enzyme immunometric assay of thyroliberin (trh)
TRANSCRIPT

ELSEVIER Journal of Immunological Methods 198 (1996179-85
JOURMl OF lMMUwOw6IcAL METHOUS
Enzyme immunometric assay of thyroliberin ( TRH)
Emmanuelle Etienne a ,bChristophe CrCminon a, Jacques Grassi ‘, Dominique Grouselle , Jacques Roland ‘, Philippe Pradelles a’ *
’ Commissariat b I’Energie Atomique, Senlice de Pharmacologic et d’immunologie. CE/ Saclay, 91191 Gif sur Ywtte, France b INSERM U 159, Centre Paul Broca, 2 Ter rue d’Al&ia, 75014 Paris, France
’ &ire d’Immunochimie Analytique, Dgpartement d’lmmunologie, Institut Pasteur (CNRS URA D 1961 and Unic:ersite Pierre et Marie Curie). 25 rue du Dr Roux, 75724 Paris Cedex 15. France
Received 13 June 1996; accepted 9 July 1996
Abstract
An enzyme immunometric assay of thyroliberin (TRH) using monoclonal antibodies and a derivatization procedure is described. This assay, named SPIE-IA, involves a four step procedure after chemical derivatization of TRH and biological samples by diazotized APEA. Step I: derivatized TRH was immunocaptured by a monoclonal anti-TRH antibody coated on a 96-well microtiter plate. Step 2: after washing, derivatized TRH was cross-linked via its amino group to the wells using glutaraldehyde. Step 3: washing and treatment with NaOH. Step 4: measurement of bound TRH using a monoclonal anti-TRH antibody labeled with acetylcholinesterase. The minimal detectable concentration was 0.1 pmol/ml: with a coefficient of variation less than 10% in the 0.156-10 pmol/ml range. This assay is 26-fold more sensitive and more specific than the competitive enzyme immunoassay using the same monoclonal capture antibody, derivatized TRH and TRH-acetylcholinesterase conjugate as tracer. Good correlation was observed between SPIE-IA and a sensitive competitive enzyme immunoassay using polyclonal antibodies.
Keywords: TRH; Enzyme immunometric assay; Monoclonal antibody; Acetylcholinesterase; Hapten; Derivatization
1. Introduction
Our laboratory has developed various enzyme immunoassays (EIAs) of different molecules using acetylcholinesterase as label (Grassi and Pradelles, 19911, and has recently described and patented a new
Abbreviations: TRH, thyroliberin; EIA, enzyme immunoassay;
SPIE-IA, solid phase immobilized epitope immunoassay; APEA,
2(4-amino-phenyllethyl amine; AChE, acetylcholinesterase; NSB,
nonspecific binding; MDC, minimum detectable concentration;
CV, coefficient of variation; MW, molecular weight.
* Corresponding author. Tel.: +33-l-69 08 72 99: Fax: +33-1-
69 08 59 07.
procedure for immunometric assays of haptens using a single antibody (Pradelles, 1995). This procedure, called SPIE-IA for ‘solid phase immobilized epitope immunoassay’, has been used to measure small molecules containing an amino group, as substance P (Pradelles et al., 1994), thyroxine (Pradelles et al., 1994; Etienne et al.. 19951, endothelin (Pradelles et al., 1994), leukotriene C4 (Volland et al., 19941, and
heptapeptide (Ezan et al., 1995). The general princi- ple involves four sequential steps. First, excess spe- cific antibodies are coated on a microtiter plate to insure the full capture of hapten after the immuno- logical reaction. Second, excess cross-linking reagent (glutaraldehyde or disuccinimidyl suberate) or UV
0022-1759/96/$15.00 Copyright 0 1996 Elsevier Science B.V. All rights reserved. PII SOO22- 1759(96100154-8

N
diazotized APEA N diazotized benzidine * *
- - Y
H2d W
NY
derivatized TRH N
N
antlgenlc carrier ---. ~~ ._._-_---
Fig. I. Molecular design of TRH derivatization for immunogen
preparation (right) and amino group addition to TRH (left).
