Étude de l'inhibition de la protéolyse trypsique de la β-lactoglobuline a par le saccharose...
TRANSCRIPT

34 o BIOCHIMICA ET BIOPHYSICA AC]-A
BBA 35244
I~TUDE DE L ' I N H I B I T I O N DE LA PROTI~OLYSE TRYPSIQUE DE LA
fi-LACTOGLOBULINE A PAR LE SACCHAROSE ET QUELQUES AUTRES
SUCRES. MISE EN I~VIDENCE D'UN CHANGEMENT DE CONFORMATION
DE LA f i -LACTOGLOBULINE A PAR LIAISON AVEC LE SACCHAROSE
J E N N Y C L E M E N T - M E T R A L * , * " ~;T J E A N N I N E YON
Laboratoire de Nutrition Humaine de l'lnstitut National de la Santd et de la Recherche Mddicale, Hdpital Bichat, Paris, el Laboratoire de Biologie Phvsico-Chimique, Facultd des Sciences d'Orsay, 9I-Orsay (France)
(Regu le 20 mars, 1968)
SUMMARY
The inhibition of tryptic proteolysis of ~-lactoglobulin A by sucrose and some other sugars. Evidence of a conformational change of fi-lactoglobuline A by linkage with sucrose
We were able to show that the inhibition caused by sucrose on tryptic (EC 3.4.4.4) hydrolysis of fl-lactoglobulin A is due to the fixation of sucrose on this protein and to the existence of strong stabilizing interactions.
The kinetic analysis, based on the steady-state approximation assuming a two- step scheme, shows that the total non-competitive inhibition occurs only with native protein substrate. Two molecules of sucrose are involved in the process, and the binding sites on the 2/2 molecule are not identical nor independent. As the presence of sucrose modifies only that part of the/3-1actoglobulin molecule that is active (total inhibition), we may say that changes in reaction rate are in inverse proportion to the saturation of the substrate with the inhibitor. Therefore we plot (v -- vd/v, in place of log Y / ( I - - Y ) , (v,, v = reaction rate with or without inhibition, Y = "fractional satu- ration" of the substrate with the inhibitor, i.e. concentration of inhibitor bound per mole and per site of substrate). The n parameter of the so-called Hill plot, in terms of log (v -- vi)/v~ versus log [I[] (I = concentration of inhibitor), is found to be 2.7. There- fore there must be some inter-molecular interactions associated with strong intra- molecular stabilizing interactions. The fl-lactoglobulin A dimer is known to dissociate between pH 6 and pH 9 into two monomers (molecular weight 17 500). The calculation for the dimer dissociation linked to the binding of two moles of sucrose per mole of protein gives the same 2. 7 value for n, assuming a wide separation of the individual
Abr6v ia t ion : TSAME, p-toluenesulfonyl-L-arginine m6thy le s t e r ch lo rhydra te . * Adresse ac tue l le : Labora to i r e de P h o t o s y n t h b s e du C.N.R.S., 9 t -G i f - su r -Yve t t e France.
*" Ce t r a v a i l fa i t pa r t i e d ' une Th6se de I )oc to ra t d 'Etat-4"s-Sciences Phys iques sou tenue le 28 ju in 1966 d e v a n t la Facu l t~ des Sciences d 'O r s a y 91, France, enregis t r6e au C.N.R.S. sous le No. A 1195.
Biochim. Biophys. Acta, 16o (1968) 34o-359

/~-LACTOGLOBULINE A ET SACCHAROSE 341
equilibrium curves of the dimer and the monomer with the inhibitor. Empirical and theoretical results are thus in good agreement.
These kinetic results are confirmed by a thermodynamic study. The increase of the activation free energy for the 5o% -inhibited enzymatic reaction is calculated to be 4oo cal, whereas the experimentally determined minimal value of the total free energy of interaction, realised in completely saturating the macromolecule by the ligand, is 76o cal. The thermodynamic study indicates, on the other hand, that the forces involved are of the hydrophobic type and accompanied by a negative variation of entropy. This suggests the occurrence of a conformational change which is all the more plausible since interactions in a macromolecule often go with changes in the tert iary and quaternary structure. Several recent studies show further that changes in conformation of proteins results from reversible combination with non-polar substances.
The conformational change of/~-lactoglobulin A induced by sucrose is made apparent, in the absence of any enzymatic reaction, by titration and difference spectros- copy curves. I t is closely related to the inhibition of the proteolysis of/5-1actoglobulin A, since the value of the sucrose concentration which gives the half transformation is the same (o. 7 M) as for the 5O~o inhibition. The observed phenomenon is not due to dis- sociation of the macromolecule alone and corresponds to a more compact structure of the protein.
I t is noteworthy that sucrose, which is so largely employed in studies on the native structure of proteins, is not inert and causes a transconformation of/~-lacto- globulin A.
INTRODUCTION
L'inhibition de la prot6olyse trypsique de la fl-lactoglobuline par les selsl, 2 avait d~jA montr6 l'existence d'interactions de diverses natures s'exer~ant sur la prot6ine substrat. La fl-lactoglobuline dont la structure est particuli~rement bien 6tudi6e subit des changements de conformation sous diverses influences 3-7 et en particulier par fixation de moMcules non polairesS, 9. Enfin, le r61e des sucres dans la protection des prot6ines contre la d6naturation par la chaleur est connu depuis longtemps 1°-12.
Nous avons donc 6tudi6 : (I) l 'inhibition de la prot6olyse de la ~-lactoglobuline A par le saccharose et montr6 dans ce travail, en nous basant sur un sch6ma de rdaction
deux 6tapes que l'inhibition observ6e, du type non comp~titif total est li6e ~ la nature prot~ique du substrat. Le saccharose ne se lie pas ~t l 'enzyme mais au substrat, si celui-ci est une prot6ine et deux mol6cules de sucre se fixent par mol6cule de fl-lactoglobuline.
(2) L'~tude thermodynamique a montr6 que la valeur de l'6nergie libre minimum totale moyenne d'interaction sucre-lactoglobuline est le double de la variation d'~nergie libre correspondant ~ la demi-inhibition. L'inhibition serait don c enti~rement due la liaison saccharose lactoglobuline et/t l 'existence de fortes interactions st abilisatrices au sein de la fl-lactoglobuline.
(3) Ind@endamment de l '6tude enzymatique, l '6tude physico-chimique de la liaison sucre fl-lactoglobuline nous a permis de mettre en ~vidence un changement de conformation de la fl-lactoglobuline A. Les interactions sont ~ la fois inter- et intra- mol6culaire; l ' interaction inter-moldculaire correspond ~ une dissociation du dim~re (35 500).
Biochim. Biophys. Acta, 16o (1968) 340-359

3 4 2 j . ( L E M E N T METRAI. , j . V()X
METHODES
Les mesures d'hydrolyse enzymatique ont 6t6 faites par la m6thode potentio- m6trique continue ~ pH constant 13' ces mesures ainsi que les titrages sont effectu6es /t l'aide d 'un pH-m6tre E.I.L. type Vibron 33 B. Nous avons utilis6 la m6thode de titrage continu la,15, sous atmosph6re d'azote, avec addition de soude ou d'acide chlorhydrique 0.5 M (Merck).
La trypsine (Worthington) est dialys6e contre HC1 o.oi M jusqu'/~ 61imination compl6te du MgSO 4 pr4sent dans la pr6paration commerciale. L'a-chymotrypsine (Worthington) est utilis6e en solution dans HC1 O.Ol M. La fl-lactoglobuline A native a 6t6 pr6parde au laboratoire par la mfthode de PALMER 16 modifidelL Nous avons utilis6 comme fi-lactoglobuline A ddnatur6e par la chaleur la forme "f" de BRIGGS ET HULL is. Ces pr6parations ont 6t6 d6crites ant6rieurement en d6tail 2,19. Le TSAME (p-toluene- sulfonyl-L-arginine m6thylester chlorhydrate) est un produit de Mann Research Laboratory,.
Les inhibiteurs que nous avons employds sont: D(+)-saccharose, produit Merck No. 7652 ou 7653; a-D(+)-glucose, produit anhydre Mann Research Labora- tory ; I)(+)-maltose, produit Light.
Les concentrations de prot6ine sont mesur6es par leur absorbance g 280 m# en Kplmg/ml 0.96 pour la fl-lactoglobuline A; prenant pour coefficient d'extinction: ~lcm - -
1.58 pour la trypsine; et 2.oo pour l'a-chymotrypsine. Nous avons employ6 des saccharoses de 3 sources diff6rentes (Billaut; Merck
No. 7652 et 7653, qualit6 pour ampoules), avec des r6sultats identiques et parfaitement reproductibles.
Ni les impuretfs du saccharose ni la variation de viscosit6 ou de constante di61ectrique du milieu ne peuvent 6tre responsables de l'annulation de la vitesse de prot6olyse. En effet les concentrations en anions pr6sents dans les 6chantillons de saccharose (o.2 A 1. 5 M) restent dix mille fois plus petites que celles qui, dans les m6mes conditions de pH et de temp6rature, sont sans influence sur la vitesse 2. La concen- tration maximum en cuivre peut varier de 5.4" l°-55~ 4o.4 • io 5 M mais c'est une activa- tion de la vitesse que l'on observe avec une concentration de 3.5" lO-5 M (voir bibl. 2o). La pr6sence d 'EDTA (sans influence sur la prot6olyse trypsique de la lactoglobuline seule) augmente syst6matiquement l'inhibition due au saccharose d'environ lO% ce qui n'est pas significatif mais pourrait correspondre ~ une 16g6re activation due aux m6taux lourds. La viscosit6 du saccharose ne peut pas non plus ~tre invoqu6e pour rendre compte de l'inhibition observ6e puisque l'effet est invers6 lorsqu'on op6re avec des substrats diff6rents (TSAME, lactoglobuline native ou d6natur6e). De mSme la vitesse de l'hydrolyse trypsique de la lactoglobuline augmente quand la constante di61ectrique diminue 2t. Bien que la variation de constante di61ectrique soit dans nos exp6riences du m~me ordre de grandeur que celle des exp6riences de YON, le ph6nom6ne d'inhibition que nous observons ne peut, en toute rigueur lui ~tre attribu6, puisque les exp6riences 6tant par ailleurs, en tous points identiques, on observe dans un cas une forte activation, et dans l'autre, une inhibition totale.
