erythropoietin induces raf-1 activation and raf-1 is required for
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 266, No. 23, Issue of August 15, pp. 14964-14969, 1991 Printed in U. S. A.
Erythropoietin Induces Raf-1 Activation and Raf-1 Is Required for Erythropoietin-mediated Proliferation*
(Received for publication, May 20, 1991)
Michael P. Carroll$$, Jerry L. Spivakll, Marilyn McMahonII , Nadine WeichII , Ulf R. Rapp**, and W. Stratford May$ $$ From the $Johns Hopkins Oncology Center and the TDepartment of Medicine, Division of Hematology, Johns Hopkins University School of Medicine, Baltimore, Maryland 21231, the IIR. W. Johnson Pharmaceutical Research Institute, Johnson and Johnson, Raritan, New Jersey 08869, and the **Laboratory of Viral Carcinogenesis, National Cancer Institute, National Institutes of Health, Frederick Cancer Research and Developmental Center, Frederick, Maryland 21 702
Erythropoietin mediates the rapid phosphorylation of Raf-1 in the murine cell lines HCD-57 and FDC-Pl/ ER, which proliferate in response to this cytokine. Phosphorylation occurs at both serine and tyrosine residues and as such is similar to the Raf-1 phosphoryl- ation seen after interleukin-3 (IL-3), granulocyte-mac- rophage colony-stimulating factor, and interleukin-2 stimulation in other murine cell lines. Such data sug- gest that these growth factors may share a common mechanism(s) of Raf-1 phosphorylation. Furthermore, in association with Raf-1 phosphorylation, erythropo- ietin induces a 2-3-fold increase in Raf-1 kinase activ- ity as measured in immune complex kinase assays in vitro. Finally, a c-raf antisense oligodeoxyribonucleo- tide, which specifically decreases intracellular Raf- 1 levels, also substantially inhibits both erythropoietin and IL-3-directed DNA synthesis. Together, these re- sults provide evidence that activated Raf-1 is a neces- sary component of erythropoietin and IL-3 growth signaling pathways.
Erythropoietin is a lineage-specific hematopoietin which stimulates the growth and differentiation of red blood cell precursors (1). These developmental phenomena are initiated by the binding of erythropoietin to its specific cell surface receptor, which has recently been cloned (2). Interestingly, this receptor belongs to a unique hematopoietic growth factor receptor family which also includes those for IL-3,’ IL-2, IL- 4, IL-6, IL-7, G-CSF, and GM-CSF (3). The mechanism(s) by which this growth signal, generated at the plasma mem- brane, is transmitted to the cell nucleus is unknown but may involve regulation of the phosphorylation status of crucial intermediate proteins. Recent studies demonstrate that eryth- ropoietin rapidly mediates phosphorylation of membrane- associated proteins at both tyrosine and serine residues in a
Grants CA 44649, CA 47993, DK 16702, and T32 HL07525. The costs *This research was supported by National Institutes of Health
of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “adver- tisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
§ Fellow of the Leukemia Society of America. $$ Scholar of the Leukemia Society of America. To whom corre-
spondence should be addressed Johns Hopkins Oncology Center, Johns Hopkins University School of Medicine, 424 N. Bond St., Baltimore, MD 21231.
The abbreviations used are: IL, interleukin; G-CSF, granulocyte colony-stimulating factor; GM-CSF, granulocyte-macrophage colony- stimulating factor; CSF-1, colony-stimulating factor-1; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis.
murine cell line which proliferates in response to the hormone (4). It has also been found that erythropoietin mediates the dephosphorylation of serine residue(s) on a 43-kDa integral membrane protein in both normal and leukemic murine eryth- roid cells (5). These findings support a role for protein phos- phorylation in the erythropoietin signal transduction mecha- nism(s).
The c-raf protooncogene encodes an ubiquitously expressed 74-kDa cytosolic protein, Raf-1, with serine/threonine protein kinase activity (6, 7). Recent studies suggest that Raf-1 may play a regulatory role in the transmission of a variety of mitogenic stimuli (8, 9). Thus, polypeptide growth factors including IL-3, GM-CSF, IL-2, platelet-derived growth factor, CSF-1, epidermal growth factor, and insulin stimulate rapid activation of the Raf-1 intrinsic protein kinase function in association with Raf-1 phosphorylation (10-16). Further, the viral homolog, v-raf, can induce factor-independent growth in murine myeloid cell lines when combined with v-myc (17). Additionally, v-raf and v-myc together induce murine ery- throleukemias and lymphomas (18). These data, suggesting that growth factor receptors, including three which share homology with the erythropoietin receptor, may employ Raf- 1 as a cytosolic second messenger and the observation that v- ruf is erythroleukemogenic, prompted an investigation of the role of Raf-1 in erythropoietin-mediated growth signaling. Herein we describe results which demonstrate that physio- logic concentrations of erythropoietin induce rapid phos- phorylation and activation of Raf-1. Furthermore, experi- ments using an antisense oligodeoxyribonucleotide provide evidence that Raf-1 is a necessary component in erythropoi- etin-induced proliferation.
EXPERIMENTAL PROCEDURES
Cell Lines and Culture Media-The erythropoietin-dependent mu- rine erythroid cell line HCD-57 was grown and maintained as de- scribed (19). The FDC-P1-derived cell line FDC-Pl/ER expresses the murine erythropoietin receptor and is dependent on either IL-3 or erythropoietin for growth as described below.
