ethanol extracts from hemerocallis citrina attenuate the decreases of brain-derived neurotrophic...
TRANSCRIPT

Journal of Ethnopharmacology 144 (2012) 328–334
Contents lists available at SciVerse ScienceDirect
Journal of Ethnopharmacology
0378-87
http://d
Abbre
factor;
HCE, et
test; 5-n Corr
E-m
journal homepage: www.elsevier.com/locate/jep
Ethanol extracts from Hemerocallis citrina attenuate the decreasesof brain-derived neurotrophic factor, TrkB levels in rat induced bycorticosterone administration
Li-Tao Yi a,n, Jing Li a, Huo-Chen Li b, Ying Zhou b, Bi-Fang Su a, Ke-Feng Yang b,Meng Jiang b, Yan-Ting Zhang a
a Department of Chemical and Pharmaceutical Engineering, College of Chemical Engineering, Huaqiao University, Xiamen 361021, Fujian province, PR Chinab Department of Biotechnology and Bioengineering, College of Chemical Engineering, Huaqiao University, Xiamen 361021, Fujian province, PR China
a r t i c l e i n f o
Article history:
Received 5 April 2012
Received in revised form
27 August 2012
Accepted 6 September 2012Available online 18 September 2012
Keywords:
Hemerocallis citrina
Antidepressant-like effect
Corticosterone
Brain-derived neurotrophic factor
TrkB
41/$ - see front matter & 2012 Elsevier Irelan
x.doi.org/10.1016/j.jep.2012.09.016
viations: ANOVA, analysis of variance; BDNF
DA, dopamine; FST, forced swimming test; G
hanol extracts of Hemerocallis citrina; NA, nor
HT, serotonin; TST, tail suspension test
esponding author. Tel./fax: þ86 592 6162302
ail addresses: [email protected], litaoyi@hqu
a b s t r a c t
Ethnopharmacological relevance: Hemerocallis citrina, a traditional herbal medicine, has been used for
the improvement of behavioral and emotional status in Eastern-Asia countries.
Aim of the study: Our previous studies have demonstrated that the ethanol extracts of H. citrina flowers
(HCE) reversed the behavioral alterations and monoamine neurotransmitter dysfunctions in stressed
mice. However, the relation of its antidepressant-like action with neurotrophic molecular expressions
remains unknown.
Materials and methods: To clarify this, we explored the effect of HCE (32.5, 65, 130 mg/kg, p.o.) on the
behavior, brain-derived neurotrophic factor (BDNF) and its receptor (TrkB) in depression-like rats
induced by exogenous administration of the stress hormone corticosterone (40 mg/kg, s.c.).
Results: It was observed that repeated administration of corticosterone induced an elevation on the
serum corticosterone levels, which caused the abnormalities observed in the sucrose preference test
and forced swimming test (FST). Administration of HCE (65 and 130 mg/kg) reversed the changes above
and up-regulated the BDNF and TrkB receptor protein expressions in the brain region of frontal cortex
and hippocampus.
Conclusion: These findings confirm that HCE produce an antidepressant-like effect in corticosterone-
induced depression-like model of rats and this effect is at least partly mediated by BDNF-TrkB signaling
in the frontal cortex and hippocampus.
& 2012 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Hemerocallis citrina Baroni (Liliaceae) which is widely grown inChina, Japan and Korea has been used for food and medicinalpurposes for thousands of years. According to the phytochemicaland pharmacological studies, H. citrina mainly contains flavo-noids, polyphenols and essential oils that contribute to the herb’sbiological effects, such as antibacterial (Zhan et al., 2005), anti-oxidant (Lang and Luo, 2007) and nitrite-eliminating (Fu et al.,2009) activity. In addition, the effects of H. citrina such asrelieving gloom and improving sleeping were recorded in thefamous pharmaceutical text of ‘‘Compendium of Materia Medica’’,
d Ltd. All rights reserved.
, brain-derived neurotrophic
R, glucocorticoid receptor;
adrenaline; OFT, open-field
.
.edu.cn (L.-T. Yi).
the most complete and comprehensive medical book ever writtenin the history of traditional Chinese medicine. Clinically, H. citrina
has been widely used for the treatment of depressive disorders inChina (Chen et al., 2008).
In previous studies, we found that the ethanol extracts ofH. citrina (HCE) administration significantly reduced the immo-bility time in both the forced swimming test (FST) and tailsuspension test (TST) without accompanying changes in locomo-tor activity in the open-field test (OFT) in mice (Gu et al., 2012).Furthermore, we also demonstrated that HCE enhanced serotonin(5-HT) and noradrenaline (NA) levels in the frontal cortex andhippocampus as well as elevated dopamine (DA) levels in thefrontal cortex (Gu et al., 2012). However, until now, the neuro-trophic molecular expression underlying the antidepressant-likeeffect of HCE remains unclear.
Neurotrophic factors are critical regulators of the formationand plasticity of neuronal networks (Lee and Kim, 2010). Brain-derived neurotrophic factor (BDNF), one of important neuro-trophic factors, has also been implicated in the etiology of major
L.-T. Yi et al. / Journal of Ethnopharmacology 144 (2012) 328–334 329
depression and the mechanism of antidepressant treatment(Numakawa et al., 2010). Clinical study demonstrates that depres-sion is associated with reduced brain BDNF and its receptor (TrkB)levels and that the reductions can be restored up to the normalvalue by antidepressant treatment (Thompson et al., 2011).Animal studies have also demonstrated clear evidence that BDNFsignaling through TrkB is involved in the mechanisms of action ofantidepressant agents (Wang et al., 2010; Kutiyanawalla et al.,2011). BDNF and TrkB levels can therefore be useful markers forantidepressant-like response.
In the present study, we used a rat depression model by repeatedcorticosterone injections to investigate the antidepressant-like effectof HCE. Furthermore, the study explored whether the HCE couldreverse the loss of BDNF and its receptor TrkB mRNA and proteinlevels in the rat brain regions of frontal cortex and hippocampus.