Les mesures spectrophotom6triques sont faites au spectrophotom6tre Cary 14. Le dim6thyl-sulfoxyde est purifi6 par distillation sous vide au moment de l'emploi. La solution mod61e d'acides amin6s est pr6par6e A partir de N-ac6tyltyrosine 6thylester (Calbiochem, A grade), N-ac6tyltryptophane 6thytester et N-ac6tylphfnylalanine
Biochim. Biop]~ys. 3cla, t6o ( t968) 34 ° 359

/~-LACTOGLOBULINE A ET SACCHAROSE 343
6thylester (Mann Research Lab.). Pour ~viter les erreurs de volume dues A la viscosit6 des solutions de saccharose, nous avons pr6ffr~ aux pipettes, des seringues de type B.D. Cornwall.
Rt~SULTATS ET DISCUSSION
Formulation des donndes cindtiques L'hydrolyse trypsique des esters labiles et des substrats sp6cifiques de synth6se
se repr6sente g6n~ralement selon un sch6ma du type"
kl k2 k3 E ~ S ~-- t in ~ ES" + P[ ~ E ~ t),z
k ~ k 2 k a
avec E enzyme, S -- substrat, ES = complexe enzyme-substrat, ES' un inter- m6diaire chimique, probablement l '"acyl-enzyme" mis en 6vidence par de nombreux auteurs22-24, P1 et P2 les produits de la r6action.
L'existence de l'acyl-enzyme pour les substrats sp6cifiques est contest~e par quelques auteurs 25:6 et l'on a mis en 6vidence un plus grand nombre d'interm~diaires dans l'hydrolyse de quelques substrats simples 27. Cependant, et bien que l'on n'en ait encore aucune preuve dans le cas des prot6ines, on peut admettre qu'un schdma A deux 6tapes est applicable A leur hydrolyse.
La pr6sence de saccharose dans le milieu rdactionnel entralne une inhibition de la prot~olyse trypsique de la fl-lactoglobuline A (bibl. 28). Dans les conditions o/1 il n 'y a pas encore d'activation par le substrat (ph6nom~ne observ6e A la lois au cours de l 'hydrolyse des esters et des lactoglobulines A et B et pour lequel les cin6tiques d'hy- drolyse ne sont plus interpr6tables selon les lois classiques 29) les rdsultats exp6rimen- taux montrent que le sch6ma r~actionnel traduisant le fait que l'inhibiteur se lie au substrat (et non A l'enzyme) et que cette liaison entraine une inhibition totale est le seul A d6crire d'une mani~re satisfaisante les donn6es obtenues.
Ce schema est le suivant:
h i k2 /}3 E + S ~ E S - - - - > E S " -->E + P
!p k -[ IF IF
S I E S I E S ' I
L'expression de la vitesse initiale est"
1 .... rS1 = J v ~ avec [1~ >> [S] >> [E]:
(I ~- [ / ] /K i ) (IS] ~- Kin)
k2k3[Et] k3(k-1 + k2) V Km
k2 + k3 (k2 + ka) kl
[S] = concentration du substrat, [IJ de l'inhibiteur, [Etl totale de l'enzyme, et K, la constante de dissociation des diff~rents compos6s d'association avec l'inhibiteur.
Lorsque deux mol6cules d'inhibiteur se fixent sur une mol6cule de substrat le sch6ma est modifi6 comme suit:
Biochim. Biophys. Acta, 16o ([968) 340-359

344 j . CLEIHENT-METRAI~, J. YON
k~ k 2 k 3 E ~ ES" - ~ E S " ~ E £ P t~ /~, , I! IF
tq il ~I lq ~I tq F,[ ElS; t-l,q"
iI i~ l
EI~ EI,2S" EI.2S"
6 exprime la variation apparente d'affinit6 de Ki du fait de la liaison de la premiere molficule.
vlsj
Si l 'on envisage une interaction entre les sites de fixation de l ' inhibiteur sur la mol6cule de lactoglobuline, la th6orie g6n6rale des fonctions li6esS°, sa permet de d6finir un para- m+tre n tel que
n = ct In { I Z V ) / d In ~11
Y "saturat ion fractionnelle" de la macromol6cule par I c'est-/~-dire concentrat ion d ' inhibiteur li6 par mole et par site.
Ce param~tre est directement li6 ~ l'6nergie libre moyenne d ' interact ion entre les sites; il est facile de montrer que lorsqu'il y a des interactions stabilisatrices entre sites d 'une m~me mol6cule n > I, si ces interactions sont suffisamment fortes pour que toutes les formes interm6diaires soient n6gligeables n tend vers q, le nombre de groupes de la macromol6cule capables de fixer I ; s i n > q, il faut envisager des inter- actions entre mol6cules diff6rentes comme par exemple une dissociation polym~re- monom+re.
Dans l 'hypoth~se oh l ' inhibition de la prot6olyse provient de la liaison de l'inhi- biteur avec le substrat, il est raisonnable de penser que la vitesse de la r6action est directement li6e au degr6 de saturat ion du substrat par l ' inhibiteur (dans le cas de l ' inhibition totale dvidemment). Nous exprimerons donc in Y / ( - I Y) par l 'expression 6quivalente In (v vi)/vi oh, vi est la vitesse en prdsence de I .
Rdsultats expdrimentaux Nous avons t radui t nos exp6riences dans six rep%sentations graphiques, d6crites
en d6tail par WEBB 32. Les expdriences consistent ~ mesurer la vitesse initiale soit ~ [S~ constant et [Ij variable, soit ~ [SI] variable et [I] constant, pour une m~me concen- trat ion d 'enzyme (rapport6e ~ I mg/ml), dans les conditions suivantes : pH 7, tem- p6rature 35 °, force ionique o.I. La concentrat ion du substrat [Sj varie de 1. 4 5~ 38 rag/ ml ; celle du saccharose [I~ de o ~ 61o mg/ml (soit 1.78 M). On exprime la vitesse en ml de NaOH o.I M par min. par ml de solution pour I mg/ml de trypsine.
I1 est possible d'utiliser de plus fortes concentrat ions en substrat (allant jusqu'/~ IOO mg/ml), mais il faut alors tenir compte des variations de force ionique dues aux fortes quantit6s de soude ndcessaires pour amener le pH initial ~ 7. I1 convient d 'op6rer en NaC1 0.2 M et d 'employer de la soude 0.2 M (voir bibl. 33). C'est pourquoi nous n 'avons pas d@ass~ 38 rng/ml.
Les repr6sentations utilis~es s2 sont les representations classiques: A, I/V en fonction de I/[SI ;B, [S]/ven fonction de [SI; C, ven fonction de v/[S~ ; D, I/Ven fonction
Biochim. Biophys..4cta, 16o (1908) 34 ° 359

fl-LACTOGLOBULINE A ET SACCHAROSE 345
2C
v
3 2 -~og EI]
Fig. I. Variat ion de la vitesse initiale de prot6olyse de la fl-lactoglobuline A par la t rypsine (O) ou par l ' a -chymotryps ine (Q) en fonction de la concentrat ion en saccharose EI! pour une concentra t ion fixe en subs t r a t (lactoglobuline) de 6 mg/ml. En ordon~e la vitesse v e s t exprim4e en nil de NaOH o.I M par min par ml de solution et pour i mg/ml de trypsine. En absciss one a portd le cologarithme d4cimal de la concentrat ion en saccharose exprim6e en mg/ml, pH 7, 35 °, force ionique o.I.
de [I 1 ; E, I/i en fonction de I/EIJ ; (i = pourcentage d' inhibition -- I - - vi/v ; v, - - vitesse en pr6sence de l 'inhibiteur) ; F, EI] (I - i ) / i en fonction de ES!.
La confrontat ion des six repr6sentations nous montre que l ' inhibition apparem- ment du type non comp6tit if total met en jeu plus d 'une mol6cule d' inhibiteur. 11 est impor tan t tout d 'abord de d6terminer sur quelle prot6ine, enzyme on substrat , se fixe le saccharose.
L' inh ib i t eur se lie au substrat et non ~ l ' enzyme
Nous observons une inhibition tout ~ fait analogue en changeant d 'enzyme, alors qu'elle disparalt to ta lement ou en partie lorsqu'on change de substrat. Seuls les substrats prot6iques paraissent affect6s par la pr6sence de saccharose.
Rdsultats compards avec la t ryps ine et l 'a-chymotrypsine . La Fig. I repr6sente la vitesse initiale de prot6olyse trypsique ou a-chymotrypsique en fonction de l ' inverse du logarithme de la concentrat ion en saccharose, la concentrat ion en substrat 6tait 6 mg par ml dans les deux cas.
Les deux courbes sont absolument comparables et donnent des pourcentages d ' inhibit ion du mSme ordre de grandeur pour des concentrations identiques en inhibi- teur, comme le montre le Tableau I. La concentrat ion de saccharose qui produit une inhibition de 50% est du m~me ordre de grandeur pour les deux enzymes. 10. 5 -- 0.8 M pour l ' a -chymotryps ine; I0. 5 = 0. 7 M pour la trypsine.
T A B L E A U I
V A L E U R S D U P O U R C E N T A G E D ' I N H I B I T I O N i - - I Vi/V , E N F O N C T I O N D E LA C O N C E N T R A T I O N E N
S A C C H A R O S E P O U R LA P R O T ] ~ O L Y S E T R Y P S I Q U E O U ( ~ - C H Y M O T R Y P S I Q U E D E L A ~ - L A C T O G L O B U L I N E A
v, est la vitesse en presence de saccharose.
fl-Lactoglobuline .4 Saccharose i(%) (mg/ml) (hi )
Trypsine a-Chymotryp- sine
6.04 0.45 23 27 6.04 0.8 58 51 6.04 1. 5 ioo 83
Biochim. Biophys. dcta, 16o (1968) 340-359

346 j . { ' I . E M E N T - M E T I ( A L , J . Y t ) X
T A B L E A U 1I
V A L E U R S D U P O U R C E N T A G E I ) ' I N H I B I T I O N i 1 i'i,/?, I )E LA I ,ROTI~2OLYSE T R Y P S I Q U E DI.2 LA [;~
L A C T O G L O B U L I N E A D t ~ N A T U R I ~ E P A R LA C H A L E U R ( F O R M E " f " {BIHL. 2 0 ) ) , P O U R D I F F t ~ . R E N T E S VA
L E U R S D E LA C O N C E N T R A T I O N E N S A C C H A R O S E
v, - v i t e s s e e n p r e s e n c e d e s a c c h a r o s e .
