evidence for a synergistic effect of the hiv-1 envelope protein gp120 and brain-derived neurotrophic...
TRANSCRIPT

Ž .Brain Research 815 1999 349–357
Research report
Evidence for a synergistic effect of the HIV-1 envelope protein gp120Ž .and brain-derived neurotrophic factor BDNF leading to enhanced
expression of somatostatin neurons in aggregate cultures derived from thehuman fetal cortex
Ayalla Barnea a,), Jodie Roberts a, Raymond H. Ho b
a Department of Obstetrics and Gynecology, The UniÕersity of Texas Southwestern Medical Center at Dallas, 5323 Harry Hines BlÕd.,Dallas, TX 75235-9032, USA
b Department of Cell Biology, Neurobiology and Anatomy, The Ohio State UniÕersity, Columbus, OH 43210-1239, USA
Accepted 13 October 1998
Abstract
Ž .Changes in the expression of somatostatin SRIF have been observed in the brains of HIV encephalitis. Since gp120 is thought to playa major role in AIDS-associated abnormalities in the brain, we addressed the question: Does gp120 alter the functional expression ofhuman fetal SRIF neurons in culture and if so, is this effect fetal-age dependent? Aggregate cultures, obtained from cortices of nine
Ž .fetuses 14.9–20.7 weeks , were exposed for 7 days to BDNF or BDNFqgp120; BDNF induced production of SRIF during thesubsequent 24–48 h was assessed. Similar effects of BDNF and gp120 were observed in the 9 brain-cultures. A 7-day exposure to BDNF
Ž .alone led to a significant increase in SRIF production ps0.014 , whereas exposure to gp120 alone did not. Co-exposure to BDNF andŽ .gp120 led to an increase in BDNF-induced SRIF production which was significantly greater than that after BDNF alone ps0.006 .
These effects were BDNF- and gp120-dose dependent and they were not accompanied by changes in DNA content of the aggregates norin lactate dehydrogenase activity in the medium; indicating that gp120 did not lead to a major loss of cell integrity. These results are
Ž .consistent with a synergistic effect of BDNF and gp120 leading to enhanced functional expression of the signalling pathway s mediatingBDNF induction of SRIF production; an effect expressed by fetal brains throughout the 2nd trimester of gestation. Thus, this culture
Ž .system can serve as a model to study the mechanism s underlying the early interactions between gp120 BDNF in the developing humanbrain. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Peptidergic neuron; Neurotrophin; SRIF; Neuropeptide Y; BDNF; gp120; Brain
1. Introduction
AIDS patients suffer from a wide range of CNS-relatedŽsymptoms e.g., anxiety, drowsiness, subcortical cognitive
.changes, loss of memory and concentration , which canprogress to severe cognitive impairments, AIDS dementia
Ž w x.complex for reviews see Refs. 21,33 . Moreover, a lossŽ .of brain cells cortical, hippocampal to various degrees
has been observed in brains of adult and pediatric AIDSw xpatients 13,18,20 . However, the mechanisms underlying
these neurological changes are poorly understood. Recentstudies have implicated SRIF neurons in HIV encephalopa-thy. In the cortex, an increase in the number of neurons
) Corresponding author. Fax: q1-214-648-2674; E-mail:[email protected]
expressing preproSRIF-mRNA was noted in autopsy ofw xchildren with HIV encephalitis 14 and in an early stage
w xof SIV infection in the monkey 15 . A decrease in thenumber of SRIF neurons and the amounts of immunoreac-tive SRIF was noted in autopsy of adult AIDS cases with
w xencephalitis 19 . Thus, the question arises: Are thesedifferences in the fate of SRIF neurons age-dependent ordoes an increase in the expression of SRIF precede ademise of SRIF neurons in the HIV infected brain?
The current view is that secretory products ofmacrophagesrmicroglia, particularly gp120, are the majorcauses of the AIDS-associated abnormalities in the brainŽ w x.reviewed in Ref. 30 . Consistent with this view is thefinding of apoptosis in HIV infected human fetal braincultures 1–2 weeks after the time of peak virus productionw x45 . However, the issue of whether gp120 is the primary
0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 98 01098-1

( )A. Barnea et al.rBrain Research 815 1999 349–357350
inducer of neuronal death in the human brain is stillcontroversial. One group reported that gp120 by itself didnot induce neuronal death in cultured human fetal brain
w xcells 42,43,51 , whereas another group reported that it didw x29 . It should be noted that the age of the donor fetuses
Žvaried between these two studies 15–18 weeks and 8–10.weeks, respectively , raising the possibility that suscepti-
bility to gp120 is fetal-age dependent. To begin to under-stand the role of gp120 in the human fetal brain, weaddressed the question: Does gp120 alter the functionalexpression of human fetal SRIF neurons in culture and ifso, is this effect fetal-age dependent? Utilizing an aggre-gate culture system derived from human fetal cortical
w xtissue 1 , we assessed the effects of gp120 on SRIFproduction under two functional states: basal culture-con-dition and activated by BDNF.
2. Material and methods
2.1. Materials
Ž .DNase I was from Boehringer Indianapolis, IN , cul-Ž .ture reagents from Gibco Grand Island, NY , ITSq
Žserum-substitute from Collaborative Research Bedford,. 125 125 ŽMA , I-NPY and I-SRIF were from NEN Boston,. 3 Ž .MA , methyl H thymidine 48 Cirmmol from Amer-
Ž .sham Arlington Heights, IL , synthetic NPY and SRIFŽ .from Bachem Torrance, CA , and other reagents were
from Sigma. Recombinant HIV-1 gp120 was kindlySF2Ž .provided by Chiron Emeryville, CA and human recombi-
Žnant BDNF was kindly provided by Amgen Thousand. w xOaks, CA . The SRIF-antiserum 23 and the NPY-anti-
w xserum 9 were raised in our laboratories.
2.2. Cell culture
Human fetal brains, obtained from elective abortions,were provided by Advanced Bioscience ResourcesŽ .Alameda, CA . Fetal age ranged from 14–21 menstrual
Žweeks and only cortical tissue was used. All DMEM cat..no. 12430 solutions contained 5 mlrml antibioticranti-
mycotic and 40 mM glucose. Dissociation of the tissue andw xcell culture were carried out as previously described 1
Žwith slight modifications. Diced tissue f1r2 cortex of.18 weeks fetus was incubated at 37 C for 20 min in
Ž .CarMg-free Hanks solution CMFH and 20 min in 6 mlŽ .trypsin 0.0625% in CMFH . Then, 50 ml DMEM contain-
ing 20 mgrml DNase and 10% fetal calf serum wereadded, cells were gently dissociated using a plastic pipette,56 ml DMEM was added and cells were pelleted. Cells
Žwere resuspended in sITS solution DMEM containing 1%y11 y9 .ITSq , 10 M triiodothyronine and 10 M cortisol .