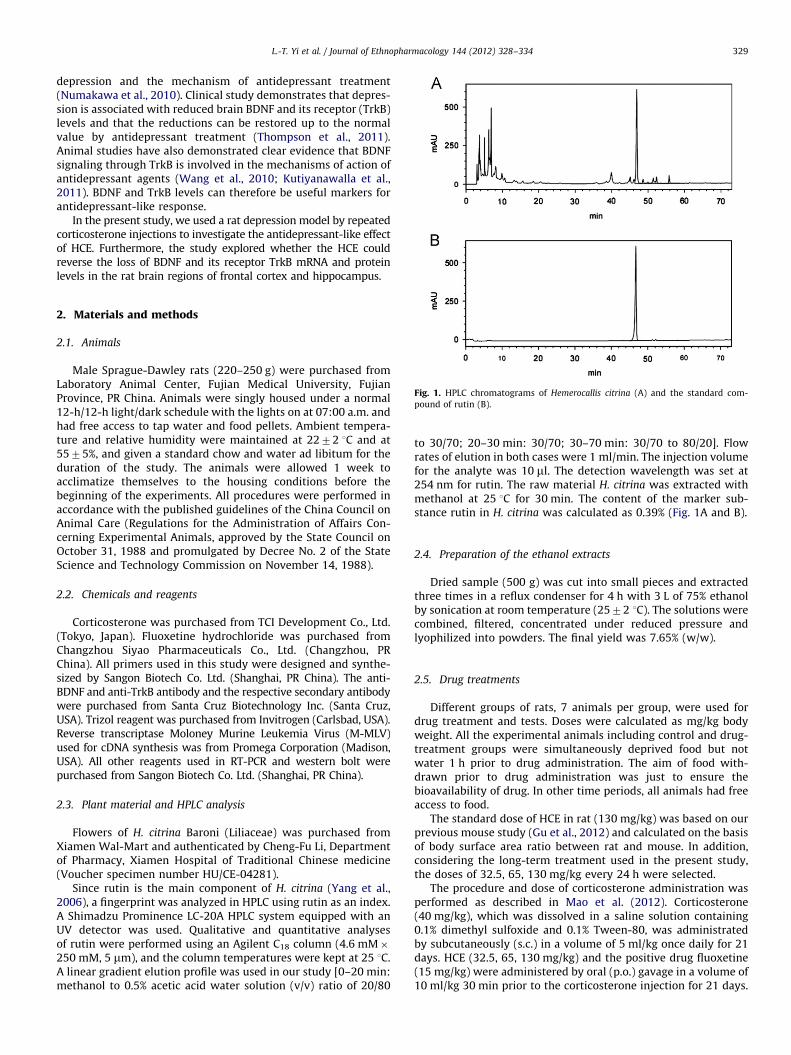
Fig. 1. HPLC chromatograms of Hemerocallis citrina (A) and the standard com-
pound of rutin (B).
2. Materials and methods
2.1. Animals
Male Sprague-Dawley rats (220–250 g) were purchased fromLaboratory Animal Center, Fujian Medical University, FujianProvince, PR China. Animals were singly housed under a normal12-h/12-h light/dark schedule with the lights on at 07:00 a.m. andhad free access to tap water and food pellets. Ambient tempera-ture and relative humidity were maintained at 2272 1C and at5575%, and given a standard chow and water ad libitum for theduration of the study. The animals were allowed 1 week toacclimatize themselves to the housing conditions before thebeginning of the experiments. All procedures were performed inaccordance with the published guidelines of the China Council onAnimal Care (Regulations for the Administration of Affairs Con-cerning Experimental Animals, approved by the State Council onOctober 31, 1988 and promulgated by Decree No. 2 of the StateScience and Technology Commission on November 14, 1988).
2.2. Chemicals and reagents
Corticosterone was purchased from TCI Development Co., Ltd.(Tokyo, Japan). Fluoxetine hydrochloride was purchased fromChangzhou Siyao Pharmaceuticals Co., Ltd. (Changzhou, PRChina). All primers used in this study were designed and synthe-sized by Sangon Biotech Co. Ltd. (Shanghai, PR China). The anti-BDNF and anti-TrkB antibody and the respective secondary antibodywere purchased from Santa Cruz Biotechnology Inc. (Santa Cruz,USA). Trizol reagent was purchased from Invitrogen (Carlsbad, USA).Reverse transcriptase Moloney Murine Leukemia Virus (M-MLV)used for cDNA synthesis was from Promega Corporation (Madison,USA). All other reagents used in RT-PCR and western bolt werepurchased from Sangon Biotech Co. Ltd. (Shanghai, PR China).
2.3. Plant material and HPLC analysis
Flowers of H. citrina Baroni (Liliaceae) was purchased fromXiamen Wal-Mart and authenticated by Cheng-Fu Li, Departmentof Pharmacy, Xiamen Hospital of Traditional Chinese medicine(Voucher specimen number HU/CE-04281).
Since rutin is the main component of H. citrina (Yang et al.,2006), a fingerprint was analyzed in HPLC using rutin as an index.A Shimadzu Prominence LC-20A HPLC system equipped with anUV detector was used. Qualitative and quantitative analysesof rutin were performed using an Agilent C18 column (4.6 mM�250 mM, 5 mm), and the column temperatures were kept at 25 1C.A linear gradient elution profile was used in our study [0–20 min:methanol to 0.5% acetic acid water solution (v/v) ratio of 20/80
to 30/70; 20–30 min: 30/70; 30–70 min: 30/70 to 80/20]. Flowrates of elution in both cases were 1 ml/min. The injection volumefor the analyte was 10 ml. The detection wavelength was set at254 nm for rutin. The raw material H. citrina was extracted withmethanol at 25 1C for 30 min. The content of the marker sub-stance rutin in H. citrina was calculated as 0.39% (Fig. 1A and B).
2.4. Preparation of the ethanol extracts
Dried sample (500 g) was cut into small pieces and extractedthree times in a reflux condenser for 4 h with 3 L of 75% ethanolby sonication at room temperature (2572 1C). The solutions werecombined, filtered, concentrated under reduced pressure andlyophilized into powders. The final yield was 7.65% (w/w).
2.5. Drug treatments
Different groups of rats, 7 animals per group, were used fordrug treatment and tests. Doses were calculated as mg/kg bodyweight. All the experimental animals including control and drug-treatment groups were simultaneously deprived food but notwater 1 h prior to drug administration. The aim of food with-drawn prior to drug administration was just to ensure thebioavailability of drug. In other time periods, all animals had freeaccess to food.
