fluorescence in situ hybridization of metaphase chromosomes in suspension
TRANSCRIPT

int. j. radiat. biol 2001, vol. 77, no. 7, 787 ± 795
Fluorescence in situ hybridization of metaphase chromosomes insuspension
H. HE†‡, W. DENG‡§, M. J. CASSEL‡ and J. N. LUCAS‡*
(Received 24 November 2000; accepted 22 February 2001)
Abstract. for quantifying infrequent chromosome aberrationsPurpose: To describe a new method for FISH analysis of meta- induced by low-dose radiation exposures. By contrast,phase chromosomes in suspension. chromosome analysis by � ow cytometry is an attract-Materials and methods: Metaphase chromosomes in suspension ive alternative to the visual chromosome analysiswere isolated from a Chinese hamster Ö human hybrid cell line,
because the chromosomes can be analyzed at a high314-2 (1) Y, and a human cell line, GM 130B. During suspensionhybridization, speci� c chromosomes were labeled from these speed, approximately 103 chromosomes (equivalenttwo cell lines using either biotin-labeled human genomic DNA, to 20 human whole genomic cells) per second (Lucasa directly labeled human pancentromere DNA probe or a et al. 1991) . In comparison, a very experiencedchromosome #1 locus-speci� c probe. cytogenetist would take 10–20 min to analyze 20Results: The method allows, for the � rst time, recovery of large
human whole genomic cells on slides using FISH ornumbers of isolated individual hybridized chromosomes withgood morphology for both human Ö hamster hybrid and human even using an automatic metaphase � nder. In previ-cell lines. The results showed that 46–73% of the starting number ous experiments by Lucas et al. (1991) , slit-scan � owof total chromosomes can be recovered after a FISH in suspension cytometry was used to detect dicentrics. However, inprocedure. The well-preserved morphology of hybridized meta-
such analysis, translocations, insertions or inversionsphase chromosomes allowed (1) rapid detection of individualcould not be detected. It was expected that FISHhuman and hamster chromosome aberrations, (2) rapid counting
of the painted human chromosomes and (3) fast, clear detection in suspension with speci� c DNA probes to wholeof chromosome region-speci� c probes. This method o Ú ers a new chromosomes would allow those aberrations totool to assay chromosomes and DNA: it o Ú ers the possibility to be detected by � uorescence � ow cytometry, or bydevelop new techniques for sorting chromosomes based on FISH
magnetic sorting with magnetic beads binding tosignals, for early detection and screening of genetic diseasesspeci� c DNA probes (Dudin et al. 1988).and for bulk measurement of both balanced or unbalanced
chromosomal exchanges and rearrangements. FISH with repetitive PNA or speci� c DNA probesConclusion : The potential of the method described should facilitate to interphase nuclei in suspension has been reportedfast, sensitive population monitoring, and increase sensitivity of recently by several groups who were able to quantifythe measurements in chromosome-based biodosimetry.
the total genomic � uorescence intensity by � uores-cence � ow cytometry (Arkesteijn et al. 1995, Eleuteri
1. Introduction et al. 1997, Hultdin et al. 1998, Rufer et al. 1998,Takahashi et al. 1998, Truong et al. 1998) . WhileFluorescence in situ hybridization (FISH) hasthese techniques were successful in measuring thebecome a powerful, practical tool for detecting gen-genomic gains or losses of target chromosomes or DNAetic abnormality such as chromosome translocations,sequences, they are not able to identify or sort outpericentric inversions and insertions. Most frequently,individual abnormal chromosomes without genomicchromosomes are studied as mitotic metaphasesDNA content alteration.� xed on slides. However, FISH-based metaphase
For chromosome sorting and detailed individualchromosome aberration analysis by visualization ofchromosome aberration analysis with speci� c wholecells � xed on slides is still very time-consumingchromosome painting or speci� c arm painting oraccurate physical localization of DNA sequences, theisolation of metaphase chromosomes as well as*Author for correspondence at University of California, POthe preservation of their morphology is needed.Box 2573, Dublin, CA 94566, USA.However, the procedures in chromosome isolation†American Laboratory Technologies, Inc., Arlington, VA
22201, USA. and denaturation of target DNA are prone to cause‡University of California, Lawrence Livermore National chromosome morphology damage and chromosome
Laboratory, PO Box 808, Livermore, CA 94551, USA. loss (Kraus et al. 1995, Nguyen et al. 1995).§Sichuan Institute of Radiation Health and Protection,Other attempts at chromosome hybridization inSichuan Institute of Occupational Health, Chengdu, Sichuan,
610041, PR China. suspension for � ow cytometric analysis involves the
International Journal of Radiation Biology ISSN 0955-3002 print/ISSN 1362-3095 online © 2001 Taylor & Francis Ltdhttp://www.tandf.co.uk/journals
DOI: 10.1080/0955300011004754 6
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.