ŽThe viability of the cells assessed by trypan blue exclu-. 6sion was 90–95% and 5=10 cells were distributed into
Ž .25 ml Erlenmeyer flasks final culture volume 3.5 ml .
Flasks were gassed with 90% airr10% CO , capped tightly2Žand then incubated at 37 C under constant rotation 77
.rpm . Two ml medium were replaced after 5 days andthereafter at 2–3-day intervals. sITS was used as thediluent for gp120 and BDNF.
2.3. Generation of astrocyte-deficient, neuronal enrichedaggregates
w xAdopting our procedure for rat aggregate cultures 4 , 2ml culture medium was replaced on day 11 with sITS
3 Ž .containing H thymidine 5 mCirml in the culture ; subse-Ž . 3quent media changes day 13, 15, 18 did not contain H
thymidine. Immunocytochemical analysis for expression ofŽ .glial fibrillary acidic protein GFAP was carried out as
w x Ždescribed in detail 5 , using anti-GFAP Sigma, cat. no.. ŽG3893, 1:30,000 and Vector ABC elite kit Vector,. 3Burlingame, CA . Exposure to H thymidine led to a
drastic reduction in the number of cells expressing GFAPŽ . 3Fig. 1 . Accordingly, the DNA content of H-treatedaggregates was reduced to 76.0"5.1% of control-treated
Ž .aggregates mean"S.E., Ns6 separate experiments . Incontrast to DNA, SRIF content of the 3H-treated cultureswas not reduced and it was 97.7"6.2% of control aggre-gates. Thus, exposure to 3H thymidine generatedastrocyte-deficient, neuronal enriched aggregates and thefunctional expression of SRIF by these aggregates does notrequire input from astrocytes.
2.4. Experimental protocols: exposures to BDNFrgp120and assay of peptide production
Unless otherwise stated, the following pre-treatmentŽ .protocol was employed 4 groups . On day 11, the 2 ml
media change of group 1 contained gp120 to give a finalconcentration in the culture medium of 20 ngrml; on days13 and 15 the medium change contained 20 ngrml gp120;in a similar fashion, the media changes of group 2 con-
Ž .tained BDNF 40 ngrml ; the media changes of group 3Ž . Ž .contained BDNF 40 ngrml qgp120 20 ngrml ; and of
group 4 it was sITS alone. For assay of SRIF productionŽ .test , on day 18, the entire medium was replaced with
Ž . Ž .BDNF 40 ngrml, group 2 and 3 or sITS group 1 and 4and 24 h later, media and aggregates were collected forassays. Each treatment group consisted of quadruplicates.
2.5. Assays
w xSRIF and NPY 10 were extracted from the media andaggregates and peptides were quantified by RIA. Bindingof 125I SRIF to our SRIF antiserum is displaced equally
Ž .well by SRIF and SRIF-28 but not by SRIF-28 1–12 .SRIF was assayed using 100 ml sample, synthetic SRIF as
125 w xthe standard and I SRIF as the tracer 6 . NPY wasassayed using synthetic hNPY as the standard and 125I
w xpNPY as the tracer 9 . DNA content of the aggregates was
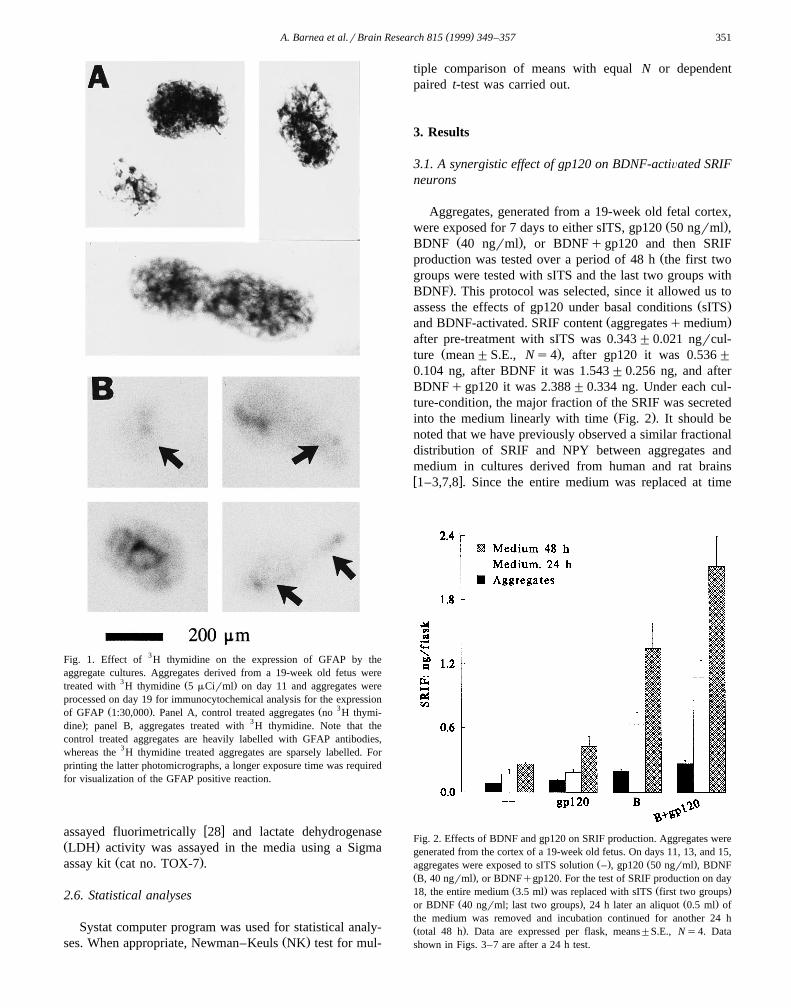
( )A. Barnea et al.rBrain Research 815 1999 349–357 351
Fig. 1. Effect of 3H thymidine on the expression of GFAP by theaggregate cultures. Aggregates derived from a 19-week old fetus were
3 Ž .treated with H thymidine 5 mCirml on day 11 and aggregates wereprocessed on day 19 for immunocytochemical analysis for the expression
Ž . Ž 3of GFAP 1:30,000 . Panel A, control treated aggregates no H thymi-. 3dine ; panel B, aggregates treated with H thymidine. Note that the
control treated aggregates are heavily labelled with GFAP antibodies,whereas the 3H thymidine treated aggregates are sparsely labelled. Forprinting the latter photomicrographs, a longer exposure time was requiredfor visualization of the GFAP positive reaction.
w xassayed fluorimetrically 28 and lactate dehydrogenaseŽ .LDH activity was assayed in the media using a Sigma
Ž .assay kit cat no. TOX-7 .