[~-Lactoglobuline A Saccharose i ddnaturde ( 3 l ) (%) (mg/ml)
5 . 9 4 0. 4 I o 5 , 9 4 0 .8 22 5 . 9 6 o . 7 2 2 .98 0 .7 24
Rdsultats compards avec d'autres substrats. Nous obtenons des rdsultats tr6s diffd- rents lorsque nous changeons de substrat. Nous avons utilis6 la fl-lactoglobuline A d6natur6e par la chaleur sous sa forme "f" et le TSAME.
Avec la prot6ine d6natur6e nous obtenons une inhibition beaucoup plus faible, diminu6e de plus de moiti6 (Tableau II). Cette diminution ne peut ~tre like qu'/~ la modification de la structure de la protdine substrat puisque routes choses sont 6gales par ailleurs.
Avec le TSAME l'exp6rience est faite dans les m~mes conditions que prdcddem- ment. La seule pr&aution suppl6mentaire consiste ~t mesurer la ddrive due A l'hydrolyse spontande, non enzymatique du TSAME, A cette temperature et ce pH. I1 n'y a aucune inhibition par le saccharose, au contraire une ldg~re activation, r6sumde darts le Ta- bleau III qui parait presque disparaltre aux fortes concentrations de substrat. A peine significative aux faibles concentrations de substrat, elle correspond peut-~tre ~t un abaissement de la constante di6lectrique du milieu qui est dans ces exp6riences d'en- viron IO unit6s; ces r6sultats sont en accord avec ceux d'autres auteurs sur l'effet des variations de la constante di6lectrique a4.
T A B L E A U 111
V A L E U R S D E L ' A C T I V A T I O N D U E A U S A C C H A R O S E D A N S L A P R O T E O L Y S E T R Y P S I Q U E D U T S A M E
( p H 7, 35 °)
£'~ccharose "I'.';A .111£ Pourcentage (:l l) ( M ) d'aclivation
o . 4 5
o .8
2 . 1 0 4 ~ 2 8
5. t o - I -- 20 t o 4 ~ ~3
5 . t o a " 4 t o 2 o
5 . 1 0 2 O
2 . 1 o I -I 33 5 . t o a 4 26
IO -a + 2 4 . 5 . 10 -3 @ 2 I
1 o 2 + 9
5 . t o 2 + 6
Biochim. Biopkys. A c/a, 160 ( t 968) 3 4 0 - 3 5 9
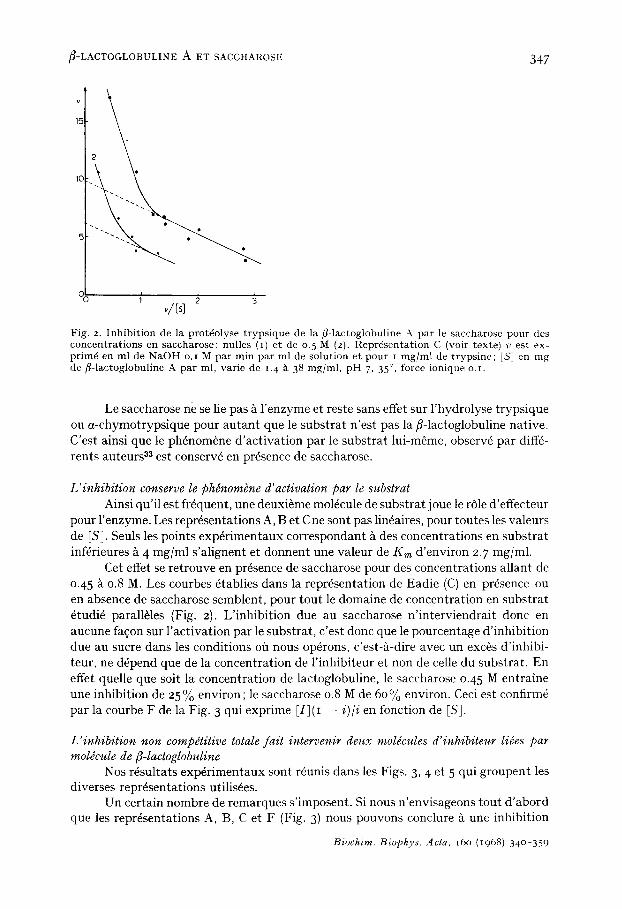
~-LACTOGLOBULINE A ET SACCHAROSE 347
v '
15
10
2 1
• •
4 ) 7 4E~]
Fig. 2. Inhibi t ion de la prot6olyse t ryps ique de la fl-lactoglobuline A par le saccharose pour des concentra t ions en saccharose: nulles (i) et de o. 5 M (2). Repr6senta t ion C (voir texte) v est ex- prim6 en ml de NaOH o.i M par min par ml de solution et pour i mg/ml de t rypsine; IS] en mg de /~-lactoglobuline A par ml, varie de t.4 k 38 mg/ml, p H 7, 35 °, force ionique o.t.
Le saccharose ne se lie pas A l'enzyme et reste sans effet sur l'hydrolyse trypsique ou a-chymotrypsique pour autant que le substrat n'est pas la fl-lactoglobuline native. C'est ainsi que le ph6nom+ne d'activation par le substrat lui-m~me, observ6 par diff6- rents auteurs 33 est conserv6 en pr6sence de saccharose.
L'inhibition conserve le phdnombne d'activation par le substrat Ainsi qu'il est frequent, une deuxi+me mol6cule de substrat joue le r61e d'effecteur
pour l'enzyme. Les repr6sentations A, Bet C ne sont pas lin6aires, pour toutes les valeurs de [S]. Seuls les points exp6rimentaux correspondant h des concentrations en substrat inf6rieures A 4 mg/ml s'alignent et donnent une valeur de Km d'environ 2.7 mg/ml.
Cet effet se retrouve en presence de saccharose pour des concentrations allant de 0.45 A 0.8 M. Les courbes 6tablies dans la repr6sentation de Eadie (C) en pr6sence ou en absence de saccharose semblent, pour tout le domaine de concentration en substrat 6tudi6 parall+les (Fig. 2). L'inhibition due au saccharose n'interviendrait donc en aucune fa~on sur l 'activation par le substrat, c'est donc que le pourcentage d'inhibition due au sucre dans les conditions off nous op6rons, c'est-A-dire avec un exc+s d'inhibi- teur, ne d6pend que de la concentration de l'inhibiteur et non de celle du substrat. En effet quelle que soit la concentration de lactoglobuline, le saccharose 0.45 M entraine une inhibition de 25 % environ; le saccharose 0.8 M de 60% environ. Ceci est confirm6 par la courbe F de la Fig. 3 qui exprime [I](I -- i)/i en fonction de IS].
L'inhibition non compdtitive totale fait intervenir deux moldcules d'inhibiteur li~es par moldcule de fl-lactoglobuline
Nos r6sultats exp6rimentaux sont r6unis dans les Figs. 3, 4 et 5 qui groupent les diverses repr6sentations utilis6es.
Un certain nombre de remarques s'imposent. Si nous n'envisageons tout d 'abord que les repr6sentations A, B, C et F (Fig. 3) nous pouvons conclure ~ une inhibition
Biochim. Biophys. Acta, 16o (1968) 34 ° 359
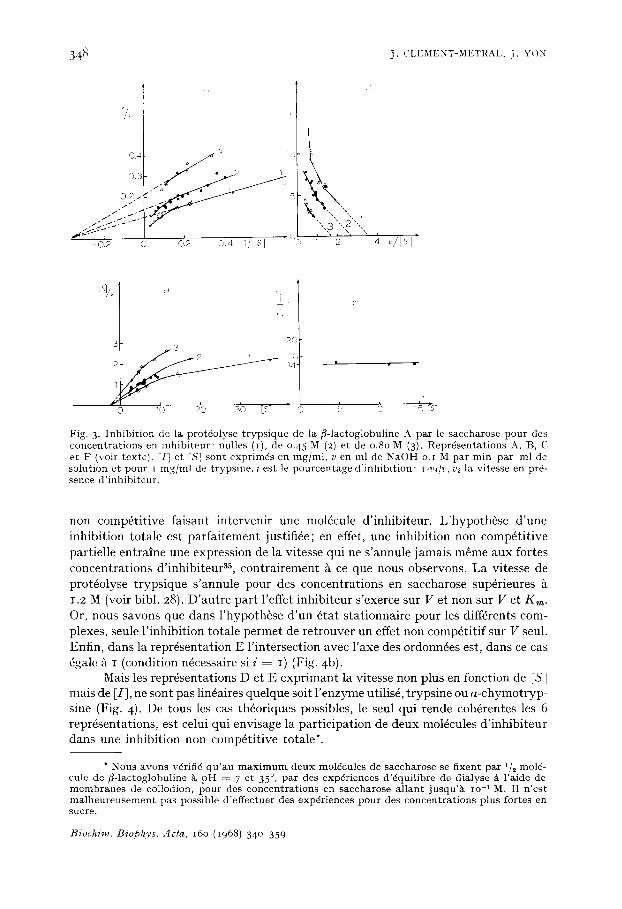
348 j . CLEMENT-METRAL, J. YON
r
. i " I-~..~%c.- ,,, "",,, ,
..:.-..;;; -"-:-"; - ] . " "'-,3 "..? ".: i ~ ' ~ ~ | ~ i - C ; ! i "~ , ", "
- 0 . 2 0 o . 2 0 . 4 , ," I s ] , , ! 2
i
4 ~/k,s]
3
2 J
1
0
i !
R C 3
" 16
i i
"c, 2o sb Is; c
Fig. 3. I n h i b i t i o n de la protdolyse tryps ique de la /3-1actoglobul ine A p a r le saccharose pour des concentrat ions en inhibiteur: nulles ( i ) , de o.45 M (2) e t de o.8o M (3). R e p r 6 s e n t a t i o n s A, B, C et F (voir t e x t e ) . [I] e t IS] s o n t exprimds en m g / m l , v e n m l de N a O H o . i M par rain par m l de s o l u t i o n e t p o u r t i n g / m l de trypsine, i e s t le pourcentage d' inhibi t ion : I-V,/V, vi la vitesse en pr6- sence d'inhibiteur.
non compfti t ive faisant intervenir une mol~cule d'inhibiteur. L'hypoth6se d'une inhibition totale est parfaitement justifi6e; en effet, une inhibition non comp6tit ive partielle entraine une expression de la vitesse qui ne s'annule jamais m~me aux fortes concentrations d'inhibiteur 3s, contrairement ~ ce que nous observons. La vitesse de prot4olyse trypsique s'annule pour des concentrations en saccharose sup6rieures 1.2 M (voir bibl. 28). D'autre part l'effet inhibiteur s'exerce sur V et non sur V et Kin. Or, nous savons que dans l'hypoth~se d'un 6tat stationnaire pour les diff6rents com- plexes, seule l'inhibition totale permet de retrouver un effet non comp6titif sur V seul. Enfin, dons la repr6sentation E l'intersection avec l'axe des ordonn~es est, dons ce cas ~gale ~ I (condition n~cessaire si i - - I) (Fig. 4b).