The standard dose of HCE in rat (130 mg/kg) was based on ourprevious mouse study (Gu et al., 2012) and calculated on the basisof body surface area ratio between rat and mouse. In addition,considering the long-term treatment used in the present study,the doses of 32.5, 65, 130 mg/kg every 24 h were selected.
The procedure and dose of corticosterone administration wasperformed as described in Mao et al. (2012). Corticosterone(40 mg/kg), which was dissolved in a saline solution containing0.1% dimethyl sulfoxide and 0.1% Tween-80, was administratedby subcutaneously (s.c.) in a volume of 5 ml/kg once daily for 21days. HCE (32.5, 65, 130 mg/kg) and the positive drug fluoxetine(15 mg/kg) were administered by oral (p.o.) gavage in a volume of10 ml/kg 30 min prior to the corticosterone injection for 21 days.

L.-T. Yi et al. / Journal of Ethnopharmacology 144 (2012) 328–334330
2.6. Sucrose preference test
The sucrose preference test was employed herein to evaluateanhedonia, one of the core symptoms of major depression inhumans. The test was performed as described by Mao et al.(2012). Briefly, the sucrose preference test was carried out 24 hafter the last drug treatment. Before the test, the rats were trainedto adapt to sucrose solution (1%, w/v) by placing two bottles ofsucrose solution in each cage for 24 h; then one of the bottles wasreplaced with water for 24 h. After the adaptation procedure, therats were deprived of water and food for 24 h. The sucrosepreference test was conducted at 10:00 a.m. The rats were housedin individual cages and given free access to the two bottlescontaining 100 ml of sucrose solution (1%, w/v) and 100 mlof water, respectively. After 60 min, the volume (ml) of both theconsumed sucrose solution and water were recorded, and sucrosepreference was calculated as sucrose preference (%)¼sucroseconsumption (ml)/[sucrose consumption (ml)þwater consump-tion (ml)]�100%.
2.7. Forced swimming test (FST)
The FST used was the same as described in detail elsewhere(Porsolt et al., 1978), with some modification. Briefly, the test wasdone by placing a rat in a glass cylinder (46 cm in height, 20 cm indiameter) filled with 30 cm high water (2572 1C). Water wasreplaced between every trial. Two swimming sessions wereconducted: an initial 15 min pretest, followed by a 5 min test24 h later. A rat was considered immobile whenever it remainedfloating passively in the water and only making slight movementsto keep its head above the water line. The test sessions wererecorded by a video camera and scored by an observer blind totreatment.
2.8. Blood sampling and tissue extraction
Animals were killed by decapitation one day after FST. Toavoid fluctuations on hormone levels due to circadian rhythm,animals were bled at 12:00–13:00 p.m. on the day of sacrifice. Thebrain regions of frontal cortex and hippocampus were isolatedimmediately, and then stored at �80 1C for later analysis of BDNFand TrkB mRNA and protein levels.
2.9. Serum corticosterone assay
Blood was collected on ice and separated in a refrigeratedcentrifuge at 4 1C (4000� g for 10 min). Serum was stored at –20 1C until assays were performed. Serum corticosterone levelswere measured using a commercial kit (Enzo Life Sciences,Plymouth Meeting, USA) based on enzyme immunoassay(Crowther, 1995).
2.10. RT-PCR
The procedure of RT-PCR was performed as described in detailelsewhere (Prediger, 2001; Pan et al., 2010), with some modifica-tions. Total RNA was isolated from frontal cortex or hippocampususing Trizol reagent following the manufacturer’s instructions.The concentration and purity of RNA were measured by theoptical density at 260 and 280 nm using spectrophotometer.Reverse transcription was performed with 1 mg RNA using M-MLV reverse transcriptase for cDNA synthesis. Amplification ofcDNA by PCR was performed in 25 ml reactions containing 8 mlcDNA, 1 ml forward and 1 ml reverse primers (10 mm), 2.5 mlPCR�10 buffer containing MgCl2, 0.5 ml dNTP mixture (10mM),0.5 ml Taq polymerase (2.5 U) and 11.5 ml sterile ddH2O. Primers
used for BDNF was: 50-TGTGACAGTATTAGCGAGTGGGT-30 and50-CGATTGGGTAGTTCGGCATT-30, that used for TrkB was50-CTTATGCTTGCTGGTCTTGG-30 and 50-GGGTATTCTTGCTGCTC-TCA-30. In general, PCR was performed with a preheating cycleat 94 1C for 5 min, denaturation, annealing and elongation werecarried out at 94 1C for 30 s, at 50 1C for 30 s (BDNF) or 59 1C for60 s (TrkB), and at 68 1C for 15 s (BDNF) or 72 1C for 35 s (TrkB),respectively. The reactions were repeated for 40 (BDNF) or 38(TrkB) cycles. The PCR products were resolved by a 1.5% agarosegel electrophoresis and quantified by the Bio-Rad ChemiDoc XRSGel Documentation system and Bio-Rad Quantity One software(Hercules, USA). The results were normalized to the mRNAexpression level of GAPDH in each sample.
2.11. Western blot
The procedure of western blot was performed as described indetail elsewhere (Blancher and Jones, 2001; Chen et al., 2010),with some modifications. Brain samples were homogenized ina lysis buffer containing 50 mM Tris–HCl (pH7.4), 1 mM EDTA,150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1%SDS, 1 mM trichostatin A, phosphatase inhibitor cocktail. Thehomogenates were centrifuged at 14000� g for 20 min at 4 1C,and the supernatants were collected and stored at �20 1C untilfurther use. The protein concentration was determined by a BCAassay. Total proteins were separated by SDS-PAGE and transferredto a PVDF membrane. Following blocking in 3% BSA/TBST at roomtemperature for 1 h, the membranes were incubated with theappropriate primary antibodies at 4 1C overnight (anti-BDNF:1:500, anti-TrkB: 1:1000). After being washed with TBST for threetimes, the membranes were incubated with an HRP-labeledsecondary antibody (1:4000). The blots were washed again forthree times by TBST buffer and the immunoreactive bands weredetected by using the enhanced chemiluminescence method.Western blot bands were scanned and subsequently analyzeddensitometrically with Bio-Rad Quantity One software (Hercules,USA). The results were normalized to the protein expression levelof GAPDH in each sample.