788 H. He et al.
use of FISH on chromosomes suspended within gel the day of use as follows: 5 ml of each of the stocksolutions I–III was combined with 35 ml of distilledmicrodrops (Nguyen et al. 1995) . This procedure,
however, results in most chromosomes becoming water and the pH adjusted to 7.2. A total of 50 ml ofb-mercaptoethanol and 0.06 g of digitonin wereball-shaped and thereby reducing its usefulness as a
tool to detect chromosome aberrations. added, and the bu Ú er was incubated at 37 ß C for45 min. Next, 25 ml of stock concentrations ofMacas et al. (1995) reported their protocol of
primed in situ DNA labeling for plant species and it spermine (0.4 m) and spermidine (1.0 m) were added,and the bu Ú er was � lter sterilized.was expected to provide a chromosomal sample for
� ow cytometry. However, to date, no such informa- Cells obtained from colcemid treatment weretransferred to conical tubes (one 50 ml tube pertion on animal and human species has been
published. T-150 or T-75 � ask) and centrifuged for 10 min atapproximately 250 g. The supernatant was removedHere, the authors report their new protocols for
in situ hybridization of whole chromosomes in suspen- and cells were resuspended in 5 ml of hypotonicbu Ú er (75 mm KCl, 0.2 mm spermine, 0.5 mmsion, which yield high recovery percentages of hybrid-
ized chromosomes with well-preserved morphology. spermidine) and swelling was allowed to proceed for25 min at room temperature, after which time thecells were centrifuged at 250 g for 6 min and the2. Materials and methodssupernatant removed. Cells were then resuspended
2.1. Cell culture in 1 ml of chromosome isolation bu Ú er and placedon ice for 10 min. Cells were vortexed vigorously forHuman GM 130B cells, a lymphoblast-derived cell30 s and placed on ice for 5 min.line, (Mutant Cell Repository, Camden, NJ, USA)
were grown in RPMI 1640 medium supplemented2.2.2. Chinese hamster Ö human hybrid cell line: Thewith 15% fetal calf serum, 1% l-glutamine and 1%mitotic cells were pelleted by centrifugation (350 g,penicillin/streptomycin solution, then blocked in col-10 min). The hypotonic treatment with a solution ofcemid for 12–16 h. Cells were then taken immediately10 mm Tris, 5 mm MgCl2 , 10 mm NaCl and 0.1 mginto the chromosome isolation procedure. TheRNase/ml at pH 7.5 was applied for 15 min at 37 ß C.Chinese hamster Ö human hybrid cell line 314-2 (1)After centrifugation (350 g, 10 min), the hypotonicY (with human chromosome #3 only) was grown insolution was removed. The chromosome pellet wasAlpha Medium supplemented with 5% fetal calfresuspended in cold hexylene glycol chromosomeserum, 1% l-glutamine and 1% penicillin/streptomy-isolation bu Ú er, as described by Stoehr et al.cin solution in plastic � asks. Growth medium in(1982) (25 mm Tris-HCl bu Ú er, pH 7.5; 0.75 mculture � asks was replaced by fresh medium con-hexandiol-(1,6); 0.5 mm CaCl2-2H2O). The chromo-taining 0.12 mg colcemid/ml. Cells were blockedsome suspension obtained was kept on ice for 5 min,in this medium for 6 h. Mitotic cells were harve-then treated with ultrasound for 1 min in an icebath.sted by shake-o Ú , and taken immediately into theIsolated chromosomes remained storable for monthschromosome isolation procedure.at 4 ß C.
A 20 ml sample was removed and combined2.2. Isolation of metaphase chromosomes in suspension with 10 ml of 4,6-diamidino-2-phenylindole (DAPI)
(0.25 mg/ml; Sigma, St Louis, MO, USA). TheBecause of the di Ú erences in the response of humanstained chromosome suspension was transferred to aand hamster Ö human cell lines to chromosome iso-hemocytometer and the number of chromosomeslation procedures, separate protocols had to bewas counted by the authors’ modi� ed method.developed for each cell line.
2.2.1. Human GM 130B cell line: The isolation proced- 2.3. Preparation of the probesure was adopted from the method described byBlumenthal et al. (1979) with modi� cations. Isolation Human genomic DNA was labeled by nick transla-
tion with biotin-16-dUTP. A BioNickTM labeling kitbu Ú er: 15 mm Tris base, 80 mm KCl, 20 mm NaCl,2 mm EDTA, 0.5 mm EGTA, 0.2 mm spermine, (GIBCO BRL) was used according to the manu-
facturer’s protocol. Human pancentromere DNA0.5 mm spermidine, 14 mm b-mercaptoethanol 0.1%digitonin. speci� c probe was ampli� ed by DOP-PCR using a
commercial kit (Boehringer Mannheim Gmbh.,Stock concentrations of the � rst � ve ingredientswere prepared as follows: (I) 150 mm Tris base, Germany) and labeled with tetramethylrhodamine-
6-dUTP (Boehringer Mannheim). Chromosome #1800 mm KCL, 200 mm NaCl, (II) 20 mm EDTA, (III)5 mm EGTA. The isolation bu Ú er was prepared on locus-speci� c DNA pUC1.77 was isolated from the
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.

