fiziologija 1
DESCRIPTION
skripta za fiziologiju I psihologijaTRANSCRIPT

Skripta za Fiziologiju 1

by J.M., Ninche, Sandra, Ranagol, gen2010
I DEOOBJAŠNJENJE:
1. Times New Roman: materijal iz stare skripte
2. Arial: prepisano iz različitih sveski ili rukopisa
3. Comic Sans MS: materijal sa predavanja
4. Microsoft Sans Serif: skripta gen 2010
1. Neuron (građa, razvoj) / 2. Funkcionalni značaj pojedinih delova neurona
Neuron je osnovna genetska, strukturalna i funkcionalna jedinica nervnog sistema. Spososbni su da primaju i
prosleđuju informaciju. Nastali su iz spoljnog klicinog lista, ektodermalnog su porekla. Stvaraju se tokom
embrionalnog života, a pred kraj veliki broj propada, jer se nisu dobro formirali (Apoptoza=genetski
programirana smrt nervne ćelije, odvija se tokom celog života, određenim tempom.)
Pre rođenja postoji veliki broj nervnih ćelij u našem nervnom sistemu, od čega velikik broj propada, zbog
bolje diferencijacije preostalih. To je važno za dalji razvoj.
Glavni delovi neurona: telo, akson, dendriti. Neuronska tela ulaze u sastav sive mase CNS, dok aksoni čine
belu masu CNS. Dendriti služe za prijem informacija.
1. Neuron (građa, razvoj)
Centralni nervni sistem čine mozak i kičmena moždina.

by J.M., Ninche, Sandra, Ranagol, gen2010
Centralni nervni sistem čine: 1.Neuroni
2. Glijalne ćelije
3. Krvni sudovi
4. Moždana tečnost
Nervni sistem je ektodermalnog porekla, nastaje iz trećeg klicinog lista.
Nervna ćelija (neuron) je osnovna genetska, strukturna i funkcionalna jedinica CNS, stvaraju se u tokuembriogeneze (od začeća do fetalnog perioda, tada su još nezrele i nisu strogo diferencirane), posle rođenjasazrevanje neurona se nastavlja, međutim gledano u toku života broj neurona se stalno smanjuje. Nervnećelije se ne dele i ne obnavljaju (osim neurona hipokampa i mirisnog puta).
Radjamo se sa oko 100 biliona nervnih ćelija. Muskarci i žene se razlikuju po ovom broju i kod žena su sitnijei složenije građe.
Nervna ćelija je jedinstvena struktura na kojoj se razlikuje:
1.Telo nervne ćelije (neuronsko telo, soma ili perikarion) 2. Dva tipanastavaka- dendriti i aksoni
Neuronska tela ulaze u sastav sive mase CNS-a i perifernih ganglija (spinalne i vegetativne ganglije), dokaksoni ulaze u sastav bele mase CNS-a, a takođe čine i periferne živce. Nervne ćelije sa ove dve vrstenastavaka zovu se heteropolarne i one su karakteristične za kičmenjake. Beskičmenjaci obično imajuneurone sa jednim razgranatim aksonskim nastavkom. To su unipolarni neuroni. Nervne ćelije su veomaraznokolikog oblika i to je jedna od njihovih izrazitih osobina. Smatra se da se u mozgu kičmenjaka nalaziukupno 100 milijardi nervnih ćelija, koje se međusobno znatno razlikuju po veličini i broju dendrita, po veličinisome, dužini i veličini aksona, broju sinaptičkih završetaka. Neki su polarni, pseudopolarni, a najviše imamultipolarnih neurona (izgrađeni od some, jednog aksona i veoma razgranatog dendritskog stabla). Overazlike čine da neuroni različitih delova nervnog sistema reaguju različito na prispele signale, zbog čega iobavljaju različite funkcije. Neuronsko telo moze biti različitog oblika: zvezdastog, okruglog, piramidalnog.Sadrži jedro sa jednim, ili retko sa dva jedara. Citoplazma sadrži: neurotransmitere, i mikrotubule, Nisslovatelašca sa endoplazmatičnim retikulomom, Goldžijev aparat, mitohondrije, a može i da sadrži i granule sapigmentom. Treba istaći da su neurotubule, neurofilamenti i mikrotubule končaste strukture proteinskogsastava i da verovatno vrše transport materije kroz akson.
Dendriti - Na neuronsko telo se često nadovezuje veliki broj dendrite, po pravilu razgranatih i kratkih, alimogu da budu i znatno duži, kao sto je slučaj sa tzv. apikalnim dendritima piramidalnih ćelija velikog mozga.Na povrsini dendrita se često nalaze sićušna protoplazmatska ispupčenja koja se nazivaju trnasti izražaji izbog njih je površina dendrita hrapava. Broj trnastih ispupcčnja varira i on se povćcava tokom embrionalnograzvica nervnog sistema, što se uzima kao dokaz da se u tom periodu uspostavljaju veze (sinapse) izmeđuneurona. Pomoću trnastih izraštaja povećava se površina membrane dendrita što je značajno za njihovesinaptičke veze. Zajedno sa neuronskim telom dendriti čine aferentni deo neurona, pošto sa njima najčešćestupaju u kontakt druge nervne ćelije ili receptori.
Aksoni - Prema dužini aksona razlikuju se 2 kategorije neurona:
Goldži I- nervna vlakna ove kategorije ulaze u sastav dugačkih ushodnih i nishodnih puteva kičmenemoždine i u sastav perifernih živaca. Dužina ovakvih aksona može da pređe i 1m.
Goldži II- neuroni sa kratkim aksonima, nalaze se unutar pojedinih delova CNS, gde stvaraju:
1. asocijativne veze (pomocu kojih se povezuju delovi iste strukture CNS)
2. komisuralne veze (koje spajaju 2 simetrične polovine iste nervne strukture), mogu ući i u sastav kratkihprojekcionih veza (koje povezuju razne delove CNS).

by J.M., Ninche, Sandra, Ranagol, gen2010
Akson je izgrađen od aksoleme i od aksoplazme sa neurotubulima i neurofilamentima.
Morfološki i fiziološki gledano akson ima: a) aksonski brežuljak (deo some iz koje izvire akson)
b) inicijalni segment (na kome nastaju akcioni potencijali)
c) aksonsko stablo (koji prevodi akcione potencijale i kojise grana na brojne aksonske kolaterale sa aksonskim završecima).
d) završno grananje (teledendrion-veliko drvo, i tu senalaze sinaptički dugmići tj. neurotransmiteri)
U blizini aksonskog zadebljanja započcinje mijelinski omotač nervnog vlakna- proteinsko- lipidni omotač kojinervnim vlaknima daje karakterističnu belu boju sa sedefastim sjajem. Aksoni beskičmenjaka nisumijelizovani. Od debljine ovog omotača zavisi dijametar nervnih vlakana, a funkcija mijelinskog omotača jeda služi za ograničavanje nervnih impulsa u okviru jednog neurona, sprečavajući stimulaciju susednihparalelnih nervnih vlakana. Prisustvo i izolatorska svojstva mijelinskog omotača obezbeđuju skokovitoprenošenje nervnih impulsa kod kičmenjaka. Mijelinski omotač se na odredjenim razmacima prekidastvarajući Ranvijeova suženja. Aksoni koji se nalaze van CNS-a imaju jos jedan omotač- nurilemu ili Švanovomotač koji se sastoji iz niza ćelija sa po jednim jedrom. Izmedju dva Ranvijeova suženja nalazi se po jednaŠvanova ćelija koja obuhvata po jedan segment mijelinskog omotača. Inače, Švanove ćelije vršemetaboličku funkciju. Pokazano je da tokom embrionalnog i postnatalnog razvića ove ćelije produkujumijelinski omotač. Aksoni CNS-a nemaju Švanov omotač. Funkciju Švanovih ćelija tj. stvaranje mijelina vršiastroglija- vrsta glijalnih ćelija. Akson u blizini svog završnog dela gubi mijelinski omotač i neurilemu. Tuzapočinje grananje nervnog vlakna na veliki broj protoplazmatičnih končića često zadebljanih na svojimkrajevima. Ova zadebljanja se nazivaju terminalni dugmići (čvorići ili završne nožice). Oni stvaraju vezu sasledećom nervnom ćelijom ili sa efektorom. Sa aksonskih završetaka nervni impuls prelazi na sledećujedinicu (na sledeću nervnu ili mišićnu ćeliju) i zato se o aksonu govori kao o eferentnom delu neurona.Proces prenošenja nervnog impulsa sa jednog neurona na drugi ili sa neurona na efektor naziva sesinaptička transmisija.
Centralni nervni sistem čine:
1) neuroni 2) glijalne ćelije 3) krvni sudovi 4) moždana tečnost
Ektodermalnog je porekla, iz 3. klicinog lista. Nervna ćelija, neuron je osnovna genetska,
strukturna i funkcionalna jedinica CNS, stvara se u toku embriogeneze i stalno propadaju.
Rađamo se sa oko 100 biliona nervnih ćelija. Ona se sastoji od: tela nervne ćelije (perikarion ili
soma) i dva tipa nastavaka dendrita i aksona (neurit).
Neuronska tela ulaze u sastav sive mase CNS i perifernih ganglija, dok aksoni ulaze u
sastav bele mase CNS, a takođe čine i periferne živce. Nervne ćelije sa ove dve vrste nastavaka
nazivaju se heteropolarne i one su karakteristične za kičmenjake, a beskičmenjaci obično imaju
neurone sajednim veoma razgranatim aksonskim nastavkom koji se nazivaju unipolarnim.
TELO NEURONA ili soma čini svega oko 10% ukupne mase neurona. Ona je njihov glavni
metabolički centarjer se u njoj nalaze jedro, endoplazmatski retikulum i Goldži aparat, u kojima
se odvija intezivna sinteza proteina. Zbog íntezivne sineteze proteina, hromatin neurona je
relativno razmotan. Osim što je metabolički centar neurona, somaje njegovo receptivno polje jer
se, osim na dendritima i na njoj formira veliki broj sinapsi. Some neurona mogu da budu
organizovane u funkcionalne grupe koje u CNS nazivamo jedrima (nukleusima) a kada su deo
perifernog nervnog sistema zovemo ih ganglijama.
DENDRIT f kratki produžeci nervne ćelije, čine oko l/3 zapremine mozga, nalaze se na raznim

by J.M., Ninche, Sandra, Ranagol, gen2010
delovima nervne ćelije i granaju se u vidu stabla. Na njima se nalaze trnoliki izdanci (spine), koji
povećavaju površinu dendrita i omogućavaju brojne sinapse. Nemaju sposobnost generisanja
akcionog potencijala, njihova osnovna uloga je u prijemu različitih informacija od drugih
neurona. Dendriti su glavno mesto ulaska impulsa u neuron, odnosno predstavljaju njihovo
glavno receptivno polje.
AKSON - je izdanak some. izgrađen je od aksoleme i aksoplazme sa neurotubulima i
neurofilamentima. Morfološki i fiziološki gledano, akson ima: aksonski brežuljak, inicijalni
segment, aksonsko stablo koje se grana na brojne aksonske kolaterale sa aksonskim završecima.
Akson je glavno mesto izlaska informacija, jer neuron uspostavlja sinaptičke komunikacije
uglavnom preko njegovih završetaka.
U blizini aksonskog zadebljanja započinje mijelinski omotač nervnog vlakna - proteinsko
lipidni omotač koji nervnim vlaknima daje karakterističnu belu boju sa sedefastim sjajem.
Aksoni beskičmenjaka nisu mijelizovana. Prisustvo i izolatorska svojstva mijelinskog omotača
obezbeđuju skokovito prenošenje nervnih impulsa kod kičmenjaka. Mijelinski omotač se na
određenim mestima prekida, stvarajući Ranvijerova suženja. Aksoni koji se nalaze van CNS
imaju još jedan omotač, Švanov omotač, koji se sastoji iz niza ćelija sa po jednim jedrom.
Između dva Ranvijerova suženja nalazi se po jedna Švanova ćelija. lnače Švanove ćelije vrše
metaboličku funkciju , tokom embrionalnog i postnatalnog razvića ove ćelije produkuju
mijelinski omotač. Aksoni CNS nemaju Švanov omotač. Funkciju Švanovih ćelija tj. stvaranje
mijelina vrši astroglija, vrsta glijalnih ćelija.
2. Funkcionalni značaj pojedinih delova neurona
Dendriti su kratki nemijelizovani izdaci some, dužine 1-2 milimetara. Ovi nervni produžeci su najdeblji uzsomu. Manje ili više su razgranati i podsećaju na stablo. Čine aferentni spektar neurona. Na njima se nalazetrnoliki izdanci (spine), koji povećavaju površinu dendrita i omogućavaju brojne sinapse. S obzirom na to da uCNS preovlađuju akson-dendritske sinapse, dendriti su glavno mesto ulaska impulsa u neuron, odnosnopredstavljaju njihovo glavno receptivno polje. Dendriti nisu u stanju da proizvode akcione potencijale jerimaju mali broj voltažno-zavisnih natrijumovih kanala, ali ipak mogu da prenose akcione potencijale do some,pritom imaju veliki gubitak. Naime oni proizvode jonske struje dekrementno, što znači da jačina ovih strujaopada sa kvadratom rastojanja. Razlozi su sledeći :
Dendriti imaju mali prečnik i oskudnu zapreminu zbog koje sadržze mali broj jona po jedinici dužine papružaju veliki otpor proticanju jonskih struja. Dekrementno provođenje jonskih struja nastaje i usled velikogpraga dendrita, koga je teško dostići (posledica je tanke membrane, veoma propustljive za jone kalijuma ihlora) i zbog malobrojnih natrijumovih i kalijumovih voltažno-zavisnih kanala u plazma membrani (ćelijskamembrana).
Poremećaji: Suviše kratki dendriti - snižen IQ, ukoliko su dendriti duži nego što bi trebalo - štrče, nadražujuse i slede epileptični napadi.
Soma (neuronsko telo) čini pre svega oko 10% ukupne mase neurona. Ona je njihov glavni metaboličkicentar jer se u njoj nalaze jedro, endoplazmatski retikulum i Goldžijev aparat u kojima se odvija vrlointenzivna sinteza proteina. Zbog intenzivne sinteze proteina, hromatin neurona je relativno razmotan. Somapredstavlja i receptivno polje neurona, jer se i na njoj formira veliki broj sinapsi. Some neurona mogu dabudu organizovane u funkcionalne grupe koje u CNS-u nazivamo nukleusima, a kada su deo PNS-a zovemoih ganglijama.

by J.M., Ninche, Sandra, Ranagol, gen2010
Akson je izdanak some, različite dužine, uglavnom jedan u zavisnosti o kojoj nervnoj ćeliji se radi. Izgrađenje od aksoleme i aksoplazme sa neurotubulima i neurofilamentima. Morfološki i fiziološki, akson ima:aksonski brežuljak, inicijalni segment, aksonsko stablo koje se grana i četvrti deo aksona je teledendrion(razgranato drvo koje ima proširenja sinaptičke dugmiće). Akson je glavno mesto izlaska informacija jerneutron uspostavlja sinaptičke komunikacije uglavnom preko njegovih završetaka. Akson je deblji oddendrita: brže provodi biostruje (jonska struja). Dobar deo aksona poseduje mijelinski omotač koji značajnoubrzava proces biostruje, a velika zastupljenost natrijumovih i kalijumovih voltažno-zavisnih kanalaomogućava aksonima da provode biostruje bez dekrementa, odnosno gubitka jačine.
3. Vrste i značaj aksonskog transporta
1. Ortogradni transport: (od tela nervne ćelije do aksonskog terminala)
a. brzi transport: 300-400 mm/dan, brzim putuju: neurotransmiteri, šećeri, masti, proteini,
enzimi
b. spori transport: 3-4 mm/dan. Sporim putuju: vezikule, mitohondrije.
2. Retrogradni transport: (od aksonskog terminala do tela ćelije)
a. brzim putuju: enzimi, toksini, bakterije
b. sporim putuju ostarele mitohondrije i ostarele vezikule. Ovako se prenosi i besnilo i herpes.
3. Vrste i značaj aksonskog transporta
Aksonski (aksoplazmatski) transport je kretanje materijala kroz aksonski nastavak nervne ćelije. Vrši seneprekidno i izuzetno je značajan za održavanje normalnog funkcionalnog stanja neurona. Sintezamakromolekula neophodnih za celokupnu funkciju neurona se odvija najvećim delom u telu neurona, a zatimse taj materijal raspoređuje na ona mesta gde će biti upotrebljen. Neurotransport se odvija kroz unutrašnjostnervnih vlakana, zahvaljujući snopovima neurofilamenata. Kroz različite neurofilamente istog nervnog vlaknadešava se, veoma intezivan, paralelni transport različitih materija u različitim smerovima. Najintenzivnijitransport odvija se kroz aksone, dok je u dendritima on manje zastupljen i vrlo spor. Na nivou smera u komese obavlja govorimo o anterogradnom neurotransportu koji se odvija od some ka sinaptičkim završecima ili oretrogradnom neurotransportu koji podrazumeva prenos materija od aksonskog završetka do some neurona.

by J.M., Ninche, Sandra, Ranagol, gen2010
Anterogradni transport - od some ka zavrsecima nervne ćelije. Postoje dve vrste ovog transporta: brzi iliaksoplazmatski transport (i do 400mm/24h) u užem smislu (transportuje mitohondrije, vezikule saneuropeptidima i razne strukture proteinske presinaptičke membrane). Spori ili aksoplazmatski protok (sveganekoliko mm/24h) (transportuje molekule aktina, proteine, filamenta i neurotubula).
Retrogradni transport (oko 200mm/24h): kretanje materijala od završetaka ka neuronskom telu (produktidegradacije ostarelih sinaptičkih vezikula, delovi otečene plazma- membrane npr. ostareli proteini i lipidi) iliorganela koji idu na razgradnju i razni neurotrofični faktori, ali ovako se transportuju i neki neurotropni virusi(virus besnila), izazivač meningitisa, tetanusni toksini i ostali štetni agensi. Značaj aksonskog transporta: uneuronsko telo se moze ubrizgati obeleženi protein ili aminokiselina, a može se uneti i obeleženi šećer ilineurotransmiter. Zahvaljujući brzom aksoplazmatskom transportu obeleženi materijal dolazi do aksonskogzavršetka te se na taj način uz upotrebu autoradiografije može odrediti projekcija aksona sve do mesta gdeon uspostavlja kontakt sa sledećom ćelijom. Retrogradni transport se koristi za nalaženje položaja telaneurona onih aksona koji ulaze u sastav pojedinih nervnih puteva. U tu svrhu se često upotrebljava specifičnienzim- enzim peroksidaze rena koji lako uđe u aksonski završetak i transportuje se do tela neurona gde semože lokalizovati pomoću histohemijske tehnike.
Neurotransport se odvija kroz unutrašnjost nervnih vlakana, zahvaljujući snopovíma
neuroñlamenata. Kroz različite neurofilamente istog nervnog vlakna dešava se, veoma
intezivan, paralelni transport različitih materija u različitim smerovima. Najintezivniji transport
odvija se kroz aksone, dokje u dendritima on manje zastupljen i vrlo spor. Na osnovu smera u
kome se obavlja govorimo o
anterogradnom neurotransportu koji se odvija od some ka sinaptičkim završecima ili o
retrogradnom transportu koji podrazumeva prenos materija od aksonskog završetka do some neurona.
ANTEROGRADNI TRANSPORT može biti brz ili spor.
Sporim (svega nekoliko mm/dan) se na primer prenose molekuli aktina, proteini neurofilamenata i
neurotubula, dok se
brzim (i do 400mm/dan) transportuju mitohondrije, vezikule sa neuropeptidima i razni strukturni _proteini
. presinaptičke membrane.
RETROGRADNI TRANSPORT se odvija brzinom od oko 200mm/dan. Ovako se npr.
šalju ka somi produkti degradacije ostarelih sinaptičkih vezikula, delovi oštećene plazma
membrane i organela koji idu na razgradnju i razni neurotrofični faktori. Nažalost ovako se
transportuju i neki neurotropni virusi (besnilo, herpes)
4. Vrste neurona po morfološkim i funkcionalnim karakteristikama
Neuroni ne funcionišu samostalno, već su deo nekog neuronskog kola, koje obrađuje odgovarajućeinformacije, a svako neuronsko kolo je deo nekog sistema koji po funkciji može biti senzorni i motorni.
Neuroni su ekscitabilne ćelije, što znači da pod uticajem primljenih impulsa (koje prethodno integrišu), moguda proizvedu akcione potencijale i da ih preko svojih aksonskih grana, sprovedu do sinapsi posredstvomkojih menjanju ekscitabilnost nekih drugih nervnih receptorskih ili efektorskih ćelija. Treba imati u vidu da seneuroni međusobno značajno razlikuju, po ekscitabilnosti, što je posledica različitog broja raznih jonskihkanala, pumpi, receptora u njihovoj plazma- membrani, kao i neujednačene količine i vrste transmitera kojekoriste. Na osnovu funkcije neurone delimo na:

by J.M., Ninche, Sandra, Ranagol, gen2010
Senzorne (aferentne neurone)- provode akcione potencijale do centra (npr. na kičmenoj moždini i mozgu).Prenošenje informacija sa sekundarne čulne ćelije, senzitivni neuron nastao je od transformirane primarnečulne ćelije (receptorna ćcelija sa izduženim nastavkom).
Motorne (eferentne)- prenose signale od centra ka efektorima. Motoneuron- neuron koji se nadovezuje nasenzitivni ili asocijativni i stupa u kontakt sa efektorom.
Asocijativne, umetnute (interneurone)- preovlađuju u CNS-u a služe u povezivanju funkcije raznih neuronaiste ili suprotne strane tela (npr. povezuju senzorne ili motorne neurone). Osim neurona koji se ekscitirajupod uticajem biostruje primljenih sa nekog drugog neurona, postoje i samookidajući ili pejsmejkeri neuronikoji, zbog spontane jonske propustljivosti plazma membrane, imaju nestabilan membranski potencijalmirovanja, pa se ''spontano aktiviraju'' i periodično emituju te spontano nastale impulse. Praktičan značajpostojanja ovakvih neurona je u tome što daju osnovni ritam neke aktivnosti.
Prema broju nastavaka koji polaze sa tela dele se na:
1.unipolarne
2.bipolarne
3.multipolarne
Unipolarni neuroni imaju samo jedan nastavak i to akson, dok dendriti ne postoje; nalaze se u sluzokoži čulamirisa kičmenjaka. Bipolarni imaju dva nastavka, dendrit i akson, i ima ih u spinalnoj gangliji kičmenjaka.Multipolarni imaju veći broj dendrita i jedan akson; nalaze se u CNS-u kičmenjaka.
Centralni neuroni- svaki neuron CNS-a koji na bilo koji način kordinira funkcije perifernog neurona.
Periferni neuroni- sastavni element svakog refleksnog luka.
Neuroni se prema broju nastavaka koji polaze sa tela dele na :
1. unipolame (pseudounipolarne),
2. bipolarne i
3. multipolame.
Unipolarni neuroni imaju samo jedan nastavak i to akson, dok dendriti ne postoje; nalaze se u
sluzokoži čula mirisa kičmenjaka.
Bipolarni imaju dva nastavka dendrit i akson i ima ih u spinalnoj gangliji kičmenj aka.
Multipolarni imaju veći broj dendrita ijedan akson; nalaze se u CNS-u kičmenjaka.

by J.M., Ninche, Sandra, Ranagol, gen2010
Prema pravcu prenošenja nadražaja razlikuju se tri vrste neurona (po funkcionalnim karakteristikama):
1. senzitivni (aferentni)
2. motorni ( eferentni) `
3. asocijativni. (interneuroni, međuneuroni)
Senzitivni neuroni prenose nadražaj od receptora do odgovarajućih centara u CNS-u, a motorni
prenose nadražaj od centara u CNS-u do efektora. Asocijativni neuroni (umetnuti) se nalaze u
CNS-u i prenose nadražaj od senzitivnih ka motornim neuronima.
Osim neurona koji se ekscitiraju pod uticajem biostruja primljenih sa nekog drugog neurona,
postoje i samookidajući ili pejsmejkerski neuroni koji, zbog spontane jonske propustljivosti
plazma membrane, imaju nestabilan membranski potencijal mirovanja, pa se spontano aktiviraju
i periodično emituju te spontano nastale impulse. Praktičan značaj ovih neurona je što daju
osnovni ritam neke aktivnosti.
Podela po obliku: zvezdasti, okrugli, piramidalni.
Podela po morfologiji: sitni, srednji, krupni.
Postoje neuroni CNS i PNS (perifernog nervnog sistema)
5. Metode ispitivanja neurona
Neuroni još nisu u potpunosti istraženi. Elektrofiziologija- posebna grana fiziologije koja se baviproučavanjem bioelektričnih fenomena.
1. Nervno - mišićni preparat.
Stvaranje nervnih impulsa u nervnim vlaksnima, odnosno u celom nervu, može da se izučava indirektno (toje najjednostavniji način, tj. registrovanjem kontrakcije onog mišića koga inervišu ispitivana nervna vlakna,odnosno živac. Pri dejstvu efikasnog stimulusa, u nervnim vlaknima se stvaraju nervni impulsi koji seprenose na mišić i izazivaju njegovu kontrakciju koja se pomoću adekvatnih metoda može registrovati. Zaoglede ove vrste upotrebljava se izolovani nervno- mišićni preparat iz zadnjeg ekstremiteta žabe (životinjekoje nemaju stalnu telesnu temperaturu, jer za njihovo održavanje u funkcionalnom stanju nisu neophodniposebni uslovi). Kontrakcija mišića koja je izazvana elektrostimulacijom njegovog nerva može se registrovatipomoću mehaničkog sistema. Tetiva izolovanog mišića se veže za jedan kraj pokretne poluge (miografskihpoluga) čiji se drugi zašiljen kraj dovede u kontakt sa aparatom za registrovanje (kimografom), pri svojojkontrakciji mišić povuče miografsku polugu i ona na hartiji ostavlja zapis (miogram). Valjak kimografskogregistratora se okreće oko svoje ose određenom brzinom pomoću elektromotora. Na taj način iz zapisa semože odrediti amplituda pojave kao i njene vremenske karakteristike. U novije vreme se za registracijumišićnih kontrakcija upotrebljavaju mehano električni pretvarači (npr. merne trake). Pošto se mehaničkapojava (mišićna kontrakcija) pretvara u električnu, ona se registruje na višekanalnom pisaču. Na sličan načinse mogu registrovati i druge mehaničke pojave kao sto je: krvni pritisak, promena zapremine grudnog koša,pokreti oka.
2. Izolovani nerv
Stvaranje nervnih impulsa može se registrovati i direktnim merenjem bioelektričnih potencijala, odnosnoakcionih potencijala: U takvim eksperimentima se koriste nervi izolovani iz organizma. Nerv se postavi nadva para metalnih elektroda od kojih prvi par služi za elekrostimulaciju (stimulacione elektrode). Drugi parelektroda služi za registraciju bioelektričnih fenomena (registrujuće ili eksplanatorne elektrode). Stimulacioneelektrode se povezuju sa izvorom elekrične struje (odnosno sa izlaznim stepenom stimulatora), aregistrujuće elektrode se povezuju sa ulaznim stepenom mernog instrumenta.
3. Izolovano nervno vlakno

by J.M., Ninche, Sandra, Ranagol, gen2010
Bioelektrične pojave se često registruju i kod pojedinačnih nervnih vlakana izolovanih iz živaca uz primenuspecifične disekcione tehnike. Pored toga što se debela nervna vlakna relativno lako izdvajaju iz nerva, unjihovu unutrašnjost mogu se uvesti mikroelektrode (staklene kapilarne čestice), pomoću kojih se vršsiintracelularno registrovanje bioelektričnih pojava.
4. Elektrostimulacija
Ako se u eksperimentalne svrhe živci ili pojedinačni aksoni izdvoje iz organizma, oni gube svoje prirodneizvore ekscitacije i na njih se mora primeniti veštački stimulus (mehanički, hemijski). Ipak, najčešće se koristielektrostimulacija (stimulacija električnom strujom) zbog sledecćih razloga.
A) električna struja lako dovodi do ekscitacije tako da količina upotrebljene energije može biti sasvim mala.
B) Ako pravilno dozirana, elektro- stimulacija ne oštećuje tkivo (ne izaziva u njemu neinverzibilne promene).
C) Električna struja je prirodni pratilac stvaranja i prostiranja nervnog impulsa.
Elektrotiziologija je posebna grana fizíologije koja se bavi proučavanjem bioelektričnih
fenomena.
1. Nervno-mišićni preparat - Stvaranje nervnih impulsa u nervnim vlaknima, odnosno u
celom nervu, može da se izučava indirektno. Za oglede ove vrste upotrebljava se izolovani'
nervno-mišićni preparat iz zadnjeg ekstremiteta žabe.
2. Izolovani nerv - Stvaranje nervnih impulsa može se registrovati i direktno merenjem
bioelektričnih potencijala, odnosno akcionih potencijala. U takvim eksperimentima se koriste
nervi izolovani iz organizma. New se postavi na dva para metalnih elektroda od kojih prvi par
služi za elektrostimulaciju, a drugi par elektoda služi za registraciju bioelektričnih fenomena.
Stimulatome elektrode se povezuju sa izvorom električne struje, a registrujuće elektrode se
_povezuju sa ulaznim stepenom mernog instrumenta.
3. Izolovano nervno vlakno - Bioelektrične pojave često se registruju i kod pojedinih
nervnih vlakana izolovanih iz živaca uz primenu specifične disekcione tehnike. Pored toga što se
debela nervna vlakna relativno lako izdvajaju iz nerva, u njihovu unutrašnjost mogu se uvesti
mikroelektrode pomoću kojih se vrši intracelulamoregistrovanje bioelektričnih pojava.
4. Elektrostimulacija - Ako se u eksperimentalne svrhe živci ilipojedinačni aksoni izdvoje
iz organizma, oni gube svoje prirodne izvore ekscitacije i na njih se mora primeniti veštački
stimulus, nejčešće se koristi elektrostimulacija zbog sledećih razloga: _
a) električnastruja lako dovodi do ekscitacije tako da količina energije može biti sasvim mala
b) ako je pravilnodozirana elektro-stimulacija ne oštećuje tkivo.
c) električna strujaje prirodni pratilac stvaranja i prostiranja nervnog impulsa
Katodni osciloskop - Služi za registrovanje i merenje bioelektričnih potencijala.
Pomoću njega se mogu meriti brze promene potencijalne razlike reda veličine mil<ro i mili
sekunde, što je neophodno pri merenju akcionih potencijala. Osnovni deo katodnog osciloskopa
je katodna cev:
O katoda

by J.M., Ninche, Sandra, Ranagol, gen2010
O anoda
O sistem za regulaciju kretanja snopa elektrona u horizontalnom i vertkalnom pravcu
6. Pojam i značaj apoptoze
Apoptoza je genetski programirana fiziološka smrt nervne ćelije. Propadanje neurona po stadijumima; iakovrlo intenzivno transkribuju DNK, veruje se da najveći deo neurona ne može da se replikuje, pa su to jedinećelije organizma koje se značajno ne umnožavaju (izuzetak su olfaktivni neuroni sluznice nosa koji žive 60dana, a obnavljaju se iz bazalnih ćelija). Ako imamo u vidu da se neuroni neprekidno i nepovratno gubeapoptozom, razumljivo je opadanje njihovog broja tokom života. Tako ljudi posle 40 godina života dnevnoizgube izmedju 1000- 300.000 neurona. Tokom rasta i razvoja jedinke, raste i razvija se njen nervni sistemali se to ne dešava na način povećavanja ukupnog broja neurona, jer ove ćelije posle prvih mesecipostnatalnog života ne mogu značajno da se razmnožavaju, već se dalji razvoj nervnog sistema odvija naračun uvećanja veličine postojećih neurona, tj. razgranjavanjem njihovih produžetaka i umnožavanjemsinapsi. (Predstavlja kontrolisan način ćeliskog umiranja, mehanizam za eliminaciju nepoželjnih ćelija)Muškarci imaju veći broj nervnih ćelija od žena, ali su kod žena složenije i tako se to kompenzuje.
Apoptoza se javlja u fiziološkim i patološkim procesima u organizmu. Apoptoza je ograničena na pojedinačnećelije, kada sama ćelija sprovodi programiranu smrt. Apoptoza se sreće kao deo normalnih procesa rasta irazvoja, npr u toku embrionalnog i fetalnog perioda, organogeneze, kada se uklanjaju tkiva i ćelije koja nisupotrebna, a obavila su svoju ulogu. U patološkim slučajevima takođe dolazi do aktiviranja programiranećelijske smrti. Ćelije koje su inficirane virusom izvršavaju ćelijsko samoubistvo. Kada se usled dejstva nekogštetnog faktora kao što su zračenje, infekcija, hipoksija, tumorska mutacija, ćelija ošteti u tolikoj meri da jepopravka nemoguća, a ćelija još živi, pokreće se apoptoza i ovakve ćelije izumiru. To je jedan vidodbrambene reakcije da bi se sprečio nastanak defektnih ćelija koje mogu dovesti do npr. tumora. Uopštenemogućnost ćelije da izvrši apoptozu je jedan od bitnijih faktora u nastanku tumora.
Apoptoza je aktivan proces, za koji je potrebna sinteza enzima i proteina koji učestvuju u njemu. To je proceskoji zahteva utrošak energije, i kod zdrave osobe je uslovljen sa dve vrste gena: geni koji donose odluku kojeneurone treba uništiti i geni koji razvijaju neurone. Programirana ćelijska smrt zapisana je u ćelijskomgenomu i može se aktivirati:
1.dejstvom aktivišućih supstanci koje se vezuju za receptore ć.membrane ili jedra
2.odsustvom faktora rasta, citokina, hormona koji su neophodni za preživljavanje ćelija
3.dejstvom faktora koji izazivaju oštećenje ćelije
Neurotrofini –usporavaju ili produžavaju propadanje neurona
Nekroza – nepovratno propadanje neurona
Apoptoza je homeostazni proces i pomoću njega se stvara sklad integriteta. Procesom apoptozemitohondrije počinju da propadaju na dva načina: spolja i iznutra.
1. Spolja- za receptore na ćeliji vežu se određene materije koje otpočinju propadanje ćelije spolja.
2. Iznutra- realizuje se tako što mitohondrija počinje da reaguje kako ne treba.
Apoptoza je oblik programirane ćelijske smrti koji se javlja u fiziološkim i patološkim
procesima u organizmu. Apoptozaje ograničena na pojedinačne ćelije, kada sama ćelija sprovodi
programiranu smrt. Apoptoza se sreće kao deo normalnih procesa rasta i razvoja, npr. u toku
embrionalnog i fetalnog perioda, organogeneze, kada se uklanjaju tkiva i ćelije koja nisu

by J.M., Ninche, Sandra, Ranagol, gen2010
potrebna, a obavila su svoju ulogu. U patološkim procesima takođe dolazi do aktiviranja
programirane ćelijske smrti. Ćelije koje su inicirane virusom izvršavaju ćelijsko samoubistvo.
Kada se usled dejstva nekog štetnog faktora kao što su zračenje, infekcija, hipoksija, tumorska
mutacija, ćelija" ošteti u tolikoj meri da je popravka nemoguća, a ćelija još živi, pokreće se
apoptoza i ovakve ćelije izumiru. To je jedan vid odbrambene reakcije da bi se sprečio nastanak
defektnih ćelija koje mogu npr. dovesti do tumora. Uopšte nemogućnost ćelije da izvrši apoptozu
je jedan od bitnijih faktora u nastanku tumora.
Apoptoza je aktivan proces, za koji je potrebna sinteza enzima i proteina, koji učestvuju u
njemu. Programirana ćelijska smrtje zapisana u ćelijskom genomu i može se aktivirati:
1. dejstvom aktivišućih supstanci, koje se vezuju za receptore ćelijske membrane ili jedra
2. odsustvom faktora rasta, citokina, hormona koji su neophodni za preživljavanje ćelija
3. dejstvom faktora koji izazivaju oštećenje ćelije.
7. Pojam biološke reakcijeMišići i nervi imaju sposobnost da aktivnim procesom odgovore na promene iz spoljašnje
sredine→nadražljivost (nadražljivost mišića je njihova sposobnost da prime draži). Reakcija živih sistema na
nadražljivost javlja se kao razdraženje ili inhibicija.
Razdraženje: rezultat nekog dejstva na živi sistem.
Fiziološki mir: odsustvo promena ili stimulusa.
Svaka reakcija ima određeni intenzitet i vreme trajanja, kao i vreme "oporavka", da bi nerv mogao ponovo da
reaguje na draž.
Nervne ćelije se ponašaju prema zakonu "sve ili ništa".→ stimulus ispod donjeg praga ne dovodi do reakcije
("ništa"), dok stimulus sa intenzitetom iznad donjeg praga izaziva maksimalno mogući efekat ("sve").
7. Pojam biološke reakcije
Nervni sistem coveka (kao i njegovih evoluitivno blizih i daljnih predaka) obezbedjuje adaptivne reakcije nafaktore sredine; sto je neophodan uslov za odrzavanje integriteta organizma. Zbog toga sto su izlozenetokom citavog zivota promenama, koje se stalno odigravaju kako u spoljasnoj sredini tako i u njima samima,jedinke moraju biti sposobne da na promene sredine reaguju svrsishodno i brzo, tj. adaptivno. Usloznjavanjeorganizma koje se zbivalo tokom evolucije pratio je i proces usloznjavanja sto se jos znacajnije usavrsavanjanjihovih adaptivnih mehanizama. Reakcija organizama na promene sredine moze biti:
1. kompenzatorna, sto ce reci da se suprostavi promenama.
2.Organizam moze da reaguje i svojom akcijama pomocu kojih izaziva promene u sredini svog opstanka. Ireakcije i akcije organizma imaju svoj duboki bioloski smisao koji je adaptivnog karaktera. Obezbedjujuciopstanak organizma u slozenoj i promenljivoj sredini, nervni sistem uslovljava kako reakcije tako i akcijeorganizma koje ako je rec o coveku, ukljucuju i svesno ponasanje. Treba naglasiti da covekove svesne akcijenemaju iskljucivo bioloski smisao. Socijalno ponasanje coveka ima svoje bioloske korene, ali se odvija iprema specificnim zakonima. Reakcija organizma na uslove sredine zavisi od njegove sposobnosti da otkrije(detektuje) promene koje se odigravaju u svakom trenutku. Pored toga, organizam mora da ima i elementa(organe) koji omogucavaju izvrsenje reakcije. Kod viseceliskih organizama funkciju detektora promena usredini imaju specializovane celije koje se nazivaju culne ili receptorne. One na stimuluse reaguju naspecifican nacin, a ishod te specificne reakcije je aktivacija sledece kategorije specijalizovanih celija. One supovezane sa receptorom i to su neuroni. Neuroni u organizmu vrse funkciju provodnika, a posto imajusposobnost da primljene informacije brzo i bez izoblicenja prenesu do izvrsioca reakcije (efektora, a to sumisicne celije- kontrahuju se, ili zlezdane celije- zapocinju lucenje).

by J.M., Ninche, Sandra, Ranagol, gen2010
Nervni sistem čoveka obezbeđuje adaptivne reakcije na faktore sredine što je neophodan
uslov za održavanje integriteta organizma. Pošto su svi organizmi, među njima i čovekov,
izloženi tokom celog života promenama koje se stalno odigravaju kako u spoljašnjoj sredini tako
i u njima samma, jedinke moraju da reaguju svrsishodno i brzo, tj. adaptivno. Mehanizmi koji
obezbeđuju adaptivne reakcije na uslove opstanka ugrađeni su u sva živa bića, od
najprimitivnijih do najsloženijih. Reakcije organizma na promene sredine mogu biti
kompenzatorne, da se suprostavljaju promenama, a takođe organizam može reagovati i svojim
akcijama pomoću kojih izaziva promene u sredini svog opstanka. I reakcije i akcije organizma
imaju svoj duboki biološki smisao koji je adaptivnog karaktera. Reakcija organizma na uslove
sredine zavisi u prvom redu od njegove sposobnosti da detektuje promene koje se odigravaju u
svakom trenutku. Pored toga organizam mora da ima i elemente koji omogućavaju izvršenje
reakcije. Kod višećelijskih organizama to su specijalizovane ćelije koje se nazivaju čulne ili
receptorne ćelije.
8. Draž (karakteristike praga) Draž=stimulus
Draženje=delovanje stimulusa
Nadražaj=akcioni potencijal (ili signal), ono što se prostire u nervnim ćelijama. Nadražaj može
biti i sam draž, kada deluje na drugu nervnu ćeliju.
Draž uvek nosi sa sobom neku energiju. Mogu delovati
direktno (u kontaktu)
distantno (sa daljine)
mislim da bi ovde najbolje bilo za svakoga, da ja lepo koristim pojam stimulus
3 bitne karakteristike draži:
1. intenzitet: do stvaranja nervnog impulsa dolazi samo ako intenzitet dostiže nivo praga, tek tada
se registruje akcioni potencijal.
2. trajanje (tj. vreme delovanja draži)
3. brzina uvođenja: ako brzina uvođenja nije dovoljno velika, onda dolazi do adaptacije. Kada je
vreme uvođenja spora, onda su istovremeno otvoreni i Na i Ka kanali. Ka joni izlaze, Na joni
ulaze istovremeno, i zbog toga se ne može postići onih kritičnih 25 mV→ ne formira se akcioni
potencijal→ → i ovo se naziva akomodacija prema tome, akomodacija se dešava, kada se draž
uvodi jako sporo.
Mora da postoji određeni odnos između intenziteta i trajanja, da bi došlo da nadražaja. Ako je
intenzitet preveliki, a vreme delovanja stimulusa premali, neće doći do nadražaja.

by J.M., Ninche, Sandra, Ranagol, gen2010
Stimulus ili draž može biti bilo koja promena u sredini. Draž utiče na receptore (na odgovarajući receptor).
Vrste draži: fizičke, hemijske, mehaničke, termijske...
Donji prag draži: je minimalan intenzitet, koji je potreban, da bi receptor registrovao draž.
Gornji prag draži: je onaj prag, preko čega već receptor ne može da registruje draž.
Diferencijalni prag: je potrebna promena, da bi se osetila razlika u draži (u njenom intenzitetu).
8. Draž (karakteristike praga)
Prema naopštijoj definiciji, stimulus (nadražaj ili draž) je bilo koja promena spoljašnje sredine u kojoj senalazi razdražljivo tkivo, odnosno tkivo sposobno da reaguje na tu promenu. Svaka promena spoljašnjesredine ne mora da bude efikasna.
I Da bi tkivo reagovalo, stimulus mora biti dovoljnog intenziteta, on mora da postigne određeni nivo.Minimalan intenzitet dovoljan da dovede do reakcije je prag nadražaja. Nadražaj čiji je intenzitet manji odpraga su potporni (subliminarni) i nisu efikasni.
II Drugi uslov koji promenu sredine čini efikasnom jeste da se promena odigra dovoljno brzo i da trajedovoljno dugo. Suviše spore i preterano kratkotrajne promene ne mogu imati karakter efikasnog stimulusa.
Draž - spoljašnja i objektivno postojeća energija koja deluje na naša čula. Postoje razne vrste draži: fizičke ihemijske, mehaničke, termičke. Mišići i nervi imaju sposobnost da odgovaraju aktivnim procesom napromene iz spoljašnje sredine i ta osobina se naziva razdražljivost. Reakcija na nadražaj moze da se završiili razdraženjem ili inhibicijom (aktivnim kočenjem specifične funkcije). Kod živih bica potrebno je da sediferenciraju 3 nivoa stanja funkcionalne aktivnosti.
1. Fiziološki mir (odsustvo manifestnih pojava, specifične pojave, kretanja)
2. Proces razdraženja (javlja se kao rezultat nekog dejstva na živu tvorevinu)
3. Proces inhibicije (aktivan proces koji se ispoljava u slabljenju ili prestanku specifičnih funkcionalnihmanifestacija).
Korisno vreme je određeno vreme dejstva nadražaja (vreme za koje se vrši reobaza), duže od hronaksije.Reobaza je minimum intenziteta nadražaja koji je potrebno zadovoljiti kako bi došlo do akcionog potencijala.Hronaksija je najmanje vreme koje je neophodno kako bi došlo do procesa razdražaja i pri dejstvu dvostrukihreobaza, uslovljava formiranje akcionog potencijala. Akomodacija je pojava da se razdražljiva tkivaprilagođavaju na nadražaj koji se sporo uvećava (neurofiziološka osnova za akomodaciju su promene navoltažno- zavisnim kanalima). Vrlo često na organizam deluje serija nadražaja, bilo da deluju istovremeno,bilo da slede jedna drugu, gde svaka izaziva određeni odgovor. Da bi se sprovele reakcije, organizam jedužan diferencirati nadražaje i to ne samo po njihovom kvalitetu, vec i po intenzitetu i dužini dejstva. To sedefiniše kao diferencijalni prag - minimalna razlika intenziteta ili dužine dejstva između dva nadražaja, koja jedovoljna da izazove novi nivo aktivnosti tkiva, organa.
Zakon sve ili ništa - Do stvaranja nervnog impulsa dolazi samo u slučaju kada intenzitet impulsa dostignevrednost praga, tek tada se može registrovati akcioni potencijal. Bodvic je utvrdio da postoji zakoniti odnosizmeđu reakcije ekscitabilnog tkiva i intenziteta stimulusa. Na stimulus čiji je intenzitet sublimiran tkivo nereaguje (tj. ne dolazi do kontrakcije), dok na stimulus na nivou praga tkivo reaguje maksimalno i svakopovećanje jačine stimulusa ne izaziva veću kontrakciju mišića. Bodvic je formulisao zakon koji je nazvao sveili ništa. Tvrdio je da je on primenljiv na sva ostala ekscitabilna tkiva (on je eksperimentisao sa srčanimmišićem), što se pokazalo kao tacno. Prema ovom zakonu subliminalni stimulusi ne dovode do reakcijeekscitabilnog tkiva - ništa, dok stimulusi na nivou praga izazivaju maksimalno mogući efekat - sve. Amplitudaakcionog potencijala ne povećava se sa povećanjem intenziteta stimulusa. Stimulus intenziteta ispod pragane izaziva akcioni potencijal, dok stimulus na nivou praga izaziva akcioni potencijal maksimalne amplitude.Na taj način stimulus čiji je intenzitet znatno iznad praga ne dovodi do povećanja amplitude akcionogpotencijala niti se menja brzina kojom se nervni impuls prostire kroz akson.
Nervni impuls se prostire kroz nervno vlakno bez dekrementa.

by J.M., Ninche, Sandra, Ranagol, gen2010
Prema najopštijoj definiciji stimulus ili nadražaj (draž) je bilo koja promena spoljašnje
sredine u kojoj se nalazi razdražljivo (ekscitabilno) tkivo, odnosno tkivo sposobno da reaguje na
tu promenu. Svaka promena spoljašnje sredine ne mora da bude efikasna.
1. Da bi tkivo reagovalo, stimulus mora da bude dovoljnog inteziteta, on mora da postigne određeni nivo.
Minimalni intezitet stimulusa dovoljan da dovede do reakcije je prag nadražaja. Nadražaji čiji je intezitet
manji od praga su potražni (sublimirani) i nisu efikasni.
2. Drugi uslov koji promenu sredine čini
efikasnom jeste da se promena odigra dovoljno brzo i
3. da traje dovoljno dugo. Suviše spore i
preterano kratkotrajne promene ne mogu imati karakter efikasnog stimulusa.
Draž je spoljašnja objektivno postojeća energija koja deluje na naša čula. Postoje razne
vrste draži: fizičke, hemijske, mehaničke, termičke...
Kod živih organizama potrebno je da se diferenciraju 3 nivoa stanja funkcionalne
aktivnosti:
1. fiziološki mir (odsustvo manifestnih pojava specifične aktivnosti, kretanja, sekrecije),
2. proces razdraženja (javlja se kao rezultat nekog dejstva na živu tvorevinu) i
3. proces inhibicije (aktivan proces koji se ispoljava u slabljenju ili prestanku
specifičnih fnkcionalnih manifestacija).
Korisno vreme (KV): draž ako deluje
Reobaza je minimalni intezitet draži koji da bi došlo do akcionog potencijala.
Hronaksija je najmanje vreme kojeje neophodno da bi se javio proces razdraženja i pri
dejstvu dvostrukih reobaza, uslovljava formiranje akcionog potencijala.
Akomodacija je pojava gde se razdražljiva tkiva prilagođavaju na nadražaj koji se sporo
uvećava.
9. Građa ćelijske membrane Svi neuroni međusobno komuniciraju
Akcioni potencijal: bioelektrična promena, može da se prostire i da deluje na neke druge ćelije
(nerv, mišić, žlezda...)
neuroni mogu biti unipolarni, bipolarni, multipolarni
nervna ćelija ima svoj omotač - ćelijsku membranu
nervna ćelija je ŽENSKOG RODA
Neurolema: omotač nervne ćelije, ćelijska memrana
Tri sloja→ 2 sloja masti

by J.M., Ninche, Sandra, Ranagol, gen2010
1 sloj proteina
Proteini imaju funkciju kanala, nosača, pumpe. Mogu biti: integralni ili periferni.
U ćelijskoj membrani su ukomponovani specijalizovane molekule, uglavnom proteinske prirode
(jonski kanali i receptori za neurotransmitere) i sa njima povezani enzimi.
Proteini koji dopiru u ekstracelularni prostor obezbeđuju specifična mesta "prepoznavanja"
(receptorski proteini). Od njih zavisi intercelularna komunikacija pomoću neurotransmitera i
hormona.
Ćelijska membrana se stalno obnavlja na račun novosintetizovanih supstanci u ćeliji.
Membrana je selektivno propustljiva i kroz nju se vrši razmena materijala i elektrolita između
spoljašnje sredine i ćelije, posebno prolaskom kroz jonske kanale.
Jonski kanali se dele na:
o Stalno otvorene - stalni transport je omogućen
o Regulisane
Proteini imaju funkciju kanala, nosača, pumpe. Proteini mogu biti integralni ili periferni.
Proteini koji ulaze u ekstracelularni prostor obezbeđuju specifična mesta "prepoznavanja"
(receptorski proteini). Od njih zavisi intercelularna komunikacija pomoću neurotransmitera i
hormona.
Ćelijska membrana je selektivno propustljiva. Kroz nju se vrši razmena materijala i elektrolita između
spoljašnje sredine i ćelije, posebno prolaskom kroz jonske kanale.
Jonski kanali se dele na:
o stalno otvoreni jonski kanali - omogućen je stalni transport
o regulisani jonski kanali
regulisane ligantima
zavisni od mehaničkog dejstva
9. Građa ćelijske membrane
Ćelijska membrana je selektivno propustljiva dinamička struktura koju sačinjavaju 2 komponente. Prva,osnovna komponenta se sastoji iz fosfolipidnog dvosloja u kojem su locirane hidrofobne lipidne grupe i onesu povezane sa hidrofilnim fosfatnim grupama. Tu se nalaze specijalizovani molekuli, uglavnom proteinskeprirode, kao što su jonski kanali receptori za neurotransmitere i sa njima povezani enzimi. Proteini koji dopruu ekstracelularni prostor obezbeđuju specifična mesta prepoznavanja (receptorski proteini) od kojih zavisi iintercelularna komunikacija pomoću neurotransmitera i hormona. Membrana se stalno obnavlja na računnovosintetisanih supstanci u ćeliji. Kroz ćelijsku membranu vrši se razmena materijala i elektrolita izmeđuspoljašnje sredine i ćelije posebno prolaskom kroz jonske kanale.
Na unutrašnju stranu membrane nadovezuju se specifične končaste proteinske strukture koje činemembranske skeletne proteine u koje spada i sinapsin, čija je uloga intercelularno prepoznavanje isignalizacija, kao i transport. Proteini se dele na lipo i gliko-proteine. Oni se u odnosu na položaj i funkcijudele na strukturne, na one koji su funkcionalni kao pumpe, kanali i receptori.