irradiation is used to cross-link hapten covalently to coated antibody through amino groups. Third, the
hapten is released from the antibody binding site by use of a dissociating/denaturing agent, as NaOH. Fourth, hapten covalently linked to the solid phase is revealed by incubation with excess anti-hapten anti- body tracer (Fab’ fragment labeled with acetylcholin- esterase) followed by the addition of enzyme sub-
strate and chromogen. Due to the use of excess reagents (capture and tracer antibody) to promote formation of immunological complexes, each assay
is 70-200 times more sensitive than conventional competitive enzyme immunoassay using the same antibody and acetylcholinesterase-hapten conjugate as tracer. Accuracy, specificity and validity were also very satisfying.
The aim of our laboratory is now to extend this procedure to a large variety of short haptens. espe- cially those devoid of amino groups. As an illustra- tion, we describe here for the first time an enzyme immunometric assay of thyroliberin (TRH, 362 Da). TRH (pyro-Glu-His-Pro-NH,) (Fig. 11 is a neu- ropeptide first isolated from hypothalamus (Baler et al., 1969: Burgus et al., 19691 and subsequently
found ubiquitously in the central nervous system and digestive tract of many species (Morley, 19791. Vat+ ous competitive radio- and enzyme immunoassays have already been described (see references in Grouselle et al., 19901 and, more recently, mono-
clonal anti-TRH antibodies have been obtained (Ro- land et al.. 1992). We report here the development of SPIE-IA for TRH. Use is made of a derivatizing agent. which leads to two parallel effects: the intro- duction of an amino group into the molecule, allow- ing efficient cross-linking during SPIE-IA, and an increase in the affinity of derivatized hapten for the monoclonal antibodies (capture and tracer antibody). Sensitivity and specificity were determined and SPIE-IA and a conventional competitive enzyme im- munoassay using TRH-acetylcholinesterase conju- gate as tracer and rabbit anti-TRH antiserum were correlated.
2. Materials and methods
2.1. Appurutus
EIA and SPIE-IA were performed with special- ized microtitration equipment (washer, dispenser and plate reader) from Labsystem (Helsinki, Finland). Microplates (Maxisorp) were from Nunc (Denmark).
Unless otherwise stated, all reagents were from Sigma (St. Louis, MO). TRH was from Calbiochem (USA), TRH-OH from Bachem (Switzerland). [Phe’l-TRH from K.G. Reichter (Budapest), [Glu]‘- TRH, [~-M~-H~~‘]-TRH, [ 1 -Me-His’]-TRH. [Phe’]-
TRH, [GIU?]-TRH and pyro-Glu-His were from Peninsula (UK). 2(4-amino-phenyllethyl amine (APEA) was from Aldrich (St. Quentin, France). Acetylcholinesterase (AChE; EC 3.1.1.7) was puri- fied from electric eel (Electrophorus electricus) by affinity chromatography (Massoulie and Bon, 1976) and used in the G4 form for TRH and monoclonal antibody labeling (Pradelles et al., 19891. Chro- mogenic substrate for AChE (Ellman’s reagent) was a mixture of acetylthiocholine (substrate) and dithionitrobenzene (chromogen1 as previously de- scribed (Pradelles et al.. 1989).

E. Etienne et al. /Journal of Immunological Methods I98 f 1996) 79-85 81
2.3. Immunochemicals
Purified mouse monoclonal anti-TRH antibodies (#T200 and #T575) used in competitive EIA and SPIE-IA were prepared as previously described (Ro- land et al., 1992). Assays were correlated using
rabbit polyclonal anti-TRH antiserum (#4B17) as previously reported (Grouselle et al., 1978). In both cases, immunogens were prepared by covalent link- age of TRH to antigenic carrier using bis-diazotized benzidine (Fig. 1).