Chemicals and Radioisotopes-Ortho[32P]phosphoric acid, [T-”P] ATP, 1251-labeled staphylococcal protein A, lZ5I-erythropoietin, and [3H]thymidine were obtained commercially (Amersham Corp.). Syn- thetic murine IL-3 was a generous gift of Dr. I. Clark-Lewis (Univer- sity of British Columbia). Recombinant human erythropoietin was provided by Ortho Biotech. All other chemicals were reagent grade and were obtained from commercial sources unless otherwise speci- fied.
Construction of pEEG/neo/ER-A 1570-base pair XbaIIKpnI frag- ment was used as a source of the murine erythropoietin receptor cDNA (2). The fragment was incubated with T, DNA polymerase to create blunt ends. It was then inserted into the EcoRI site of the eukaryotic expression vector pEEG/HCMV/neo (20), which had been
14964
Erythropoietin Induces Raf-1 Activation 14965
filled in using the Klenow fragment of Escherichia coli DNA polym- After 2 weeks in drug-containing medium, the cells were transferred erase 1. to medium which had G418 and recomhinant human erythropoietin
Transfection and Exprc.ssion-The murine IL-Mependent cell (10 units/ml) in place of WEHI-3-conditioned medium. Stahle, single line, FDC-P1 (21), does not express the erythropoietin receptor cell clones expressing the erythropoietin receptor, as determined hy message and is not dependent on erythropoietin for growth as deter- their ability to grow in the presence of erythropoietin, were isolated. mined hy Northern hlot analysis and cell proliferation assay (data The cells, FDC-Pl/ER, were maintained in RI'MI 1640 medium not shown). FDC-P1 cells were suspended at a concentration of 10' containing 10%) fetal calf serum and ervthropoietin (0.5 units/ml). cells/ml in ice-cold phosphate-buffered saline. Plasmid DNA (8.2 pg) Equilibrium Binding Assa,v.s-Growth lactor was withdrawn from was added to the cuvettes containing 800 pl of cell suspension, and logarithmic phase FDC-Pl/ER cells 16 h prior to the assay. Cells the cuvettes were incubated on ice for 15 min. The DNA-cell mixture were resuspended in hinding huffer consisting of 0.02"A sodium azide was then suh.iected to 500 microfarads and 350 V field strength (875 and 0.2% hovine serum alhumin (w/v) in RPMI 1640, pH 7.4. T W O X V/cm) with a Rio-Rad gene pulser. After a 10-min incubation on ice, 10'' cells in 100 p1 of hinding huffer containing "'I-erythropoietin the contents ofthe cuvettes were transferred directly to culture flasks were incubated with or without 0.2 p M (100-fold o r more excess) with RI'MI 1640 medium containing 10% fetal calf serum and 10% unlabeled erythropoietin a t 37 "C for 3 h. The reaction mixtures were WEHI 4 conditioned medium as a source of IL-3. Forty-eight h after layered on a 10% sucrose solution, centrifuged, and frozen on dry ice. translection the cells were centrifuged and resuspended in medium The hottoms of the tuhes containing the frozen cell pellets were cut containing G418 (400 pg/ml) and 10% WEHI-.?-conditioned medium. off and counted in a y-counter. Results were corrected for the non-
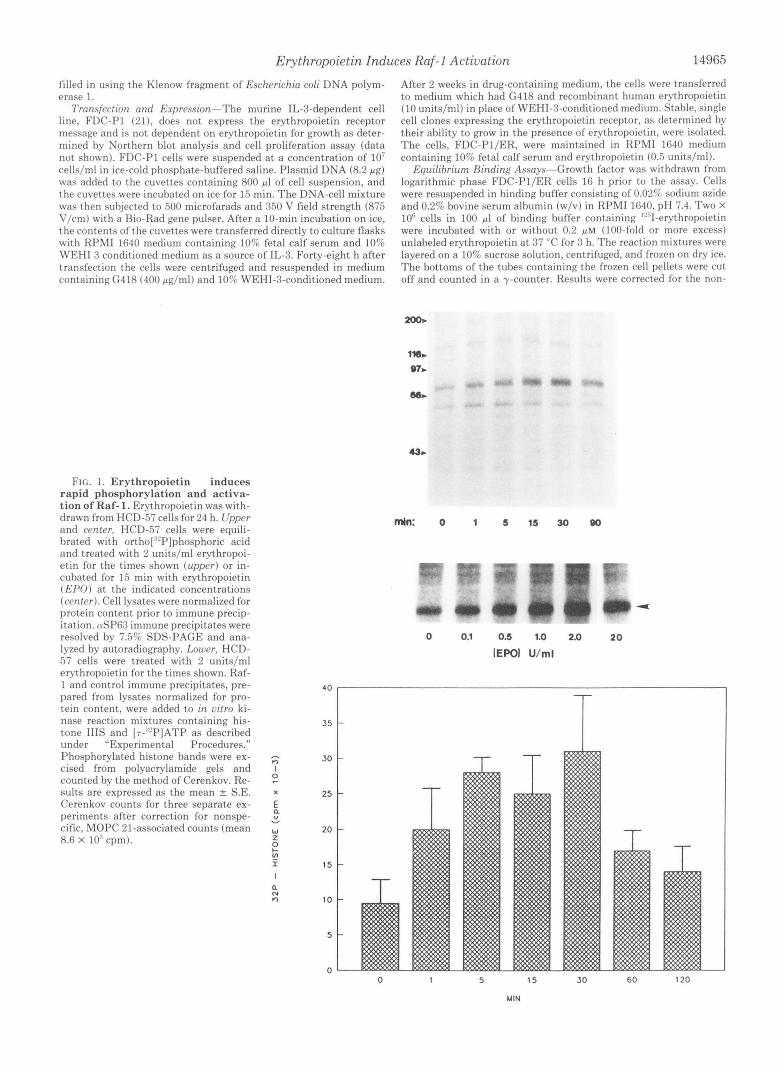
FIG. 1. Erythropoietin induces rapid phosphorylation and activa- tion of Raf- 1. Erythropoietin was with- drawn from HCD-57 cells for 24 h. Upper and center, HCD-57 cells were equili- brated with ortho[:"P]phosphoric acid and treated with 2 units/ml erythropoi- etin for the times shown (upper) or in- cuhated for I 5 min with erythropoietin (E,'PO) at the indicated concentrations (center). Cell lysates were normalized for protein content prior to immune precip- itation. CUSP63 immune precipitates were resolved by 7.5% SDS-PAGE and ana- lyzed by autoradiography. Lower, HCD- 57 cells were treated with 2 units/ml erythropoietin for the times shown. Raf- 1 and control immune precipitates, pre- pared from lysates normalized for pro- tein content, were added to in vitro ki- nase reaction mixtures containing his- tone Ills and [r-"'PIATP as described under "Experimental Procedures." Phosphorylated histone hands were ex- cised from polyacrylamide gels and counted by the method of Cerenkov. Re- sults are expressed as the mean & S.E. Cerenkov counts for three separate ex- periments after correction for nonspe- cific, MOPC 21-associated counts (mean 8.6 X 10.' cpm).