789FISH of metaphase chromosomes in suspension
plasmid pUC1.77 (Cooke and Hindley 1979) . Total methanol:acetic acid � xative to the isolation bu Ú ersuspension containing the chromosomes while gentlyisolated plasmid DNA was directly labeled in
FluoroRed (Amersham, Piscataway, N.J., USA) using mixing. The chromosomes were then centrifugedand the isolation bu Ú er removed. A total of 1 mla standard random priming kit (GIBCO, BRL,
Gaithersburg, MD, USA). � xative was added drop-wise while vortexing thesample on the lowest setting. The sample was thenincubated at room temperature for 30 min, centri-2.4. Chromosome in situ hybridization in suspension fuged, and the supernatant removed and replacedwith 1 ml of fresh � xative.As with chromosome isolation above, the two cell
lines required development of di Ú erent hybridization Isolated human chromosomes were hybridized asdescribed above with the following modi� cations: (1)procedures.2.5 ml of directly labeled chromosome #1 speci� cpUC1.77 probe without Cot-1 DNA was used; (2)2.4.1. Chinese hamster Ö human hybrid cell line: Isolated
chromosomes were � xed in methanol:acetic acid hybridization proceeded for 2.5 h at 37 ß C; (3) post-hybridization washes consisted of one 5 min wash at(3:1) prior to hybridization. This � xation step is
crucial to chromosome stability. The sample con- 37 ß C in the 0.1 Ö SSC wash bu Ú er.taining about 3.5 Ö 106 isolated chromosomes wasallocated into a 5 ml Falcon tube (Becton Dickinson, 2.5. Detection of recovery of hybridized chromosomeFranklin Lakes, NJ, USA), and 500 ml of meth-
After the post-hybridization wash, the hybridizedanol:acetic acid (3:1) was added. After 10 min, thechromosomes were resuspended in a solution con-� x solution was removed by centrifugation (350 g,sisting of 30 ml 2 Ö SSC bu Ú er and 70 ml DAPI. The10 min), and the chromosome pellet was resuspendedstained chromosome suspension was transferred to ain 160 ml prewarmed hybridization mixture (40%hemocytometer and the number of recovered chro-deionized formamide, 4 Ö SSC, 2 Ö Denhardt’s solu-mosomes was counted. The initial number of chro-tion). Next, 20 ml (0.36 mg) biotinylated human gen-mosomes for each FISH in suspension reaction wasomic DNA probe, 15 ml (0.27 mg) directly labeled� xed. After each FISH in suspension reaction, thehuman pancentromere DNA probe and 5.3 mlremaining chromosomes were resuspended in a cer-(1 mg/ml) hamster Cot-1 (GIBCO) were added totain volume and the chromosome concentration wasthe hybridization mixture. The chromosomes weredetermined using a hemocytometer. Thus, the initialdenatured for 5 min at 73 ß C in a waterbath, thennumber of chromosomes used for each reaction andplaced on ice for 5 min. The chromosomes werethe remaining number in each sample after eachhybridized overnight at 43 ß C in a shaking waterbath.reaction was known. This allowed us the comparisonFor post hybridization washing, chromosomes wereof chromosome loss for di Ú erent FISH in suspensionpelleted by centrifugation at 350 g for 10 min, washedprocedures and protocols.in 500 ml of prewarmed washing solution I (2 Ö SSC,
500 mg/ml bovine serum albumin (Amersham LifeScience Inc., Piscataway, N.J., USA), 5 mm EDTA) 2.6. Detection of hybridization signalfor 25 min at 43 ß C in a shaking waterbath. After the
After the post-hybridization wash and centri-second centrifugation, the chromosomes were resus-fugation, the hybridized chromosomes were re-pended into washing solution II (0.1 Ö SSC,suspended in 15 ml 2 Ö SSC and 22 ml DAPI,500 mg/ml bovine serum albumin, 5 mm EDTA) anddropped on a slide, immediately covered by apassed through the second wash procedure for 25 mincoverslip, observed under a � uorescence microscopeat 43 ß C in a shaking waterbath. The washing solutionand then photographed while still in suspensionwas removed by centrifugation at 350 g for 10 min.under a coverslip.The chromosomes were resuspended in 70 ml PNM
bu Ú er for 5 min. Then, 1.2 ml avidine-FITC wasadded to the suspension and the suspension was 3. Results and discussionincubated 20 min at 30 ß C in a shaking waterbath. 3.1. Fluorescence in situ hybridizationAfter centrifugation the chromosomes were washedtwice in 500 ml PN bu Ú er, and centrifuged again. 3.3.1. Chinese hamster Ö human cell line: Metaphase
chromosomes were isolated from the ChineseThe centrifugations always took place at 350 g for10 min. hamster Ö human hybrid cell line 314-2 (1) Y, which
contains only human chromosome #3 in a hamsterbackground. Isolated chromosomes were hybridized2.4.2. Human GM 130B cell line: Isolated chromo-
somes were � xed by slowly adding 20 drops of 3:1 in suspension to biotin-labeled human genomic DNA
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.