2.4. Enzyme labels
TRH-AChE conjugate was obtained as previously described (Grouselle et al., 1990). Monoclonal anti- TRH antibodies (#T200 and #T575) were purified as described elsewhere (Roland et al., 1992). One (#T200) was used as capture antibody and the other
(#T575) as tracer antibody after labeling by covalent coupling of its Fab’ fragments to the maleimido derivative of the G4 form of AChE using N-succi-
nimidyl-4-c N-maleimidomethyl) cyclohexane-l- carboxylate as previously described (Grassi et al.,
1989).
2.5. Derivatization procedure
TRH standard, TRH analogs and biological sam- ples were derivatized before enzyme immunoassay. 0.05 M APEA (200 ~1) in 1 N HCI was added to 100 ~1 of 0.12 M NaNO,. The diazotization reaction
was allowed to proceed for 15 min in an ice bath. Reaction mixture (IO ~1: 333 nmol of diazotized APEA) was added to 100 ~1 of each concentration
of standards or biological samples diluted in EIA buffer containing 0.5 N NaOH pH > 10. The reac- tion was allowed to proceed for 15 min in an ice bath. Diazotized APEA reacted with albumin to form orange azo compounds. Excess diazotized APEA was neutralized by addition of 860 ~1 of EIA buffer containing 0.1 M phenol. After 5 min of reaction the mixture was adjusted to pH 7 by addition of 30 ~1 of
1 N HCl.
2.6. Competitive EIA
EIA was performed in 0.1 M phosphate buffer, pH 7.4, containing 0.15 M NaCl, lo-’ M EDTA,
0.1% bovine serum albumin, and 0.01% sodium azide (Pradelles et al., 1989). Briefly, 96-well mi- crotiter plates were coated with goat affinity-purified antibodies specific for mouse IgGs (Immunotech, France) or mouse purified monoclonal anti-rabbit
IgGs @PI-Bio, France). To each well were added 50 ~1 of standard, buffer or sample (all derivatized or not), 50 ~1 of enzymatic tracer (2 Ellman U/ml), and 50 ~1 of diluted antibody. After 18 h incubation at 4°C the wells were washed and Ellman’s reagent (200 ~1) was dispensed into each well. After 60 min of enzymatic reaction, the absorbance at 414 nm was measured in each well.
2.7. SPIE-IA
The principle of SPIE-IA for TRH is illustrated in Fig. 2. The routine assay was performed in EIA buffer as previously described (Volland et al., 1994)
for leukotriene C,. Briefly, each step (Fig. 2) was preceded by a washing procedure (Pradelles et al., 1989) using 0.01 M phosphate buffer (pH 7) contain- ing 0.05% Tween 20. Microtiter plates were coated with monoclonal anti-TRH antibody (#T200) as de- scribed elsewhere (Pradelles et al., 1994). Step 1 Immunological capture. Incubation for 18 h
at 4°C of derivatized standards or biological samples diluted in EIA buffer.
Step 1 : immunological capturs Step 2 : covalent immobilization
washing and cross-linking
reaction
darivatized TRH
J
washing and NaOH addition
Step 4 : immunological capture with tracer end visualization I -
Step 3 : epitop-2 liberation I
Fig. 2. General principle of SPIE-IA of TRH.

Step 2 Epitope immobili:ution. 100 ~1 of 0.1 M phosphate buffer, pH 7, containing 2.5% glu- taraldehyde were added to each well. After 5 min reaction at 22°C and washing, 250 ~1 of 0.1 M NaBH, in water were added and the reaction was allowed to proceed for 5 min in order to reduce the remaining aldehyde groups of glutaraldehyde bound to the plate.
Step 3 Epitope release. 250 ~1 of 1 N NaOH were added to each well for a 2 min incubation.
Step 4 Enzymatic tracer udditiorl and calorimetric ussu_v. 100 ~1 of monoclonal Fab’ anti-TRH antibody (#T575) labeled with acetylcholin- esterase (5 Ellman U/ml) diluted in EIA buffer were added to each well and the im-
munological reaction was allowed to proceed for 18 h at 4°C. After soaking with washing buffer for 30 min, Ellman’s reagent (200 ~1)
was added to each well and the enzymatic reaction was allowed to proceed for 30 min
before reading the absorbance at 4 14 nm.