Mais les repr6sentations D et E exprimant la vitesse non plus en fonetion de IS] mais de [I], ne sont pas lin6aires quelque soit l 'enzyme utilis6, trypsine ou a-chymotryp- sine (Fig. 4). De tous les cas th6oriques possibles, le seul qui rende coh6rentes les 6 repr6sentations, est celui qui envisage la participation de deux mol6cules d'inhibiteur dans une inhibition non eomp6titive totale*.
* N o u s a v o n s v6rif i6 qu'au m a x i m u m d e u x m o l e c u l e s de saccharose se f ixent par 1/2 mol6- c u l t de /3-1ac toglobul ine 5, p H . - 7 e t 35 °, par des exp6riences d'~quilibre de dialyse & l ' a i d e de m e m b r a n e s de co l lod ion , pour des concentrat ions en saccharose a l lant jusqu'5` i o 1 M. I1 n'est m a l h e u r e u s e m e n t pas possible d'effectuer des exp6riences pour des concentrat ions plus fortes en sucre .
Biochim. Biophys. Acta, 16o (1968) 34 ° 359
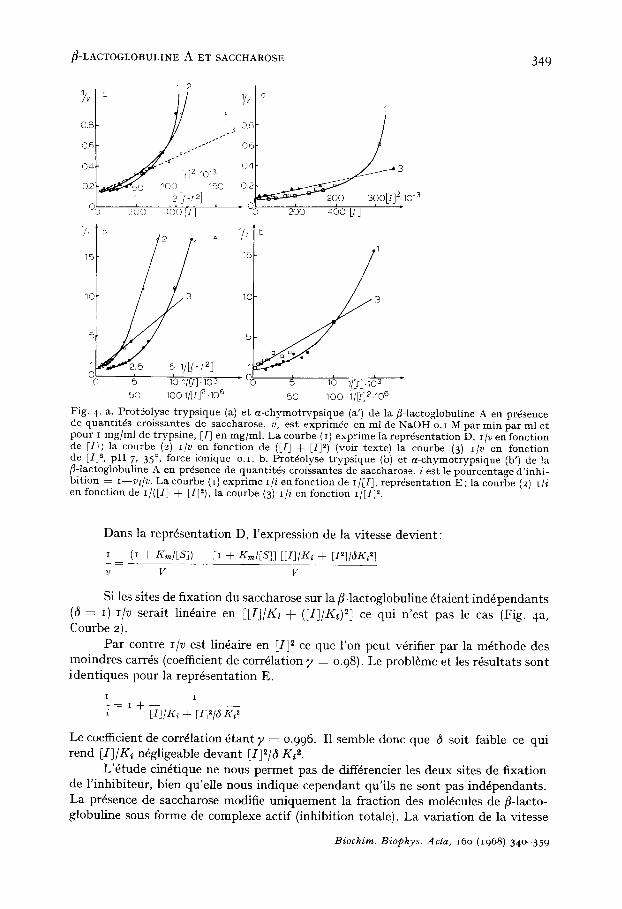
~-LACTOGLOBULINE A ET SACCHAROSE 349
1 2
0.8 3
0.6
04 2.10-3
0.2 ~ ' ~ % 0 ~00 15C I 2 [Z+I 2]
0 C ' - 2~'0 40'0 r j ]
15 / 2 / I a
5
o ~ ~5 W].1o3 5o loo 1/It] 2.1o 6
q/v
0.~
O,E
04
0.2
yi
1.5
10
5
11
o'
1
211o_3 200 40o [z]
b'
1
16 ~IOd.1& 50 l o o 1/[r]24o 6
Fig. 4. a. Prot~olyse t ryps ique (a) e t a - chymot ryps ique (a') de la f l - lactoglobuline A en pr4sence de quant i t4s croissantes de saccharose, v, est exprim4e en ml de NaOH o.i M par rain par ml et pour I mg/ml de trypsine, [I] en mg/ml. La courbe (i) expr ime la repr4senta t ion D, I/v en fonct ion de [I]; la courbe (2) i/v en fonct ion de ([I] + [I]~) (voir texte) la courbe (3) i/v en fonct ion de [I] 2, pH 7, 35 °, force ionique o . I . b . Prot4olyse t ryps ique (b) et a -chymot ryps ique (b') de la fl-lactoglobuline A en pr6sence de quant i t6s croissantes de saccharose, i e s t le pourcentage d' inhi- bi t ion = i --vi /v . La courbe (I) expr ime i l l en fonct ion de I/[I], repr4sentat ion E; la courbe (2) Ill en fonct ion de I/([I] + [I]~), la courbe (3) I/i en fonct ion I /[I j I.
Dans la repr6sentation D, l'expression de la vitesse devient :
I (I + Km/[S]) [I + Km/[S]] [[I]/K~ + [I~]/6K, ~] +
v V V
Si les sites de fixation du saccharose sur la fl-lactoglobuline 6taient ind6pendants (~ ---- I) I/V serait lin6aire en [[I]/Ki + (II]/K~) 2] ce qui n'est pas le cas (Fig. 4 a, Courbe 2).
Par contre I/V est lin6aire en [I] 2 ce que l'on peut v6rifier par la m6thode des moindres carr6s (coefficient de corr61ation y = 0.98 ). Le probl6me et les r6sultats sont identiques pour la repr6sentation E.
i i
i [ I ] /K , + [112/6 K , 2
Le coefficient de corr61ation 6tant ? ~ 0.996. I1 semble donc que 6 soit faible ce qui rend [I]/K, n6gligeable devant [I]2/~ K, 2.
L'6tude cin6tique ne nous permet pas de diff4rencier les deux sites de fixation de l'inhibiteur, bien qu'elle nous indique cependant qu'ils ne sont pas ind6pendants. La pr6sence de saccharose modifie uniquement la fraction des moMcules de fl-lacto- globuline sous forme de complexe actif (inhibition totale). La variation de la vitesse
Biochim. Biophys. dcta, 16o (1968) 340-359
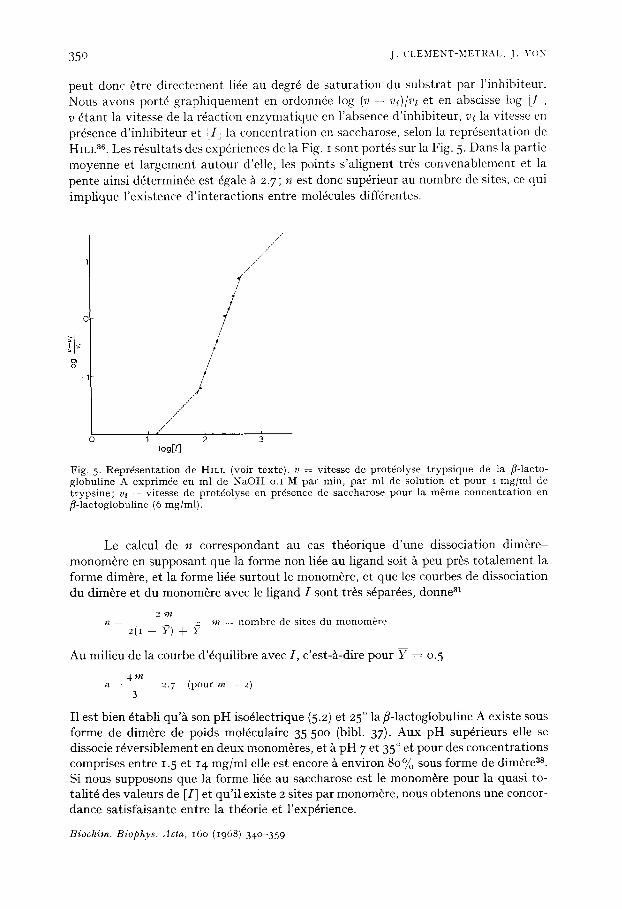
3 5 0 J . C I . E M E N T - b l E T R A I . , J . v o x
peut done fitre d i rec tement lide au degr4 de sa tu ra t ion du subs t ra t par l ' inhibi teur . Nous avons port6 g raph iquemen t en ordonnde log (v - v.~)/v, et en abscisse log [ I ' , v 6 tant la vitesse de la r6act ion enzymat ique en l 'absence d ' inh ib i teur , v, la vitesse en pr6sence d ' i nh ib i t eu r et ~Ii la concent ra t ion en saccharose, selon la repr6senta t ion de HILL a~. Les rdsul ta ts des exp6riences de la Fig. I sont port6s sur la Fig. 5. Dans la par t ie moyenne et l a rgement au tour d'elle, les points s ' a l ignent tr6s convenablement et la pente ainsi d6termin6e est 6gale ~ 2.7; ~z est donc sup6rieur au nombre de sites, ce qui impl ique l 'exis tence d ' in te rac t ions entre mol6cules diff6rentes.
g -I
/ /" /"
/, ,/ /.
/" 7"
/, / / /
/¢" ,/
I d ~ 3
log[f]
Fig . 5- R e p r d s e n t a t i o n d e HILL (vo i r t e x t e ) , v - - v i t e s s e d e p r o t 4 o l y s e t r y p s i q u e d e l a ]~- lacto- g l o b u l i n e A e x p r i m d e e n m l d e N a O H o . I M p a r m i n , p a r m l de s o l u t i o n e t p o u r i m g / m l d e t r y p s i n e ; v, - - v i t e s s e d e p r o t d o l y s e e n p r d s e n c e d e s a c c h a r o s e p o u r l a m 6 m e c o n c e n t r a t i o n e n / ~ - l a c t o g l o b u l i n e (6 m g / m l ) .
Le calcul de n cor respondant au cas th6orique d 'une dissociat ion dim~re monom6re en supposan t que la forme non li6e au l igand soit ~ peu pros to t a l emen t la forme dim~re, et la forme li6e su r tou t le monom~re, et que les courbes de dissociat ion du dim6re et du monom~re avec le l igand I sont tr~s s6par6es, donne 3t
2 m n m == n o m b r e d e s i t e s d u m o n o m 6 r e
20 - ?) + ?