by J.M., Ninche, Sandra, Ranagol, gen2010
Ćelijska membrane je lipoproteinska struktura koja je sačinjena od dva lipidna i jednog belančevinastogsloja. Belančevine su strukturne ili periferne, i one koje učestvuju u građi mogu imati funkciju kanala, pumpeili nosača enzima. Oko ćelijske membrane nalazi se omotač neurolema.
Lipidne dvosloj se ne meša sa ekstra-intra celularnom tečnošću. On obrazuje barijeru za kretanje molekulavode i hidrosolubilnih supstanci između ekstra i intra celularne tečnosti. Mali broj supstanci može proći krozlipidni dvosloj, a sa druge strane molekuli proteina imaju sasvim drugačija transportna svojstva, oni stvarajualternativni put kroz ćelijsku membranu.
Ćelijska membrana je dinamička struktura koju sačinjavaju dve komponente. Prva,
osnovna komponenta sastoji se od lipidnog dvosloja, tu se nalaze specijalizovani molekuli,
uglavnom proteinske prirode, kao što su jonski kanali i receptori za neurotransmitere i sa njima
povezani enzimi. Proteini koji dopiru u ekstracelularni prostor obezbeđuju specifična mesta
prepoznavanja od kojih zavisi i intracelularna komunikacija pomoću neurotransmitera i
hormona. Membrana se stalno obnavlja na račun novosintetizovanih supstancija u ćeliji. Kroz
ćelijsku membranu vrši se razmena materijala i elektrolita između spoljašnje sredine i ćelije,
posebno prolaskom krozjonske kanale.
Na unutrašnju stranu membrane nadovezuju se specifične končaste proteinske strukture
koje čine membranske skeletne proteine u koje spada i sinapsin.
Razmena supstanci sa okolinom
Ćelijska membrana je selektivno propustljiva što ćeliji omogućava da kontroliše razmenu supstanci i takoštiti stalnost sastava unutrašnje sredine.
Postoje dva osnovna načina kojima ćelija obavlja razmenu sa okolnom sredinom:
1. transport malih molekula kroz membranu (kroz kanale u proteinima ili pomoću proteina nosača, kojise ne može uočiti pod mikroskopom);
2. citoza - unošenje i izbacivanje krupnih molekula u čemu sama membrana aktivno učestvuje, što semože uočiti pod mikroskopom; procesi se nazivaju endocitoza i egzocitoza.
[uredi] Transport molekula kroz membranu
Transport materija kroz membranu (A-F), endocitoza i egzocitozaTransport kroz membranu može biti:

by J.M., Ninche, Sandra, Ranagol, gen2010
aktivan i
pasivan.
[uredi] Pasivan transport
Pri pasivnom transportu molekuli se kreću kroz membranu zahvaljujući razlici u koncentraciji sa jedne i drugestrane membrane, odnosno iz sredine sa većom u sredinu sa manjom koncentracijom (niz hemijskigradijent), pri čemu se energija ne troši. Oblici pasivnog transporta su:
osmoza,
difuzija i
olakšana difuzija.
Difuzija je transport molekula kroz membranu iz sredine sa većom u sredinu sa manjomkoncentracijom sve dok se koncentracije ne izjednače. Supstance rastvorljive u lipidima čiji sumolekuli mali prolaze brzo kroz membranu slobodnom (prostom) difuzijom (tako se prenose gasoviO2 i CO2 u plućima i tkivima). Supstance koje nisu rastvorljive u lipidima prolaze procesom olakšanedifuzije.
Olakšana difuzija je transport supstanci koje se ne rastvaraju u lipidima, niz hemijski gradijent, alipomoću molekula nosača (zato se naziva olakšana difuzija). Molekul nosač je protein koji prolazikroz lipidni dvosloj i sadrži mesto za koje se molekul vezuje. Olakšanom difuzijom se transportujeglukoza. Ovaj način transporta regulišu hormoni. Na primer olakšanu difuziju glukoze regulišeinsulin.
Osmoza je difuzija vode kroz membranu. Molekuli vode se kreću iz sredine sa manjomkoncentracijom (tu je više vode) u sredinu sa većom koncentracijom rastvorenih supstanci (tu jevode manje). Pritisak koji je potreban da bi se sprečila osmoza je osmotski pritisak. Osmotskipritisak je veći što je koncentracija rastvora veća i obrnuto. Ako se ćelije nalaze u rastvoru, voda ćetežiti da prođe kroz ćelijsku membranu u cilju da se izjednače količine vode sa obe stranemembrane. Tako, ako ćeliju stavimo u hipotoničan rastvor (rastvor koji je manje koncentracije uodnos na samu ćeliju) doći će do ulaženja vode u ćeliju, do njenog bubrenja i prskanja. Uhipertoničnom rastvoru (koncentrovaniji od rastvora u samoj ćeliji), voda će izlaziti iz ćelije i ona ćese smežurati. Zbog toga je veoma važno da se održava stalan osmotski pritisak, odnosno količinavode u organizmu što se naziva osmoregulacija. Kod sisara se osmoregulacija vrši preko bubrega –višak vode iz krvi prelazi u mokraću.
[uredi] Aktivan transport
Aktivan transport se vrši nasuprot hemijskom gradijentu – supstance se transportuju iz sredine sa manjom usredinu sa većom koncentracijom, pomoću proteina nosača i uz utrošak energije.
Energija za odvijanje aktivnog transporta dobija se hidrolizom ATP-a u ADP. Proteini nosači rade kao pumpei kao enzimi ATP-aze (adenozin trifosfataze) jer katalizuju razlaganje ATP-a. Najbolje proučena je Na,Kpumpa koja nasuprot hemijskom gradijentu, aktivno ispumpava Na+ iz ćelije, a upumpava К+ u ćeliju. (Na+ jevanćelijski jon – ima ga više van, nego u ćeliji; К+ je obrnuto – unutarćelijski jon).
Natrijum-kalijum pumpa ima glavnu ulogu u stvaranju membranskog potencijala životinjskih ćelija.Membranski potencijal ili potencijal mirovanja je električni napon koji postoji između unutrašnje (negativnonaelektrisane) i spoljašnje stane (pozitivno naelektrisane) membrane nervne i mišićne ćelije u stanjumirovanja, odnosno pre njihovog nadraživanja. Kada se ove ćelije nadraže, onda potencijal mirovanjapostaje akcioni potencijal – nervi impuls, pri kome se obrće polarizovanost membrane tako da unutrašnjastrana postaje pozitivno, a spoljašnja negativno naelektrisana.)
[uredi] Endocitoza i egzocitoza
Makromolekuli kao što su proteini, polisaharidi, polinukleotidi ili čak čitave ćelije (npr. bakterije) ne moguprolaziti kroz membranu ni jednim od navedenih tipova transporta. Unošenje i izlučivanje velikih molekulaobavlja se aktivnim učešćem membrane pri čemu ona obrazuje vezikule, a procesi se nazivaju:

by J.M., Ninche, Sandra, Ranagol, gen2010
endocitoza i
egzocitoza.
Proces unošenja makromolekula i rastvora naziva se endocitoza, dok se izbacivanje specifičnih proizvodaćelije ili nekih drugih materijala u vanćelijsku sredinu naziva egzocitoza.
Endocitoza
Pri endocitozi dolazi prvo do udubljenja na membrani u koje ulazi supstanca koja treba da se unese u ćeliju.Udubljenje se sve više spušta u unutrašnjost ćelije, a zatim se ivice membrane spoje i oko unete materije seobrazuje vezikula (videti:lizozom). Endocitoza obuhvata dva procesa:
fagocitozu - unošenje krupnih čestica (grč. phagein = jesti) i
pinocitozu - unošenje rastvorenih materija (grč. pino = piti).
Dok skoro sve vrste ćelija neprekidno unose tečnosti i rastvorene molekule pinocitozom, fagocitozu mogu davrše samo posebne ćelije – fagocite (npr. leukociti vrše fagocitozu bakterija).
Egzocitoza
Procesom egzocitoze u vanćelijsku sredinu dospevaju supstance koje ćelija treba da odstrani (suprotnoendocitozi). Takvi su hormoni i enzimi čija se funkcija ispoljava izvan ćelije koja ih je sintetisala, kao što npr.ćelije pankreasa izlučuju insulin. Oko supstance, koja treba da se izbaci van ćelije, se obrazuje vezikula.Vezikula se kreće ka površini ćelije, njena membrana se ujedinjuje sa ćelijskom membranom, a sadržajvezikule biva izbačen.
Smatra se da su mnogim vrstama ćelija endocitoza i egzocitoza međusobno povezani procesi i da na tajnačin ugrađivanje membrane vezikule ne dovodi do povećanja površine ćelijske membrane. Vezikulepredstavljaju transportni sistem i u samoj ćeliji tako što se prenose između različitih organela.
10. Transportni sistemi ćelijske membrane (kanali, receptori) Ka jon voli fiziološki mir i spokoj. Ka jon može lako da prolazi kroz ćelijsku membranu (ć.m. voli
Ka jon, a ne voli Na jon :) )
Na jon privlači molekule vode, i zato ona ne može da prolazi kroz ć.m..
ćelijska molekula je lipoproteinska struktura, to znači da ć.m. ima dva lipidna i jedan proteinski
sloj
integralni proteini: funkcija kanala, funkcija pumpe ili nosači pumpe.
integralni proteini mogu biti kanali.
ć.m. je dinamička struktura, jer se neprekidno formira i neprekidno menja
kanali se razlikuju, nazive dobijaju na osnovu toga šta nose
stalno otvoreni kanali se nalaze u integralnim proteinima, svi kanali su u integralnim proteinima
kanal može da se otvara promenom napona ili npr. ligant-vezivanjem.
prolaz kroz lipidni deo zavisi od rastvorljivosti u lipidima (mastima). Npr. alkohol je rastvorljiv u
mastima, zato alkohol prolazi kroz masti (lipide)

by J.M., Ninche, Sandra, Ranagol, gen2010
voda isto prolazi kroz ć.m..
Ćelijska membrana je selektivno propustljiva. Kroz nju se vrši razmena materijala i elektrolita između
spoljašnje sredine i ćelije, posebno prolaskom kroz jonske kanale.
Jonski kanali se dele na:
o stalno otvoreni jonski kanali - omogućen je stalni transport
o regulisani jonski kanali
regulisane ligantima
zavisni od mehaničkog dejstva
Uslovi za prolazak materijala: rastvorljivost u mastima (izuzetak je voda! Njene molekule su dovoljne
male.)
Ćelijska membrana u stanju mirovanja propušta K jone, a ne propušta druge jone.

by J.M., Ninche, Sandra, Ranagol, gen2010
Jonski kanali su transmembranske proteinske tvorevine od integralníh proteina koji u
potpunosti probijaju membranu povezujući njenu intra i ekstracelulamu površinu. Jonski kanali
uglavnom služe za difuziju jedne određene vrste jona, ali ima i onih koji nisu tako strogo
selektivni već propuštaju više različitih vrstajona. Svi jonski kanali propuštaju i vodu.
Podela prema vrsti jona čiju difuziju omogućavaju:
1. Natrijumovi kanali - su transmembranski proteini čije subjedinice u fosfolipidnom dvosloju
grade pore kroz koje mogu difundovati hidratisani joni Na. U vreme električnog mirovanja
plazma membrane Na difunduje kroz svoje kanale iz ekstracelulame sredine ka unutrašnjosti
ćelije, zahvaljujući jako izraženom hemijskom i električnom gradijentu. Negativno naelektrisanje
koje dominira uz unutrašnju površinu membrane, privlači katjone Na i tako dodatno olakšava
njihovu difuziju u ćeliji.
2. Kalijmovi kanali - su transmembranski proteini, organizovani tako da formiraju centralnu
poru kroz koju mogu proći kalijumovi joni. U vreme električnog mirovanja kalijum difunduje iz
ćelija kroz tzv. Pasivne kalijumove kanale, na bazi povoljnog koncetracijskog gradijenta, ali je ta
difuzija spora jer višak pozitivnog naelektrisanja koja tada dominira uz spoljašnju površinu
membrane odbija K katjone - nepovoljan koncetracij ski gradijent.
3. Hloridni kanali - su jedini poznati anjonski kanali. Imaju pozitivno naelektrisanu
unutrašnjost kojom privlače negativno naelektrisane hloride da difunduju iz međućelijske
tečnosti ka unutrašnjosti ćelija. Bez obzira na povoljan koneetracijski gradijent, difuzija hlorida
je spora u vreme električnog mirovanja plazma membrane, jer tada na unutrašnjoj strani
membrane dominira negativno naelektrisanje koje odbija negativno naelektrisane hloride.
Podela prema načinu regulacije progustljivosti:
1. Pasivni (ili kontinuirano otvoreni jonski kanali) - su spontano otvoreni tako da omogućavaju
kontinuiranu difuziju odgovarajućih jona kroz membranu. U plazma membrani većine ćelija
postoje pasivni kanali za K koji obezbeđuju najveći deo vrednosti membranskog potencijala
mirovanja, tako što kroz njih K sporo idifunduje iz ćelije ka ekstraeelularnoj sredini, pa ove
kanale nazivamo sporo curećim.
2. Aktivni (ili regulisani) jonski kanali - imaju strogo kontrolisanu prolaznost, jer se njihova vrata otvaraju i
zatvaraju samo na odgovarajući signal. Prisustvo različitih vrsta aktivnih kanala u ćelijskim
membranama uspostavlja pojavu selektivne preambilnosti, koja je različita u odnosu na vreme i
na vrstu jona. Na osnovu načina regulisanja prolaznosti aktivni jonski kanali mogu biti:

by J.M., Ninche, Sandra, Ranagol, gen2010
a) Ligand - zavisni, otvaraju se vezivanjem specifične hemijske materije - liganda za
odgovarajući receptor.
b) Voltažno ili potencijal - zavisni jonski kanali imaju voltažne senzore koji se aktiviraju pri
tačno određenoj vrednosti potencijala membrane. Ovi kanali su isto proteini. Mogu menjati svoj oblik→ tj. mogu da se
konformaciono promene→vrata se menjaju
c) Kanali zavisni od mehaničke sile - su mehanosenzitivni, otvaraju se pod uticajem neke
mehaničke sile.
11. Vrste transporta kroz ćelijsku membranuPostoje dve osnovne vrste: pasivan i aktivan transport. Pri pasivnom transportu materije
se kreću iz sredine sa većom u sredinu sa manjom koncentracijom (niz hemijski gradijent), pri
čemu se energija ne troši. Oblici pasivnog transporta su osmoza, difuzija i olakšana difuzija.
1. Difuzija (prosta difuzija) je transport materija kroz membranu iz sredine sa većom u sredinu sa manjom
koncentracijom sve dok se koncentracije ne izjednače. Materije rastvorljive u lipidima i malog
molekula prolaze brzo kroz membranu slobodnom (prostom) difuzijom (tako se prenose gasovi
02 i CO2 u plućima i tkivima). Materije koje nisu rastvorljive u lipidima prolaze procesom
olakšane difuzije.
materija se kreće sa mesta veće koncetracije na mesto manje koncetracije
ovo je u skladu sa koncetracionim gradijentom
brzina difuzije zavisi od razlike koncetracija sa jedne i druge strane membrane

by J.M., Ninche, Sandra, Ranagol, gen2010
2. Olakšana difuzija je transport materija, koje se ne rastvaraju u lipidima, niz hemijski
gradijent, ali pomoću molekula nosača (zato se naziva olakšana difuzija). Molekul nosač je
protein koji prolazi kroz lipidni dvosloj i sadrži mesto za koje se materija vezuje. Olakšanom
difuzijom se transportuje glukoza. Ovaj način transporta regulišu hormoni pa tako olakšanu
difuziju glukoza reguliše insulin.
Olakšana difuzija se odvija uz pomoć nosača, kroz kanale
nosač vezuje ono što treba da bude transportovano, i tako prenosi
za ovo se koriti energija ( a za to je izvor ATP: adenozin-tri-fosfat)
ako nosaš ne dobije energiju, onda je to olakšana difuzija??????????????????????????
kanali se neprekidno menjaju
3. Osmoza je difuzija vode kroz membranu. Molekuli vode se kreću iz sredine sa manjom
koncentracijom (tu je više vode) u sredinu sa većom koncentracijom rastvorenih materija (tu je
vode manje). Pritisak koji je potreban da bi se sprečila osmoza je osmotski pritisak. Osmotski
pritisakje veći štoje koncentracija rastvora veća i obrnuto. Ako se ćelije nalaze u rastvoru, voda
će težiti da prođe kroz ćelijsku membranu u cilju da se izjednače količine vode sa obe strane
membrane. Postoji osmoska membrana.
Osmoza je kretanje rastvarača sa mesta manje koncetracije na mesto veće koncetracije. Cilj
ovakvog kretanja je da se izjednači pritisak sa obe strane membrane.
nosači mogu biti
1. uniport: vezuje uvek istu supstancu, ili jednu molekulu i prenosi je kroz ćelijsku membranu
uvek u jednom pravcu.
2. simport: učestvuje u mehanizmima ko-transporta (što znači 2 ili više vrsta materije se
prenose.
neophodno je da se vežu 2 materije za njega unutra, da bi se prenelo sa spoljne
strane ćelije
nosi 2 ili više supstance u istom smeru, uvek sa mesta manje koncetracije na mesto
veće koncetracije.

by J.M., Ninche, Sandra, Ranagol, gen2010
troši ATP.
3. antiport: vezuje najmanje 2 supstance (prvi u ekstracelularnom prostoru, a drugi unutar
ćelijskom prostoru)
tad ostvaruje mehanizme kontra transporta (prenosi molekule jedne u jednom
pravcu, a druge u drugom pravcu, uvek nasuprot koncetracionom gradijentu. Dakle:
kontratransport je transport u oba smera.
Aktivan transport se vrši kroz ćelijsku membranu nasuprot hemijskom gradijentu
materije se transportuju iz sredine sa manjom .u sredinu sa većom koncentracijom, pomoću
proteina nosača i uz utrošak energije. Energija za odvijanje aktivnog transporta dobija se
hidrolizom ATP-a u ADP. Proteini nosači rade kao pumpe i kao enzimi ATP - aze (adenozin
trifosfataze) jer katalizuju razlaganje ATP -a.
Najbolje proučena je Na - K pumpa koja nasuprot
hemijskom gradijentu, aktivno ispumpava jone Na iz ćelije, a upumpava jone K+ u ćeliju.
KaNa pumpa vezuje 3 jona Na i 2 jona Ka. Ovako Na jon izlazi napolje iz ćelije, a Ka joni ulaze u
ćeliju. Na joni privlače molekule vode→ i ovako se čuva volumen ćelije
Natrijum-kalijum pumpa ima glavnu ulogu u stvaranju membranskog potencijala životinjskih
ćelija. Slika dole: NaKa pumpa
Aktivan transport sa mesta MANJE koncetracije na mesto VEĆE koncetracije, uz pomoć
nosača, nosači su uvek proteini. Aktivan transport je uvek nasuprot koncetracionom gradijentu
Uvek zahteva energiju. Za to se koristi ATP.
Uvek postoji kompetencija za nosača (ko prvi, njemu devojka)

by J.M., Ninche, Sandra, Ranagol, gen2010
Ovaj transport može biti:
o primarni aktivni transport: koristi ATP. Primer za ovo je K+Na+ pumpa. Uvek se troši
energija kod aktivnog primarnog transporta.
o sekundarni aktivni transport: koristi energiju, koja se nagomilava mehanizmima
primarnog aktivnog transporta
11. Vrste transporta kroz ćelijsku membranu
Kroz membranu se sprovodi selektivni transport materija koji može biti: pasivni i aktivni.
Pasivni transport kroz membranu u ćeliju ili iz nje u spoljašnjost se sprovodi bez potrošnje energije. Osnovnioblici pasivnog transporta su osmoza, difuzija i olakšana difuzija. Osmoza je difuzija vode kroz membranu.Molekuli vode se kreću iz sredine sa manjom koncentracijom (tu je više vode) u sredinu sa većomkoncentracijom rastvorenih materija (tu je manje vode). Pritisak koji je potreban da bi se sprečila osmoza jeosmotski pritisak i on je veći što je koncentracija rastvora veća i obrnuto. Ako se ćelije nalaze u rastvoru vodaće težiti da prođe kroz ćelijsku membranu u cilju da se izjednači količina vode sa obe strane membrane.Osnovni mehanizam koji obezbeđuje prodiranje materije u ćeliju je difuzija. Ona je podeljena u dva posebnaciklusa, a to su jednostavna i olakšana difuzija. Jednostavna difuzija se odvija kroz međuprostore lipidnogdvosloja, pri čemu nije potrebno da ono što treba da bude transportovano bude vezano za proteinskenosače. Jednostavna difuzija se odvija sa mesta veće ka mestu menje koncentracije (niz hemijski gradijent).Olakšana difuzija zahteva interakciju molekula ili jona sa proteinskim nosačima koji pomažu njihovoprolaženje kroz membranu. Ona se odvija kroz kanale, a nosači se zovu uniport nosači jer prenose materijesamo u jednom pravcu.
Nosači:
1) Uniport- nosi 1 supstancu
2) Simport- nosi istovremeno bar 2 supstance u istom smeru, učestvuje u mehanizmu kotransporta
3) Antiport- nosi supstance u suprotnim pravcima
Olakšana difuzija se odvija u pravcu gradijenta. Olakšanom difuzijom se transportuje glukoza i ovaj načintransporta regulišu hormoni pa tako olakšanu difuziju glukoze reguliše insulin.
Aktivni transport (Na K pumpa)- transport sa mesta male koncentracije na mesto veće koncentracije(odnosno, protiv koncentraciskog gradijenta) Ovaj transportni sistem se odvija zahvaljujući energiji ATP izavisi od prenosa materije pomoću proteinskih nosača Na-K pumpa neprekidno odražava razlikukoncentracije Na i K sa jedne i druge strane membrane kao i uspostavljanje negativnog električnogpotencijala unutar ćelije.
Odstranjenje Na iz unutrašnjosti ćelije vrši se pomoću Na pumpe koja je jedan od različitih vidova aktivnogtransporta. Aktivnost natrijumske pumpe obezbeđuje nisku koncentraciju jona Na u ćeliji i njihovokoncentrisanje sa spoljašnje strane. Smatra se da se aktivni transport Na vrši pomoću enzima ATP-aza, kojiulazi u sastav membrane nerve ćelije Na unutrašnjoj strani membrane enzim vezuje Na⁺ a zatim ga prenosina njenu spoljašnju stranu i tu ga otpušta. Ova pumpa- molekul, u isto vreme na spoljašnjoj strani membranevezuje K⁺ jone suprotno od koncentracionog gradijentu, vraća ih u unutrašnjost ćelije, opet pomoću aktivnogtransporta. To bi pored ostalih faktora takođe doprinosilo odražavanju visoke koncentracije K⁺ u aksoplazmi.Zato se enzim pomoću koga se opisani transport ustvari naziva N⁺ K⁺ ATP-aza a sam aktivni transport NA⁺K⁺ pumpa. Kako se ovaj transport vrši nasuprot koncentracionom gradijentu, ovaj proces zavisi od utroškaenergije. Ako se na neki način inhibira enzim ATP-aza i spreči sinteza visoko energetskih fosfatnih jedinjenjakao što je adenozin tri fosfat (ATP) i arginin (kod beskičmenjaka), koji su glavni energetski izvori, dolazi doinaktivacije pumpnog mehanizma, a ako inaktivacija pumpe traje duže, neuron se depolariše i ne može višeda generiše nervne impulse. Važna uloga pumpe je da očuva volumen ćelije. Pumpa ispumpa 3 jona Na⁺ aunese 2 K⁺. Ima 5 aktivnih mesta.

by J.M., Ninche, Sandra, Ranagol, gen2010
12. Bioelektrične pojave (membranski potencijal)SLEDEĆEM MATERIJALU MOŽDA NIJE OVDE MESTO, ALI JE PREDAVANJE IVETIĆKE BILO
PRILIČNO KONFUZNO OVOG PUTA, PA JEDNOSTAVNO NE ZNAM GDE DA UGLAVIM.
extracelularno: van ćelije
intracelularno: unutar ćelije
ćelijska membrana preferira Ka jone, ovi joni jako lako ulaze u ćeliju. Zato Ka jona ima puno više
unutar ćelije, nego van ćelije.
Ka jon je pozitivno naelektrisana.
Na jona ima puno više spolja, van ćelije
U uslovima mira, ćelijska membrana je pozitivnija sa spoljašnje strane, nego sa unutrašnje
strane, za oko 60 mV. Ovo je potencijal mirovanja.
Ćelijska membrana je ženski rod, i preferira Ka jone.
Ćelijska membrana može gubiti naelektrisanost nekada
Ako na ćelijsku membranu deluje određena draž, onda ć.m. može postati osetljivija na Na jone, i
zbog toga Na joni će lakše prolaziti kroz ć.m., nego Ka joni. Ovo se zove depolarizacija (kada su
otvoreni Na kanali)
i ovako se dakle dolazi do gubitka polarizovanosti ć.m, polarizovanost postaje 0 mV, pa prelazi u
pozitivno, nastaje takozvani šiljak.
Preduslov je: kritični nivo depolarizacije, što iznosi 25 mV.
Kada se potencijal mirovanja promeni za 25 mV, to znači da će doći do formiranja akcionog
potencijala.

by J.M., Ninche, Sandra, Ranagol, gen2010
Akcioni potencijal je... bla.
Repolarizacija: vraćanje ćelijske membrane u stanje koje je bilo pre delovanja draži.
PREDAVANJE IVETIĆKE PONOVO JAKO KONFUZNO, PA STAVLJAM OVDE.
doprinos difuznog potencijala Ka - ako su potencijali jedini uzrok potencijala mirovanja sa
unutrašnje strane vlakana iznosi 94mV????????????????????????
doprinos dif. Na - u normalnom vlaknu, propustljivost za Ka je oko 100 puta veća nego za Na,
sam difuzni potencijal nastaje zbog difuzije Ka i Na stvorio bi difuzni potencijal ok -86mV?????
doprinos Na/Ka pumpe- dodatna -4mV membranskog potencijala??????????????
broj voltažno zavisnih kanala
Ranvijerova suženja: 2000-12000
površina mijelina <25
soma: 50-75
Inicijalni segment: 350-500
Membrana svih neurona je polarizovana: oko njene spoljašnje i unutrašnje strane nalaze se pozitivno i
negatinovo naelektrisane čestice. Spoljašnja strana je pozitivna, unutrašnja strana je negativna.
Membranski potencijal je nejednaka distribucija naelektrisanih čestica na spoljašnjoj i unutrašnjoj strani
membrane. Zbog ovoga postoji potencijalna razlika između aksoplazme i ekstracelularne tečnosti. U
uslovima kada nervna ćelija nije aktivirana, potencijal mirovanja je -60 -75 mV. Kada draž deluje na nevrnu
ćeliju, dešava se depolarizacija.
12. Bioelektrične pojave (membranski potencijal)
Bioelektrični fenomeni su kontrolni sistemi koji sinhronizuju funkcije najvećeg broja ćelija našeg tela. Unutarćelije i u međućelijskoj tečnosti vlada „princip elektroneutralnosti“, što znači da je broj anjona i katjonapribližno jednak sa obe strane ćelijske membrane. U vanćelijskoj sredini preovlađuju natrijumovi joni i hloridi,a u ćeliji joni kalijuma i organski anjoni. Zato u vreme električnog mirivanja, kada ne stvaraju i ne provodebioelektrične signale, između intracelularne i ekstracelularne površine membrane postoji razlika unaelektrisanju (potencijalska razlika). Ona predstavlja osnovnu polarisanost membrane, a poznata je kaomembranski potencijal mirovanja (MPM) ili transmembranski potencijal (TMP). Najčešće je to potencijalkalijuma koji u miru prolazi kroz svoje sporo-cureće kanale i ide ka međućelijskoj tečnosti. MPM različitihćelija ima različitu vrednost, ali je za isti tip ćelija konstantan. Razlozi za postojanje MPM-a su: selektivna
propustljivost plazma membrane, Donnan-ov efekat nedifuzibilnih anjona u aktivnost Na⁺/Ka⁺ pumpi.
Selektivna propustljivost (permeabilnost) membrane znači da je ona u različito vreme, na različitim mestimapropustljiva za različite jone, usled različitih tipova jonskih kanala, nosača i/ili pumpi koje se u njoj nalaze.Tako je u vreme električnog mirovanja plazma membrana slabo propustljiva za fosfate, sulfate, jonemagnezijuma i kalcijuma, a veoma je propustljiva za jone kalijuma i natrijuma (u nekim slučajevima i zahloride), koji su zato odgovorni za njenu osnovnu polarisanost, tj. za postojanje MPM-a. Ovo znači da je zakalijum otvoren daleko veći broj kanala nego za natrijum, pa je difuzija kalijumovih jona kroz plazmamembranu oko 50 puta veća od difuzije natrijuma. Kalijumovi joni, tada prolaze u međućelijsku tečnost krozsvoje pasivne kanale zbog povoljnog koncentracijskog gradijenta. Kalijumovi joni prolaze sami, bez anjona

by J.M., Ninche, Sandra, Ranagol, gen2010
koji ostaju uz membranu i doprinose nejednakoj distribuciji anjona i katjona uz spoljašnju i unutrašnjupovršinu membrane. Upravo ova difuzija kalijumovih jona kroz sporo-cureće jonske kanale daje najveći deovrednost MPM-a.
Donnan-ov efekat nedifuzibilnih anjona daju molekuli i joni koji ne mogu prolaziti kroz ćelijsku membranu jernisu liposolubilni, a u membrani nema jonskih kanala ili nosača za njihov transport. To su veliki proteinski,fosfatni i sulfatni anjoni ćelije. Tokom difuzije kalijumovih jona ka međućelijskoj tečnosti, u vreme MPM-a, oviintracelularni anjoni kreću za katjonima kalijuma, ali pošto ne mogu proći kroz plazma membranu, ostaju utankom sloju uz njenu unutršnju površinu i svojim negativnim nabojem ne dozvoljavaju katjonima kalijumakoji su izašli da se udalje iz uskog sloja uz spoljašnju površinu plazma membrane. Na opisan način sepozitivno naelektrisani joni grupišu uz spoljašnju površinu plazma membrane, dok se sa njene unutrašnjestrane nagomilavaju anjoni, čime se postiže veliki deo polarisanosti plazma membrane u vreme MPM-a.
Natrijum-kalijumove pumpe su elektrogenične jer ne menjaju jon natrijuma za jon kalijuma, već za svaka 3natrijumova jona koja izbace u međućelijsku tečnost vrate u ćeliju 2 kalijumova jona. One tako doprinose dau uskom sloju, koji prekriva spoljašnju površinu olazma membrane, ima više pozitivnih jona nego sa
intracelularne strane membrane. Na⁺/Ka⁺-pumpe imaju veliki značaj za održavanje MPM-a, jer su
neophodne za vraćanje repolarisane membrane u stanje mirivanja (polarisanosti). Natrijum-kalijum ATP-aze(pumpe) su integralni transportni membranski proteini koji omogućavaju održavanje visoke unutarćelijskekoncentracije kalijuma, uz nisku koncentraciju natrijuma. Na njenoj ekstracelularnoj površini nalaze se dvamesta za vezivanje kalijuma, a na ekstracelularnoj strani su tri mesta za natrijum.
Hipopolarisanost i hiperpolarisanost membrane
Hipopolarisanost i hiperpolarisanost su promene potencijala plazma membrane koje omogućavaju promenuekscitabilnosti (razdražljivosti) ćelije. Tako je hipopolarisanost stanje smanjene polarisanosti plazmamembrane. Javlja se usled njene pojačane propustljivosti za jone natrijuma i/ili kalcijuma koji, zbog povoljnogkoncentracijskog i električnog gradijenta, intenzivnije prolaze u ćeliju u vreme MPM i tako smanjuju razliku unaelektrisanju između spoljašnje i unutrašnje površine plazma membrane. Zato se potencijal membranepribližava pražnoj vrednosti, pa raste ekscitabilnost, tj. povećava se mogućnost da ćelija proizvede akcionipotencijal. Međutim, ako se ne dostigne pražna vrednost neće nastupiti akcioni potencijal, a hipopolarizacijamembrane će biti poništena istim mehanizmima kojima se održava vrednost MPM-a. Hiperpolarisanost jestanje suprotno hipopolarisanosti jer podrazumeva povećanje polarisanosti plazma membrane. Jedan odnačina da se ona postigne je povećanje propustljivosti za kalijumove jone u vreme MPM-a, kada zbogpovoljnog koncentracijskog gradijenta oni izlaze iz ćelije i gomilaju svoje pozitivno naelektrisanje u uskompojasu uz spoljašnju površinu plazma membrane, čineći je više polarisanom. Drugi način je da membranapostane pojačano propustljiva za hloride koji tada, zbog izrazitog koncentracijskog gradijenta, ulaze u ćeliju igomilaju negativno naelektrisanje sa unutrašnje strane plazma membrane, što se takođe manifestujepovećanjem polarnosti. Treći način je da se smanji propustljivost ćelijske memrane za jone natrijuma i/ilikalcijuma a da se istovremeno zadrži povećana propustljivost za kalijumove jone koja redovno postoji uvreme MPM-a, tako da nema mogućnosti da natrijumovi i/ili kalcijimovi joni svojim ulaskom kompenzujuizlazak kalijumovih jona kroz sporo-cureće kanale, i zato raste količina pozitivnog naelektrisanja uzspoljašnju površinu plazma membrane.
Ravnoteža potencijala je izjednačavanje sila elektrohemijskog (teži da nađe K⁺) i koncentracionog (vuku K⁺u spoljašnju sredinu) gradijenta na određenom nivou potencijalne razlike.
Unutar ćelije i u međućelijskoj tečnosti vlada princip elektoneutralnosti, što znači da je
broj anjona i katjona približno jednak sa obe strane ćelijske membrane. Pritom, u vanćelijskoj
sredini preovlađuju natrijumovi joni i hloridi, dok u ćeliji dominiraju joni kalijuma i organski
anjoni. Ovi joni su tako raspoređeni da u tankom sloju uz spoljašnju površinu plazma membrane
dominiraju katjoni, dok sa unutrašnje strane preovlađuju anjoni. Zato u vreme električnog

by J.M., Ninche, Sandra, Ranagol, gen2010
mirovanja, kada ne stvaraju i ne sprovode bioelektrične signale, između intra i ekstracelularne
površine membrane skoro svih životinjskih ćelija postoji razlika u naelektrisanju koja predstavlja
polarisanost membrane, odnosno membranski potencijal mirovanja ili transmembranski
potencijal. Razlozi za postojanje MPM su:
l. Izbirljiva propustljivost membrane(selektivna preambilnost) znači da je ona u različito
vreme i na različitim mestima propustlj iva za različite jone, usled različitih tipova jonskih kanala
ili pumpi koji se u njoj nalaze (veoma propustljiva zajone Na i K).
2 .Donnan-ov efekat nedifuzibilnih anjona daju molekuli ijoni koji ne mogu difundovati
kroz ćelijsku membranu jer nisu liposolubilni a u membrani nema jonskih kanala ili nosača za
njihov transport.
3. Natrijum - kalijumove pumpe su elektrogenične jer ne menjaju jon natrijuma za jon
kalijuma, već svaka 3 natrijumova jona koja izbace u međućelijsku tečnost vrate u ćeliju 2
kalijumova jona. One tako doprinose da u uskom sloju koji prekriva spoljašnju površinu plazma
membrane ima više pozitivnih jona nego sa intracelulame strane membrane i imaju veliki značaj
u održavanju vrednosti MPM, jer su neophodne za vraćanje repolarisane membrane u stanje
mirovanja - polarisanosti.
Hipopolarisanost: stanje smanjene polarisanosti plazma membrane, usled njene pojačane
propustljivosti za jone Na i K i tako smanjuju razliku u naelektrisanju između spoljašnje i
unutrašnje površine plazma membrane
Hiperpolarisanost je stanje suprotno hipopolarisanosti, jer podrazumeva povećanje
polarisanosti plazma membrane odnosno povećanu propustljivost za K jone koji pojačano izlaze
iz ćelije.
Membranski potencijal postoji na ćelijskoj membrani gotovo svih ćelija. Nastaje usled različite koncenracije
jona sa obe strane ćelijske membrane, kao i različite propustljivosti ove membrane za jone. Membranski
potencijal je bitan za nastanak i prenošenje nervnih impulsa, kao i za membranski transport.
Membranski potencijal izazvan difuzijom
Koncentracja jona kalijuma je intracelularno veoma visoka, dok je ekstracelularno niska. Usled toga se javljaveliki koncentracioni gradijent i joni kalijuma prirodno teže da difunduju iz unutražnjosti ćelije u spoljašnjusredinu. Natrijum je glavni jon ekstracelularne sredine, dok ga intracelularno ima samo u niskoj koncentraciji.Joni natrijuma prirodano teže da difunduju u unutrašnjost ćelija.
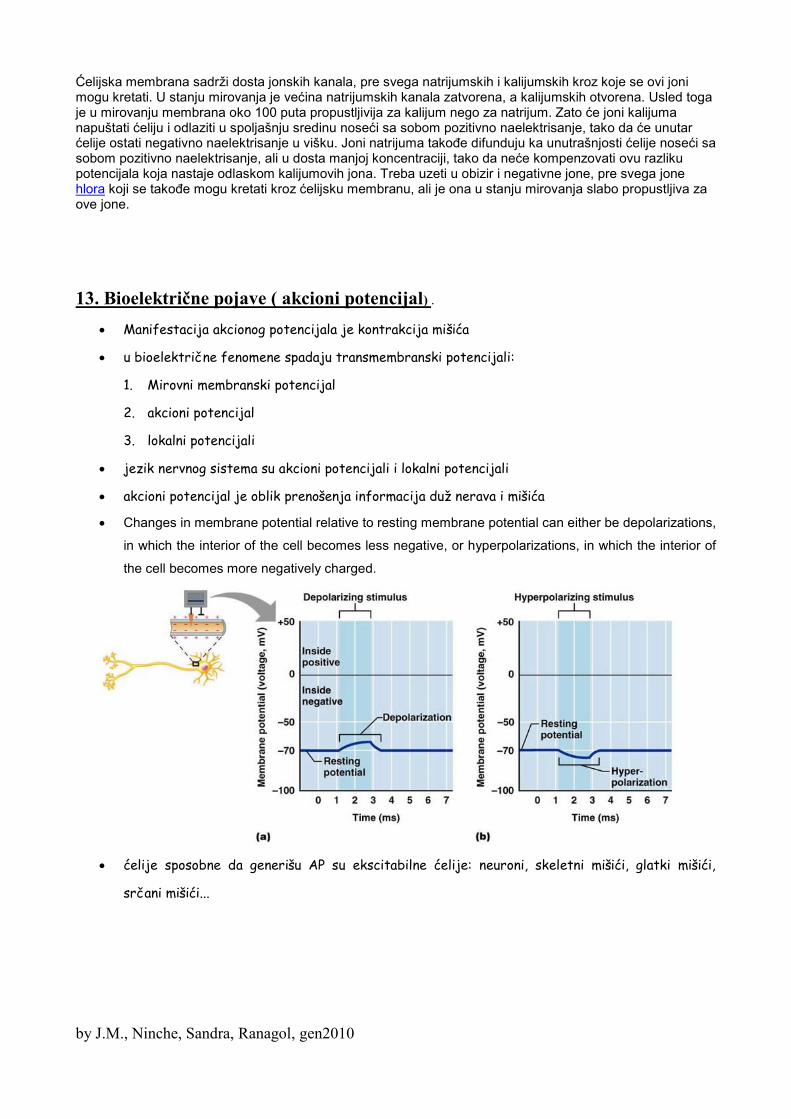
by J.M., Ninche, Sandra, Ranagol, gen2010
Ćelijska membrana sadrži dosta jonskih kanala, pre svega natrijumskih i kalijumskih kroz koje se ovi jonimogu kretati. U stanju mirovanja je većina natrijumskih kanala zatvorena, a kalijumskih otvorena. Usled togaje u mirovanju membrana oko 100 puta propustljivija za kalijum nego za natrijum. Zato će joni kalijumanapuštati ćeliju i odlaziti u spoljašnju sredinu noseći sa sobom pozitivno naelektrisanje, tako da će unutarćelije ostati negativno naelektrisanje u višku. Joni natrijuma takođe difunduju ka unutrašnjosti ćelije noseći sasobom pozitivno naelektrisanje, ali u dosta manjoj koncentraciji, tako da neće kompenzovati ovu razlikupotencijala koja nastaje odlaskom kalijumovih jona. Treba uzeti u obizir i negativne jone, pre svega jonehlora koji se takođe mogu kretati kroz ćelijsku membranu, ali je ona u stanju mirovanja slabo propustljiva zaove jone.
13. Bioelektrične pojave ( akcioni potencijal) .
Manifestacija akcionog potencijala je kontrakcija mišića
u bioelektrične fenomene spadaju transmembranski potencijali:
1. Mirovni membranski potencijal
2. akcioni potencijal
3. lokalni potencijali
jezik nervnog sistema su akcioni potencijali i lokalni potencijali
akcioni potencijal je oblik prenošenja informacija duž nerava i mišića
Changes in membrane potential relative to resting membrane potential can either be depolarizations,
in which the interior of the cell becomes less negative, or hyperpolarizations, in which the interior of
the cell becomes more negatively charged.
ćelije sposobne da generišu AP su ekscitabilne ćelije: neuroni, skeletni mišići, glatki mišići,
srčani mišići...

by J.M., Ninche, Sandra, Ranagol, gen2010
apsolutni refraktorni period?
relativni refraktorni period?
Akcioni potencijal
Nervni signali se prenose putem akcionog potencijala. Akcioni potencijal nastaje naglom promenompotencijala mirovanja. Ovaj proces se zove depolarizacija. Akcioni potencijal obično traje kratko i završavase vraćanjem potencijala u stanje potencijala mirovanja-repolarizacija.
Depolarizacija predstavlja naglu promenu mirovnog potencijala. U tom trenutku se otvaraju brzi natrijumski ispori kalcijumski kanali, a zatvaraju kalijumski tako da pozitivni joni natrijuma i kalcijuma ulaze velikombrzinom u ćeliju. Sada je propustljivost membrane za jone natrijuma oko 5 000 puta veća nego za jonekalijuma. Oni nose sa sobom pozitivno naelektrisanje, tako da potencijal unutar ćelije postaje pozitivniji.
Kada se ove vrednosti sada unesu u Goldmanovu jednačinu dobija se vrednost akcionog potencijala oko 0mV (nekad malo manje, nekad više).
Natrijumski kanali se ubrzo zatvaraju, a kalijumski ponovo otvaraju čime se potencijal vraća u pređašnjestanje. Ovaj proces je repolarizacija.

by J.M., Ninche, Sandra, Ranagol, gen2010
Faze AP:
1. Mirovanje
2. Depolarizacija
3. Repolarizacija
AP traje od 1 ms do 2 ms
vrste AP:
šiljati AP: bez gubitka snage, amplituda i trajanje ostaju isti
AP sa platoom: traje oko 250 ms, otvaraju se spori kanali Ka
onemogućuje tetanizaciju?????????????

by J.M., Ninche, Sandra, Ranagol, gen2010
13. Bioelektrične pojave (akcioni potencijal)
Akcioni potencijal predstavlja promenu naelektrisanja membrana. Nervne, mišićne i žlezdane ćelije suekscitabilne, što znači da se osnovna polarisanost njihove plazma membrane lako može narušiti, u smisluhipopolarizacije i hiperpolarizacije. Kada hipopolarisanost plazma membrane dostigne vrednost pražnogpotencijala nastaje akcioni potencijal (AP). To je „biološka ili jonska“ struja (biostruja ili impuls) koja služi zameđućelijske komunikacije, tj. za prenos informacija sa jedne ne drugu ćeliju. AP je oblik prenošenjainformacija duž membrane nerava i mišića, i u svakodnevnom jeziku se naziva impuls ili nadraženje.
Svaki monofazni AP se sastoji iz faze depolarizacije i faze repolarizacije. Pritom se AP različitih ćelijarazlikuju jer su njihove membrane različito propustljive za različite jone.
Faza depolarizacije
Da bi nastupila ekscitacija ćelije neophodno je da ulaz natrijumovih jona u nju bude 5-20 puta veći od izlaskajona kalijuma i/ili ulaska hloridnih jona. To se dešava kada ekscitabilna ćelija primi signal a neke druge ćelijeili sa receptora nekog čula, a u retkim slučajevima do toga dolazi spontano (kod samookidajućih ćelija).Spontano, posredstvom odgovarajućeg liganda, usled dejstva neke mehaničke sile ili promene voltažeotvaraju se kanali za natrijum i/ili kalcijum, pa ovi joni pojačano ulaze u ćeliju i smanjuju razliku unaelektrisanju (polarisanost) između spoljašnje i unutrašnje površine njene plazma membrane. Tako sepotencijal membrane približava pražnoj vrednosti, odnosno nastupa hipopolarisanost, koja pripada fazidepolarizacije a poznata je kao prepotencijal.
Pražni potencijal, prag ili nivo okidanja predstavlja minimalni potencijal potreban za otvaranje voltažno-zavisnih natrijumovih i/ili kalcijumovih kanala. Samo ako je prepotencijal dovoljno veliki da postigne voltažupraga, on po zakonu „sve ili ništa“ automatski omogućava okidanje ekscitabilne ćelije, tj. nastanak akcionogpotencijala.
Depolarizacija se definiše kao ulazak natrijumovih i/ili kalcijumovih jona u ćeliju, pri čemu se potencijalplazma membrane menja sa -75 mV (MPM) na oko +15 mV (maksimalno do +55 mV. Na zapisu se toregistruje kao uzlazna faza AP. Ovde treba imati na umu da prava depolarizacija nastupa kada se izgubipolarisanost i membrana se zbilja depolariše, tako da više nema razlike u naelektrisanju između njenespoljašnje i unutrašnje površine (0 mV). Deo AP koji ima vrednost iznad 0 mV označava se kao prebačenipotencijal ili kao obrnuta polarisanost plazma membrane (reverzija polariteta) jer tada njena spoljašnjapovršina, koja je u vreme MPM-a imala višak pozitivnog, ima višak negativnog naelektrisanja, dok uzunutrašnju površinu dominira pozitivno naelektrisanje koje je uneo natrijum.
Pri depolarizaciji se, zbog ulaska jona natrijuma i/ili kalcijuma, po spoljašnjoj površini plazma membrane širitalas negativnog a po unutrašnjij talas pozitivnog naelektrisanja, što obično predstavlja naredbu ćeliji zaodgovarajuću akciju. Kada su u pitanju neuroni „akcija“ je izazivanje hipopolarizacije ili hiperpolarizacijejednog ili više neurona, efektorskih ili receptorskih ćelija sa kojima oni sinaptički komuniciraju, dok kodmišićnih ćelija pod akcijom podrazumevamo kontrakciju.
Vrste AP:
1) Šiljati - od početka do kraja nervnog vlakna
2) Sa platoom - depolarizacija se vrši regulisanjem natrijumovih kanala, a kada se dostigne vrh otvarajuse spori kalijumovi kanali. Plato traje do 250 milisekundi. Primer je srčani mišić.
Akcioni potencijali mogu da budu: veliki, mali, visoko ili niskofrekventni, dugotrajni, kratkotrajni (u neuronimatraju samo 1 msec.), depolarišući ili hiperpolarišući. Pritom svi AP iste ćelije imaju identičnu amplitudu itrajanje,ali su im frekvence i pauze koje se javljaju između njih vrlo promenljive, što se koristi u međućelijskojsignalizaciji.
Kada jedna elektroda za registrovanje AP stoji neposredno uz unutrašnju a druga uz spoljašnju površinuplazma membrane dobija se zapis klasičnog šiljatog monofaznog AP, ali ako obe regulišuće elektrode stojesa iste strane plazma membrane regidtruje se bifazni AP. Za razliku od zapisa šiljatog AP, na kome serazlikuju uzlazna depolarizaciona i silazna repolarizaciona faza, na zapisu bifaznog AP razlikuju se:pozitivan otklon, izoelektrična linija, negativan ozklon i još jedna izoelektrična linija na kraju.

