se trate de moléculas polares intrínsecas (como el agua) o moléculas en las que el campo eléctrico ha inducido un cierto momento dipolar. Por tanto, resulta necesario conocer el comportamiento de un dipolo eléctrico en presencia de un campo externo. Teniendo en cuenta que en un campo eléctrico uniforme las cargas positiva y negativa del dipolo experimentan fuerzas iguales en módulo pero dirigidas en direcciones opuestas (fig. 17.11), sobre el dipolo actúa un momento que lo hace girar orientándolo en la dirección del campo ( p ║
E ).
Fig. 17.11. Un dipolo en un campo eléctrico uniforme experimenta un par de fuerzas que tienden a alinearlo paralelamente al campo.
17.4.2 Constante dieléctricaConsideremos el efecto de un campo eléctrico sobre un material no con-
ductor, también llamado dieléctrico, tal como el vidrio, plástico o madera. (Los materiales conductores y la corriente eléctrica serán estudiados en el capítulo 18). Supongamos que dicho material se sitúa en un campo eléctrico uniforme, tal como sucedería al situarlo entre las placas de un condensador plano (fig. 17.12).

14 Fundamentos físicos de los procesos biológicos
En ausencia de campo eléctrico, o bien las moléculas carecen de momento dipolar, o bien, aun teniendo un momento dipolar permanente, estarán orien-tadas aleatoriamente (fig. 17.12.a). En presencia del campo
E0 el material se polariza, ya sea generando un momento dipolar inducido, ya sea orientando los dipolos permanentes, desplazándose las cargas positivas en la dirección del campo y las negativas en dirección opuesta (fig. 17.12.b). En el interior del dieléctrico estos desplazamientos se compensan, pero en los extremos del dieléctrico aparecen sendos excesos de carga +δq y –δq. En consecuencia, el material polarizado crea a su vez un campo
Epol que, en el interior del mate-rial, lleva la dirección contraria al campo externo y el campo resultante
Etot , suma del campo externo y el debido a la polarización, es, por tanto, inferior al campo externo (fig. 17.12.c). La presencia de un dieléctrico reduce el valor del campo eléctrico.
E E E Etot pol= + <0 0
Fig. 17.12. El campo eléctrico en el interior de un medio dieléctrico se reduce por efecto de su polarización.
El factor en que se reduce el campo eléctrico depende de la facilidad de polarización de los átomos o moléculas del material, y se cuantifica por medio de la magnitud denominada constante dieléctrica (κ), definida a partir de la relación:
E E Etot = <0 0/ κ
de donde se deduce que:κ >1
Para calcular el campo eléctrico creado por una carga Q en el interior de un dieléctrico basta con sustituir el valor de la permitividad dieléctrica del

Fuerzas, campos y potenciales eléctricos 15
vacío (ε0) por la permitividad dieléctrica del material (ε), que se obtiene de multiplicar la permitividad del vacío por la constante dieléctrica.
ε κ ε= × 0 (17.8)
Así, el campo eléctrico de un condensador plano con un material dieléc-trico en su interior puede calcularse a partir de la ec. (17.7), sustituyendo la permitividad dieléctrica del vacío (ε0) por el valor de la permitividad dieléc-trica del dieléctrico (ε):
E u u K u EN N e N= =
=
( ) =
σε κ
σε κ
π σκ
1 1 4 1
00 (17.9)
Análogamente, el campo eléctrico creado por una carga puntual Q, en un medio dieléctrico, se obtendría de forma similar, a partir de la ley de Coulomb:
E Q
rQr
Qr
Etot o= = =
=
14
14
1 14
12
02
02πε π κε κ πε κ( )
(17.10)
Tal como hemos indicado anteriormente, el campo eléctrico se reduce en un factor igual a la constante dieléctrica.
En la tabla 17.1, se resumen los valores de la constante dieléctrica de algunos materiales de interés. Nótese, en particular, el alto valor de la cons-tante dieléctrica del agua (κ = 80). Esto es debido a su momento dipolar per-manente, que, por efecto de un campo eléctrico, da lugar a su orientación produciendo una polarización macroscópica elevada. Esto da lugar a una reducción de la intensidad de las interacciones eléctricas en disolución, que juega un papel muy importante en Química y Biología, ya que apantalla y aísla las cargas, facilitando la disociación de las moléculas.
Tabla 17.1. Constante dieléctrica de algunos materiales
Material Constante dieléctrica (κ)vacío 1aire 1,00059
papel ~ 3,5vidrio 5-10
plásticos 2-20aceite 2
membrana celular 7-9agua 80

