elämän energiatalous
TRANSCRIPT

2.2. Elämän energiatalous
Vesikin on kiinnostava molekyyli
Jatkossa tulemme tutustumaan ilmiöihin, joiden ymmärtämiseksi on
tärkeätä ensin tutustua vesimolekyylin ominaisuuksiin.
Vesimolekyyli muistuttaa rakenteeltaan Mikki-Hiirtä. Happiatomi
muodostaa Mikki-Hiiren kasvot ja vedyt toimivat sen korvina. Vetyjen ja
hapen väliset sidoselektronit eivät liiku atomien ympärillä tasaisesti, vaan
ne viettävät suurimman osan ajastaan happiatomin ympärillä. Syy tähän
”epädemokraattiseen” asiaintilaan on happiatomin selvästi vetyä
suuremmassa elektronegatiivisuudessa (ilmiön syistä voit neuvotella
fysiikanopettajasi kanssa).
Elektronien epätasainen jakautuminen aiheuttaa varauseron
vesimolekyylin vety- ja happipuolen kesken. Se puoli, missä vedyt
sijaitsevat, on varaukseltaan positiivinen verrattuna happipuoleen, mikä
taas on varaukseltaan negatiivinen. Näin yksittäisestä vesimolekyylistä
tulee pienoismagneetti plus- ja miinusnapoineen (kuva 8).

H 2 O
Kuva 8. Vesimolekyylin rakennemalli
Magneettisuutensa vuoksi joukko vesimolekyylejä ryhmittyy mielellään
ketjuksi. Ilmiö havaitaan vedessä pintajännityksenä. Veden
magneettisuutta hyödyntävät kasvit vesitaloudessaan ja kaikki solut
kalvorakenteittensa (solukelmu, tumakotelo, solulimakalvosto)
muodostamisessa.
Monet elämän ilmiöt tarvitsevat protoneita
Vetyatomi on atomeista kaikkein yksinkertaisin (kuva 9) . Sen ytimessä
sijaitsee yksi positiivinen hiukkanen nimeltä protoni (H+ ). Protonia
kiertää yksi negatiivinen hiukkanen nimeltä elektroni (e-). Aiemmin jo
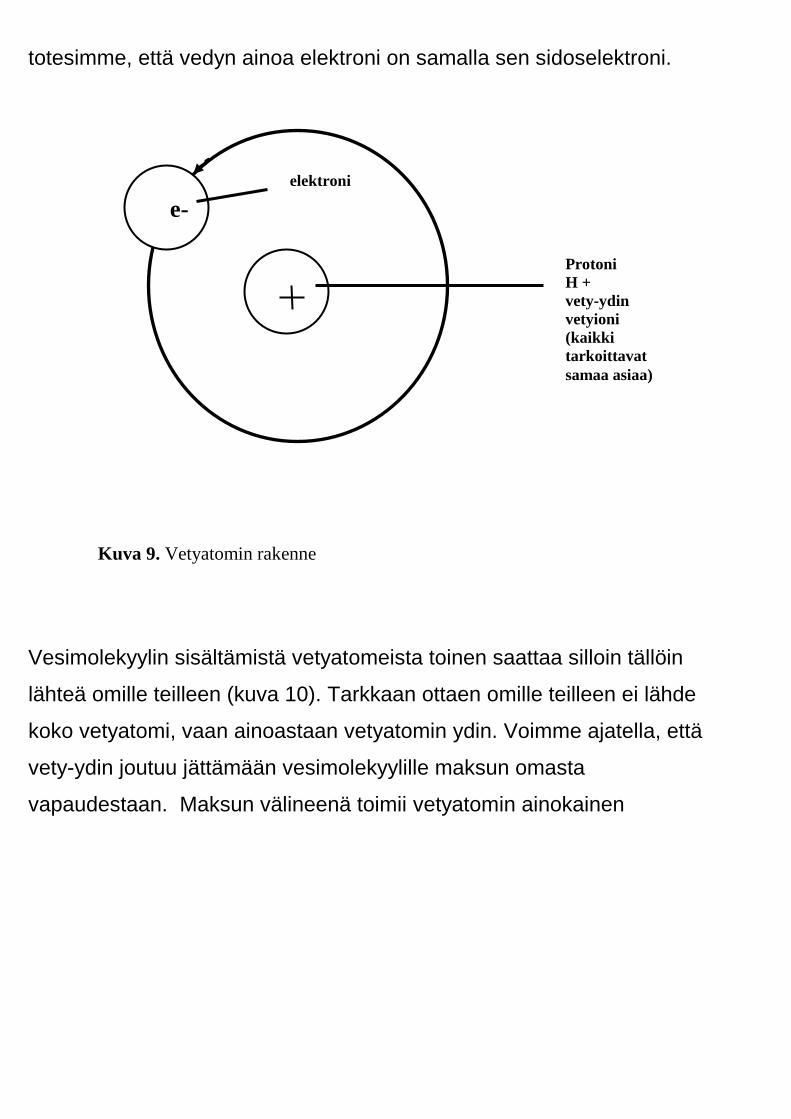
totesimme, että vedyn ainoa elektroni on samalla sen sidoselektroni.
Protoni
H +
vety-ydin
vetyioni
(kaikki
tarkoittavat
samaa asiaa)
Kuva 9. Vetyatomin rakenne
e-
elektroni
Vesimolekyylin sisältämistä vetyatomeista toinen saattaa silloin tällöin
lähteä omille teilleen (kuva 10). Tarkkaan ottaen omille teilleen ei lähde
koko vetyatomi, vaan ainoastaan vetyatomin ydin. Voimme ajatella, että
vety-ydin joutuu jättämään vesimolekyylille maksun omasta
vapaudestaan. Maksun välineenä toimii vetyatomin ainokainen