2.8. Calculations
The competition curves were expressed in terms of B/B, X 100 as a function of the logarithm of the dose. B and B, represent the bound enzyme activity measured in the presence or absence of competitor, respectively. Fitting of the standard curves and calcu- lations of TRH content in biological samples were performed using a linear log-logit (B/B,,) transfor- mation. Unless otherwise stated, all measurements for standards or samples were made in duplicate, and in quadruplicate for B. values. Nonspecific binding (NSB) was determined using an incubation mixture in which the specific antibody was replaced by 50 ~1 of EIA buffer. The minimum detectable concentra- tion (MDC) was taken as the mean of eight concen- trations of analyte inducing a significant decrease (3 standard deviations) in B,,
SPIE-IA curves were expressed in terms of ab- sorbance (AU) versus analyte concentration. Fitting of the standard curves and calculations of TRH content in biological samples were performed using Eliot software (Immunotech. France). Unless other- wise stated, all measurements for standards or sam- ples were made in duplicate, and in octuplicate for NSB. The minimum detectable concentration was
taken as the concentration of analyte inducing a significant increase (3 standard deviations) in NSB value. The precision profile was established on the
basis of standard curves for which eight replicates of each standard concentration were used. The results were expressed in terms of the coefficient of varia- tion (% CV> versus the analyte concentration.
2.9. Specificity measurements
The specificity of each assay was checked by testing its capacity to detect compounds structurally related to TRH. by establishing for each of these compounds the corresponding standard curves. Re- sults were expressed in terms of percentage cross-re- activity (% CR), arbitrarily defined as the ratio (%I of the concentration of TRH and TRH-related com- pounds producing a 0.5 AU signal for SPIE-IA, or a 50% decrease in B, for competitive EIA.
2. IO. Biological sample preparation and correlutiw .study
Adult Wistar rats hypothalami and hypophyses (IFFA Credo, France) were treated as previously described (GrouselIe et al., 1990). Three and two pools of five and six hypothalami each, respectively,
and one pool of 22 neurohypophyses, were used for the correlation study. The extracts were resuspended with 0.5 ml of EIA buffer and 100 ~1 of each mixture was assayed directly or after derivatization by diazotized APEA as described above. Concentra- tions of TRH equivalents measured by SPIE-IA were
compared with those found by conventional competi- tive enzyme immunoassay using rabbit anti-TRH antiserum as previously described (Grouselle et al., 1990).
3. Results
3. I. Sewitir$v and precision