2.6. Statistical analyses
Systat computer program was used for statistical analy-Ž .ses. When appropriate, Newman–Keuls NK test for mul-
tiple comparison of means with equal N or dependentpaired t-test was carried out.
3. Results
3.1. A synergistic effect of gp120 on BDNF-actiÕated SRIFneurons
Aggregates, generated from a 19-week old fetal cortex,Ž .were exposed for 7 days to either sITS, gp120 50 ngrml ,
Ž .BDNF 40 ngrml , or BDNFqgp120 and then SRIFŽproduction was tested over a period of 48 h the first two
groups were tested with sITS and the last two groups with.BDNF . This protocol was selected, since it allowed us to
Ž .assess the effects of gp120 under basal conditions sITSŽ .and BDNF-activated. SRIF content aggregatesqmedium
after pre-treatment with sITS was 0.343"0.021 ngrcul-Ž .ture mean"S.E., Ns4 , after gp120 it was 0.536"
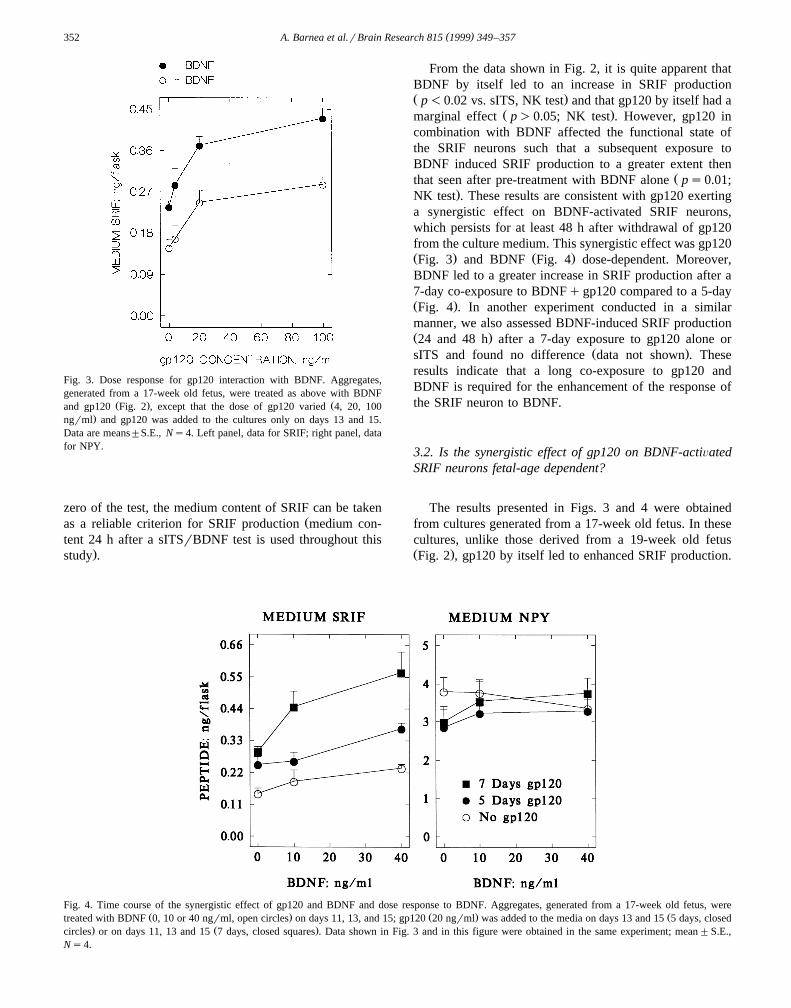
0.104 ng, after BDNF it was 1.543"0.256 ng, and afterBDNFqgp120 it was 2.388"0.334 ng. Under each cul-ture-condition, the major fraction of the SRIF was secreted
Ž .into the medium linearly with time Fig. 2 . It should benoted that we have previously observed a similar fractionaldistribution of SRIF and NPY between aggregates andmedium in cultures derived from human and rat brainsw x1–3,7,8 . Since the entire medium was replaced at time
Fig. 2. Effects of BDNF and gp120 on SRIF production. Aggregates weregenerated from the cortex of a 19-week old fetus. On days 11, 13, and 15,
Ž . Ž .aggregates were exposed to sITS solution – , gp120 50 ngrml , BDNFŽ .B, 40 ngrml , or BDNFqgp120. For the test of SRIF production on day
Ž . Ž .18, the entire medium 3.5 ml was replaced with sITS first two groupsŽ . Ž .or BDNF 40 ngrml; last two groups , 24 h later an aliquot 0.5 ml of
the medium was removed and incubation continued for another 24 hŽ .total 48 h . Data are expressed per flask, means"S.E., Ns4. Datashown in Figs. 3–7 are after a 24 h test.