2.12. Statistical analyses
All data were expressed as mean7S.E.M. To compare experi-mental and control groups, we used one-way ANOVA, followed bypost-hoc Dunnett’s test. A value of Po0.05 was consideredstatistically significant for analysis.
3. Results
3.1. Effects of HCE on the sucrose preference
The effects of HCE on the sucrose preference in rats exposed torepeated corticosterone administration were given in Fig. 2.Corticosterone administration induced a significant decrease inthe sucrose preference (Po0.05). 65 and 130 mg/kg HCE reversedthe reduction to the baseline value (Po0.05, Po0.01, respec-tively). Fluoxetine, the positive drug, also increased the sucrosepreference in the rat exposed to corticosterone (Po0.05).
3.2. Effects of HCE on the immobility time
Repeated corticosterone treatment (s.c.) caused a significantelevation of immobility time in the rat FST compared with thecontrols (Po0.05). Long-term treatment with HCE (65 and130 mg/kg) and fluoxetine (15 mg/kg) for 21 days significantly
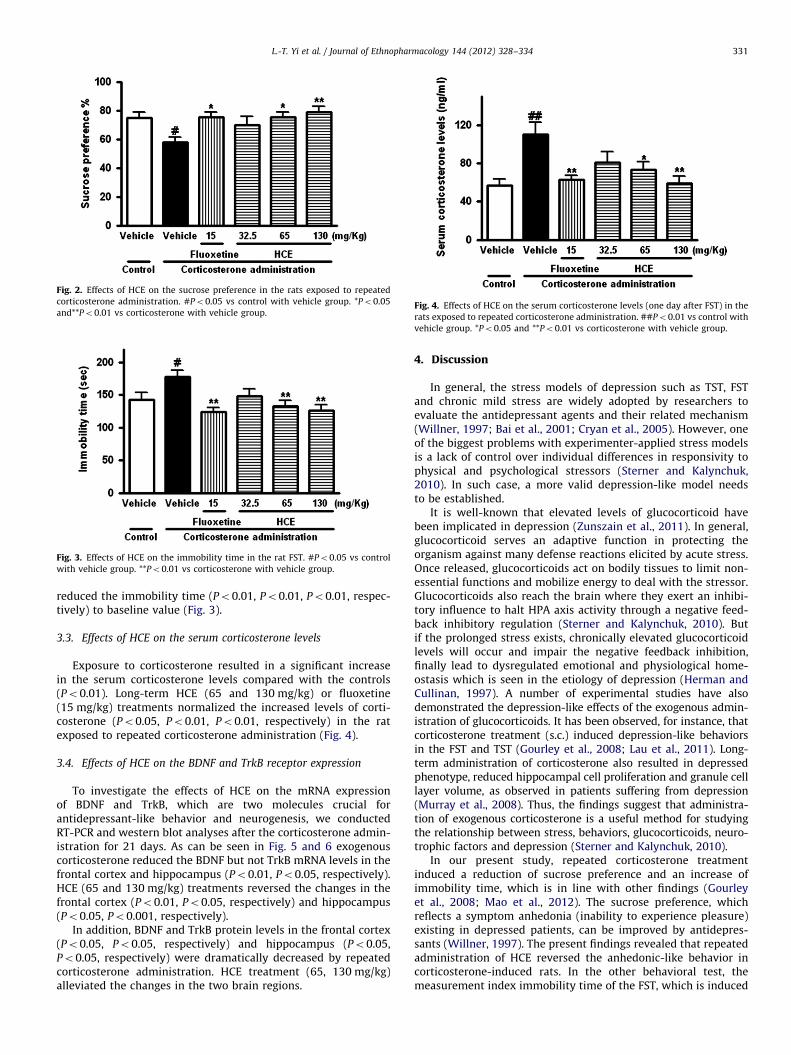
Fig. 3. Effects of HCE on the immobility time in the rat FST. #Po0.05 vs control
with vehicle group. **Po0.01 vs corticosterone with vehicle group.
Fig. 4. Effects of HCE on the serum corticosterone levels (one day after FST) in the
rats exposed to repeated corticosterone administration. ##Po0.01 vs control with
vehicle group. *Po0.05 and **Po0.01 vs corticosterone with vehicle group.
Fig. 2. Effects of HCE on the sucrose preference in the rats exposed to repeated
corticosterone administration. #Po0.05 vs control with vehicle group. *Po0.05
and**Po0.01 vs corticosterone with vehicle group.
L.-T. Yi et al. / Journal of Ethnopharmacology 144 (2012) 328–334 331
reduced the immobility time (Po0.01, Po0.01, Po0.01, respec-tively) to baseline value (Fig. 3).
3.3. Effects of HCE on the serum corticosterone levels
Exposure to corticosterone resulted in a significant increasein the serum corticosterone levels compared with the controls(Po0.01). Long-term HCE (65 and 130 mg/kg) or fluoxetine(15 mg/kg) treatments normalized the increased levels of corti-costerone (Po0.05, Po0.01, Po0.01, respectively) in the ratexposed to repeated corticosterone administration (Fig. 4).