Routine standard curves for TRH measurement by competitive EIA using monoclonal antibody are pre- sented in Fig. 3.
Sensitivity was greater with APEA-TRH deriva- tive, with an MDC of 2.6 pmol/ml compared with
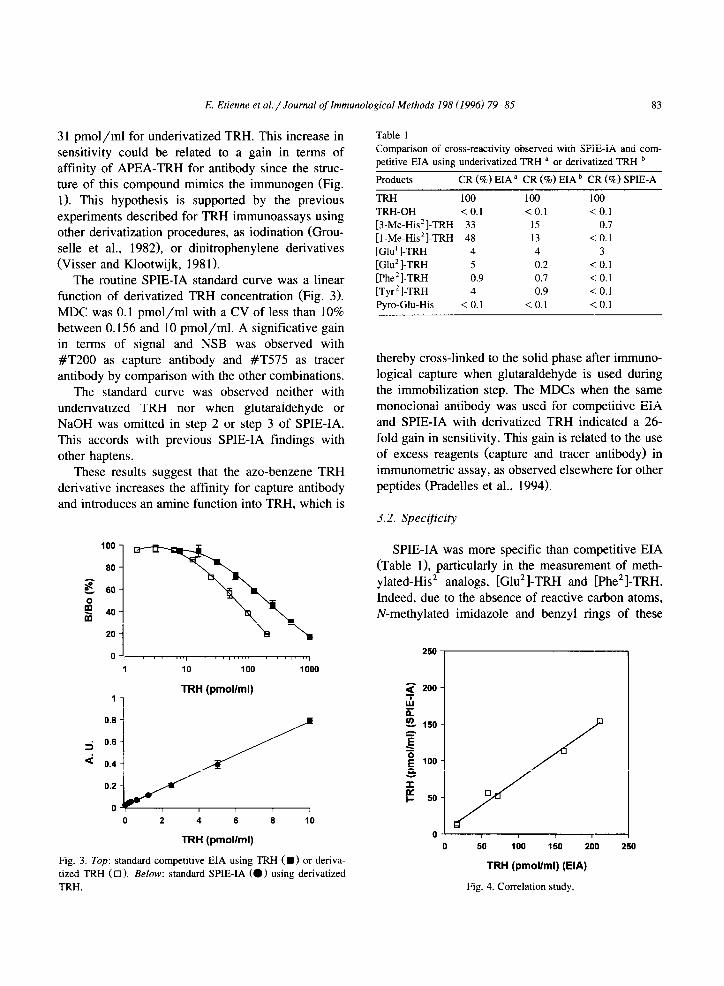
E. Etienne et al. / Journal of Immunological Methods 198 (19961 79-85 83
31 pmol/ml for underivatized TRH. This increase in
sensitivity could be related to a gain in terms of affinity of APEA-TRH for antibody since the struc- ture of this compound mimics the immunogen (Fig.
1). This hypothesis is supported by the previous experiments described for TRH immunoassays using other derivatization procedures, as iodination (Grou- selle et al., 1982), or dinitrophenylene derivatives (Visser and Klootwijk, 1981).
The routine SPIE-IA standard curve was a linear function of derivatized TRH concentration (Fig. 3). MDC was 0.1 pmol/ml with a CV of less than 10% between 0. I56 and 10 pmol/ml. A significative gain in terms of signal and NSB was observed with
#T200 as capture antibody and #T575 as tracer antibody by comparison with the other combinations.
The standard curve was observed neither with underivatized TRH nor when glutaraldehyde or NaOH was omitted in step 2 or step 3 of SPIE-IA. This accords with previous SPIE-IA findings with other haptens.
These results suggest that the azo-benzene TRH derivative increases the affinity for capture antibody and introduces an amine function into TRH, which is
100 -
a0 -
g _ 60- 0 2 40
20 -
10 100
TRH (pmollml)
0 2 4 6 a 10
TRH (pmollml)
Fig. 3. Top: standard competitive EIA using TRH ( W) or deriva-
tized TRH ( 0 1. Below: standard SPIE-IA (0) using derivatized
TRH.
Table 1
Comparison of cross-reactivity observed with SPIE-IA and com-
petitive EIA using underivatized TRH a or derivatized TRH b
Products CR (%) EIA a CR (%I EIA b CR f%) SPIE-A
TRH 100
TRH-OH <O.l
[3-Me-His’]-TRH 33
[ 1 -Me-His* I-TRH 48
[Glu’ ]-TRH 4
[Glu*]-TRH 5
[Phe’]-TRH 0.9
[Tyr * I-TRH 4
Pyro-C&-His <O.l
100
< 0.1
15
13
4
0.2
0.7
0.9
< 0.1
100
<O.l
0.7 <O.l
3 < 0.1
< 0.1
< 0.1
<O.l
thereby cross-linked to the solid phase after immuno-
logical capture when glutaraldehyde is used during the immobilization step. The MDCs when the same monoclonal antibody was used for competitive EIA and SPIE-IA with derivatized TRH indicated a 26- fold gain in sensitivity. This gain is related to the use of excess reagents (capture and tracer antibody) in immunometric assay, as observed elsewhere for other
peptides (Pradelles et al., 1994).
3.2. Specificity
SPIE-IA was more specific than competitive EIA (Table l), particularly in the measurement of meth- ylated-His* analogs, [Glu’]-TM-I and [Phe’]-TRH. Indeed, due to the absence of reactive carbon atoms, N-methylated imidazole and benzyl rings of these
2607
TRH (pmollml) (EIA)
Fig. 4. Correlation study.