4 0
35
h n 30 I 0 r
X 25
E, v
W 20 z ? 9
15 I a n 10 N
5
0
mln: 0 1 5 15 30 90
"" 4
0 0.1 0.5 1.0 2.0 20
IEPOI U/ml
T T
T
0 1 5 15 30 60 I20
MIN

14966 Erythropoietin Induces Raf-1 Activation
. P0,F
. OrWn
EPO - + FIG. 2. Erythropoietin (EP0)-induced Raf-1 phosphoryla-
tion occurs at both serine and tyrosine residues. Phosphorylated Iiaf-1 from HCD-57 cells was excised from polyacrylamide gels and subjected to phosphoamino acid analysis as described under “Exper- imental Procedures.” Dashed circles indicate the positions of the ninhydrin-stained standards. P-srr, phosphoserine; P-thr, phospho- threonine; 1’-tyr, phosphotyrosine.
specific binding of ’“”I-erythropoietin which occurred in the presence of excess unlabeled ligand.
Metabolic Labeling and Immunoprecipitation-FDC-Pl/ER cells were growth factor-deprived and equilibrated with ortho(:”P]phos- phoric acid as described for FDC-P1 cells (10). HCD-57 cells were labeled using a modification of this protocol. Briefly, HCD-57 cells were washed three times in RPMI 1640 and incubated in RPMI 1640 containing 3Ori fetal hovine serum without erythropoietin for 24 h. Cells were then washed three times in phosphate-free RPMI 1640 and incubated at 1 X 10’ cells/ml in phosphate-free medium contain- ing 305 dialyzed fetal bovine serum and 100 pCi/ml ortho[:”P] phosphoric acid for 60 min. Radiolabeled cells were treated with erythropoietin for the times indicated, washed, and lysed as described (10). Lysates were clarified by centrifugation at 8,740 X g for 15 min and protein content determined as described (10). The clarified lysates were normalized for protein content, and Raf-1 was precipi- tated using a Raf-1 peptide antiserum, (uSP63, adsorhed to staphylo- coccal protein A-Sepharose as described (10). Immune complexes were eluted and resolved by SDS-PAGE. Phosphoproteins were ana- lyzed by autoradiography using Kodak X-Omat film.
In Vitro Kinase Rractions-FDC-Pl/ER or HCD-57 cells were deprived of growth factor for 24 h and treated with erythropoietin for the times indicated. Immune precipitates were prepared from cell lysates using anti-Raf monoclonal antibodies, PRRl and URP30S3, or MOPC 21, an isot-ype-matched monoclonal control (Sigma) ad- sorhed to protein G-Sepharose beads as described (22). Washed immune complex-containing beads were suspended in 100 p1 of 50 mM Tris (pH 7.2) containing 50 mM NaCI, 1% glycerol (v/v), 0.1% Triton 5-100 (v/v), 1 mM dithiothreitol, 20 mM MgCI,, and 50 pg of histone 111-S (Sigma) as a substrate. After the addition of 20 pCi of [r-.”P]ATP, the reactions were carried out for 30 min at 37 “C and were terminated hy boiling in SDS sample buffer. Supernatants were resolved by SDS-PAGE, and the phosphorylated histone bands were excised and counted by the method of Cerenkov.
Preparation ofO(igodeol.yribonucleotides-Antisense(5”TCCCTG- TATGTGCTCCAT-3‘) and sense (5”ATGGAGCACATACAGGGA- 3’) oligodeoxyrihonucleotides corresponding to codons 1-6 of murine c-raf or a randomly generated nonsense oligomer control (5’- TTTTTGCACCAGCTTGCC-3’) with the same overall base compo- sition as the antisense oligomer were prepared as descrihed using a Milligen 7500 synthesizer and purified by reverse phase high pressure liquid chromatography at the DNA Synthesis Core Facility, Depart- ment of Biochemistry, Johns Hopkins University (23). Aliquots of purified oligomers dissolved in ammoniated water were evaporated to
dryness in a Speed Vac concentrator (Savant) and resuspended in RPMI 1640 immediately prior to use.