3.4. Effects of HCE on the BDNF and TrkB receptor expression
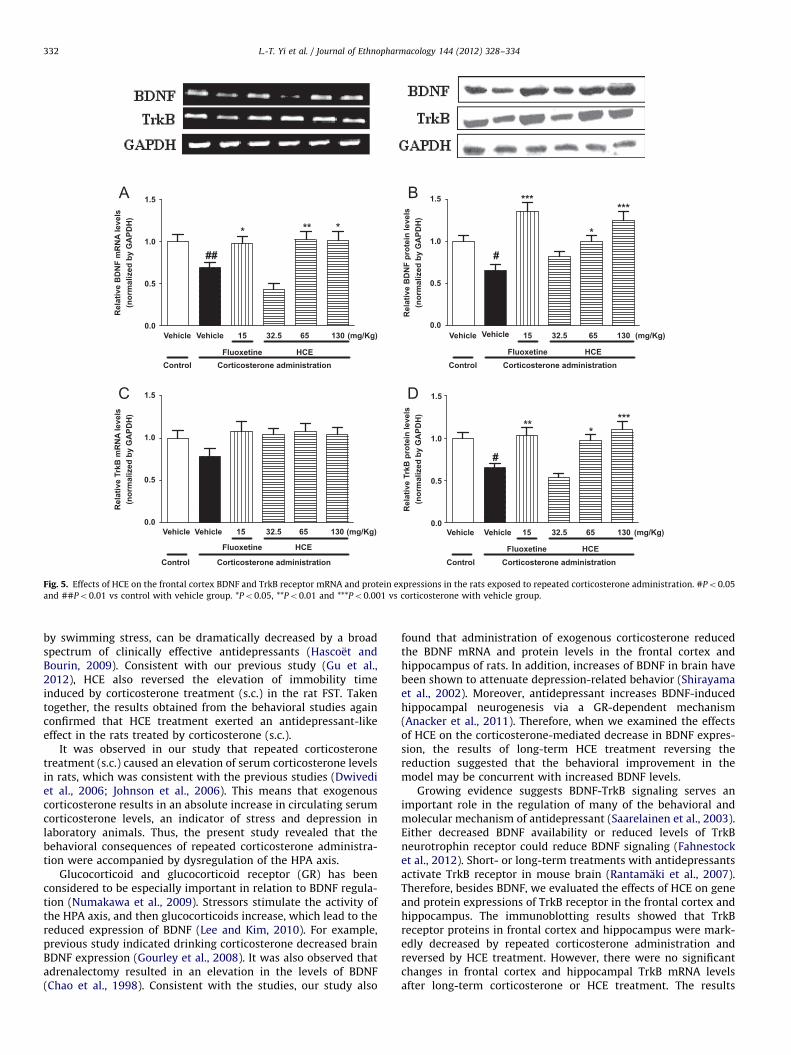
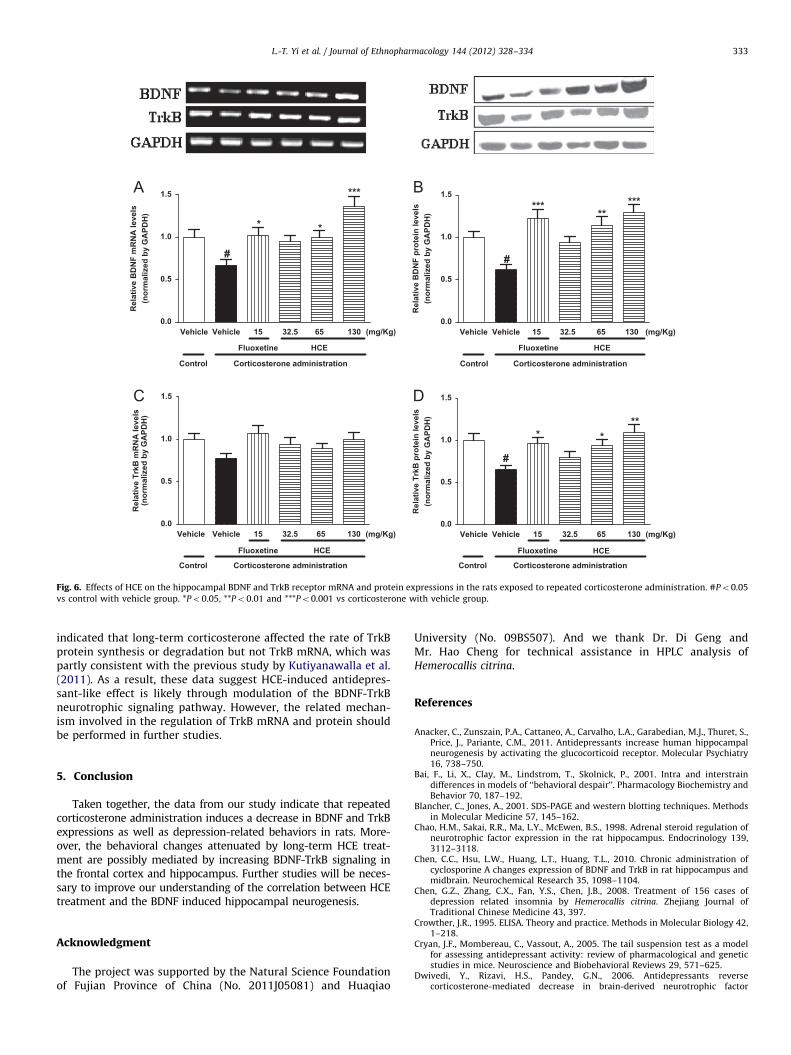
To investigate the effects of HCE on the mRNA expressionof BDNF and TrkB, which are two molecules crucial forantidepressant-like behavior and neurogenesis, we conductedRT-PCR and western blot analyses after the corticosterone admin-istration for 21 days. As can be seen in Fig. 5 and 6 exogenouscorticosterone reduced the BDNF but not TrkB mRNA levels in thefrontal cortex and hippocampus (Po0.01, Po0.05, respectively).HCE (65 and 130 mg/kg) treatments reversed the changes in thefrontal cortex (Po0.01, Po0.05, respectively) and hippocampus(Po0.05, Po0.001, respectively).
In addition, BDNF and TrkB protein levels in the frontal cortex(Po0.05, Po0.05, respectively) and hippocampus (Po0.05,Po0.05, respectively) were dramatically decreased by repeatedcorticosterone administration. HCE treatment (65, 130 mg/kg)alleviated the changes in the two brain regions.
4. Discussion
In general, the stress models of depression such as TST, FSTand chronic mild stress are widely adopted by researchers toevaluate the antidepressant agents and their related mechanism(Willner, 1997; Bai et al., 2001; Cryan et al., 2005). However, oneof the biggest problems with experimenter-applied stress modelsis a lack of control over individual differences in responsivity tophysical and psychological stressors (Sterner and Kalynchuk,2010). In such case, a more valid depression-like model needsto be established.