by J.M., Ninche, Sandra, Ranagol, gen2010
Svaka depolarisana ćelija mora da se repolariše. Repolarizacija otpočinje kada se završi depolarizacija iotvore se voltažno-zavisni kalijumovi kanali, zbog promene voltaže.
Repolarizacija se definiše kao izlazak kalijimovih jona iz ćelije, pri čemu se po spoljašnjoj površini plazmamembrane širi talas pozitivnog a po unutrašnjoj talas negativnog naelektrisanja. Ona obično predstavljanaredbu ćeliji da prestane sa započetom akcijom. Izlazak kalijima dovodi do promene polarisanostimembrane, tako što izaziva gomilanje pozitivnog naelektrisanja uz unutrašnju površinu plazma membrane.
Na čoveka deluje draž a nadražaj nastaje.
Draž ili draženj jemogućnost nastanka nadražaja (to je AP).
Da bi se uslovilo formiranje akcionog potencijala draž mora imati tri karakteristike.
1. Intenzitet- označava se kao reobaza i to je najniži intentitet neophodan za akcioni potencijal. Dražintenziteta reobaze deluje u vremenu (korisno vreme) i formira se akcioni potencijal.
2. Trajanje
3. Brzina uvođenja- ako nema brzine uvođenja doći će do adaptacije
Nervne, mišićne i žlezdane ćelije su ekscitabillne, što znači da polarisanost njihove
plazma membrane u vreme električne neaktivnosti (MPM) lako može da se naruši, u smislu hipo
ili hiperpolarizacije. Pritom je pravilo da hiperpolarisanost smanjuje a hipopolarisanost povećava
razdražljivost.
Kada hipopolarisanost plazma membrane dostigne vrednost pražnog potencijala nastaje
AKCIONI POTENCIJAL. To su biološke ili jonske struje koje služe za međućelijske
komunikacije, odnosno za prenos informacija sa jedne na drugu ćeliju. Akcioni potencijal
predstavlja brze prolazne promene vrednosti MPM, koje kada započnu moraju spontano i da se
završe. Svaki monofazni akcioni potencijal sastoji se _ iz faze depolarizacije i faze
repolarizacije. '
FAZA DEPOLARIZACIJE: u to vreme, membrana iznenada postaje veoma preambilna za
natrijumove jone dozvoljavajući dotok ogromnog broja natrijumovih jona u unutrašnjost aksona.
Normalno polarisano stanje od -90 mV gubi se zbog brzog porasta potencijala u pozitivnom
pravcu. To se naziva depolarizacija. Kod debelih nervnih vlakana, membranski potencijal u
stvari prebacuje nulti nivo i postaje nešto malo pozitivan, ali kod nekih tanjih vlakana kao i u
mnogobrojnim neuronima CNS, potencijal se samo približava nultom nivou i ne prebacuje se u
pozitivnu vrdnost.
FAZA REPOLARIZACIJE: počinje na vrhu akcionog potencijala. Pošto je membrana postala
visoko preambilna za natrijumove jone, natrijumovi kanali počinju da se zatvaraju, a kalijumovi
kanali se otvaraju više nego normalno. Onda, brza difuzija kalijumovih jona u spoljašnju sredinu
ponovo uspostavlja normalan negativan membranski potencijal mirovanja. To se naziva
repolarizacija membrane.
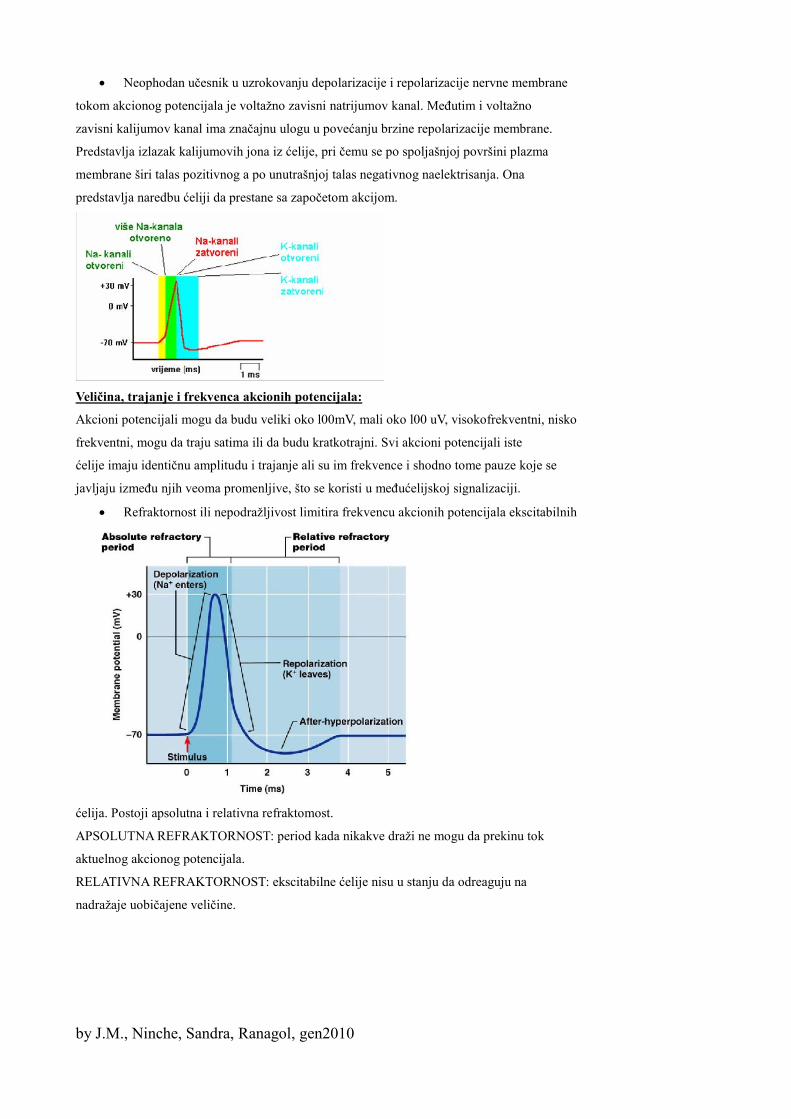
by J.M., Ninche, Sandra, Ranagol, gen2010
Neophodan učesnik u uzrokovanju depolarizacije i repolarizacije nervne membrane
tokom akcionog potencijala je voltažno zavisni natrijumov kanal. Međutim i voltažno
zavisni kalijumov kanal ima značajnu ulogu u povećanju brzine repolarizacije membrane.
Predstavlja izlazak kalijumovih jona iz ćelije, pri čemu se po spoljašnjoj površini plazma
membrane širi talas pozitivnog a po unutrašnjoj talas negativnog naelektrisanja. Ona
predstavlja naredbu ćeliji da prestane sa započetom akcijom.
Veličina, trajanje i frekvenca akcionih potencijala:
Akcioni potencijali mogu da budu veliki oko l00mV, mali oko l00 uV, visokofrekventni, nisko
frekventni, mogu da traju satima ili da budu kratkotrajni. Svi akcioni potencijali iste
ćelije imaju identičnu amplitudu i trajanje ali su im frekvence i shodno tome pauze koje se
javljaju između njih veoma promenljive, što se koristi u međućelijskoj signalizaciji.
Refraktornost ili nepodražljivost limitira frekvencu akcionih potencijala ekscitabilnih
ćelija. Postoji apsolutna i relativna refraktomost.
APSOLUTNA REFRAKTORNOST: period kada nikakve draži ne mogu da prekinu tok
aktuelnog akcionog potencijala.
RELATIVNA REFRAKTORNOST: ekscitabilne ćelije nisu u stanju da odreaguju na
nadražaje uobičajene veličine.

by J.M., Ninche, Sandra, Ranagol, gen2010
14. Bioelektrične pojave (postakcioni potencijal)
14. Bioelektrične pojave (postakcioni potencijal)
II faza repolarizacije
To kod većine ekscitabilnih ćelija počinje na vrhuncu AP, u momentu u kome voltaža iznosi između +15 i+30 mV. Usled otvaranja ovih kanala dolazi do brze difuzije kalijumovih jona iz ćelije u međućelijsku tečnost,što se na zapisu AP-a vidi kao naglo vraćanje potencijala na vrednost MPM-a. Na zapisu monofaznog APzapaža se brza početna difuzija kalijumovih jona koja traje sve do vraćanja voltaže na vrednost pražnogpotencijala. Potom se usporava difuzija kalijuma, usled sve nepovoljnijeg koncentracijskog i električnoggradijenta, što se registruje u vidu negativnog naknadnog potencijala (negativnog postpotencijala). Još uveknije neutralisano celokupno negativno naelektrisanje koje je zaostalo uz spoljašnju površinu plazmamembrane od prethodne depolarizacije. Negativan postpotencijal traje sve do uspostavljanja MPM-a. Potomse difuzija kalijuma nastavlja, ali je sve sporija, zbog sve slabijeg koncentracijskog i električnog gradijenta.Taj deo repolarizacije, tokom koje potencijal pada ispod vrednosti MPM-a, označavamo kao pozitivannaknadni potencijal ili pozitivan postpotencijal. Konačno se celokupna spoljašnja površina membranenaelektriše pozitivno, a unutrašnja negativno, tj. potencijal membrane se vreti na vrednost MPM-a.
Refraktornost (nepodraživost) limitira frekvencu AP ekscitabilnih ćelija. Postoji apsolutna i relativnarefraktornost. Apsolutna refraktornost vlada od momenta postizanja pražnog potencijala, tokomdepolarizacije, pa sve dok se tokom repolarizacije potencijal ne vrati na pražnu vrednost. To je period kadanikakve draži ne mogu prekinuti tok aktuelnog AP. Relativna refraktornost vlada kada ekscitabilne ćelije nisusposobne da odreaguju na nadražaje uobičajene jačine. Međutim, ako tada prime stimulus koji je dovoljnojak da dovede potencijal njihove plazma membrane do pražne vrednosti proizvešće AP.
15. Periferni nervi- građa i podela Vrste neurona:
1. Senzorni neuroni: primaju draži, tu nastaje AP, što se usmeravaju ka centrali,
koriste aferentna vlakna.
2. Motorni neuroni: tela neurona su u CNS, oni nose naredbe od CNS do mišića, koriste
eferentna vlakna.
3. Interni neuroni:
periferni nervi su sačinjeni od aferentnih i eferentnih vlakana, građeni su od motornih
neurona i senzornih neurona.
3 vrste perifernih nerava:
1. Motorni nervi
2. Senzitivni nervi
3. Mešoviti nervi
Vlakna u perifernom nervnom sistemu:
1. Mijelinska vlakna: poseduju mijelinski omotač (ovo čine Švanove ćelije), izanci se
obmotavaju, odvojeni su Ranvijerovim suženjima. Razdraženje se prostire skokovito,
jer je mijelin izolator. Što je mijelin deblji, brže prenosi razdraženje. Mijelinski

by J.M., Ninche, Sandra, Ranagol, gen2010
omotač stvaraju oligodendrociti.
2. Amijelinska vlakna: spolja se nalazi neurolema. Prolaz razdraženja je jako spora
broj mijelinskih vlakana je daleko veća od amijelinskih vlakana
razlika između neurona i glija
1. Neuroni formiraju AP, glije ne formiraju AP
2. Kod neurona postoje hemijske sinapse, kod glija ne postoje hemijske sinapse
3. Glija ima mnogo više, nego neurona
periferni nervni su praktično nezamorljivi, i koriste jako malo energije
15. Periferni nervi-građa i podela
Periferni nerv ili živac je složena tvorevina koja se sastoji od velikog broja nervnih vlakana koja su povezanavezivnim tkivom. Pojedinačna vlakna u nervu obavijena su izolacionim vezivno-tkivnim omotačemENDONEURIJUMOM. Preme funkciji razlikujemo 3 vrste nervnih vlakana:
1) Senzorna (senzitivna, aferentna, uzlazna) prenose impulse od receptora ka višim delovimanervnog sistema gde se doživljavaju kao određene senzacije.
2) Motorna (eferentna, silazna) prenose impulse od CNS-a da skeletnih mišića (efektora) ,tj.mišićnih vlakana kontrolišući tako voljne pokrete čoveka.
3) Autonomna (vegetativna) su eferentna po funkciji, kontrolišu svojim impulsima glatke mišićevisceralnih organa, žlezdane aktivnosti.
Senzorni i motorni neuroni učestvuju u građi perifernih nerava.
Periferni nervi se mogu podeliti u 2 velike grupe:
1) Spinalni nervi- potiču iz medule spinalis
2) Kranijalni nervi- potiču iz moždanog stabla, tj. iz baze mozga (lobanjski, ima 12 pari)
Periferni nervi su većinom mešoviti.
Vlakna u okviru perifernog NS mogu biti:
1) Mijelinska koja zauzimaju 2/3
2) Amijelinska koja zauzimaju 1/3
Mijelinska vlakna (vlakna A tipa) poseduju mijelinski omotač koga u perifernom NS proizvode i luče Švanovećelije, a u CNS-u to rade oligodendrociti. Mijelinski omotač se sastoji od većeg br. namotaja dvostrukeplazma membrane Švanove ćelije, filovanih međućelijskim matriksom i mijelinom. Broj namotaja ovogomotača kreće se i do stotinu i često je višestruko deblji od samog aksona. Zbog omotača ova vlakna subele boje i deblja su od nemijelinskih. Na istom vlaknu obično se nalazi oko 500 Švanovih ćelija koje surazdvojene Ranvierovim suženjima - čvorićima. Švanova ćelija obmotava 1-3 mm nervnog vlakna, ondanastupa Ranvierovo suženje koje je oko 1000 puta kraće. Mijelinska vlakna su deblja i imaju duže Švanovećelije, pa zato pružaju manji otpor provođenju impulsa. Njihova Ranvierova suženja su dosta udaljenameđusobno, pa su skokovi AP veliki i on se brzo prenosi.
Amijelinska vlakna (nemijelinirana, vlakna C tipa) nemaju mijelinski omotač, već je nekoliko njih utisnuto uŠvanovu ćeliju im trasira put ali se ne obavija oko njih u više slojeva i ne luči značajne količine mijelina.Odsustvo mijelinskog omotača uzrok je sive boje ovih nervnih vlakana. Kroz amijelinska vlakna AP putujesporo.
Periferni nervi su praktično nezamorni i ispituje se njihova osetljivost na pritisak (najosetljivija su mijelisananervna vlakna) i anestetike.

by J.M., Ninche, Sandra, Ranagol, gen2010
Živci ili nervi su skupovi aksona čija se neuronska tela nalaze u CNS ili u perifemim
ganglijama. Nerv je skup aksona koji pripadaju različitim neuronima, a prostiru se u istom
pravcu. U jednom nervu obično ima od 100 do nekoliko hiljada nervnih vlakana. Sva provode
impulse izolovano, potpuno nezavisno od ostalih, zato što su pojedinačna vlakna u nervu
obavijena izolacionim vezivnotkivnim omotačem - ENDONEURIJUMOM. U sastavu istog
nerva mogu se nalaziti senzitivna i motorna nervna vlakna, oni se nazivaju mešoviti nervi i u
njima se informacije, odnosno nervni impulsi kreću u dva smera: od receptora ka CNS` i od CNS
ka efektoru. Perifemi živci su većinom mešoviti.
Neuronska tela nervnih vlakana senzitivnih živaca nalaze se van CNS, a neuronska tela
motoneurona su unutar CNS.
Prema delu CNS u kome su smeštena neuronska tela aksona koji ulaze u njihov sastav,
nervi se dele na:
1. spinalne (leđne) i
2. kranijalne (lobanjske).
Spinalnih živaca kod čoveka ima 30, a
lobanjskih 12 pari. Spinalni živci su uvek mešoviti, dok neki od kranijalnih živaca mogu biti
samo senzitivni ili samo motorni. I leđni i lobanjski pored somatskih nervnih vlakana koja
inervišu poprečno - prugaste mišiće, mogu da sadrže i vlakna koja inervišu viscelarnu
muskulaturu. Nervna vlakna koja inervišu mišiće viscelarnih (unutrašnjih) organa, nalaze se u
sastavu vegetativnog nervnog sistema.
16. Wallerova degeneracija i regeneracija
Ako se izvrši aksotomija, odnosno, presecanje perifernog živca I na taj način se nervna vlakna odvoje odsome, relativno brzo dolazi do degenerativnih promena u distalnom delu aksona, odnosno delu koji jeizgubio vezu sa telom neurona. U isto vreme se u proksimalnom delu aksona, tj. onom koji je ostao povezansa somom, odigravaju se procesi koji omogućavaju ponovo srastanje oba dela nervnog vlakna, odnosnoregeneraciju neurona. Kod sisara, regeneracija presečenih aksona moguća je samo u perifernom nervnomsistemu. Soma kod nižih kičmenjaka (npr. gušteri) dolazi do potpune regeneracije aksona i u perifernom i ucentralnom nervnom sistemu. Procese koje prate degeneraciju i regeneraciju neurona u perifernom nervnomsistemu opisao je Waller i po njemu se degeneracija presečenih nerava naziva Wallerova degeneracija.Degenerativni procesi u distalnom delu presečenog aksona ispoljavaju se u vidu morfoloških i funkcionalnihpromena. Morfološke promene se najpre uočavaju na mijelinskom omotaču a zatim i u aksoplazmi.
Mijelinski omotač najpre promeni svoj izgled i u preiodu od 8-32 dana iščezava i njega absorbuju lokalnifagociti koji potiču od Švanovih ćelija. Na isti način reaguje i aksoplazma. U procesu degeneracije aksona,broj Švanovih ćelija se povećava i na kraju od distalnog dela nervnog vlakna preostane samo nurilema.Funkcionalna sposobnost aksona se u toku degeneracije gubi u roku od 3-5 dana. Paralelno sadegeneracijom distalnog dela nervnog vlakna mogu se uočiti jasne promene u njegovom proksimalnom delukoje omogućavaju regeneraciju aksona.

by J.M., Ninche, Sandra, Ranagol, gen2010
Na kraju proksimalnog dela javljaju se brojni protoplazmični izdanci i tu se umnožavaju Švanove ćelije. Akoje rastojanje između dva rasečena dela aksona malo, do 3 mm, ako je sačuvano vezivno tkivo živca, jedanod protoplazmičnih izraštaja proksimalnog dela aksona urasta u praznu neurilemu i tako dolazi do srastanjanervnog vlakna. Ako je rastojanje između proksimalnog i distalnog dela veće, može doći do formiranja otokasličnog tumoru koji se naziva neuronom, koji sprečava regeneraciju. U CNS-u viših kičmenjaka presečeniaksoni ne izrastaju. Razlog tome može biti glijalni ožiljak koji blokira regeneraciju. Takođe, misli se da sumetaboličke sposobnosti centralnih neurona nedovoljne da produkuju materijal potreban za regeneraciju.
U toku regeneracije se vrši neprekidno aksoplazmatsko kretanje od some ka nervnom vlaknu (anterogradnitransport) i to približno onom brzinom kojom regenerisani deo aksona raste. Postavljena je i teorija premakojoj dva dela nervnog vlakna srastaju na osnovu specifičnih hemijskih signala. Takođe se razmatramogućnost učešća sličnih hemijskih signala u embrionalnom razviću nervnog sistema. Njihova priroda nijedo kraja definisana ali se smatra da je reč o membranskim proteinima male molekulske mase. PoredWallerovih degenerativnih procesa koji zahvataju distalni deo presečenog vlakna, u novije vreme otkrivenesu i degenerativne promene koje zahvataju proksimalni deo presečenog neurona označene kao retrogradnadegeneracija i prostiru se od prvog Ranvierovog prstena ili do samog neuronskog tela gde se primećuju ucitoplazmi, jedru, i Nislovim granulama. Utvrđeno je da se degenerativne promene događaju i kod ovihneurona sa kojima stvaraju sinapsu presečena nervna vlakna. Reakcija tih postsinaptičkih ćelija, tj.transsimpatička reakcija ukazuje na to da se pomoću sinaptičke veze, pored nervih impulsa, napostsinaptičku ćeliju prenose I trofički uticaji.
Wallerova regeneracija se može odvijati do 6 meseci, nakon toga ne može da dođe do
regeneracije!
1 nerv = 1000-e vlakana
inervacija---funkcija poremećena---pogrešan tok regeneracije
Ako se slučajno ili namerno u eksperimentalne svrhe, izvrši aksotomija, odnosno
presecanje perifernog živca i na taj način se nervna vlakna odvoje od some, relativno brzo dolazi
do degenerativnih promena u distalnom delu aksona, odnosno delu koji je izgubio vezu sa telom
neurona. U isto vreme u proksimalnom delu aksona, tj. onom koji je ostao povezan sa somom,
odigravaju se procesi koji omogućavaju ponovno srastanje oba dela nervnog vlakna, odnosno
regeneraciju neurona.

by J.M., Ninche, Sandra, Ranagol, gen2010
Degenerativni procesi u distalnom delu presečenog aksona ispoljavaju. se u vidu
morfołoških i funkcionalnih promena. Morfološke promene se najpre uočavaju na mijelinskom
omotaču, a zatim i u aksoplazmi. Pošto prvo na karakterističan način promeni svoj izgled,
mijelinski omotač u periodu od 8-32 dana iščezava, njega apsorbuju lokalni fagociti koji potiču
od Švanovih ćelija. Na isti način razgrađuje se i aksoplazma. U procesu degeneracije aksona broj
Švanovih ćelija se poveća i na kraju od distalnog nervnog vlakna preostane samo prazna
neurilema. Što se tiče funkcionalne sposobnosti alćbna i ona se gubi u roku od oko 3-5 dana.
Paralelno sa degeneracijom distalnog dela nervnog vlakna mogu se uočiti i jasne promene
u njegovom proksimalnom delu koje omogućavaju regeneraciju aksona. Na kraju tog dela
aksona javljaju se brojni protoplazmatični izdanci i tu se umnožavaju Švanove ćelije. Ako je
rastojanje između dva presečena dela aksona malo i ako je sačuvano vezivno tkivo živca, jedan
od protoplazmatiinih izraštaja proksimalnog dela aksona urasta u praznu neurilemu i tako dolazi
do srastanja nervnog vlakna. U slučaju kadaje rastojanje izmeđuproksimalnog i distalnog dela
aksona veće, može doći do formiranja otoka sličnog tumom koji se naziva neuroma i koji
sprečava regeneraciju.
17. Zakoni provođena razdraženja nervnim vlaknimaNervi, tj. nervna vlakna poseduju visok stepen nadražljivosti i sprovodljivosti tj. sposobnosti
da na dejstvo nadražaja odgovara procesom razdraženja, tj. depolarizacijom i pojavom A.P.
ZAKON OBOSTRANOG SPROVOÐENJA - tj. zakon dvosmernosti. U uslovima
eksperimentalnog nadraživanja nervnog vlakna Babužin je pokazao da se proces razdraženja"
rasprostire u oba smera od mesta dejstva nadražaja. U neurofiziologiji se govori o rasprostiranju
procesa razdraženja u aferentnom-centripetalnom pravcu pojedinačnih nervnih vlakana (od
perifemih receptomih sistema u CNS), kao i eferentnom-centrifugałnom pravcu (iz nervnih
centara na periferiji ka efektomim organima). U prirodnim uslovima u organizmu proces
razdraženja se sprovodi uvek u jednom smeru.
ZAKON FIZIOLOŠKE CELINE NERAVA - ZAKON INTEGRITETA- prema ovom zakonu
koji podrazumeva anatomsku i fiziološku celinu, tj. integritet membrane, ako se bilo kojim
agensom naruši ovaj integritet nervnog vlakna, prestaće da se sprovodi proces depolarizacije i
generisanje A.P. Nervno vlakno se u tom slučaju ponaša kao da je presečeno, tj. kao da je
razoren anatomski integritet. Npr. ako je u_ pitanju mešoviti nerv, nastaje anestezija i paraliza
onih delova tela koje inerviše presečeni nerv.
ZAKON IZOLOVANOG SPROVOĐENJA IMPULSA- periferni nerv se sastoji iz velikog broja
pojedinačnih nervnih vlakana pri čemu se u isto vreme proces razdraženja rasprostire u jednim
vlaknima centripetalno a u drugim centrifugalno, pri čemu ovaj proces razdraženja koji se
sprovodi preko jednog nervnog vlakna nikada ne prelazi na susedno nervno vlakno. Ukoliko bi

by J.M., Ninche, Sandra, Ranagol, gen2010
razdraženje prelazilo na susedno nervno vlakno tada bi reagovali razni mišići što bi dovelo do
rastrojstva pokreta.
BRZINA SPROVOĐENJA NADRAŽAJA-dugo se verovalo daje brzina prostiranja razdraženja
po nervu skoro jednaka brzini rasprostiranja svetlosti. Helmholc je ustanovio da je brzina
rasprostiranja mnogo manja, a takođe je utvrđeno da je ona i različita kod raznih bioloških vrsta,
pa i kod organizama iste vrste. Najbržu sprovodljivost imaju motorni nervi kod čoveka (60-
l20m/s), a najsporija su nervna vlakna koja sprovode recepciju, kao i vegetativna nervna vlakna.
Sva nervna vlakna provode impulse IZOLOVANO – nezavisno od ostalih, jer su pojedinačna vlaknaobavijena izolacionim vezivnotkivnim omotacem-endoneurijumom. Perineurijum je omotač skupa nervnihvlakana koja se pružaju u istom pravcu (fascikulus) a fascikulusi su udruženi u nerv koji obavija omotačepineurijum.
JEDNOSMERNO provođenje – Aferentna vlakna prenose akcione potencijale od receptora ka centrima aeferentna ih prevode od centara ka efektorima. Sinapse određuju smer provođenja impulsa kroz nervnavlakna (in vivo). Impulsi idu isključivo ka presinaptičkoj membrani a sa nje se prenose na postsinaptičkumembranu hemijske sinapse. Obrnut smer se retko dešava u organizmu. DVOSMERNI prenos impulsa semože registrovati izolovanjem nervnog vlakna van organizma (in vitro) I podraživanjem strujom njegovogsredišnjeg dela, što nije uobičajeno u živom organizmu (osim u slučaju akson-aksonskih sinapsi), pa seaferentna vlakna podražuju od receptora a eferentna iz centra. Ovo se primenjuje samo kod fizikalne terapijekada želimo stimulacijom nervnih vlakana istovremeno da podstaknemo I centar I efektor.
KONTINUIRANO – prenos a.p. duž nemijelinisanih nervnih vlakana. Svaki deo vlakna mora posebno razvitia.p. (mora se depolarizovati a potom i repolarizovati), moraju da zahvate svaki deo membrane ponaosob.Proces depolarizacije privući NA+ sa susednog polarisanog dela koji je MPM i time mu smanjuje polarnost,izaziva depolarizacuju dok se on automatski repolariše. Zbog ovakvog načina provođenja kod kojegdepolarizacija i repolarizacija moraju zahvatiti svaki deo membrane nervnog vlakna ponaosob, a.p. seprenose sporo (0.5-2m/s)- razlozi: tanji aksoni (zato imaju manju količinu jona pa pružaju veći odpor),nemijelisana vlakna.
SKOKOVITO (saltatorna kondukcija) - mijelisana vlakna - impuls zahvata jedno po jedno susednoRanvijerovo suženje (skakanje biostruja sa jednog na drugo Ranvijerovo suženje). Proces: depolarisanoRanvijerovo suženje pasivno privlači NA+ jone sa susednog polarizovanog Ranvjerovog suženja koje jeMPM i njemu smanjuje polarnost i izaziva depolarnost, on se automatski repolariše. Brz prenos - razlozi:veliki prečnik (debljina mijelina) i dužina rastojanja.
18. Klasifikacija nervnih vlakana
Vrsta vlakna Prečnik (μm) Brzina (m/s) FunkcijeA-α 13-22 70-120 α-motoneuron, primarni završeci u mišićnom vretenu,
Goldžijev tetivni organ, za dodir
A-β 8-13 40-70 dodir, sekundarni završeci u mišićnom vretenu
A-γ 4-8 15-40 dodir, pritisak, γ-motoneuron
A-δ 1-4 5-15 bol, grub dodir, pritisak, temperatura
B 1-3 3-14 preganglijska vlakna
C 0,1-1 0,2-2 bol, dodir, pritisak, temperatura, postganglijska vlakna

by J.M., Ninche, Sandra, Ranagol, gen2010
Po funkciji nervna vlakna svrstavamo u aferentna i eferentna.
Aferentna ili ushodna nervna vlakna provode impulse od receptora ka višim delovima
nervnog sistema. Često ih nazivamo senzornim mada sva aferentna vlakna nisu senzitivna, već
su to samo ona koja prenose informacüe do svesnih delova CNS-a. Za razliku od aferentnih
somatskih nervnih vlakana koja prenose informacije do svesti (npr. bol, dodir, ukus, miris), kroz
aferentna vlakna vegetativnog nervnog sistema pretežno se prenose informacije koje ne dopiru
do svesti (npr. visina krvnog pritiska, koncentracija kiseonika i ugljen-dioksida u krvi).
Eferentna, motorna ili nishodna nervna vlakna provode impulse od centra ka efektorima.
Ako pripadaju somatskom nervnom sistemu služe za kontrolu pokreta voljnih, tj.
poprečnoprugastih mišića, a kada su deo vegetativnog nervog sistema kontrolišu nevoljne
miokardske, glatkomišićne ili žlezdane ćelije.
Nervi su obično mešoviti, što znači da sadrže aferentna i eferentna vlakna. U mešovite spadaju
svi spinalni nervi, Vagus (X kranijalni nerv);koji sadrži ushodna i nishodna parasimpatička
vlakna vegetativnog nervnog sistema, kao i vlakna somatskog nervnog sistema koja nose
impulse iz kaudałnih delova usne duplje i vlakna koja kontrolišu poprečno prugaste mišića zida
jednjaliaiiničiće koji učestvujuu gutanju i govoru.
Podelapo histološkoj građi i načinu provođenja struja:
Mijelisana ili vlakna tipa A poseduju mijelinski omotač, koga u perifemom nervnom sistemu
proizvode i luče_Švanove ćelije, dok u CN S to rade oligodendrociti. Mijelinski omotač se sastoji
iz .većeg broja namotaja dvostruke plazma membrane Švanovih ćelija. Zbog prisustva ovog
omotača mijelinska nervna vlakna imaju belu boju i deblja su od nemijelinisanih.
Nemijelisana, anrijelisana ili vlakna tipa C nemaju mijelinski omotač, već je nekoliko njih
utisnuto u Švanovu ćeliju. Odsustvo omotačaje uzrok sive boje ovih nervnih vlakana.
Podela po debljini i po brzini provođenja biostruja:
Svrstavamo ih u A, B i C tip, a sva vlakna tip A grupišemo u podtipove: Aα, Aβ, Aγ i Aδ.
Pritom klasifikacija na A, B i C tip važi samo za eferentna, odnosno sva motorna vlakna, dok se
kod aferentnih koristi podela na vlakna tipa I, II, III i IV.
19. Karakteristike i vrste A nervnih vlakana
Aalfa – najdeblja, imaju najduže Švanove ćelije, pružaju najmanji otpor provođenja impulsa, kod njih sunajudaljenija Ranvierova suženja od drugih, pa su skokovi a.p. veliki i brzo se prenosi (čak 70-120m/s). Ovavlakna služe za kontrolu aktivnosti najpreciznijih poprečno-prugastih mišića.
Abeta - tanja su, bliže postavljene Švanove ćelije, sporije provode a.p.(30-75m/s), služe kao senzorna vlaknaza prenos informacija iz čula dodira.
Agama - još su tanja nego prethodna, još sporija (15-30m/s). Takva su motorna vlakna koja kontrolišupočetnu zategnutost intrafuznih vlakana mišićnih vretena.
Adelta - najtanja u organizmu. Njihove su najkraće Švanove ćelije, pa su im najučestalija Ranvijerovasuženja, prenose a.p. u veoma sitnim skokovima i imaju najmanju brzinu provođenja (5-30m/s). Npr. bol,hladno, dodir.

by J.M., Ninche, Sandra, Ranagol, gen2010
-tipu B pripadaju mijelinisana preganglijska simpatička i parasimpatička vlakna
-tip C obuhvata nemijelinisana vlakna.
Senzorna vlakna tipa I odgovaraju po debljini i po brzini provođenja impulsa motornim vlaknima tipa Aalfa. Iavlakna polaze sa receptorskih delova intrafuznih vlakana mišićnih vretena, Ib polaze sa Goldžijevih tetivnihreceptora, a samo su malo tanja od Ia i neznatno sporija od njih.
Senzorna vlakna tipa II odgovaraju po debljini i po brzini provođenja impulsa motornim vlaknima tipa Abeta.
Senzorna vlakna tipa III odgovaraju motornim vlaknima Adelta, a tip IV analogan je nemijelinisanim Cmotornim vlaknima.
U klasifikaciji aferentnih vlakana nema onih koji odgovaraju po debljini i po brzini provođenja impulsamotornim Agama i B vlaknima, jer u organizmu kičmenjaka nema ovakvih aferentnih vlakana.
Vlakna tipa Aα su najdeblja i imaju najduže Švanove ćelije pa pružaju najmanji otpor
provođenju impulsa. Ranvijerova suženja ovih vlakana su najviše udaljena od drugih, što
uslovljava velike skokove akcionog potencijala i omogućava im prenos brzinom od čak 70 -
120 m/s. lz ovih razloga Aα vlakna služe za kontrolu aktivnosti najpreciznijih poprečno
prugastih mišića.
Vlakna tipa Aβ su tanja i imaju bliže postavljene Švanove ćelije nego vlakna tipa Aα, pa
sporije provode biostruje (30 -75 m/s). Služe kao senzorna vlakna za prenos informacija iz
čula dodira.
Vlakna tipa Aγ su još tanja od Aβ i još sporije provode biostruje (15-30 m/s). Takva su
motorna vlakna koja kontrolišu početnu zategnutost intrafuznih vlakana mišićnih vretena.
Vlakna tipa Aδ su najtanja mijelisana vlakna u organizmu. Njihove Švanove ćelije su
najkraće pa su im Ranvijerova suženja najučestalija i zato prenose AP u veoma sitnim
skokovima, tako da imaju najmanju brzinu provođenja među mijelisanim vlaknima (5-30)
npr. bol, hladno, dodir.
Tipu B pripadaju mijelisana preganglijska i parasimpatička vlakna.
Tip C obuhvata nemijelisana vlakna.
Senzorna vlakna tipa I odgovaraju po debljini i po brzini provođenja impulsa motornim
vlaknima tipa Aδ.
Ia vlakna polaze sa receptorskih delova intrafuznih vlakana mišićnih vretena,
Ib polaze sa Goldžijevih tetivnih receptora, a samo su malo tanja od Ia i neznatno
sporija od njih.
Senzorna vlakna tipa II odgovaraju po debljini i po brzini provođenja impulsa motomim
vlaknima tipa Aβ.
Senzorna vlakna tipa III odgovaraju motomim vlaknima Aδ,
a tip IV analogan je nemijelisanim C motomim vlaknima.

by J.M., Ninche, Sandra, Ranagol, gen2010
U klasifikaciji aferentnih vlakana nema onih koji odgovaraju po debljini i po brzini
provođenja impulsa motornim Aγ i β vlaknima, jer u organizmu kičmenjaka nema ovakvih
aferentnih vlakana.
20. Razlike nervnih vlakana u odnosu na hipoksiju, pritisak i anestetike1. Hipoksija _ nedostatak kiseonika U tkivu "
Najosetljivija vlakna -tip B
Manje osetljiva - tip A
Najmanje osetljiva -tip C
2. Pritisak
Najosetljivija vlakna -tip A
Manje osetljiva - tip B
Najmanje osetljiva - tip C
3. Anestetici
Najosetljivija vlakna - tip C
Manje osetljiva - tip B
Najmanje osetljiva - tip A
21. Složene karakteristike akcionog potencijala perifernog nerva složeni AP: manifestacija različitih vlakana
Akcioni potencijal nervnog vlakna ima izgled šiljatog monofaznog akcionog potencijala,
na kome prvo uočavamo fazu depolarizacije a zatim i repolaizacije. Međutim, akcioni potencijal
nerva se bitno razlikuje od akcionih potencijala pojedinačnih nervnih vlakana, jer je nerv skup
nervnih vlakana različite debljine a samim tim i različite brzine provodenja impulsa. Prlikom
registrovanja akcionog potencijala sa celog nerva dobija se karakterističan zapis složenog
monofaznog akcionog potencijala, odnosno zbimi zapis akcionih potencijala, istovremeno
stimulisanih, pojedinačnih vlakana istog nerva. Prag nervnih vlakana se razlikuje pa zato na
ovom zapisu razlikujemo više vrhova, a svaki od njih odgovara jednom tipu vlakna. Na zapisu
složenog akcionog potencijala prvo uočavamo vrh mijelisanih Aα, zatim Aβ, Aγ, B i konačno
registrujemo zapis akcionog potencijla onih potencijala najsporijih nemijelisanih C vlakana.
Visina svakog vrha zavisi od broja vlakana određenog tipa u ispitivanom nervu, a ako u nekom
nervu ne postoje određeni tipovi vlakna na zapisu njegovog AP-a nedostaju vrhovi.
21. Složene karakterisitike AP perifernog nerva
AP nervnog vlakna ima izgled šiljatog monofaznog AP, na kome prvo uočavamo fazu depolarizacije a zatim irepolarizacije. AP nerva se bitno razlikuje od AP pojedinačnih nervnih vlakana, jer je nerv skup nervnih

by J.M., Ninche, Sandra, Ranagol, gen2010
vlakana različite debljine a samim tim i različite brzine provođenja impulsa. Prilikom registrovanja AP sacelog nerva dobija se karakterističan zapis složenog monofaznog AP, tj. zbirni zapis AP, istovremenostimulisanih, pojedinačnih nervnih vlakana istog nerva. Prag nervnih vlakana se razlikuje pa zato na ovomzapisu razlikujemo više vrhova, a svaki od njih odgovara jednom tipu vlakna. Na zapisu složenog akcionogpotencijala prvo uočavamo vrh mijelinisanih Aalfa, zatim Abeta, Agama, B i konačno registrujemo zapis APonih potencijala najsporijih nemijelinisanih C vlakana. Visina svakog vrha zavisi od broja vlakana određenogtipa u ispitivanom nervu, a ako u nekom nervu ne postoje određeni tipovi vlakna na zapisu njegovog APnedostaju vrhovi.
1. Amplituda AP pojedinačnih nervnih vlakana je konstantna a kod živca je promenljiva. Stimulus čiji jeintenzitet na nivou praga izaziva kod nerva (AP) manje amplitude, dok promena stimulusa jačine iznad pragadovodi do povećanja AP. Primenom stimulusa sve vecćg intenziteta moze se ustanoviti da porast amplitudeAP živca nije beskonačan. Pri određenoj jačini stimulusa, amplituda AP dostiže svoju maksimalnu vrednost iona se više ne menja. Tada živac počne da se ponaša po zakonu "sve ili ništa", tako da je odstupanje od togzakona bilo samo prividno. To se može objasniti razlikama koje postoje u fiziološkim karakterisitkamaaksona u sastavu pojedinih nerava, u prvom redu razlikama u pogledu njihove ekscitabilnosti. Recimo, akose na nerv primeni stimulus malog intenziteta, samo najekscitabilnija nervna vlakna- vlakna A alfa nerava ćegenerisati nervni impuls. Sa porastom intenziteta stimulusa, nervni impuls se stvara i u manje ekscitabilnimnervnim vlaknima i najzad će i najmanje ekscitibilna vlakna da stvaraju AP. Akcionom potencijalu živca kogagenerišu najekscitibilna nervna vlakna, aksoni pridodaju na taj način što oni koji se odnose na manjeekscitibilna nervna vlakna, nervni impulse pridodaju porastu ukupne amplitude AP živca. Kada se dostignemaksimalni intenzitet stimulusa, tj. takav intenzitet koji aktivira i najmanje ekscitabilna nervna vlakna usastavu istog živca, onda se registruje AP maksimalne amplitude. Porast amplitude AP živca priangažovanju sve većeg broja nervnih vlakana u njegovom sastavu uslovljen je time da razlika potencijalaizmeđu dela nerva zahvaćenog nervnim impulsom i ostalih delova postaje utoliko veći što je veći brojangažovanih jedinica.
2. AP živca razlikuje se od AP pojedinačnih nervnih vlakana i po svom obliku. Kada na nerv deluje stimulusmaksimalnog intenziteta tj. kada su ekscitirana sva nervna vlakna u njegovom sastavu, AP je složen i sastojise iz nekoliko talasa A alfa, beta, gama delta, B i C. Razlog za složeni oblik AP živca je nejednaka brzinaprostiranja nervnih impulsa kod nervnih vlakana različitih kategorija. Složeni AP živca sastojaće se iz svihpomenutih talasa samo ako su u njemu prisutni aksoni svih kategorija. Ukoliko nisu, AP ce biti jednostavniji-registrovaće se manji broj talasa. Intenzitet stimulusa mora biti dovoljan da aktivira vlakna svih kategorija.
22. Glijalne ćelije (astrociti - poreklo, građa i uloge)
Glijalne ćelije su potporne ćelije CNS-a i PNS-a, koje okružuju neurone. Neuroglija se deli na makrogliju(krupne ćelije), mikrogliju (sitne) i epitelne. Čine ½ zapremine moždanog tkiva, ima ih više nego neurona,sitnije su i manje razgranate. Nemaju akson, samo somu i dendrite (iz kojih po potrebi oslobađaju

by J.M., Ninche, Sandra, Ranagol, gen2010
neurotransmitere). Na membrani imaju mnogo receptora za neurotransmitere, faktore rasta, hormone...Mogu se razmnožavati za razliku od neurona. Ima više vrsta glija: astrociti, mikroglija, oligodendrociti,Švanove ćelije...
Astrociti – najkrupniji, najbrojniji, zvezdoliki. Svojim produžecima obavijaju neurone, sinapse i moždanekapilare. Velikim delom se nalaze između neurona i spoljašnjeg zida kapilara meke moždane ovojnice, anjihovi produžeci završavaju na bazalnoj membrani kapilara, u vidu papučice gradeći hematoencefalnubarijeru. Međusobno su povezani prisnim vezama (ne dozvoljavaju pasivan prelazak hidrosubilnih materija izkrvi u neurone) i električnim sinapsama tj. pukotinastim vezama(i sa neuronima), koje učestvuju u održavanjukoncentracije K jona i u koordinaciji glija ćelija. Formiraju mrežu koja usmerava migraciju neurona i rukovodinjihovim rastom jer proizvode neurotrofične supstance. Poseduju depoe glikogena i oslobađaju glukozu zaokolne neurone, pružaju im potporu i služe za električnu izolaciju, a predstavljaju i ćelije odbrane jer se mogutransformirati u makrofage. Učestvuju u procesima učenja i pamćenja. Obavijaju centralne sinapse, uklanjajui reutilizuju njihove neurotransmitere, regulišu koncentraciju K jona u međućelijskom prostoru mozga, jer supropustljivi za njih. Ne razmnožavaju se (kod zdravog mozga) ali u slučaju oštećenja, njihov broj sepovećava na mestu povrede i hipertrofiraju im produžeci da bi ispunili deo gde su mikroglije fagocitovaleoštećeno tkivo.
Glija ćelije su potporne ćelije CNS i perifernog nervnog sistema koje okružuju neurone,
ima ih daleko više nego neurona, one čine % zapremine moždanog tkiva. Za razliku od neurona
koji ne mogu značajno da se razmnožavaju, glija ćelije to mogu.
Astrociti su najkrupniji i najbrojniji gliociti, to su velike zvedolike ćelije, svojim
produžecima obavijaju neurone, sinapse i moždane kapilare. Povezani suag ipukotinastim
vezama, tj. električnim sinapsama, koje učestvuju u održavanju koncentracije K jona i u
koordinaciji glija ćelija. Astrociti formiraju mrežu koja usmerava migraciju neurona i rukovodi
njihovim rastom jer proizvode neurotrofične supstance. Oni pružaju potporu neuronima i služe
im' za električnu izolaciju, a predstavljaju i ćelije odbrane. Smatra se da astrociti učestvuju u
procesima vezanim za učenje i upamćivanje.
23. Glijalne ćelije ( oligodendrociti - poreklo, građa i uloge)

by J.M., Ninche, Sandra, Ranagol, gen2010
Oligodendrociti - sitniji i manje brojniji od astrocita. Ektodermalnog su porekla. Ime dobili po oskudnomdendritskom stablu, (Dendron-drvo, grane; oligo-malo). Osetljivi (u slučaju virusnog oboljenja sledidemijelinizacja). Imaju nutritivnu ulogu, ishranjuju okolne neurone. Popunjavaju mala oštećenja u CNS-u.Formiraju mijelinski omotač oko nervnih vlakana u CNS-u. Oligodendrocit različitim produžecima obavijapedesetak aksona ili okružuje isti akson na više mesta i proizvodi njegov mijelinski omotač.
Oligodendrociti se dele i imaju nekoliko produžetaka. Ne obnavljaju se lako, a i kada se obnove to nije upotpunosti.
Glija ćelije su potpoľnć ćelije CNS i perifemog nervnog sistema koje okružuju neurone,
ima ih daleko više nego neurona, one čine %z zapremine moždanog tkiva. Za razliku od neurona
koji ne mogu značajno da se razmnožavaju, glija ćelije to mogu.
Oligodendrociti su dobili ime po svom oskudnom dendritskom stablu. Ove glija ćelije su
sitnije i manje brojne od astrocita. [maju nutritivnu ulogu' jer ishranjuju okolne neurone, a
popunjavaju i mala oštećenja u CNS-u. Pored toga, oligodendrociti formiraju mijelínski omotač
oko nervnih vlakana CNS-a.
24. Švanove ćelije - građa i uloge
Obavijaju periferne sinapse i tako ih izoluju od okolnog međućelijskog matriksa. Ulaze u sastav neurona,tako što pružaju potporu nemijalisanim i mijelisanim nervnim vlaknima. Nemijelisana vlakna samo uranjaju uŠvanove ćelije, dok duž mijelisanih nervnih vlakana ove ćelije formiraju namotaje i luče mijelin, tj. grademijelinski omotač i održavaju njegovu stabilnost. Postiču funkciju neurona tako što proizvode i oslobađajucilijarni neurotrofični faktor i što stimulišu nervna vlakna, da u aksolemi istaknu receptore za faktor rastanerava (neophodan za njihov opstanak i razvoj). Mogu se razmnožavati (za razliku od neurona), njihovaproliferacija se ubrzava posle uspostavljanja kontakta sa aksonom.
Jedna Švanova ćelija stvara jednu mijelinsku košuljicu. Nalaze se između dva Ranvierova suženja. Imajumetaboličku funkciju.
Švanove ćelije obavijaju perifme sinapse, izolujući ih tako od okolnog međućelijskog
matriksa. Ove ćelije ulaze u sastav nerava, tako što pružaju potporu nemijelisanim i mijelisanim
nervnim vlaknima. Nemijelisana vlakna samo uranjaju u Švanove ćelije, dok duž mjelisanih

by J.M., Ninche, Sandra, Ranagol, gen2010
nervnih vlakana ove ćelije formiraju namotaje i luče mijelin, odnosno grade mijelínski omotač i
održavaju njegovu stabilnost. Stimulišu nervna vlakna da istaknu receptore za faktor rasta nerava
i proizvode i oslobađaju cilijarni neurotroñčni faktor. Švanove ćelije mogu da se razmnožavaju
za razliku od neurona.
25. Mikroglija (poreklo i uloge)
Mikroglija celije
Su sitne glija ćelije sa nekoliko razgranatih produžetaka. Potiču od matičnih ćelija hematopoeze (od stemćelija), mezodermalnog porekla, iz srednjeg klicinog lista. One su periciti kapilara, tj. izlaze iz krvnog suda.
Smeštene su u CNS-u, a aktivno se pokreću, uvećavaju i razmnožavaju tek u zapaljenom ili degenerisanomnervnom tkivu. Primarna uloga im je fagocitoza neurona podleglih apoptozi (fiziološka programirana smrtćelije). Po potrebi mogu fagocitovati oštećeno nervno tkivo i eventualne mikroorganizme koji uspevaju daprodru u njega, tako da njihov broj raste u toku infektivnih oboljenja nervnog sistema. Ova uloga uvrštavamikroglija ćelije u tkivni makrofagni sistem (tms).
Glija ćelije su potporne ćelije CNS i perifemog nervnog sistema koje okružuju neurone,
ima ih daleko više nego neurona, one čine % zapremine moždanog tkiva. Za razliku od neurona
koji ne mogu značajno da se razmnožavaju, glija ćelije to mogu.
Mikroglija ćelije su sitne glija ćelije sanekoliko razgranatih produžetaka. Uglavnom su
staticionirane na jednom mestu, a pokreću se, uvećavaju i razmnožavaju tek u zapaljenom ili
degenerisanom nervnom tkivu. One potiču od matičnih ćelija hematopoeze a primama uloga im
je fagocitoza neurona podleglih spontanoj apoptozi, mada po potrebi mogu fagocitovati oštećeno
nervno tkivo i eventualne mikroorganizme koji uspeju da prodru u njega, tako da njihov broj
raste u toku infektivnih oboljenja nervnog sistema.