Mrasurement of pH/Thvrnidine Incorporation-Log phase FDC- P l /ER cells were washed three times in KPMI 1640 and plated at 5 X 10’ cells/well in 96-well dishes in serum-free RPMI 1640. Oligomers were added at one-half the final concentration indicated, and cells were incubated at 37 “C for 14 h. The additional dose of the oligomer was added for 5 h followed by the addition of 0.1 units/ml ervthro- poietin or 1.0 pg/ml IL-3. Following an additional 6-h incubation, cells were pulsed with 1 pCi/well [“Hlthymidine for 16 h. Cells were then hypotonically lysed and passed over a glass fiber filter using the Mini-MASH II apparatus (Whittaker M.A. Rioproducts), and [“HI thymidine incorporation was determined by scintillation counting as described (10). The IL-3 and erythropoietin proliferation assays (Fig. 3, lower) were performed as outlined above using the indicated con- centrations of growth factor in the ahsence of oligomers.
Western Rhtting-2 X 10’ log phase FDC-Pl/ER cells were washed, suspended in serum-free KPMI, and treated with oligomers at times 0 and 14 h as ahove. Cell lysates were prepared after a total of 19 h incubation time and normalized for protein content as ahove. Raf-1 immunoprecipitates prepared using the tuSP63 antiserum under conditions of antibody excess were resolved by SDS-PAGE as above. The resolved proteins were electrophoret.ically transferred to nitro- cellulose membranes and probed with tuSP6S antiserum, and Raf-1 was detected using ‘“”I-labeled staphylococcal protein A as descrihed (10).
Phosphoamino Acid Ana!vsis-’”P-Labeled Raf-1 was excised from polyacrylamide gels, and phosphoamino acid analysis was performed as described (10).
RESULTS AND DISCUSSION
Erythropoietin Induces Rapid Phosphorylation and Activa- tion of Raf-I-Addition of erythropoietin to growth factor- deprived HCD-57 cells results in rapid phosphorylation of Raf-1 (Fig. 1, upper). The increase in phosphorylation, evi- dent l min after erythropoietin addition, peaks after 15-30 min and declines by 90 min. This rapid and transient phos- phorylation is similar to that seen with IL-3 and GM-CSF (10) suggesting that Raf-1 may have a central, early regulatory role in hematopoietic growth. Furthermore, Raf-1 phosphoryl- ation occurs a t physiologic doses of erythropoietin. Phos- phorylation is evident after treatment with 0.1 units/ml eryth- ropoietin and is maximal a t 2.0 units/ml (Fig. 1, center). Importantly, erythropoietin induces activation of the Raf-1 kinase function in a time course which parallels phosphoryl- ation. Histone 111-S phosphorylation by immunoprecipitated Raf-1 is increased 2-fold 1 min after erythropoietin addition, increased over 3-fold by 30 min, and is diminished to near base-line levels by 120 min (Fig. 1, lower). Together, phos- phorylation and activation of Raf-1 represent a potential rapid regulatory mechanism for erythropoietin-mediated pro- liferation.