16 Fundamentos físicos de los procesos biológicos
17.5 Interacción electrostática y energía17.5.1 Energía potencial eléctrica
Estamos ya familiarizados con los conceptos de trabajo y energía, que se definieron en el capítulo 5, y con las ventajas que ofrece su utilización para describir el movimiento bajo la acción de fuerzas conservativas. Y las fuerzas electrostáticas son conservativas.
La fuerza que experimenta una carga q, en presencia de otra carga Q, queda expresada por la ley de Coulomb
F K qQ r ue r= ( )( )2 , cuya forma es totalmente análoga a la ley de Newton de interacción gravitatoria
F GmM r ur= −( )( )2 , pudiendo obtenerse una de la otra sin más que intercambiar –GmM por Ke q Q.
En consecuencia, podemos aplicar a la interacción electrostática todo lo dicho en el Volumen 1 en relación con el trabajo y energía de las fuerzas gravitatorias. En particular, podemos asegurar que las fuerzas entre cargas eléctricas son fuerzas conservativas. Recordemos que esto significa que cuando una carga q se desplaza en el campo eléctrico creado por la carga Q (fig. 17.13) desde un punto (1) a otro punto (2), el trabajo W realizado por la fuerza eléctrica, sobre la carga q, puede expresarse como la diferencia de energía potencial U entre los dos puntos. Sólo depende, por tanto, de las posi-ciones inicial y final de la carga y no del camino seguido para pasar de una a otra posición.
W Fdr U r U r= = ( ) − ( )∫
1
2
1 2 (17.11)
W Fdr U r U r= = ( ) − ( )∫
1
2
1 2
Fig. 17.13. Las fuerzas eléctricas son conservativas, por lo que el trabajo realizado sobre la carga q, desplazándose desde un punto (1) a otro (2), puede expresarse como la dife rencia
de energía potencial eléctrica entre los dos puntos.

Fuerzas, campos y potenciales eléctricos 17
La energía potencial electrostática se puede obtener de forma inmediata, a partir de la ec. (17.11) utilizando para la fuerza la ley de Coulomb, ec. (17.1), e integrando, con lo que se obtiene:
U r K qQ
re( ) = (17.12)
Recordemos que, como se vio en el capítulo 5, la energía potencial está definida salvo una constante, lo que permite elegir el origen de energías. En la expresión (17.12) se ha hecho uso de esto, tomando como origen de energía potencial electrostática la situación en que las cargas se encuentran infinita-mente alejadas:
U r( )→ ∞ = 0
La tabla 17.2 resume las analogías entre magnitudes gravitatorias y electrostáticas.
Tabla 17.2. Analogías entre magnitudes gravitatorias y electrostáticas
Magnitud Interacción gravitatoria Interacción electrostática
Fuerza L. de Newton
F GmM r ur= −( )2L. de Coulomb
F K qQ r ue r= ( )2
Magnitud característica Masa, m Carga, q
Energía potencial U G mM r= − U K qQ re=
Potencial V G M r= − V K Q re=
17.5.2 Potencial eléctricoComo hemos visto, la energía potencial de una carga q en un punto es
proporcional al valor de dicha carga, es decir, si colocamos en el mismo punto una carga doble tendrá el doble de energía y una carga triple adquirirá un valor triple en su energía.
Por ello resulta útil separar este efecto de aquellos otros que dependen exclusivamente del campo (de la carga que lo crea y de la posición), utilizando para ello el concepto de potencial eléctrico (V) o energía potencial por unidad de carga.
V r
U rq
( ) =( ) (17.13)