( )A. Barnea et al.rBrain Research 815 1999 349–357352
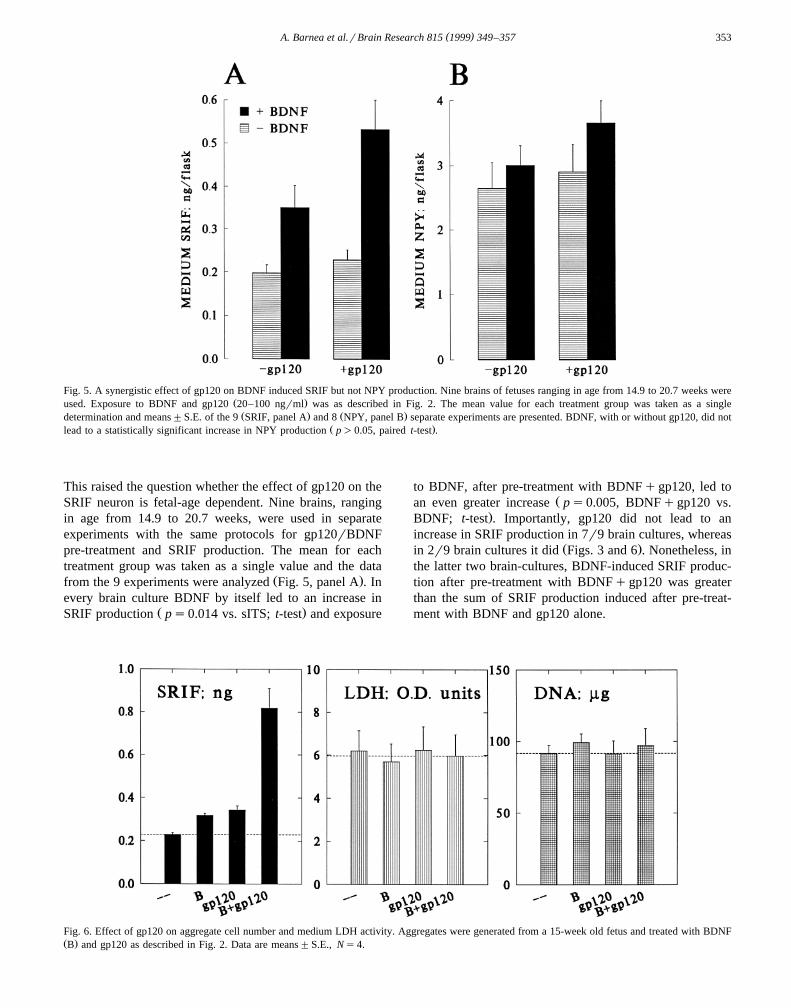
Fig. 3. Dose response for gp120 interaction with BDNF. Aggregates,generated from a 17-week old fetus, were treated as above with BDNF
Ž . Žand gp120 Fig. 2 , except that the dose of gp120 varied 4, 20, 100.ngrml and gp120 was added to the cultures only on days 13 and 15.
Data are means"S.E., Ns4. Left panel, data for SRIF; right panel, datafor NPY.
zero of the test, the medium content of SRIF can be takenŽas a reliable criterion for SRIF production medium con-
tent 24 h after a sITSrBDNF test is used throughout this.study .
From the data shown in Fig. 2, it is quite apparent thatBDNF by itself led to an increase in SRIF productionŽ .p-0.02 vs. sITS, NK test and that gp120 by itself had a
Ž .marginal effect p)0.05; NK test . However, gp120 incombination with BDNF affected the functional state ofthe SRIF neurons such that a subsequent exposure toBDNF induced SRIF production to a greater extent then
Žthat seen after pre-treatment with BDNF alone ps0.01;.NK test . These results are consistent with gp120 exerting
a synergistic effect on BDNF-activated SRIF neurons,which persists for at least 48 h after withdrawal of gp120from the culture medium. This synergistic effect was gp120Ž . Ž .Fig. 3 and BDNF Fig. 4 dose-dependent. Moreover,BDNF led to a greater increase in SRIF production after a7-day co-exposure to BDNFqgp120 compared to a 5-dayŽ .Fig. 4 . In another experiment conducted in a similarmanner, we also assessed BDNF-induced SRIF productionŽ .24 and 48 h after a 7-day exposure to gp120 alone or
Ž .sITS and found no difference data not shown . Theseresults indicate that a long co-exposure to gp120 andBDNF is required for the enhancement of the response ofthe SRIF neuron to BDNF.
3.2. Is the synergistic effect of gp120 on BDNF-actiÕatedSRIF neurons fetal-age dependent?
The results presented in Figs. 3 and 4 were obtainedfrom cultures generated from a 17-week old fetus. In thesecultures, unlike those derived from a 19-week old fetusŽ .Fig. 2 , gp120 by itself led to enhanced SRIF production.
Fig. 4. Time course of the synergistic effect of gp120 and BDNF and dose response to BDNF. Aggregates, generated from a 17-week old fetus, wereŽ . Ž . Žtreated with BDNF 0, 10 or 40 ngrml, open circles on days 11, 13, and 15; gp120 20 ngrml was added to the media on days 13 and 15 5 days, closed
. Ž .circles or on days 11, 13 and 15 7 days, closed squares . Data shown in Fig. 3 and in this figure were obtained in the same experiment; mean"S.E.,Ns4.
( )A. Barnea et al.rBrain Research 815 1999 349–357 353
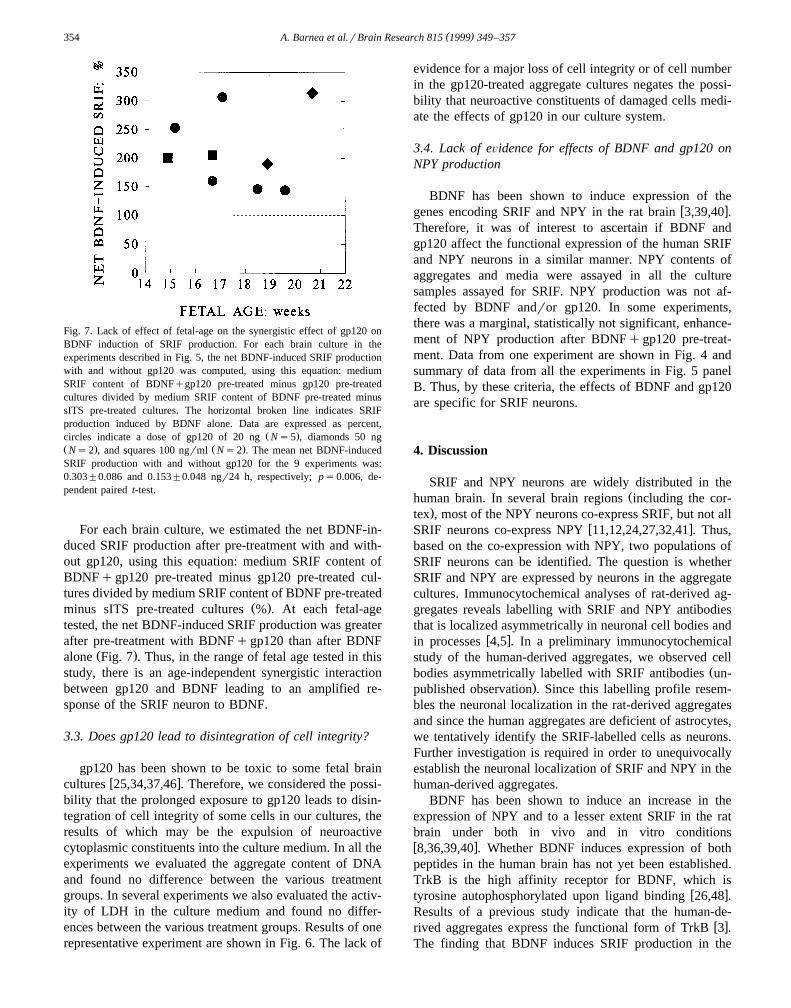
Fig. 5. A synergistic effect of gp120 on BDNF induced SRIF but not NPY production. Nine brains of fetuses ranging in age from 14.9 to 20.7 weeks wereŽ .used. Exposure to BDNF and gp120 20–100 ngrml was as described in Fig. 2. The mean value for each treatment group was taken as a singleŽ . Ž .determination and means"S.E. of the 9 SRIF, panel A and 8 NPY, panel B separate experiments are presented. BDNF, with or without gp120, did not
Ž .lead to a statistically significant increase in NPY production p)0.05, paired t-test .