by J.M., Ninche, Sandra, Ranagol, gen2010
26. Sinapse u CNS morfologija (hemijske i električne)
Sinapsa- funkcionalni kontakt izmedju 2 ćelije od kojih jedna mora biti neuron preko koje se prenosispecifična nervna informacija. Sinapse predstavljaju međućelijske komunikacije. To su funkcionalne jedinicenervnog sistema specijalizovane za prenos biostruja.
Kod ljudi su uglavnom sve sinapse hemijske koje služe za prenos signala u CNS-u. U njima prvi neuronsekretuje u sinapsu neurotransmiter. Neurotransmiter zatim deluje na receptorske protein u membranesledećeg neurona i može da ga ekscitira ili inhibira. 80 do 90 odsto ovih sinapsi je na dendritimaekscitatornog tipa, a 10 do 20 odsto je na telu neurona inhibitornog tipa. Izrazito važna karakteristika im ješto uvek prenose signale u jednom smeru, od neurona koji ispušta neurotransmiter (presinaptički neuron) doneurona na koji transmiter deluje (postsinaptički neuron). To je pricip jednostavnog provođenja kroz hemijskesinapse.
Najčešće prenose nervne informacije sa aksona na telo neurona.
Mogu biti različitog funkcionalnog značaja.
1. Da ugase nevažne informacije, a propuste jake signale
2. Šire važne signale
3. Sužavaju informaciju
Dele se na: 1. Hemijske (najbrojnije i najznačajnije)
2. električne (imaju specifične karakteristike)
3. mešovite(na jednom neuronu i hemijske i električne sinapse)
Hemijske sinapse- Sprovode nadražaj od aksona jednog neurona do dendrita drugog neurona. Plastične ifleksbilne: obezbeđuju varijabilnost u
prenošenju nervnih impulsa. Ostvaruju se uz posredovanje hemijske materije-
neurotransmitera, koga sintetiše i oslobađa presinaptička celija. Ove
sinapse su obavijene glija ćelijama (koje ih izoluju od okolnog tkiva i
sprečavaju osipanje neurotransmitera).

by J.M., Ninche, Sandra, Ranagol, gen2010
Postoje tri vrste hemijskih sinapsi :
1. aksodendritske (ima ih 80 – 90 %)
2. aksosomatske (nalaze se na telu naurona i ima ih 10 %)
3. aksoaksonske (1 % )
Presinaptička membrana - je deo plazma membrane koji prekriva površinu
sinaptičkog završetka presinaptičkog neurona, u njoj se nalaze brojni
voltažno- zavisni kanali za Na, K, Ca, kalcijumove natrijum- kalijum,
natrijum- kalijumove pumpe, receptori za neurotransmitere i transporteri
potrebni za rentilizaciju transmitera. Delovi presinaptičke membrane na
kojima ima najviše kalcijumovih voltažno- zavisnih jonskih kanala i uz koje
se nadovezuju brojne vezikule sa neurotransmiterom, poznati su kao "aktivne
zone" ili "gusta tela".
Sinaptički završeci su konkretni delovi ćelija koji međusobno sinaptički komuniciraju. Nazivaju se i završničvorići ili sinaptičke nožice. U njima se nalaze mitohondrije koje obezbedjuju ATP neophodan za rentilizacijui deponovanje neurotransmitera. Tu su i brojna
skladišta neurotransmitera u vidu vezikula koje se tokom prenosa biostruja
prazne u sinaptičku pukotinu egzocitozom. Sinapticke vezikule mogu biti male(40-60nm) i velike(70-250nm).
Tu se nalaze i molekuli kalmodulina, protein koji se aktivira vezivanjem četiri kalcijumova jona. Napresinaptičkoj membrani pojedinih hemijskih supstanci, mogu se naći i receptori i neki od kotransmitera(neuromodulatora) koji ne učestvuju direktno u transmisiji
biostruja, samo modifikuju efekte oslobođenog neurotransmitera. Takođe, na
presinaptičkoj pukotini nekih sinapsi nalaze se i autoreceptori za koje se
vezuju molekuli neurotransmitera sinapsi. Fiziološka opravdanost njihovog
postojanja je u tome što posle vezivanja neurotransmitera menjaju intenzitet
dalje egzocitoze, šime menjaju ekscitabilnost sinapse.
Sinaptička pukotina- je prostor širine 20-55 nm koji se nalazi između pre i
postsinaptičke membrane ćelijske sinapse. Ispunjen je međućelijskim
matriksom sa obiljem mukopolisaharida i glikoproteina koji mu daju sunđersku
strukturu, pogodnu za difuziju neurotransmitera sa sinaptičke ka
postsinaptičkoj membrani( moguća je i u suprotnom smeru, ali to je ređi

by J.M., Ninche, Sandra, Ranagol, gen2010
slučaj).
Postsinaptička membrane –sadrži receptore (stalno se obnavljaju i njihov broj se menja u zavisnosti odaktivnosti neurona).
Sinapse su kontakt između dve nervne ćelije, preko kojeg se prenosi specifična nervna informacija. Postoje
3 vrste sinapsi:
1. Hemijska sinapsa: prenošenje informacija se vrši sa neurotransmiterima. Češće su od električnih
sinapsi. Kako radi: povećava se akcioni potencijal i nervni impuls dolazi do presinaptičke ćelije. Na
njenim krajevima su vezikule, iz kojih se izlučuju neurotransmiteri. Neurotransmiteri ulaze u
membranu postsinaptičke ćelije, i izazivaju reakciju.
2. Električna sinapsa: dominiraju posebno kod beskičmenjaka. Sprovode brze informacije. Nema
neurotransmitera! Da ih nema, gubili bi smo ravnotežu.
3. Mešovita sinapsa
Nervni signali se prenose sa jednog neurona na drugi preko interneuralnih spojeva koji se
nazivaju sinapse. One su funkcionalne jedinice nervnog sistema specijalizovane za prenos
biostruja. Po načinu prenosa biostruja mogu biti hemijske i električne.
Kod ljudi su uglavom sve sinapse koje služe za prenos signala u CNS hemijske sinapse.
U njima prvi neuron sekretuje u sinapsu hemijsku supstancu nazvanu neurotransmiter. Ovaj
neurotransmiter zatim deluje na receptorske proteine u membrani sledećeg neurona i može da ga
ekscitira, ili inhibira ili pak da modifikuje njegovu osetljivost na neki drugi način. 80-90% ovih
sinapsi je na dendritima ekscitatirnog tipa, a 10-20% ja na telu neurona, inhibitomog tipa. Imaju
jednu izrazito važnu karakteristiku koja ih čini veoma pogodnim za prenos signala u CNS: one
uvek prenose signale u jednom smeru, od neurona koji sekretuje transmiter, nazvan presinaptički
neuron, do neurona na koji transmiter deluje nazvan postsinaptički neuron. To je princip
jednosmernog provođenja kroz hemijske sinapse, sasvim drugačiji od provođenja kroz električne
sinapse koje mogu provoditi signal u oba smera. Delovi:
1. Presinaptički deo - deo plazma membrane koji prekriva površinu sinaptičkog
završetka presinaptičkog neurona. U njoj se nalaze brojni voltažno zavisni kanali za Na, Cl, K,
zatim kalcijumove, natrijum-kalcijumove i natrijum-kalijumove pumpe, receptori za
neurotransmitere i transmiteri potrebni za reutilizaciju transmitera.
2. Sinaptička pukotina - ekstracelulama tečnost, slična likvoru, pogodna za difuziju
neurotransmitera sa presinaptičke ka postsinaptičkoj membrani.
3. Postsinaptička membrana - sadrži receptore ( stalno se obnavljaju i njihov broj se
menja u zavisnosti od aktivnosti neurona).
Električne sinapse karakteriše postojanje direktnih kanala za provođenje električne struje

by J.M., Ninche, Sandra, Ranagol, gen2010
sa jednog neurona na drugi. Većinu tih kanala grade mali proteini tubulame strukture nazvani
pukotinaste veze koje dozvoljavaju slobodno kretanje jona od unutrašnjosti jedne ćelije ka
sledećoj. U CNS je nađeno nekoliko pukotinastih veza, ali njihovo značenje nije poznato. S
druge strane u viscelarnim glatkim mišićima akcioni potencijali se prenose sa jednog glatkog
mišićnog vlakna na drugi i preko pukotinastih veza.
sinapsa: osnovni kontakt između 2 neurona, način prenošenja informacija neuroglijalne ćelije su u stalnoj vezi međusobno 2 vrste sinapse:
o brze: bez stvaranja akcionog potencijalao hemijske sinapse
neurotransmiter ima tačni oblik u receptoru, uklapa se u receptor kao ključ u bravu receptori žive oko 8 dana. Izbacuju se kao rogovi puža. Akcioni potencijal (AP) ima amplitudu→ Ca++ jon→ izlučenje (difuzija) neurotransmitera→
vezuje se za receptor (sve ovo traje oko 1ms)→ 2 vrste:o Kanalotrpone Na+ jon AKTIVIRAN→ dobija se depolarizacija od 1 mV, i ovo se zove
APSP (aktivirajući post sinaptički potencijal)o Metabotropne (menjaju metabolizam, INHIBIRAN) K+ izlazi , Cl jon ulazi, i ovo se zove
IPSP (Inhibitorni post sinaptički potencijal) sve ovo brzo traje neurotransmiter treba i skoniti sa receptora
o razgrađuje se sa enzimimao može i da se vrati nazado ide u glijalne ćelije, poslužiće kao rezerva za ubuduće
dok je tamo (gde?) Ca++ non-stop se ispuštaju transmiteri. Dakle, treba se rešiti Ca++. Bitno jesve vratiti na staro.
27. Postsinaptička membrana i procesi na njojTransmisija sa presinaptičke ćelije na postsinaptičku ćeliju je jednosmerna! Postsinaptička membrana je deo
plazma membrane postsinaptičke ćelije. Poseduje aktivne zone sa velikom gustinom, jonske kanale i između
ostalog: receptore. Po vezivanju odgovarajućeg neurotransmitera, receptori aktiviraju primarne/sekundarne
kanale (aktiviraju se jonski kanali) Ako se otvore kanali Na = depolarizacija. Ako K+ joni izlaze, a ulaze Cl-
joni= hiperpolarizacija. Procesi:
1. Jednosmerna transmisija (Egzocitoza: pražnjenje neurotransmitera)
2. Postsinaptički potencijal (inhibitorni, ekscitatorni)
3. Sumacija potencijala (prostorni i vremenski)
4. Adaptacija
5. Zamor
6. Rentilizacija (neurotransmiteri se vraćaju u sinaptički završetak, vrši se razgradnja)

by J.M., Ninche, Sandra, Ranagol, gen2010
Električne sinapse (porozne, pukotinaste veze, cevaste veze ili međućelijski mostovi)- su mesta namembrani na kojima jonski kanali jedne ćelije direktno naležu na jonske kanale druge ćelije, krozmeđućelijski prostor, ne širi od 2-3 nm.
Karakteriše ih postojanje direktnih kanala za provođenje el.struje sa jednog na drugi neuron. Većinu tihkanala grade mali proteini tubularne strukture nazvani pukotinaste veze koje dozvoljavaju slobodno kretanjejona od unutrašnjosti jedne ćelije ka sledećoj.
Jonske struje (depolarizacione i hiperpolarizacione) se prenose direktno sa membrane jedne na membranudruge ćelije a joni direktno difunduju kroz njihove kanale bez prethodnog prolaska kroz međućelijski matriks,tako da je tu otpor proticanju jonske struje oko 400 puta manji nego na ostalom delu plazma membrane.Električnim sinapsama nisu neophodni neurotransmiteri, u njima nema sinaptičkog zadržavanja i prenosbiostruje je trenutan. Ovde se radi o tzv efaptičkom ili elektroničkom provođenju biostruja, kod koga suposrednici sami joni. Električne sinapse ne poseduju sinaptičku pukotinu, budući da uglavnom provodeimpulse dvosmerno, ne može se jasno definisati šta je njihova presinaptička, a šta postsinaptičkamembrana. Za razliku od hemijskih sinapsi, koje mogu biti ekscitacijske ili inhibicijske, električne sinapse sugotovo isključivo ekscitacijske , jer retko prenose hiperpolarizaciju.
Električne sinapse nazivamo i neksusima, jer su njihovi pojedinačni međućelijski mostovi (koneksoni)izgrađeni od 6 proteinskih subjedinica – koneksina. Koneksoni jedne ćelije adheriraju na koneksone susedne,formirajući tunel sa centralnom porom koja se zatvara prilikom porasta koncentracije kalcijumovih jona ućeliji, pri padu ph vrednosti i promeni voltaže plazma membrane.
El.sinapse mogu provoditi signal u oba smera za razliku od hemijskih.
Broj električnih sinapsi je manji od broja hemijskih sinapsi, a smatra se da ih ukupno ima manje od1%. Ove sinapse postoje samo u delovima koji brzo, sinhronizovano okidaju , kao što su: lateralnavestibularna jedra, jedra malog mozga, hipotalamusna suprahijazmatska i jedra zaduzena za sekrecijuodgovarajućih rilizing hormona, inspiratorni, apneustički, presorni centar, centri za rad srca, centar za glad, ineki neuroni mrežnjače. Sve ćelije ovih delova povezane su električnim sinapsama tako da grade električnisincicijum.
Post sinaptička membrana je deo plazma membrane postsinaptičcke ćelije koji pripada hemijskoj sinapsi.Ona poseduje aktivne zone sa velikom gustinom jonskih kanala, jonskih pumpi i receptora zaneurotransmiter. Broj funkcionalnih receptora za neurotransmitter vrlo je promenljiv a regulisan je Up/downmehanizmom. To znači da je senzitivnost postsinaptičke membrane smanjena kada su receptori zaneurotransmiter uvučeni (down), a pojačana je kad su oni istaknuti u membrane (Up). Po vezivanju

by J.M., Ninche, Sandra, Ranagol, gen2010
odgovarajućeg transmitera, receptori postsinaptičke membrane aktiviraju primarne ili sekundarne ligand-zavisne jonske kanale. Kod brzih sinapsi neurotransmiter se vezuje za receptor koji je istovremeno primarniligand-zavisni jonski kanal, tj jonotropni receptor. Spore sinapse koriste neuropeptidne neurotransmitere, kojinisu u stanju da sami otvore ili zatvore odgovarajući jonski kanal već to rade indirektno, preko metabotropnihreceptora, aktivacijom G-proteina i odgovarajućeg intracelularnog mesendžera. Na ovaj način otvoreni jonskikanali mogu duže da ostanu aktivni, tako da obezbeđuju trajanje sinaptičke transmisije sekundama,minutima, ili čak satima.
'Postsinaptička membrana je deo plazma membrane postsinaptičke ćelije koji pripada
hemijskoj sinapsi. Ona poseduje aktivne zone sa velikom gustinom jonskih kanala, jonskih
pumpi i receptora za neurotransmiter. Broj funkcionalnih receptora za neurotransmiter vrlo je
promenljiv a regulisan je up-down mehanizmom, što znači da je senzitivnost postsinaptičke
membrane smanjena kada su receptori za neurotransmiter uvučeni, a pojačana je kad su oni
istaknuti. Po vezivanju odgovarajućeg neurotransmitera receptori postsinaptičke membrane
aktivirju primame ili sekudame ligand-zavisne jonske kanale. Spore sinapse koriste
neuropeptidne neurotransmitere, koji nisu u stanju sami otvore ili zatvore odgovarajući jonski
kanal već to rade indirektno, aktivacijom odgovarajućeg sekundarnog menadžera. Na ovaj način
jonski kanali mogu duže da ostanu aktivni, pa obezbeđuju trajanje sinaptičke transmisije
sekundama, minutima ili čak satima.
U zavisnosti od vrste receptora za neurotransmiter u njihovoj postsinaptičkoj membrani, a
sa aspekta uticaja na sinaptičku transmisiju centralne hemijske sinapse mogu biti: ekscitacijske
ili inhibijske. U CNS vlada pravilo da se ekscitacijske sinapse nalaze na dendritima dok su
inhibicijske sinapse pretežno locirane na somi i aksonu.
Ekscitacijske ili sinapse tipa l - posle vezivanja neurotransmitera za receptore pojačava
se propustljivost njihove njihove postsinaptičke membrane za Na i/ili Cl jone a u nekim
slučajevima se zatvaraju sporo-cureći kalijumovi i/ili hloridni kanali. Usled toga javlja se
prolazna lokalna hipopolarizacija, odnosno EPSP. On približava potencijal postsinaptičke
membrane pražnoj vrednosti i tako olakšava okidanje postsinaptičkog neurona; Međutim jedan
EPSP ne može izazvati okidanje jer kratko traje i ima malu voltažu. Zato je neophodno da se
EPSP-i postsinaptičke membrane sumiraju, jer samo tako integrisani imaju šanse da dovedu do
nastanka A.P.
lnhibicijske ili sinapse tipa Il - po vezivanju neurotransmitera za receptore javlja se
prolazna lokalna hiperpolarizacija postsinaptičke membrane, odnosno IPSP, jer neurotransmiter
pojačava propustljivost ligand-zavisnih hloridnih i kalijumovih kanala ili pak blokira ulazak Na
i/ili Cl u postsinaptički neuron. Zato IPSP manje ili jače otežavaju dalji prenos biostruja.
Akcioni potencijal (AP) ima amplitudu→ Ca++ jon→ izlučenje (difuzija) neurotransmitera→vezuje se za receptor (sve ovo traje oko 1ms)→ 2 vrste:
o Kanalotrpone Na+ jon AKTIVIRAN→ dobija se depolarizacija od 1 mV, i ovo se zoveAPSP (aktivirajući post sinaptički potencijal)
o Metabotropne (menjaju metabolizam, INHIBIRAN) K+ izlazi , Cl jon ulazi, i ovo se zoveIPSP (Inhibitorni post sinaptički potencijal)
sve ovo brzo traje neurotransmiter treba i skoniti sa receptora

by J.M., Ninche, Sandra, Ranagol, gen2010
o razgrađuje se sa enzimimao može i da se vrati nazado ide u glijalne ćelije, poslužiće kao rezerva za ubuduće
dok je tamo (gde?) Ca++ non-stop se ispuštaju transmiteri. Dakle, treba se rešiti Ca++. Bitno jesve vratiti na staro.
28. Hemijske sinapse (vrste i uloge brzo - dejstvujućih transmitera)Karakteristike hemijskih sinapsi:
1. Jednosmerno provođenje
2. Sinapsa je jako zamorljiva
3. Fino doziranje informacija
4. Usporavanje procesa prenošenja (0,5 ms) u odnosu na akson→kroz akson bukvalno pretrči
informacija
5. Velika osetljivost na smanjivanje kiseonika
6. Alkaloza (izbacivanjem CO2 izdisanjem dolazi do alkaloze→ povećava razgradljivost
7. Zakiseljavanje krvi ACIDOZA
Neurotransmiteri se dele na dve grupe:
1. Brzo dejstvujući transmiteri
a. su neurotransmiteri prvog reda
b. to su male molekule, i deluju za 10-15ms
c. 80% prenosa se radi sa ovom vrstom
d. zadatak: funkcionisanje organizma
2. Sporo dejtvujući transmiteri
Neurotransmiteri su jedinjenja koja se oslobađaju iz nervnih završetaka pod uticajem
akcionih potencijala i stupaju u interakciju sa odgovarajćim receptorima okolnih struktura kako
bi proizvela specifične fiziološke odgovore. Postoje dve grupe transmitera: neurotransmiteri i
neuropeptidi. .
Neurotransmiteri su sinaptički transmiteri male molekulkse mase koji brzo deluju. Oni se
sintetišu direktno u nervnom završetku (sinaptički završetak) i skladište u sinaptičkim
vezikulama.
1. Acetil - holin -je neurotransmiter perifernog i centralnog nervnog sistema. Uglavnom
ima ekscitatorni efekat, ali na nekim perifernim parasimpatičkim nervnim završecima može

by J.M., Ninche, Sandra, Ranagol, gen2010
imati i inhibitorno dejstvo. Propadanje holinergičkih neurona u centralnom sistemu povezano
je sa nastankom Alchajmerove bolesti, odnosno demencije.
-aktivirajuće dejstvo, bitan je za pamćenje
2. Amini:
- noradrenalin - učestvuje u sinaptičkoj transmisiji između postganglijskih vlakana
sinaptičkog dela vegetativnog NS i glatkih mišića, tj. efektora. Pomoću njega vrši se i
sinaptička transmisija u kičmenoj moždini i mozgu. Prisutni su i u moždanoj strukturi i u nižim
delovima moždanosg stabla. Od noradrenalina zavise mnoge funkcije CNS kao što su
raspoloženje, stepen živahnosti organizma, memorija i raspoloženje→dobro raspoloženje
- dopamin - je uglavnom neurotransmiter u okviru bazalnih ganglija, mada ga ima u
olfaktornom bulbusu, u refleksnom luku koji reguliše povraćanje i dr. Parkinsonova bolest
je uslovljena smanjenom koncentracijom dopamina u mozgu, a šizofrenija povećanjem
dopamina u mozgu.→stanje budnosti
- adrenalin - poboljšava raspoloženje (ako se nalazi van CNS kao noradrenalin luči ga srž
nadbubrega, ali tada funkcioniše kao hormon)
- histamin - neuroni koji sadrže histamin nalaze se u nekim jedrima hipotalamusa, njegov
promet se menja u stresnim situacijama - pojačava budnost
- serotonin - može da bude i ekscitami i inhibitomi transmiter, nalazi se u moždanom stablu,
smanjenje izaziva depresivnost, serotonin izaziva dobro raspoloženje, pojačan apetit,
budnost i podstiče sve seksualne funkcije. →dobro raspoloženje
3. Aminokiseline (mogu biti inhibirajuće kao što su GABA i Glicin, ili aktivirajuće kao Glutamat, Aspartat)
- gama-aminobuterna kiselina je isključivo inhibitorni neurotransmiter i najčešći je
inhibitorni neurotransmiter u mozgu, receptori su GABA A i GABA B (inhibišu ih
benzodijazepini)
- glicin -jedan od inhibitomih neurotransmitera kičmene moždine, produžene moždine i
moždanog stabla.
- glutamat - ekscítator, ubrzava ćelijsku smrt, neurotransmiter za brzi bol
4. Gasoviti transmiteri
- azot - oksid - ekscitator i vazodilatator
Brzodejstvujući neurotransmiteri imaju male molekule i spadaju u neurotransmitere prvog reda (osnovni su).Deluju u roku od 10-15msek. 80 do 90 odsto neurotransmitera spadaju u ovu grupu.
1.Acetilholin - prvootkriveni neurotransmiter.

by J.M., Ninche, Sandra, Ranagol, gen2010
Pomoću njega se vrši transmisija nervnih impulsa sa svih postganglijskih nervnih vlakana parasimpatičkogdela autonomnog nervnog sistema na efektore. Ovaj neurotransmiter se izlučuje i na završecima svihpreganglijskih vlakana autonomnog sistema i na taj način se pomoću acetilholina vrši transmisija nervnihimpulsa u svim autonomnim ganglijama. Sva nervna vlakna koja na svojim završecima izlučuju ACH nazivajuse holinergična. ACH je
neurotransmiter kojim se vrši sinapsa u mnogim delovima CNS-a, neuromišićna sinapsa, u mnogimmoždanim strukturama. Kao i drugi neurotransmiteri i ACH se pod dejstvom nervnog impulsa oslobađa izsinaptičkih vezikula, difunduje u sinaptički prostor i pošto se veže za postsinaptičku membranu, odnosno zaACH receptor na njoj, on ostvari svoje dejstvo koje se sastoji u aktiviranju jonskih kanala, propustljivih zajednovalentne katjone NA i K+. Ako dolazi do lokalne promene polarizovanosti, odnosno stvaranjasinaptičkog potencijala ACH, kao ni drugi neurotransmiteri on ne ostaje trajno vezan za receptorskemolekule postsinaptičke membrane. On se posle izvršenog dejstva otklanja kako bi postinaptička membranamogla da se aktivira pod dejstvom sledećeg nervnog impulsa. Kod holinergičnih sinapsi, enzimskadegradacij, je osnovni mehanizam pomoću koga se otklanja ACH receptor. Receptorski molekuli zaacetilholin nisu sasvim identični kod svih holinergičnih sinapsi. Postoje tzv. nikotinski receptori (npr. receptorimotorne ploče) i muskarinski receptori. Dakle, receptori nisu uvek istovetni i ta raznolikost uslovljava razlikeu efektu holinergične transmisije. Procesi pamćenja i učenja. Alzheimerova bolest - posledica degeneracijeholinergičnih neurona.
2.Biogeni amin (monoamin)
a) noradrenalin - učestvuje u sinaptičkoj transmisiji između postaganglijskih vlakana sinaptičkog dela VNS iglatkih mišića, tj. efektora. Pomoću noradrenalina vrši se sinaptička transmisija u kičmenoj mozdini i mozgu.Prisutni su i u moždanoj strukturi i u nižim delovima moždanog stabla. Od noradrenalina zavise mnogefunkcije CNS (zavisi od raspoloženja i živahnosti organizma, balans memorije i raspoloženja).
b) dopamin - neuroni koji sadrže ovaj neurotransmiter nalaze se u 3 moždana regiona:
-nigro - strialni sistem
-mezolimbički sistem
-hipotalamus
Sinaptičke ganglije VNS-a sadrže dopamin kao i retina a pomoću njega se vrši
sinaptička transmisija u pojedinim delovima nervnog sistema. Emocionalno ponašanje, raspoloženje,motorika, pokreti. Parkinsonova bolest je uslovljena smanjenom koncentracijom dopamina u mozgu, ašizofrenija povećanjem dopamina u mozgu.
c) adrenalin - u CNS-u (pojedinim delovima mozga) utiče na stepen aktiviteta. Ako se nalazi van CNS-a, kaoi noradrenalin, luči ga srž nadbubrega, ali tada funkcioniše kao hormon.
d) histamin - Neuroni koji sadrže histamin nalaze se u nekim jedrima hipotalamusa. Njegov promet se menjau stresnim situacijama- povećava budnost.

by J.M., Ninche, Sandra, Ranagol, gen2010
e) serotonin - može da bude i ekscitatorni i inhibitorni neurotransmiter. Nalazi se u moždanom stablu.Smanjenje izaziva depresiju – emocionalno stanje, spavanje, budnost, doživljaj bola, pojačava apetit,podstiče seksualne funkcije, podstiče libido.
3. Aminokiseline
Se dele na aktivirajuće i inhibirajuće. Aktivirajuće su glutamat (on se čuva u glijalnim ćelijama), aspartat (imatri vrste receptora).
Inhibirajuće su:
a) GABA- aminobuterna kiselina- je osnovni inhibitorni transmiter, centralnog nervnog sistema; nalazi se uneuronima bazalnih ganglija koji se projektuju na supstancu nigru, a i inhibitorni neuroni malog mozga sugabaminergični. Na GABA deluju alkoholi i enzodijazepini koji potenciraju inhibitorno dejstvo.
b) Glicin jedan od inhibitornih transmitera kičmene moždine produžene moždine i moždanog stabla.
c) Glutamat - ekscitatorni neurotransmiter (takvo dejstvo se pripisuje i aspartatu koji je sličan glutamatu).Prisutan u kičmenoj moždini, kod određenih neurona malog mozga, a vezuje se i za specifične receptorerazličcitih moždanih struktura (hipokampusa). S obzirom na moguću ulogu hipokampusa u pamćenju, efektuglutamata se posvećuje posebna pažnja.
Najpotentniji neurotoksin prilikom trauma na mozgu.
- Gasoviti transmiteri
Azot oksid - aktivirajuće dejstvo.
29. Hemijske sinapse (vrste i uloge sporo - dejstvujućih transmitera)-neuropeptidi
Neurotransmiteri se dele na dve grupe:
1. Brzo dejstvujući transmiteri
2. Sporo dejtvujući transmiteri
a. peptidi
b. mogu delovati minutima, satima, danima
c. prenose se aksonskim transportom, zato se i zovu sekundarni neurotransmiteri (ali isto
su toliko važni, kao primarni neurotransmiteri)
d. kada odrade posao, razgrađuju se
e. ovi neurotransmiteri daju individualnost, dok su primarni transmiteri zaduženi za
funkcionisanje organizma
i. Oksitocin: hormon koji dovodi do porođaja, kontrakcije materice, i koja je
odgovorna za empatiju (jako je bitna za empatiju), ali i za izgradnju duboke

by J.M., Ninche, Sandra, Ranagol, gen2010
emocionalne veze između majke i deteta
ii.Vazopresin: utiče na pamćenje
iii. Adenozin: dovodi do spavanja. Peptidni hormon, i ako se blokiraju njegovi
receptori (npr. sa kofeinom), onda se ne može spavati.
iv. Supstanca P: ?
Neuropeptidi su sasvim različita grupa transmitera. Njihova sinteza je različita, a dejstva
su obično spora, a i u drugim pogledima sasvim različita od onih koje izazivaju malomolekulski
transmiteri. Neuropeptidi se sintetišu kao integralni deo velikih proteinskih molekula na
ribozomima u telu nervne ćelije. Proteinski molekuli se odmah transportuju u endoplazmatski
retikulum tela nervne ćelije, zatim endoplazmatski retikulum, a zatim Goldžijev aparat delujući
zajedno ostvaruju dva procesa. Prvo, enzimski cepaju prvobitni protein u manje fragmente i
potom ih oslobađaju bilo kao sam neuropeptid, bilo kao njegov prekusor. Drugo, Goldžijev
aparat pakuje neuropeptid u minijaturne transmiterske vezikule koje se oslobađaju u citoplazmu.
Potom se transmiterske vezikule transportuju celim putem do samih krajeva nervnih vlakana.
Nošene strujanjem aksoplazme putuju sporo, svega nekoliko centimetara na dan. Na kraju ove
vezikule oslobađaju svoj transmiter kao odgovor na A.P., ali se vezikula ponovo ne koristi već
nastupa autoliza. Zbog ovakvog težeg načina stvaranja neuropeptida, obično se mnogo manja
količina oslobađa u odnosu na količinu malomolekulskih transmitera. Međutim to se delimično
kompenzuje činjenicom da su neuropeptidi uopšteno hiljadu i više puta potentniji od
malomolekulskih transmitera, a imaju i znatno duže dejstvo. Neuropeptidi se klasiñkuju prema
mestu sekrecije, pa tako postoje:
l. Hipotalamusni - tireotropin, somatostatin
2. Hipofizni - beta - endorfin, prolaktin, hormon rasta, vazopersin, oksitocin, tireotropní hormon
3.Peptidi koji deluju u crevima i mozgu - supstanca P, insulin, neurotenzin, glukagon, leucin-
enkefalin
4.Iz drugih tkiva - karnozin, peptidi spavanja, bradikinín, kalcitonin
29. Hemijske sinapse (vrste i uloge sporo-dejstvujućih transmitera)
Neuropeptidi se sintetišu kao integralni deo velikih proteinskih molekula na ribozomima u telu nervne ćelije.Proteinski molekuli se odmah transportuju u Endoplazmatski retikulum tela nervne ćelije, zatim Goldžijevaparat delujući zajedno ostvaruju dva procesa. Prvo enzimski cepaju prvobitni protein u manje fragmente ipotom ih oslobađaju kao sam neuropeptid ili kao njegov prekursor. Drugo, Goldžijev aparat pakujeneuropeptid u minijaturne transmitorske vezikule koje se oslobađaju u citoplazmu. Potom se transmiterskevezikule transportuju celim putem do samih krajeva nervnih vlakana. Nošene strujanjem aksoplazme putujusporo, svega nekoliko centimetara na dan. Na kraju ove vezikule oslobađaju svoj transmitter kao odgovor naAP, ali se vezikula ponovo ne koristi već nastupa autoliza. Zbog ovakvog težeg načina stvaranjaneuropeptida, obično se mnogo manja količina oslobađa u odnosu na količinu malomolekulskih transmitera.Međutim to se delimično kompenzuje činjenicom da su neuroprptidi uopšteno 1000 i više puta potentniji odmalomolekulskih transmitera, a imaju i znatno duže dejstvo. Imaju specifične puteve, stvaraju se uneuronima a prenose se aksonskim transportom, i zovu se sekundarni neurotransmiteri. Neuron ih može

by J.M., Ninche, Sandra, Ranagol, gen2010
stvoriti od 10 do 20, razgrađuju se do aminokiselina. Daju nam individualnost. Sporo-dejstvujućineurotransmiteri imaju velike molekule.
To su neuropeptidi i klasifikuju se prema mestu sekrecije:
1. Hipotalamusni (tu spadaju tireotropin i somatostatin)
2. Hipofizni (tu spadaju beta-endorfin, prolaktin, hormon rasta, vazopresin, oksitocin, tireotropni hormon)
3. Peptidi koji deluju u crevima i mozgu (supstanca P, insulin, neurotenzin, glukagon, leucinenkefalin)
4. Iz drugih tkiva (karnozin, peptidi spavanja, bradikinin i kalcitonin)
Peptidi koji aktivno utiču na nerve nazivaju se neuropeptidi. Njihov efekat na neuron traje duže od efektaneurotransmitera.
- različiti nivoi složenosti peptida
specifičan put - stvaranje protoplazme jedra
Neuromodulatori - izazivaju hiperpolarizaciju - oplemenjivanje brzih efekata
neurotransmitera.
Stvaranje: hipotalamus, hipofiza, digestivni trakt, mozak...
1. Vazopresin - hipofiza, krvni pritisak. Vazopresin ima značajne funkcije za izlučivanje tečnosti i utiče napamćenje.
2. Oksitocin - hipofiza, mentalne funkcije, empatija, pamćenje, afilijaciju
3. Supsanca P - retikularna formacija, hipokampus, kičmena moždina - velike
doze supstance P izazivaju spavanje, anesteziju, a male dovode do vazodilatacije, stimulišu kontrakcijuglatkih mišića tankog creva, bronhija... Supstanca P služi za bolnu recepciju.
4. Opioidi - endogenog porekla su enkefalini, endorfini, dinorfin. Služe kao neurotransmiteri ineuromodulatori. Izazivaju presinaptičku inhibiciju u putevima prenosa bolnih nadražaja.
Adenozin je hormon peptidne vrste koji dovodi do spavanja. Ako se blokiraju njegovi receptori (ima 3 vrstereceptora) ne može se spavati - on se blokira kofeinom.

by J.M. & Ranagol, gen 2010 1
II DEOBroj Nepoznata Pašićka Kratko
pitanja reč "~ a đe je" objašnjenjereceptor 2 detektuje promenu u sredini, i aktiviraju senzitivne neuroneefektori 2 mišićne ili žlezdane ćelije
senzitivni neuron 2 nadovezuje se na receptor, i stupa u kontakt sa motoneuronom
motoneuron 2nadovezuje se na senzitivni neuron, i stupa u kontakt saefektorom
refleksni luk 2Osnovna funkcionalna jedinica nervnog sistema. Čine je:receptor, senzitivni neuroninterneuron, motoneuron, efektor.
interneuron 2Ćelija, koja se umeće između senzitivnog neurona imotoneurona.
ekscitabilnost 15Nervna ćelija je ekscitabilna, odnosno nadražljiva. Provodi nervniimpuls.
ekscitacija aktiviranjeinhibicija kočenje
2-3 aksoplazmatski transport 14 kretanje materijala kroz akson, u cilju regeneracije n. vlakana2-7 amplifikacija 144 povećanje reakcije
EPSP 144 ekscitatorni sinaptički potencijalIPSP 76 Inhibitorni postsinaptički potencijal
prostorna sumacija 144 Istovremena aktivacija dva posebna presinaptička neurona.
vremenska sumacija 145Kada se isti presinaptički neuron aktivira sa većim brojemstimulusa
2-8 paralelna veza 146 crtežoscilatorna veza 146 crtež
2-10 presinaptički neuron pre sinapse
divergencija 68Nervni impuls se prenosi sa jednog presinaptičnog na veći br.postsinaptičnog neurona
postsinaptički neuron posle sinapsesinapsa 66 Mesto gde se stvara veza između dva neurona i efektorne ćelijesinapsa 66 transmisija, prenošenje nervnog impulsa
sinaptička veza 66 Stvara se između presinaptičkog i postsinaptičkog neurona
akso-somatska 72... sinapsa: akson presinaptičkog neurona stupa u vezu sa telompostsinaptičkog neurona.
akso-dendritska 72...sinapsa: aksonski završetak presin. neurona stupa u vezu sadendritem postsin. neurona.
akso-aksonska 72 ...sinapsa: akson se priključuje na aksonneuron nervna ćelija
2-11 konvergencija 68Više presinaptičkih neurona stupaju u kontakt sa jednimpostsinaptičkim neuronomtj., koji konvergiranju na isti postsinaptički neuron
ili: prenošenje nervnog impulsa sa više presinaptičnih na 1postsinaptični neuron
2-12 recipročna inervacija 176Antagonističko dejstvo senzorne stimulacije na dve kategorijeefektora (Šerington)
2-18 inhibicija 151 crtež

by J.M. & Ranagol, gen 2010 2
OBJAŠNJENJE:
1. Times New Roman: materijal iz stare skripte
2. Arial: prepisano iz različitih sveski ili rukopisa
3. Comic Sans MS: materijal sa predavanja
4. Microsoft Sans Serif: skripta gen 2010
1. Mozak kao biokompjuter
Mozak se smatra biokompjuterom jer se funkcije mozga odnose na održavanje homeostaze (oscilovanjevrednosti u skladu sa životnim okolnostima, a kada se poremeti dolazi do alostaze – bolesnog stanja).Homeostaza se održava na sledeće načine :
1. Određuje i reguliše rad svih organa čineći celinu
2. Adaptira naš organizam na stalno promenljive uslove, obuhvatajući najsloženije mentalne funkcije :mišljenje, svest, pažnja
3. Koordinira 1. i 2. funkciju a to znači da sve što mislimo možemo voljno da realizujemo.
Kada su naučnici širom sveta konstruisali prve elektronske računare, vrlo brzo su uočena
mnoga svojstva ovih mašina koja su zajednička sa nervnim sistemom. Prvo, svi oni imaju ulazna
kola koja se mogu uporediti sa senzornim delom nervnog sistema, i izlazna kola koja se porede
sa motornim delom nervnog sistema. U vezama koje postoje između ulaza i izlaza nalaze se
mehanizmi koji obavljaju različite vrste računskih operacija.
U jednostavnim računarima izlazne signale direktno kontrolišu ulazni signali,
funkcionišući na način sličan jednostavnim refleksima kičmene moždine. Međutim, u složenij im
kompjuterima, izlaz određuju zajedno ulazni signali i informacije koje su već uskladištene u
memoriji kompjutera, što je slično višesloženim refleksima i mehanizmu obrade u višim
delovima nervnog sistema. Kako su kompjuteri postali sve komplikovaniji, bilo je neophodno
dodati još jednu jedinicu, nazvanu centralna jedinica za programiranje, koja je određivala
redosled svih operacija. Ova jedinica je analogna mehanizmu u našem mozgu koji nam
dozvoljava da usmerimo svoju pažnju prvo na jednu misao, a zatim na drugu, i tako redom sve
dok ne nastanu složeni redosled misli ili pokret.
Činjenica da osnovne komponente računara za opštu upotrebu odgovaraju onima u
nervnom sistemu ljudi, ukazuje na to da je mozak u osnovi kompjuter koji neprestano skuplja
senzorne informacije koje koristi zajedno sa uskladištenim informacijama za proračun
svakodnevnog toka telesnih aktivnosti.
2. Refleksna delatnost CNS-a Ako učestvuje CNS, onda to nije reflex.
Pojam refleksa je uveo Dekart negde oko 1600

by J.M. & Ranagol, gen 2010 3
1800 Šerington
1904-05 Pavlov je najviše otrkio o refleksima, dobio je Nobelovu nagradu. Po Pavlovu, svaka
aktivnost organizma je refleksna aktivnost. Refleks je odgovor na draži, koje dolaze iz
spoljašnje ili unutrašnje sredine.
Prema jednoj (neispravnoj) definiciji: refleks je nesvesna reakcija na draž u kojoj učestvuje i
nervni sistem. Ovo ne valja, jer su i mentalne aktivnosti refleksi, a mentalne aktivnosti su
svesne. Dakle: postoje SVESNE i NESVESNE reakcije.
Refleksna delatnost ima značajnu ulogu u integracionim procesima funkcionalne
organizacije CNS-a, a odvija se na principu mehanizma refleksnog luka, koji je osnovna
funkcionalna jedinica CNS, od koje zavisi neposredna reakcija organizma na promene sredine.
Kičmena moždina je centar refleksnih reakcija a termin refleks je prvi upotrebio Dekart u XVII
veku. Za ostvarivanje refleksnih reakcija neophodne su sve komponente refleksnog luka:
1. RECEPTOR
2. AFERENTNI PUT
3. CENTAR
4. EFERENTNI PUT
5. EFEKTOR
Refleksne reakcije nisu vezane isključivo za kičmenu moždinu, već se centri za neke od
njih nalaze i u drugim višim delovima CNS. Za više delove CNS-a vezju se složenije, a za niže
delove CNS-a manje složeni refleksi. U tom smislu može se govoriti o hijerarhijskoj organizacij
nervnog sistema.
Refleksno vreme - vreme koje protekne od momenta dejstva draži do pojave refleksne reakcije. Refleksno
vreme je neophodno vreme da se na stimulus dobije odgovor.
Reakciono vreme - vreme koje protekne od momenta zadavanja nekog zadatka do momenta njegovog
izvršenja.
o Prosto reakciono vreme: npr. ispitanik na svetlosni znak mora da pritisne taster.
o Kompleksno (složeno) reakciono vreme: uključen je veći deo nervnog sistema u taj
zadatak. Npr: rešavanje nekog zadatka, gde je uključeno i čitanja i razmišljanje,
odlučivanje.
Složeno izborno reakciono vreme: reakciono vreme koje je potrebno za
donošenje izbora.
Razlika između reakcionog vremena i refleksnog vremena:
o reakciono vreme je širi pojam od refleksnog vremena. Tu učestvuje svest i kora velikog
mozga.
o kod refleksnog vremena nema svesti, i ne učestvuje kora velikog mozga.

by J.M. & Ranagol, gen 2010 4
Refleksi predstavljaju osnovni model funkcionisanja nervnog sistema i osnovne su jedinice integrisanenervne aktivnosti. Predstavljaju reakciju na dejstvo draži uz obavezno učešće nervnog sistema. Definisemoih kao automatske, nesvesne (nevoljne) reakcije efektora koje se javljaju kao odgovor na draženjeodgovarajućih receptora. Zato su refleksi najprimitivniji poznati oblik ponašanja. Izuzev aksonskog refleksa,svi ostali refleksi se realizuju uz posredovanje refleksnog luka koji ima 5 osnovniih komponenti: receptore,aferentni put, centar, eferentni put i efektore, i predstavlja morfološku bazu receptora.
3. Morfo - funkcionalne karakteristike refleksnog lukaRefleksi predstavljaju osnovni model funkcionisanja nervnog sistema. Definišemo ih kao
automatske, nesvesne reakcije efektora koje se javljaju kao odgovor na podraživanje
odgovarajućeg receptora. Refleksi su najprimitivniji poznat oblik ponašanja koji se ostvaruje uz
posredovanje refleksnog luka, koji ima 5 osnovnih komponenti.
1. Receptori se nalaze na samom početku refleksnog luka, a uloga im je da prime adekvatnu draž i prevedu je u
akcioni potencijal.
2. Aferentni (ushodni ili senzorni) put prima te akcione potencijale do
3. centra, koji ih obrađuje i potom šalje informacije koje su odgovor na njih takođe u vidu akcionog potencijala
preko
4. eferentnog (nishodnog ili motornog) puta, ka
5. efektorima koji treba automatski da odreaguju na primljenu draž. Efektori mogu biti:
a. poprečnoprugaste,
b. glatkomišićne,
c. miokardske ili žlezdane ćelije.
Refleksni luk može da bude
1. monosinaptički,
2. polisinaptički ili
3. aksonski.
MONOSINAPTIČKI QPROST) REFLEKSNI LUK - ima svih 5 komponenti, njegovi
receptori i aferentni putevi pripadaju senzomim, eferentni put motornim neuronima, a u centru
postoje direktne sinaptičke komunikacije između ova dva tipa neurona. Prenos akcionog
potencijala je veoma brz, pa su i reakcije kod ovih refleksa najbrže moguće. U organizmu
domaćih životinja i ljudi samo miotatički refleksi imaju monosinptički refleksni luk, dok su svi
ostali refleksi polisinaptički.
POLISINAPTIČKI REFLEKSNI LUK (imaju 2 ili više sinapsi)- osim 5 osnovnih komponenti, ima i
interneurone, čiji broj može da varira od dva do nekoliko stotina. Ovi interneuroni se nalaze u centru i
grade centralne sinapse između senzornih i motomih neurona. .Ukupno vreme centralnog
kašnjenja je veće ukoliko refleksni luk ima više interneurona, a samim tim je i refleksna reakcija sporija.

by J.M. & Ranagol, gen 2010 5
AKSONSKI REFLEKS -je jedinstven po tome što ne funkcioniše pomoću klasičnog
refleksnog luka (nema 5 osnovnih komponenti). On je zapravo najprostiji autonomni (vegetativni
ili viscelarni) refleks, koji nema centralnu integraciju, on je lokalni refleks. Cilj ovog refleksa je
da u vreme delovanja mehaničkih, hemijskih ili inflamatornih nadražaja poveća protok krvi u
podraženom delu kože, pri čemu se smanjuje oštećenje tkiva a samim tim i osećaj bola koji ga
prati.
3. Morfo-funkcionalne karakteristike refleksnog luka
Refleksni luk je osnovna funkcionalna jedinica nervnog sistema od koje zavisi neposredna reakcijaorganizma na promene sredine. Refleksni luk ima 5 osnovnih komponenti: receptore, aferentni put, centar,eferentni put i efektore. Receptori i aferentni put pripadaju senzornim neuronima, eferentni motornimneuronima, a u centrima postoje direktne sinaptičke veze između ova dva tipa neurona.
Receptori se nalaze na samom početku refleksnog luka i predstavljaju specijalno adaptirane nervnećelije. Uloga im je da prime adekvatnu draž i prevedu je u akcione potencijale.
Aferentni (ushodni,senzorni) put prenosi te akcione potencijale od periferije do određenog centra nervnogsistema.
Nervni centar ili sinapsa može biti manje ili više složen. Ako je aktivirana jedna sinapsa, to je monosinaptičkirefleks, a ako je više sinapsi, polisinaptički refleks, koji može da ima puno sinaptičkih kontakata.
Eferentni put spada u motorne neurone. To je put kojim se informacija iz nervnog centra prenosi doperifernog organa (efektora).
Efektori mogu da budu:
-poprečnoprugaste
-glatkomišićne
-miokardske
-žlezdane ćelije
Na osnovu refleksnog luka refleksi mogu da budu monosinaptički ili polisinaptički, a postoji i jedan specifičnireflex koga nazivamo aksonskim.
Monosinapticki (prost ) refleks funkcioniše pomoću monosinaptičkog refleksnog luka, sastavljenog od svih 5navedenih osnovnih komponenti. Njegovi receptori i aferentni put pripadaju senzornim, a eferentni putmotornim neuronima. Monosinaptički refleksni luk je dobio takvo ime jer u centru refleksa njegovi senzornineuroni direktno sinaptički komuniciraju sa motornim neuronima (imeđu njih postoje centralne hemijskesinapse). Vreme sinaptičkog zadrzavanja (centralno kašnjenje) tj. prenos biostruje traje 0.5 do 1.0 ms.Reakcije kod ovog refleksa su najbrže moguće, zbog direktne komunikacije između senzornih i motornihneurona.
U refleksnom luku MR postoje i periferne hemijske sinapse koje povezuju eferentni put sa efektorima nakojima takođe dolazi do sinaptičkog zadržavanja. Ukupno refleksno vreme mr od momenta podraživanja dopojave reakcije iznosi između 9 i 24 ms. U organizmu ljudi i životinja samo miotatički refleksi imaju MR luk,dok su svi ostali refleksi polisinaptički.
Polisinapticki (slozeni) reflex funkcioniše pomoću polisinaptičkog ref luka, koji osim 5 osnovnih komponentiima i interneurone čiji broj varira od 2 do nekoliko stotina. Svi interneuroni se nalaze u centru i gradecentralne hemijske sinapse između senzornih i motornih neurona. Kako broj neurona nije isti, nije isto ninjihovo vreme centralnog kašnjenja-ukupno vreme centralnog kašnjenja je veće ukoliko ref luk ima više

by J.M. & Ranagol, gen 2010 6
interneurona, pa je i refleksna reakcija sporija. Izuzev miotatičkog i aksonskog refleksa svi ostali refleksi uorganizmu ljudi i domaćih životinja su polisinaptički.
Aksonski reflex je jedinstven po tome sto nema 5 osnovnih komponenti. To je najprostiji autonomni(vegetativni, viceralni ) reflex, jedini koji nema centralnu integraciju, pa se može reći da je to lokalnireflex. Aktivira se podraživanjem kožnih receptora za bol i toplotu, čiji se impuls prenosi aferentnim putem(ortodromno) ka kičmenoj moždini. Ovi refleksi poseduju i antidroman prenos dela akcionih potencijala kojine putuju ka kičmenoj moždini, vec skreću sa ushodnog puta, preko kolaterale aferentnog vlakna izavršavaju na krvnim sudovima dela kože čciji su nociceptori i termoreceptori podraženi.
Aksonski reflex se manifestuje vazodilatacijom krvnih sudova i porastom kapilarne propustljivosti upodraženom delu kože. Ovi efektori nastaju tako što završetak antidromne aksonske grane podstiče susednemastocite na oslobađanje histamina i supstance P, što prouzrokuje pojavu lokalnog crvenila.
Cilj ovog refleksa je da u vreme delovanja mehaničkih, hemijskih ili inflamatornih draži poveća dotok krvi upodraženom delu kože i da tako u njemu intenzivira, ne samo proizvodnju nego I drenažu međućelijsketečnosti.
Refleksno vreme je najkraće vreme koje protekne od momenta dejstva stimulusa do početka refleksnereakcije, njegova dužina zavisi od toga koliko je sinapsi u CNS-u uključeno u obradu i ne mora obavezno dauključuje svest.
Reakciono vreme je složeniji proces, i predstavlja vreme koje protekne od momenta zadavanja zadatka domomenta njegovog izvršenja. Uključuje obavezno našu svest, koru hemisfere velikog mozga. Može bitiprosto i složeno. Prosto se ispituje preko reakciometra i ono predstavlja reakciju ispitanika na nekijednostavan zadatak. Složeno se dobija kada se ispitaniku daje zadatak da izvrši diferencijaciju određenogstimulusa i samo na njega reaguje.
Refleksni luk je nervni put koji nadražaj pređe od mesta delovanja draži preko senzitivnih nerava do sivemase kičmene moždine i potom preko motornih nerava do efektornih organa. S obzirom da se kod reflesnogluka impulsi ne prenose do centara u mozgu, potrebno je da protekne manje vremena od momentadelovanja draži do reakcije. To omogućava osobi da se brzo i bez uticaja volje prilagodi na promene uokruženju, ali takođe pomaže i regulaciju procesa unutar organizma što doprinosi očuvanju homeostaze.
Glavne komponente refleksnog luka su:
receptor (koji preko čulnih ćelija prima nadražaj i pretvara ga u akcioni potencijal),
aferentno nervno vlakno (koje prenosi akcioni potencijal od receptora do refleksnog centra),
centar refleksne radnje (u kome aferentno vlakno gradi sinapsu sa eferentnim vlaknom iliinterneuronom),
eferentno nervno vlakno (koje prenosi nervni impuls do odgovarajućeg efektora) i
efektor (mišić ili žlezda).
Dve osnovne vrste refleksnog luka su monosinaptički i polisinaptički luk.
Kod monosinaptičkog luka ne postoji interneuron, odnosno sinapsa se uspostavlja između senzitivnog imotornog neurona. Ovakav refleksni luk se sreće kod patelarnog refleksa, gde se kao odgovor na udaracčekićom u predeo ispod čašice kolena javlja trzaj noge.
Kod polisinaptičkog luka postoji jedan ili više neurona između aferentnog i eferentnog neurona. Na taj načinse ostvaruje stimulacija ili inhibicija refleksne radnje prilikom prelaska signala od aferentnog premaeferentnom vlaknu unutar kičmene moždine.