18 Fundamentos físicos de los procesos biológicos
El valor del potencial a una distancia r de una carga puntual Q se obtiene de forma inmediata a partir de la ecuación (17.13):
V r K Q
re( ) = (17.14)
y cuando hay varias cargas basta con sumar el potencial creado por cada una de ellas:
V r V r K Qrtotal i
N
ei
i
N
( ) ( )= =∑ ∑1 1
siendo ri la distancia de cada carga al punto donde se calcula el potencial.
Conociendo el potencial V(r) se puede determinar inmediatamente (ec. 17.13) la energía potencial U(r) que adquiere una carga cualquiera q en ese punto:
U r qV r( ) = ( )
El trabajo realizado por las fuerzas eléctricas, cuando una carga q se desplaza entre los puntos r1 y r2, vendrá dado por la diferencia de energía potencial entre ellos, que también se puede expresar en función de la diferencia de potencial:
W Fdr U r U r q V r V r= = ( ) − ( ) = ( ) − ( )( )∫
1
2
1 2 1 2 (17.15)
Ecuación que podemos reformular en términos del campo eléctrico (
E F q= / ):
Edr V r V r1
2
1 2∫ = ( ) − ( ) (17.16)
Si consideramos una región del espacio donde haya un campo eléctrico uniforme, como entre las placas de un condensador plano, por ejemplo, y nos desplazamos una distancia ∆
r en la dirección del campo, podemos integrar inmediatamente la ecuación anterior obteniendo
E dr E r r V r V r V r V r1
2
2 1 1 2 2 1∫ = −( ) = ( ) − ( ) = − ( ) − ( )
es decir:
E r V∆ ∆= − (17.17a)
o bien:

Fuerzas, campos y potenciales eléctricos 19
E V
r= −
∆∆
17.17b)
donde el signo – indica que el campo eléctrico está dirigido hacia potenciales decrecientes (fig. 17.14).
Fig. 17.14. El campo eléctrico está dirigido siempre hacia potenciales decrecientes.
La expresión (17.16) indica que, al igual que sucede con la energía poten-cial, son las diferencias de potencial (V(r1) – V(r2)) las que tienen sentido físico, pudiendo elegirse un origen de potenciales arbitrario. En el caso de cargas puntuales (ec. 17.14), se toma el infinito como cero de potenciales, pero no siempre se hace así. Por ejemplo, al estudiar el potencial de membrana (capítulo 19) se adopta habitualmente como origen de potencial el valor del medio extracelular (Ve = 0).
La unidad de potencial eléctrico es el voltio (V) y corresponde, como indica la ecuación (17.13), al potencial que existe en un punto donde al situar una carga de un culombio ésta adquiere una energía potencial de un julio.
1 Voltio = 1 Julio / 1 Culombio
En el lenguaje habitual se habla de “voltaje” cuando se quiere indicar cuál es la diferencia de potencial entre dos puntos, siendo ésta la forma habitual de describir las características eléctricas en muchas ocasiones cotidianas. Así, se dice que el voltaje en un enchufe es de 220 voltios, con lo que se quiere indicar que entre los dos terminales del mismo existe esa diferencia de potencial. Si ambos se conectan mediante un conductor, la carga se mueve de uno a otro debido a la diferencia energética entre dichos puntos. Si de cimos que un dispo sitivo funciona a 1,5 voltios, queremos decir que ésa es la diferencia de potencial adecuada a la que deben conectarse los terminales correspondientes.

20 Fundamentos físicos de los procesos biológicos
Observación. Electronvoltio (eV). La descripción de los fenómenos atómicos y moleculares se fundamenta en la interacción electrostática entre protones y electrones, mientras que a nivel celular resulta necesario describir el movimiento de iones (Na+, K+, Cl–, Ca2+...). En todos estos casos se trata de describir el comportamiento de partículas con cargas cuya magnitud es del orden de la carga fundamental (e = 1,6 × 1019 C).
Cuando una carga igual a la fundamental se mueve entre puntos con una diferencia de potencial de 1 voltio, adquiere una energía que se denomina electronvoltio (eV), y cuya equivalencia en el SI es:
1 1 6 10 1 1 6 1019 19eV J= ×( )× ( ) = ×− −, ,
El electronvoltio es una unidad de energía adecuada, por tanto, para muchos procesos atómicos, moleculares y celulares, en los que se estudia el movimiento de cargas de algunos electrones en diferencias de potencial de voltios. Así, la energía del estado fundamental del átomo de hidrógeno es de 13,6 eV, la afinidad electrónica del f luor es de 3,4 eV, la energía de un fotón de luz visi ble varía entre 1,7 eV y 3,0 eV, la energía liberada en la hidrólisis del ATP es de 0,3 eV/molécula (en condiciones estándar), etc.
En otros ámbitos de aplicación, implicando órdenes de magnitud diferentes, también se expresa la energía en eV o, mejor dicho, sus múltiplos. Por ejemplo, las diferencias de potencial implicadas en los tubos de vacío como los televisores y tubos de rayos catódicos son del orden de kilovoltios (kV), por lo que las energías que alcanzan los electrones allí acelerados se expresan en kiloelectronvoltios (1 keV = 103 eV). De forma similar, las energías implicadas en procesos nucleares se indican en megaelectronvoltios (1 MeV= 106 eV).
Observación. Unidades de campo eléctrico. La unidad de campo eléctrico en el SI es el NC1, pero en la práctica se expresa corrientemente en función del potencial, a partir de la ecuación (17.17), con la equivalencia:
1 1NC
Vm
=
17.5.3 Capacidad En el caso de un condensador plano, el campo eléctrico (E) es uniforme
y perpendicular a las placas, pudiendo expresarse en función de la diferencia