This raised the question whether the effect of gp120 on theSRIF neuron is fetal-age dependent. Nine brains, rangingin age from 14.9 to 20.7 weeks, were used in separateexperiments with the same protocols for gp120rBDNFpre-treatment and SRIF production. The mean for eachtreatment group was taken as a single value and the data
Ž .from the 9 experiments were analyzed Fig. 5, panel A . Inevery brain culture BDNF by itself led to an increase in
Ž .SRIF production ps0.014 vs. sITS; t-test and exposure
to BDNF, after pre-treatment with BDNFqgp120, led toŽan even greater increase ps0.005, BDNFqgp120 vs.
.BDNF; t-test . Importantly, gp120 did not lead to anincrease in SRIF production in 7r9 brain cultures, whereas
Ž .in 2r9 brain cultures it did Figs. 3 and 6 . Nonetheless, inthe latter two brain-cultures, BDNF-induced SRIF produc-tion after pre-treatment with BDNFqgp120 was greaterthan the sum of SRIF production induced after pre-treat-ment with BDNF and gp120 alone.
Fig. 6. Effect of gp120 on aggregate cell number and medium LDH activity. Aggregates were generated from a 15-week old fetus and treated with BDNFŽ .B and gp120 as described in Fig. 2. Data are means"S.E., Ns4.
( )A. Barnea et al.rBrain Research 815 1999 349–357354
Fig. 7. Lack of effect of fetal-age on the synergistic effect of gp120 onBDNF induction of SRIF production. For each brain culture in theexperiments described in Fig. 5, the net BDNF-induced SRIF productionwith and without gp120 was computed, using this equation: mediumSRIF content of BDNFqgp120 pre-treated minus gp120 pre-treatedcultures divided by medium SRIF content of BDNF pre-treated minussITS pre-treated cultures. The horizontal broken line indicates SRIFproduction induced by BDNF alone. Data are expressed as percent,
Ž .circles indicate a dose of gp120 of 20 ng Ns5 , diamonds 50 ngŽ . Ž .Ns2 , and squares 100 ngrml Ns2 . The mean net BDNF-inducedSRIF production with and without gp120 for the 9 experiments was:0.303"0.086 and 0.153"0.048 ngr24 h, respectively; ps0.006, de-pendent paired t-test.
For each brain culture, we estimated the net BDNF-in-duced SRIF production after pre-treatment with and with-out gp120, using this equation: medium SRIF content ofBDNFqgp120 pre-treated minus gp120 pre-treated cul-tures divided by medium SRIF content of BDNF pre-treated
Ž .minus sITS pre-treated cultures % . At each fetal-agetested, the net BDNF-induced SRIF production was greaterafter pre-treatment with BDNFqgp120 than after BDNF
Ž .alone Fig. 7 . Thus, in the range of fetal age tested in thisstudy, there is an age-independent synergistic interactionbetween gp120 and BDNF leading to an amplified re-sponse of the SRIF neuron to BDNF.
3.3. Does gp120 lead to disintegration of cell integrity?
gp120 has been shown to be toxic to some fetal brainw xcultures 25,34,37,46 . Therefore, we considered the possi-
bility that the prolonged exposure to gp120 leads to disin-tegration of cell integrity of some cells in our cultures, theresults of which may be the expulsion of neuroactivecytoplasmic constituents into the culture medium. In all theexperiments we evaluated the aggregate content of DNAand found no difference between the various treatmentgroups. In several experiments we also evaluated the activ-ity of LDH in the culture medium and found no differ-ences between the various treatment groups. Results of onerepresentative experiment are shown in Fig. 6. The lack of
evidence for a major loss of cell integrity or of cell numberin the gp120-treated aggregate cultures negates the possi-bility that neuroactive constituents of damaged cells medi-ate the effects of gp120 in our culture system.
3.4. Lack of eÕidence for effects of BDNF and gp120 onNPY production
BDNF has been shown to induce expression of thew xgenes encoding SRIF and NPY in the rat brain 3,39,40 .
Therefore, it was of interest to ascertain if BDNF andgp120 affect the functional expression of the human SRIFand NPY neurons in a similar manner. NPY contents ofaggregates and media were assayed in all the culturesamples assayed for SRIF. NPY production was not af-fected by BDNF andror gp120. In some experiments,there was a marginal, statistically not significant, enhance-ment of NPY production after BDNFqgp120 pre-treat-ment. Data from one experiment are shown in Fig. 4 andsummary of data from all the experiments in Fig. 5 panelB. Thus, by these criteria, the effects of BDNF and gp120are specific for SRIF neurons.
4. Discussion
SRIF and NPY neurons are widely distributed in theŽhuman brain. In several brain regions including the cor-
.tex , most of the NPY neurons co-express SRIF, but not allw xSRIF neurons co-express NPY 11,12,24,27,32,41 . Thus,
based on the co-expression with NPY, two populations ofSRIF neurons can be identified. The question is whetherSRIF and NPY are expressed by neurons in the aggregatecultures. Immunocytochemical analyses of rat-derived ag-gregates reveals labelling with SRIF and NPY antibodiesthat is localized asymmetrically in neuronal cell bodies and
w xin processes 4,5 . In a preliminary immunocytochemicalstudy of the human-derived aggregates, we observed cell
Žbodies asymmetrically labelled with SRIF antibodies un-.published observation . Since this labelling profile resem-
bles the neuronal localization in the rat-derived aggregatesand since the human aggregates are deficient of astrocytes,we tentatively identify the SRIF-labelled cells as neurons.Further investigation is required in order to unequivocallyestablish the neuronal localization of SRIF and NPY in thehuman-derived aggregates.