by J.M. & Ranagol, gen 2010 7
4. Klasifikacije refleksaPo načinu formiranja
a) Urođeni (bezuslovni) refleksi - sisanje , gutanje, lučenje pluvačke, kijanje, kašalj, plač, mokrenje. Ovo su
- jednostavniji refleksi,
- javljaju se pri rođenju,
- karakteristično je za datu vrstu.
- Zadatak: održavanje jedinke i vrste.
b) Stečeni (uslovni) refleksi - uče se tokom šivota, npr. svesna regulacije mikcije i defekacije. Individualni su,
karakteristično je za jedinku. Vrste uslovnih refleksa (što je jednako=vrste učenja)
1. Klasično uslovljavanje: adaptiranje na stalno promenljive uslove sredine
2. Instrumentalno uslovljavanje:
a. mora da postoji motiv
b. jedinka sama preduzima akciju
c. učenje na greškama i pokušajima
Podela na osnovu efektora koji učestvuju u refleksnoj reakciji
a) Somatski (spinalni) - efektori su poprečno-prugasti mišići, a efekat je pomeranje nekog
dela tela, npr. pateralni i Ahilov refleks. Nazivaju se još i spinalni jer im se najniži centri
nalaze u sivoj masi kičmene moždine
b) Viscelarni- podrazumevaju refleksne reakcije unutrašnjih organa, efektori su
glatkomišićne ili žlezdane ćelije, npr. pupilami, kamelami, refleks, kašalj, kijanje,
lučenje pljuvačke i želudačnog soka. U viscelame reflekse spadaju još i neuro - endokrini
refleksi kojim se oslobađaju odgovarajuđi hormoni iz endokrine žlezdane ćelije ( refleks
naviranja mleka).
Podela po biološkoj osnovi
a) lshrambeni refleksi- sve refleksne reakcije povezane sa ishranom: uzimanje hrane, varenje, izbacivanje nepotrebnog
materijala. Poremećaji: anoreksija, bulemija, prejedanje.
b) Odbrambeni refleksi- brane nas od štetnih uticaja, štite život, npr.: bežanje ili agresija
c) Polni refleksi- vezane za ostvarivanje polnog akta, podizanje dece
d) Orientacioni - aktiviranje kada se nešto desi neočekivano. OIR= Orientaciono istraživački refleks.
e) Statokinetički refleks - za održavanje ravnoteže, adekvatni pokreti.
i) Homeostazni - obezbeđuju da organizam zadrži stalnost svog unutrašnjeg sastava
Podela po tome gde se nalazi centar
a) U kičmenoj moždini - najjednostavniji refleksi .

by J.M. & Ranagol, gen 2010 8
b) U produženoj moždini - žvakanje, gutanje, disanje, treptanje , povraćanje
c) U srednjem mozgu - regulacija širine zenice
d) U međumozgu - održavanje telesne težine, soli, metabolizam, reakcije vezane za emocije
e) U kori velikog mozga - sve svesne reakcije
Podela po tome, gde se nalazi receptor
a) Eksteroreceptivni - zvuk, svetlost, miris, ukus
b) lnteroreceptivni - refleksi unutrašnjih organa
c) Proprioreceptivni - u mišićima, tetivama i zglobovima
Podela po tome kakav efekat dobijamo
a) Motorni
b) Vazomotorni - realizuju se proširenjem ili suženjem krvnih sudova
c) Sekretorni - sekrecija žlezda
5. Pojam receptornog polja
Uža definicija – skup neurona i sinapsi u mozgu gde stimulacije dovode do verifikovane reakcije organizma.
Šira definicija – sve sinapse i neuroni koji su na bilo koji način uključeni u neki vid ponašanja. Postoji velikibroj centara zbog velike količine informacije. Receptivno polje čini deo receptora koji aktivira ili inhibiraodređen neuron. Receptivna polja pojedinačnih neurona mogu biti različite veličine i ona se mogu preklapatisa poljima drugih neurona tako da isti deo receptora, tj. pojedine delove receptora može „videti“ veći brojneurona. Naročito detaljno su analizirana receptivna polja neurona genikulatnog tela. Ustanovljeno je da senervna vlakna gangliskih neurona koja u sastavu optičkog nerva dospevaju do talamusa, raspoređujusistematski na 6 slojeva neurona od kojih neki primaju aksone iz ipsilateralnog, a drugi iz kontralateralnogoka. Sem toga nije ustanovljeno da pojedinačnim neuronima pripadaju određeni kružni delići retine koji moguimati veće ili manje dimenzije. Tako su receptivna polja talamičkih neurona slična onima koje imaju ganglijskineuroni same retine. I funkcionalna organizacija je slična kao kod nekih genikulatnih neurona, osvetljivanjesredišta izaziva ekscitaciju, a osvetjlivanje perifernog polja inhibira spontanu bioelektričnu aktivnost. Dakle,ako svetlost prodre u centar receptivnog polja, a periferni deo ostane u mraku, neuron se maksimalnoekscitira. Na taj način ako se receptivno polje osvetli tačkom na tamnoj podlozi dolazi do maksimalneeksitacije talamičnog neurona.
Značaj receptivnog polja je u tome što do promene spontana bioelektrične aktivnosti dolazi samo onda kadase stimulus primeni na receptorno polje. Receptorna polja su organizovana po principu centar – periferija, štoznači da će u zavisnosti od toga koji deo receptivnog polja se nadražuje doći do ekscitacije tj inhibicije paposledično do povećanja ili smanjenja frekvencije nervnih impulsa.
Receptomo polje je deo receptora koji aktivira ili inhibia određeni neuron. Receptivo
polje pojedinačnih neurona mogu biti različite veličine i mogu se preklapati sa poljima drugih
neurona, tako da isti deo receptora, odnosno pojedine delove receptora može “videti” veći broj
neurona.

by J.M. & Ranagol, gen 2010 9
Značaj receptivnog polja je u tome što do promene spontane bioelektrične aktivnosti
dolazi samo onda kada se stimulus primeni na receptomo polje. Receptoma polja su
organizovana po principu centar - periferija, što znači da će u zavisnosti od toga koji deo se
receptivnog polja podražuje doći do ekscitacije, tj. inhibicije pa posledično do povećanja ili
smanjenja frekvencije nervnh impulsa.
6. Nervni centar-morfološke karakteristike, definicije
Podela nervnih centra:
1. N.C. u užem smislu: onaj skup neurona_____________________________
2. N.C. u širem smislu: skup svih neurona u našem mozgu, koji su angažovani da se reši neki zadatak.
Tu postoji već integrativna funkcija.
Pod terminom „nervni centar“ podrazumevaju se grupe neurona u CNS-u koje vrše istu funkciju. Tako senaziva sveukupnost neurona koji su neophodni za sprovođenje, ostvarivanje celokupnog refleksa ilisprovođenje regulacije neke funkcije organizma. Ti nervni centri odgovaraju određenim reakcijama.
Lokalizacija nervnih centara u CNS-u određuje se pomoću nekoliko metoda:
-metoda nadraženja (pojačavanjem, isticanjem njegove aktivnosti)
-metoda razdraženja ili ekstirpacije (uništavanjem osnovne i regulatorne funkcije nernog sistema)
-elektrofiziološke metode (EEG)
U refleksnom aktu organizma kao celine koja je složena, uzimaju se češće ne samo pojedine grupe neuronakoje su smeštene u usko ograničenom delu mozga, nego i mnogi drugi neuroni koji su široko razbacani počitavom CNS-U.
Karakteristike:
1) Jednostavno sprovođenje procesa razdraženja za razliku od nervnog vlakna u kome je nemogućeobostrano sprovođenje. U nervnim centrima talas razdraženja se prenosi samo u jednom pravcu, odreceptivnog neurona preko medijatora, a on se nalazi samo u presimpatičkom delu. Nema ga upostsimpatičkom delu, te je sprovođenje nervnih impulsa kroz sinapse moguće samo od presinaptičkemembrane ka postsinaptičkoj, a ne obrnuto. Sinapsa radi kao zalistak.
2) Usporeno sprovođenje procesa razdraženja – u osnovi sprovođenja procesa razdraženja impulsa ležedva različita neurofiziološka mehanizma: bioelektrični (sprovođenje A.P. po nervnom vlaknu) i hemijski(prenošenje kroz sinapsu pomoću medijatora). Prvi mehanizam se sprovodi jako velikom brzinom, 100-140m/sec, dok je drugi mehanizam hiljadu puta sporiji. To usporenje nastaje jer je potrebno da se odviju sviprocesi u sinapsi od momenta pojave presinaptičkog impulsa do pojave u postsimpatičkoj membrani. Tajinterval se zove sinaptičko zadržavanje, a po sinapsi iznosi 5 ms.
3) Zavisnost refleksnog odgovora od jačine i družine nadraženja – sam refleksni odgovor je intenzivniji štoje intenzitet draži veći i što duže traje a isto tako i što je veći broj receptora zahvaćen u refleksogenom polju.Time se povećava broj refleksa koji pristižu u nervni centar.
4) Sumacija procesa razdraženja
5) Hemijske sinapse služe za fino i tačno doziranje informacija
6) Velika zamorljivost sinaptičkog prenošenja nastaje iz više razloga: iscrpljuju se vezikule, receptori napostsinaptičkoj membrani prestaju da reaguju na neurotransmitere, ponestaje energije
7) Velika osetljivost sinapsi na smanjen novo kiseonika (anoksija)

by J.M. & Ranagol, gen 2010 10
7. Sumacija procesa razdraženja u nervnom centru (vrste, značaj) Nervni sistem ne treba posmatrati sa aspekta akcija-reakcija (kao što je to Pavlov radio) nego
sa aspekta REZULTATA reakcije. Rezultat može biti adekvatna, poluadekvatna ili neadekvatna.
homeo-kineza (pojam koji se nadovezuje na homeostazu) pokazuje da ni jedan parametar našeg
funckionisanja nije konstantan, nego da postoji oscilacija.
alostaza: kada funckija iskoči iz fizioloških granica, van normalnih vrednosti
uslovi za pravilnu integraciju funkcija (tj. uslovi da bi mozak radio kako treba):
o Mozak mora biti adekvatno obavešten (npr. o događajima iz spoljašnje ili unutrašnje
sredine)
o Ova obaveštenja moraju da idu i kvalitativno i kvantitatinvo adekvatno. (da idu
određenim putem. Ovaj princip se zove princip označenog puta. Ako razdraženje ne ide
određenim putem, dobićemo pogrešan utisak, npr osećaj svetlosti u slučaju bola)
o Obrada obaveštenja se vrši u nervnim centrima. Nervni centri su grupa neurona. Ako se
nervni centri međusobno dobro koordiniraju, onda mozak odlično radi.
zona okidanja: ovde imamo najviše sinapsi, najviše AP
oko zone okidanja imamo zonu facilitacije: ova zona se može pretvoriti u zonu okidanja. Ovako,
pomoću ove zone se povećava ili smanjuje broj AP.
Neuron prima, sinhrono ili potpuno nezavisno informacüe sa mnogo drugih neurona
posredstvom brojnih ekscitacijskih i inhibicijskih sinapsi - konvergencija sinapsi. Njihovom
aktivacüom se na membrani postsinaptičkog neurona razvijaju prolazni lokalni sinaptički
potencijali u vidu EPSP-a i {PSP-a. Amplituda svakog od njih je mala, tako da ih repolarizacione
sile lako poništavaju, pa je potrebno da se sumiraju kako bi bili u stanju da značajno promene
ekscitabilnost postsinaptičkog neurona. Sumacija je moguća na bilo kom delu plazma membrane
neurona ali što se ona dešava dalje od aksonskog brežuljka i inicijalnog segmenta dolazi do
većeg gubitka jer su upravo ovi delovi neurona najpodražljiviji jer posedju veliki broj voltažno-
zavisnih kanala, pa se EPSP-i i IPSP-i sumiraju baš tu gde najlakše mogu izazvati A.P.'
(intergrativna ili triger zona).

by J.M. & Ranagol, gen 2010 11
Zbog veoma provodljivog rastvora elektrolita u unutrašnjosti neurona, svaka promena
potencijala u bilo kom delu intracelulame tečnosti some izaziva gotovo identičnu promenu
potencijala u svim ostalim unutrašnjim delovima some neurona što ima veoma važnu ulogu u
sumacji signala.
Efekat' sumacije zavisi i od dužine njenog trajanja, kao i od broja EPSP-a i lPSP-a
sumiranih u određenom periodu, tako da sumacija može biti prostoma i/ili vremenska.
PROSTORNA QSPACUALNAQ SUMACIJA - podrazumeva udruživanje postsinaptičkih
potencijala sinapsi koji okidaju istovremeno a nalaze se na različitim delovima postsinaptičkog
neurona. Ovaj oblik sumacijeje u CNS daleko više zastupljen nego vremenska sumacija. _
VREMENSKA (TEMPORALNA) SUMACIJA - dešava se kada frekventno okidanje
iste sinapse dovodi do nastanka većeg broja EPSP-a i/ili IPSP-a koji se javljaju jedan za drugim i
tako udruženi menjaju potencijal postsinaptičkog neurona.
7. Sumacija procesa razdraženja u nervnom centru
Ako se stimulacija senzitivnog neurona S1 vrši stimulusom čiji je intenzitet na novou praga za motoneuron Aon će se aktivirati i doći će do odgovarajuće refleksne reakcije. Ali, ako je u isto vreme, iz nekog razloga pragmotoneurona B veći od praga neurona A, zajednički neuron B se neće aktivirati ali se u njemu mogu stvoritiekscitatori sinaptički potencijal (EPSP). Ako se stimulus istog intenziteta zatim primeni i na sinaptički S2 ulaz,doći će samo do aktivacuje C neurona, pošto B neće dospeti do novoa paljenja. Ako se međutim, neuron S1i S2 stimulišu istovremeno, motoneuron B može da se aktivira jer se dva pojedinačna EPSP mogu sumirati itako dovesti membranu do nivoa paljenja. Umesto reakciji koja bi odgovarala aktivaciji samo 2 neurona(A,B), izaziva se reakcija koja odgovara aktivaciji tri neurona (A,B,C). Dakle, pri istovremenoj stimulaciji S1 iS2 neurona u zajedničkom motoneuronu B može doći do sumacije, reakcija na stimulus je povećana štoodražava amplifikaciju ulaznog signala. Opisani oblik sumacije je rezultat istovremene aktivacije dvaposebna presinaptička neurona i naziva se prostorna sumacija.
Drugi oblik sumacije je vremenska sumacija. Do nje dolazi kada se isti presinaptički neuron aktivira učesalo,tj. sa većim brojem stimulusa u jedinici vremena. Češći oblik je prostorna sumacija do koje može doći priaktivaciji različitih i čak prostorno veoma udaljenih presimpatičkih neurona. Vremenska sumacija ima manjiznačaj u aktivaciji centralnih neurona, verovatno zato što je ovaj mehanizam modulacije ekscitabilnostisinapse manje pogodan, pošto promena ekscitabilnosti traje relativno kratko.
Subliminarni rub - zajednički neuroni koji isti senzitivni neuron ili interneuron dovode do povećaneekscitabilnosti. U određenim uslovima se aktivira i dovodi do sumacije.

by J.M. & Ranagol, gen 2010 12
8. Postaktivnost. Paralelne i oscilatorne veze (funkcionalni značaj) post pražnjenje: posle dejstva stimulusa mi još uvek doživljavamo... (npr. uticaj emotivno važnog
sna na ostatak dana, kada nam se tokom tog dana vraćaju slike i predstave iz tog sna. Ili kada
sretnemo nekog davno viđenog starog prijatelja, pa posle toga ne možemo skoro da prestanemo
da mislimo na neke zajedničke događaje iz detinstva)
veća labilnost reverberantnog sistema: prividno osoba može da misli na više stvari istovremeno
(Napoleon)
ako se nalazimo u ekscitiranom stanju, stimulusi dovode do većeg odgovora, veće reakcije.
Normalno stanje osobe dovodi do normalnog odgovora. U inhibiranom stanju osoba daje
smanjeni odgovor u odnosu na očekivano.
____________________________________________________________________________
Post aktivnost: neko ponašanje nastavlja da se odvija i onda, kada već nema draži, koja je
provocirala to ponašanje
tokom post aktivnosti jedan nervni impuls ne prođe samo jednom, nego nastavlja da kruži u
nervnom sistemu.
Na ovaj način se ostvaruje dugotrajna funkcionalnost nekog dela mozga, npr.: održavanje
budnosti tokom dana.
Kruženje razdražnje u jednom krugu se naziva reverberacija.
Ova reverberacija može da prestanke kada:
o sinapse se zamore - potroše se neurotransmiteri
o kada dođe neki drugi stimulus, pa zakoči ovo kruženje
Primer za reverberaciju osim budnosti: npr. ako sanjamo nešto što je za nas emocionalno bitno,
pa nam se taj san non stop vraća u svest tokom dana. Kada sretnemo neku bitnu i dragu osobu,
pa posle toga danima razmišljamo i prisećamo o nekim zajedničkim i davnim događajim

by J.M. & Ranagol, gen 2010 13
Postaktivnost je proces koji nastavlja da se odvija i posle delovanja draži, produženje neke refleksne i drugeradnje.
Složene sinaptičke veze koje se stvaraju u CNS-u ne dovode samo do prostornog širenja refleksne ili drugereakcije već se ono može i značajno produžiti. To zavisi od specifičnih neuronskih veza na osnovu kojihjednostruka aktivacija može izazvati mnogostruku aktivaciju istog motoneurona ili drugih centralnih neurona.Specifične interneuronske veze (od kojih zavise takvi efekti) nazvane su lančanim vezama. Postoje dvaoblika takvih veza:
1) Paralelne veze
Interneuroni različitog nivoa povezani su sa različitim motoneuronom. Paralelne veze se ostvaruju pomoćuaksonskih kolaterala specifičnih interneurona. Jedna od aksonskih grana takvih internurona stvara sinapsesa sledećim neuronima i izaziva njegovu eksictaciju. Druga aksonska kolaterala istog interneurona može dastvara ekscitatornu sinapsu sa sledećim interneutonom, tj. interneuronom sledećeg reda. On takođe možebiti ekscitiran i da vrši sinapsu sa istim neuronom – motoneuronom, može da se aktivira neposredno pomoćuinterneurona prvog reda i posredno pomoću interneurona drugog reda. Međutim, interneuton drugog reda,kao i prvi neuron u sastavu ove neuronske mreže, može imati dve aksonske grupe. Druga kolateralainterneurona drugog reda može da stupi u sinaptički kontakt sa interneuronom trećeg reda čiji akson stvara itreću eksictatornu sinapsu sa završnim neuronom u kome bi se stvorila tri akciona potencijala što bi npr.dovelo do trostruke kontrakcije iste motorne jedinice. Da bi došlo do ponovljenje aktivacije završnog neuronamreže, trajanje sinaptičkog zadržavanja mora pri prelasku nervnog impulsa sa interneurona na završnineuron mora da premaši trajanje refraktornog perioda (što i jeste slučaj u neuronskom kolu čak i sa dvaneurona). Sinaptičko zadržavanje je tu dovoljno dugačko te se zato u završnom neuronu mreže posle prvogmože generisati i drugi nervni impuls. Ako je završni neuron motoneuron, njegova višestruka aktivacija ćeizazvati refleksnu reakciju koja traje duže od stimulacije senzitivnog neurona. U slučajevima kada se uneuronskoj mreži umetne i veći broj interneurona aktivnost motoneurona (tj. drugih neurona) može bitiznačajno produžena.
2) Oscilatorne veze (reverberantni krugovi)
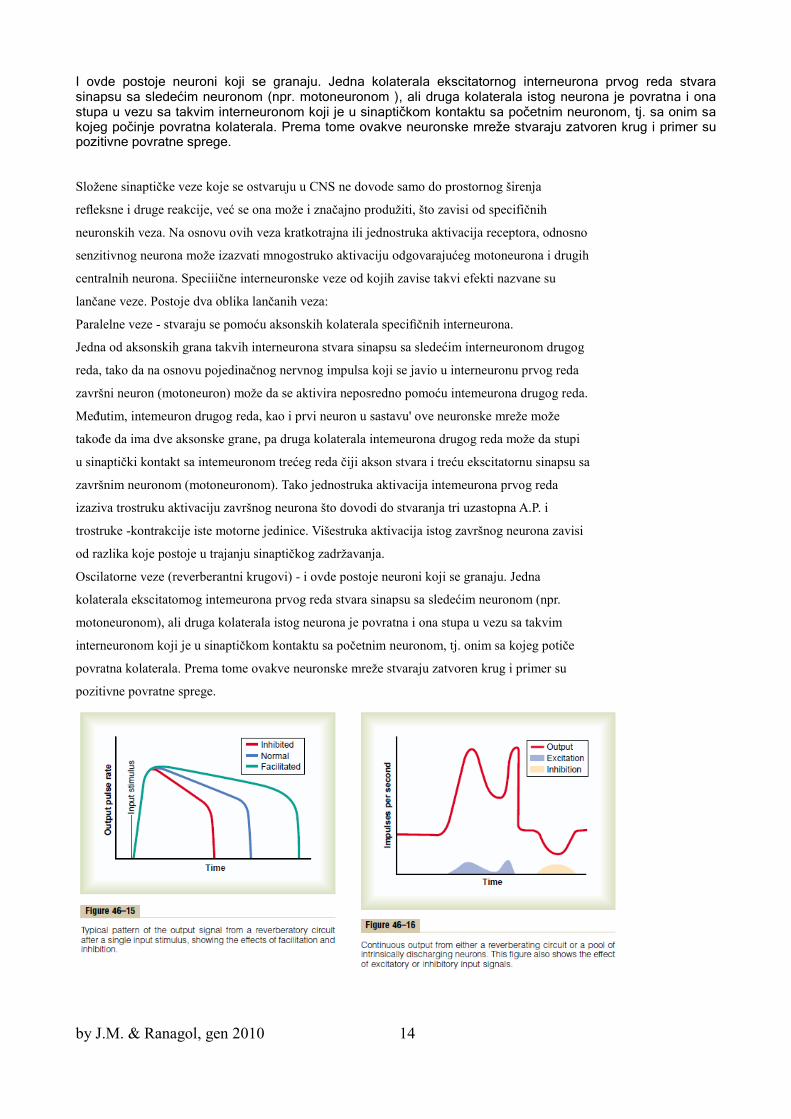
by J.M. & Ranagol, gen 2010 14
I ovde postoje neuroni koji se granaju. Jedna kolaterala ekscitatornog interneurona prvog reda stvarasinapsu sa sledećim neuronom (npr. motoneuronom ), ali druga kolaterala istog neurona je povratna i onastupa u vezu sa takvim interneuronom koji je u sinaptičkom kontaktu sa početnim neuronom, tj. sa onim sakojeg počinje povratna kolaterala. Prema tome ovakve neuronske mreže stvaraju zatvoren krug i primer supozitivne povratne sprege.
Složene sinaptičke veze koje se ostvaruju u CNS ne dovode samo do prostornog širenja
refleksne i druge reakcije, već se ona može i značajno produžiti, što zavisi od specifičnih
neuronskih veza. Na osnovu ovih veza kratkotrajna ili jednostruka aktivacija receptora, odnosno
senzitivnog neurona može izazvati mnogostruko aktivaciju odgovarajućeg motoneurona i drugih
centralnih neurona. Speciiične interneuronske veze od kojih zavise takvi efekti nazvane su
lančane veze. Postoje dva oblika lančanih veza:
Paralelne veze - stvaraju se pomoću aksonskih kolaterala specifičnih interneurona.
Jedna od aksonskih grana takvih interneurona stvara sinapsu sa sledećim interneuronom drugog
reda, tako da na osnovu pojedinačnog nervnog impulsa koji se javio u interneuronu prvog reda
završni neuron (motoneuron) može da se aktivira neposredno pomoću intemeurona drugog reda.
Međutim, intemeuron drugog reda, kao i prvi neuron u sastavu' ove neuronske mreže može
takođe da ima dve aksonske grane, pa druga kolaterala intemeurona drugog reda može da stupi
u sinaptički kontakt sa intemeuronom trećeg reda čiji akson stvara i treću ekscitatornu sinapsu sa
završnim neuronom (motoneuronom). Tako jednostruka aktivacija intemeurona prvog reda
izaziva trostruku aktivaciju završnog neurona što dovodi do stvaranja tri uzastopna A.P. i
trostruke -kontrakcije iste motorne jedinice. Višestruka aktivacija istog završnog neurona zavisi
od razlika koje postoje u trajanju sinaptičkog zadržavanja.
Oscilatorne veze (reverberantni krugovi) - i ovde postoje neuroni koji se granaju. Jedna
kolaterala ekscitatomog intemeurona prvog reda stvara sinapsu sa sledećim neuronom (npr.
motoneuronom), ali druga kolaterala istog neurona je povratna i ona stupa u vezu sa takvim
interneuronom koji je u sinaptičkom kontaktu sa početnim neuronom, tj. onim sa kojeg potiče
povratna kolaterala. Prema tome ovakve neuronske mreže stvaraju zatvoren krug i primer su
pozitivne povratne sprege.

by J.M. & Ranagol, gen 2010 15
9. Zamorljivost nervnih centara Postoji određen stepen aktivnosti većine neurona unutar neurološkog ansambla, a to se zove
TONUS NERVNIH CENTARA. Tonus nervnih centara je osobina neurona, da su ovi neuroni
spontano aktivni i onda kada nema nekih evidentnih dejstava.
Ovaj tonus može biti više ili manje izražen u vidu spontano generisanih aktivnih potencijala u
jednoj jedinic vremena.
Tonus nervnih centara je važan zbog BRZINE REAGOVANJA.
Tonus nervnih centara obezbeđuje našu adekvatnu i brzu reakciju (npr održavanje ravnoteže
prilikom spoticanja)
Velika ekscitacija dovodi do velike promene (moduliranja) tonusa nervnog centra
Inhibicija dovodi do promene tonusa N.C. u negativnom smislu. Ovaj tonus se odnosi ne samo na
mišiće, nego na našu celu spremnost na aktivnost.
Zamaranje nervnih centara karakteriše ceo nervni sistem (ovo se odnosi i na pojedinačnu
skupinu neurona, ali i na ceo nervni sistem). CNS se brzo zamara→ → i treba da se brzo zamara
jer tako štiti naše organe-mišiće od propadanja. Zamaranje je reverzibilno (može da se odmori),
ali propadanje organa nije reverzibilno.
Nisu svi delovi mozga jednako aktivni. Angažovane grupe neurona se prve zamore, dok su ostale
grupe neurona pošteđene.
Exhaustirani generalni nervni sistem: jako iscrpljene osobe, koje su prešle preko nekog teškog
perioda u životu. Ovi treba da se puno odmaraju, spavaju. Na taj način se oporavlja nervni
sistem.
Ako smo zamorili jednu grupu nervnih centara npr. učeći fiziologiju, onda tu grupu nervnih
centara možemo da odmorimo tako, što ćemo učiti statistiku (koja aktivira neke sasvim druge
nervne centre)
Npr. ako odemo u muzej, prve slike će nam biti jako lepe i interesantne, ali na kraju ćemo se
jako zamoriti, i sve slike će nam biti iste. Ovde opet radi samo jedan ograničeni broj neurona,
koji se zamara.
Zamor nervnih centara ogleda se u zamoru hemijskih sinapsi, kao zaštita od prevelike
stimulacije. Manifestuje se prvo usporenjem, a potom i prekidom sinaptičke transmisije, a obično
se javlja prilikom frekventnog i/ili dugotrajnog okidanja sinapse, uglavnom kada se istroše
raspoložive količine neurotransmitera a reutilizacija nije dovoljna da to kompenzuje. On može
nastati i usled progresivne inaktivacije receptora na postsinaptičkoj membrani. U nekim

by J.M. & Ranagol, gen 2010 16
slučajevimaje zamor sinapsi posledica nagomilavanja kalcijumovih jona u sinaptičkm završetku,
jer oni aktiviraju kalcijum-zavisne kalijumove kanale presinaptičke membrane i tako
omogućavaju difuziju kalijumovih jona u međućelijsku tečnost, usled čega dolazi do
hiperpolarizacije presinaptičke membrane tako da se zaustavlja dalja transmisija.
10. Proces divergencije u CNS (iradijacija)
Divergencija je pojam širenja razdraženja, nastaje kod dugog i jakog stimulusa.
Razdraženje se može širiti toliko, da može nastati generalizacija (veliki epileptični napad)
kod divergencije (iradijacije) izlaz je veći od ulaza
Pod divergencijom se podrazumeva prenošenje nervnih impulsa sajednog presinaptičkog
na veći broj postsinaptičkih neurona. Divergentne veze karakterišu sve delove kičmene moždine,
a takođe i sve delove mozga. U kičmenoj moždini posoje divergentne veze koje se stvaraju na taj
način što se aksoni senzitivnih neurona granaju i stupaju u sinaptičku vezu sa više intemeurona
ili motoneurona, zahvaljujući čemu dolazi do širenja refleksne reakcije. Prema tome u CNS
divergentne veze u stvari obezbeđuju proces amplifikacije pri kom izlaz (npr. refleksna reakcija)
postaje veći od ulaza ( aktivacije senzornih receptora, odnosno senzitivnih neurona).
Divergentne veze se stvaraju i u samim moždanim strukturama, takve veze su izrazite
između neurona malog mozga, a takođe između međumozga (talamusa) i kore prednjeg mozga.
11. Konvergencija u nervnom sistemu (funkcionalni značaj) Konvergencija je suprotan pojam od divergencije.

by J.M. & Ranagol, gen 2010 17
konvergencija znači okupljanje ili sabiranje
ulaz je veći od izlaza.
Pojedine postsinaptičke neurone u CNS može da aktivira više presinaptičkih neurona koji
na njih konvergiraju. Na mnoge motoneurone kičmene moždine konvergira veliki broj
senzitivnih neurona, a naročito intemeurona. Sa interneuronima sive mase kičmene moždine
stvaraju sinapsu mnogi drugi neuroni čija su tela locirana ivan kičmene moždine, u drugim
delovima mozga.
Konvergencija mnogih senzitivnih neurona na iste motoneurone u kičmenoj moždini
posledica je kvantitativnih odnosa između nervnih ćelija jedne i druge kategorije. Informacije,
odnosno nervni impulsi koji potiču iz čula dospevaju do kore velikog mozga na taj način što se
prenesu prvo na tzv. kortikalna relejna jedra talamusa. Između talamusa i kore velikog mozga
stvaraju se i divergentne i konvergentne veze. Tako se sjedne strane informacije sa pojedinačnih
talamičkih neurona prenose na više kortikalnih neurona. S druge strane, na iste neurone u sastavu
senzomih zona korteksa telencefalona konvergira više talamičnih neurona. Od različitih
informacija kortikalni neuroni odaberu one koje u datom momentu imaju funkcionalni smisao.
Osnova konvergencije je sumacija postsinaptičkih potencijala na nervnu ćeliju.
Značaj – odgovarajuća reakcija efektora na različite informacije iz većeg broja receptora.
Postoji konvergencija iz jednog i različitih izvora:
l) Multisenzorna - ostvaruje se iz različitih senzomih izvora (iz oka, uva, miris...)
2) Multibiološka - vezana su za hranu, vodu, polne funkcije, odbranu
3) Senzobiološka - istovremeno se na neuronu okupe informacije i iz senzornih i
bioloških sistema, bez nje nema izgradnje uslovnih refleksa-ovo je najvažnije. Ovo je uslov za izgradnju uslovnih
refleksa, uslov za adaptaciju.
12. Recipročna inervacija (spojena inervacija)
ovo je sninonim za istovremenu negativnu indukcija
kod spojene inervacije se dešava sledeće: jedan ekscitirajući ulaz daje 2 različita izlaz
jedan razdraženi neuron direktno aktivira neke efektore, a inhibira neke druge efektore.
ovo je bitno kod fleksije ekstremiteta
fleksija je refleksno savijanje ekstremiteta
Šerington je ` registrovao kontrakciju dve kategorije mišića u toku refleksne fleksije
ekstremiteta i utvrdio da se ekstenzori relaksiraju proporcionalno kontrakciji fleksora. Ako se
kontrahuje manji broj motornih jedinica fleksora, relaksira se i adekvatni broj ekstenzomih
jedinica. Na taj način se dakle može zaključiti da pri refleksu fleksüe mišići ekstenzori imaju

by J.M. & Ranagol, gen 2010 18
funkciju antagonista u odnosu na fleksorne mišiće koji u ovom refleksu učestvuju kao sinergisti.
Proporcionalno, ali suprotno stanje dve kategorije mišića obezbeđuje normalno, koordinisano i
glatko odvijanje ovog refleksa kao i svih drugih refleksnih reakcija odnosno pokreta. Stimulacija
receptora ima suprotan efekat na dve kategorije motoneurona: aktivacija senzitivne komponente
refleksnog luka izaziva ekscitacüu motoneurona sinergista, a istovremeno dolazi do inhibicije
motoneurona antagonista. Antagonističko dejstvo senzorne stimulacije na dve kategorije efektora
Šerington je nazvao recipročna inervacija. Recipročna inervacfia je pored divergencije, sumacije
i konvergencija veoma značajan mehanizam na kome se zasniva. koordinacija refleksa.
Suprotne efekte postignute stimulacijom receptora kože na dve grupe mišića Šerington je nazvao recipročnainervacija. Ova pojava uslovljena je divergencijom senzitivnih neurona. Jedna grana aksona senzitivnogneurona uspostavlja sinapsu direktno s motoneuronima mišića fleksora i u njima izaziva akcione potencijale.Druga grana vrši sinapsu s umetnutim neuronima koji inhibiraju motoneurone ekstenzora.
U suštini, ektenzori relaksiraju proporcionalno kontrakciji fleksora. Pri refleksu fleksije, mišići ekstenzori imajufunkciju antagonista u odnosu na fleksorne mišiće koji u ovom refleksu učestvuju kao sinergisti.Proporcionalno, ali suprotno stanje dve kategorije mišića obezbeđuje normalno koordinisano i glatkoodvijanje ovog refleksa. Aktivacija senzitivne komponente refleksnog luka izaziva ekscitaciju motoneuronasinergista i istovremeno inhibiciju antagonista.
Recipročna inervacija je pored divergencije, sumacije i konvergencije veoma značajan mehanizam na komese zasniva koordinacira refleksa.
13. Princip povratne sprege(veze) princip povratne aferentacije nam dozvoljava da vidimo, koliko smo efikasni u tome, što
radimo. (feedback)
postoje dve vrste povratne aferentacije:
o PRAVA POVRATNA AFERENTACIJA se zove još i multiparametarska→ feedback
na svim čulima, da li je izvršen zadatak
o LAŽNA POVRATNA AFERENTACIJA:
pranje zuba kao dijeta
ugrađen balon u želucu, što stvara osećaj sitosti
fantomski ud (gde se može pomoći hirurškim odstranjivanjem zaostalog
nervnog centra, koji ustvari šalje te lažne signale postojanja uda, tj. signale
fantomskog bola)
Pod pojmom povratne sprege podrazumeva se takva veza pojedinih elemenata u sistemu
gde izlaz utiče na aktivnost ulaza. Može biti pozitivna ili negativna. Kod pozitivnih izlaz
pojačava (ampliñcira) ulaz, a dok kod negativnih dozati do inhibicije-implifikacije i imaju
presudan značaj za znaje o uspešnosti. U biološkim sistemima česte su negativne povratne
sprege. Povratna sprega je tehničko matematički pojam,~ali koristan i za razumevanje mnogih
bioloških procesa, među njima i procesa u CNS.
0 primer pozitivne povratne sprege je neuronska mreža koja se naziva reverberantni krug ili

by J.M. & Ranagol, gen 2010 19
oscilatorna veza.
Pod pojmom povratne sprege podrazumeva se veza pojedinih elemenata u sistemu gde izlaz utiče naaktivnost ulaza. Može biti pozitivna i negativna. Kod pozitivne, izlaz pojačava (amplificira) ulaz, dok kodnegativnih dolazi do inhibicije (implifikacije) ulaza, i one imaju presudi značaj za znanje o uspešnosti i čestesu u biološkim sistemima.
Povratna sprega je tehničko-matematički pojam, ali koristan za razumevanje mnogih procesa, među njima iprocesa CNS-a. Primer pozitivne povratne sprege je neuronska mreža koja se naziva reverberantni krug ilioscilatorna veza.
Oscilatorna veza se stvara između neurona kore velikog mozga i talamusa, a takođe je i u experimentima naizolovanim ostacima kore prednjeg mozga dokazan fenomen reverberacije.
Razmatrajući funkcionalan značaj reverberantnih interneuronskih mreža u CNS-u, neki naučnici supretpostavili da bi se fenomen memorije mogao objasniti neprekidnim kruženjem nervnih impulsa unutar istepopulacije neurona. Na taj način bi se jednom primljena informacija mogla trajno zadržati u CNS-u. Međutim,ova dinamička teorija je danas pobijena, bar kada se radi o konsolidovanom pamćenju. Za razliku odneučvršćenog, skorašnjeg (nekonsolidovanog) pamćenja koje se relativno lako gubi, konsolidovanopamćenje odoleva različitim, često drastičnim tretmanima organizma, kao što su elektrošokovi velike jačine,duboke hipotermije i ekstremno duboke amnezije, a takvi postupci onemogućavaju stvaranje i prostiranjenervnih impulsa u svim, pa i u centralnim neuronima. U ovakvim situacijama, ponavljana aktivacija neurona uoscilatornom kolu bi se prekinula, te bi se tako i gubilo pamćenje ako bi ono zavisilo od reverberacije tj.stalnog kruženja impulsa u okviru iste neuronse populacije.
14. Princip dominante ( hijerarhija motivacija ) Nas vode brojne motivacije kroz život
Ona motivacija, koja je u datom momentu najznačajnija, ona će rukovoditi, koji će centri u
nervnom sistemu biti najrazdražljiviji.
Najrazdražljiviji nervni centri će se i najlakše aktivirati
Ako dominira socijalna motivacija nad biološkim motivima (motiv postignuća, imitiranje nekoga...)
to ne valja. Primer: anoreksija zbog socijalnih činioca.
U CNS u svakom momentu našeg svesnog života postoje odgovarajući delovi mozga koji
su aktivniji u odnosu na druge delove. Te delove mozga aktivira naša dominírajuća potreba,
odnosno naša dominírajuća motivacija, sve naše funkcije podređene su principu dominante.
Odlike dominante:
1. Povećena razdražljivost nervnih centara
2. Stabilnost procesa razdraženja u vremenu (prostomih)
3. Sposobnost za funkciju drugih razdraženja
4. lnercija dominantnog žarišta razdraženja
Osnovnu odliku dominante predstavlja sposobnost čvrstog održavanja procesa razdraženja u
vremenu. Retko je odsutna bilo koja dominanta. Stanje bez dominante je stanje slabog
razdraženja. Ono se javlja kod sportista u stanju potpune opuštenosti. Dominanta se javljakao
fiziološka osnova pažnje što je izraženija, sve intezivnije inhibira druge tekuće reflekse.

by J.M. & Ranagol, gen 2010 20
Hijerarhija motiva:
l .Biološki
život
ishrana
produženje vrste - polne motivacije
2.Socijalni
motiv postignuća
plastičnost motivacije - zamenjivanjejednog motiva drugim
15. Princip opšteg krajnjeg puta Princip krajnjeg puta se naziva i levak Šeringtonav
Ovde se radi o integrativnoj funkciji mozga, gde mozak radi tako, da u skladu sa dominirajućom
motivacijom, stvara dominatne oblike razdraženja, i tako zacrtava određeni cilj. To je taj levak.
Uprkos činjenici da mi doživljavamo različite modalitete osećaja, nervna vlakna prenose
samo impulse. Svako nervno vlakno završava u specifičnoj tački centralnog nervnog sistema, pa
je stoga tip osećaja koji se doživi pri stimulaciji nervnog vlakna determinisan mestom u nervonm
sistemu do kojeg to vlakno vodi. Na primer, ako se stimuliše vlakno za bol, čovek oseća bol bez
obzira na to koji tip draži deluje na vlakno. Ova specifičnost nervnih vlakana za prenos jednog
modaliteta osećaja naziva se principom označenog puta.
Princip označenog puta se sastoji iz kombinacije signala koji putuje i draži određenog intenziteta, kvaliteta imodaliteta. Odgovor može biti okidanje ili faciliranje.
16. Plastičnost nervnih centara plastičnost znači promenljivost, znači prilagodljivost nervnog sistema
Plastičnost može biti
1. Fiziološki fenomen adaptacije na uslove života (učenje)
2. Regeneracija nakon lezije CNS - izrastanje aksona i dendrita (ne stvaranje novih neurona! To se
dešava isključivo kod receptora mirisa i ukusa!)
delovi hromozoma → → → kod bolesne osobe se menjaju (fragilni su) prave kapice (telomere)
oni čuvaju hromozom (nešto kao čarapa na nozi :) )→ zbog stresa telomere propadaju→pa i
hromozom propada, tj. psihološki strada
psihoterapija dovodi do repariranja genetskog materijala→ pojačavaju se ove telomere→što se
može postignuti psihoterapijom. Dakle, psihoterapijom mi menjamo i strukturu neurona.
epigenetski delovi (prazna mesta kod hromozoma) su osnova plastičnosti na genetskom nivou

by J.M. & Ranagol, gen 2010 21
dakle, psihoterapijskim radom možemo da štitimo hromozom. Suština psihoterapije: epigenetski
uticaj.
Plastičnost nervnih centara prevashodno se odnosi na plastičnost hemijskih sinapsi. Plastičnost iliadaptibilnost mozga, tj vikarisanje njegovih funkcija je sposobnost projekcionih zora kore velikog mozga dase adaptira na promene koje nastaju u toku života. Prilagodljivost je kompenzacija funkcija. Nervni sistem jeprilagodljiv za mnoge informacije. Takođe je dinamičan i prilagodljiv – ako dođe do razaranja nekih delovaCNS-a drugi delovi preuzimaju njihovu ulogu.
Biološki mehanizam pamćenja i zaboravljanja se zasniva na plastičnosti. Odvijaju se tokom čitavog života,naročito za vreme komunikacije i fizičke aktivnosti. Kroz učenje dolazi do usložnjavanja strukture i stvarajuse novi engrami pamćenja. Mehanizam stvaranja do danas nije poznat.
Obuhvata i neurone i gliju, i centralni i periferni nervni sistem.
Hemijske sinapse nisu statički spojevi koji prenose neizmenjene akcione potencijale sa presinaptičkog napostsinaptički neuron, nego pritom i moduliraju oblik bioimpulsa u smislu olakšavanja (facilitacije),postetanične potencijacije, ili pojave zamora (depresije) sinapsi. Svi oblici modulacije su naročito izraženi utoku visokofrekventne (tetanične) stimulacije, kada amplituda akcionih potencijala postepeno raste -facilitacija transmisije, da bi potom počela da opada - depresija transmisije (više opada što je veća frekvencastimulacije). Plasticitet sinaptičke transmisije impulsa uglavnom zavisi od intenziteta egzocitozeneurotransmitera, a on je upravo proporcionalan količini difundovanih kalcijumovih jona u sinaptičkizavršetak. U uslovima visokofrekventne stimulacije, inače spore kalcijumove pumpe ne stignu da ispumpajusav višak kalcijumovih jona. Zato se oni nagomilavaju u citoplazmi sinaptičkog završetka i dovode dopojačane i produžene egzocitoze neurotransmitera, koja može trajati od nekoliko minuta do sat vremena. Izovog razloga, pojedinačni stimulus dat po prestanku frekventne stimulacije produkuje amplitude akcionihpotencijala veće od svih prethodnih (posttetanična potencijacija).
Plastičnost ili adaptibilnost mozga odnosno vikarisanje njegovih funkcija je sposobnost
projekcionih zona kore velikog mozga da se adaptira na promene koje nastaju tokom života.
Naime većina kortikalnih reprezentacija receptora u SKVM i predstavništa pokreta u MKVM
nije isključivo nasleđena i zauvek datajerje moguće da se tokom života neki receptori i/ili voljni
mišići koriste intezivnije pa se povećava površina njihovog predstavništva u senzomoj, tj.
motomoj kori velikog mozga. Ovo potvrđuje da učenje dovodi do strukturnih promena u kori,
koje se ogledaju u promenama senomih i/ili motornih mapa. Tako npr. osoba koja je ostala bez
ruku može da nauči da piše prstima stopala ili držeći olovku u ustima. Uklonjeno predstavništvo
bilo kog dela tela se brzo ofonni na novom mestu u kori, a kod većih oštećenja, kada je npr.
zahvaćeno predstavništvo dela tela u jednoj hemisferi, redovno dolazi do proširivanja
predstavništva za taj deo tela u suprotnoj hemisferi. Veoma važna sposobnost CNS za održavanje
njegove integracijske uloge je vikarisanje njegovih funkcija, zahvaljujući čemu, oštećene ili
izgubljene funkcije bivaju kompenzirane drugim funkcijama.
17. Kompenzacija funkcija u CNS-u
CNS u osnovi ima dve funkcije:
1. one povezuje receptore sa udaljenim efektorima

by J.M. & Ranagol, gen 2010 22
2. u CNS-u se odvijaju složeni integracioni procesi od kojih zavisi kako koordinisano i svrsishodnokretanje tako i svi drugo oblici ponašanja što uključuje i svesno ponašanje čoveka.
CNS je dinamičan i prilagodljiv. Ako dođe do razaranja nekih delova CNS-a ostali delovi preuzimaju njihovufunkciju. Nadoknađenje funkcija posle oštećenja mozga se odvija na dva načina :
1. stvaranjem bočnih ogranaka ostalih neurona i nadomešćuje se oštećenje
2. aktiviranjem mirujućih neurona (može se javiti greška pa će se naći zalutali nervi u oporavku).
18. Inhibicija u CNS-u (vrste i značaj)
Pojavu inhibicije opisao je Šerington. Aktivacija i inhibicija su suprotni procesi, međusobno neodvojivi.
Inhibicija je aktivan nervni proces, na makronivou se manifestuje u vidu smanjenja ili prestanka nekedelatnosti, a na mikronivou predstavlja izraz smanjene razdražljivosti neurona koja se manifestujesmanjenjem broja generalisanih ak.potencijala ili njegovim potpunim prestankom.
Prema Šeringtonu nervni impulsi koji stižu sa perifernih receptora u CNS mogu imati dvojako dejstvo:aktivirajuće (ekscitatorno) i kočeće (inhibitorno).
Proces koji dovodi do smanjenja ili do prestanka aktivnosti centralnih neurona zove se centralna inhibicija.
Tipovi inhibicije:
1.DIREKTNA (postsinaptička) se javlja u postsinaptičkoj membrani neurona, a pod dejstvom specijalnoginhibitornog medijatora. U CNS-u postoje specijalni inhibitorni neuroni, tj. jedna vrsta interneurona u čijimzavršecima se izlučuje inhibitorni medijator GABA, koji uslovljava hiperpolarizaciju membrane motoneurona,tj. stvaranje IPSP, čime se membrana udaljuje od nivoa paljenja I prestaju da se generišu ak.potencijali(mišić se ne kontrahuje). Nervni impulsi kada pristignu u inhibitorni neuron izazivaju u njima procesrazdraženja, u njemu se javlja ak.potencijal koji se rasprostire duž njegovog aksona, ali taj akson pri dolaskuak.potencijala na presinaptički završetak izlučuje ne razdražujući, nego inhibitorni medijator. Javlja se kratkahiperpolarizacija postsinaptičke membrane sledećeg beurona i dobijamo IPSP.
2.DIREKTNA (presinaptička) se javlja pred sinaptičkim kontaktom neposredno u presinaptičkoj oblasti.Aksonalni završeci inhibitornog neurona formiraju akso-aksonsku sinapsu na aksonskom završetku drugenervne ćelije i tako blokira proces razdraženja u njoj, jer kad se aktivira on luči inhibitorni transmiter. Takodolazi do hiperpolarizacije aksonskog završetka neurona koji inače stvara ekscitatornu sianpsu sa sledećomnervnom ćelijom. Akcioni potencijal se smanjuje kasnije a poslediaca smanjenja amplitude akcionogpotencijala je smanjenje ulaska Na I K.
3. Inhibicija pomoću negativen povratne sprege (povratna) - Ovaj tip inhibicije utvrđen je kod alfamotoneurona u kičmenoj moždini, do čije inhibicije dolazi na račun aktivacije specifičnih interneurona koji sezovu Renšou neuroni.
Pojedini motoneuroni u sivoj masi kičmene moždine odlikuju se time što se njihovi aksoni granaju. Neke odaksonskih kolaterala su povratne i stvaraju sinapsu sa Renšou neuronima . Sa druge strane, taj isti neuronstupa u sinaptički kontakt sa telom onog istog neurona čija aksonska kolaterala stvara ekscitatornusinaptičku vezu sa Renšou neuronom. Ovde se radi o povratnoj sprezi koja se stvara između motoneurona Injegove kolaterale tj. inhibitornih neurona.
Inhibicijske sinapse ili sinapse tipa II imaju sve delove uobičajene za hemijsku sinapsu (asimetričan spljoštenbuton sa pljosnatim ili ovalnim vezikulama, manja aktivna zona i uža sinaptička pukotina (oko 20nanometara) nego kod ekscitatijskih sinapsi. Inhibicija neke ekscitacijske sinapse se postiže sehiperpolarizacijom njene presinaptičke ili postsinaptičke membrane, a može se ostvariti i jakom aktivacijom

by J.M. & Ranagol, gen 2010 23
specifičnih enzima koji razlažu njen neurotransmiter. U zavisnosti od toga ona može biti presinaptička ilipostsinaptička.
Presinaptička inhibicija nastaje pražnjenjem inhibicijske sinapse koja se nalazi ispred presinaptičkogzavršetka. Pretežno se odvija preko akson-aksonskih sinapsi, koje posredstvom neurotransmitera GABA iliglicina, izazivaju manje ili jače izraženu lokalnu hiperpolarizaciju. Usled toga potpuno se zaustavlja ili samoumanjuje frekvenca akcionih potencijala na putu ka sinaptičkom završetku. Osim presinaptičke inhibicijepostoji i presinaptička facilitacija. Uglavnom se ostvaruje posredsvom akson-aksonskih sinapsi, ali u ovomslučaju ona dovodi do manje ili jače izražene presinaptičke membrane.
Postsinaptička inhibicija u zavisnosti od stepena postignute hiperpolarizacije, opada ili čak privremenopotpuno gubi ekscitabiilnost postsinaptičkog neurona. Poseban oblik postsinaptičke inhibicije je Renshawinhibicija koja nastaje u alfa motornim neuronima prednjih rogova sive mase kičmene moždine. Ona služi zakontrolu poprečnoprugastih mišića, tj. preciznosti i snage voljnih pokreta. Može se reći da ova vrsta inhibicije,zapravo, filtrira impulse do optimalne frekvence za mišićnu kontrakciju. Ona spada u recipročnu inhibiciju jeralfa neuron sam sebe hiperpolariše i na taj način smanjuje broj porečnoprugastih mišića koji učestvuju upokretu. Time se dodatno povećava precišnost pokreta.
Značaj inhibicije leži u tome što omogućava da se postigne velika preciznost voljnih pokreta.
19. Metode ispitivanja funkcija CNS-a (razaranje, presecanja, ekstirpacija) Postoji 3 osnovne vrste izučavanja CNS:
1. Metode razaranja, presecanja, ekstirpacije
2. Metode stimulacije
3. Neurofiziološke metode
svaka metoda ima svoje dobre i loše strane, tek kombinacija svih metoda daje punu sliku
Kod metode razaranja, presecanja, ekstirpacije se napravi oštećenje, pa se prati koje se
funkcije oštećuju. Dakle, gledaju se prouzrokovane lezije (kako na ljudima ne možemo da
uzrokujemo povrede...) i simptomi kod razaranja.
Mogu biti:
o Grube metode: vađenje hemisfere malog mozga, presecanje kičmene moždine
o Fine metode: X zraci, protonski zraci
Razaranje se može koristiti i kod terapije
Pravi se oštećenje nervnog sistema I prati se koje su funkcije nestale. Gube se i velike i male funkcije.Koriste se lezije.
Prva ispitivanja strukture i funkcija CNS-a vršena su neuroanatomskim i neurofiziološkim metodamarazaranja ili presecanja pojedinačnog nerva, nervnog vlakna, odnosno tkiva, kao i ekstirpacijom tj. potpunimodstranjivanjem određenog strukturnog dela oglednog mehanizma (organa, tkiva, tumora) i potomposmatranjem i zapisivnjem nastalih promena. Vršeni su na životinjama I ljudima.
Razaranje– razaranje delova CNS-a različitim postupcima, prati se šta nedostaje.