sidoselektroni.
H 2 O
Kuva 10. Veden hajoaminen protoniksi ja OH-
ioniksi (veden autoprotolyysi).
e-
H+
OH -
Edellä kuvatusta veden hajoamisesta käytetään nimitystä veden
autoprotolyysi (suomennettuna ”itsestään tapahtuva protonin luovutus”).
Sen seurauksena vesimolekyylistä syntyy kaksi erimerkkistä hiukkasta:
positiivinen protoni H+ ja negatiivinen OH- (kuva 10).
Pian tulet huomaamaan, että protonit ovat solun energiatalouden
perusta. Protoneiden olemassaoloon perustuu myös kasvien
ravinnetalous ja happosateiden aiheuttamat haitat. Tässä joitakin vesi- ja
protonikatsaukseni syitä..
Silloin tällöin vastaasi tulee molekyylikaavoja, joissa esiintyy plus- tai
miinusmerkkejä. Ne ovat kohtia, joista on irronnut tai joihin on kiinnittynyt
protoneita tai elektroneita varauksen ilmoittama määrä. Varauksellisia
atomeja tai molekyylikaavoja ei siis tarvitse sen kummemmin arastella.

Mihin solut tarvitsevat energiaa?
Proteiinit ovat solun työkaluja. Kuten tavallisetkin työkalut, eivät
proteiinitkaan yleensä toimi itsestään, vaan tarvitsevat käyttövoimaa.
Käyttövoima on peräisin auringosta.
Vain harvat ja valitut proteiinit pystyvät hyödyntämään auringon
säteilyenergiaa sellaisenaan. Siksi valon sisältämä energia on sidottava
tavallisillekin proteiineille käyttökelpoiseen muotoon,
energiamolekyyleiksi.
Energiamolekyyleistä tärkeimmät ovat ATP ja sokerit. Tutustumme vasta
myöhemmin näiden molekyylien rakenteeseen. Säilyvyydeltään ja
käyttöominaisuuksiltaan ne eroavat selvästi toisistaan. ATP hajoaa hyvin
nopeasti aivan itsestäänkin. Se säilyy käyttökelpoisena vain n. 20
sekuntia. Sokerit säilyvät hyvin (ajattelepa vaikka kaikkia kesämökkien
hyllyillä vuosikausia seisoneita ikivanhoja sokeripaketteja).
Energiahävikkiä välttääkseen eliöt varastoivat energiaa sokereina. Kun
energiaa tarvitaan (proteiinien käyttövoimaksi siis), sokerimolekyylejä
käsitellään tavalla, jonka tuloksena syntyy ATP:tä. Näin on meneteltävä
siksi, että jostakin syystä proteiinit pystyvät hyödyntämään energiaa vain
ATP-muotoisena (tämä toteamus ei kuitenkaan ole aivan ehdoton).
Menetelmä, jolla sokereita tuotetaan, on nimeltään yhteyttäminen.
Menetelmä, jolla ATP:tä tuotetaan on nimeltään soluhengitys. Alla (kuva
11) on uudelleen jo aikaisemmin geenien toiminnan yhteydessä esillä

ollut kaavio, joka ilmentää soluissa tavattavien yhdisteiden välisiä
riippuvuussuhteita. Proteiinit sijaitsevat kaaviossa keskellä, sillä ne
varsinaisesti aiheuttavat kaikki elämälle ominaiset toiminnot.
Energia-aineenvaihduntaan liittyvien aineiden kierrot näkyvät kuvan
oikeassa laidassa.