It is well-known that elevated levels of glucocorticoid havebeen implicated in depression (Zunszain et al., 2011). In general,glucocorticoid serves an adaptive function in protecting theorganism against many defense reactions elicited by acute stress.Once released, glucocorticoids act on bodily tissues to limit non-essential functions and mobilize energy to deal with the stressor.Glucocorticoids also reach the brain where they exert an inhibi-tory influence to halt HPA axis activity through a negative feed-back inhibitory regulation (Sterner and Kalynchuk, 2010). Butif the prolonged stress exists, chronically elevated glucocorticoidlevels will occur and impair the negative feedback inhibition,finally lead to dysregulated emotional and physiological home-ostasis which is seen in the etiology of depression (Herman andCullinan, 1997). A number of experimental studies have alsodemonstrated the depression-like effects of the exogenous admin-istration of glucocorticoids. It has been observed, for instance, thatcorticosterone treatment (s.c.) induced depression-like behaviorsin the FST and TST (Gourley et al., 2008; Lau et al., 2011). Long-term administration of corticosterone also resulted in depressedphenotype, reduced hippocampal cell proliferation and granule celllayer volume, as observed in patients suffering from depression(Murray et al., 2008). Thus, the findings suggest that administra-tion of exogenous corticosterone is a useful method for studyingthe relationship between stress, behaviors, glucocorticoids, neuro-trophic factors and depression (Sterner and Kalynchuk, 2010).
In our present study, repeated corticosterone treatmentinduced a reduction of sucrose preference and an increase ofimmobility time, which is in line with other findings (Gourleyet al., 2008; Mao et al., 2012). The sucrose preference, whichreflects a symptom anhedonia (inability to experience pleasure)existing in depressed patients, can be improved by antidepres-sants (Willner, 1997). The present findings revealed that repeatedadministration of HCE reversed the anhedonic-like behavior incorticosterone-induced rats. In the other behavioral test, themeasurement index immobility time of the FST, which is induced
0.0
0.5
1.0
1.5
*** *
HCE
##
Vehicle Vehicle 15 32.5 65 130
FluoxetineControl Corticosterone administration
Rel
ativ
e B
DN
F m
RN
A le
vels
(nor
mal
ized
by
GA
PDH
)
0.0
0.5
1.0
1.5
(mg/Kg)(mg/Kg)
(mg/Kg)
*
******
HCE
#
Vehicle Vehicle 15 32.5 65 130
FluoxetineControl Corticosterone administration
Rel
ativ
e B
DN
F pr
otei
n le
vels
(nor
mal
ized
by
GA
PDH
)
0.0
0.5
1.0
1.5
HCE
Vehicle Vehicle 15 32.5 65 130
Fluoxetine
Control Corticosterone administration
Rel
ativ
e Tr
kB m
RN
A le
vels
(nor
mal
ized
by
GA
PDH
)
0.0
0.5
1.0
1.5
(mg/Kg)
*** ***
HCE
#
Vehicle Vehicle 15 32.5 65 130
FluoxetineControl Corticosterone administration
Rel
ativ
e Tr
kB p
rote
in le
vels
(nor
mal
ized
by
GA
PDH
)
Fig. 5. Effects of HCE on the frontal cortex BDNF and TrkB receptor mRNA and protein expressions in the rats exposed to repeated corticosterone administration. #Po0.05
and ##Po0.01 vs control with vehicle group. *Po0.05, **Po0.01 and ***Po0.001 vs corticosterone with vehicle group.
L.-T. Yi et al. / Journal of Ethnopharmacology 144 (2012) 328–334332
by swimming stress, can be dramatically decreased by a broadspectrum of clinically effective antidepressants (Hascoet andBourin, 2009). Consistent with our previous study (Gu et al.,2012), HCE also reversed the elevation of immobility timeinduced by corticosterone treatment (s.c.) in the rat FST. Takentogether, the results obtained from the behavioral studies againconfirmed that HCE treatment exerted an antidepressant-likeeffect in the rats treated by corticosterone (s.c.).
It was observed in our study that repeated corticosteronetreatment (s.c.) caused an elevation of serum corticosterone levelsin rats, which was consistent with the previous studies (Dwivediet al., 2006; Johnson et al., 2006). This means that exogenouscorticosterone results in an absolute increase in circulating serumcorticosterone levels, an indicator of stress and depression inlaboratory animals. Thus, the present study revealed that thebehavioral consequences of repeated corticosterone administra-tion were accompanied by dysregulation of the HPA axis.
Glucocorticoid and glucocorticoid receptor (GR) has beenconsidered to be especially important in relation to BDNF regula-tion (Numakawa et al., 2009). Stressors stimulate the activity ofthe HPA axis, and then glucocorticoids increase, which lead to thereduced expression of BDNF (Lee and Kim, 2010). For example,previous study indicated drinking corticosterone decreased brainBDNF expression (Gourley et al., 2008). It was also observed thatadrenalectomy resulted in an elevation in the levels of BDNF(Chao et al., 1998). Consistent with the studies, our study also
found that administration of exogenous corticosterone reducedthe BDNF mRNA and protein levels in the frontal cortex andhippocampus of rats. In addition, increases of BDNF in brain havebeen shown to attenuate depression-related behavior (Shirayamaet al., 2002). Moreover, antidepressant increases BDNF-inducedhippocampal neurogenesis via a GR-dependent mechanism(Anacker et al., 2011). Therefore, when we examined the effectsof HCE on the corticosterone-mediated decrease in BDNF expres-sion, the results of long-term HCE treatment reversing thereduction suggested that the behavioral improvement in themodel may be concurrent with increased BDNF levels.