by J.M. & Ranagol, gen 2010 24
Presecanje – ogleda se u presecanju različitih delova CNS-a, najstarija metoda, posmatranje funkcija kojeprestaju sa izvršenim ogledom.
Ekstirpacija (udaljivanje) – razdvajanje delova CNS-a.
Primer – prefrontalna lobotomija (hirurško presecanje veze između frontalnog područja i ostalih delova mozgakao tretman mentalnih bolesti, sideaffects.)
20. Metode ispitivanja funkcija CNS-a (stimulacije) stimulišemo određenu nervnu strukturu i pratimo kakve efekte dobijamo
električna stimulacija je bolja od hemijske stimulacije, jer:
o blaga struja odgovarajuče jačine i napona imitira akcione potencijale
o može se dozirati frekvencija, jačina struje, napon
primer za korišćenje elektro-stimulacije: lečenje depresije→ elektrošok resetuje mozak
(nestane depresija)→ na neko vreme ponestanu određeni neurotransmiteri→ pa posle ponovo
treba elektrošok...
hemijska stimulacija može biti problematična, zbog:
o može imati različita dejstva na različite ljude
o sa preteranom dozom se može naneti šteta
o sa premalom dozom ne znamo šta se dobija
Ako deluje odgovarajući stimulus na receptor, u nervnim vlaknima se stvaraju nervni impulsi. Ovi nervni impulsi se
prenose na mišiće, i stvaraju kontrakcije. Ove kontrakcije se mogu registrovati pomoći adekvatnih metoda.
Stimulacije se mogu vršiti na više načina:
1. Mehanički (pritiskom)
2. Elektrostimulacija
3. Magnetna stimulacija (najsavršenija metoda)
4. Stimulacija hemijskim materijalima
20. Metode ispitivanja funkcija CNS-a (stimulacije)
Stimulišu se određene nervne strukture i prate se promene funkcija. Uglavnom se koristi elektrostimulacija,a postoji i stimulacija hemijskim materijama. Pri dejstvu stimulusa se u nevnim vlaknima stvaraju nervniimpulsi. Pored električne i hemijske, postoji i mehanička i magnetna stimulacija.
Velika grupa metoda, suprotna metodama presecanja, udaljavanja i razaranja.
Stimulisanje pokreće na aktivnost.
Električna :
Cešća - razlozi : (prednost električne struje )
1. lako se dovodi i dozira
2. imitira AP

by J.M. & Ranagol, gen 2010 25
3. u dovoljnoj dozi ne oštećuje tkivo i izaziva povratno dejstvo
Lokalizacija elektroda :
na povrsini, unutar strukture
Primena : mapiranje korteksa
Hemijska :
ređa -razlozi:
1. ne može se lako neutralisati dejstvo unesene materije
2. predoziranje
3. metabolisanje hemijske materije : drugačiji efekat
Mehanička – najjednostavnija, a magnetna je najsavršenija. Magnetna je zamenila električnu jer jejednostavnija i bezbolna.
21. Metode ispitivanja funkcija CNS-a (elektrofiziološke ili neurofiziološke
metode) zasnivaju se na tome, da se registruju bioelektrični procesi, koji se odvijaju unutar nervnog
sistema
ovi bioelektrični ili neurofiziološki procesi se dele na više grupa:
1. Elektroencefalogram (EEG): suma bioelektrične aktivnosti mozga (EEG je sumarna
bioelektrična aktivnost, registruje se više hiljada neuronske aktivnosti). Može se koristiti
duže vreme, nije škodljivo. Dakle, stave se elektrode, i snimaju se bioelektrični potencijali.
Analiza se vrši kompjuterski, sa specijalnim programom.
2. Izazvani (evocirani) potencijali
3. Registracija aktivnosti samih neurona. Vrši se hirurškim putem, kod životinja, sa
mikroelektrodom. Registruju se akcioni potencijali neurona.
4. Intracelularna metoda: uvede se elektroda u sam neuron. Daje podatke o elektrofiziologiji
ćelije.
Savremena ispitivanja funkcija CNS obuhvataju različite elektrofiziološke metode koje se
okvirno mogu svrstati u :
Elektroencefalografija -je posebna neurofiziološka metoda koja registruje moždanu električnu
aktivnost preko elektroda smeštenih na poglavini (što je najčešći vid korišćenja) ili subduralno
odnosno unutar moždanog tkiva. Rezultujući dijagram je poznat kao elektroencefalogram (EEG)

by J.M. & Ranagol, gen 2010 26
koji je, ranije, preko posebnih pera osetljivih na izmenu napona, iscrtavan na papiru, a u
sadašnjici češće zastupljen u vidu digitalnog crteža korišćenjem računara. EEG beleži moždane
talase koji su prema frekvenciji podeljeni na delta talase (aktivnost do 4 herca), teta talase (od 4
do 8 herca), alfa talase (od 8-12 herca) i beta talase (preko 12 herca). Pojedina normalna
(budnost, spavanje, dremanje) i patološka stanja (epilepsija, encefalopatija, moždani udar ili
druge strukturne lezije mozga) imaju svoj specifičan EEG zapis, zahvaljujući kojem je EEG
uvršten kao neophodna dijagnostička metoda u ispitivanja epilepsije. Nažalost, EEG ima svoja
značajna ograničenja. Ona se odnose na prostorna (EEG beleži električnu aktivnost sa svega l/3
moždane kore) i vremenska (EEG se u standamim uslovima beleži u vremenskom trajanju od
dvadeset .minuta)ograničenja. Ova ograničenja se delom mogu prevazići upotrebom tzv. metoda
aktivacije (hipeventilacija, fotostimulacija) ili produženjem snimanja u specifičnim fiziološkim
stanj ima (spavanje nakon dužeg perioda nespavanja).
PET, fMRI, EP...
Registracija bioelektričnih procesa
EEG -> elektroencefalografija
Registruje moždanu el. aktivnost preko elektroda smeštenih na poglavini (najčešće) ili subduralno (unutarmoždanog tkiva). Rezultat je dijagram koji prikazuju elektroencefalografske talase.
CT je precizno snimanje glave, bolje za koštane delove nego za moždanu masu.
registrovanje neprekidne sumirane, lokalne, spontane bioelektricne aktivnosti
dopunsko ispitivanje kore velikog mozga
Elektroencelografski talasi
Nedostaci : samo 2/3 kore se ispituje, ne može se upotrebiti za ispitivanje subkortikalnih odnosa i definisanjenekog poremecaja.
Postoje još i PET (pozitron emisiona tomografija), fMRI (funkcionalna magnetna rezonanca), EP (evociranipotencijali) i CT (komjuterizovana tomografija).
22. Evocirani potencijali (vizuelni, auditivni, somatosenzorni,kognitivni)
nop... :(
Evocirani potencijali su promene u električnoj aktivnosti nervnog sistema izazvane nekim događajem ilipobudom. Koriste se u dijagnostici. Izazivaju se vizuelnom, auditivnom, ili magnetnom stimulacijomsenzornih ili motornih puteva.
Analize izazvanih potencijala u senzornim i motornim zonama.
Vrste:
1. Vizuelni (VEP) - najveći evocirani potencijal, najlakše se snima i najosetljiviji je na promene uzrokovaneneurološkim oboljenjima.Mogu se izazvati različitim vizuelnim stimulacijama.
2. Auditivni (AEP) - za ispitivanje funkcije akustičnog puta počev od unutrašnjeg uha, do kortikalnogakustičnog centra u temporalnom režnju.

by J.M. & Ranagol, gen 2010 27
3. Somatosenzorni (SSEP) - ispitivanje funkcije senzornog sistema na nivou perifernih nerava, senzitivnihspinalnih korenova, kičmene moždine, talamusa i senzitivne kore.
4. Kognitivni - relativno novi elektrofiziološki test za ispitivanje poremećaja kognitivnih funkcija. Primena u dgneuroloških i psihijatrijskih oboljenja odraslih i dece.
Evocirani potencijali (EP) predstavljaju promene u električnoj aktivnosti nervnog sistema
izazvane nekim događajem odnosno pobudom. Danas su EP postali standardna dijagnostička
metoda u klinikama. U kliničkim ispitivanjima EP se izazivaju vizuelnom, auditivnom,
magnetskom ili električnom stimulacijom senzomih ili motomih puteva. U širem smislu pod EP
se podrazumeva i odziv mozga kao rezultat kognitivnih procesa i mehanizma _pripremanja koji
prethodi motornim akcijama. Q i
l.Vizuelni (VEP) - ovo je najveći evocirani potencijal, pa se najlakše snima i najosetljiviji je na
promene uzrokovane neurološkim oboljenjima. Mogu se izazvati različitim uobičajnim i
neubičajnim vizuelnim stimulacijama.
2. Auditivni(AEP) - za ispitivanje funkcije akustičnog puta, počev od unutrašnjeg uva do kortikalnog akustičnog centra
u temporalnom režnju.
3. Somatosenzorni (SEP) - koriste se za ispitivanje funkcije senzitivnog sistema na nivou
perifernih nerava, senzitivnih spinalnih korenova, kičmene moždine, talamusa i senzitivne kore.
4. Kognitivni - predstavlja relativno novi elektrofiziološki test za ispitivanje poremećaja
kognitivnih funkcija. lma primenu u dijagnostici neuroloških i psihijatrijskih oboljenja, kako
odraslih, tako i dece.
23. Ostale metode ispitivanja CNS-a (Rtg, MR, MRA, MR spect, PET)
CT: kompjuterizovana tomografija. Za desetak minuta se može dobiti snimak mozga po
slojevima. Mogu se videti i krvni sudovi. Manje steti a daje bolju sliku
Rtg - rengensko zračenje. To su elektromagnetni talasi, koji mogu da prodiru kroz telo, i mogu se posle detektovati.
Rentgensko zračenje se može koristiti za dijagnostiku, ali i za terapiju, npr.: uništavanje malignih stanica.
MR - magnetna rezonanca, je postupak konstruisanja slike visoke rezolucije struktura živog
mozga, na osnovu merenja valova koje emituju atomi vodonika aktivirani radiofrekventnim
valovima u magnetnom polju. Snaga aparata MR tj. magnetnog polja meri se u jedinicama tesla,
a jedan teslaje 10 000 gausa. MR niskog intenziteta je ispod 0,5 tesla, srednja efikasnost je od
0,5 do 1 tesla, I visoko izražena iznad 1 tesla.
Magnetna rezonanca daje 10 puta bolju rezoluciju i precizniji snimak nego CT. Organizam se
izlaže magnetnim zracima. Slika se dobija kompjuterskim putem, sa odbijanjem magnetnih
talasa. Problem: ovo ne može svako da radi. Često se uradi CT, pa ako je potrebno, onda se radi
magnetna rezonanca. Gleda se gde je najveća moždana aktivnost. Dosta je skupa metoda.

by J.M. & Ranagol, gen 2010 28
MRA - magnetno rezonantna angiografija, radi se o novijem dijagnostičkom postupku koji
nije štetan i invazivan, a daje dobre rezultate posebno u prikazu krvnih žila mozga i vrata, u
kombinaciji sa MR-om tih regija.
MR spect - magnetno rezonantna spektroskopija, iz odabranih delova tkiva mozga kao i
patološki promenjenih tkiva se dobijaju spektri/signali pomoću kojih se može, kao i kod obične
spektroskopije zaključiti o molekulima koji se nalaze u tkivu. Iako je ponegde rutinska metoda
ipak se retko koristi.
PET- pozitronska emisiona tomografija, predstavlja tehniku neuroodslikavanja koja se
zasniva na merenju razlika u protoku krvi koje su povezane sa moždanom aktivnošću. Njome se
detektuju pozitivno naelektrisane čestice koje emituje radioaktivna supstancija ubrizgana u
krvotok ispitanika, čime se omogućava vizualizacija aktivnih zona u mozgu
izotopska metoda, jako je skupa. Ubrzgava se određeni materijal pacijentu.
Poligraf (detektor laži): aparat, koji meri reakciono vreme, puls, znojenje na dlanu, srčanu frekvenciju.

by J.M. & Ranagol 1
III DEOOBJAŠNJENJE:
1. Times New Roman: materijal iz stare skripte
2. Arial: prepisano iz različitih sveski ili rukopisa
3. Comic Sans MS: materijal sa predavanja
4. Microsoft Sans Serif: skripta gen 2010
I. Razvoj nervnog sistema
CNS se razvija iz trećeg klicinog lista
CNS se razvija u embrionalnom periodu
iz kože se izdvaja nervna ploča -> nervni usek-> nervna cev-> iz te cevi se razvija ceo CNS
teratogeni lekovi dovode do problema kod razvoja mozga. Između ostalog i svi antileptici suteratogeni.
miostatin je hormon koji određuje količinu mišića kod embriona
1. Razvoj nervnog sistema
Nervni sistem kičmenjaka se tokom embriogeneze stvara od ektoderma iz trećeg klicinog lista.
1. Stadijum :
Diferencirana ćelijska struktura na leđnoj strani ( nastaje do 18-od dana starosti kod čoveka ). Na prednjem (anteriornom ) kraju nervne ploče ubrzo po njenom stvaranju javljaju se posebna ektodermalna zadebljanja -plakode. Od njih se kasnije formiraju čulni organi ( vid, sluh, miris) i deo jedara kranijalnih živaca.
2. Stadijum :
Nervna cev : nervna ploča se uporedo sa rastom embriona izdužuje i u isto vreme ona počinje da seinvagnira ( ulubljuje ) spuštajući se postepeno svojim srednjim delom prema unutrašnjosti tela, dok njenizadebljani rubovi ostaju uzdignuti. Ovaj proces dovodi do sve većeg približavanja rubova nervne ploče, takoda se na kraju oni spoje i oko 25-og dana embrionalnog razvića formira se nervna cev, od koje će se zatimrazvijati svi delovi CNS-a kičmenjaka (nervni sistem cevastog tipa). Zadebljali rubovi nervne ploče se tokomnjenog ulubljavanja prema unutrašnjosti tela odvajaju i zaostaju u međuprostoru izmedju nervne ploče iektoderma od koga će postati koža. Tako se stvara nervna kresta (ganglionarna ploča), materijal za kasnijuizgradnju spinalnih ganglija i nekih ganglija autonomnog nervnog sistema.
Stvaranje nervne ploče u cev se odvija po sledecem redosledu :
1. najpre se rubovi ploče susreću u vratnom predelu
2. zatim počinje spajanje u glavenom i repnom pravcu

by J.M. & Ranagol 2
3. na oba svoja kraja nervna cev ostaje još izvesno vreme otvorena i preko prednjeg otvora (neuropora) onakomunicira sa spoljašnjom sredinom, dok zadnji otvor ostaje u vezi sa zadnjim crevom.
Diferenciranje nervne cevi započinje neposredno po njenom stvaranju na taj način što se prednji rastralni krajcevi proširuje u mešak, arhencephalon (pra mozak), od koga se u daljem toku razvića stvaraju svi drugidelovi mozga. Zadnji kaudalni kraj cevi ostaje sužen i on se tokom embriogeneze izdužuje i od njega nastajekičmena moždina.
3. Stadijum :
Diferencijacija prvobitnog moždanog mehura u 3 nova proširenja, odnosno mehura (kod čoveka se dostiže učetvrtoj nedelji starosti embriona) :
a. prosencephalon- prednji mozak
b. mesencephalon- srednji mozak
c. rhombencephalon- rombasti zadnji mozak
4. Stadijum :
Formiranje svih 5 moždanih mehurova koji čine mozak odraslih kičmenjaka krajem 4-e i pocetkom 5-enedelje života čoveka.
Prosecephalon se deli na :
Telencephalon - prednji mozak
Diencephalon - među mozak
Mesencephalon - srednji mozak se ne deli
Rombencephalon se deli :
Metencephalon- zadnji mozak
Myelencephalon-na završni mozak se nadovezuje kičmena moždina
U toku embriogenalnog razvića mozak trpi značajne transformacije, kako zbog prostorne preraspodelemoždane mase, tako i zbog nejednakog rasta pojedinih njegovih delova. Telencephalon se pomera znatnounapred i deli se u dve hemisfere, nesrazmerno više narasta od drugih moždanih struktura. Paralelno sestvara 5 moždanih mehurova a kasnije nastupaju i druge promene u građi mozga.
Razviće mozga u stvari započinje burnom deobom, odnosno proliferacijom nervnih ćelija a zatim i njihovomdiferencijacijom. Posle toga dolazi do migracije, nervne ćelije se na osnovu genetskog koda po određenomrasporedu razmeštaju na tačno ona mesta koja će zauzeti u odraslom nervnom sistemu. Zatim dolazi domaturacije tj. sazrevanja nervnih ćelija koje uspostavljaju striktno definisane međusobne sinaptičke veze kaoi veze sa receptorima i efektorima.
U procesu embriogeneze u njegovog poodmakloj fazi, dolazi i do odumiranje velikog broja nervnih ćelija kojesu nastale u prvoj fazi razvića, odnosno faze proliferacije nervnih ćelija. Odrasli nervni sistem sadrži određenbroj ćelija jer po pravilu po završenoj embriogenezi ne dolazi više do deobe nervnih ćelija. Pravilna

by J.M. & Ranagol 3
diferencijacija i migracija nervnih ćelija tokom razvića važan je preduslov za pravilno formiranje i za normalnufunkciju nervnog sistema odraslih jedinki.
Raspored nervnih ćelija - u toku rane embriogeneze diferenciraju se i neke kategorije glijalnih ćelija kojeulaze u sastav CNS-a gde vrše potporu i metaboličku funkciju. Dolazi do odumiranja tj. popadanja neuronausled neuspostavljanja veza i funkcija.
Nervni sistem kičmenjaka se tokom embriogeneze stvara od ektoderma, iz trećeg klicinog lista.
I stadijum
Kod čoveka l8 dana starosti, ektodermalni ćelijski sloj se na leđnoj strani embriona
duž središnje linije zadeblja i na taj način se stvara posebno diferencirana ćelijska struktura koja
se naziva nervna ploča. Na prednjem (rostralnom) kraju nervne ploče ubrzo po njenom
stvaranju javljaju se posebna ektodermalna zadebljanja koja se zovu plaktode od koji se kasnije
formiraju čulni organi i deo jedara kranijalnih živaca.
II stadijum
U toku daljeg embrionalnog razvića nervna ploča se uporedo rastom embriona izdužujei
u isto vreme ona počinje da invagira (ulubljuje), i oko 25-og dana embrionalnog razvijanja formira
se nervna cev, od koje će se zatim razviti svi delovi CNS-a kičmenjaka koji se stoga i naziva
nervni sistem cevastog tipa. Zadebljali rubovi nervne ploče se tokom njenog ulubljivanja prema
unutrašnjosti tela odvajaju i zaostaju u međuprostoru između nervne ploče i ektoderma od koga
će postati koža. Tako se stvara nervna kresta ili gangiionarna ploča, materijal za izgradnju
spinalnih ganglija i nekih ganglija autonomnog nervnog sistema. Diferencijacija nervne cevi
započinje neposredno po njenom stvaranju na taj način što se prednji rostralni kraj cevi proširuje
u mešak koji se naziva arhencephalon (pramozak) od koga se u daljem toku razvića stvaraju
svi drugi delovi mozga. Zadnji kaudalni (repni) kraj cevi, ostaje sužen, ali se izdužuje i od njega
nastaje kičmena moždina.

by J.M. & Ranagol 4
III stadijum
U četvrtoj nedelji starosti embriona dolazi do diferencijacije prvobitnog moždanog
mehura u tri nova proširenja, odnosno mehura. To su prosencephalon (prednji mozak),
mesencephalon (srednji mozak) i rhombencephalon (rombasti zadnji mozak)
IV stadijum
lz stupnja trodelnog mozga prelazi se vrlo brzo, već krajem četvrte i početkom pete
nedelje od začetka čoveka, u stadijum kada se formiraju svih pet moždanih mehurova koji čine
mozak odraslih kičmenjaka. U tom procesu pgqsencefalon se deli u dva dela: u telencephalon
(prednji mozak) i diencephlon (međumozak). Srednji mozak se ne deli, dok od
rhombencephalona postaju dva meška: metencephalon (zadnji mozak) i myelencephalon
(završni mozak). Na završni mozak se nadovezuje kičmena moždina.
U toku daljeg embrionalnog razvića mozak trpi i druge značajne transformacije kako
zbog prostorne preraspodele moždane mase, tako i zbog nejednakog rasta njegovih delova.
Razviće mozga u stvari započinje burnom deobom poliferacijom nervnih ćelija, a zatim
njihovom diferencijacijom. Posle toga nervne ćelije se na “osnovu genetskog koda po
određenom redosledu razmeštaju na tačno ona mesta koja će zauzeti u adultom (odraslom)
nervnom sistemu. Posle tog procesa, migracije, dolazi do maturacije tj. sazrevanja nervnih
ćelija koje uspostavljaju striktno definisane međusobne sinaptičke veze kao i veze sa receptorima
i efektorima. U procesu embriogeneze, u njegovoj poodmakloj fazi dolazi do odumiranja
velikog broja nervnih ćelija, jer adultni nerni sistem sadrži određen broj nervnih ćelija. Pravilna
diferencijacija i migracija nervnih ćelija tokom razvića važan je preduslov za pravilno formiranje
i za normalnu funkciju nervnog sistema odraslih jedinki.
by J.M. & Ranagol 5
Celokupno nervno tkivo čoveka potiče od dela ektoderma koji se naziva neuroektoderm koji prvobitno uobliku ploče procesom neurulacije postaje nervna cev od koje potiče veći deo nervnog tkiva:
od glavenih dve trećine postaće mozak;
od kaudalne trećine nastaje kičmena moždina;
od nervnog kanala (šupljine nervne cevi) nastaće moždane komore i centralni kanal kičmenemoždine;
od neuroepitelnih ćelija koje zadržavaju sposobnost deobe i daju dve osnovne vrste ćelija:
1. neuroblaste od koji se nastati nervne ćelije;
2. glioblasti od kojih će postati glijske ćelije.
2. Morfološka podela nervnog sistemaFunkcija nervnog sistema je usaglašavanje organizma sa njegovom
okolinom i
usaglašeno delovanje organizma kao celine. Cevastog je tipa i
sastoji se od:
1. Centralnog nervnog sistema (CNS) - čine ga nervne ćelije
koncentrisane u nervnim centrima
koje čine mozak i kičmena moždina. Mozak predstavlja proširenje
prednjeg dela nervne cevi i
sastoji se iz: prednjeg (velikog) mozga, medumozga, srednjeg
mozga, malog mozga i produžene
moždine.
2. Perifernog nervnog sistema- čine ga nastavci nervnih ćelija
koji povezuju nervne centre sa

by J.M. & Ranagol 6
periferijom (čulima i organima koji reaguju na primljene draži - efektorima), čine ga dva dela:
cerebospinalni i simpatički jautonomnij nervi sistem. Cerebrospinalní nervni sistem sačinjavaju:
glaveni nervi- polaze sa mozga i ima ih 12 pari kod čoveka i spinalni nervi - polaze sa kičmene
moždine, kod čoveka ih ima 31 par, dok autonomni nervni sistem inerviše unutrašnje organe:
crevni, respiratorni, krvni, urogenitalni sistem kao i žlezde sa spoljašnjim i unutrašnjim
lučenjem.
Nervni sistem čoveka: CNS (crveno) i PNS (plavo)
Funkcionalna podela:
1. Autonomni i
2. Somatski nervni sistem.
Strukturna podela:
1. Periferni : sve senzitivna i motorna vlakna spinalnih i kranijalnih nerava, sve periferne ganglije, zatimsimpatička i parasimpatička uglavnom postganglijska vlakna.
2. Centralni : veliki i mali mozak, moždano stablo i kičmena mozdina, mozak : prednji (veliki mozak imeđumozak), srednji, zadnji (mali mozak, produžena moždina )
Funkcija nervnog sistema je u saglašavanju organizma sa njegovom okolinom i usaglašeno delovanjeorganizma kao celine. Cevastog je tipa i sastoji se od CNS-a i perifernog nervnog sistema.
Centralni nervni sistem – čine ga nervne ćelije koncentrisane u nervnim centrima koje čine mozak i kičmenamoždina. Mozak je proširenje prednjeg dela nervne cevi i sastoji se iz prednjeg (velikog) mozga,međumozga, srednjeg, malog mozga i produžene moždine.
Periferni nervni sistem – čine ga nastavci nervnih ćelija koji povezuju nervne centre sa periferijom (čulima iorganima koji reaguju na primljene draži tj. efektorima). Čine ga dva dela, a to su cerebrospinalni i simpatičkiodnosno autonomni nervni sistem. Cerebrospinalni sačinjavaju glaveni nervi – polaze sa mozga i ima ih 12pari kod čoveka i spinalni nervi – polaze sa kičmene moždine, kod čoveka ih ima 31 par. Autonomni nervnisistem inerviše unutrašnje organe: crevni, respiratorni, krvni, urogenitalni, endokrine i egzokrine žlezde.
3. Vrsta i uloga ovojnica nervnog sistema
CNS je obavijen vezivnim opnama (moždanicama) kojih kod sisara
ima tri:
1. tvrda - spoljašnja, ka lobanji;
2. paučinasta - srednja, nežna, rastresita i bogata limfom;
3. meka - unutrašnja, naleže na mozak i sadrži krvne sudove koji

by J.M. & Ranagol 7
prodíru u unutrašnjost mozga.
Prostor između moždanica je ispunjen likvorom (cerebrospinalna tečnost) tako da ceo CNS pliva
u tečnosti i na tej način se štiti od udara o tvrdu površinu lobanje i kičmeníh pršljenova.
3. Vrste i uloga ovojnica nervnog sistema
Svi delovi CNS-a obavijeni su sa 3 različite ovojnice ili opne:
1. Tvrda dura mater, je spoljašnja, nalazi se uz kost lobanje, izuzetno bolna - to su veliki slivovi
2. Paučinasta arachnoidea - je u sredini - velika količina krvnih sudova, nežna, rastresita i bogata limfom(likvorom)
3. Meka pia mater – unutrašnja, naleže direktno na mozak i sadrži krvne sudove koji prodiru u unutrašnjostmozga.
Prostor izmedju meke i paučinaste opne se naziva subarahnoidalni prostor i ispunjen je cerebrospinalnomtečnošću (likvorom) tako da ceo mozak pliva u tečnosti i na taj način se štiti od udara o tvrdu površinulobanje i kičmenih pršljenova. Subarahnoidalni prostor svih delova CNS-a je jedinstven i na nekim mestimase proširuje u tzv. cisterne, rezervoare cerebrospinalne tečnosti. Subarahnoidalni prostor je u tesnoj vezi samoždanim komorama.
Moždane opne čoveka
Crtež moždanih opni čoveka (čeoni presek mozga): 1 -subduralni prostor; 2 - subarahnoidni prostor; 3 -meka moždanica; 4 - paučinasta moždanica; 5 - tvrda moždanica; 6 - kora velikog mozga
CNS čoveka je, kao i kod drugih sisara, obavijem sa tri moždane opne: tvrdom, paučinastom i mekom. Osimšto ga obavijaju, ove opne mu pružaju potporu, štite ga i pregrađuju njegovo tkivo. Upala moždanih opninaziva se meningitis, bolest koja može imati ozbiljne posledice.

by J.M. & Ranagol 8
4. Likvor - nastanak, količina, tok, funkcije elektroliti i belančevine
normalno oko 140-150 ml
dnevno se obnovi 4-8 puta
uloga: zaštita pri traumama i potporna uloga mozgu.
ima i transportnu ulogu, a i čisti nervni sistem
količina likvora osciluje zbog bolesti
sa godinama neuroni propadaju, a zbog toga količina likvora raste (a interkranijalni pritisak
ostaje isti)
Moždane komore, centralni kanal kičmene mozdine i subarahnoidalni prostor ispunjeni su sa liquorcerebrospinalis, to je potpuna bistra tečnost, puna elektrolita, u kojoj nema belančevina.
Nastanak: likvor luče horioidni pleksusi, on je tvorevina kapilarnih krvnih sudova moždanih komora. Likvor sekontinuirano proizvodi i uklanja resorpcijom kroz arahnoidalne resice.
Kolicina: prevashodno zavisi od osmolarnosti krvi, a manje od visine krvnog pritiska, jer osmolarnost krviodređuje intenzitet transporta natrijumovih jona u likvor, a samim tim definiše prelazak hlorida i vode. 95%likvora proizvede najbolje razvijeni horioidni pleksus lateralnih moždanih komora ( leva I i desna II hemisferavelikog mozga ). Preostalih 5% likvora nastaje u III i IV komori. Količina likvora odraslog čoveka iznosi 140 –150 ml. Dnevno se obnavlja 4-8 puta, njena količina tokom života osciluje.
Tok: likvor se pomera neprekidno pokretima treplji ependimalnih ćelija. Razlika u pritisku omogućava likvoruda teče iz lateralnih moždanih komora. Odatle preko Silvijevog akvadukta otiče ka četvrtoj moždanoj komoripa iz nje ide u siternu magna iz koje otiče u dva smera: jedan deo prelazi u subarahnoidalni prostor kojiobavija mozak i kičmenu moždinu a drugi deo se uliva u centralni kanal kičmene moždine.
Ako se cirkuacija iz bilo kojih razloga onemogući dolazi do poremecaja funkcije CNS-a. Cirkulaciju likvoraomogućavaju različiti faktori, među njima i pokreti glave.
Funkcije :
1. Amortizer- ne dozvoljava mozgu i kičmenoj moždini da udare o kost pri naglim pokretima.
2. Prevenira fizička oštećenja mozga, kičmene moždine, korena kranijalnih i spinalnih nerava i omogućavada udarac u glavu istovremeno pomera ceo mozak tako da se u momentu udara nijedan njegov deo neulubljuje, mozak i kičmena mozdina plivaju u lobanji i kičmenom kanalu, čime se obezbeđuje da se u tečnojsredini likvora velika stvarna masa mozga ponaša kao neuporedivo lakša.
3. Održavanje homeostaze međućelijskog prostora CNS-a tako što pogoduje odstranjivanju metabolika,neuroendokrinih materija i viška neurotransmitera.
4. Raznosi hormone i neke peptide po tkivu nervnog sistema.
5. Regulacija disanja, jer porast parcijalnog pritiska ugljen-dioksida u ovoj tečnosti podstiče aktivacijurespiratornog centra.

by J.M. & Ranagol 9
6. Transportna uloga
7. Predstavlja i protivtežu naglim promenama krvnog pritiska do kojih dolazi u određenim fiziološkimsituacijama.
Likvor vrši i određeni pritisak u subarahnoidalnom prostoru nervnog sistema i centralnom kanalu kičmenemoždine- intrakranijalni pritisak.
Sastav: bistra tečnost u kojoj su rastvorene minimalne količine glukoze, proteina, ćelijskih elemenata(limfociti, monociti, deskvamirani epitel) aminokiseline, joni ...
Likvor se koristi kao dijagnostički materijal za utvrđivanje postojanja meningitisa. Uzima se lumbalnompunkcijom.
Moždane komore i centralni kanali kičmene moždine, kao i subarahnoidni prostor,
ispunjeni su tečnošću specifičnog sastava koja se naziva likvor ili cerebrospinalna tečnost.
Cerebrospinalnu tečnost luče horoidni pleksusi, tvorevine kapilarnih krvnih sudova moždanih
komora. lzlučena cerebrospinalna tečnost prelazi iz lateralnih komora u treću moždanu komoru i
najzad preko silvijevog akvadukta u četvrtu komoru i odavde delimično u središnji kanal
kičmene moždine, a takođe i u subarahčidni prostor. Cirkulaciju cerebrospinalne tečnosti
omogućavaju različiti faktori među njima i pokreti glave. Ako se cirkulacija iz bilo kojih razloga
onemogući dolazi do poremećaja funkcija CNS.
Likvor je potpuno bistra tečnost u kojoj su rastvorene minimalne količine glukoze,
proteina i različitih anorganskih jona inače prisutnih u krvi, a takođe se u njoj nalazi mali broj
belih krvnih zrnaca. Količina likvora odraslog normalnog čoveka iznosi oko 135ml i ona ima
zaštitnu ulogu : moždano tkivo pliva u tečnosti i time je zaštićeno odpotresa. Osim toga likvor
predstavlja i protivtežu naglim promenama krvnog pritiska do kojih dolazi u određenim
fiziološkm situacijama.
5. Krvno - moždana barijera (građa, značaj) ne postoji samo u hipotalamusu i u retikularnoj formaciji
omogućava da mozak ne bude intoksiciran
krvnu-moždanu barijeru grade endotelne ćelije krvnih sudova
oko njih su astrociti, hranljive materije koje prođu barijeru idu dalje
kičmeno-moždana barijera je uslovna: ovde lako prolaze sve mikrosolubilne materije, i
nastavljaju ka mozgu
o Mikrosolubilne materije: kiseonik i ugljen-dioxid lako prolaze.
o Olakšanom difuzijom prolaze aminokiseline i glukoza

by J.M. & Ranagol 10
jedine ćelije koje ne zahtevaju transport glukoze su nervne ćelije
Nalazi se između krvi i neurona. Grade je:
1. Endotel i bazalne membrane kapilara meke moždane ovojnice
2. Okolni Astrociti, čiji produžeci skoro potpuno obavijaju zid moždanih kapilara
Razvoj krvno moždane barijere se završava u prvim godinama života. Prolazak materijala
kroz krvno-moždanu barijeru obrnuto je proporcionalan njihovoj veličini, a upravo je
proporcionalan liposolubilnosti. Zato imali i liposolubilni molekuli (voda, glukoza, kiseonik,
ugljen- dioksid, alkohol, narkotici...)
Kroz ovu barijeru mogu da prođu i molekuli koji nisu
liposolubilni (npr. aminokiseline), ali se to dešava samo kada u ćelijskim strukturama krvno
moždane barijere postoje specifični transportni proteini za njih.
Hematoencefalna barijera je vrlo značajna za opstanak i funkcionisanje nervnog tkiva jer
održava konstantnu sredinu oko neurona, poseb o osetljivih na promene kalijumovih,
kalcijumovih, magnezijumovih, vodonikovih jona i glikoze. Uz to ona štiti nervni sistem od
endotoksina (npr. od bilirubina) i egzotoksina, prevenira prelazak neurotransmitera
moždanih sinapsi u krv i ne dozvoljava ulazak u neurone hormonima proteinske i peptidne
prirode.
5. Krvno-moždana barijera
To je višestruka barijera, nalazi se između krvi i mozga, mozga i likvora i krvi i likvora. Nema je na dva mesta– hipotalamus i retikularna formacija (centar za povraćanje). U hipotalamusu ne postoji jer on održavahomeostazu svih materija u organizmu. Grade je endotelne ćelije kapilara koje imaju tesne veze i astrocitikoji se nalaze oko njih. Ona je uslovna. Lako prolaze sve solubilne materije (lepkovi). Nekada propada, kaonpr. zbog encephalitisa. Moždane infecije su uglavnom hronične.
Grade je:
1. Endotel i bazalne membrane kapilara meke moždane ovojnice
2. Okolni astrociti čiji produžeci skoro potpuno obavijaju zid moždanih kapilara ( nožice astrocita pružajukapilarima mehaničku potrporu i ne dozvoljavaju im da se rašire pri porastu krvnog pritiska, između njih ikapilarnog zida obrazuju se prisne veze koje u potpunosti prekrivaju površinu kojom astrociti naležu nakapilarni zid).
Razvoj krvno-moždane barijere završava se u prvim godinama života (ova barijera obezbeđuje homeostazneuslove i štiti neurone od promene raznih parametara krvi ).
Prolazak materija kroz krvnu-moždanu barijeru je obrnuto proporcionalan njihovoj veličini a direktno jeproporcionalan liposolubinosti. Zato mali i liposolubilni molekuli (voda, glukoza, kiseonik, ugljen-dioksid,alkohol, steroidni hormoni, anestetici, narkotici) brzo difunduju preko krvno-moždane barijere u neurone.Kroz ovu barijeru mogu da prođu i molekuli koji nisu liposolubilni ali se to dešava samo kada u ćelijskimstrukturama krvno-moždane barijere postoje specifični transportni proteini za njih. Veliki molekuli soli žucnihkiselina i proteina prolaze sporo ili uopšte ne prolaze kroz ovu barijeru i hormoni, proteinske, peptidne prirode

by J.M. & Ranagol 11
koji su u krvi vezani za specifične proteinske nosače uopšte ne mogu da prođu kroz ovu barijeru, ne prolazeni acetilholin a i neurotoksična žučna boja bilirubin sve dok je u krvi vezana za albumine, ne mogu da prođuni vodnikovi joni krvi, a ni amonijak ( osim pri oštećenju jetre ).
Delovi CNS-a u kojima nama hematocefalne barijere su : neurohipofiza i eminencija medijana, hipotalamus,horoidni spletovi, lokus ceruleus, cirkumventrikulani organi, endokrine žlezde, epifiza i adenohipofiza kojefunkcionalno i ne pripadaju organima nervnog sistema. Delovi gde nema ove barijere: hemoreceptivne zone.
Značaj hematocephalne barijere :
Vrlo važan za opstanak i funkcionisanje nervnog tkiva jer :
1.Održava konstantu sredinu sredinu oko neurona (posebno osetljivih na promene koncentracije K, Ca, i Hjona i glikoze krvi)
2. Štiti nervni sistem od endotoksina ( npr. od bilirubina ) i egzotoksina
3. Prevenira prelazak neurotransmitera moždanih sinapsi u krv.
4. Ne dozvoljava ulazak hormona proteinske i peptidne prirode
Krvno-moždana barijera
Za efikasno funkcionisanje CNS-a neophodna je stabilna unutrašnja sredina koju obezbeđuje krvno-moždana (hemato-encefalitična) barijera. Ona se ostvaruje na tri nivoa:
funkcionalnom, osobinom endotelskih ćelija kapilara mozga da spreče kretanje molekula izmeđuarterijske krvi i neurona;
morfološkom, endotelske ćelije su u direktnom kontaktu sa glijalnim ćelijama;
biohemijskom, u krvnim sudovima mozga je prisutna velika količina enzima koji razlažuneurotransmitere i tako sprečavaju da oni iz krvi pređu u moždano tkivo.
Ova barijera ne postoji kod novorođenčadi već se obrazuje u toku prve godine života što se vremenskipiklapa sa pojavom glijalnih ćelija.
6. Vaskularizacija moždanog tkivaOko l5% minutnog volumena srca ide u moždani krvotok. Ovaj krvotok spada u
terminalne, pa začepljenja kranijalnih krvnih sudova (moždani infarkt) mogu biti veoma opasna.
Važna karakteristika moždanih kapilara je njihova manja propustljivost u odnosu na ostale
kapilare kontinuiranog tipa, jer ih sa svih strana podupiru stopala astrocita koja im pružaju
fizičku potporu i sprečavaju njihovo prekomomo istezanje u slučju porasta krvnog pritiska.
Meka moždana ovojnica vrlo labavo prijanja uz zid moždanih krvnih sudova tako da između nje
i neurona postoje perivaskularni prostori. Oni grade specijalizovani limfatični sistem mozga,
preko koga se odstranjuju mrtvi leukociti i ostali produkti infekcije. Budući da u mozgu nema
pravih limfnih sudova male količine proteina krvi, koji redovno izlaze u intersticijum, napuštaju
moždano tkivo uglavnom kroz te prostore i prelaze u likvor subarahnoidealnog prostora.