BDNF has been shown to induce an increase in theexpression of NPY and to a lesser extent SRIF in the ratbrain under both in vivo and in vitro conditionsw x8,36,39,40 . Whether BDNF induces expression of bothpeptides in the human brain has not yet been established.TrkB is the high affinity receptor for BDNF, which is
w xtyrosine autophosphorylated upon ligand binding 26,48 .Results of a previous study indicate that the human-de-
w xrived aggregates express the functional form of TrkB 3 .The finding that BDNF induces SRIF production in the

( )A. Barnea et al.rBrain Research 815 1999 349–357 355
aggregates in a dose-dependent manner implicates thetyrosine kinase signalling pathway in mediating BDNFaction on the SRIF neuron. An interesting question iswhether the effects of BDNF are manifested in the popula-tion of SRIF neurons co-expressing NPY. The lack ofeffect of BDNF by itself or with gp120 on NPY produc-
Ž w x.tion this study and Ref. 3 is suggestive that the effectsof BDNF seen in this study manifest in SRIF neurons thatdo not co-express NPY. If on the other hand, the SRIFneurons do co-express NPY, these results would suggestdifferences in the signalling pathways mediating BDNFinduction of SRIF and NPY production in the humanbrain.
There are two schools of thought regarding the neuro-toxicity of gp120, based on results obtained from studiesutilizing rat- and human-derived brain cultures. One view
wis that gp120 by itself induces neuronal death 25,29,x Ž .34,37,50 . The other view is that gp120 requires a partner s
w xfor inducing neuronal death, e.g., glutamate 16,31 , solu-w xble products of HIV-infected macrophages 42,43,51 . In
our culture system, gp120 does not lead to a significantdecrease in the aggregate DNA content nor to an increase
Ž .in LDH activity in the medium Fig. 6 . Hence, a majorloss of cells andror cell integrity is not evident. Likewise,gp120 does not alter DNA content and LDH activity in
Žaggregate cultures derived from the rat fetal cortex unpub-.lished observation . It should be emphasized that gp120
leads to a massive loss of cells in the rat brain cultures, inw xthe range of 60–90% of the total cells 25,34,37,46 . On
the other hand, HIV infection of human brain culturesleads to apoptosis of a small population of cells, about
w x10% of the total 45 . Our quantitative analyses for DNAand LDH cannot discriminate a change of 10% or less.Therefore, one cannot exclude the possibility that gp120leads to apoptosis of a select small population of cellswhich does not include the SRIF neuron.
The novel finding in this study is that gp120 actssynergistically with BDNF to enhance the functional stateof the SRIF neuron, although gp120 by itself does not doso. Two lines of evidence support the proposition that thesynergistic effect is manifested at the level of the tyrosinekinase signalling pathway mediating BDNF induction ofSRIF production. One, BDNF-induced SRIF production issignificantly greater after co-exposure to BDNFqgp120compared to BDNF alone. Two, this enhanced inductionby BDNF persists for at least 48 h after withdrawal ofgp120 from the culture medium. Importantly, the synergis-tic effect of gp120 and BDNF is expressed in every brainculture derived from fetuses ranging in gestational agefrom 14.9 to 20.7 weeks. The critical question is therelevance of these findings to the AIDS-associated changesin the brains of HIV infected people. The inhibitory effect
Ž .of SRIF on growth hormone GH secretion from thepituitary is well established. SRIF has also been implicatedin a wide range of brain functions, acting primarily as an
Ž w x.inhibitory agent for recent reviews, see Refs. 44,49 . It is
now becoming apparent that SRIF neurons are affected byHIVrgp120. An increase in the number of SRIF express-ing neurons has been observed in the brains of children
w xwith HIV encephalitis 14 and of SIV infected monkeysw x15 . A decrease in plasma GH has been observed in the ratafter in vivo administration of gp120, which may be due,
w xin part, to elevated levels of SRIF 38 . The question ariseswhether the in vivo elevation in SRIF expression is broughtabout by BDNF?
BDNF belongs to the neurotrophin family of peptides,which includes NGF, NT-3, and NT-4r5. Recent studiesdemonstrate that activated macrophagesrmicroglia expressone or more of these neurotrophins. For example, mi-
w xcroglia express NT-3 in vivo 17 and BDNF, NT-3, andw xNGF in vitro 17,22,35 . Importantly, expression of BDNF
in activated microglia of HIV-1 encephalitis has recentlyw xbeen demonstrated 47 . Thus, it is tempting to speculate
that activated microglia in the brains of HIV infectedpeople secrete BDNF, which, in turn, interacts synergisti-cally with gp120 to enhance the functional expression ofthe SRIF neuron. We propose that our aggregate culturesystem can serve as a model to study the mechanismsunderlying the early interactions between BDNF and gp120in the developing human brain.
Acknowledgements
The authors thank Chiron for the generous supply ofrecombinant gp120 and Amgen for the generous supply ofrecombinant human BDNF. The technical assistance ofMichael Slaughter and Abdul Qayyum is highly appreci-ated. This study was supported by NIH Research Grant
Ž .NS32207 A.B. .
References
w x1 N. Aguila-Mansilla, A. Barnea, Human fetal brain cells in aggregateculture: a model system to study regulatory processes of the devel-
Ž .oping human neuropeptide Y NPY producing neuron, Int. J. Dev.Ž .Neurosci. 14 1996 531–539.
w x2 N. Aguila-Mansilla, B.B. Little, R.H. Ho, A. Barnea, Differentialpotencies of cocaine and its metabolites, cocaethylene and ben-zoylecgonine, in suppressing the functional expression of somato-statin and neuropeptide Y producing neurons in cultures of fetal
Ž .cortical cells, Biochem. Pharmacol. 54 1997 491–500.w x3 A. Barnea, N. Aguila-Mansilla, H.T. Chute, A.A. Welcher, Compar-
ison of neurotrophin regulation of human and rat neuropeptide YŽ .NPY neurons: induction of NPY production in aggregate culturesderived from rat but not from human fetal brains, Brain Res. 732Ž .1996 52–60.
w x4 A. Barnea, N. Aguila-Mansilla, G. Lu, R.H. Ho, Opposite effects ofŽ .astrocyte-derived soluble factor s on the functional expression of
fetal peptidergic neurons in aggregate cultures: enhancement ofneuropeptide Y and suppression of somatostatin, J. Neurosci. Res.