Au mil ieu de la courbe d '6qui l ibre avec I , c 'es t-5-dire pour Y = 0.5
4 m n = - = 2. 7 ( p o u r m = 2)
3
n e s t bien dtabl i qu '~ son p H isodlectrique (5.2) et 25 ° la/5-1actoglobuline A existe sous forme de dim&re de poids mol6culaire 35 50o (bibl. 37)- Aux p H sup6rieurs elle se dissocie rdvers ib lement en deux monom~res, et ~ p H 7 et 35 ° et pour des concent ra t ions comprises entre 1. 5 et 14 mg/ml elle est encore 5 environ 80% sous forme de dim&re as. Si nous supposons que la forme lide au saccharose est le monom~re pour la quasi to- ta l i td des valeurs de [I] et qu ' i l existe 2 sites pa r monom~re, nous obtenons une concor- dance sa t i s fa i san te ent re la th4orie et l 'exp6rience.
Biochim. Biophys. Acta, 16o ( I968) 3 4 o - 3 5 9

--LACTOGLOBULINE A ET SACCHAROSE 3 5 I
F.tude thermodynamique Les variations d'6nergie libre, d'enthalpie et d'entropie, pour une temp6rature
donn6e, sont li6es aux constantes d'6quilibre K et aux constantes de vitesse k, par les relations :
AG = z lH -- T A S = --2. 3 R T log K ; A G * = 3 H * T A S * = - -2 . 3 R T log h_k k T
R = constante des gaz, h = constante de Planck, k = constante de Boltzmann. Les variations de K et k avec T permettent de d6terminer AH et AH* par les
relations classiques :
d(log tf) d(log k) z lH = 2. 3 R - - - - ; A H * + R T -- --2.3 R - -
d( i /T) d ( i /T )
Dans une r6action enzymat ique/ t ~tapes successives, l'effet global de la tem- p6rature est la somme des effets s6pards sur chacune des 6tapes que l 'analyse cin6tique ne permet pas de diff6rencier. Les constantes exp6rimentales Km et V n 'ont de signi- fication physique que dans certains cas favorables o£1 l 'une des 6tapes est limitante. I1 leur correspond des constantes thermodynamiques exp6rimentales:
AG *v , A H * v , A S * v , A G x m, A H K . v A S u . ,
Les constantes thermodynamiques correspondant k Km sont difficilement inter- pr6tables puisque Km est une expression complexe renfermant plusieurs constantes.
Cependant Km est constant en fonction de la temp6rature, dans le cas de l 'hy- drolyse de la/3-1actoglobuline native seule ~ et en pr6sence de saccharose ~ deux con- centrations diff6rentes (comme nous l 'avons v6rifi6), alors que V varie; on peut donc repr6senter log ven fonction de I/T au lieu de log V (Fig. 6). La pente des deux droites V
20
10
£2 3:3 3:4 d/r ) "Io 3 "K
Fig. 6. R e p r 6 s e n t a t i o n semi - logar i thmique . E n ordonn6e la v i tesse de protdolyse t r y p s i q u e de la f l - lactoglobuline A seule (cercles pleins) ou en pr6sence de saccharose o. 7 M, expr im6e en ml de N a O H o.I M pa r m i n pa r ml de so lu t ion et pour I m g / m l de t ryps ine . E n abscisse l ' inverse de la t e m p 6 r a t u r e absolue, p H 7, force ionique o.I.
Biochim. Biophys. Acta. i6o (1958) 340-359

3 5 2 ] . C L E M E N T - M E T R A L , J . YON
T A B I . E : k U IX, '
VALEUR [)ES CONSTANTES TttERMO1)YNAMIQUES CORRESPONI)ANT ~t V, a 3 5 ET p H 7
l . es v a l e u r s d e z I H # s o n [ c a l c u l d e s it p a r t i r d e s p e n t e s d e s c o u r b e s d e l a F ig . 6, d o n t les c o e f f i c i e n t s d e r~Sgression s u i v i s d e l e u r d c a r t - t y p e s o n [ : p o u r la /1 l a c t o g l o b u l i n e seu le (A 6.8 _- 1.5), p o u r l a / ~ q a c t o g l o b u l i n e + s a c c h o r a s e o. 7 M (B ~ : 4.5 - o .8) . L ' a n a l y s e d e c o v a r i a n c e d o r m e F = 7 .372 q u i c o r r e s p o n d k P o . o l 5 cc q u i e s t t r 6 s s i gn i f i c a t i f . P o u r le c a l c u l d e . l ( / * v o i r t e x t e .
AH* (ca//mole) A(;* (cal/mole) AS* (u.e./mole)
/ ~ - L a c t o g l o b u l i n e A seu le 3o 5oo ~ 6 o o o I9 9 0 o 34 / 5 - L a c t o g l o b u l i n e ~ s a c c h a r o s e o. 7 M 2o 30o c 3 o o o 2o 3oo o
obtenues, l 'une sans inhibi teur , l ' au t r e avec une concentra t ion en saccharose o. 7 M nous pe rme t de calculer ZIH* dans les deux cas (Tableau IV).
Si le sucre 6tai t li6 ~ la prot6ine pa r des liaisons hydrog6nes, ZIH* augmen te ra i t en pr6sence de sucre, puisqu ' i l diminue, il y a donc p robab lemen t s tab i l i sa t ion pa r fo rmat ion de l iaisons hydrophobes .
Puisque nos exp6riences ont su r tou t fit6 faites dans la zone o/1 la vitesse n 'es t pas lin6aire en fonction de v~ [S:, nous avons pris pour va leur de la cons tante de vitesse k une va leur moyenne de o.o 7 s ec 1, dans le calcul de zIG*. Les aut res cons tantes ont pour va leur : poids mol6culaire de la t ryps ine : 24 5oo ; h - - 6.62. IO 27 ; k ~ 1.38. IO 16 ; a = o.38; (voir bibl . 2, 19) on adme t que le nombre to ta l de liaisons rompues pa r mol6cule de fl- lactoglobuline est de 2o. a = coefficient de dissociat ion de la r6action
NHa+ -~ N H 2 + H +. Une concent ra t ion en saccharose o. 7 M, qui correspond 5 une inhibi t ion de
5o% ent ra lne une var ia t ion de zIG* de 4oo calories et une impor t a n t e var ia t ion nfga- t ive d ' en t rop ie zIS*. Nous pouvons th fo r iquemen t calculer l '6nergie libre to ta le d ' in- te rac t ion sucre lactoglobuline, pour la macromol6cule compl6tement saturde de ligand, b~ pa r t i r de la repr6senta t ion di te de HILL. L ' ex t r a po l a t i on des a sympto t e s de pente I effectu6e sur la Fig. 5 donne pour valeur min imum de l '6nergie to ta le d ' i n t e rac t ion sucre--lactoglobuline 76o calories. Cette valeur compar fe ~ celle de 4oo calories calcul6e pour la demi inhib i t ion est un a rgument en faveur de l 'hypoth6se selon laquelle l ' in- h ib i t ion est ent i6rement due/~ la liaison saccharose lactoglobul ine et ~ l 'exis tence de fortes in terac t ions s tabi l i sa t r ices au sein de la molfcule de laetoglobuline.
La var ia t ion n6gat ive d ' en t rop ie d ' a c t i va t i on de la rdact ion de prot6olyse lorsqu 'el le s 'effectue en pr6sence de saccharose t r a du i t la d ispar i t ion d 'un cer ta in nombre de degr4s de l ibert6 p robab lemen t li6 ~t un changement de conformat ion de la lactoglobuline.
Nous avons donc 6tudi4 l 'effet du saccharose sur la s t ruc ture de la fi-lactoglo- bul ine en l 'absence de tou te r6action enzymat ique 5~ l ' a ide des courbes de t i t rage et de la spec t rophotomdt r ie de diff6rence.
Courbes de titrage de fl-lactoglobuline A La liaison est tr~s rapide, au plus 6gale ~ une seconde, l imitde pa r le t emps de
m~lange. Les courbes de t i t rages de fl- laetoglobuline A ont 6td effectu6es en absence et en
prdsence de sucre ~ diff6rentes concentra t ions de l 'une et de l ' aut re , en solut ion dans
Biochim. Biophys. Acta, 16o ( I968) 3 4 o - 3 5 9

fl-LACTOGLOBULINE A ET SACCHAROSE 353
NaC1, pour une force ionique de o.I. NOZAKI, BUNVILLE ET TANFORD 39, reprenant le travail de CANNAN, PALMER ET KIBRICK 4°, ont retrouv6 des courbes expdrimentales identiques bien que l'interprdtation quantitative puisse ~tre ldg+rement modifid. C'est pourquoi nous avons fait les calculs selon CANNAN, en prenant pour poids moldculaire de la protdine 40 ooo, ce qui entralne une l~g~re translation de la courbe, sans incon- vdnient s'il ne s'agit tout d'abord que d'observer un changement relatif. Nous avons ainsi n~glig6 la correction due ~ la liaison des ions C1- et Na +. Selon CARR ~1 et NOZAKI, B~:XVlLLE ET TANFORD 39, les ions C1- ne se lient pas ~t la protdine dans la zone de pH qui nous intdresse, par contre le pH isodlectrique de la lactoglobuline diminue quand on lui ajoute du KC1, indiquant ainsi qu'elle poss+de des sites de forte affinit6 pour K + (voir bibl. 42) ; il en est de m~me pour les ions Na + entre pH 6 et 9 (voir bibl. 43). Nous~n'en avons eependant pas tenu eompte, ne sachant pas quelle aurait 6td la correction en prdsence de saccharose et nos courbes t~moins sont donc les m~mes que celles de CANNAN. Elles expriment Z la charge nette en fonction du pH. Les courbes qui cor- respondent au titrage en pr6sence de saccharose sont corrigdes ~t l'aide des courbes de titrage du saecharose seul, qui montrent que le saccharose peut ~tre assimild ~t un diacide faible dont les pK seraient pK1 < 3; pK2 > 9.5.