molecules cannot be chemically reactive toward an electrophilic substitution reaction with diazotized APEA. [Glu2]-TRH and [Phe’]-TRH have been re- cently isolated from rabbit prostate (Cockle et al.. 1989) and human seminal fluid (Khan et al.. 1992).
3.3. Correlatim studies
Good correlation was observed ( r = 0.995) for the TRH content of rat tissues using SPIE-IA and competitive assay (Fig. 41.
4. Discussion
During the development of SPIE-IA, immunologi- cal capture and covalent epitope immobilization were critical steps. To insure efficient capture, the solid phase coated capture must have a high affinity for analyte. In most cases, monoclonal antibodies for short haptens have poor affinities when compared with polyclonal antibodies, as observed for TRH (Roland et al., 1992). On the other hand. the hapten must bear a chemical group suitable for chemical cross-linking to the coated capture antibody. This has been observed in our laboratory by use of homo-bi- functional cross-linkers, as glutaraldehyde or dis-
sucinimidyl suberate, for various haptens possessing an amino group. In addition. this chemical group must be located in an area of the hapten structure where chemical change cannot dramatically alter the immunoreactivity of the hapten with tracer antibody. When this group and the chemical modification in- volved are similar to those used for the immunogen preparation, it is even possible to increase the in- munoreactivity. As previously observed for various substances as cyclic nucleotides. prostaglandins. his- tamine and serotonin (Ezan et al., 1987). chemical derivatizations of short haptens before competitive immunoassay sometimes increase assay sensitivity. TRH bears no amino group and monoclonal anti-TRH antibodies have poor affinity (Roland et al.. 1992). We therefore decided to introduce an amino group via diazotized APEA following the same route previ- ously used during immunogen preparation with bia- diazotized benzidine (Grouselle et al., 1978).
APEA is well known as a chemical substrate of monoamine oxidase (Reyes-Parada et al.. 1994) but
has never been described. as for as we know. as a derivatizing reagent for histidine side chains in pep- tides and proteins. In our hands, diazotized APEA used in large excess quite effectively introduced amino groups into TRH. This method may allow SPIE-IA of small molecules containing imidazole or phenolic groups if the same kind of derivatization has been used for immunogen preparation. These results now prompt us to investigate SPIE-IA of small molecules as histamine (MW 111 Da). for example. by the use of structurally related derivatiz- ing agents for immunogen synthesis and cross-lin- king.
Acknowledgements
This work was supported by grants from the Commissariat a 1’Energie Atomique (France).
References
Baler, J.. Enzman. F.. Folkers, K.. Bowers. C.Y. and Schally,
A.V. ( 1969) The identity of chemical and hormonal properties
01’ the thyrotropin releasing hormone and pyroglutamyl-histi-
Jyl-prolinc amide. Biochem. Biophys. Res. Commun. 37. 705.
Burgus. R.. Dunn. T.F.. Desidrro. D. and Guillemin. R. (1969)
Structure molPculaire du facteur hypothalamique hypophya-
iotrope TRF d’origine ovine: mise en evidence par
\pectrom&rie de masse de la sdquence PGA-His-Pro-NH,.
C.R. Acad. Sci. (Paris) 269. 1870.
Cockle. S.M.. Aitken, A., Beg. F. and Smyth. D.G. (1989) A
novel peptide. pyroglutamyl-glutamylproline amide. in the
rabbit prostate complex. structurally related to thytropin-rc-
leasing hormone. J. Biol. Chem. 264, 7788.
Etienne. E.. Creminon. C.. Lamourette. P.. Grasi. J. and Pradelle\.
Ph. ( 199.5) Enzyme immunometric assay for t.-thyroxine usins
direct ultraviolet irradiation. Anal. Biochem. 225. 34.
Eran. E.. Mama&, S.. Rougeot. C. and Dray. F. (1987) Strategic\
for developing specific and Eensitive hapten radioimmunoas-
say\. In: C. Patron0 and B.A. Peskar (Eds.), Radioimmunoas-
hay in Basic and Clinical Pharmacology. Springer-Verlag.
New York. p. 133.
Ezan. E.. Tarradr. T.. CaLenave. C.. Ardouin. T., Genet, R..