Erythropoietin-induced Raf-I Phosphorylation Occurs at Both Serine and Tyrosine Residues-Interestingly, erythro- poietin mediates significant phosphorylation of both serine and tyrosine residues on Raf-1 (Fig. 2). This finding mimics those observed with the other hematopoietic growth factors, IL-3, GM-CSF, and IL-2 (10, 11). However, this is in contra- distinction to the Raf-1 phosphorylation induced by some other mitogenic stimuli, such as the T-cell activators, platelet- derived growth factor, epidermal growth factor, insulin, and CSF-1, all of which have been observed to induce serine phosphorylation predominately or exclusively (12-16, 24). The significance underlying Raf-1 phosphorylation is un- known, although some evidence directly implicates it in the Raf-1 activation mechanism. Thus, tyrosine phosphorylation of Raf-1 in vitro has been associated with an increase in Raf- 1 kinase activity (25). However, tyrosine phosphorylation is apparently not necessary for Raf-1 activation since mitogens which mediate only serine phosphorylation also induce Raf-1
Erythropoietin Induces Raf-1 Activation 14967
‘1
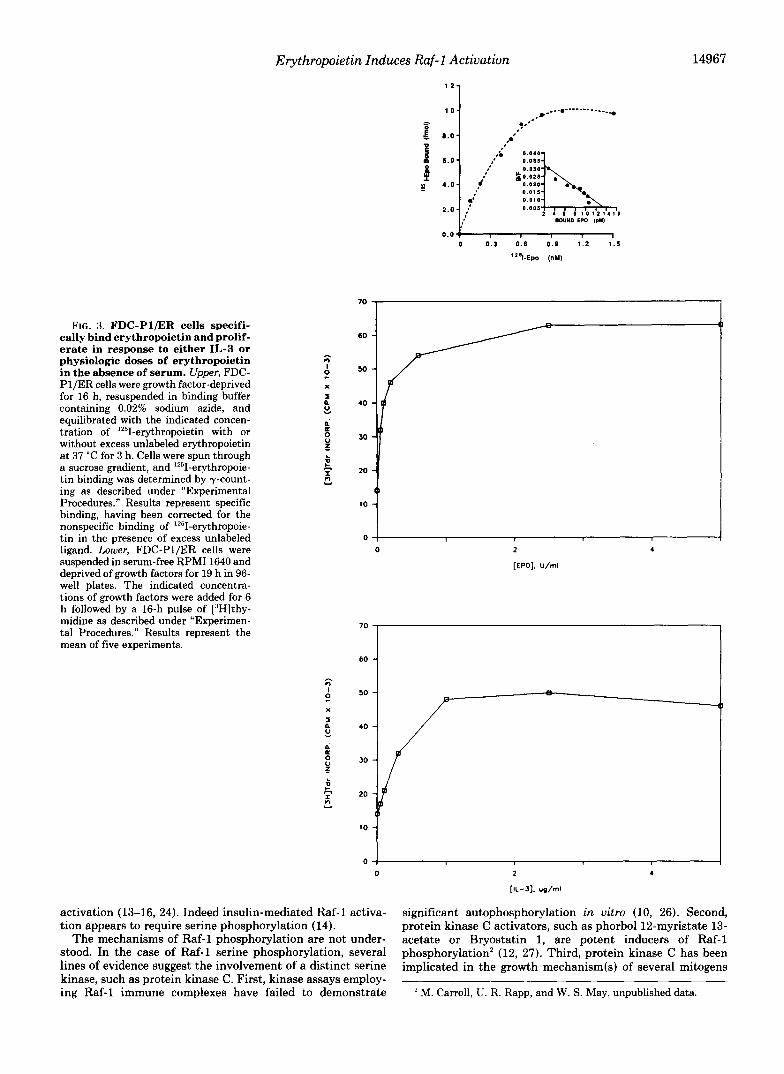
FIG. 3. FDC-Pl/ER cells specifi- cally bind erythropoietin and prolif- erate in response to either IL-3 or physiologic doses of erythropoietin in the absence of serum. Upper, FDC- Pl/ER cells were growth factor-deprived for 16 h, resuspended in binding buffer containing 0.02% sodium azide, and equilibrated with the indicated concen- tration of 1251-erythropoietin with or without excess unlabeled erythropoietin a t 37 “C for 3 h. Cells were spun through a sucrose gradient, and 1’51-erythropoie- tin binding was determined by y-count- ing as described under “Experimental Procedures.” Results represent specific binding, having been corrected for the nonspecific binding of lZ5I-erythropoie- tin in the presence of excess unlabeled ligand. Lower, FDC-Pl/ER cells were suspended in serum-free RPMI 1640 and deprived of growth factors for 19 h in 96- well plates. The indicated concentra- tions of growth factors were added for 6 h followed by a 16-h pulse of [3H]thy- midine as described under “Experimen- tal Procedures.” Results represent the mean of five experiments.
I 0 X
I 0 v
i 0
z - P X Y
6 . 0 -
4 . 0 - {
.’.
2.0- : ?‘ 2 4 8 8 1 0 1 2 1 4 1 8
:::::q 0.010
0.015
BOUND EPO IpY)
0.0 I 0 0 . 3 0.6 0.0 1 . 2 1.5
”61-Epo (nM)
70
60
50
40
x)
20
10
0
0 2 4
[EPO]. u/ml
60
’” I
0 1 0 2 4
[IL-31. uq/ml
activation (13-16, 24). Indeed insulin-mediated Raf-1 activa- significant autophosphorylation in vitro (10, 26). Second, tion appears to require serine phosphorylation (14). protein kinase C activators, such as phorbol 12-myristate 13-
The mechanisms of Raf-1 phosphorylation are not under- acetate or Bryostatin 1, are potent inducers of Raf-1 stood. In the case of Raf-1 serine phosphorylation, several phosphorylation2 (12, 27). Third, protein kinase C has been lines of evidence suggest the involvement of a distinct serine implicated in the growth mechanism(s) of several mitogens kinase, such as protein kinase C. First, kinase assays employ- ing Raf-1 immune complexes have failed to demonstrate ’ M. Carroll, U. R. Rapp, and W. S. May, unpublished data.