Ž .50 1997 605–617.w x5 A. Barnea, E. Anthony, G. Lu, G. Cho, Morphological differentia-
tion of neuropeptide Y neurons in aggregate cultures of dissociated

( )A. Barnea et al.rBrain Research 815 1999 349–357356
fetal cortical cells: a model system for glia–neuron paracrine interac-Ž .tions, Brain Res. 625 1993 313–322.
w x6 A. Barnea, G. Cho, A. Hajibeigi, M.C. Aguila, P. Magni, Dexam-ethasone induced accumulation of neuropeptide Y by aggregatingfetal brain cells in culture: a process dependent on the developmen-
Ž .tal-age of the aggregates, Endocrinology 129 1991 931–938.w x7 A. Barnea, G. Cho, G. Lu, Role for glial cells in regulating the
Ž .functional expression of neuropeptide Y NPY neurons in aggregateŽ .cultures derived from fetal brain cells, J. Neurosci. Res. 38 1994
459–467.w x8 A. Barnea, G. Cho, G. Lu, M. Mathis, Brain-derived neurotrophic
factor induces functional expression and phenotypic differentiationof cultured fetal neuropeptide Y producing neurons, J. Neurosci.
Ž .Res. 42 1995 638–647.w x9 A. Barnea, O. Cramer, G. Cho, Truncated insulin-like growth factor-
I: A potent inducer of neuropeptide Y production by aggregateŽ .cultures of fetal brain cells, Endocrine J. 1 1993 11–17.
w x10 A. Barnea, A. Hajibeigi, G. Cho, P. Magni, Regulated productionand secretion of immunoreactive neuropeptide Y by aggregating
Ž .fetal brain cells in culture, Neuroendocrinology 54 1991 7–13.w x11 C. Bouras, P.J. Magistretti, J.H. Morrison, J. Constantinidis, An
immunohistochemical study of pro-somatostatin-derived peptides inŽ .the human brain, Neuroscience 22 1987 781–800.
w x12 V. Chan Palay, Somatostatin immunoreactive neurons in the humanhippocampus and cortex shown by immunogoldrsilver intensifica-tion on vibratome sections: coexistence with neuropeptide Y neu-rons, and effects in Alzheimer-type dementia, J. Comp. Neurol. 260Ž .1987 201–223.
w x13 L.A. Civitello, Neurologic complications of HIV infection in chil-Ž .dren, Pediatr. Neurosurg. 17 1991 104–112.
w x14 A. Da Cunha, M. Mintz, L.E. Eiden, L.R. Sharer, A neuronal andneuroanatomical correlate of HIV-1 encephalopathy relative to HIV-1encephalitis in HIV-1-infected children, J. Neuropathol. Exp. Neu-
Ž .rol. 56 1997 974–987.w x15 A. Da Cunha, D.M. Rausch, L.E. Eiden, An early increase in
somatostatin mRNA expression in the frontal cortex of rhesusmonkeys infected with simian immunodeficiency virus, Proc. Natl.
Ž .Acad. Sci. USA 92 1995 1371–1375.w x16 V.L. Dawson, T.M. Dawson, G.R. Uhl, S.H. Snyder, Human im-
munodeficiency virus type 1 coat protein neurotoxicity mediated bynitric oxide in primary cortical cultures, Proc. Natl. Acad. Sci. USA
Ž .90 1993 3256–3259.w x17 S. Elkabes, E.M. DiCicco-Bloom, I.B. Black, Brain microgliarmac-
rophages express neurotrophins that selectively regulate microglialŽ .proliferation and function, J. Neurosci. 16 1996 2508–2521.
w x18 I.P. Everall, P.J. Luthert, P.L. Lantos, Neuronal loss in the frontalw x Ž .cortex in HIV infection see comments , Lancet 337 1991 1119–
1121.w x19 L. Fox, M. Alford, C. Achim, M. Mallory, E. Masliah, Neurodegen-
eration of somatostatin-immunoreactive neurons in HIV encephalitis,Ž .J. Neuropathol. Exp. Neurol. 56 1997 360–368.
w x20 H.A. Gelbard, H.J. James, L.R. Sharer, S.W. Perry, Y. Saito, A.M.Kazee, B.M. Blumberg, L.G. Epstein, Apoptotic neurons in brainsfrom paediatric patients with HIV-1 encephalitis and progressiveencephalopathy, Neuropathology and Applied Neurobiology 21Ž .1995 208–217.
w x21 I. Grant, J.H. Atkinson, The evolution of neurobehavioral complica-Ž .tions of HIV infection, Psychol. Med. 20 1990 747–754.
w x22 K. Heese, C. Hock, U. Otten, Inflammatory signals induce neu-rotrophin expression in human microglial cells, J. Neurochem. 70Ž .1998 699–707.
w x23 R.H. Ho, L.R. DePalatis, Substance P immunoreactivity in themedian eminence of the North American opossum and domestic
Ž .fowl, Brain Res. 189 1980 565–569.w x24 J.P. Hornung, N. de Tribolet, I. Tork, Morphology and distribution
of neuropeptide-containing neurons in human cerebral cortex, Neu-Ž .roscience 51 1992 363–375.
w x25 P.K. Kaiser, J.T. Offermann, S.A. Lipton, Neuronal injury due toHIV-1 envelope protein is blocked by anti-gp120 antibodies but not
Ž .by anti-CD4 antibodies, Neurology 40 1990 1757–1761.w x26 R. Klein, V. Nanduri, S.A. Jing, F. Lamballe, P. Tapley, S. Bryant,
C. Cordon-Cardo, K.R. Jones, L.F. Reichardt, M. Barbacid, ThetrkB tyrosine protein kinase is a receptor for brain-derived neu-
Ž .rotrophic factor and neurotrophin-3, Cell 66 1991 395–403.w x27 N.W. Kowall, R.J. Ferrante, M.F. Beal, E.P. Richardson, M.V.