by J.M. & Ranagol 12
Osim što lokalno reguliše sopstveni protok krvi, moždano tkivo učestvuje i u kontroli
visine sistematskog krvnog pritiska, tako što se .pri smanjenom prilivu krvi u mozak aktivira
personalni centar i pokreće vazokonstrikciju krvnih sudova velikog krvotoka, što rezultira
porastom sistematskog krvnog pritiska - ishemijska reakcija mozga.
Krvotok mozga je specifičan, tokom godina dolazi do suženja krvnih sudova. Krv u mozak dolazi na 2 različitanačina:
1. Karotidna arterija – leva i desna karotida. 1 litar krvi/min prođe kroz mozak, 350 ml po svakojkarotidi/min, ukupno 700 ml.
2. Vertebrobazilarni sliv – 250 ml/min
Na bazi mozga se formira šestougao krvnih sudova koji se zove Vilisov šestougao ili prsten. Karotide dajuarteriju cerebri mediu, one se spajaju u arteriju cerebri lateralis.
Vilisov šestougao – kod njega su dva krvotoka povezana, ne dozvoljava insuficijenciju krvotoka. Kodmladih osoba zbog stresa, endotel krvnih sudova se odvaja i krv zatvara volumen krvnog suda. Stavlja sestent (žičana metalna cev koja se postavlja u krvni sud, prianja uz zidove i drži ga otvorenim) ili se radi balondilatacija. Mozak se štiti jer ima autoregulaciju krvotoka, 60 – 170 mmHg. Normalan krvni pritisak 80 – 120mmHg. U moždanom stablu se nalaze osnovni centri regulacije krvotoka, respiracije itd (čvor života) iz kogse vrši regulacija krvotoka. Noradrenenergična vlakna sužavaju krvne sudove mozga.
Sinhrono regulisanje se odvija lokalno – u delovima mozga koij su aktivni oslobađaju se materije kojelokalno šire krvne sudove.
Holinergična vlakna šire krvne sudove. Penumbra – delovi neurona gde su neuroni poluživi. Kora velikogmozga bez cirkulacije može da izdrži 2,5 min, moždano stablo 10-20 min, a kičmena moždina 20-30 min. Ukičmenoj moždini se nalaze najelementarniji nervni centri. Mozak je zaštićen krvno-moždanom barijerom.
Oko 15% minutnog volumena srca ide u moždani krvotok. Ovaj krvotok spada u terminalne što znači dazačepljenja kranijalnih krvnih sudova (moždani infarkti) mogu biti veoma opasna. Važna karakteristikamoždanih kapilara je njihova manja propustljivost u odnosu na ostale kapilare kontinuiranog tipa jer ihpodupiru astrociti koji sprečavaju njihovo prekomerno istezanje u slučaju porasta krvnog pritiska.
Meka moždana ovojnica vrlo labavo prijanja uz zid moždanih krvnih sudova. Između nje i neuronapostoje perivaskularni prostori koji grade specijalizovani limfatični sistem mozga, preko koga se odstranjujumrtvi leukociti i ostali produkti infekcije.
Krvotok mozga nastaje spajanjem a.vertebralis u a. basilaris koja se na bazi mozga povezuje sa dveunutrašnje karotidne arterije dajući Willsov most. Od njega se odvajaju arterije za ishranu moždanog tkivatako što prve 2/3 mozga uglavnom ishranjuju krvni sudovi nastali grananjem unutrašnjih karotidnih arterija

by J.M. & Ranagol 13
dok zadnju trečinu (pons, produženu možzdinu i mali mozak) prokrvljuju ogranci bazilarne arterije. Venskakrv mozga skuplja se u venske sinuse, a iznosi se putem jugularnih vena.
Vazokonstrikcija i vazodilatacija moždanog krvotoka su uglavnom dobro lokalno kontrolisaneautoregulacijom, tako da se protok krvi kroz mozak ne menja pri variranju sistemskog krvnog pritiska od 4-26kPa ali kada padne ispod 4kPa vazomotorni krvotok reaguje vazodilatacijom.
Vazokonstrikcija je slabije izražena u mozgu nego u ostalim organima i uglavnom nastaje usled lokalnognakupljanja metabolita.
Vazokonstrikcijske i vazodilatacijske materije krvi ne prolaze kroz hematocephalnu berijeru jer nisuliposolubilne.
Iako pod normalnim uslovima preovlađuje autoregulacija moždanog krvotoka, u vreme intenzivnog mišićnograda, kada sistemski krvni pritisak značajno raste, simpatička stimulacija postaje dovoljno jaka da izazoveznačajnu konstikciju cerebralnih arterija kojom se mozak štiti od porasta sistemskog krvnog pritiska. Tokomstresnih stanja, paralelno sa opštom vazodilatacijom mozdanih vazomotora usled koje nastupa nesvestica, au težim situacijama dolazi i do potpunog gubitka svesti.
Osim sto lokalno reguliše sopstveni protok krvi, moždano tkivo učestvuje i u regulaciji visine sistemskogkrvnog pritiska, tako što se pri smanjenom prilivu krvi u mozak aktivira presom centar i pokrećevazokonstrikciju krvnih sudova velikog krvotoka što rezultira porastom sistemskog krvnog pritiska -ishemijska reakcija mozga.
Dva sistema:
Arterijski :Karotidni -> 300-350 ml/min ( X2 )veliki mozakmedju mozaksrednji mozak
Vratni -> 300 ml/minkičmena moždinaprodužena moždinaponsmali mozak
Venski -> venski sinusiodlike: autoregulacija -> lokalnog krvnog pritiska, sistemskog krvnog pritiska.60-170 mmHg
7. Metabolizam mozgaMozak čini 2% ukupne mase tela, a moždani metabolizam troši čak 20% minutnog
volumena srca. Moždano tkivo ima aeroban "metabolizam, tako da zahteva: velike količine
kiseonika, ali njegova potrošnja nije konstantna već je uslovljena stepenom aktivnosti različitih
moždanih struktura. Mozak nema mogućnost deponovanja kiseonika. Neuroni su osetljivi na
hipoksiju, najosetljivija je kora i subkorteks, a najmanje je osetljiva kičmena moždina.
Metabolizam mozga poseduje autoregulaciju, neuroni su okruženi sa oko tri kapilara i

by J.M. & Ranagol 14
vaskulamo su zaštićeni od izumiranja.
Pri maksimalnom angažovanju metabolizma, CNS se udvostručuje, usled jakog
intenziviranja aktivnosti brojnih jonskih pumpi koje rade isključivo na račun energije raspada
ATP-a za čije dobijanje su potrebni oksidacioni procesi.
Metabolizam mozga obavljaju likvor i krvno moždana barijera. On je veoma intenzivan. Neuron ne posedujeglikogen i nema anaerobni metabolizam. Mozak iznosi 2% telesne težine, metabolizam mozga u stanjumirovanja iznosi 20 % ukupnog metabolizma. Od 170g glukoze, mozak troši 100g. Protok krvi je 45-50ml/100g moždanog tkiva. Krupni neuroni su obezbeđeni od izumiranja jer dobijaju po 3 kapilara, a sitnineuroni leže u spletu kapilara.
Iako mozgu pripada svega 2-2,5% telesne mase ljudi, moždani metabolizam u uslovima mirovanja čini oko15% ukupnog metabolizma.
Moždano tkivo ima aeroban metabolizam, tako da zahteva velike količine kiseonika, ali njegova potrošnja nijekonstantna vec je uslovljena stepenom aktivnosti različitih moždanih struktura. Mozak nema mogućnostdeponovanja kiseonika. Količina kiseonika koja se dobija sagorevanjem glikoze iz astrocita potroši se otprilikeveć za dva minuta.
Posle desetak sekundi anoksije javlja se nesvestica, a kada se nakon dve-tri minute istroše glikogenskerezerve astrocita prestaje metabolizam, dok smrt nastupa posle svega 2-5 min (pri niskim temperaturama tose dešava nesto kasnije). Pritom je somatski deo nervnog sistema osetljiv na hipoksiju a naročito brzo seoštecuju kora i subkorteksi, a najmanje je osetljiva kičmena moždina.
Pri maksimalnom angažmanu metabolizma funkcija CNS se udvostručuje usled jakog intenziviranja velikogbroja jonskih pumpi koje rade isključivo na račun energije raspada ATP-a za čije dobijanje su potrebnioksidativni procesi. Neuroni su zavisni od glukoze krvi jer imaju male glikogenske rezerve (depo u astrocitimaje dovoljan za svega 90 sekundi rada mozga). Oni katabolišu glukozu pretežno aerobnom glukolizom ikrebsovim ciklusom, a svega 3-5% glukoze metabolišu pentozofosfatnim putem. Kora velikog mozgametaboliše i masne kiseline iz kojih može dobiti od 60% ukupne energije, zatim laktate koji maksimalnodaju 35% energije akcionih potencijala i u sinaptičkoj transmisiji.
U mozgu postoji i vrlo intenzivan anabolizam (stvaranje materija), a posebno je izražena sinteza strukturnih ifunkcionalnih proteina i mnoštvo drugih molekula koji učestvuju u regulaciji polarnosti, nastanku AP-a i usinaptičkoj transmisiji.
8. Morfologija kičmene moždine kičmena moždina je dugačka 42-45 cm
prostire se u kičmi do drugog lumbalnog pršljena (a ne u celoj dužini kičme)
liquor se uzima tamo, gde nema kičmene moždine (ispod 2. lumbalnog pršljena)
k.m. je uvek segmentirane građe
prečnik k.m. je oko 1 cm
k.m. ima 31 segmenata:

by J.M. & Ranagol 15
1. 8 vratnih segmenata
2. 12 grudnih
3. 5 lumbalnih
4. 5 sakralnih
5. 1 repni
1811.-1822.- formulisan je Bel-Magendijev zakon (C. Bell, Škotska, F. Magendi, Francuska). Autori
su pokazali da senzitivna nervna vlakna (aferentna) vode do kičmene moždine kojoj se priključuju
kao deo leđnog spleta kičmenih nerava, dok motorna vlakna izlaze iz kičmene moždine kao deo
ventralnog spleta kičmenih nerava.
k.m. se posle presecanja potpuno inaktivira, ovo se zove SPINALNI ŠOK. U slučaju vršenja
eksperimenta sa k.m. životinja, treba sačekati neko vreme, da bi k.m. počela samostalno da radi
posle spinalnog šoka sledi faza hiperrefleksije. Tada nervni
centri i releksni lukovi već rade, ali rade neadekvatno i
neodmereno.
svaki pokretni deo tela ima svoj segment, kome šalje svoje
signale.
Dermatomi su poprečni odresci tela, koju opslužuje isti spinalni
nerv.
levo i desno prepokrivanje aferentnih funkcija nije podeljena
tačno sa linijom, dakle postoji prepokrivanje. Ako se pacijent
žali da ne oseća polovinu tela "tačno od polovine" to ne može biti organskog porekla - zbog
prepokrivanja. To znači da onda tu imamo psihičke razloge.
kod bočnih rogova (presek k.m., oblik leptira, negde na krajevima krila "leptira") se nalaze
centri simpatikusa
u sakralnim segmentima (od S2 do S4) se nalaze centri parasimpatikusa. Parasimpatikus
kontroliše npr. mokrenje ili defekaciju. Povreda ili disfunkcionalnost parasimpatikusa može
dovesti do nevoljnog mokrenja ili defekacije. Ovi ljudi su psihologu najteži pacijenti.
interneuroni povezuju različite nervne centre
k.m. je centar refleksa. Zaštitni refleksi su uvek polisinaptički.
refleksi istezanja mišića: tu su receptori mišićna vretena: oni gledaju izduženost mišića,
detekcija dužine mišića.
tetivni refleksi su osetljivi na napon mišića

by J.M. & Ranagol 16
Medulla spinalis je deo centralnog nervnog sistema koji je smešten u kičmenom kanalu,
kod čoveka zauzima samo gomje dve trećine kičmenog kanala. Ukupna dužina kičmene možldine
čoveka kreće se u proseku od 40-45cm, dijametar l-l.5cm, a težina joj iznosi oko 30w.
Valjkastog je oblika, nejednake debljine, najšira je na vratnom segmentu, a najuža na repnom
segmentu gde se završava šupljina kroz koju cirkuliše likvor.
Kičmena moždina se može podeliti na: vratni deo, grudni deo, slabinski deo i krstačni
deo.
Dve uzdužne brazde na lednoj i trbušnoj strani dele kičrnenu moždinu na desnu i levu
polovinu. Kičmena moždina je segmentirana zbog koštanih pršljenova, gde svakom odgovara po
jedan par od spinalnih živaca. Kod čoveka ima 31 pari spinalnih živaca: 8 pari vratnih, 12 pari
grudnih, 5 pari lumbalnih, 5 pari krstačnih i jedan par trtičnih.
Raspored sive i bele mase je obmut u odnosu na veliki mozak - bela masa je spolja, a
siva se nalazi unutra. Siva masa na poprečnom preseku daje oblik latiničnog slova H, naime, ona
obrazuje jedan par leđnih ijedan par trbušnih rogova. Sa leđnih rogova polaze senzitivna, a sa
trbušnih motoma nervna vlakna pa se po izlasku iz kičmene moždine sjedinjuju u zajednički
nerv. Belu masu čine uzlazna i silazna nervna vlakna kojima se provode nadražaji ka mozgu kao
i impulsi od mozga ka raznim organima. _Kičmena moždina je prema tome sprovodnik nadražaja
u rav nervna v a na ao ` " " ` ' a)..
Osim toga, onaje centar za mnoge proste reflekse.
Kičmena moždina se prostire celom dužinom tela. Idući ka zadnjem kraju tela ona se sužava. Cilindričnogje oblika i ima centralni kanal u sredini. Sa obe stane kičmene moždine polaze nervi koji su segmentalnoraspoređeni – ima ih onoliko koliko i kičmenih pršljenova.

by J.M. & Ranagol 17
Presek kičmene moždine
Raspored sive i bele mase je obrnut u odnosu na veliki mozak – bela masa je spolja, a siva se nalazi unutra.Siva masa na poprečnom preseku daje oblik latiničnog slova H, naime, ona obrazuje jedan par leđnih i jedanpar trbušnih rogova. Sa leđnih rogova polaze senzitivna, a sa trbušnih motorna nervna vlakna pa se poizlasku iz kičmene moždine sjedinjuju u zajednički nerv. Belu masu čine uzlazna i silazna nervna vlaknakojima se provode nadražaji ka mozgu kao i impulsi od mozga ka raznim organima. Kičmena moždina jeprema tome sprovodnik nadražaja u pravcu mozga (uzlazna nervna vlakna) kao i impulsa koji se šalju odmozga (silazna vlakna). Osim toga, ona je centar za mnoge proste reflekse.
8. Morfologija kičmene moždine

by J.M. & Ranagol 18
Medulla spinalis je najstariji deo CNS-a smešten u kičmenom kanalu van lobanje. Zauzima 2/3 zapremineCNS-a. Pokretljiva je u svom kanalu. Ukupna dužina kod čoveka kreće se 40 – 45 cm, a širina 1-1,5cm,težine 30g. To je izdužzena struktura valjkastog oblika. Prostire se od vratnog do lumbalnog segmentakičmenog kanala. Ona je nejednake debljine: najšira na vratnom segmentu, najtanja na repnom segmentugde završava kičmenim končićima. U sredini se nalazicentralna šupljina kroz koju cirkuliše likvor. Dve uzdužnebrazde na leđnoj i trbuđnoj strani dele kičmenu moždinuna desnu i levu polovinu. Segmentirana je zbog koštanihpršljenova na 31 segment (pršljen) gde svakomsegmentu odgovara 1 par spinalnih nerava: vratni 8,grudni 12, lumbalni 5, krsni 5, trtični 1.
Sastoji se od 2 sloja, sive i bele mase.
Raspored sive i bele mase je obrnut u odnosu na velikimozak – bela masa je spolja, a siva iznutra.
1. Siva masa : centralni, unutrašnji deo. Napoprečnom preseku daje oblik latiničnog slova H (oblikleptira koji daje zadnje rogove – ulaz za aferentnavlakna i prednje rogove – izlaz za motorna vlakna), čijekrakove popunjava bela masa. Krakovi sive mase sezovu stubovi. Postoje tri para stubova sive mase odkojih se prednji zove columna ventralis ili anterior, zadnjicolumna dorsalis ili posterior a bočni par se zovecolumna lateralis. Neuroni sive mase se grupišu u 9 slojeva ili lamina.
Iz kičmene moždine polazi 31 par mešovitih spinalnih živaca koji sadrže motorna i senzorna vlakna. Vezaizmeđu kičmene moždine i spinalnih živaca ostvaruje se pomoću dorzalnih i ventralnih korenova spinalnihživaca. Dorzalni korenovi sadrže senzitivna (aferentna), a ventralni motorna (eferentna) vlakna, te su spinalninervi mešoviti i sadrže obe kategorije vlakana.
Spinalna ganglija se nalazi na dorzalnim korenovima spinalnih nerava i ona sadrži neuronska telasenzornih neurona.
Siva masa obrazuje 1 par leđnih i 1 par trbušnih rogova. Sa leđnih rogova polaze senzitivna, a sa trbušnihmotorna nervna vlakna, pa se po izlasku iz kičmene moždine sjedinjuju u zajednički nerv. Čine je telaasocijativnih neurona i motoneurona, dok su tela senzitivnih neurona van kičmene moždine u spinalnojgangliji, zatim glija celije i nemijalisana nervna vlakna.
2. Bela masa : nalazi se spolja i okružuje sivu masu. Čine je uzlazna i silazna nervna vlakna kojim sesprovode nadražaji ka mozgu kao i impulsi od mozga ka raznim organima. Kičmena moždina je prema tome,sprovodnik nadražaja u pravcu mozga (uzlazna nervna vlakna), kao i impulsa koji se šalju od mozga (silaznavlakna).To su pretežno mijelinizirani aksoni čija se neuronska tela nalaze u sivoj masi same moždine ili udrugim moždanim strukturama. Nervna vlakna bele mase grupišu se u snopove i pomoću njih se stvarajuveze kako između pojedinih segmenata same kičmene moždine, tako i između moždine i viših delova CNS-a.Aksonski snopovi koji povezuju različite segmente kičmene moždine čine asocijativne puteve koji mogu bitiushodni ili nishodni u zavisnosti od toga da li se impulsi prenose ka nižim ili višim segmentima od onih ukojma se nalaze neuronska tela datih aksona. Asocijativne puteve čine snopovi relativno kratkih nervnih

by J.M. & Ranagol 19
vlakana. Nervna vlakna pomoću kojih se ostvaruje veza između moždine i viših delova CNS-a su relativnodugačka. Oni se zovu putevi ili tractusi. Oni mogu biti:
1. Ascedentni ili ushodni – prenose informacije sa periferije u pojedine delove mozga.
2. Descedentni ili nishodni – dele se na dve velike grupe:
- Piramidalni (kortikospinalni) – povezuju koru velikog mozga sa kičmenom moždinom. Postojedva piramidalna puta – piramidalis i anterior. Oni potiču iz precentralnog gyrusa cortexa telencephalona.Većina piramidalnih nerava su mijelinizirana. Nervni impulsi koji se prenose ovim vlaknima obezbeđujukontrolu specifičnih voljnih pokreta.
- Ekstrapiramidalni putevi – aksoni u sastavu ovih puteva vode poreklo iz različitih subkortikalnihstruktura od kojih su mnoge u vezi sa motornim zonama kore prednjeg mozga i značajni su za podešavajepoložaja tela i udova.
9. Funkcije kičmene moždine. Kičmena moždina kao najstariji deo CNS je vrlo važna, jer je ona posrednik aferentnim
putevima, i tako omogućava da mi doživimo različite osećaje, i na njih da nadogradimo
percepciju (spaja se elementarni osećaj sa pamćenjem).
Osećaji iz kože i potkožnih tkiva su bazični osećaji. Bazično je zato, jer se na ovo najviše
oslanjamo. Postoji analigoja između kože i ćelijske membrane jednoćelijskog organizma. I zato
je važna kičmena moždina.
Kičmena moždina je centar refleksnih reakcija. Refleksi kičmene moždine se mogu klasifikovati na više načina:
Prema broju segmenata
1. Segrnentalni - pojedinačni segmenti kao centar: zavise od pojedinačnih segmenata
2. Intersegrnentalni - više segmenata kao centar, angažuju se VIŠE segmenata
Prema položaju receptora
1. Površinski - koža i površina tela: izazivaju se stimulacijom kože ili površine tela
2. Dubinski - mišići i tetive: nagla konstrakcija mišića pri istezanju tetive
3. Viscelarni -unutrašnji organi: npr.: sekrecija pljuvačke pri unošenju hrane u usta
Prema kategoriji efektora pomoću kojih se ostvaruju
1. Somatski
1. refleksi fleksije (tetivni i miotatički) - koordinacija
2. refleksi ekstenzije - zaštitna uloga
3. refleksi ukrštene ekstenzije - mišićni tonus
4. postulami - podupiranje i koordinacija pokreta očiju
5. lančani - refleks hodanja, ritmička smena fleksije i ekstenzije

by J.M. & Ranagol 20
6. centralni program - aktivacija serije neurona u sastavu određenih neuronskih mreža koje su genetski
programirane
2. Viscelarni
1. refleks širenja zenice
2. refleks vazomotorike
3. refleks disanja .
4. refleks kontrole defekacije i mikcije
Masovni refleksi - istovremeno aktiviranje svih segmenata autonomnih refleksa
9. Funkcija kičmene moždine
Kičmena moždina je centar refleksnih reakcija, dve osnovne funkcije : refleksna i sprovodna.
Refleksna funkcija se sprovodi preko nervnih centara motornih i vegetativnih refleksa. Specijalnimotoneuroni inervišu respiratornu muskulaturu, međurebarne mišiće i dijafragmu i tako se obezbeđujurespiratorni pokreti. Vegetativni neuroni inervišu sve unutrašnje organe. Važni su i anospinalni centri kojiregulišu defekacije, vezikospinalni centar koji reguliše refleks mikcije, centri za regulaciju seksualnih funkcija.
Sprovodna funkcija se manifestuje u sprovođenju impulsa sa periferije ka višim delovima CNS-a, kao i usprovođenju impulsa iz viših delova kičmene moždine.
Aferentni putevi kičmene moždine povezuju razne receptore trupa, vrata i extremiteta sa delovima CNS-a.
1. Tractus spinotalamicus sprovodi impulse za osećaj dodira, bola i termičke osetljivosti.
2. Tractus spinobulbotalamicus sprovodi impulse za čulne utiske tonusa mišića, položaja extremiteta izglobova, a delimično i sa kožnih receptora.
3. Tractus spinocerebelaris sprovodi impulse za složene pokrete.
Eferentni putevi povezuju srednji mozak i preoduženu moždinu sa motoneuronima kičmene moždine a prekonjih skeletne mišiće trupa i extremiteta:
Tractus kortikospinalis sprovodi impulse za voljne pokrete.
Tractus rubrospinalis sprovodi impulse za regulaciju tonusa extenzora.
Centar refleksnih radnji – stereotipni pokreti (najjednnostavniji) i niži integrativni centar motornihfunkcija.
Podela :
Prema segmentima : segmentalni- pojedinačni segmenti
intersegmentalni- više segmenata kao centar (refleks hodanja)
Prema receptorima : Povrsinski - koža
Dubinski - mišići i tetive
Visceralni - unutrašnji organi

by J.M. & Ranagol 21
Prema efektorima :
refleksi fleksije ( tetivni i miotatički ) - koordinacija
refleksi ekstenzije - zaštitna uloga
refleksi ukrštanja ekstenzije - mišićni tonus
postularni - podupiranje i koordinacija pokreta očiju
lančani - refleks hodanja, ritmička smena fleksije i ekstenzije
centralni program = aktivacija serije neurona (generatori obrasca) u sastavu određenih neuronskihmreža koje su genetski programirane.
Visceralni - niži centri simpatikusa i parasimpatikusa - refleks širenja zenica, refleks vazomotorike(poprečno-prugasti mišići prilikom hlađenja,zagrevanja), refleks disanja, refleks intenstinalnog trakta, reflekskontrole defekacije i mikcije.
Masovni refleksi -> istovremeno aktiviranje svih segmenata autonomnih refleksa (hiperfunkcija)
Spinalni šok -> stanje totalne arefleksije posle preseka kičmene moždine
10. Funkcionalni značaj centralnog i perifernog motoneuronaPerifemi neuron je sastavni deo svakog refleksnog luka, nalazi se van CNS.
Periferni motonenroni
motoneuron prednjih rogova kičmene moždine
alfa motoneuron
kod lezije se javlja: lokalizovana slabost, hipotrofija, hiporefleksija ¡ hipotonus
Centralni neuron je svaki neuron koji na bilo koji način koordinira funkciju periferije, tj.perifernog neuronli.
Centralni rnotoneuron
iz kore, moždanog stabla, retikularne formacije
kod lezije se javlja: globalna slabost, nema hipotroñje, zatim hiperrefleksija, hipertonus, patološki refleksi
11. Morfološke karakteristike moždanog stabla na k.m. se nadovezuje moždano stablo.
Moždano stablo čine : produžena moždina, varolijev most i srednji mozak (ovo je najmanji deo,
1-1,5 cm)
dakle, moždano stablo je u lobanji, to je produžetak k.m.
moždano stablo je dugačko 6-6,5 cm

by J.M. & Ranagol 22
mali mozak sa moždanim stablom čine ZADNJI MOZAK
moždano stablo povezuje k.m. sa svim ostalim delovima CNS, i to obostrano, tj. u oba pravca
Funkcije moždanog stabla:
1. Tu se nalaze brojni centri životno važnih funkcija: npr. regulisanje brzina rada srca, kijanje,
kašljanje. Zato se ovo naziva i čvor života.
2. Funkcija ravnoteže tela, ili statika. (Ovo je centar podsvesne ravnosteže, centar svesne
ravnoteže se nalazi u parijetalnom režnju.) Ako čovek nema ravnotežu, to ga čini
izbezumljenim. Ravnotežu reguliše
o vestibulatorni sistem (najvažnije)
o vizuleni sistem: isto utiče na ravnotežu
o somatski sistem, tj. dodir i pritisak
sumacija ili konvergencija svih ovih informacija i impulsa čini ravnotežu
3. Funkcije retikularne formacije: budnost, spavanje, aktivirajuća i inhibirajuća dejstva
4. Ostale funkcije: treptanje... itd.
_______________________________________________________________________
Moždano stablo čine : produžena moždina, varolijev most i srednji mozak
PRODUŽENA MOŽDINA (medula oblongata) - direktno se nadovezuje na kičmenu
moždinu i ima oblik zaobljene kupe čiji je uži kraj na nivou velikog otvora na dnu lobanje. To je
struktura duga oko 2.5 cm, čija se glavnina nalazi na podu IV moždanekomore. Sastoji se iz sive
i bele mase i u njoj prestaje njihov pravilan raspored kakav je :postojao u kičmenoj moždini,
mada je siva masa i dalje locirana centralno. Njenu sivu masu grade motorna jedra VIII-Xll
kranijalnog nerva, nukleus i kuneatus, neka jedra retikulame formacije i jedra donje olive, dok je
bela masa nastavak ushodnih i nishodnih puteva kičmene moždine. Osim ovako organizovane
sive i bele mase, u produženoj moždini se nalaze još i mreža sive i bele mase koja pripada
retikularnoj formaciji.
VAROLIJEV MOST ( pons Varoli) - povezuje produženu moždinu i srednji mozak. Varolijev
most ima ulogu uodržavanju ravnoteže i položaja glave i tela u prostoru pošto prima impulse sa
receptora aparata za održavanje ravnoteže srednjeg uha. Takođe moždani most usklađuje mimiku
lica, prima nađražaje iz kože lica, uha i zuba, sadrži centre za žvakanje, sisanje, kao i centre
odbrambenih refleksa za suzenje i treptanje. Takođe u Varolijevom mostu se nalazi i centar koji
prekida spontan udisaj i time reguliše frekfencu disanja.
SREDNJI MOZAK - naročito je jako razvijen kod nižih kičmenjaka. Jednom uzdužnom

by J.M. & Ranagol 23
brazdom podeljen je na dva dela, samo kod sisara postoji i poprečna brazda koja ga deli na 4
režnja. U srednjem mozgu nema komora već se nalazi Silvijusov kanal koji povezuje lll i IV
komoru. Duž njegove osnove teku snopovi vlakana koji povezuju prednji
mozak sa produženom
moždinom i malim mozgom.
srednji mozak je 1,5 cm veličine. On je najmanji deo
moždanog stabla, ali je istovremeno i jedna od
najsloženijih struktura
nastaje iz treće moždane komore
Silvije kanal promera je oko 1 mm

by J.M. & Ranagol 24
Moždano stablo je centar brojnih refleksa i kroz njega prolazi čitav niz puteva u oba smera i on povezujekičmenu moždinu sa drugim delovima mozga. Moždano stablo je čvor života i u njemu je smeštena iretikularna formacija. Njega čine produžena moždina, Varolijev most i srednji mozak.
1. Produzena mozdina (medula oblongata)
Direktno se nadovezuje na kičmenu moždinu i ima oblik zaobljene kupe čiji je uži kraj na nivou velikog otvora(foramen magnum) na dnu lobanje. To je struktura duga oko 2,5 cm čija se glavnina nalazi na podu četvrtemoždane komore. Sastoji se iz sive mase (koja je iznutra) i bele mase (koja je spolja).
Siva masa: sastoji se uglovnom iz neuronskih tela koja su velikim delom grupisana u jedru. Tu se nalazemotorna jedra sledećih kranijalnih živaca : 12 nervus hypoglossus, 11 nervus accessorius, 10 nervus vagus, 9nervus glassopharingeus, i 8 nervus statoacusticus.
Senzitivna jedra kranijalnih živaca su slična spinalnim ganglijama i sadrže neuronska tela.
U sivoj masi se nalaze i druga filogenetski nova jedra kao sto su nukleus cuneatum gracilis a tu su i jedraretikularne formacije i jedra donje olive.
Bela masa: Tu se nalaze prominentne piramide, vlakna MLF, spinotektalni, spinotalamički, i spinocerebralnitraktusi. Nju čine snopovi nervnih vlakana kao nastavaka ushodnih i nishodnih puteva kičmene moždine kojaprolaze kroz produženu moždinu i oni koji povejuku medulu sa drugim delovima mozga ili sa kičmenommoždinom.
Na ventralnoj strani produžene moždine su ispupčenja koja se zovu piramide, postoji brazda koja deliproduženu moždinu na levu i desnu polovinu i na njenim bočnim stranama su ovalna zadebljanja olive. Uproduženoj moždini se nalazi jos i mreža sive i bele mase koja pripada retikularnoj formaciji. Sam početakčetvrte moždane komore se naziva romboidna jama.
2. Varolijev most (pons Varoli)
Povezuje produženu moždinu i srednji mozak. Ima ulogu održanja ravnoteže i položaja glave i tela u prostorupošto prima impulse sa receptora vestibularnog aparata unutrašnjeg uha. Takođe, moždani most usklađujemimiku lica, prima nadražaje iz kože lica, uha i zuba, sadrži centre za žvakanje, sisanje i centre odbrambenihrefleksa za suzenje i treptanje. Takođe, u Varolijevom mostu se nalazi centar koji prekida spontan udisaj itime reguliše frekvenciju disanja. To je ventralni deo zadnjeg mozga koji se proteže sve do moždanihkrakova. Dužina ponsa iznosi 2,5 do 3 cm. Na ventralnoj i lateralnoj površini mosta izraziti su poprečnisnopovi nervnih vlakana pomoću kojih se ostvaruju veze između ponsa i malog mozga.
Ponsu se priključuju delovi VI i VII kranijalnog živca, nervus trigeminus (V zivac) koji je je najveći kranijalniživac i njegova jedra se nalaze takođe u ponsu.
Dorzalna strana ponsa je okrenuta prema četvrtoj moždanoj komori (obično kao i dorzalna strana produženemoždine).
Sastiji se od sive i bele mase. Strukture retikularne formacije zalaze i u pons.
3. Srednji mozak (mezencephalon)
Deo mozga koji je tokom embroogeneze trpeo najmanju transformaciju i do kraja života ostaje jedinstven.Čine ga nervne strukture koje su sačuvale valjkasti oblik sa uzanom središnjom šupljinom - Silvijevimakvaduktom koji povezuje III i IV moždanu komoru. Dorzalna strana mezencefalona je tectum mesencephali.Ventralni deo čine pedunculli cerebri ( moždani kraci ).

by J.M. & Ranagol 25
12. Funkcije produžene moždine i ponsaProdužena moždina se često naziva čvor života, jer njena povreda dovodi do zastoja u
disanju, što u krajnjoj liniji uslovljava smrt. U njoj se nalaze:
l. Vitalni centri - vazomotorni, inspiratorni, ekspiratorni i centri za regulaciju rada
srca
2. Centri većine digestivnih refleksa - žvakanja, gutanja, lučenja pljuvačke,
pokreti želuca i lučenje želudačnog soka, sisanje, podrigivanje
3. Mesto izlaska poslednjih kranijalnih nerava VIII-XII
4. Centri nekih zaštitnih refleksa: kijanje, kašljanje, povraćanje, lučenje suza i
centar komealnog refleksa
5. Bulbarni centar za defekaciju -- koordinira defekaciju sa ostalim digestivnim i
drugim refleksima, a usklađuje je i sa radom drugih centara
6. Centar za fonaciju - kontroliše formiranje glasa u larinsku
7. Održavanje statike tela i regulacija kinetike tela-poremećenost ravnoteže ozbiljno remeti mentalni život
čoveka
8. Budno sta12. Funkcija produžene moždine i ponsa
Čine zadnji deo moždanog stabla i kao i kičmena moždina, imaju sprovodnu i refleksu ulogu.
Sprovodna uloga – vezana je za prolaženje ushodnih i nishodnih puteva koji povezuju kičmenu moždinu savelikim mozgom.
Refleksna funkcija – ogleda se u tome što se u produženoj moždini nalaze vitalni refleksni centri (centarsrčane delatnosti i disanja, a tu je smešten i vazomotorni centar koji reguliše krvni pritisak i širenje krvnihsudova).
Produžena moždina se često naziva čvor zivota jer njena povreda dovodi do zastoja u disanju, što u krajnjojliniji uslovljava smrt.
1. Vitalni centri :
Vazomotorni
Inspiratorni
Ekspiratorni
Centri za regulaciju rada srca
2. Centri većine digestivnih refleksa
- žvakanje, gutanje, lučenje pljuvačke, pokreti želuca i lučenja želudačnog soka, sisanje podrigivanje.
3.Mesto izlaska poslednjih kranijalnih nerava ( VIII-XII )
4. Centri nekih zaštitnih refleksa :

by J.M. & Ranagol 26
kijanje, kašljanje, povraćanje, lučenje suza i centar kornealnog refleksa
5. Bulbarni centri za defekaciju :
koordinira defekaciju sa ostalim digestivnim i drugim refleksima a usklađuje je i sa radom drugih centara(mesojedima posebno važni respiratorni centri, jer moduliraju disanje tokom defekacije)
6. Centar za fonaciju
kontroliše formiranje glasa u larinksu.
7. Održavanje statike tela i regulacija kinetike tela
8. Budno stanje i spavanje.
13. Morfo-funkcionalni pregled moždanih nerava
POGLEDAJ J.M. SKRIPTU, TAMO JE NEŠTO SKRAĆENIJE
13. Morfo-funkcionalni pregled moždanih nerava
I. Nervus olfactorius- mirisni nerv koji služi za obradu i prijem mirisa. Sadrži samo senzitivnanervna vlakna koja su u stvari nastavci primarnih čulnih ćelija mirisnog epitela u nosnoj šupljini. Oni seprojektuju na bulbus olfactorius (mirisnu kvržicu u mozgu) i posle izvršene sinapse olfaktorni put se završavana filogenetski starim delovima kore velikog mozga.
II. Nervus opticus - optički nerv. Čisto senzitivni i sadrži samo specijalnu somatsku aferentnukomponentu. Sastoji se od aksona ganglijskog neurona mrežnjače. Posle delimičnog ukrštanja u optičkojhijazmi nervna vlakna ovog nerva se projektuju na lateralno genikulatno telo talamusa, zatim se aksoni noveserije neurona završavaju na primarnoj vidnoj zoni kore velikog mozga. Manji deo vlakana ganglijskogneurona se projektuje na mezencefalon.
III. Nervus oculomotorius- pokreće spoljašnje mišiće oka. Uglavnom motorni nerv ali se unjegovom sastavu nalaze i neka senzitivna vlakna. Kompleksno jedro ovog živca prima projekcije iz mnogihdelova mozga.
a) opštu somatsku eferentnu komponentu čine aksoni koji inervišu poprečno-prugaste mišiče, pokretačeoka, osim dva mišića (superior obliguus i lateralis rectus).
b) opšta visceralna eferentna komponenta ovog živca sadrži preganglijska parasimpatička vlakna kojainervisu sfinger pupile od koga zavisi suženje zenice, parasimpatička vlakna inervišu i cilijarne mišiće.
c) Opštu somatsku aferentnu komponentu čine aksoni koji prenose nervne impulse iz proprio receptoraonih očnih mišića koje inervišse III nerv.
IV. Nervus trohlearis - inerviše unutrašnje mišiće oka. Uglavnom motorni nerv, ali sadrži i nekasenzitivna nervna vlakna.
a) opštu somatsku eferentnu komponentu živca čine aksoni koji potiču iz motornog jedra i inervišumusculus superior obliguus, mišićni pokretač oka.
b) opštu aferentnu komponentu čine aksoni u vezi sa proprioceptorima tog očnog mišića.
V. Nervus trigeminus - najveći moždani nerv. Mešovit nerv, najveći kranijalni nerv čija suvlakna rasprostranjena po ustima, nosnoj šupljini i prednjoj polovini lobanje. Motorna komponenta inervišemišiće za žvakanje i mišiće srednjeg uha. Senzitivni deo inerviše celo lice.

by J.M. & Ranagol 27
VI. Nervus abducens - odvodi oko u stranu. Većim delom motorni nerv.
a) opštu somatsku eferentnu komponentu čine nervna vlakna koja inervišu očni mišić lateralis rectus.
b) opšta somatska aferentna komponenta prenosi nervne impulse iz proprio receptora istog mišića.
VII. Nervus facialis - inerviše žlezde za lučenje pljuvačke, stvaranje suza i prednje 2/3 jezika.Većim delom je motorni nerv koji inerviše lice, ali on sadrži i manju količinu senzitivnih vlakana.
VIII. Nervus statoacusticus - deo nerva koji se nalazi u mostu (pons) i zadužen je za održavanjeravnoteže. Senzitivni nerv čiju specijalnu aferentnu komponentu čine nervna vlakna koja prenose informacijeiz receptora unutrašnjeg uha. Deo nerva u produženoj moždini nosi informacije o zvuku. Nervna vlakna kojaprenose nervne impulse iz receptora vestibularnog aparata ulaze u sastav nervus vestibularis i projektuju sena nucleus vestibularis u produženoj moždini. Nervus cochlearis prenosi informacije koje potiču izfonoreceptora i aksoni u sastavu tog istog živca se projektuju na nucleus cochlearis produžene moždine, azatim dalje do auditivnih zona kore velikog mozga.
IX. Nervus glassopharingeus - prima osećaj ukusa iz poslednje trećine jezika, motorno inervišezavršnu pljuvačnu žlezdu. Mešovit ali uglavnom senzitivni živac čija je funkcija povezivanje jezika i dušnika.
a) opšta somatska aferentna komponenta sadrži relativno mali broj aksona koji prenose nervne impulseiz receptora kože u oblasti uha i iz auditivnog kanala.
b) opšta visceralna komponenta sadrži relativno mali broj aksona koji prenose nervne impulse izreceptora za dodir i bolzadnje 2/3 jezika, dušnika, mekog nepca i ušnog kanala. prenosi i informacijeneophodne za održavanje krvnog pritiska, srčanog ritma i za disanje.
c) specijalna visceralna aferentna komponenta se sastoji iz aksona u vezi sa gustatornim receptorimaposlednje trećine jezika.
d) specijalnu visceralnu eferentnu komponentu čine moždana nervna vlakna koja inervišu m.stylopharyingeus.
X. Nervus vagus - nerv lutalica, od ždrela do trbušne duplje, inerviše srce, pluća, ždrelo,grkljan, spoljašnje uho. Mešoviti živac.
a) opšta somatska aferentna komponenta se sastoji od aksona koji prenose nervne impulse iz kožniheksteroceptora oko uha i zadnjeg dela ušnog kanala.
b) opšta visceralna aferentna komponenta sadrži nervna vlakna koja prenose informacije iz receptora udušniku, jednjaku, grudnom košu i trbušnoj duplji.
c) specijalnu visceralnu aferentnu komponentu čine nervna vlakna koja prenose informacije iz gustatornihreceptora nepca prvo do nucleusa solitarisa, a zatim dalje do kore velikog mozga.
d) opšta visceralna eferentna komponenta se sastoji iz preganglijskih parasimpatičkih vlakana kojainervišu visceralne organe grudnog koša uključujući i srce i inervaciju trbušne duplje, uključujući organe zavarenje.
e) specijalna visceralna eferentna komponenta sadrži nervna vlakna koja inervišu visceralnu muskulaturumekog nepca, dušnika i jezika.
XI. Nervus acessorius - čisto motorni nerv. Specijalna visceralna komponenta sastoji se odnervnih vlakana koja se spajaju sa vagusom i inervišu delimično muskulaturu za pokretanje glave i ramena imišića od kojih zavise fonacija i gutanje. Kranijalni deo ovog nerva potiče iz nucleusa ambiguusa i on inervišemišiće laringsa. Spinalni deo potiče iz prvog i sedmog cervikalnog segmenta kičmene moždine i vršiinervaciju mišića glave.
XII. Nervus hypoglossus - inerviše mišiće jezika. Motorni nerv. Opštu somatsku eferentnukomponentu čine nervna vlakna koja inervišu somatsku muskulaturu jezika (omogućavaju njegove pokrete).

by J.M. & Ranagol 28

by J.M. & Ranagol 29
14. Funkcije srednjeg mozga srednji mozak je 1,5 cm veličine. On je najmanji deo moždanog stabla, ali je istovremeno i jedna
od najsloženijih struktura
nastaje iz treće moždane komore
Silvije kanal promera je oko 1 mm
PAG stvara unutrašnje opijate mozga, odgovoran je za smanjenje bolova. Dakle, stvaraju se
takvi neurotransmiteri (enkefalin), koji smanjuju bol.
Mezencefalon ima niz važnih i složenih funkcija koje su tesno povezane
sa funkcijama
drugih delova moždanog stabla.
1. Učestvuje u sistemu pomoću koga se moduliše ekscitabilnost
spinalnih motoneurona za
anti gravitacionu muskulaturu od koje zavisi uspravan stav tela
2. Ima udela i u refieksnom uspravljanju

by J.M. & Ranagol 30
3. Udeo u centralnom programu za lokomociju koji je lociran u kičmenoj moždini u vidu
specifičnih neuronskih mreža
4. Sadrži složeni jedarski kompleks III kranijalnog nerva (nervus okulomotorius) od koga
zavise pokreti oka i IV kranijalni nerv (nervus trohlearis) .
5. Sadrži crveno i cmo jedro - sistem regulisanja motorike
6. Refleksi podešavanja oka i glave
Informacije: vidnih, slušnih i kožnih stimulusa
7. Automodifikacija senzacije bola
14. Funkcije srednjeg mozga
On je najmanja moždana struktura, nalazi se oko Silvijevog akvadukta, odnosno između međumozga iVarolija. Ima sprovodnu i refleksu funkciju.
Sprovodna – sprovođenje kroz njega aferentnih i eferentnih puteva koji povezuju kičmenu moždinu iproduženu moždinu sa kranijalnim delovima CNS-a, tj malim mozgom, jedrima talamusa, bazalnimganglijama i preko njih sa korom velikog mozga.
Refleksne funkcije su mnogobrojne i raznovrsne: refleks konvergencije, zenica na svetlo i akomodaciju,orijentacioni zvučni refleks, refleks gutanja i žvakanja (supstantia nigra).
Supstanitia nigra učestvuje u regulaciji plastičnog tonusa i regulaciji preciznih, finih pokreta prstiju ruke.
Crveno jedro (nucleus ruber) igra važnu ulogu u regulaciji tonusa.
Mezencefalon ima niz važnih i složenih funkcija koje su tesno povezane sa funkcijama drugih delovamoždanog stabla. Učestvuje u sistemu pomoću koga se moduliše ekscitabilnost spinalnih motoneurona zaantigravitacionu muskulaturu od koje zavisi uspravan stav tela. Ima udela i u refleksima uspravljanja.
Udeo u centralnom programu za lokomociju tj. pokretanje koji je lociran u kičmenoj moždini u viduspecifičnih neuronskih mreža.
Sadrži složen jedarski komleks III kranijalnog nerva (nervus oculomotorius) od koga zavise pokreti oka, i IVkranijalni nerv (nervus trohlearis). Sadrži crveno i crno jedro, sistem regulisanja motorike.
Refleksi podešavanja oka i glave.
Informacije: vidnih, slušnih i kožnih stimulusa.
Automodifikacija senzacije bola.
Srednji mozak
Srednji mozak se sastoji iz krovne pločice i pedunuculus cerebri, a između njih se nalazi Silvijev kanalispunjen cerebrospinalnom tečnošću koji povezuje treću i četvrtu moždanu komoru. Na krovnoj pločici senalaze dva para kvržica: gornje i donje. Gornje kvržice su zadužene za refleksne pokrete očiju, kao ikoordinaciju pokreta očiju i glave. Iako se vizuelne informacije koje srednji mozak prima od očiju prenosepreko talamusa do kore velikog mozga, gornje kvržice na određene vizuelne nadražaje mogu da reaguju bezučešća kore velikog mozga. U donjim kvržicama se prvi put sastaju sve akustičke informacije, a zatim seprosleđuju kori velikog mozga na obradu. I one u nekim situacijama mogu samostalno da reaguju na nekezvučne nadražaje. Pedunuculus cerebri je deo tzv. sistema nagrađivanja. On je uključen u veoma važannačin učenja koji nam pomaže da preživimo. Taj sistem se aktivira kada ispunjavamo neke funkcije koje suod vitalnog značaja (ako jedemo kada smo gladni ili pijemo kada smo žedni i sl.), a zauzvrat mozak nasnagrađuje prijatnim osećanjima koja nas uče da te aktivnosti treba da ponovimo. Neke droge, kao što je

by J.M. & Ranagol 31
kokain, direktno aktiviraju ovaj sistem i na taj način pružaju zadovoljstvo i smatra se da je to razlog zbogkojeg se stvara zavisnost.
15. Retikularna formacija
Nalazi se u srednjem delu moždanog stabla, raspoređena je višecentralno
Retikularna formacija se naziva i mrežolika formacija
Sastoji se od ogromnog broja sitnih neurona, i neštomanjeg broja krupnijih neurona, koji su razbacani umoždanom stablu. Manjim delom, neki su objedinjena ujedra.
Retikularnoj formaciji nalazi i LOCUS COEROLUS: lučinoradrenalin. On je osnovni izvor noradrenalina. A ovoima izrazita aktivirajuća dejstva.
Neuroni retikularne formacije su osetljivi naneurotransmitere i produkte metabolizma.
Neuroni retikularne formacije imaju spontanu aktivnost, što je osnova budnog stanja
Retikularna formacija je jedna vrlo aktivna struktura, aktivira stanje budnosti
u donjem delu moždanog stabla (kod nižih struktura) se nalaze jedra srednje linije, koje lučeserotonin. Serotonin (neurotransmiter) ima inhibitorno dejstvo, što izaziva npr. spavanje.
postoji jedna vrsta klackalice: ili dominira inhibitorno dejstvo nižih struktura (tj. jedarasrednje linije), pa se onda luči serotonin, pa zbog toga imamo inhibitorno dejstvo (inhibicijubudnosti) (konkretno: spavanje)
ILI,
ako se ne luči serotonin, onda dominira stanje budnosti.
dakle retikularna formacija je centar i spavanja i budnog stanja (zavisi koji deo r.f. dominira)
ako se desi nešto nepredviđeno, to će probuditi veliki deo aktivirajućih neurona u r.f., i imaćemoorientaciono-istraživačko ponašanje, koje je suštinski važno za naš život. Povećaće se aktivnostmoždane kore, povećaće se budnost i pažnja.
Retikularna formacija je moždana struktura u centru moždanog stabla koju grade naizmenično postavljenislojevi sive i bele mase. Ona zapravo predstavlja složenu mrežu neuronskih kola senzornih i motornih puteva,koja se prostire kroz produženu moždinu, pons, i srednji mozak, a daje izlaze ka višim centrima u talamusu,hipotalamusu, senzornoj i limbičkoj kori i ka nižim centrima u kičmenoj moždini. U retikularnoj formaciji senalazi oko stotinu jedara. Retikularna formacija pokazuje neprekidnu spontanu aktivnost, iako su za njenoodržavanje bitne i stalne stimulacije iz raznih aferentnih puteva.