Kuva 2. Mistä elämässä on kysymys?
Rasva-aineet eli tapahtumien näyttämönä toimivat
kalvorakenteet esim. solukelmu, solulimakalvosto,
tumakotelo, mitokondrioiden ja viherhiukkasten
sisältämät kalvostot
Rasva-aineiden tuotanto
Proteiinisynteesi eli
proteiinien tuotanto
- tapahtuu kahden
proteiinityypin toimesta:
1) DNA:n ”lukijat”
(RNA-polymeraasi)
2) Proteiinien
rakentajat
(ribosomit)
- proteiinien tekemiseen
tarvitaan siis proteiineja
→ muna-kana -ongelma
Yhteyttäminen
tapahtuu kasvien
viherhiukkasissa
auringon valossa
Sokerit eli
energiavarasto
Soluhengitys
tapahtuu
mitokondrioissa
CO2
H2O
O2
ATP eli
proteiinien
käyttövoima
Proteiinit eli solun
työkalut
- yhdenlainen työkalu
pystyy tekemään vain
yhdenlaisen työn
DNA:n kopiointi eli DNA-
replikaatio
- tapahtuu muutaman eri
proteiinin toimesta, joista
tärkein on DNA-
polymeraasi
- geenit ovat DNA:sta
koostuvia proteiinien
rakenneohjeita, joten nämä
proteiinit rakentavat omia
rakenneohjeitaan → muna-
kana –ongelma
- DNA-replikaatio on
lisääntymisen edellytys ja
perusidea
Nukleiinihapot DNA ja RNA
- DNA:sta koostuvat geenit eli
proteiinien rakenneohjeet
- RNA on yksittäisestä geenistä
tehtävä lyhytikäinen nauhoite
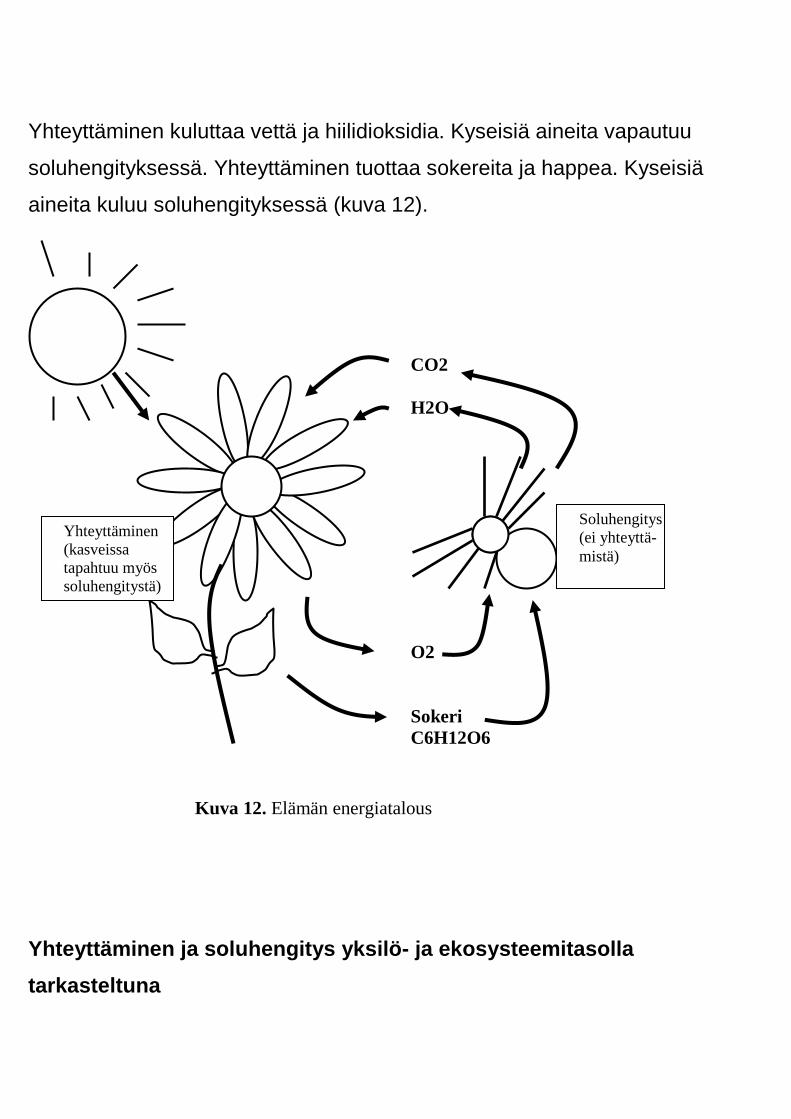
Yhteyttäminen kuluttaa vettä ja hiilidioksidia. Kyseisiä aineita vapautuu
soluhengityksessä. Yhteyttäminen tuottaa sokereita ja happea. Kyseisiä
aineita kuluu soluhengityksessä (kuva 12).
CO2
H2O
O2
Sokeri
C6H12O6
Yhteyttäminen
(kasveissa
tapahtuu myös
soluhengitystä)
Soluhengitys
(ei yhteyttä-
mistä)
Kuva 12. Elämän energiatalous
Yhteyttäminen ja soluhengitys yksilö- ja ekosysteemitasolla
tarkasteltuna