Sofroniew, A.C. Cuello, J.B. Martin, Neuropeptide Y, somatostatin,and reduced nicotinamide XX XX adenine dinucleotide phosphatediaphorase in the human XX XX striatum: a combined immunocyto-chemical and enzyme histochemical XX XX study, Neuroscience 20Ž .1987 817–828.
w x28 C. Labarca, K. Paigen, A simple, rapid, and sensitive DNA assayŽ .procedure, Ann. Biochem. 102 1980 344–352.
w x29 A. Lannuzel, J.V. Barnier, C. Hery, H.V. Tan, B. Guilbert, F. Gray,J.D. Vincent, M. Tardieu, Human immunodeficiency virus type 1and its coat protein gp120 induce apoptosis and activate JNK andERK mitogen-activated proteins kinases in human neurons, Ann.
Ž .Neurol. 42 1997 847–856.w x30 J.A. Levy, Pathogenesis of human immunodeficiency virus infec-
Ž .tion, Microbiol. Rev. 57 1993 183–289.w x31 S.A. Lipton, N.J. Sucher, P.K. Kaiser, E.B. Dreyer, Synergistic
effects of HIV coat protein and NMDA receptor-mediated neurotoxi-Ž .city, Neuron 7 1991 111–118.
w x32 G.W. Mathern, T.L. Babb, J.K. Pretorius, J.P. Leite, Reactivesynaptogenesis and neuron densities for neuropeptide Y, somato-statin, and glutamate decarboxylase immunoreactivity in the epilep-
Ž .togenic human fascia dentata, J. Neurosci. 15 1995 3990–4004.w x33 J.E. Merril, I.Y. Chen, HIV-1, macrophages, glial cells, and cy-
Ž .tokines in AIDS nervous system disease, FASEB J. 5 1991 2391–2397.
w x34 O. Meucci, R.J. Miller, gp120-Induced neurotoxicity in hippocampalpyramidal neuron cultures: protective action of TGF-b1, J. Neurosci.
Ž .16 1996 4080–4088.w x35 T. Miwa, S. Furukawa, K. Nakajima, Y. Furukawa, S. Kohsaka,
Lipopolysaccharide enhances synthesis of brain-derived neu-Ž .rotrophic factor in cultured rat microglia, J. Neurosci. Res. 50 1997
1023–1029.w x36 K. Mizuno, J. Carnahan, H. Nawa, Brain-derived neurotrophic factor
promotes differentiation of striatal GABAergic neurons, Dev. Biol.Ž .165 1994 243–256.
w x37 W.E. Muller, H.C. Schroder, H. Ushijima, J. Dapper, J. Bormann,gp120 of HIV-1 induces apoptosis in rat cortical cell cultures:
Ž .prevention by memantine, Eur. J. Pharmacol. 226 1992 209–214.w x38 S.E. Mulroney, K.J. McDonnel, C.B. Pert, M.R. Ruff, Z. Resch,
W.K. Samson, M.D. Lumpkin, HIV gp120 inhibits the somatotropicaxis: a possible GH-releasing hormone receptor mechanisms for thepathogenesis of AIDS wasting, Proc. Natl. Acad. Sci. USA 95Ž .1998 1927–1932.
w x39 H. Nawa, Y. Bessho, J. Carnahan, S. Nakanishi, K. Mizuno, Regula-tion of neuropeptide expression in cultured cerebral cortical neurons
Ž .by brain-derived neurotrophic factor, J. Neurochem. 60 1993 772–775.
w x40 H. Nawa, M.A. Pelleymounter, J. Carnahan, Intraventricular admin-istration of BDNF increases neuropeptide expression in newborn rat
Ž .brain, J. Neurosci. 14 1994 3751–3765.w x41 W.Y. Ong, L.J. Garey, Distribution of GABA and neuropeptides in
the human cerebral cortex. A light and electron microscopic study,Ž .Anat. Embryol. 183 1991 397–413.
w x42 L. Pulliam, B.G. Herndier, N.M. Tang, M.S. McGrath, Humanimmunodeficiency virus-infected macrophages produce soluble fac-tors that cause histological and neurochemical alterations in cultured
Ž .human brains, J. Clin. Invest. 87 1991 503–512.w x43 L. Pulliam, D. West, N. Haigwood, R.A. Swanson, HIV-1 envelope
gp120 alters astrocytes in human brain cultures, Aids Res. HumanŽ .Retrovir. 9 1993 439–444.

( )A. Barnea et al.rBrain Research 815 1999 349–357 357
w x44 M. Schindler, P.P. Humphrey, P.C. Emson, Somatostatin receptorsŽ .in the central nervous system, Prog. Neurobiol. 50 1996 9–47,
w x w xReview 255 refs .w x45 B. Shi, U.D. Girolami, J. He, S. Wang, A. Lorenzo, J. Busciglio, D.
Gabuzda, Apoptosis induced by HIV-1 infection of the centralnervous system, J. Clin. Invest., Vol. 98, 1996, pp. 1979-1980.
w x46 P. Sindou, P. Couratier, F. Esclaire, C. Yardin, A. Bousseau, J.Ž .Hugon, Prevention of HIV coat protein gp120 toxicity in cortical
Ž .cell cultures by riluzole, J. Neurol. Sci. 126 1994 133–137.w x47 V. Soontornniyomkij, G. Wang, C.A. Pittman, C.A. Wiley, C.L.
Achim, Expression of brain-derived neurotrophic factor protein inactivated microglia of human immunodeficiency virus type 1 en-
Ž .cephalitis, Neuropathol. Appl. Neurobiol., In Press 1998 .w x48 S.P. Squinto, T.N. Stitt, T.H. Aldrich, S. Davis, S.M. Bianco, C.
Radziejewski, D.J. Glass, P. Masiakowski, M.E. Furth, D.M. Valen-zuela et al., trkB encodes a functional receptor for brain-derivedneurotrophic factor and neurotrophin-3 but not nerve growth factor,
Ž .Cell 65 1991 885–893.w x49 L. Vecsei, E. Widerlov, Brain and CSF somatostatin concentrations
in patients with psychiatric or neurological illness. An overview,Ž . w xActa Psychiatr. Scand. 78 1988 657–667, Review .
w x50 P. Wu, P. Price, B. Du, W.C. Hatch, E.F. Terwilliger, Directcytotoxicity of HIV-1 envelope protein gp120 on human NT neu-
Ž .rons, Neuroreport 7 1996 1045–1049.w x51 M.C. Yeung, L. Pulliam, A.S. Lau, The HIV envelope protein gp120
is toxic to human brain-cell cultures through induction of inter-Ž .leukin-6 and tumor necrosis factor-alpha, AIDS 9 1995 137–143.