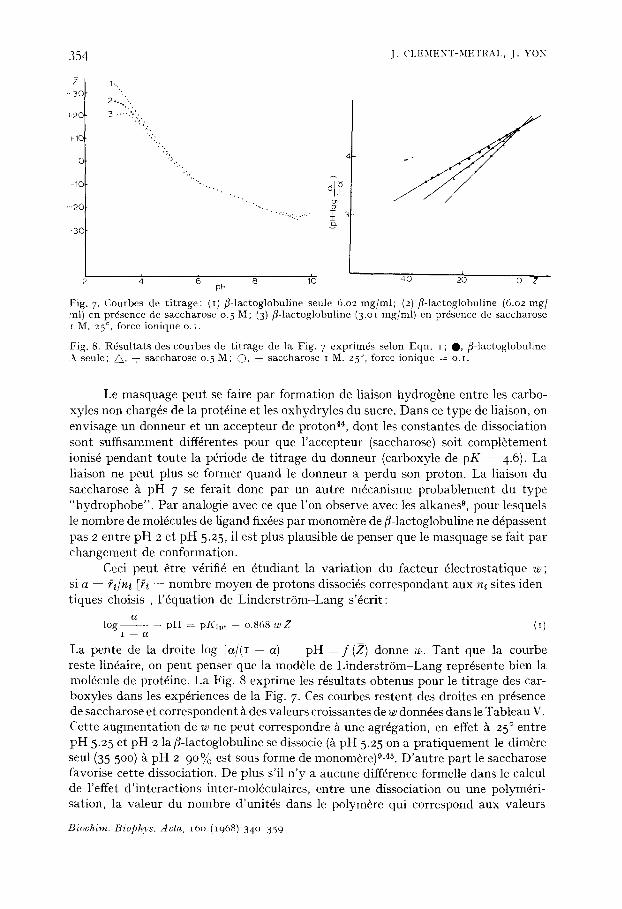
La Fig. 7 pr6sente 3 courbes de titrage de/3-1actoglobuline A effectudes: l'une sans saccharose, les deux autres pour des concentrations en protdine EPI et en sucre [I! telles que le rapport [PI/I[~ soit constant. Elles sont r6versibles jusqu'~t pH 9.5.
La courbe I prdsente les trois rdgions classiques: (a) Une rdgion de pKin t = 4.6 dans laquelle ne sont titr6s que les carboxyles dont
le nombre total est 52 pour une moldcule de poids mol6culaire de 35 500. Les deux a-COOH n'apparaissent pas sdpar~s des autres.
(b) Une r~gion centrale faiblement tamponnde dans laquelle sont titr6s les deux carboxyles de pKint -- 7.3, les a-NH 2 et les groupes imidazoles (pKint = 7.3)- La moldcule subit un changement de conformation autour de pH 7.5, qui est lid l'ionisation de deux carboxyles anormaux 4. Ce ph6nom+ne se retrouve sur les courbes 2 e t 3 .
(c) Une r6gion alcaline de pKint ~ 9.95 qui correspond aux groupes e-NH 2 et aux ph6nols. Les groupes guanidiques dont le pK est aux environs de 12 ne sont pas titrds sur nos courbes. Les anomalies que nous observons ne sont dues ni ~t des modi- fications de force ionique, ni / t u n changement de constante didlectrique, ni 5~ des valeurs anormales des pKint. En effet il n 'y a pas de variation de force ionique due au saccharose car les volumes de base ou d'acide ajout6s sont tout ~ fait du m~me ordre de grandeur en pr6sence ou en absence de saccharose. Pour une variation de D de 80 ~t 18, on peut observer thdoriquement une variation de pK de 4 a 5 unitds. Les courbes ne sont pas modifi6es dans ce sens (en particulier pas de modification du pK des COOH). De plus la variation de eonstante didlectrique due au saccharose est faible (AD _< 20).
Les courbes 2 et 3 de la Fig. 3 pr6sentent des anomalies dans la partie acide et dans la partie alcaline. Entre pH 6 et pH 8. 5 elles sont identiques aux courbes de titrage de ~-lactoglobuline A seule, les pKint des groupes carboxyles, imidazoles et a-NH 2 sont conservds. On peut attribuer la forme des courbes, entre pH 3 et pH 6 ~ un "masquage" des carboxyles par liaison avec le saccharose ou par changement de con- formation. Bien que nous n'ayons pu atteindre les r6gions acides extr~naes, ~ pH 3 le nombre de COOH masquds dans les conditions de la courbe 3 atteint 15 par dim~re de/3-1actoglobuline (35 5oo), il pourrait ~tre de 20 ~ 25 au maximum/t pH I.
Biochim. Biophys. Acta, 16o (t968) 34 ° 359
354
2
..30
~2o
flO
0
-10
, 20
.30
j . CI.EMt~NT-METRAL, J. YON
1- . ,
2 ° ° o i ' " .
? ...~:o:'.
i I ,IZ: I
I ; I
' " ' , . . . . .
~I iI 1
" ' . . . .
" ' . . . .
1 I" I . ii :2 ~II I I ii" T
#o & Ib 4'0 2& ~) y p~
Fig. 7. Courbes de t i trage: (i) fl-lactoglobuline seule 6.02 mg/ml; (2) fl-lactoglobuline (6.02 mg] rnl) en pr6senee de saccharose 0.5 M; (3) fl-lactoglobuline (3.ol mg/ml) en prdsence de saccharose
. o t M. 27 , force ionique o.i.
Fig. 8. R6sultats des courbes de t i trage de la Fig. 7 expriln~s selon Eqn. i ; O, fl-lactoglobuline seule; • , + saccharose o.5 M; C), + saccharose I M. 25 °, force ionique = o.r.
Le masquage peut se faire par formation de liaison hydrog6ne entre les carbo- xyles non charg6s de la protfine et les oxhydryles du sucre. Dans ce type de liaison, on envisage un donneur et un accepteur de proton 44, dont les constantes de dissociation sont suffisamment diff6rentes pour que l'accepteur (saccharose) soit compl6tement ionis6 pendant toute la p6riode de titrage du donneur (carboxyle de pK 4.6). La liaison ne peut plus se former quand le donneur a perdu son proton. La liaison du saccharose ~ pH 7 se ferait done par un autre m6canisme probablement du type "hydrophobe". Par analogie avec ce que l'on observe avec les alkanes 9, pour lesquels le nombre de mol6cules de ligand fix6es par monom~re de fi-lactoglobuline ne d6passent pas 2 entre pH 2 et pH 5.25, il est plus plausible de penser que le masquage se fait par changelnent de conformation.
Ceci peut 6tre v6rifi6 en 6tudiant la variation du facteur 61ectrostatique w; si a = ~i/n~ [~ = n o m b r e moyen de protons dissoci6s correspondant aux n~ sites iden- tiques choisis], l '6quation de Linderstr6m-Lang s'dcrit:
C ¢
log . . . . . p H = pKin t 0.868 w ~ (I) I C~
La pente de la droite log [ a / ( I - - a)] pH f (Z) donne w. Tant que la courbe reste lin6aire, on peut penser que la mod6le de Linderstr6m-Lang repr6sente bien la mol6cule de prot6ine. La Fig. 8 exprime les r6sultats obtenus pour le titrage des car- boxyles dans les exp6riences de la Fig. 7. Ces courbes restent des droites en pr6sence de saccharose et correspondent/~ des valeurs croissantes de w donn6es dans le Tableau V. Cette augmentation de w ne peut correspondre ~ une agr6gation, en effet ~t 25 ° entre pH 5.25 et pH 2 la fl-lactoglobuline se dissocie (5 pH 5.25 on a pratiquement le dim~re seul (35 500) 5~ pH 2 90% est sous forme de monom~re)°, 45. D'autre part le saccharose favorise cette dissociation. De plus s'il n 'y a aucune diff6rence formelle dans le calcul de l'effet d'interactions inter-mol6culaires, entre une dissociation ou une polym6ri- sation, la valeur du nombre d'unit6s dans le polym~re qui correspond aux valeurs
Biochim. Biophys. dcta, 16o (I968) 34 ° 359

fl-LACTOGLOBULINE A ET SACCHAROSE 355
T A B L E A U V
VALEURS DE, ZZ) (Eqn, I), CORRESPONDANT AU TITRAGt~ DES COOH, pKlnt 4.5 8
f l -Lactoglobul ine A seule 0.0346 f l -Lactoglobul ine + saccharose 0. 5 M 0.0438 f l -Lactoglobul ine + saccharose I M o.o581
exp6rimentales est 2, or le seul polym6re connu, ~ basse temp6rature seulement, est l 'octam6re et non le t6tram6re.
L 'augmentat ion de la valeur de w semble donc en rapport avec une structure plus compacte de la mol6cule de/5-1actoglobuline qui entralnerait un masquage d 'un certain nombre de carboxyles. On peut rapprocher ce r6sultat d 'un travail de Lo- VRIENa6: la fixation d'un maximum de IO mol6cules de sulfate dod6cylique de sodium par mol6cule de s6rum albumine bovine ~ des pH alcalins, entraine une augmentation de w qui varie parall61ement ~ une diminution du volume hydrodynamique. Ceci n 'est pas surprenant: il est bien connu que la ~-lactoglobuline peut sous des influences di- verses prendre une conformation plus compacte. Le changement de conformation observ6 entre pH 4 et pH 6 (voir bibl. 5) Q ~ N ; celui qui accompagne l'ionisation des 2 carboxyles anormaux entre pH 6. 5 et pH 9.5 (voir bibl. 4) N -+ R correspondent une structure de plus en plus compacte de Q ~ R. La liaison d'hydrocarbures entraine de m4me un changement de conformation de la fl-lactoglobuline qui lui donne une structure plus compacte s.
Les carboxyles ne peuvent 6tre dissimul6s sous forme charg6e 47, ils le sont plus probablement par formation de paires neutres avec les groupes e-NH3+ par exem- ple 5, ce qui est peut-fitre apparent sur la courbe 3 de la Fig. 7, bien qu'en raison de la d6naturation irr6versible de la fl-lactoglobuline, nous ne puissions effectuer de titrage au del~ de pH 9.5.