Growing evidence suggests BDNF-TrkB signaling serves animportant role in the regulation of many of the behavioral andmolecular mechanism of antidepressant (Saarelainen et al., 2003).Either decreased BDNF availability or reduced levels of TrkBneurotrophin receptor could reduce BDNF signaling (Fahnestocket al., 2012). Short- or long-term treatments with antidepressantsactivate TrkB receptor in mouse brain (Rantamaki et al., 2007).Therefore, besides BDNF, we evaluated the effects of HCE on geneand protein expressions of TrkB receptor in the frontal cortex andhippocampus. The immunoblotting results showed that TrkBreceptor proteins in frontal cortex and hippocampus were mark-edly decreased by repeated corticosterone administration andreversed by HCE treatment. However, there were no significantchanges in frontal cortex and hippocampal TrkB mRNA levelsafter long-term corticosterone or HCE treatment. The results
0.0
0.5
1.0
1.5
(mg/Kg)
**
***
HCE
#
Vehicle Vehicle 15 32.5 65 130
Fluoxetine
Control
Rel
ativ
e B
DN
F m
RN
A le
vels
(nor
mal
ized
by
GA
PDH
)
0.0
0.5
1.0
1.5
(mg/Kg)
***** ***
HCE
#
Vehicle Vehicle 15 32.5 65 130
Fluoxetine
Control
Rel
ativ
e B
DN
F pr
otei
n le
vels
(nor
mal
ized
by
GA
PDH
)
0.0
0.5
1.0
1.5
(mg/Kg)
HCE
Vehicle Vehicle 15 32.5 65 130
Fluoxetine
Control
Rel
ativ
e Tr
kB m
RN
A le
vels
(nor
mal
ized
by
GA
PDH
)
0.0
0.5
1.0
1.5
(mg/Kg)
****
HCE
#
Vehicle Vehicle 15 32.5 65 130
Fluoxetine
Control
Rel
ativ
e Tr
kB p
rote
in le
vels
(nor
mal
ized
by
GA
PDH
)
Corticosterone administration Corticosterone administration
Corticosterone administration Corticosterone administration
Fig. 6. Effects of HCE on the hippocampal BDNF and TrkB receptor mRNA and protein expressions in the rats exposed to repeated corticosterone administration. #Po0.05
vs control with vehicle group. *Po0.05, **Po0.01 and ***Po0.001 vs corticosterone with vehicle group.
L.-T. Yi et al. / Journal of Ethnopharmacology 144 (2012) 328–334 333
indicated that long-term corticosterone affected the rate of TrkBprotein synthesis or degradation but not TrkB mRNA, which waspartly consistent with the previous study by Kutiyanawalla et al.(2011). As a result, these data suggest HCE-induced antidepres-sant-like effect is likely through modulation of the BDNF-TrkBneurotrophic signaling pathway. However, the related mechan-ism involved in the regulation of TrkB mRNA and protein shouldbe performed in further studies.
5. Conclusion
Taken together, the data from our study indicate that repeatedcorticosterone administration induces a decrease in BDNF and TrkBexpressions as well as depression-related behaviors in rats. More-over, the behavioral changes attenuated by long-term HCE treat-ment are possibly mediated by increasing BDNF-TrkB signaling inthe frontal cortex and hippocampus. Further studies will be neces-sary to improve our understanding of the correlation between HCEtreatment and the BDNF induced hippocampal neurogenesis.
Acknowledgment
The project was supported by the Natural Science Foundationof Fujian Province of China (No. 2011J05081) and Huaqiao
University (No. 09BS507). And we thank Dr. Di Geng andMr. Hao Cheng for technical assistance in HPLC analysis ofHemerocallis citrina.
References
Anacker, C., Zunszain, P.A., Cattaneo, A., Carvalho, L.A., Garabedian, M.J., Thuret, S.,Price, J., Pariante, C.M., 2011. Antidepressants increase human hippocampalneurogenesis by activating the glucocorticoid receptor. Molecular Psychiatry16, 738–750.
Bai, F., Li, X., Clay, M., Lindstrom, T., Skolnick, P., 2001. Intra and interstraindifferences in models of ‘‘behavioral despair’’. Pharmacology Biochemistry andBehavior 70, 187–192.
Blancher, C., Jones, A., 2001. SDS-PAGE and western blotting techniques. Methodsin Molecular Medicine 57, 145–162.
Chao, H.M., Sakai, R.R., Ma, L.Y., McEwen, B.S., 1998. Adrenal steroid regulation ofneurotrophic factor expression in the rat hippocampus. Endocrinology 139,3112–3118.
Chen, C.C., Hsu, L.W., Huang, L.T., Huang, T.L., 2010. Chronic administration ofcyclosporine A changes expression of BDNF and TrkB in rat hippocampus andmidbrain. Neurochemical Research 35, 1098–1104.
Chen, G.Z., Zhang, C.X., Fan, Y.S., Chen, J.B., 2008. Treatment of 156 cases ofdepression related insomnia by Hemerocallis citrina. Zhejiang Journal ofTraditional Chinese Medicine 43, 397.
Crowther, J.R., 1995. ELISA. Theory and practice. Methods in Molecular Biology 42,1–218.
Cryan, J.F., Mombereau, C., Vassout, A., 2005. The tail suspension test as a modelfor assessing antidepressant activity: review of pharmacological and geneticstudies in mice. Neuroscience and Biobehavioral Reviews 29, 571–625.
Dwivedi, Y., Rizavi, H.S., Pandey, G.N., 2006. Antidepressants reversecorticosterone-mediated decrease in brain-derived neurotrophic factor

L.-T. Yi et al. / Journal of Ethnopharmacology 144 (2012) 328–334334
expression: differential regulation of specific exons by antidepressants andcorticosterone. Neuroscience 139, 1017–1029.
Fahnestock, M., Marchese, M., Head, E., Pop, V., Michalski, B., Milgram, W.N.,Cotman, C.W., 2012. BDNF increases with behavioral enrichment and anantioxidant diet in the aged dog. Neurobiology of Aging 33, 546–554.
Fu, M.R., Chen, Q.M., Mao, L.C., 2009. Suppression of nitrosation by extracts ofnightlily (Hemerocallis citrina) flowers. Food Science 30, 114–119.
Gourley, S.L., Wu, F.J., Kiraly, D.D., Ploski, J.E., Kedves, A.T., Duman, R.S., Taylor, J.R.,2008. Regionally specific regulation of ERK MAP kinase in a model of
antidepressant-sensitive chronic depression. Biological Psychiatry 63,353–359.
Gu, L., Liu, Y.J., Wang, Y.B., Yi, L.T., 2012. Role for monoaminergic systems in theantidepressant-like effect of ethanol extracts from Hemerocallis citrina. Journalof Ethnopharmacology 139, 780–787.