14968 Erythropoietin Induces Raf-I Actiuation
100 I
I
i 1
00
70
60
50
40
30
20
10
0
1 2 3
5.0 10 I OLlW I. uM
15
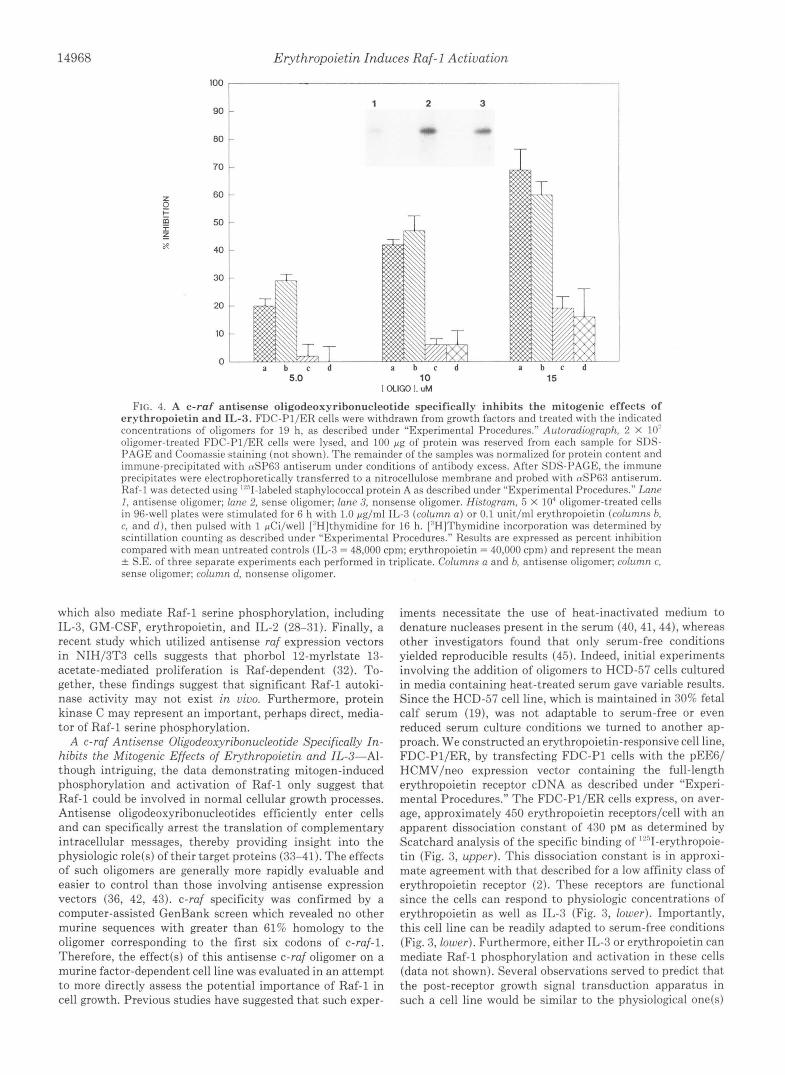
F I ~ . 4. A c-raf antisense oligodeoxyribonucleotide specifically inhibits the mitogenic effects of erythropoietin and IL-3. FDC-Pl/ER cells were withdrawn from growth factors and treated with the indicated concentrations of oligomers for 19 h, as described under “Experimental Procedures.” Autorfldiogrflph, 2 X 1 0 ’ oligomer-treated FDC-PI/ER cells were lysed, and 100 gg of‘ protein was reserved from each sample for SDS- PAGE and Coomassie staining (not shown). The remainder of the samples was normalized for protein content and immune-precipitated with nS1’63 antiserum under conditions of antihody excess. After SDS-PAGE, the immune precipitates were electrophoretically transferred to a nitrocellulose memhrane and prohed with tuSP6.7 antiserum. Raf-1 was detected using ”,.’I-labeled staphylococcal prot,ein A as descrihed under “Experimental Procedures.” I m c I , antisense oligomer; lane 2. sense oligomer; lane 3, nonsense oligomer. Histogram, 5 X 10” oligomer-treated cells in 96-well plates were stimulated for 6 h with 1.0 pg/ml IL-3 (column n ) or 0.1 unit,/ml erythropoietin (columns h. c, and d ) , then pulsed with 1 pCi/well [“Hlthvmidine for 16 h. [“HIThymidine incorporation was determined by scintillation counting as descrihed under “Experimental Procedures.” Results are expressed as percent inhihition compared with mean untreated controls (11,-3 = 48,000 cpm; erythropoietin = 40,000 cpm) and represent the mean rf: S.E. of three seDarate exueriments each Derformed in triplicate. Columns a and b, antisense oligomer; column c. sense oligomer; column d, nonsense oligomer.
which also mediate Raf-1 serine phosphorylation, including IL-3, GM-CSF, erythropoietin, and IL-2 (28-31). Finally, a recent study which utilized antisense ruf expression vectors in NIH/3T3 cells suggests that phorbol 12-myrlstate 13- acetate-mediated proliferation is Raf-dependent (32). To- gether, these findings suggest that significant Raf-1 autoki- nase activity may not exist in vivo. Furthermore, protein kinase C may represent an important, perhaps direct, media- tor of Raf-l serine phosphorylation.
A c-ruf Antisense Oligodeoxyribonucleotide Specifically In- hibits the Mitogenic Effects of Erythropoietin and IL-3”Al- though intriguing, the data demonstrating mitogen-induced phosphorylation and activation of Raf-1 only suggest that Raf-1 could be involved in normal cellular growth processes. Antisense oligodeoxyribonucleotides efficiently enter cells and can specifically arrest the translation of complementary intracellular messages, thereby providing insight into the physiologic role(s) of their target proteins (33-41). The effects of such oligomers are generally more rapidly evaluable and easier to control than those involving antisense expression vectors (36, 42, 43). c-ruf specificity was confirmed by a computer-assisted GenBank screen which revealed no other murine sequences with greater than 61% homology to the oligomer corresponding to the first six codons of c-ruf-1. Therefore, the effect(s) of this antisense c-ruj oligomer on a murine factor-dependent cell line was evaluated in an attempt to more directly assess the potential importance of Raf-1 in cell growth. Previous studies have suggested that such exper-
iments necessitate the use of heat-inactivated medium to denature nucleases present in the serum (40,41,44), whereas other investigators found that only serum-free conditions yielded reproducible results (45). Indeed, initial experiments involving the addition of oligomers to HCD-57 cells cultured in media containing heat-treated serum gave variable results. Since the HCD-57 cell line, which is maintained in 30% fetal calf serum (19), was not adaptable to serum-free or even reduced serum culture conditions we turned to another ap- proach. We constructed an erythropoietin-responsive cell line, FDC-PI/ER, by transfecting FDC-PI cells with the pEE6/ HCMV/neo expression vector containing the full-length erythropoietin receptor cDNA as described under “Experi- mental Procedures.” The FDC-PI/ER cells express, on aver- age, approximately 450 erythropoietin receptors/cell with an apparent dissociation constant of 430 PM as determined by Scatchard analysis of the specific binding of ’“’I-erythropoie- tin (Fig. 3, upper). This dissociation constant is in approxi- mate agreement with that described for a low affinity class of erythropoietin receptor (2). These receptors are functional since the cells can respond to physiologic concentrations of erythropoietin as well as IL-3 (Fig. 3, lower). Importantly, this cell line can be readily adapted to serum-free conditions (Fig. 3, lower). Furthermore, either IL-3 or erythropoietin can mediate Raf-1 phosphorylation and activation in these cells (data not shown). Several observations served to predict that the post-receptor growth signal transduction apparatus in such a cell line would be similar to the physiological one(s)