Yhteyttäminen ja soluhengitys pyörittävät koko maailmantaloutta. Vain
kasvit ja eräät muut yhteyttävät (omavaraiset eli autotrofiset) eliöt
pystyvät sitomaan auringon energiaa sokereiksi. Kaikki muut
(toisenvaraiset eli heterotrofiset) eliöt ovat riippuvaisia yhteyttämisessä
syntyvistä sokereista. Koko ekosysteemin toiminta perustuu
yhteyttämiseen ja soluhengitykseen.
Riippuvuussuhteen ymmärrät pysähtymällä pohtimaan, mikä liikuttaa
polkupyörää, autoa tai käkikelloa. Kaikissa niissä on kyse omasta
lihastyöstäsi (joka vaatii ATP:tä, siis sokereita). Autossa työ rajoittuu
tosin polkimien painamiseen ja ajatteluun. Yllättäen kuitenkin myös
bensiinin sisältämä energia on alun perin yksisoluisten merenpohjaan
vajonneiden levien yhteyttämällä tuottamaa sokeria eli auringon
säteilyenergiaa.
Huomaa, että edellä oleva kuva 12 havainnollistaa elämän
energiataloutta paitsi yksilö- myös ekosysteemitasolla.
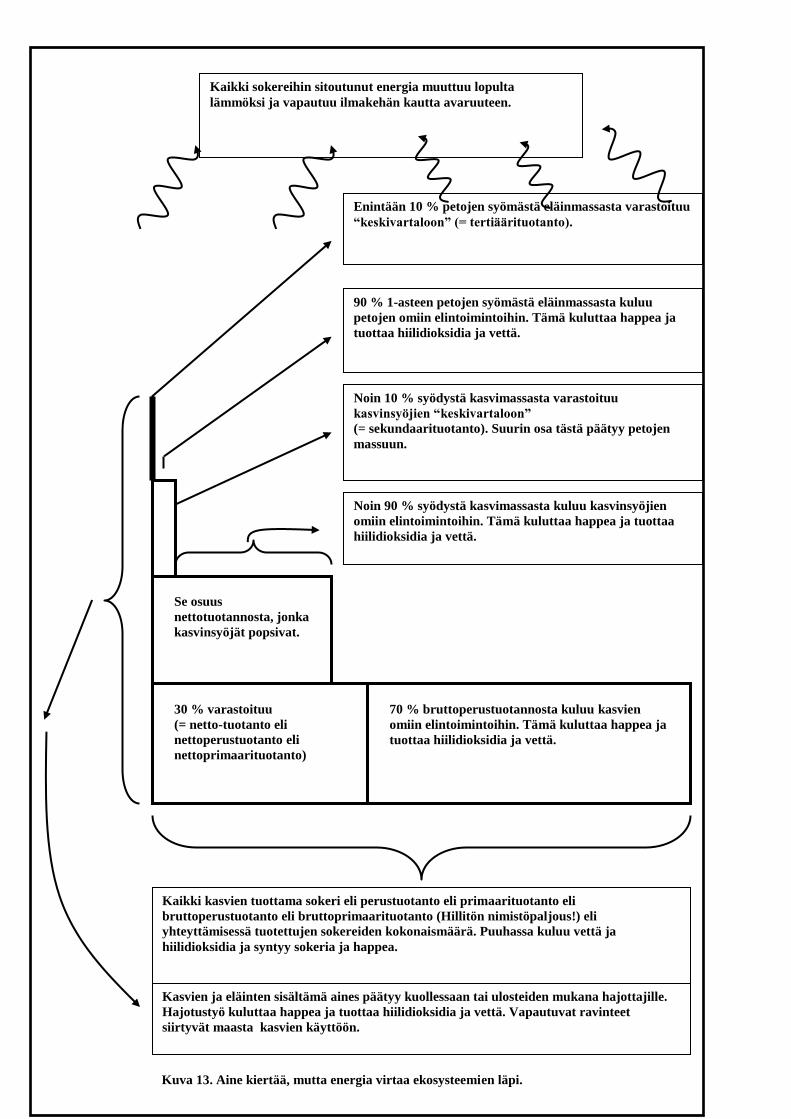
Seuraavakin kaavio (kuva 13) havainnollistaa omalla tavallaan, miten
elollisessa luonnossa aineet kiertävät, mutta energia virtaa eliöiden läpi.
Tässä kaaviossa näkökulma on ekologinen. Auringon säteilyenergia
viivähtää meissä vain hetkisen sokereiden muodossa ja päätyy lopulta
elintoimintojemme tuottamana lämpöenergiana avaruuteen. Sama
kohtalohan olisi auringon säteilyenergialla ilman meitäkin. Kaaviossa on
myös termejä, joita ekologit käyttävät elämän energiataloutta
kuvatessaan. Kaaviossa käyttämäni kirjasinkoko on sen verran pieni,
että ainakin pienikokoiselta näyttöruudulta voi tekstien lukeminen tuottaa
hankaluuksia. Suosittelen tällöin kaavion tulostamista.
Kaikki sokereihin sitoutunut energia muuttuu lopulta
lämmöksi ja vapautuu ilmakehän kautta avaruuteen.
Enintään 10 % petojen syömästä eläinmassasta varastoituu
“keskivartaloon” (= tertiäärituotanto).
90 % 1-asteen petojen syömästä eläinmassasta kuluu
petojen omiin elintoimintoihin. Tämä kuluttaa happea ja
tuottaa hiilidioksidia ja vettä.
Noin 10 % syödystä kasvimassasta varastoituu
kasvinsyöjien “keskivartaloon”
(= sekundaarituotanto). Suurin osa tästä päätyy petojen
massuun.
Noin 90 % syödystä kasvimassasta kuluu kasvinsyöjien
omiin elintoimintoihin. Tämä kuluttaa happea ja tuottaa
hiilidioksidia ja vettä.
Kaikki kasvien tuottama sokeri eli perustuotanto eli primaarituotanto eli
bruttoperustuotanto eli bruttoprimaarituotanto (Hillitön nimistöpaljous!) eli
yhteyttämisessä tuotettujen sokereiden kokonaismäärä. Puuhassa kuluu vettä ja
hiilidioksidia ja syntyy sokeria ja happea.
30 % varastoituu
(= netto-tuotanto eli
nettoperustuotanto eli
nettoprimaarituotanto)
70 % bruttoperustuotannosta kuluu kasvien
omiin elintoimintoihin. Tämä kuluttaa happea ja
tuottaa hiilidioksidia ja vettä.
Se osuus
nettotuotannosta, jonka
kasvinsyöjät popsivat.
Kasvien ja eläinten sisältämä aines päätyy kuollessaan tai ulosteiden mukana hajottajille.
Hajotustyö kuluttaa happea ja tuottaa hiilidioksidia ja vettä. Vapautuvat ravinteet
siirtyvät maasta kasvien käyttöön.
Kuva 13. Aine kiertää, mutta energia virtaa ekosysteemien läpi.