Fuerzas, campos y potenciales eléctricos 21
de potencial entre las mismas (V) y su separación (d), a partir de la ecuación (17.17).
E = V/d
Pero, a su vez, podemos expresarlo en función de la carga de sus placas (ecuación 17.9):
E = =
σε κ
σε
1
0
donde σ = Q/S es la densidad superficial de carga y κ la constante dieléctrica del material existente entre las placas (si fuese el vacío o aire, κ ≈ 1).
Por tanto, igualando ambas expresiones obtenemos:
Vd
QS
= =σε ε
o bien:
Q S
dV=
ε (17.18)
Es decir, existe una proporcionalidad entre la carga Q en las placas de un condensador plano y la diferencia de potencial V entre ellas. El coeficiente de proporcionalidad entre carga y potencial recibe el nombre de capacidad C.
Q C V= (17.19)
Y, por tanto, la capacidad de un condensador plano es:
C S
dSd
= =ε
κ ε0 (17.20)
La capacidad del condensador plano depende de la geometría, concreta-mente del área de las placas S y de su separación d, así como de las propiedades dieléctricas del material entre las placas, a través de su constante dieléctrica κ.
Si almacenamos cargas en un condensador con una geometría diferente, también existe una relación de proporcionalidad entre la carga acumulada y la diferencia de potencial que se genera (ec. 17.19), donde la capacidad ten-dría otra expresión diferente de la (17.20) aunque dependerá igualmente de la geometría del condensador y de la constante dieléctrica del medio.

22 Fundamentos físicos de los procesos biológicos
La unidad de capacidad en el SI es el faradio (F), que a partir de la expresión (17.19) se define como la capacidad de un condensador cuyas placas tienen una carga de 1 culombio cuando entre sus placas hay una diferencia de potencial de 1 voltio. Es decir:
1 11
F CV
=
El faradio es una unidad muy grande, por lo que en la práctica se utilizan habitualmente sus submúltiplos, tales como el microfaradio 1 µF = 10–6 F y el picofaradio 1 pF = 10–12 F.
Conexión/asociación de condensadores
Los condensadores suelen utilizarse conectados entre sí, existiendo dos formas básicas de hacerlo: en paralelo y en serie.
Cuando dos condensadores se conectan tal como aparece en la figura 17.15 se dice que están conectados en paralelo. Las líneas continuas representan conexiones mediante conductores de resistencia despreciable, uniendo puntos que están a un mismo potencial (lo que quedará justificado en el capítulo siguiente), por lo que se puede considerar que ambos condensadores están conectados a puntos comunes A y B, es decir, con la misma diferencia de potencial entre las placas: V = VA – VB.
Si las capacidades de los condensadores son C1 y C2, las cargas Q1 y Q2 de cada uno serán:
Q C V1 1= , Q C V2 2=
La carga total es, por tanto:
Q Q Q C V C V C C V= + = + = +( )1 2 1 2 1 2
Es decir, equivale a un único condensador con capacidad equivalente (C) igual a la suma de las capacidades:
C C C= +1 2 (17.21)