790 H. He et al.
probe. After avidin-FITC staining of chromosomes human cell line were found to be less tolerant of thedenaturation conditions used for the hybridizationin suspension, a large number of hybridized meta-
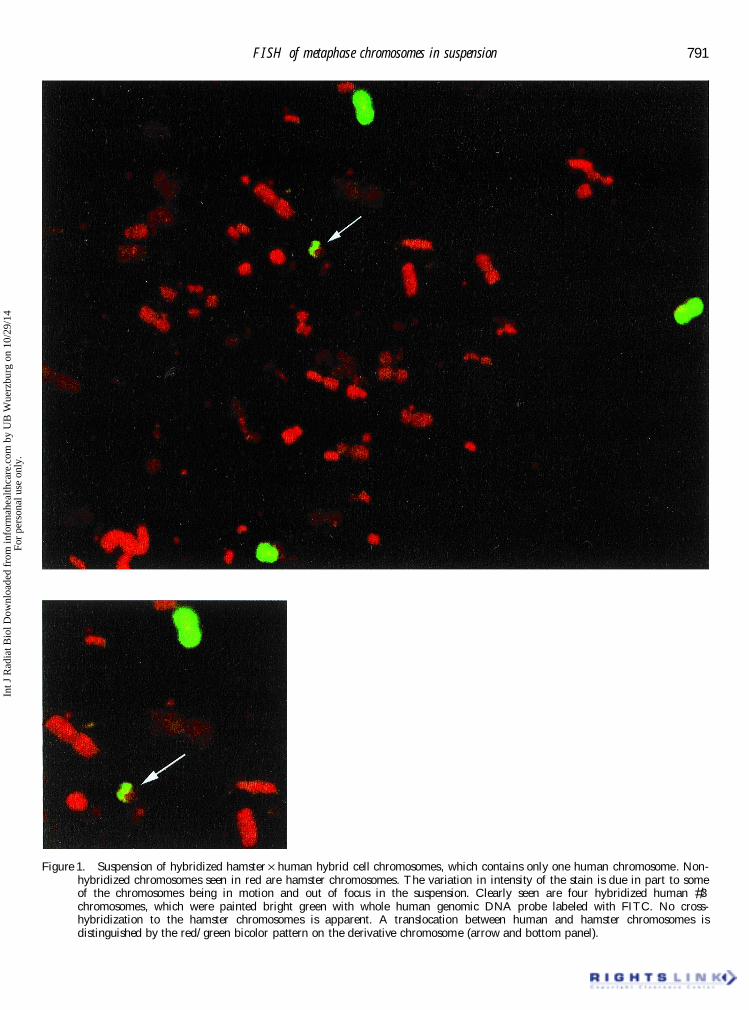
phase chromosomes with good morphology were procedure. Inclusion of a protease inhibitor in theisolation bu Ú er and 3:1 methanol:acetic acid � xationobtained (see � gure 1). Seen in that � gure in red are
hamster chromosomes and in green are human #3 immediately after isolation greatly improved the post-hybridization morphology of chromosomes isolatedchromosomes. No cross-hybridization to the hamster
chromosomes is apparent. A translocation between from human cell lines. Chromosomes isolated fromhuman cell lines in this manner could be stored forhuman and hamster chromosomes is distinguished
by the red/green bicolor pattern on the derivative up to 4 weeks prior to hybridization with minimalloss of morphology following hybridization.chromosome (� gure 1: arrow). The variation in � uo-
rescent intensity of the hamster chromosomes is duein part to some of the chromosomes being in motion 3.2. Recoveryand out of focus in the suspension.