Yhteyttäminen ja soluhengitys soluelinten tasolla tarkasteltuna
Soluissa esiintyy kaksi energia-aineenvaihduntaan erikoistunutta
soluelintä: mitokondrio ja viherhiukkanen. Kumpaakin näistä tavataan
yhteyttävissä soluissa. Viherhiukkasten sisältämä lehtivihreä saa
yhteyttävät solut näyttämään vihreiltä.
Kaikissa muissa aitotumallisissa eliöissä paitsi kasveissa on pelkkiä
mitokondrioita. Viherhiukkaset puuttuvat.
Yksittäisissä soluissa näitä soluelimiä voi olla valtavasti. Esimerkiksi
lihassoluissa mitokondrioita suorastaan tungeksii. Viherhiukkasten ja
mitokondrioiden välistä työnjakoa havainnollistaa kuva 14.

ATP
ATP
CO2
H2O
O2
Sokeri
C6H12O6
Yhteyttäminen
- viherhiukka-
sissa, vain
kasveissa ja
eräissä
Protoctistoissa
esim. levissä
Soluhengitys
- aitotumallisten
eliöiden (siis myös
kasvien)
mitokondrioissa,
bakteereilla
tapahtuu solun
ulkokalvolla
Kuva 14. Elämän energiatalous.
Viherhiukkasen ja mitokondrion välinen
tehtävänjako.
Viherhiukkasten ja mitokondrioiden rakenne
Mitokondriot ja viherhiukkaset muistuttavat rakenteeltaan kovasti
toisiaan. Kumpikin soluelin koostuu ulko- ja sisäkalvosta.
Mitokondrioiden sisäkalvo on löysillä poimuilla. Niitä kutsutaan kristoiksi.
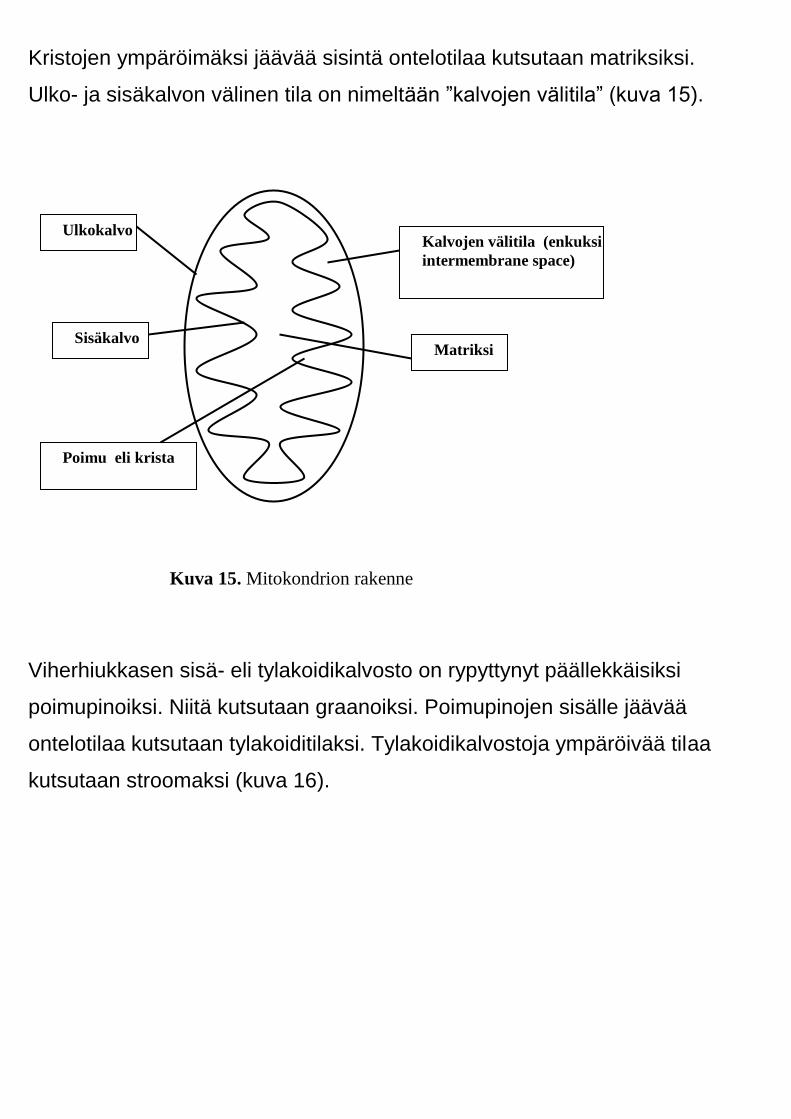
Kristojen ympäröimäksi jäävää sisintä ontelotilaa kutsutaan matriksiksi.
Ulko- ja sisäkalvon välinen tila on nimeltään ”kalvojen välitila” (kuva 15).
Kalvojen välitila (enkuksi
intermembrane space)
Poimu eli krista
Matriksi
Kuva 15. Mitokondrion rakenne
Ulkokalvo
Sisäkalvo
Viherhiukkasen sisä- eli tylakoidikalvosto on rypyttynyt päällekkäisiksi
poimupinoiksi. Niitä kutsutaan graanoiksi. Poimupinojen sisälle jäävää
ontelotilaa kutsutaan tylakoiditilaksi. Tylakoidikalvostoja ympäröivää tilaa
kutsutaan stroomaksi (kuva 16).

Tylakoidikalvosto
eli graana Strooma
Tylakoiditila
Kuva 16. Viherhiukkasen rakenne
Ulkokalvo
Sisäkalvo
Kumpikin elin on kooltaan bakteerin luokkaa ja sisältää pienen
rengasmaisen DNA-jakson. Rengasmaista DNA-jaksoa kutsutaan