Grassi. J.. Grognet. J.M. and Pradelles. Ph. (1995) Immune-
metric aasay of BN 52080. a heptapeptide C-terminal analogue
of horbin. Peptidea 16. 449,
(3a\\i. J. and Pradelles, Ph. (1991) Compound lahelled by the
acetylcholinesterase of Elrctrophonrs rlrctricw. its prepara-
tion and its use as tracer or marker in enzymoimmunological
determinations. United States Patent No. 5.04730.
GrassI. J., Frobert. Y.. Pradelles. Ph.. Chercuitte. F., Gruaz, D..

E. Etienne et al. / Journal of Immunological Methods I98 (1996) 79-85 85
Dayer, J.M. and Poubelle, P. (1989) Production of monoclonal
antibodies against interleukin-la and -lb. J. Immunol. Meth-
ods 123. 193.
Grouselle, D., Faivre-Bauman, A. and Tixier-Vidal, A. (19781 A
radioimmunoassay for thyroliberin (TRH). Comparison with a
TRH radioreceptor assay. Neurosci. Lett. 7, 7.
Grouselle. D., Tixier-Vidal, A. and Pradelles, Ph. (19821 A new
improvement of the sensitivity and specificity of radioim-
munoassay for thyroliberin. Application to biological samples.
Neuropeptides 3. 29.
Grouselle. D.. Tixier-Vidal. A. and Pradelles. Ph. (1990) Enzyme
immunoassays for thyroliberin (TRH) and TRH-elongated
peptides in mouse and rat hypothalamus. Neuropeptides 17.
155.
Khan, Z., Aitken, A., Del Rio Garcia, J. and Smyth, D.G. (1992)
Isolation and identification of two neutral thyrotropin releasing
hormone-like peptides, pyroglutamylphenylalanineproline
amide and pyroglutamylglutamine-proline amide. from human
seminal fluid. J. Biol. Chem. 267. 7464.
Massoulie, J. and Bon. S. (19761 Affinity chromatography of
acetylcholinesterase. The importance of hydrophobic interac-
tions. Eur. J. Biochem. 68, 531.
Morley. J.E. (1979) Extrahypothalamic thyrotropin releasing hor-
mone (TRH). Its distribution and its functions. Life Sci. 25.
1539.
Pradelles, Ph. (1995) Immunometric determination of an antigen
or hapten. United States Patent No. 5, 476, 770.
Pradelles, Ph., Grassi. J.. Chabardes. D. and Guiso. N. (1989)
Enzyme immunoassays of adenosine cyclic 3’,5’-monophos-
phate and guanosine cyclic 3’.5’-monophosphate using acetyl-
cholinesterase. Anal. Chem. 61. 447.
Pradelles, Ph., Grassi, J., CrCminon, C., Boutten. B. and Mamas,
S. (19941 Immunometric assay of low molecular weight hap-
tens containing primary amino groups. Anal. Chem. 66, 16.
Reyes-Parada, M., Scorza, M.C., Silveira, R., Dajas, F., Costa, G.,
Tipton, K.F. and Cassels, B.K. (19941 Monoamine oxidase
inhibitory effects of some 4-aminophenethylamine derivatives.
Biochem. Pharmacol. 47, 1365.
Roland, J., Grouselle, D.. Gourdji, D., Tixier-Vidal, A. and
Cazenave, P.A. (1992) Properties of monoclonal antibodies to
thyroliberin (TRH) induced by different immunogens: compar-
ison with pituitary TRH receptor. Mol. Immunol. 29. 463.
Visser, T.J. and Klootwijk, W. (1981) Approaches to a markedly
increased sensitivity of the radioimmunoassay for thyrotropin-
releasing hormone by derivatization. Biochem. Biophys. Acta
673. 454.
Volland, H., Vulliez Le Normand, B., Mamas, S., Grassi. J..
Creminon. C., Ezan, E. and Pradelles, Ph. (19941 Enzyme
immunometric assay for leukotriene C4. J. Immunol. Methods
175, 97.