by J.M. & Ranagol 32
1. U retikularnoj formaciji produžene moždine iponsa nalaze se vitalni centri:
- Kardioakceleratorni i kardioinhibitorni centarkoji kontroliše rad srca.
- Presorni i depresorni centar koji regulišekrvni pritisak
- Inspiratorni, ekspiratorni, apneustički ipneumotaksički centar za kontrolu disanja ikontrolu dahtanja kod životinja.
2. U retikularnoj formaciji su i jedra kranijalnihnerava koja kontrolišu gutanje, povraćanje i lučenjepljuvačke.
3. U retikularnoj formaciji se nalazi kompleks polisinaptičkih puteva sluha, vida, mirisa, somatskog ivisceralnog senzibiliteta, koji gradi tzv. retikularni aktivacioni sistem za regulaciju budnosti i spavanja.Smatrana je centrom budnosti jer aktivira i održava svest. U zadnjem delu retikularne formacije su neki odcentara za spavanje, odnosno Rafe jedra koja izazivaju ne-REM fazu i locus coeculus koji dovodi do REM fazespavanja.
4. Retikularna formacija je mesto sinaptičkog prekida anterolateralnog sistema i sistema dorzalnihkolumni, zatim ushodnih visceralnih puteva koji završavaju u hipotalamusu i limbusnoj kori, a u retikularnojformaciji se sinaptički prekidaju i vlakna koja polaze iz malog mozga i bazalnih ganglija. Zato retikularnuformaciju smatramo dobro informisanim delom CNS-a kako o spoljašnjoj sredini tako i o unutrašnjostiorganizma.
5. Kroz retikularnu formaciju prolaze i neka eferentna vlakna. Kroz retikularnu formaciju prolaze i u njojse sinaptički prekidaju simpatička i parasimpatička vlakna za kontrolu rada unutrašnjih organa. Iz jedraretikularne formacije polaze nishodni putevi i ona daje izlaz za mali mozak, moždano stablo i bazalneganglije. Prisustvo motornih jedara i njihovih nishodnih puteva čine retikularnu formaciju delomekstrapiramidnog sistema koji menja aktivnost alfa i gama motornih neurona i učestvuje u recipročnojkontroli što znači da smanjuje tonus fleksora u vreme pojačanog tonusa ekstenzora i obrnuto.
16. Građa međumozga
moždano stablo se nadovezuje na prednji mozak

by J.M. & Ranagol 33
Prednji mozak se sastoji iz dva dela:
o međumozak (se nastavlja ili utapa)
o u veliki mozak
Veliki mozak ima levu i desnu hemisferu
osnovni delovi međumozga su: talamus, hipotalamus i epifiza
suština međumozga je treća moždana komora
Epifiza luči melatonin, koji je važan za spavanje
Talamus je homogena, jajolika parna struktura. Ima 40 jedra. Bela masa ga deli na tri dela.
o lateralni deo
o prednji deo: ovo se zove limbički talamus. Ovo je povezan sa limbičkim sistemom i samim
tim i sa ispoljavanjem emocija.
o medijalni deo
Talamus je osnovni relej svih aferentacija, koja dolaze o koru mozga. Ovde se integrišu svi
signali, pre nego što dospeju u koru mozga. Ovo se odnosi i na senzorne i na
nesenzorne(motorički signali) (osim detekcije mirisa) informacije.
Osim ovoga, Talamus je centar tmule, žareće boli (hronična bol). Ova hronična bol može jako da
izmeni mentalno funkcionisanje čoveka.
Tokom embriogeneze se stvara od prosencephalona.Centar za vid i nevoljne pokrete. Nadovezuje se naprednji kraj moždanog stabla, prekriven je hemisferama velikog mozga. Centralna šupljina međumozga je IIImoždana komora oko koje je smeštena moždana masa. Međumozak je ključna struktura za razumevanjefunkcionalne organizacije celog centralnog nervnog sistema.
TALAMUS (na dorzalnoj strani III moždane komore). Podrazumeva dve jajolike strukture sive mase kojegrade krov i stranice III moždane komore a povezane su masom intermediom. Oko talamusa se nalazikapsula interna koja ga odvaja od bazalnih ganglija. Kroz nju prolaze aferentni i eferentni putevi kojipovezuju talamus sa korom velikog mozga, talamus je izgrađen iz više jedara među kojima su:
a. Anteriornab. Posteriorno-lateralnac. Ventroanteriornad. Ventroposteriornolateralnae. Ventrolateralnaf. Medijalnag. Dorzolateralnah. Pulvinari. Zadnja talamičkaj. Retikularna jedra
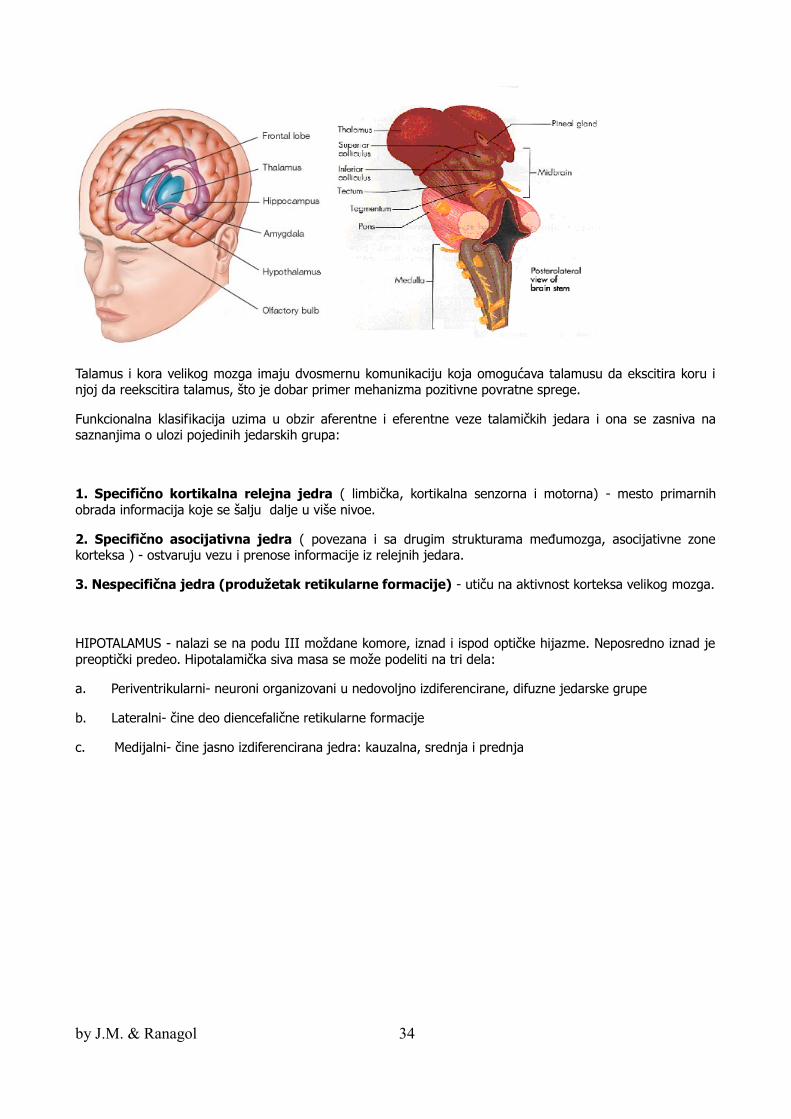
by J.M. & Ranagol 34
Talamus i kora velikog mozga imaju dvosmernu komunikaciju koja omogućava talamusu da ekscitira koru injoj da reekscitira talamus, što je dobar primer mehanizma pozitivne povratne sprege.
Funkcionalna klasifikacija uzima u obzir aferentne i eferentne veze talamičkih jedara i ona se zasniva nasaznanjima o ulozi pojedinih jedarskih grupa:
1. Specifično kortikalna relejna jedra ( limbička, kortikalna senzorna i motorna) - mesto primarnihobrada informacija koje se šalju dalje u više nivoe.
2. Specifično asocijativna jedra ( povezana i sa drugim strukturama međumozga, asocijativne zonekorteksa ) - ostvaruju vezu i prenose informacije iz relejnih jedara.
3. Nespecifična jedra (produžetak retikularne formacije) - utiču na aktivnost korteksa velikog mozga.
HIPOTALAMUS - nalazi se na podu III moždane komore, iznad i ispod optičke hijazme. Neposredno iznad jepreoptički predeo. Hipotalamička siva masa se može podeliti na tri dela:
a. Periventrikularni- neuroni organizovani u nedovoljno izdiferencirane, difuzne jedarske grupe
b. Lateralni- čine deo diencefalične retikularne formacije
c. Medijalni- čine jasno izdiferencirana jedra: kauzalna, srednja i prednja

by J.M. & Ranagol 35
Limbički sistem je jedinstven sistem sastavnjen iz hipotalamusa i filogenetski starih delovakore velikog mozga i to je jedinstven funkcionalni sistem.
EPIFIZA (krov III moždane komore)- endokrina funkcija i malatamin hormon.

by J.M. & Ranagol 36
17. Funkcije međumozga (Talamus)
Sve senzorne informacije moraju biti obrađene u talamusu, sve osim mirisnih informacija. U relejnim jedrimase vrše složene obrade informacija (ona su povezana sa primarnim senzornim zonama koje služe zaformiranje adekvatnih elementarnih senzacija). Pomoću asocijativnih jedara ostvaruju se uzajamne vezepojednih delova talamusa. Retikularna jedra regulišu stepen aktivnosti kore hemisfera velikog mozga. Odtalamusa zavisi kakav će afektni efekat imati određena senzacija – da li će izazvati osećanje dobrobiti ilinelagodnosti. Povrede talamusa mogu izazvati veoma neprijatne senzacije kao što su ako osećanjespontanog bola. Tu je centar tuge, bola, pri čemu se javlja velika depresija.
1. Relejna uloga - podrazumeva da se sve aferentne somatske informacije, deo visceralnih signala,podaci iz čula vida, sluha, ukusa i mirisa, na putu ka kortikalnim centrima sinaptički prekidaju usibkortikalnim talamičkim jedrima.
2. Integrativna - talamus prispele informacije integriše i zaključuje šta to receptori trenutno poručuju.
3. Značajan za svesnu percepciju (tumačenje) signala sa receptorasomatskog i dela visceralnogsenzibiliteta.
4. Modulira primljene informacije u saradnji sa asocijativnom korom, tako što pojačava onekoje su trenutno bitne, dok nebitne informacije umanjuje ili čak potpuno gasi, dajući jedinki sposobnost dase skoncentriše na ono što je bitno u datom momentu.
5. Centar dugotrajnog pamćenja - sposobnost traženja i nalaženja upamćenog.
6. Projekcioni sistemi:
a. Nespecifični: prima podatke iz aferentnih puteva prekinutih u retikularnoj formaciji, zatim se rejelnajedra ovog projekcionog sistema nespecifično projektuju u neokorteksu i podižu njegov opšti nivo budnosti.
b. Specifični: čine vlakna koja polaze iz jasno definisanih talamičkih jedara koja se prekotalamokortikalne radijacije projektuju u tačno definisanim kortikalnim centrima sa ciljem da povećaju pažnjuprema određenim informacijama.
7. Kroz talamus prolaze i u njemu se sinaptički prekidaju motorni putevi koji povezuju malipozak i bazalne ganglije sa korom velikog mozga.
Talamus predstavljaju dve ovalne strukture sive mase koje grade krov i stranice treće moždane komore. Utalamusu se nalaze mnoga jedara u kojima se sinaptički prekidaju mnogi putevi koji se projektuju ka kori

by J.M. & Ranagol 37
velikog mozga i služe za održanje njene budnosti. Sa druge strane i iz kori velikog mozga polaze puteviprema talamusu.
Talamus ima relejnu, integrativnu i modulatorsku funkciju. Njegova relejna uloga podrazumeva da se sveaferentne informacije koje idu ka centralnom nervnom sistemu (bol, dodir, termičke draži), i svesni dubokisenzibilitet na pragu kortikalnim centrima, sinaptički prekidaju u talamičkim jedrima. Ovo omogućavatalamusu da ih integriše i zaključi šta to receptori poručuju senzornoj kori velikog mozga. Zato je i samtalamus bez senzorne kore značajan za svesnu percepciju signala sa receptora somatskog senzibiliteta.Osim ovoga talamus može i da moduliše primljene informacije u saradnji sa asocijativnom korom, tako štopojačava one koje su trenutno bitne dok nebitne informacije umanjuje ili ih totuno gasi omogućujući da sejedinka skoncetriše na ono sto joj je bitno u određenom momentu.
18. Funkcije međurnozga (Hipotalamus)-OVO NEĆE BITI NA ISPITU, IZJAVILA JE
IVETIĆKA
Hipotalamus ima endokrinu ulogu jer proizvodi neke neurohormone i .reguliše sintezu i
oslobađanje hormona adenohipofize u krv. On je glavni izlazni p' ut limbusnog sistemai 'jedan od
centara "vegetativnog nervnog ~sisten1a, l<Oj¡. funkcioniše kao viši centar za kontrolu. rada
_unutrašnjih organa jer koordinira viscelarne refleksečiji centri su 'umoždanom stablu ( zakrvni
pritisak, disanje, rad srca i dr.) i kičmenoj moždini (zadefekaciju, mikciju, polne reflekse i dr.).
Hipotalamus održava homeostazu tako što reguliše -lučenje skoro svih hormona ikontroliše tonus
simpatičkih i parasimpatičkih vlakana. U hipotalamusu se nalaze centri za glad, sitost, žeđ,
termoregulaciju, polne nagone' i reproduktivne "funkcije, kao i za regulaciju ponašanja koje
obezbeđuje zadovoljenje ovih potreba. On učestvuje u kontroli emocija i u njemu su biološki sat
i centri za budnost i spavanje.
Hipotalamus je deo međumozga (diencephalona), nalazi se u sredini limbusnog sistema i povezan je sa svimnjegovim nivoima, kao i sa mnogim drugim delovima mozga. Pored viših centara, hipotalamus šaljeinformacije ka hipofizi i u preganglijske neurone simpatikusa i parasimpatikusa koji se nalaze u tegmentumumezencephalona.
Hipotalamus ima nekoliko funkcija:
1. Neuroendokrina uloga se ogleda u proizvodnji nekih neurohormona, kao i u kontroli endokrineaktivnosti hipofize putem stimulišućih (rilizing)i inhibišućih (inhibiting) hormona, tj faktora. Hipotalamuspripada difuznom neuroendokrinom sistemu (DNES) jer proizvodi neurohormone : oksitocin, vazopresin,neurotenzin, endorfine, enkefaline idr. Rilizing i inhibiting hormoni regulišu sintezu i oslobađanje hormonaadenohipofize u krv. Neki kontrolisš jedan, a neki dva adenohipofizna hormona. Svi rilizing hormonihipotalamusa su neuropeptidi.
2. Kontrola rada unutrasnjih organa: Hipotalamus i limbusna kora se nazivaju vegetativnim(visceralnim) mozgom jer posredstvom hormona, simpaticčkih i parasimpatičkih vlakana regulišu radunutrašnjih organa na osnovu podataka iz njih samih i spoljašnje sredine. Hipotalamus svojim hormonimareguliše rad adenohipofize, a samim tim i celog endokrinog sistema i unutrašnjih organa. Hipotalamus senaziva i glavnim ganglionom vegetativnog nervnog sistema jer se u njemu nalaze viši simpatički iparasimpatički centri koji primaju informacije sa visceralnih organa, a iz njih polaze eferentna simpatička iparasimpatička vlakna koja završavaju na visceralnim organima i kontrolišu njihovu aktivnost. Ovi centriregulišu aktivnost kardiovaskularnog sistema (frekvenca srca, minutni volumen), respiratornog sistema(frekvenca i dubina disanja), glad, zeđ, apetit itd.

by J.M. & Ranagol 38
3. Regulacija gladi i sitosti: U hipotalamusu postoje dva centra za kontolu unosa hrane, centar za gladu ventrolateralnim i centar za sitost u ventromedijalnim jedrima. Kontrola funkcije centara za sitost i gladpoznat je kao glikostatska teorija gladi. Aktivnost centra za glad je pod kontrolom holecistokinina 8. Da bi sepokrenula ta inhibicija, potrebno je da poraste glikemija i podigne nivo insulina koji omogućava glukozi dauđe u neurone centra za sitost koji zato oslobađaju holecistokinin 8 i preko njega smanjuju tonusnuaktivnost ili čak potpuno inhibiraju neurone centra za glad. Stimulacijom neurona centra za sitost se javlja iosećaj zadovoljstva.
4. Regulacija unosa vode: Osecaj zeđi nastaje iz više razloga, aktivacijom osmo ili volumenskihreceptora, ishranom bogatom proteinima, suva sluznica usta i ždrela, i porast telesne temperature. Svakipojedinačno može izazvati osecaj zeđi, ali uglavnom deluje više faktora istovremeno. Preoptička regija,subfornikalni organ i organum vaskulorum laminae terminalis hipotalamusa poseduju osmoreceptore koji prihiperosmolarnosti krvi izazivaju osecaj zeđi i podsticu lucenje antidiuretičnog hormona (ADH).
5. Kontrola telesne temperature: hipotalamusni termoregulacioni centar dobija informacije izcentralnih termoreceptora za toplo i hladno, u neuronima preoptičke regije hipotalamusa i iz perifernihtermoreceptora u koži i krvnim sudovima i kombinuje ih u cilju podsticaja odavanja ili proizvodnje toplote.Kada se javi psihički osećaj pregrejanosti ili pada temperature, hipotalamus se uključuje u održavanjeizotermije regulacijom lučenja određenih hormona i kontrolom ponašanja u zavisnosti od trenutnetemperature.
6. Regulacija seksualnog ponasanja i reprodukcije: hipotalamus reguliše seksualno ponašanje ireprodukciju posredstvom neuronske mreže zadužene za kontrolu karakteristično ženskog ili muškogponašanja, i preko kontrole sekrecije gonadotropnog rilizing hormona čija sinteza započinje u fetalnomperiodu.
7. Regulacija budnosti i spavanja: U hipotalamusu se nalaze centri za budnost i neki od hipnogenihcentara. Kroz hipotalamus prolaze vlakna RAS-a koji se pružaju prema talamusu i senzornoj kori velikogmozga a služe za održavanje budnosti i stoga se smatra da hipotalamus nije jedan od centara budnosti ispavanja već je samo mesto prolaska vlakana koja učestvuju u njihovoj regulaciji.
8. Funkcija bioloskog sata: Suprahijazmatska hipotalamička jedra funkcionišu kao endogeni biološkisat za regulaciju aktivnosti koje se ritmički ponavljaju. Ona imaju bioelektričnu (pejsmejkersku) aktivnost samanje ili više izraženim dnevnim i noćnim varijacijama. Ova jedra su povezana sa mrežnjacom iz kojedobijaju vidne signale na osnovu kojih učestvuju u regulaciji smenjivanja budnosti i spavanja, kontroliunutrašnjih organa shodno trenutnom stanju u okolini. Bez uticaja spoljašnje sredine, i to uglavnom svetlostikoja iz nje dolazi, biološki sat bi diktirao cikluse koji bi se smenjivali na 25 sati – cirkadialni ritam. Postoje iultradialni ritmovi koji se ponavljaju na manje od 24 sata i sezonski ritmovi koji podrazumevaju različitoponašanje u različitim meterološkim uslovima.
9. Kontrola ekspresije emocija: Hipotalamusni centri za nagradu i kažnjavanje su najvažniji centri zakontrolu telesnih aktivnosti, nagona i motivacije jer se senzorni doživljaji koji ne uzrokuju nagradu ni kaznuteško pamte. Hipotalamus ostvaruje uticaj na emocije posredstvom retikularne formacije.
19. Građa malog mozgaMali mozak (lat. cerebellum) je deo mozga koji se nalazi na krovu IV moždane komore. Uloga mu je uglavnompokretanje. Ima dve hemisfere između kojih se nalazi centralni crv. Unutrašnjost ispunjava bela masa (mijelinskavlakna ili provodnici moždanog signala). Mali mozak usklađuje pokrete mišića, prima podatke o stanju u kojemsu mišići i ispravlja greške u kodu koji šalju ostali delovi mozga za pokrete.

by J.M. & Ranagol 39
nalazi se iza moždanog stabla, ispod velikog mozga, u zadnjojlobanjskoj jami
gradi rombencefalon - ovo je iz nekog razloga bitno mali mozak je zadužen za kontrolu motorike razlikuje se:
1. siva masa: na površini, gradi jedro, nalazi se okolo-kora. Čine je 3 sloja.
2. Bela masa: unutrašnjost. centralni deo se naziva crv ili Vermis Hemisfere:
1. Intermedijalni deo: distalna muskulatura2. lateralna zona: planiranje motorne aktivnosti
Organizacija malog mozga:1. Archicerebellum-Vestibulocerebellum: odgovoran za ravnotežu. Najstarija.2. Paleocerebellum-Spinocerebellum: u bliskoj je vezi sa kičmenom moždinom, dobija od k.m. jako puno
informacija. Mlađa je od Archicerebelluma. Zadužena za održavanje stava, položaja ekstremiteta3. Neocerebellum-Putocerebellum: veliki mozak nema direktne veze sa malim mozgom, nego preko ponsa.
Planira motorne radnje. Ispravlja, usklađuje, predviđa pokrete. mali mozak funkcioniše na nesvesnom nivou mali mozak ima ulogu regulatora, to radi nesvesno m.m. je uključen u motorno učenje i motorno pamćenje m.m. određuje obrt ekscitacije (npr. dokle da se diže ruka), anticipacija pokreta sve ćelije u kori m.m. su inhibitorne
Oštećenje cerebelluma: Dismetrija - promašivanje cilja Ataksija - nekoordinisani pokreti Hipermetrija - prebačaj cilja Disdijadohokinezija - poremećaj u nadovezivanju Intenacioni tremor - akcioni, pred ciljem dolazi do podrhtavanja
19. Gradja malog mozga (cerebellum)
Mali mozak se nalazi iza velikog mozga a iznad ponsa, produžene moždine i žetvrte moždane komore.Podeljen je na 3 režnja- prednji, zadnji i najstariji flokulonodularni režanj vermisa. Postoji i funkcionalnapodela prednjeg i zadnjeg režnja na centralnu usku traku od deset lobusa - vermis i dve hemisfere koje supodeljene na intermedijarnu i lateralnu zonu.
U malom mozgu se nalazi polovina svih neurona, a on daje samo 10 posto ukupne težine mozga. Njegovspoljašnji deo čini siva kora, izgrađena od tri sloja neurona, a unutrašnja bela masa je organizovana u vidustabla života – arbor vitae. Na kori su mnogobrojni sulkusi koji razdvajaju nabore . Ispod kore malog mozganalaze se parna, spontano bioelektrično aktivna subkortikalna jedra sive mase : nucleus dentatus , nc.globosus, nc. Emboliformis i nc fastigii. Nucleus dentatus je najveće jedro malog mozga, koje šaljeinformacije u motornu koru velikog mozga i nucleus ruber radi koordinacije i realizacije voljnih pokreta.Nucleus fastigii obezbeđuje održavanje ravnoteže u uspravnom položaju.

by J.M. & Ranagol 40
U mali mozak pristižu informacije iz:
Kortikoponto cerebelarnog puta koji nosi podatke o nameri
Olivocerebelarnog puta koji donosi proprioceptivne informacije
Retikulo i spinocerebelarnog puta koji nose podatke iz telesnih proprioceptora
Vestibulocerebelarnog puta koji nosi podatke iz čula ravnoteže
Kuneocerebelarnog puta koji donosi proprioceptivne podatke iz glave i vrata
Tektocerebelarnog puta koji donosi vidne i slušne informacije iz kolikula mezencefalona
Ulazna i izlazna vlakna malog mozga su najbrža koja postoje, tip I aferentna i Aα eferentna.Sva ulaznavlakna malog mozga se razdvajaju na dva dela od kojih jedan ide u duboka jedra a drugi u koru. U skorosvim delovima malog mozga nalaze se topografske prezentacije pokreta do kojih dolaze aferentni signali izodgovarajućih delova tela. One imaju dvosmernu komunikaciju sa odgovarajućim topografskim oblastimamotorne kore velikog mozga i motornim oblastima moždanog stabla a posle obrade šalju informaciju uMKVM, nucelus ruber i retikularnu formaciju.
Postoji i longitudinalna funkcionalna podela prednjeg i zadnjeg režnja na centralnu usku traku od 10lobusa vermus i dve hemisfere koje su podeljene na indermedijarnu i lateralnu zonu.
U malom mozgu se nalazi preko 50% svih neurona a daje svega ukupno 10% težine mozga. Njegovspoljašnji deo čine siva masa koja čini koru cerebeluma i sastoji se iz 3 sloja nervnih ćelija:
1. Molekularni sloj koji se nalazi spolja :
Korpaste ćelije
Stelatne ćelije (vrsta interneurona, u ovom slučaju se nalaze i paralelna vlakna- aksoni neuronačija su tela u 3. sloju kore cerebeluma i ona stvaraju sinapsu sa purkinje neuronima 2. reda)
2. Sloj purkinje ćelija
3. Unutrašnji granularni sloj (u njegov sloj ulazi i manji broj neurona tipa Goldzi II)
Na kori postoje mnogobrojni sulkusi koji razdvajaju nabore (folije- paralelno postavljene vijuge u čijim seunutrašnjostima nalaze mijelirana vlakna )
20. Funkcije malog mozga
postoji kraća varijanta u skripti J.M.
Mali mozak učestvuje u kontroli motorike, ali on ne može inicirati pokrete, vec dobija plan za njih izasocijativne kore velikog mozga, i na osnovu tih podataka koordinira aktivnost mišića kako bi pravilno vodiopokret. On je neophodan za otpočinjanje i završavanje voljnih pokreta. Kontrolom agonističkih iantagonističkih mišićnih grupa tokom voljnih pokreta on reguliše put i preciznost, odnosno cilja pokrete.Preciznost obezbeđuje korekcijom pokreta koji traju. Takođe on pomaže kori velikog mozga u planiranjunarednog u nizu pokreta, delić sekunde unapred dok aktuelni pokret još traje , stoga je neophodan zakontrolu brzih i preciznih pokreta. On ispravlja greške u pokretima i omogućava nesmetano nadovezivanjevremenski usklađenih motornih aktivnosti(glatko klizanje pokreta). Za pravilno izvođenje pokreta suneophodne i povratne informacije o promeni dužine i tonusa mišićnih celija tokom odvijanja voljnih pokreta

by J.M. & Ranagol 41
na osnovu kojih mali mozak koriguje mišićni tonus u cilju prevencije narušavanja ravnoteže – anticipacijski(feedforward) mehanizam.
Mali mozak je neophodan i za učenje i pamćenje motornih obrazaca pošto se u njemu nalaze depoi motornihengrama. On može modifikovati motorne engrame na bazi stečenog iskustva, što znaci da ovaj deomotornog sistema uči na svojim greškama, tako da svaki ponovljeni pokret koriguje sve bolje dok ne postaneadekvatan.
Dakle, on:
1.Učestvuje u kontroli motorike, ali kao ni bazalne ganglije ni on ne može da inicira pokrete već dobija planza njih iz asocijativne kore velikog mozga i na osnovu njega koordinira aktivnosti različitih mišića kako bipravilno vodio pokret :
1.1. Otpočinje i završava voljni pokret - kontrola voljne motorike ( Na početku kretanja pomažestvaranje brzih uključujućih signala za mišiće agoniste i isključuje signale za antagoniste, da bi kadaprestane kretanje obezbedio vremenski usklađene isključujuće signale za agoniste i isključujućesignale za antagoniste )
1.2. Ciljanje pokreta - regulacija mišićnog tonusa (kontrolom međuigre agonističkih i antagonističkihmišićnih grupa). Za vreme izvođenja pokreta mali mozak preko kortikopontocerebelarnog delapiramidalnog puta kontinuirano dobija podatke iz asocijativne kore velikog mozga o 'nameri', aistovremeno je informacija iz vestibularnog aparata i drugih proprioreceptora upoznat sa mišićnimtonusom, dok ga čulo dodira, sluha ili vida informišu o trenutnom položaju jedinke i njenih delovatela.
1.3. Pravi tzv. korektivne ili signalne greške (koje šalje nazad u motornu koru velikog mozga kako bikorekcijom pokreta koji traju obezbedio preciznost)
1.4. Pomaže kori velikog mozga u planiranju narednog niza u pokretu
1.5. Neophodan je za kontrolu brzih i preciznijih pokreta.
1.6.Glatko klizanje pokreta (ispravlja greške u pokretima i omogućava nesmetano nadovezivanjevremenski usklađenih motornih aktivnosti )
2. Održavanje ravnoteže ( kao i vestibularni aparat )
Za pravilno izvođenje pokreta su osim adekvatne stimulacije alfa motornih neurona neophodne i povratneinformacije o promeni dužine i tonusa mišićnih želija tokom odvijanja voljnih pokreta, na osnovu kojih malimozak koriguje mišićni tonus u cilju prevencije narušavanja ravnoteže - anticipacijski mehanizam.
3. Neophodan je za učenje i upamćivanje motornih obrazaca (uz bazalne ganglije) pošto se u njemu nalaze'depoi motornih engrama' on može da modifikuje motorne engrame na bazi stečenog iskustva, što znači daovaj deo motornog sistema uči na svojim greškama.
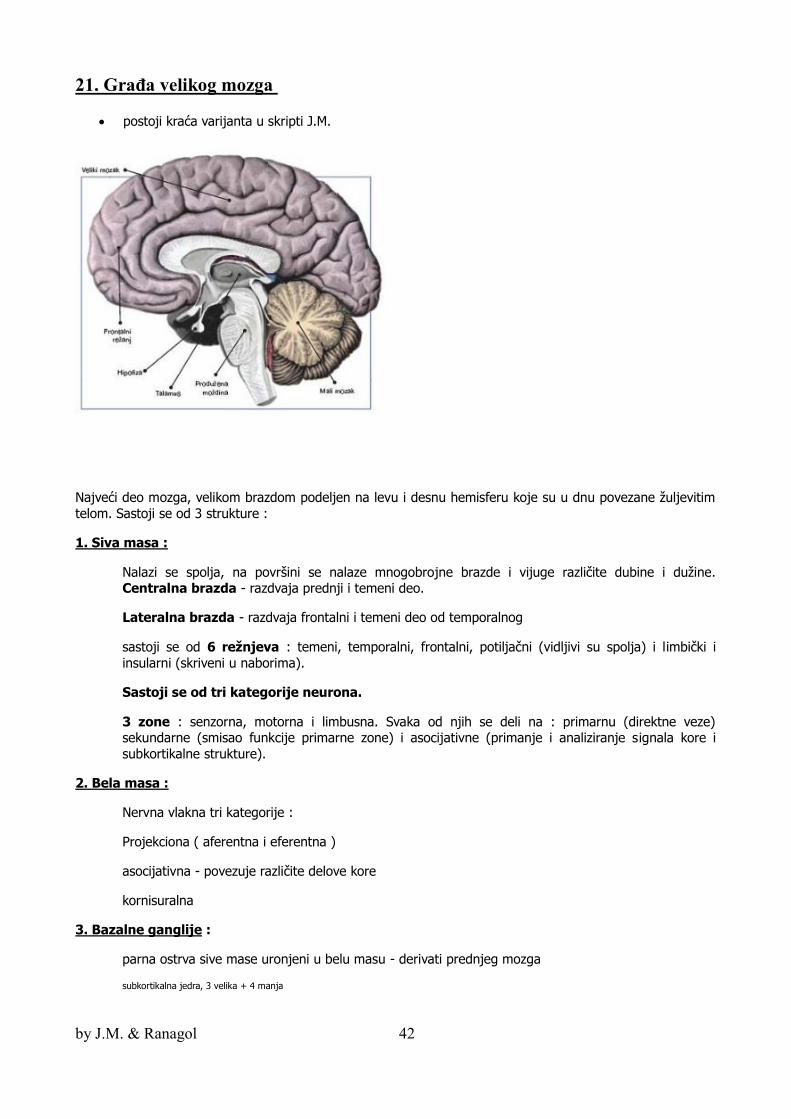
by J.M. & Ranagol 42
21. Građa velikog mozga
postoji kraća varijanta u skripti J.M.
Najveći deo mozga, velikom brazdom podeljen na levu i desnu hemisferu koje su u dnu povezane žuljevitimtelom. Sastoji se od 3 strukture :
1. Siva masa :
Nalazi se spolja, na površini se nalaze mnogobrojne brazde i vijuge različite dubine i dužine.Centralna brazda - razdvaja prednji i temeni deo.
Lateralna brazda - razdvaja frontalni i temeni deo od temporalnog
sastoji se od 6 režnjeva : temeni, temporalni, frontalni, potiljačni (vidljivi su spolja) i limbički iinsularni (skriveni u naborima).
Sastoji se od tri kategorije neurona.
3 zone : senzorna, motorna i limbusna. Svaka od njih se deli na : primarnu (direktne veze)sekundarne (smisao funkcije primarne zone) i asocijativne (primanje i analiziranje signala kore isubkortikalne strukture).
2. Bela masa :
Nervna vlakna tri kategorije :
Projekciona ( aferentna i eferentna )
asocijativna - povezuje različite delove kore
kornisuralna
3. Bazalne ganglije :
parna ostrva sive mase uronjeni u belu masu - derivati prednjeg mozga
subkortikalna jedra, 3 velika + 4 manja

by J.M. & Ranagol 43
repato jedro
putamen jedro
bledo jedro
supstanca nigra
subtalamička jedra
crveno jedro
amigdalna jedra
U procesu evolutivnog razvoja kora kao najviši deo CNS-a javlja se kasnije od ostalih delova. Deli se na dvehemisfere koje su povezane na svom dnu pomoću snopa žuljevitih tela. Pored nervnih ćelija u telencefalonusu i glijalne ćelije, a bogat je i krvnim sudovima. Sastoji se iz sive i bele mase. Siva masa je na površini i onačini korteks. Debljina korteksa varira od 1.5-4.5 mm. Kod čoveka postoje različite vrste kore. Bela masa jeiznutra, izgrađena je od mijelinskih puteva, u okviru nje su subkortikalna jedra i to su siva jedra. Svakahemisfera ima tri strane: spoljna (kojom je u kontaktu sa lobanjskom kosti), unutrašnja i donja. Svakahemisfera ima 4 režnja – frontalni (čeoni), temeni (parijetalni), potiljačni (okcipitalni) i slepoočni(temporalni). Površina nije glatka, ona je nagužvana, naborana. Uočavaju se gyrusi (udubljenja) odnosnovijuge koje su smeštene između sulcusa (žlebova). Broj sulcusa je veliki i glavni su uvek pristutni, a drugi ne.
Površina kore je 2200cm2. Korteks je sačinjen od dve vrste ćelija – piramidalnih (imaju dendrite i 1 akson) istelatnih neurona.
Vrste korteksa:
1. Neokorteks (izokorteks)
a) laminarna (stratum) organizacija korteksa, šestoslojni korteks
b) kolumnarna organizacija
2. arhikorteks – stariji, troslojan, petoslojan
a) alokorteks
b) paleokorteks
Šestoslojnu koru čine:
1. Stratum molekulare externum – radi se o molekularnim ćelijama, neuronima koji se nalaze spolja ito je najpovršniji sloj.
2. Stratum granulozum externum – spoljni granularni sloj, jedna vrsta stelatnih ćelija (spoljašnjipiramidalni sloj)
3. Stratum piramidale externum – čine ga piramidalne ćelije
4. Stratum granulozum internum – unutrašnji sloj od gusto sabijenih stelatnih ćelija, tu je prijemaferentacije u najvećem stepenu
5. Stratum piramidale internum – srednje i velike piramidalne ćelije, koje se zovu Beteove ćelije.
6. Stratum multiforme – sloj ćelija različitog oblika, najčešće vretenastog oblika

by J.M. & Ranagol 44
Od ovih lamina kreću dendritaske i aksonske veze ka drugom neuronima. Slojevi mogu biti podjednakedebljine kada pričamo o homotopičkoj organizaciji kore ali kora češće ima heterotopičku organizaciju (ufrontalnom režnju najrazvijeniji je peti stratum, a u okcipitalnom četvrti).
Kolumne su nešto što je vertikalno. Jednu kolumnu čine 5 – 10 hiljada vertikalno povezanih neurona. Njihovprečnik je 0,3 -0,5 mm, a visina 1,5 – 4,5 mm. Jedna kolumna funkcioniše individualno i do nje dolazeaferentacije iste modalnosti. Kolumne su međusobno odvojene, razdraženje se može prenositi sa jedne nadrugu. Aktivacija jedne rezultira inhibicijom njoj susednih bliskih kolumni.
Brodman je podelio koru velikog mozga na približno 52 polja. Kortikalna polja mogu biti:
1. Primarna – projekcione regije, dele se na motorna i somatosenzorna. Od primarnih motornih neuronakreću aferentacije put bazilarnih jedara. Do somatosenzornih neurona dolaze informacije iz spoljašnjeg svetai na njihovom nivou formira se osećaj.
2. Sekundarna - nalaze se oko primarnih regiona, na njih konvergiraju razdraženja i sa drugihprimarnih polja. Na nivou ovih polja vrši se sinteza osećaja, mi shvatamo, prepoznajemo. Sekundarno poljekoristi mehanizme memorije, pamćenja.
3. Tercijarna polja – najudaljeniji regioni i nalaze se samo u korteksu čoveka.
S obzirom na ova polja kora se deli na sledeće osnovne oblasti, zone ili analizatore:
1. Motorni analizator se nalazi u prednjoj vijuzi i zapravo u čeonoj oblasti. To je 4, 6, 8, 9, i 10. poljepo Brodmanu. Draženje ove zone izaziva kontrakcije skeletne muskulature na kontralateralnoj strani tela.
2. Vidni analizator – formira se u okcipitalnom režnju zahvaljujući Brodmanovom polju 17. (primarnopolje) a znamo šta smo videli zahvaljujući 18. i 19. asocijativnom polju. Pri razaranju ove oblasti nastajekortikalno slepilo gde je polje 17 u redu ali je poremećaj 18. i 19. agnozija.
3. Slušni analizator – primarno polje u temporalnom režnju. U polju 41 i 42 razumemo ono što smočuli, kao i u polju 22 koje je sekundarno.
4. Mirisni – centar je u Brodmanovom primarnom polju 11. Ako ga razorimo, nema mirisa.
5. Ukusa – primarno polje je 43. u predelu centralne brazde a okolni regioni su odgovorni zarazumevanje i prepoznavanje.
6. Ravnoteža – odgovorno je drugo primarno polje. Razaranjem primarnog polja nastaje slepilo igluvoća, a razaranjem sekundarnih agnozija (nemogućnost prepoznavanja nekog predmeta na osnovusenzornih impulsa iz ijednog čulnog organa).
7. Somestetičko polje – Brodmanovo 1,3,5,7,9 nalazi se u parietalnom režnju i pomoću njegarazlikujemo toplo, hladno, i svrab.
22. Funkcija bazalnih ganglija Veliki mozak se sastoji od leve i desne hemisfere. Veliki mozak ima sivu masu na površini (kora
mozga), i ima sive mase, koje su uronjene u belu masu. Te sive mase, koje su uronjene u belu
mase, se zovu bazalne ganglije.
bazalne ganglije imamo 3 osnovne vrste:
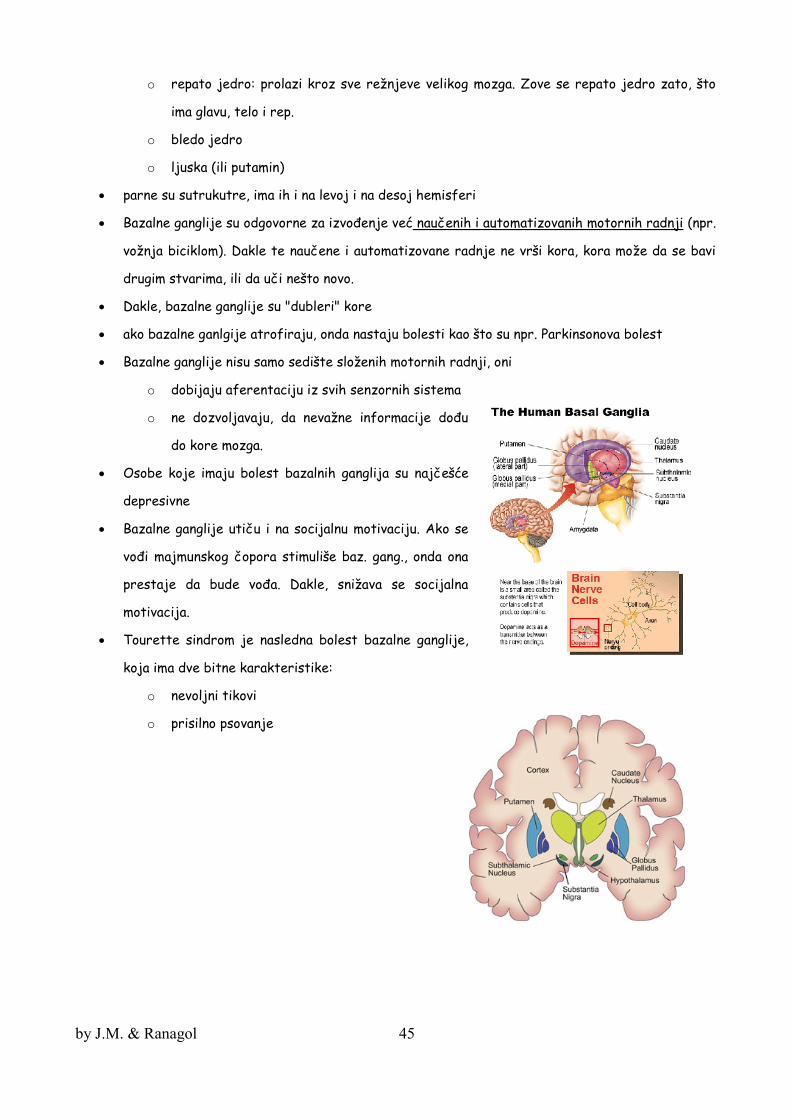
by J.M. & Ranagol 45
o repato jedro: prolazi kroz sve režnjeve velikog mozga. Zove se repato jedro zato, što
ima glavu, telo i rep.
o bledo jedro
o ljuska (ili putamin)
parne su sutrukutre, ima ih i na levoj i na desoj hemisferi
Bazalne ganglije su odgovorne za izvođenje već naučenih i automatizovanih motornih radnji (npr.
vožnja biciklom). Dakle te naučene i automatizovane radnje ne vrši kora, kora može da se bavi
drugim stvarima, ili da uči nešto novo.
Dakle, bazalne ganglije su "dubleri" kore
ako bazalne ganlgije atrofiraju, onda nastaju bolesti kao što su npr. Parkinsonova bolest
Bazalne ganglije nisu samo sedište složenih motornih radnji, oni
o dobijaju aferentaciju iz svih senzornih sistema
o ne dozvoljavaju, da nevažne informacije dođu
do kore mozga.
Osobe koje imaju bolest bazalnih ganglija su najčešće
depresivne
Bazalne ganglije utiču i na socijalnu motivaciju. Ako se
vođi majmunskog čopora stimuliše baz. gang., onda ona
prestaje da bude vođa. Dakle, snižava se socijalna
motivacija.
Tourette sindrom je nasledna bolest bazalne ganglije,
koja ima dve bitne karakteristike:
o nevoljni tikovi
o prisilno psovanje

by J.M. & Ranagol 46
Bazalne ganglije su parna ostrva sive mase smeštene u beloj masi korone radijate, odmah ispod sive masekore velikog mozga, lateralno od talamusa. One zauzimaju veliki deo dubljih oblasti cerebralnih hemisfera inazivaju se subkortikalna jedra. U njih spadaju 3 velika parna jedra, koja grade corpus striatum, a to sunucleus caudatus, putamen i globus palidus. Bazalnim ganglijama pripadaju još nucleus ruber retikularneformacije mezencefalona i amigdalna jedra limbusnog sistema.
Bazalne ganglije su, posle motorne kore, najviši centar integracije motornih aktivnosti. Ne funkcionišusamostalno, vec u vezi sa cerebralnim korteksom i kortikospinalnim putem. Ne poseduju direktne eferentneveze sa nižim motornim neuronima, vec primaju signale iz kore velikog mozga i skoro sve svoje signalevraćaju u nju.
Bazalne ganglije su pomoćni motorni sistem – one pomažu kori velikog mozga da omogući realizacijunaučenih pokreta i da planira multiple paralelne nadovezujuće pokrete kako bi bilo moguće obavljanjesmislenih motornih zadataka. Bazalne ganglije najpre dobijaju plan iz asocijativnih oblasti kore velikog mozgai zajedno sa malim mozgom na osnovu njega izrađuju program pokreta, odnosno, one razrađuju detalje iodređuju put izvođenja pokreta uz istovremenu korekciju ravnoteže.
Program pokreta prevodi abstraktnu zamisao u voljni pokret, a predstavlja set koordiniranih komandi zakontrolu mišićne aktivnosti koje su rezultat interakcije nekoliko moždanih centara. Na taj način, bazalneganglije i mali mozak omogućavaju automatsko izvršavanje sekvenci motornih programa koji su obuhvaćeninaučenim motornim planom.
Osim što prave program koji omogućava da pokret otpočne i završi se, one su potrebne i za modifikacijupostojećih obrazaca pokreta, odnosno, one usporavaju, ubrzavaju, pojačavaju ili smanjuju intenzitetpokreta, što znači da one kontrolišu i vremensko usklađivanje i dimenzije motornih obrazaca.
Subtalamička jedra bazalnih ganglija kontrolišu stereotipne (sinhrone, naizmenične, ritmične iprecizne) aktivnosti ekstremiteta, tj pokrete koji se identično ponavljaju a za koje je već napravljenprogram koji ova jedra, bez modifikacije, ponovo šalju u motornu koru. (npr. kontrola jednoličnog kasa konjapo poznatom predelu).
Bazalne ganglije su neophodne i za učenje i pamćenje motornih obrazaca jer se u njima nalazidepo motornih engrama. Osim toga, one učestvuju i u regulaciji ponašanja na taj način što mogu da otpočnuneki pokret kao reakciju na emotivne stimuluse, tj kontrolišu usputne pokrete mimike i gestikulacije kojeprate određeno raspoloženje.
Pored kontrole motorike, bazalne ganglije učestvuju i u prepoznavanju okoline.

by J.M. & Ranagol 47
FUNKCIJE:
1. Najvisi centri integracije motornih aktivnosti (posle motorne kore)
2. Predstavljaju pomoćni motorni sistem, jer ne mogu da funkcionišu samostalno, vec pomažu kori velikogmozga da omogućuju realizaciju naučenih pokreta i da planira multipne paralelne nadovezujuće pokrete,kako bi bilo moguće obavljanje smislenih motornih zadataka.
3. Izgrađuje program pokreta ( zajedno sa malim mozgom )
Dobijaju plan iz asocijativnih oblasti kore velikog mozga, odgovornih za obrasce pokreta, one razgrađujudetalje i određuju put izvođenja pokreta uz istovremenu korekciju ravnoteže, bazalne ganglije definišu kadaće pokret otpočeti i prestati, koliko će kada i koji mišići biti angažovani tokom njegove realizacije, tako dabazalne ganglije i mali mozak zajedno omogućavaju automatsko izvršavanje sekvenci motornih programa.
4. Modifikaciju postojećih obrazaca pokreta u smislu njegovog usporavanja, ubrzavanja, pojačavanja ilismanjenja intenziteta, što znači da one kontrolišu, ne samo vremensko usklađivanje vec i dimenzije motornihobrazaca.
5. Subtalamička jedra bazalnih ganglija kontrolišu stereotipne aktivnosti ekstremiteta, odnosno pokrete kojise identifikuju ponavljanjem.
6. Neophodne su za učenje i upamćivanje ( uz mali mozak ) jer se u njima nalazi 'depo motornih engrama'.
7. Kontrolišu usputne pokrete mimike i gestikulacije koji prate određeno raspoloženje.
8. Učestvuju u prepoznavanju okoline.
Chorea je bolest koja prema mišljenju velikog broja autora, nastaje pri degeneraciji kaudatusa. Ova bolest može biti i nasledna (Hantingova horea) ikarakteriše se naglim, brzim, odsečenim i neregulisanim besciljnim pokretima ekstremiteta koji podsećaju na igru uz muziku. Nevoljno se grče i mišići lica.Mišićni tonus je smanjen, te je prema tome ovo slucaj hiperkinetičkog poremecaja uz hipotoniju.
Athetosis je bolest čiji je uzrok, kako se smatra, poremećajć lenkularnih jedara, uglavnom putamena. Dolazi do ukrućenosti mišića, pri čemu su naročitokontrahovani dugački aksijalni leđni mišići. Dolazi do krutog stava i pokreti su veoma usporeni.
Balizam se karakteriše violentnim nevoljnim pokretima koji podsećaju na bacakanje tela. Veruje se da je ovaj poremećaj uslovljen lezijom subtalamičnihjedara. U slucaju jednostrane lezije pojavljuje se hemibalizam tj. balizam suprotne strane tela.
Parkinsonova bolest se karakteriše rigidnošću svih mišića i tremorom. Subjekat teško počinje da hoda iuopšte teško menja položaj tela, siromaštvo facijalnog izraza. Smatra se da je ova bolest uslovljenadegenerativnim promena supstancije nigra i striatuma.
23. Pojam i značaj subkortikalnih jedara to su sive jedra moždane mase, koja se nalaze ispod kore
Subkortikalne strukture su: bazalne ganglije (pre svega oni), talamus, hipotalamus ( i retikularna
formacija, i srednji mozak...) . Dakle svaka siva supstanca, koja se nalazi ispod kore su
subkortikalne strukture. Dakle, ovo je jako veliki pojam sam po sebi.
Smešteni su ispod korteksa telencefalona.
Pored bazilarnih ganglija u ova jedra spadaju :
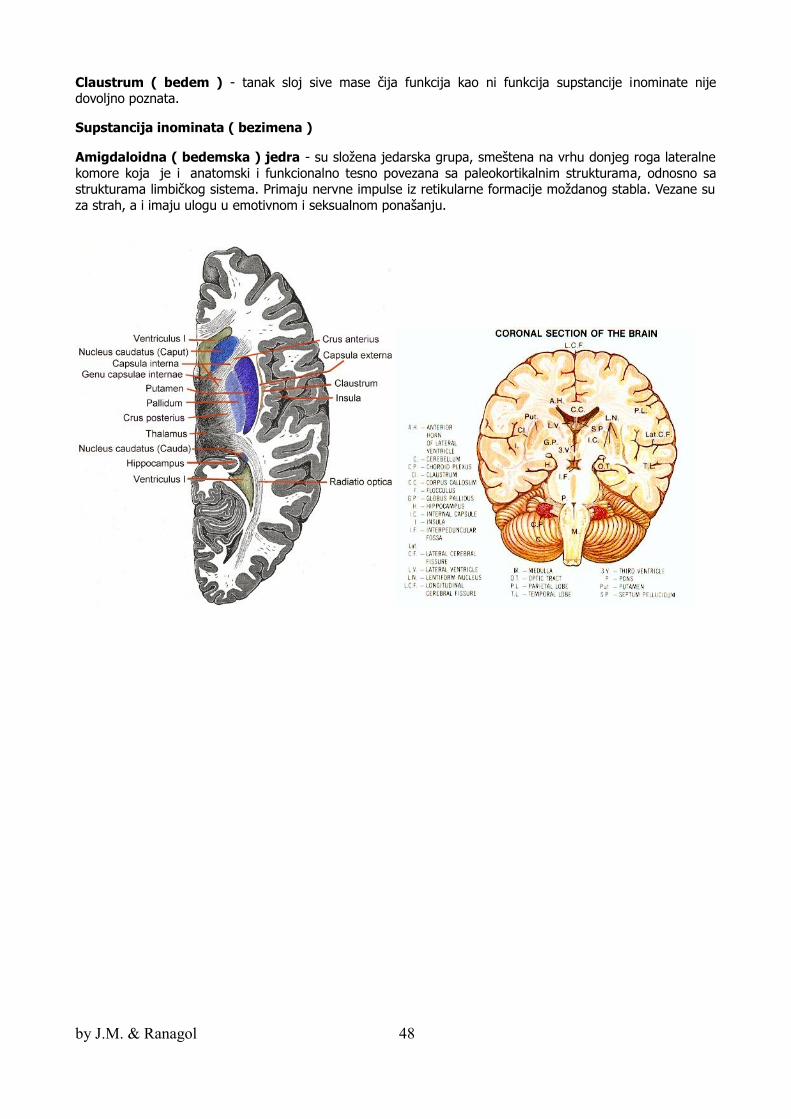
by J.M. & Ranagol 48
Claustrum ( bedem ) - tanak sloj sive mase čija funkcija kao ni funkcija supstancije inominate nijedovoljno poznata.
Supstancija inominata ( bezimena )
Amigdaloidna ( bedemska ) jedra - su složena jedarska grupa, smeštena na vrhu donjeg roga lateralnekomore koja je i anatomski i funkcionalno tesno povezana sa paleokortikalnim strukturama, odnosno sastrukturama limbičkog sistema. Primaju nervne impulse iz retikularne formacije moždanog stabla. Vezane suza strah, a i imaju ulogu u emotivnom i seksualnom ponašanju.
