Spectre de diffdrence Nous avons 6tudi6 les spectres ultraviolets de diff6rence de $-lactoglobuline A
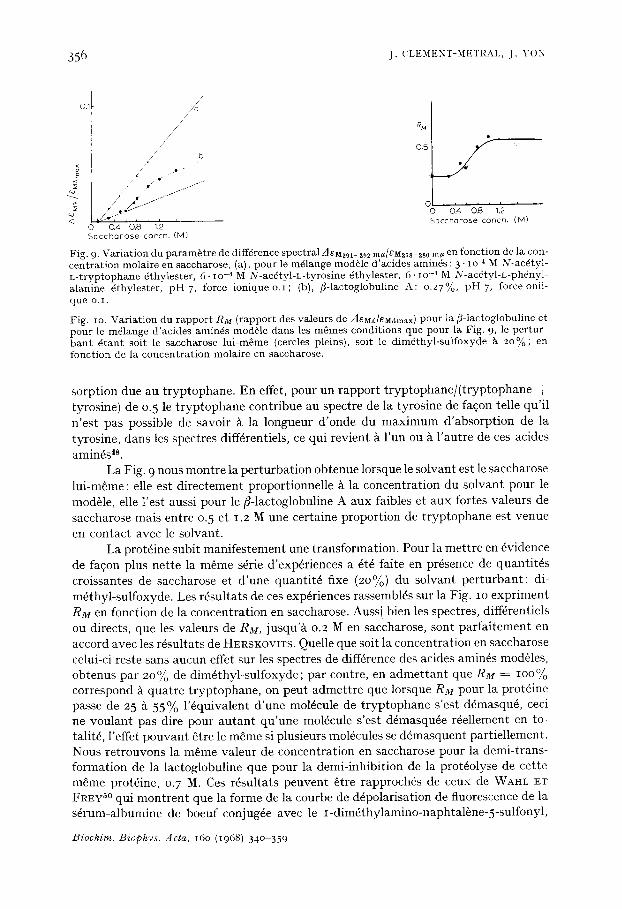
avec ou sans saccharose, par la m6thode de perturbation due au solvant introduite par HERSKOVITS ET LASKOWSKI as,49. On compare la perturbation apport4e ~ la prot6ine par un solvant donn6, ~ celle produite par le m~me solvant sur un mod61e: solution des chromophores tyrosine, t ryptophane et ph6nyl-alanine dans les m~mes pro- portions que dans la mol4cule de fl-lactoglobuline (8:4:8 pour un poids mol6culaire de 35 500). On mesure ainsi la fraction des r6sidus de la prot6ine accessible au solvant perturbant par rapport au mod61e dans lequel IOO% des r6sidus sont totalement expos6s.
Aussi bien pour le mod61e que pour la prot6ine, l 'absorbance des solutions dans NaC1 o.I M ~ )l = 278-28o m#, est d'environ 1.3" IO 3, ~ pH 7, force ionique o.I, 20 °. Les concentrations en saccharose varient de o.I ~ I. 5 M. Les r6sultats sont exprim6s soit e n d~M,t/81~Ixmax - - rapport entre la difffrence des coefficients d 'absorption molaire deMx, et le coefficient d 'absorption molaire eM Xm~x. Les indices expriment les longueurs d'onde auxquelles les maxima du spectre de diff6renee et du spectre direct sont ob- serv6s (ce rapport est 6gal ~ z lA /A ; A = l 'absorbance). RM est le rapport des valeurs de AeMX/eMXmax pour la/~-laetoglobuline rapport6es ~ celles que l'on observe pour le m61ange mod61e dans les m~mes conditions. Les maxima ehoisis sont ceux de l 'ab-
Biochim. Biophys. Acta, 16o (1968) 340-359
356 j. CLEMENT-METRAL, J. YON
0.1
u) <3
/ /
/ /
/ /
/ b
:z : .................. 0 0.4 0 .8 1.2 5 ~ c c h ( ] r o s e concn . (M)
"t/ 0.5
0 0.4 0.8 1.2 Bc lcchc l rose concn . ( M )
Fig. 9. Var ia t ion du p a r a m ~ t r e de diffdrence spect ra l AeM291-2~ mu/eM2~s-2so mu en fonct ion de la con- c en t r a t i on molaire en saccharose , (a), pour le mdlange module d 'ac ides aminds : 3 " IO-4 M N-ac~tyl - L - t r y p t o p h a n e dthyles ter , 6. IO -~ M N-acdty l -L- tyros ine dthyles ter , 6. IO -~ M N-ac6ty l -L-ph6nyl - a lan ine dthyles ter , p H 7, force i o n i q u e o . I ; (b), f l- lactoglobuline A: o .27%, p H 7, force onii- que o. i .
Fig. io. Var ia t ion du r a p p o r t RM ( rappor t des va leurs de AeM,deM,~m~x) pour la f l - lactoglobuline et pour le mdlange d 'ac ides aminds mod6le dans les m&mes condi t ions que pour la Fig. 9, le pe r tu r - b a n t d t a n t soit le s&ccharose lu i -m6me (cercles pleins), soit le d i rnd thy l - su l foxyde A ~ o % ; en fonc t ion de la concen t r a t i on molaire en saccharose .
sorption due au tryptophane. En effet, pour un rapport tryptophane/(tryptophane ÷ tyrosine) de o.5 le tryptophane contribue au spectre de la tyrosine de fa$on telle qu'il n'est pas possible de savoir ~ la longueur d'onde du maximum d'absorption de la tyrosine, dans les spectres diff6rentiels, ce qui revient ~ l'un ou ~ l'autre de ces acides amin6s 4s.
La Fig. 9 nous montre la perturbation obtenue lorsque le solvant est le saccharose lui-mfime: elle est directement proportionnelle ~ la concentration du solvant pour le module, elle l'est aussi pour le fl-lactoglobuline A aux faibles et aux fortes valeurs de saccharose mais entre o.5 et 1.2 M une certaine proportion de tryptophane est venue en contact avec le solvant.
La prot6ine subit manifestement une transformation. Pour la mettre en ~vidence de fa~on plus nette la m~me s6rie d'exp6riences a fit6 faite en pr6sence de quantit6s croissantes de saccharose et d'une quantit6 fixe (2O~o) du solvant perturbant: di- m6thyl-sulfoxyde. Les r6sultats de ces exp6riences rassembl6s sur la Fig. IO expriment RM en fonction de la concentration en saccharose. Auss i bien les spectres, diff6rentiels ou directs, que les valeurs de RM, jusqu'~ o.2 Men sacCharose, sont parfaitement en accord avec les r6sultats de HERSKOVITS. Quelle que soit la concentration en saccharose celui-ci reste sans aucun effet sur les spectres de diff6rence des acides amin6s modules, obtenus par 2o% de dim6thyl-sulfoxyde; par contre, en admettant que RM = IOO% correspond 5. quatre tryptophane, on peut admettre que lorsque RM pour la prot6ine passe de 25 ~ 55% l'6quivalent d'une mol6cule de tryptophane s'est d~masqu6, ceci ne voulant pas dire pour autant qu'une mol6cule s'est d6masqu6e r6ellement en to- talitfi, l'effet pouvant fitre le m~me si plusieurs mol6cules se d6masquent partiellement. Nous retrouvons la m~me valeur de concentration en saccharose pour la demi-trans- formation de la lactoglobuline que pour la demi-inhibition de la prot6olyse de cette m~me prot~ine, 0.7 M. Ces r~sultats peuvent ~tre rapproch~s de ceux de WAHl. ET FREY 5° qui montrent que la forme de la courbe de ddpolarisation de fluorescence de la s~rum-albumine de boeuf conjug6e avec le i-dim~thylamino-naphtal~ne-5-sulfonyl,
Biochim. Biophys. Acta, 16o (1968) 340-359

fl-LACTOGLOBULINE A ET SACCHAROSE 357
obtenue par addition de saccharose, ne peut s'expliquer que si l'on admet que la prot6ine conjugu6e subit un changement de structure sous l'action du saccharose. N ous avons pu montrer par des considerations cin6tiques que la fixation de 2 molecules de saccharose par monom+re de fl-lactoglobuline A est li6e/~ une dissociation de cette prot~ine, on peut alors se demander quelle est la part de ces deux ph6nom+nes disso- ciation et changement de conformation dans la variation des spectres de diff6rence? I1 parait difficile d'admettre comme le font WETLAUFER ET LOVRIEN s que les modification du spectre ne sont dues qu'~ des modifications de conformation, puisque TIMASHEFF, TOWNEND ET MESCANTI 6 montrent que la r6gion de dim6risation contient des zones de structure organis6e en relation directe avec le contenu en tryptophane de la moldcule ; invoquer la seule dissociation est en contradiction avec les r6sultats des courbes de titrage. La fixation du saccharose entralnerait donc ~ la lois une dissociation de la fl-lactoglobuline A et un changement de conformation. Cette mani+re de voir nous paralt tr~s largement justifi~e par tout un ensemble de travaux. Les ~quilibres de dissociation ou d'association de la {/-lactoglobuline peuvent ~tre li6s ~ des trans- conformations. C'est ainsi que la transconformation observ6e par TANFORD 4 entre pH 6 et pH 9 (N ~ R) li6e ~ l'ionisation d'un carboxyle anormal par chalne, est associ6e ~ une dissociation (N~ ~ 2R) (bibl. 7). I1 est bien connu de plus que la fixation de nombreux ligands entraine des changements de la structure tertiaire ou quaternaire de la fl-lactoglobuline: les ions H (voir bibl. 5), les m6taux4, 51 des substances non polaires comme le m6thanol 6, ou les alkanesS, ~. TIMASHEFF, TOWNEND ET MESCANTI 6
observent entre pH 4 et pH 6 un changement de conformation associ6 pour la fl-lacto- globuline A ~ l'ionisation de 2 H+, alors que pour la fl-lactoglobuline B ou Cun seul est impliqu6 par mol6cule. Ace pH les fl-lactoglobulines 6tant des dim&res constitu~s de deux chaines identiques, la variation de la fl-lactoglobuline A en fonction du pH peut r6fl6ter soit une transition simultan6e cooperative int6ressant les deux chalnes, un groupe agissant dans chaque, soit une transition dans laquelle chaque chaine est ind6pendante de l'autre et dans laquelle 2 groupes sont affect6s par chalne, donnant un total de 4 groupes par unit6 de poids mol6culaire 35 500. Puisque dans les/3-1acto- globulines B e t C les deux chaines identiques subissent la transition ind6pendamment et que les trois lactoglobulines pr6sentent de grandes similitudes, il est raisonnable de conclure avec ces auteurs que la deuxi+me hypoth+se est la plus vraisemblable. WISH- NIA ET PINDER 9 sont conduit ~ la m~me conclusion ~ savoir que les chalnes identiques sont ind6pendantes ~ la lois dans le dim+re et dans l'octam+re (pour la/3-1actoglobuline A). Nos r6sultats pr6sentent une grande analogie, d6jh 6voqude par TANFORD 42 avec ce que l'on observe avec l'hdmoglobine des mammif+res : existence de 4 sites de fixation par dim~re; dissociation dim+re-monom+re; et changement de conformation li6 aux deux ph6nom+nes pr6c6dents 3z. I1 s 'y ajoute une inhibition de la prot6olyse. Cette inhibition probablement peu sp6cifique, se retrouve avec un demi-effet du m~me ordre de grandeur pour l'a-D-glucose et le maltose (fl-D-glucopyranosyl-4fl-n-glucopyrano- side), que pour le saccharose (a-n-glucopyranosyl-2fl-n-fructofuranoside) : 10. 5 -- 0. 7 M.