It is important, when using FISH for radiation A low recovery of chromosomes after FISH insuspension compromises chromosome aberrationcytogenetics, to distinguish dicentrics from transloca-
tions. Concomitant pan-centromeric probing with analysis and cannot compete with hybridization onslides. The recovery of chromosomes followingwhole chromosome painting was introduced to make
this possible (Weier et al. 1991) . This technique hybridization in suspension was evaluated by com-paring with percentage of chromosomes recoveredallowed us the checking of the speci� city of hybridiza-
tion in suspension for hamster Ö human cell line after hybridization with the starting number of chro-mosomes before hybridization. The chromosomechromosomes also, by employing two speci� c probes
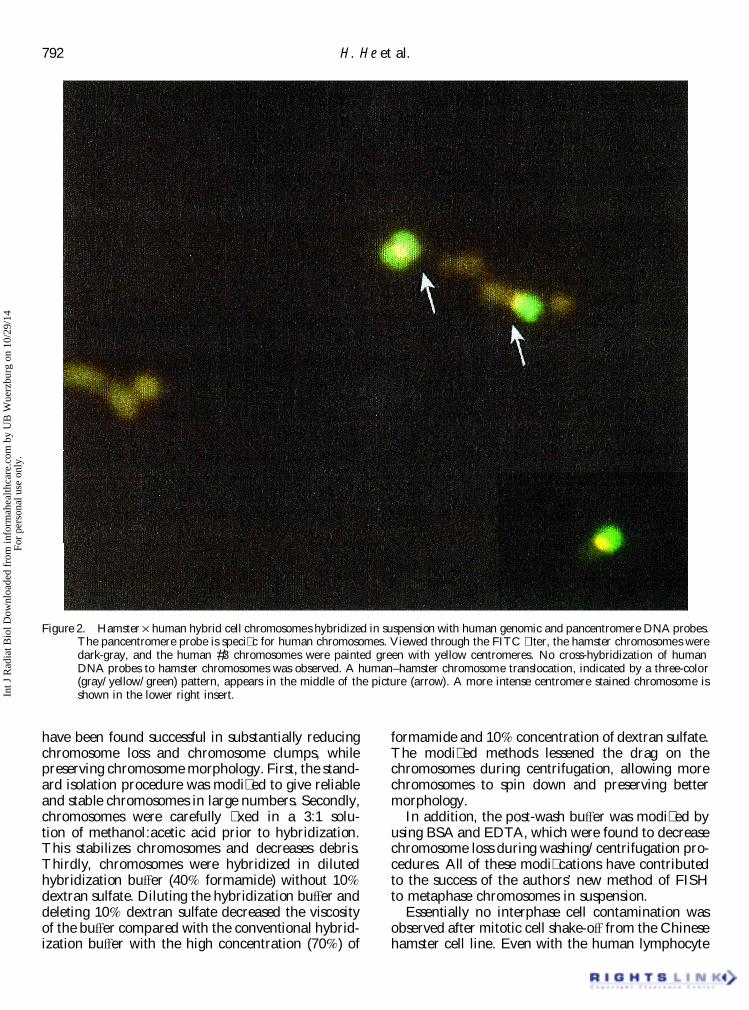
to simultaneously stain the human chromosome. A recovery after hybridization in suspension and washesby the authors’ methods ranged from 46% to 73%pancentromere probe was used, which � uoresced
red, and a human genomic DNA probe, which (table 1). Table 1 shows the reproducibility of thismethod to recover large numbers of chromosomes� uoresced green on human #3 chromosome. When
visualized simultaneously, the red and green signals after hybridization. As seen in the table, chromosomerecovery was routinely over 60%. Similar results wereproduced a yellow � uorescence at the centromeres
of chromosome #3 (� gure 2). No separated red and obtained for solution-hybridization of the human cellline GM 130B as for the hamster Ö human hybridgreen signals were observed in the 124 chromo-
somes #3 analyzed. A human–hamster chromosome cell line.Because previous studies (Dudin et al. 1987, 1988)translocation, indicated by a three-color (gray/
yellow/green) pattern, appears in the middle of did not provide quantitative information on chromo-some loss in their publications (although they did� gure 2. Also, the FISH measured frequency of
hybridized human chromosome #3 in suspension indicate substantial chromosome loss), their methodswere repeated in order to compare directly with thecorrelated well with the FISH measured frequency
of chromosome #3 on glass slides. Using the present authors’ method for chromosome recovery.Both methods were performed in parallel on aliquotshamster Ö human cell line, the number of painted
chromosomes (chromosomes with positive FISH sig- of the same chromosome suspension. In a comparat-ive study in the authors’ laboratory, 62.9% of chro-nals) after hybridization in suspension were compared
with that after conventional FISH on slides. The mosomes were recovered compared with 4.6% usingthe Dudin method (table 2). The methods of theresults showed that 5.05% (51/1015) of chromosomes
were painted after FISH in suspension and 4.8% authors resulted in more than 10 times higher chro-mosome recovery than that obtained by the methods(48/1005) were painted after conventional FISH on