plasmidiksi ja yleensä sellainen on tunnusomainen bakteereille.
Mitokondrioiden ja viherhiukkasten DNA:ssa on joukko toimivia geenejä.
Niissä on joidenkin energia-aineenvaihdunnassa tarvittavien proteiinien
rakenneohjeet. (Tällaisia ovat tässä tekstissä tuonnempana esiteltävän
elektroninsiirtoketjun proteiinit. Ne rakentuvat modulaarisesti. Kukin
elektroninsiirtäjä on nimittäin useamman pienemmän osaproteiinin
muodostama kokonaisuus, vähän niin kuin pantaisiin useita Lego-
palikoita, toistuvia moduleita, kiinni toisiinsa. Joitakin näistä
osaproteiineista koodaavat sellaiset geenit, jotka sijaitsevat tumassa.
Jotkin samaisen elektroninsiirtäjän osaproteiineista voivat olla
koodattuina geeneihin, jotka ovat mitokondrion tai viherhiukkasen
omassa DNA-molekyylissä.).

Pieni koko ja plasmidien olemassaolo viittaavat siihen, että mitokondriot
ja viherhiukkaset saattavat evolutiivisesti alun perin olla aitotumalliseen
soluun asettuneita yhteistyöbakteereita (ns. endosymbioositeoria).
Viherhiukkasten ja mitokondrioiden toiminta
Johdanto
Viherhiukkasen ja mitokondrion toiminnassa on paljon
samankaltaisuuksia. Silmiinpistävää on molempien kyky tuottaa ATP:tä
vieläpä hyvin samantapaisella menetelmällä. Tuotantotapaan
tutustumme tuonnempana.
Sekä yhteyttämiseen että soluhengitykseen sisältyy myös vaihe, jota
kutsutaan sykliseksi (renkaanmuotoiseksi). Myös tämä on hyvin
samankaltainen kummassakin soluelimessä.
Syklisyydellä tarkoitetaan sitä, että reaktiokulku tuottaa kaksi eri
lopputuotetta (kuva 17). Ensimmäinen (reaktion varsinainen lopputuote)
poistuu reaktiokulusta ensin. Myöhemmin muodostuvaa toista
lopputuotetta tarvitaan kuitenkin myös, koska se on reaktiokulun
(seuraavan kierroksen) tärkeä lähtöaine. Siksi, vaikka perimmäisenä
päämääränä onkin tuottaa kierrosta ensimmäisenä poistuvaa tuotetta, ei
reaktiokulku voi pysähtyä vielä tähän.
Alla oleva kaavio (kuva 17) kuvaa syklisen reaktiokulun yleisperiaatteen.
Sykliset reaktiokulut ovat elämälle luonteenomaisia muutenkin kuin
energia-aineenvaihdunnan osalta.