Fuerzas, campos y potenciales eléctricos 23
Fig. 17.15. Asociación de condensadores en paralelo. Nótese que en los circuitos eléctricos, un condensador se representa por el símbolo , mientras que las líneas continuas repre-
sentan conexiones mediante conductores de resistencia despreciable. En consecuencia, los condensadores están conectados a la misma diferencia de potencial y tendrán cargas difer-entes, dependiendo de su capacidad. Tal como se muestra en la parte inferior de la figura,
equivalen a un único condensador de capacidad equivalente C = C1 + C2.
Cuando dos condensadores se conectan tal como aparece en la figura 17.16 se dice que están conectados en serie. Cuando se aplica una diferencia de potencial entre los extremos A y B (V = VA – VB), la carga de ambos conden-sadores es la misma (el conjunto de la segunda placa del condensador 1 y la primera placa del condensador 2 ha de ser eléctricamente neutro).
La diferencia de potencial en los condensadores es:
V V V QCA m1
1
= − = , V V V Q
Cm B22
= − =
Siendo Vm el potencial en un punto intermedio (fig. 17.16).
La diferencia de potencial entre los extremos es:
V V V V V QC
QC
QC CA B= − = + = + = +
1 2
1 2 1 2
1 1
Es decir, el conjunto equivale a un único condensador con capacidad equivalente (C), tal que
1 1 1
1 2C C C= + (17.22)

24 Fundamentos físicos de los procesos biológicos
Fig. 17.16. Asociación de condensadores en serie. Los condensadores adquieren la misma carga y la diferencia de potencial entre las placas depende de su capacidad. Equivalen a un
único condensador de capacidad 1/C = 1/C1 + 1/C2.
17.5.4 Energía del campo eléctrico Al cargar un condensador, separando las cargas positiva y negativa una
cierta distancia d, se está realizando un trabajo en contra del campo eléctrico, almacenándose energía que puede utilizarse con posterioridad para producir movimiento de cargas. ¿Cuánto vale la energía almacenada en un condensador con carga Q y diferencia de potencial V?
Teniendo en cuenta que el potencial representa la energía por unidad de carga, podría pensarse que la energía almacenada es: Energía = Q V, pero, en realidad, la energía almacenada en el condensador se genera de forma paulatina. El proceso no se hace desplazando una carga Q contra una dife-rencia de potencial V constante, sino que la diferencia de potencial cambia de forma simultánea al cambio en la carga almacenada, manteniéndose siempre la proporcionalidad q = C V entre ellas.
Para hallar la energía del condensador hay que sumar la energía almacenada por cada incremento de carga dq que se produce. Así, en un estado de carga intermedio se transfiere una carga dq a un potencial V (fig. 17.17.a), con lo que la energía potencial de la carga se incrementa en:
d Energía V dq qC
dq( ) = =
Por tanto, al cargar el condensador desde la carga q = 0 hasta la carga final q = Q, la energía acumulada es:
Energía V dq q C dq QC
Q Q= = ( ) =∫ ∫0 0
212
/

Fuerzas, campos y potenciales eléctricos 25
Y utilizando Q = C V, se obtienen expresiones alternativas:
Energía Q
CQV CV= = =
12
12
12
22 (17.23)
Lo que también se puede obtener de forma gráfica, representando el valor del potencial en función de la carga (V = q/C), y considerando que la energía ( V dq
Q
0∫ ) equivale al área del triángulo de la figura 17.17.b, es decir, energía Q Q C= ×( )1
2, que coincide con el valor obtenido mediante el cálculo
integral.
Fig. 17.17. La energía almacenada en un condensador de capacidad C con una carga Q es igual al área bajo la curva: (q/C) dq = (1/2)Q2/C = (1/2)QV.
17.6 Propiedades eléctricas de la membrana celular Toda célula está rodeada por una membrana (membrana plasmática) que
le permite mantener su identidad, aislándola del exterior. El modelo aceptado hoy en día para la membrana celular (mosaico fluido) es común para todos los organismos vivos (animales, plantas y microorganismos) y se presenta de forma simplificada en la figura 17.18. La membrana, de un espesor aproxi-mado entre 8 y 12 nm (1 nm = 10–9 m), consta de una doble capa lipídica, con los extremos hidrofóbicos hacia el interior y las cabezas hidrofílicas hacia el exterior de la membrana y en contacto con el citosol y el medio extracelular.
La membrana celular incorpora también diferentes proteínas, algunas de ellas denominadas de canal, que permiten el intercambio de sustancias entre el citosol y el exterior, lo que resulta imprescindible para el mantenimiento de la actividad celular. La permeabilidad de la membrana resulta ser muy selec-tiva, permitiendo el paso de determinadas sustancias e impidiendo el paso de otras. Esta selectividad da lugar (capítulo 19) a que la composición iónica del citosol sea notablemente diferente del medio externo, produciéndose un exceso de iones K+ y un defecto de iones Na+ y Cl– respecto a sus concentra-ciones en el medio extracelular.