Le saccharose si largement employ6 comme une substance "inerte" dans les milieux biologiques entraine des alt6rations de structure dans une prot6ine globulaire comme la fl-lactoglobuline et le changement de conformation qui r6sulte de cette liaison est responsable de l'inhibition de la prot6olyse.
Biochim. Biophys. Acta, 16o (i968) 340-359

358 J. (;LE?dl~;NT-METRAI., J. YON
REMERCIEMENTS
Ce travail a fit~ effectu6 grace/~ une bourse de Recherches de l ' Inst i tut National de la Sant~ et de la Recherche M6dicale (I.N.S.E.R.M.).
I1 n 'aurai t pas vu le jour sans l'accueil bienveillant de Monsieur le Professeur TREMOLIERES, Directeur du Laboratoire de Nutrition Humaine de I 'I.N.S.E.R.M. /~ qui nous exprimons toute notre gratitude.
Nous sommes heureuses de remercier le Professeur TIMASHEFF des intdressantes discussions que nous avons pu avoir avec lui.
R~SUMg
Dans la premi6re partie de cet article nous avons montr6 que l'inhibition pro- duite par le saccharose sur la prot6olyse trypsique de la fi-lactoglobuline A (pH 7, 35 °) est due ~ la fixation du saccharose sur le substrat et ~ l'existence de fortes inter- actions stabilisatrices, au sein des mol6cules de celui-ci. L'analyse cin6tique, faite partir du sch6ma ~ deux &apes suivant :
E ~ - S , ~ E S - + E S " ~ E t~ P2 + P1
montre que l'inhibition, non comp&itive totale, ne se produit qu'avec un substrat prot6ique pour autant qu'il est natif et r6sulte de la liaison de deux mol&ules de saccharose par mol&ule de fl-lactoglobuline. Les sites de fixation sur cette derni~re ne sont ni identiques ni ind@endants. L'inhibition non compdtitive totale permet de remplacer l'expression log (v v d v d [vi, v = vitesse de la r6action inhibib6e ou non], par l'expression 6quivalente log ~//(I Y) [Y est la saturation fractionnelle de la mol6cule de fl-lactoglobuline par le saccharose]. La reprfsentation de HILL met alors en 6vidence l'existence non seulement d'interactions inter mais intra-mol6culaires. Ces derni6res correspondent 5~ une dissociation dim6re monom6re de la fl-lacto- globuline.
Ces r6sultats sont confirm6s par l'6tude thermodynamique. L 'augmentat ion de l'6nergie libre d'activation pour la demi-inhibition peut-@tre calcul6e, elle est de 4oo cal, alors que la valeur exp6rimentale minimum d'6nergie libre totale d'interaction saccharose fl-lactoglobuline correspondant ~ la saturation compl6te de la macro- molfcule par le ligand est de 76o cal. L'6tude thermodynamique indique d 'autre part, que les forces impliqu6es sont du type hydrophobe et correspondent ~ une variation d'entropie ndgative. Ceci sugg6re l'existence d'un changement de conformation, qui peut ~tre mis en 6vidence ind6pendamment de toute rfaction enzymatique par titrage et spectroscopie de diff6rence. Chaque chalne polypeptidique de fl-lactoglobuline est ind6pendante dans le dim&e, qui fixe par cons6quent 4 mol6cules de sucre.
I1 n'est pas sans int6r4t de noter que le saccharose si largement employ6 dans les 6tudes sur la structure des prot6ines n'est pas inerte et provoque un changement de conformation de la fl-lactoglobuline A, vers une structure plus compacte.
L'inhibition de la prot6olyse de la fi-lactoglobuline parait &re en relation directe avec ce changement de conformation puisque la concentration en saccharose qui produit la demi-conversion est la m~me (o.7 M) que celle qui entraine une inhibition de 5o%.
Biochin*. Biophys. Acla, 16o (1968) 34o-359

fl-LACTOGLOBULINE A ET SACCHAROSE 359
B I B L I O G R A P H I E
I J. YON, Biochim. Biophys. Acta, 27 (1958) I I I . 2 M. MONNOT, Th6se 3 e Cycle, Paris, 1963. 3 M. OUPONT, Thbse de Doctora t d ' E t a t 6s Sciences Physiques, Paris, 1964. 4 C. TANFORD, L. G. BUNVILLE ET Y. NOZAKI, J. Am. Chem. Soc., 81 (1959) 4o32 . 5 S. N. TIMASHEFF, L. MESCANTI, J. J. BASCH ET R. TOWNEND, J. Biol. Chem., 241 (i966) 2496. 6 S. N. TIMASHEFF, R. TOWNEND ET L. MESCANTI, J. Biol. Chem., 24I (1966) 1863. 7 D. PANTALONI, Th6se de Doctora t d ' E t a t 6s Sciences Physiques, Paris, I965. 8 D. B. \'VETLAUFER ET l~. LOVRIEN, J. Biol. Chem., 239 (1964) 596. 9 A. WTISHNIA ET T. ~V. PINDER, JR., Biochemistr% 5 (1966) I534.
io C. R. HARDT, I. F. HUDDELSON ET C. D. BALL, J. Biol. Chem., 163 (1946 ) 2 i i . I I A. BEILINSSON, Biochem. Z., 213 (1929) 299. 12 P. D. BUYER, F. G. LUM, C. A. BALLOU, J. M. LUCK ET R. G. RICE, J. Biol. Chem., 162 (1946)
181. 13 G. W. SCHXVERT, H. NEURATH, S. I~AUFMANN ET J. E. SNOKE, J. Biol. Chem., 172 (i948) 221. 14 A. "W. KENSHINGTON, darts P. ALEXANDER ET R. J. BLOCK, A Laboratory Manual of Analytical
Methods of Protein Chemistry, Vol. 2, 196o, p. 353- 15 C. TANFORD, Advan. Protein Chem., 17 (1962) 7 ° 166. 16 A. H. PALMER, J. Biol. Chem., lO 4 (1934) 359. 17 A. G. OGSTON ET D. CECIL, Biochem. J., 44 (1949) 33. I8 D. R. BRIGGS ET R. HULL, J. Am. Chem. Soc., 67 (1945) 2007. 19 J. YON, Bull. Soc. CAirn. Biol., 39 (19.57) 1163. 20 J. YON, Bull. Soc. CAirn. Biol., 42 (196o) 1263. 21 J. YON, Bull. Soc. CAirn. Biol., 42 (196o) 209. 22 M. L. BENDER, J. Ant. Chem. Soc., 84 (1962) 2582. 23 G. JOHANNIN, Th6se 3 e Cycle, Paris, 1966. 24 K. G. BRANDT ET G. P. HESS, Biochem. Biophys. Res. Commun., 22 (1966) 447- 25 A. BERNHARD, S. J. LAU ET H. NOLLER, Biochemislry, 4 (1965) 11o8. 26 T. E. BARMAN ET H. GUTFREUND, Proc. Natl. Acad. Sci. U.S., 53 (I965) 1243. 27 T. E. BARMAN ET I-I. GUTFREUND, Biochem. J., IOI (1966) 411. 28 J. CLEMENT-METRAL, J. YON ET J. TREMOLIERES, Compt. Rend., 256 (I963) 352o. 29 J. J. BECHET ET J. YON, Biochim. Biophys. Acta, 89 (1964) 117. 3 ° J. WYMAN, JR., Advan. Protein Chem., 4 (1948) 407 • 31 J. VVYMAN, JR., Advan. Protein Chem., 19 (1964) 223. 32 J. L. "vVEBB, Enzyme and Metabolic Inhibitors, Vol. i, Academic Press, New York, 1963. 33 M. MONNOT, H. WEINTRAUB ET J. YON, Bull. Soc. CAirn. Biol., 48 (1966) 959. 34 F. SEYDOUX, Th6se de Doctora t 3 e Cycle, Orsay, 1966. 35 J- M. REINER, Behavior of Enzyme Systems, Burgess, Minneapolis, Minn., 1959, p. 156. 36 A. V. HILL, J . Physiol., 4 ° (194 o) 4 p. 37 J. WITZ, S. N. TIMASHEFF ET V. LUZZATI, J. Am. Chem. Soc., 86 (1964) 168. 38 C. GEORGES ET S. GUINAND, J. Chim. Phys., 57 (196o) 606. 39 Y. NOZAKI, L. G. BUNVILLE ETC. TANFORD, J. Am. Chem. Soc., 81 (1959) 5.523. 4 ° R. K. CANNAN, A. H. PALMER ETA. C. KIBRICK, J. Biol. Chem., 142 (1942) 803. 41 C. W. CARR, Arch. Biochem. Biophys., 62 (1956) 476. 42 C. TANFORD ET Y. NOZAKI, J. Biol. Chem., 234 (1959) 2874. 43 H. P. BAKER ET H. A. SAROFF, Biochemistry, 4 (1965) 167o. 44 M. LASKOWSKI, JR. ET H. A. SHERAGA, J, Am. Chem. Soc., 76 (1954) 6305. 45 S. N. TIMASHEFF ET R. TOVCNEND, J. Am. Chem. Soc., 83 (1061) 47 o. 46 R. LOVRIEN, J. Am. Chem. Soc., 85 (1963) 3677. 47 C. TANFORD, J. Am. Chem. Soc., 79 (1957) 5348. 48 T. T. HERSKOVlTS ET M. LASKOWSKI, JR., J. Biol. Chem., 237 (1962) 2481. 49 T. T. HERSKOVlTS, J . Biol. Chem., 240 (1965) 628. 5 ° P. WAHL ET M. FREY, Compt. Rend., 262 S6rie D (1966) 2521. 51 G. PREAUX, N. BINON ET R. LONTIE, Acta Biol. Med. Ger., Suppl. 3 (1964) 277.
Biochim. Biophys. Acta, 16o (1968) 340-359