1. Muokattava
yhdiste
1. ”Kelkkamolekyyli”
4. ”Kelkkamolekyylin” esiaste
3. Valmis
reaktiotulos
2. Muokattava yhdiste liitettynä
”kelkkamolekyyliin”
Kuva 17. Syklisen reaktiokulun perusmalli
ATP:n rakenne, tuotanto ja merkitys
Tutustumme nyt elämän energiatalouteen molekyylitasolla. Aloitamme
urakkamme ATP-molekyylistä.
ATP on ainoa proteiinien käyttövoimaksi sopiva molekyyli. Siksi voimme
kuvitella, että yhteyttämisen ja soluhengityksen perimmäinen tavoite on
ATP:n tuottaminen.
ATP (adenosiinitrifosfaatti) on rakenteeltaan RNA-nukleotidi, jossa on
adeniiniemäs (kuva 18a). Nukleotidit ovat DNA:n ja RNA:n rakenneosia.
Ellet vielä ole tutustunut DNA:n ja RNA:n rakenteeseen, voit aivan hyvin
jättää sen edelleen tuonnemmaksi ja tässä yhteydessä tutustua
nukleotidiin pelkkänä sanana.

Proteiinien toiminta kuluttaa ATP:tä, jolloin se hajoaa kahdeksi osaksi:
ADP:ksi ja P:ksi. Kun ATP:tä tuotetaan, osat yhdistetään uudelleen
ATP:ksi (kuva 18b).
Riboosi
-sokeri
Fosfaatti
Fosfaatti
Fosfaatti
Adeniini-emäs
RNA:n adeniini-
emäksellä
varustettu
nukleotidi
eli AMP ADP
ATP
Kuva 18a. ATP (adenosiini-tri-fosfaatti) on Adeniini-
emäksellä varustettu RNA-nukleotidi, johon vain on liitetty
kaksi ylimääräistä fosfaattiryhmää. Yhden fosfaatin
lisääminen tuottaa ADP:n (adenosiini-di-fosfaatti).
Lähtöaineena toimivaa RNA-nukleotidia voidaan kutsua
myös nimellä AMP(adenosiini-mono-fosfaatti).