26 Fundamentos físicos de los procesos biológicos
Como consecuencia de este desequilibrio iónico, aparece una carga eléc-trica neta a ambos lados de la membrana, estando el citosol cargado negativa-mente respecto al exterior, lo que da lugar a la aparición de un campo eléctrico en la membrana y una diferencia de potencial eléctrico entre el exterior y el citoplasma, denominado potencial de membrana.
Fig. 17.18. (a) La membrana celular es una fina capa (10–8 m) dieléctrica. (b) Está com-puesta por una doble capa lipídica con los extremos hidrofóbicos hacia el interior y las
cabezas hidrofílicas hacia el exterior.
Campo eléctrico y potencial en la membrana celular
La membrana se puede considerar, desde el punto de vista eléctrico, como un condensador, aproximadamente plano, ya que su espesor es mucho menor que las dimensiones celulares. Por tanto, sus propiedades quedarán descritas por su espesor d, área S y permitividad dieléctrica ε (o la constante dieléc-trica κ = ε/ε0) y el valor del campo eléctrico y del potencial de membrana quedan determinados por el valor de la densidad superficial de carga en la membrana.
Medidas eléctricas realizadas en membranas demuestran que el campo eléctrico es del orden de:
E N C≈ −107 1
Podemos calcular también el valor de la diferencia de potencial entre el citosol y el medio extracelular, que, suponiendo de nuevo que se trata de un condensador plano, con campo uniforme en su interior y tomando como valor estándar para el espesor de la membrana d ≈ 10 nm = 10‒8 m, será:
∆V E d V mV= ≈ × = =−10 10 0 1 1007 8 ,
Se adopta como convenio tomar el potencial exterior a la célula como cero, por lo que, como el citosol se encuentra negativamente cargado respecto al medio extracelular, el potencial en el interior es del orden de:
V mVi ≈ −100

Fuerzas, campos y potenciales eléctricos 27
aunque, como veremos en el capítulo 19, este valor puede sufrir variaciones notables debido a cambios en la concentración iónica del citosol, particular-mente de iones Na+ y K+, debido a modificaciones en la permeabilidad de la membrana plasmática. Estos cambios de permeabilidad de la membrana pueden ser debidos a diferentes estímulos, tanto químicos como físicos, lo que confiere a la célula propiedades de excitabilidad, es decir, la capacidad de reaccionar ante estímulos externos. Algunas células son particularmente activas desde este punto de vista, tales como las células nerviosas, cuyo estudio abordaremos en el capítulo 19.
Extraer un ion positivo desde el interior al exterior de la célula, por ejemplo un ion K+ de carga q = +e, requiere realizar un trabajo:
W q V e V eV J= ≈ ( )× ( ) = = × × = ×− −∆ 1 0 1 0 1 1 6 10 0 1 1 6 1019 20, , , , ,
Por ello, las actividades nerviosa y muscular, que implican el flujo de iones a través de la membrana de las neuronas y células musculares respectiva-mente, requieren un aporte energético.
Capacidad
Considerando la geometría de la membrana, su capacidad eléctrica se puede expresar como:
C Sd
= ( )κ ε0
Como se verá en el capítulo 19, es habitual utilizar la capacidad específica o capacidad por unidad de área ce como la magnitud adecuada para carac-terizar la membrana.
c CS de = =
κ ε0
Si utilizamos como espesor característico de la membrana d ≈ 10 nm, y un valor de constante dieléctrica κ ≈ 9, se obtendría:
c F m F cme ≈× ×
≈ × =−
−− − −9 8 85 10
108 10 0 8
12
83 2 2, , µ
que concuerda razonablemente con el valor obtenido experimentalmente en diferentes tipos de células:
ce (experimental) ≈ 1 μF cm–2 = 10–2 F m–2