slides in human Ö hamster cell lines. The results are of Dudin et al. (1987, 1988) . The high chromosomerecovery after FISH in suspension by the authors’not signi� cantly di Ú erent ( p > 0.05).methods makes FISH in suspension a potentiallypractical tool for rapid and bulk analysis of chromo-3.1.2. Human cell line: Metaphase chromosomes were
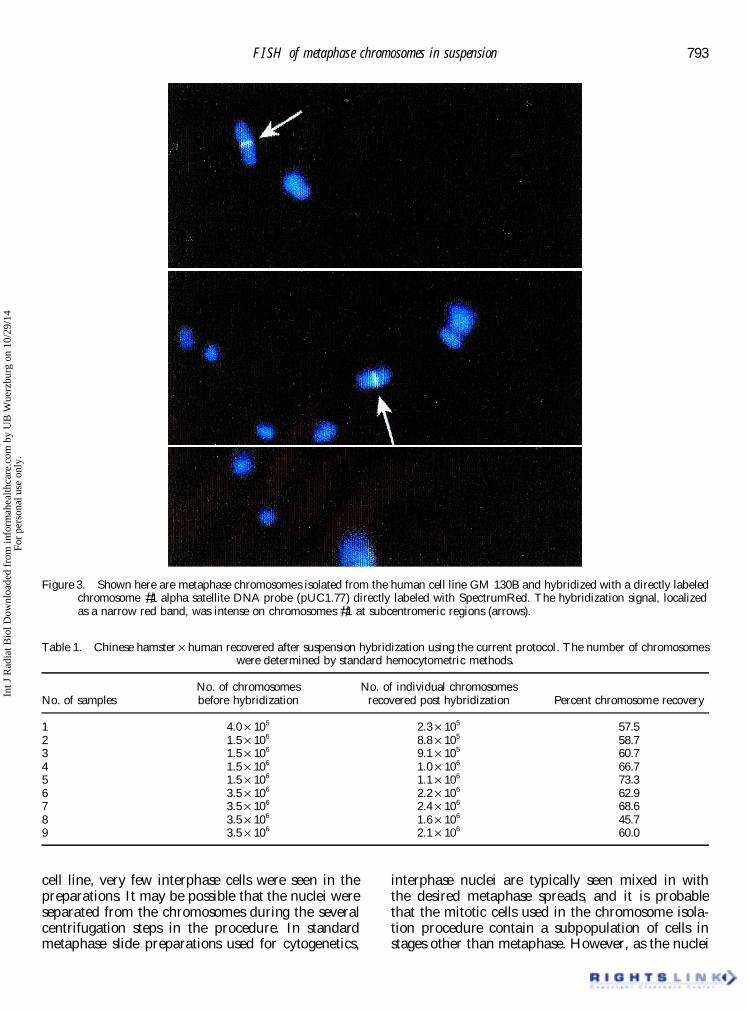
isolated from the human cell line GM 130B and some aberrations using � ow cytometry.hybridized with a directly labeled chromosome #1alpha satellite DNA probe following the optimized 3.3. Improvement in the authors’ methodsprotocol described in §2. The probe was directlylabeled with SpectrumRed. As can be seen in � gure 3, Previous attempts to perform in situ hybridization
on metaphase chromosomes in suspension werethe hybridization signal, localized as a narrow redband, was intense on chromosomes #1 at subcen- hindered by chromosome loss, breakage, and
aggregation. In the authors’ protocol, several stepstromeric regions. Chromosomes derived from the
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.
791FISH of metaphase chromosomes in suspension
Figure 1. Suspension of hybridized hamster Ö human hybrid cell chromosomes, which contains only one human chromosome. Non-hybridized chromosomes seen in red are hamster chromosomes. The variation in intensity of the stain is due in part to someof the chromosomes being in motion and out of focus in the suspension. Clearly seen are four hybridized human #3chromosomes, which were painted bright green with whole human genomic DNA probe labeled with FITC. No cross-hybridization to the hamster chromosomes is apparent. A translocation between human and hamster chromosomes isdistinguished by the red/green bicolor pattern on the derivative chromosome (arrow and bottom panel).
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.
792 H. He et al.
Figure 2. Hamster Ö human hybrid cell chromosomes hybridized in suspension with human genomic and pancentromere DNA probes.The pancentromere probe is speci� c for human chromosomes. Viewed through the FITC � lter, the hamster chromosomes weredark-gray, and the human #3 chromosomes were painted green with yellow centromeres. No cross-hybridization of humanDNA probes to hamster chromosomes was observed. A human–hamster chromosome translocation, indicated by a three-color(gray/yellow/green) pattern, appears in the middle of the picture (arrow). A more intense centromere stained chromosome isshown in the lower right insert.
have been found successful in substantially reducing formamide and 10% concentration of dextran sulfate.The modi� ed methods lessened the drag on thechromosome loss and chromosome clumps, while
preserving chromosome morphology. First, the stand- chromosomes during centrifugation, allowing morechromosomes to spin down and preserving betterard isolation procedure was modi� ed to give reliable
and stable chromosomes in large numbers. Secondly, morphology.In addition, the post-wash bu Ú er was modi� ed bychromosomes were carefully � xed in a 3:1 solu-
tion of methanol:acetic acid prior to hybridization. using BSA and EDTA, which were found to decreasechromosome loss during washing/centrifugation pro-This stabilizes chromosomes and decreases debris.