Erythropoietin Induces Raf-1 Activation 14969
employed by erythropoietin. First, it has been shown that (1987) in Oncogenes and Cancer (Aaronson, S. A,, ed) pp. 55-74, Japan
another murine IL-3-dependent cell line Ba/F3/ER, also ex- 9. Li, P., Wood, K., Mamon, H., Haser, W., and Roberts, T. (1991) Cell 6 4 , Scientific Societies Press, Tokyo
pressing the murine erythropoietin can 10. Carroll, M. P., Clark-Lewis, I., Rapp, U. R., and May, W. S. (1990) J. Biol. 479-482
proliferate in response to erythropoietin (46). Second, the Chem. 2 6 5 , 19812-19817 murine receptors for I L - ~ and erythropoietin share homology 11. Turner, B., R a w , U., APP, H., Greene, M., Dohashi, K., and Reed, J. (1991)
in their cytosolic domains suggesting that they may interact 12. Morrison, D. K., Kaplan, D. R., Rapp, U. R., and Roberts, T. M. (1988)
both IL-3 and erythropoietin mediate tyrosine phosphoryla- (1990) EMBO J. 9, 3649-3657 tion of five proteins with identical M, in a cell line which 14. Kovacina, K. S., Yonezawa, K., Brautigan, D. L., Tonks, N. K., Rapp, U.
proliferates in response to either growth factor (4). Addition 15. Blackshear, P. J., Haupt, D. M., App, H., and Rapp, U. R. (1990) J. Biol.
Raf-1 levels by as much as 65% as estimated by optical U. (1991) Mol. Cell. Biol. 11 , 913-919 densitometry ( ~ i ~ . 4). H ~ ~ ~ ~ ~ ~ , no changes in other cellular 17. Cleveland, J. L., Jamen, H. w., Bister, K., Fredrickson, T. N., Morse, H.
protein levels were seen on Coomassie-stained polyacrylamide 18. Rapp, U. R., Cleveland, J. L., Frederickson, T. N., Holmes, K. L., Morse, gels of cell lysates (data not shown). Concomitantly, these H. C., Jansen, H. W., Patschinsky, T., and Bister, K. (1985) J. Virol.
same oligomers specifically inhibited IL-3 or erythropoietin- 19. Spivak, J. L., Pham, T., Isaacs, M., and Hankins, W. D. (1991) Blood 7 7 , induced [SH]thymidine in a dose-dependent 20. Stephens, P. E., and Cockett, M. I. (1989) Nucleic Acids Res. 17, 7110 manner (Fig. 4). This observed effective dose range is similar 21. Dexter, T. M., Garland, J., Scott, D., Scolnick, E., and Metcalf, D. (1980) to the Oligomer concentrations reported to be active in Other 22. Kolch, W., Weissinger, E., Mischak, H., Troppmair, J., Showalter, S. D., systems (35,38,40,41). Viability, as assessed by light micros- Lloyd, P., Heidecker, G., and Rapp, U. R. (1990) Oncogene 5 , 713-720 copy and trypan blue exclusion, was 7 5 8 5 % for both oligo- 23. Beaucage, S. L., and Caruthers, M. H. (1981) Tetrahedron Lett. 2 2 , 1859-
mer-treated and untreated cells, indicating that the antisense 24. Siegel, J. N., Klausner, R. D., Rapp, U. R., and Samelson, L. E. (1990) J.
antisense-mediated inhibition of cell proliferation as meas- M., and Williams, L. T. (1989) Cell 5 8 , 649-657 ured by DNA synthesis and a commensurate specific decrease 26. S c h r k A. M., Copeland, T., Oroszlan, S., and Raw, U. R. (1988) Oncogene
in the intracellular Raf-1 levels, strongly suggest that Raf-1 27. Carroll, M. P., Rapp, U. R., and May, W. S. (1990) Exp. Hematol. ( N Y ) is a necessary component in 1L-3- and 28. Fields, A. P., Pincus, S. M., Kraft, A. S., and May, W. S. (1989) J. Biol. diated growth. These observations are in agreement with a Chem. 264,21896-21901 recent report which demonstrated that transfection of a 29. Adunyah, S. E., U n b , T. M., Wagner, F., and Kraft, A. s. (1991) J. B i d
antisense expression vector could induce reversion of NTH/ 30. Spangler, R., Bailey, S. C., and Sytkowski, A. J. (1991) J. Biol. Chem. 266 , 3T3 v-ruf transformants and that at least One such revertant 31. Farrar, W. L., and Anderson, W. B. (1985) Nature 315 , 233-235 clone had a blunted mitogenic response to either phorbol 12- 32. Kolch, U., Heidecker, G., Lloyd, P., and Rapp, U. R. (1991) Nature 349 ,
myristate 13-acetate Or Serum (32). These data, when taken 33. Weintraub, N., Izant, J. G., and Harland, R. M. (1985) Trends Genet. 1, together with the observed growth factor-mediated phos- phorylation and activation of Raf-l, form the basis for a 34. Neckers, L. M. (1989) in 0ligodeoxynucleotides:Antisense InhibitorsofGene
plausible erythroid growth program featuring Raf-1 as a cru- 35. Gewirtz, A. M., and Calahretta, B. (1988) Science 2 4 2 , 1303-1306 cia1 component which is activated immediately following li- 36. Stein, c. A., and Cohen, J. s. (1988) Cancer Res. 48,2659-2668
gand binding to the receptor.