28 Fundamentos físicos de los procesos biológicos
Sin embargo, algunas membranas parecen tener capacidades especí-ficas con valores muy inferiores. Nos referimos a las fibras nerviosas con mielina (véase el capítulo 9), donde la capacidad específica se reduce a ce < 0,01 µF/ cm2, lo que tiene importantes efectos en la propagación del impulso nervioso. La razón de esta disminución en la capacidad específica estriba en la naturaleza de estas células. Una fibra nerviosa con mielina, típica de los vertebrados, está envuelta por una capa aislante, correspondiente a un recubrimiento de células especializadas, en una especie de arrollamiento espiral que aumenta el espesor en cerca de un factor 200. Como la capacidad y la capacidad específica son inversamente proporcionales al espesor d, sus valores resultan divididos por ese mismo factor. Como se verá más adelante, esto supone una disminución importante en los tiempos característicos de respuesta de la membrana (capítulos 18 y 19) que se traduce en un aumento en la velocidad de propagación de los impulsos nerviosos.
La densidad superficial de carga acumulada a ambos lados de la mem-brana es:
σ = = = ≈ × × =− − − −QS
CVS
c Ed C me 10 10 10 102 7 8 3 2
lo que representa una densidad superficial de iones monovalentes:
Ne
ionesm
ionesm
= =×
≈ × =−
−
σµ
101 6 10
6 10 6 0003
1915
2 2,.
Esto supone una número de iones pequeño, comparado con las concentra-ciones habituales en el citoplasma celular, como se comprobará más adelante, equivalente a una fracción de apenas 10–5 del número total de iones. Por tanto, se puede suponer que la célula mantiene, en términos macroscópicos, la neutralidad de carga.
El valor del campo eléctrico en la membrana celular es suficientemente alto como para provocar el flujo de iones entre el citosol y el exterior de la célula, cuando la permeabilidad de la membrana así lo permite. Estos flujos debidos al campo eléctrico (gradiente de potencial) junto a los debidos a gradientes de concentraciones determinan muchas de las propiedades celu-lares, incluyendo la propagación de impulsos nerviosos, como se verá en el capítulo 19.
Observación. Peces eléctricos. Electrorrecepción. Acabamos de ver que todas las células experimentan actividad eléctrica en sus membranas (especialmente las células nerviosas y musculares). En consecuencia,

Fuerzas, campos y potenciales eléctricos 29
todos los organismos generan señales eléctricas que pueden ser detectadas externamente.
Algunos peces son electrorreceptivos, es decir, tienen la capacidad de detectar campos eléctricos, lo que a veces ha sido considerado como un “sexto sentido”. Los tiburones son un ejemplo de peces capaces de detectar a sus presas a través del campo eléctrico, lo que les permite su localización incluso si se encuentran enterradas bajo la arena. Pero el nombre de peces eléctricos suele reservarse para aquellos que no solo son capaces de detectar los campos eléctricos, sino que son “electrogénicos”, es decir, ellos mismos son capaces de generar campos eléctricos. Estos peces poseen un órgano eléctrico (OE) especializado, mediante el cual son capaces de generar descargas eléctricas, en general en forma de pulsos.
Los “peces eléctricos débiles” generan descargas débiles (E < 0,5 V/cm) que utilizan para la detección (electrolocalización) y para comunicación entre individuos (electrocomunicación), aunque también existen “peces eléctricos fuertes”, como la anguila eléctrica o el pez torpedo, capaces de generar descargas suficientemente intensas como para aturdir a sus presas.
Muchos de los peces eléctricos habitan en aguas turbias con baja visibilidad, por lo que el sentido de la electrolocalización aporta ventajas evidentes en este entorno. La electrolocalización es en cierto modo equivalente a la ecolocalización mediante ondas sonoras, utilizada por los murciélagos en el aire y por los delfines y ballenas en el agua (capítulo 21). Se trata de una loca lización activa, en la que el individuo detector se encarga también de emitir la señal que, tras rebotar o ser distorsionada por un objeto, se va a emplear para la detección. En el caso de la electrolocalización, el pez produce una descarga eléctrica, debido al desplazamiento de cargas que genera en el órgano eléctrico. Este consiste en un apilamiento de células excitables (electrocitos), derivadas de la modificación de células musculares o nerviosas, cuya disposición les permite producir señales eléctricas intensas, estando localizado en muchas especies (aunque no todas) en la región caudal (fig. 17.19).
La separación de cargas que se produce en el órgano eléctrico produce un campo eléctrico de tipo dipolar en torno al animal. El campo eléctrico es detectado por electrorreceptores, localizados principalmente a lo largo de la línea lateral del pez, alcanzando la máxima densidad en la cabeza. Por ejemplo, el Apteronotus albifrons tiene unos 15.000 electrorreceptores, que se extienden a lo largo de todo el cuerpo (12 receptores/mm2) y cuya máxima densidad (1020 receptores/mm2) se sitúa cerca de la boca del animal. Hay dos tipos de órganos electrorreceptores que difieren morfológica