Hascoet, M., Bourin, M., 2009. The forced swimming test in mice: a suitable modelto study antidepressants. In: Gould, T.D. (Ed.), Mood and Anxiety Related
Phenotypes in Mice. Neuromethods. Humana Press, New York, vol. 42, pp. 85–118.
Herman, J.P., Cullinan, W.E., 1997. Neurocircuitry of stress: central control of thehypothalamo-pituitary-adrenocortical axis. Trends in Neurosciences 20,78–84.
Johnson, S.A., Fournier, N.M., Kalynchuk, L.E., 2006. Effect of different doses ofcorticosterone on depression-like behavior and HPA axis responses to a novel
stressor. Behavioural Brain Research 168, 280–288.Kutiyanawalla, A., Terry Jr., A.V., Pillai, A., 2011. Cysteamine attenuates the
decreases in TrkB protein levels and the anxiety/depression-like behaviors inmice induced by corticosterone treatment. PLoS One 6, e26153.
Lang, N., Luo, H.X., 2007. Study for the activity of flavonoid in Hemerocallis citrina.Food Research and Development 28, 74–77.
Lau, B.W., Yau, S.Y., Lee, T.M., Ching, Y.P., Tang, S.W., So, K.F., 2011. Effect ofcorticosterone and paroxetine on masculine mating behavior: possible invol-vement of neurogenesis. Journal of Sexual Medicine 8, 1390–1403.
Lee, B.H., Kim, Y.K., 2010. The roles of BDNF in the pathophysiology of majordepression and in antidepressant treatment. Psychiatry Investigation 7,
231–235.Mao, Q.Q., Huang, Z., Ip, S.P., Xian, Y.F., Che, C.T., 2012. Peony glycosides reverse
the effects of corticosterone on behavior and brain BDNF expression in rats.Behavioural Brain Research 227, 305–309.
Murray, F., Smith, D.W., Hutson, P.H., 2008. Chronic low dose corticosteroneexposure decreased hippocampal cell proliferation, volume and inducedanxiety and depression like behaviours in mice. European Journal of Pharma-
cology 583, 115–127.Numakawa, T., Kumamaru, E., Adachi, N., Yagasaki, Y., Izumi, A., Kunugi, H., 2009.
Glucocorticoid receptor interaction with TrkB promotes BDNF-triggered PLC-gamma signaling for glutamate release via a glutamate transporter. Proceed-ings of the National Academy of Sciences 106, 647–652.
Numakawa, T., Suzuki, S., Kumamaru, E., Adachi, N., Richards, M., Kunugi, H., 2010.BDNF function and intracellular signaling in neurons. Histology and Histo-pathology 25, 237–258.
Pan, M., Li, Z., Yeung, V., Xu, R.J., 2010. Dietary supplementation of soy germphytoestrogens or estradiol improves spatial memory performance andincreases gene expression of BDNF, TrkB receptor and synaptic factors inovariectomized rats. Nutrition and Metabolism 7, 75.
Porsolt, R.D., Anton, G., Blavet, N., Jalfre, M., 1978. Behavioural despair in rats:a new model sensitive to antidepressant treatments. European Journal ofPharmacology 47, 379–391.
Prediger, E.A., 2001. Quantitating mRNAs with relative and competitive RT-PCR.Methods in Molecular Biology 160, 49–63.
Rantamaki, T., Hendolin, P., Kankaanpaa, A., Mijatovic, J., Piepponen, P., Domenici, E.,Chao, M.V., Mannisto, P.T., Castren, E., 2007. Pharmacologically diverse anti-depressants rapidly activate brain-derived neurotrophic factor receptor TrkBand induce phospholipase-Cgamma signaling pathways in mouse brain. Neu-ropsychopharmacology 32, 2152–2162.
Saarelainen, T., Hendolin, P., Lucas, G., Koponen, E., Sairanen, M., MacDonald, E.,Agerman, K., Haapasalo, A., Nawa, H., Aloyz, R., Ernfors, P., Castren, E., 2003.Activation of the TrkB neurotrophin receptor is induced by antidepressantdrugs and is required for antidepressant-induced behavioral effects. Journal ofNeuroscience 23, 349–357.
Shirayama, Y., Chen, A.C., Nakagawa, S., Russell, D.S., Duman, R.S., 2002. Brain-derived neurotrophic factor produces antidepressant effects in behavioralmodels of depression. Journal of Neuroscience 22, 3251–3261.
Sterner, E.Y., Kalynchuk, L.E., 2010. Behavioral and neurobiological consequencesof prolonged glucocorticoid exposure in rats: relevance to depression. Progressin Neuro-Psychopharmacology and Biological Psychiatry 34, 777–790.
Thompson, Ray.M., Weickert, C.S., Wyatt, E., Webster, M.J., 2011. Decreased BDNF,trkB-TKþ and GAD67 mRNA expression in the hippocampus of individualswith schizophrenia and mood disorders. Journal of Psychiatry and Neu-roscience 36, 195–203.
Wang, R., Li, Y.H., Xu, Y., Li, Y.B., Wu, H.L., Guo, H., Zhang, J.Z., Zhang, J.J., Pan, X.Y.,Li, X.J., 2010. Curcumin produces neuroprotective effects via activating brain-derived neurotrophic factor/TrkB-dependent MAPK and PI-3K cascades inrodent cortical neurons. Progress in Neuro-Psychopharmacology and Biologi-cal Psychiatry 34, 147–153.
Willner, P., 1997. Validity, reliability and utility of the chronic mild stress model ofdepression: a 10-year review and evaluation. Psychopharmacology 134,319–329.
Yang, Q., Tang, R.R., Ren, F.L., 2006. Determination on flavonoids from HemerocallisCitrina baroni. Food Science 9, 235–236.
Zhan, L.S., Li, G.R., Li, S.D., Wu, S.Y., 2005. Study on extraction of flavonids inHemerocallis Citrina Baroni and its medical effects. Journal of Nanhua Uni-versity (Medical Edition) 33, 112–114.
Zunszain, P.A., Anacker, C., Cattaneo, A., Carvalho, L.A., Pariante, C.M., 2011.Glucocorticoids, cytokines and brain abnormalities in depression. Progress inNeuro-Psychopharmacology and Biological Psychiatry 35, 722–729.