Proc. Natl. Acad. Sci. U. S. A. 8 8 , 1227-1231
Proc. Natl. Acad. Sci. U. S. A. 85,8855-8859 with an Or second messenger(s) (47). 13, Baccarini, M,, Sabatini, D, M,, App, H,, Rapp, U, R,, and Stanley, E. R,
R., and Roth, R. A. (1990) J. Biol. Chem. 2 6 5 , 12115-12118
Chem. 2 6 5 , 12131-12134 Of lo pM antisense c-ruf-l specifically decreased FDC-P1/ER 16. App, H., Hazan, R., Zilberstein, A,, Ullrich, A,, Schlessinger, J., and Rapp,
C., Ihle, J. N., and Rapp, U. R. (1986) J. Cell. Biochem. 3 0 , 195-218
55,23-33
1228-1233
J. Exp. Med. 152 , 1036-1047
1862
Biol. Chem. 2 6 5 , 18472-18480 effect was not cflotoxic' These demonstrating an 25. Morrison, D. K., Kaplan, D. R., Escohedo, J. A,, Rapp, U. R., Roberts, T.
2 , 187-193
18 , 625a (abstr.)
Chem. 266,5670-5675
681-684
426-428
22-25
Expression (Cohen, J. S., ed) pp. 211-231, CRC Press, Boca Raton, FL
37. Loke, S. L., Stein, C. A,, Zhang, X. H., Mori, K., Nakanishi, M., Suhasinghe, C., Cohen, J. S., and Neckers, L. M. (1989) Proc. Natl. Acad. Sci. U. S. A. 86,3474-3478
Acknowledgments-We are grateful to Ian Clark-Lewis for supply- 38. Zamecnik, P. C., Goodchild, J., Taguchi, Y., and Sarin, P. S. (1986) Proc.
ing IL-3, L. Mulcahy for providing the erythropoietin receptor cDNA, 39. Walder, R. Y., and Walder, J. A. (1988) Proc. Natl. Acad. Sci. U, S. A. 8 5 , Natl. Acad. Sci. U. S. A. 83,4143-4146
P. Gregory Tyler for graphic design, Barbara A. Lee for preparing the manuscript, and J~~~~~~~ Ravenel and Mary Ann Isaacs for 40. H o k J. T., Redner, A. L., and Nienhuis, A. W. (1988) Mol. Cell. Bid. 8,
5011-5015
technical assistance. 41. Wickstrom, E. L., Bacon, T. A., Gonzalez, A,, Freeman, D. L., Lyman, G. 963-973
H., and Wickstrom. E. (1988) Proc. Natl. Acad. Sci. U. S. A. 85 . 1028- REFERENCES
2. D'Andrea, A. D., Lodish, H. F., and Wong G. G . (1989) Cell 5 7 , 277-285 1. Krantz, S. B. (1991) Blood 77,419-434
3. Bazan. J. F. (1990) Proc. Natl. Acad. Sci. c. S. A. 87.6934-6938 4. Quelle, F. W., and Wojchowski, D. M. (1991) J. Biol.'Chem. 2 6 6 , 609-614 5. Choi, H.-S., Wojchowski, D. M., and Sytkowski, A. J. (1987) J. Biol. Chem.
6. Rapp, U. R., Cleveland, J. L., Bonner, T. I., and Storm, S. M. (1988) in 262,2933-2936
The Oncogene Handbook (Reddy, E. P., Skalka, A. M., and Curran, T., eds) pp. 213-253, Elsevier Science Publishers B.V., Amsterdam
7. Storm, S. M., Cleveland, J. L., and Rapp, U. R. (1990) Oncogene 5 , 345- 351
8. Rapp, U. R., Cleveland, J. L., Storm, S. M., Beck, T. W., and Hpleihel, M.
42.
43.
44.
45.
46.
47.
1032 Green, P. J., Pines, O., and Inouye, M. (1986) Annu. Rev. Biochem. 5 5 ,
van der Krol, A. R., Mol, J. N. M., and Stuitje, A. R. (1988) BioTechniques 569-597
Valtieri, M., Venturelli, D., Care, A,, Fossati, C., Pelosi, E., Lahbaye, C., 6,958-976
Mattia, G., Gewirtz, A. M., Calabretta, B., and Peschle, C. (1991) Blood 77,1181-1190
Reed, J. C., Stein, C., Subasinghe, C., Haldar, S., Croce, C. M., Yum, S., and Cohen J. (1990) Cancer Res. 50,6565-6570
Li, J.-P., D'Andrea, A. D., Lodish, H. F., and Baltimore, D. (1990) Nature 343 , 762-764
Itoh, N., Yonehara, S., Schreurs, J., Gorman, D. M., Maruyama, K., Ishii, A., Yahara, I., Arai, K., and Miyajima, A. (1990) Science 247,324-327