30 Fundamentos físicos de los procesos biológicos
y funcionalmente: los receptores “ampulares” (forma de ampolla) y “tubulares” (con forma de tubo). Los primeros están presentes en todos los peces electrosensitivos y responden a señales continuas o de baja frecuencia (50 Hz), mientras los segundos, que responden a estímulos de alta frecuencia (hasta 2.000 Hz), son característicos de los peces eléctricos activos.
En cuanto al rango de acción de la electrorrecepción, debe tenerse en cuenta (como hemos visto en la sección 3.2) que el campo eléctrico dipolar decae rápidamente con la distancia (E ~ 1/r3), por lo que su utilización tiene un alcance limitado. La electrolocalización no parece ser operativa a distancias superiores a unos pocos metros.
Fig. 17.19. Localización del órgano eléctrico en algunos de los peces eléctricos (a) pez elefante, (b) raya, (c) anguila.
17.7 Ejercicios propuestosEjercicio 17.1 En un cristal de cloruro sódico los iones Na+ y Cl– ocupan
posiciones correspondientes a una red cúbica, siendo la distancia entre iones más próximos d = 2,82 Å. Calcúlese (a) la fuerza entre dos iones Na+ y Cl
adyacentes, (b) la fuerza entre dos iones Na+ (a una distancia 2d).
Sol.: (a) 2,9 × 10‒9 N (b) 7,2 × 10‒10 N
Ejercicio 17.2 Tres cargas están alineadas, tal como se indica en la figura 17.20. La carga q1 = +2 C está a una distancia de 1 m de la carga q2 = –1 C, que, a su vez, dista 2 m de la tercera carga q3. Calcúlese el valor de esta última carga para que la fuerza total sobre q2 sea cero.
Sol.: q3 = +8 C

Fuerzas, campos y potenciales eléctricos 31
Fig. 17.20.
Ejercicio 17.3 Tres cargas se encuentran situadas como se indica en la figura 17.21. La carga q1 = +4 C está situada en el punto (–2, 0), la carga carga q2 está en el origen de coordenadas y la carga q3 = +1 C, situada en (0, –1). Calcúlese (a) el valor de la fuerza total sobre q2, (b) el módulo de la fuerza si q2 = –2 C.
Fig. 17.21.
Sol.: (a)
F K q i j Ne= × × +( )2 (b) F N= ×2 5 1010,
Ejercicio 17.4 Calcúlese la relación entre el valor de las fuerzas eléctrica y gravitatoria entre el protón y electrón en un átomo de hidrógeno.
Datos: masas del electrón y del protón. me = 9,11 × 1031 kg y Mp = 1,67 × 1027 kg ≈ 1840 me.
Sol.: F Fe G ≈ ×2 1039
Ejercicio 17.5 Al frotar una varilla plástica, ésta se carga eléctricamente y atrae un fragmento de papel, de masa m = 1 mg, que comienza a elevarse cuando la varilla se acerca a una distancia r = 0,5 cm. Si suponemos que esto es debido a que la varilla adquiere una cierta carga positiva +q y, por inducción, el papel adquiere una carga similar y de signo contrario a la varilla ( q), calcúlese (a) el valor de dicha carga q, (b) el número del exceso de electrones en el papel (N =q/e).
Sol.: (a) q = 1,6 × 10‒10 C (b) N = 109
Ejercicio 17.6 Dos cargas puntuales Q1 = +1 nC y Q2 = –2 nC (1 nC = 10–9 C) se sitúan sobre el eje X, en posiciones x1 = ‒1 m y x2 = +2 m, como indica