Thirdly, chromosomes were hybridized in diluted cedures. All of these modi� cations have contributedto the success of the authors’ new method of FISHhybridization bu Ú er (40% formamide) without 10%
dextran sulfate. Diluting the hybridization bu Ú er and to metaphase chromosomes in suspension.Essentially no interphase cell contamination wasdeleting 10% dextran sulfate decreased the viscosity
of the bu Ú er compared with the conventional hybrid- observed after mitotic cell shake-o Ú from the Chinesehamster cell line. Even with the human lymphocyteization bu Ú er with the high concentration (70%) of
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.
793FISH of metaphase chromosomes in suspension
Figure 3. Shown here are metaphase chromosomes isolated from the human cell line GM 130B and hybridized with a directly labeledchromosome #1 alpha satellite DNA probe (pUC1.77) directly labeled with SpectrumRed. The hybridization signal, localizedas a narrow red band, was intense on chromosomes #1 at subcentromeric regions (arrows).
Table 1. Chinese hamster Ö human recovered after suspension hybridization using the current protocol. The number of chromosomeswere determined by standard hemocytometric methods.
No. of chromosomes No. of individual chromosomesNo. of samples before hybridization recovered post hybridization Percent chromosome recovery
1 4.0 Ö 105 2.3 Ö 105 57.52 1.5 Ö 106 8.8 Ö 105 58.73 1.5 Ö 106 9.1 Ö 105 60.74 1.5 Ö 106 1.0 Ö 106 66.75 1.5 Ö 106 1.1 Ö 106 73.36 3.5 Ö 106 2.2 Ö 106 62.97 3.5 Ö 106 2.4 Ö 106 68.68 3.5 Ö 106 1.6 Ö 106 45.79 3.5 Ö 106 2.1 Ö 106 60.0
cell line, very few interphase cells were seen in the interphase nuclei are typically seen mixed in withthe desired metaphase spreads, and it is probablepreparations. It may be possible that the nuclei were
separated from the chromosomes during the several that the mitotic cells used in the chromosome isola-tion procedure contain a subpopulation of cells incentrifugation steps in the procedure. In standard
metaphase slide preparations used for cytogenetics, stages other than metaphase. However, as the nuclei
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.

794 H. He et al.
Table 2. Fraction of chromosomes recovered for the two aberrations, such as chromosome translocations, canmethods after suspension hybridization, as determined by be detected by a combined hybridization with probes� uorescence microscopy. The cell line used was the 1 and 2. In such cases, probes 1 and 2 are speci� chamster Ö human hybrid described in §2.
for separate DNA sequences. Magnetic beads thatNo. of chromosomes bind to probe 1 are used to separate chromosomes
that contain probe 1. After separation, the beads areBefore Recovered after cleaved from the chromosomes. From the separated
Method hybridization hybridization Recovery chromosomes hybridized with probe 1, magneticbeads that bind to probe 2 are used to separateHamster Ö human
Dudin et al. 1987a 3.5 Ö 106 1.6 Ö 105 4.6% chromosomes that contain probe 2. Thus, the � nalCurrent study 3.5 Ö 106 2.2 Ö 106 62.9% pool of separated chromosomes will be aberrant
chromosomes containing both probe 1 and probe 2.HumanCurrent study 4.0 Ö 106 2.5 Ö 106 62.5% Therefore, the aberrant chromosomes can be isolated
and sorted rapidly for bulk analyses.a Data generated in our lab using the mthod of Dudin et al.(1987).
Referencesof the cells in the undesired phases are intact, the Arkestei jn, G. J. A., Erpelinck, S. L., Martens, A. C. andchromosomes in those cells are not free to interact Hagenbeek, A., 1995, Chromosome speci� c DNA
hybridization in suspension for � ow cytometric detectionwith the chromosomes released from the mitotic cells.of chimerism in bone marrow transplantation and leuke-While it is possible that nuclei from non-mitotic cellsmia. Cytometry, 19, 353–360.can cause clumping of free chromosomes, no such
Blumenthal, A. B., Dieden, J. D., Kapp, L. N. and Sedat,phenomenon was observed in these experiments. J. W., 1979, Rapid isolation of metaphase chromosomesThis study clearly demonstrates that isolated chro- containing high molecular weight DNA. Journal of Cell
Biology, 81, 255–259.mosomes in suspension can be successfully hybridizedCooke, H. J. and Hindley, J., 1979, Cloning of human satellitein solution with a high recovery and low number of
III DNA: di Ú erent components are on di Ú erent chromo-clumps. The recovered hybridized chromosomessomes. Nucleic Acids Research, 6, 3177–3197.maintained good morphology, similar to that Dudin, G., Cremer, T ., Schardin, M., Hausmann, M., Bier,
observed on glass slide hybridization. The methods F. and Cremer , C., 1987, Methods for nucleic acidhybridization to isolated chromosomes in suspension.are reliable and show consistency from repeatHuman Genetics, 76, 290–292.experiments.
Dudin, G., Steegmayer , E. W., Vogt, P ., Schnitzer, H., Diaz,E., Howell, K. E., Cremer, T . and Cremer , C., 1988,Sorting of chromosomes by magnetic separation. Human3.4. Potential application of the methodGenetics, 80, 111–116.
Eleuteri, P ., Grollino, M. G., P omponi, D., Calugi , G.,The isolation and FISH in suspension of meta-Rocchi, M. and de Vita, R., 1997, DNA � ow cytometricphase chromosomes with good morphology, highanalysis and numerical chromosome 9 aberrationschromosome recovery and high speci� city of hybrid-detected by interphase cytogenetics in super� cial bladderization will be very valuable for physical localization TCC. European Journal of Histochemistry, 41 (suppl. 2),
of low-copy-number DNA sequences on chromo- 161–162.Hultdin, M., Gronlund, E., Norrback, K., Eriksson-somes and for rapid chromosome sorting and chro-
Lindstrom, E., Just, T . and Roos, G., 1998, Telomeremosome aberration analysis using � uorescence � owanalysis by � uorescence in situ hybridization and � owcytometry. It may be readily adapted for rapid andcytometry. Nucleic Acid Research, 26, 3651–3656.early diagnosis of genetic diseases. Because of the Kraus, M., Hausmann, M., Hartig, R., Durm, M., Haar,
ability to clearly localize a speci� c DNA sequence F. M. and Cremer , C., 1995, Non-encymatic, low temper-ature in situ hybridization of metaphase chromosomes foron a chromosome by FISH in suspension (as demon-magnetic labelling and sorting. Experimental Technique ofstrated in � gure 3), the technique is particularly usefulPhysics Experimentelle Technik der Physik, 41, 139–153.for identifying a nucleic acid sequence as well as
Lucas, J. N., Mullikin, J. C. and Gray, J. W., 1991, Dicentricchromosomal re-arrangements. chromosome frequency analysis using slit-scan � owBecause metaphase chromosomes can be released cytometry. Cytometry, 12, 316–322.
Macas, J., Dolezel, J., Gualberti , G., P ich, U., Schubert, I.from cells and hybridized with speci� c probes usingand Lucretti , S., 1995, Primer-induced labeling of peathe authors’ technique, the suspension hybridizationand � eld bean chromosomes in situ and in suspension.technique will also have important potential applica-Biotechniques , 19, 402-404.tions in magnetic sorting of individual chromosomes Nguyen, B. T ., Lazzari, K., Abebe, J., Mac, I., Chang, A.,
or speci� c DNA sequences. As an example of one of Wydner, K. L., Lawrence, J. B., Cram, L. S. andWeier, H. U., Weaver, J. C. and Bradley, D. W.,the proposed applications, the target chromosome
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.

795FISH of metaphase chromosomes in suspension
1995, In situ hybridization to chromosomes stabilized in � uorescence-activated cell sorting and � uorescence in situhybridization. Blood, 92, 4758–4763.gel microdrops. Cytometry, 21, 111–119.
Rufer, N., Dragowska, W., Thornbury, G., Roosnek, E. and Truong, K., Guilly, M. N., Gerbault-Seureau, M., Malfoy,B., Vielh, P ., Bourgeois, C. A. and Dutrillaux, B.,Lansdorp, P., 1998, Telomere length dynamics in human
lymphocyte subpopulations measured by � ow cytometry. 1998, Quantitative FISH by image cytometry for thedetection of chromosome 1 imbalances in breast cancer:Nature, Biotechnology, 16, 743–747.
Stoehr, M., Hutter, K. J., Frank, M. and Goerttler , K. I., a novel approach analyzing chromosome rearrangementswithin interphase nuclei. Lab Investigation, 78, 1607–1613.1982, A reliable preparation of mono-dispersed chromo-
some suspensions for � ow cytometry. Histochemistry , 74, Weier, H-U., Lucas, J. N., P oggensee, M., Segraves, R.,P inkel, D. and Gray, J. W., 1991, Two-color hybridiza-57–61.
Takahashi , N., Miura, I., Saitoch, K. and Miura, A. B., tion with high complexity chromosome-speci� c probesand a degenerate alpha satellite probe DNA allows1998, Lineage involvement of stem cells bearing the
philadelphia chromosome in chronic myeloid leukemia unambiguous discrimination between symmetrical andasymmetrical translocations. Chromosoma, 100, 371–376.in the chronic phase as shown by a combination of
Int J
Rad
iat B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
B W
uerz
burg
on
10/2
9/14
For
pers
onal
use
onl
y.