P
P
ADP
+
P
ATP
P
P
1) Aineiden
kuljetus
2) Liikkuminen
(supistumiskykyiset
säikeet)
A B + P A B
3) Kemiallinen työ
(= pikkumolekyyleistä
kootaan suurempia)
Mitokondrioihin ATP:n
uudelleentuotantoa varten
Kuva 18b. ATP:n käyttötavat soluissa. Solussa työtä tekevät sen
sisältämät proteiinit. Työ vaatii energiaa. Proteiinit pystyvät
hyödyntämään energiaa vain ATP:n muodossa. Työn tapahtuessa ATP
hajoaa perusosasikseen ADP:ksi ja P:ksi. Nämä kierrätetään
uusiokäyttöön mitokondrioihin, missä niistä valmistetaan uutta ATP:tä.
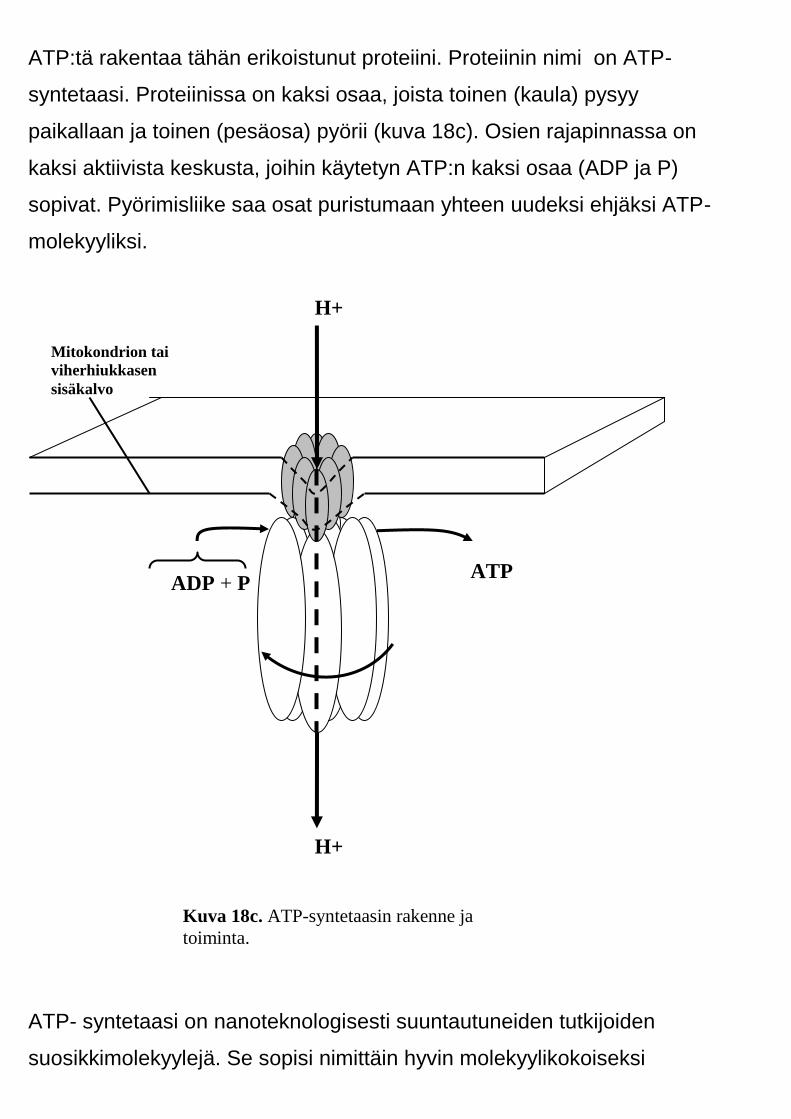
ATP:tä rakentaa tähän erikoistunut proteiini. Proteiinin nimi on ATP-
syntetaasi. Proteiinissa on kaksi osaa, joista toinen (kaula) pysyy
paikallaan ja toinen (pesäosa) pyörii (kuva 18c). Osien rajapinnassa on
kaksi aktiivista keskusta, joihin käytetyn ATP:n kaksi osaa (ADP ja P)
sopivat. Pyörimisliike saa osat puristumaan yhteen uudeksi ehjäksi ATP-
molekyyliksi.
ATP ADP + P
H+
H+
Mitokondrion tai
viherhiukkasen
sisäkalvo
Kuva 18c. ATP-syntetaasin rakenne ja
toiminta.
ATP- syntetaasi on nanoteknologisesti suuntautuneiden tutkijoiden
suosikkimolekyylejä. Se sopisi nimittäin hyvin molekyylikokoiseksi

moottoriksi tulevaisuudessa tehtäviin pienenpieniin laitteisiin. ATP-
syntetaasi on sikäli ainutlaatuinen proteiini, että sen toiminta tosiaankin
perustuu pyörimisliikkeeseen. Pyörimisliikettä elävä luonto ei
nykykäsityksen mukaan hyödynnä missään muualla.
Mikä pyörittää ATP-syntetaasia?
ATP-syntetaasimolekyylejä tavataan mitokondrioiden ja viherhiukkasten
sisäkalvolla. Molekyylin naapurissa sisäkalvolle kiinnittyneinä on myös
rivi pienempiä proteiineja. Näitä kutsutaan elektroninsiirtäjäentsyymeiksi
(kuva 19). ATP-syntetaasin pyöriminen perustuu elektroninsiirtäjien
antamaan apuun. Näiden lisäksi paikalle tarvitaan vetyatomeja.

H+
e- e- e- e-
H e-
H+
H
ADP
+
P
ATP
Elektroninsiirtä-jäentsyymi
ATP-syntetaasi
Elektroninsiirtoketjusta tuleva vetyatomi
päätyy yhteyttämisessä sokerin osaksi ja
soluhengi-tyksessä vesimolekyyliin
Vetyatomi
elektroninsiirtoketjuun
saadaan joko vedestä
(yhteyttämisessä) tai
sokerista
(soluhengityksessä)
Kuva 19. Mitokondrion ja viherhiukkasen sisäkalvon
rakenne ja ATP-tuotannon perusperiaate.
Mitokondrion tai
viherhiukkasen sisäkalvo
Perusidea on, että elektroninsiirtäjäentsyymit riistävät elektroneja
vetyatomeilta, jolloin vedystä jää jäljelle pelkkä vedyn ydin eli protoni.
Elektroninsiirtäjät toimittavat elektronit ja protonit, niin mitokondriossa
kuin viherhiukkasissakin, sisäkalvon vastakkaisille puolille.
Varaukseltaan vastakkaismerkkisinä nämä hiukkaset alkavat vetää
toisiaan puoleensa.
Sisäkalvo on niin tiivis, etteivät elektronit ja protonit voi siirtyä sen läpi.
On vain yksi paikka, missä virtausliike on mahdollinen ja siinäkin vain

yhteen suuntaan. Paikka on ATP-syntetaasi. Tämän pohjatonta pulloa
muistuttavan proteiinin läpi sopivat ainoastaan protonit. Ne alkavatkin
virrata menettämiensä elektronien luo. Protonien liike saa ATP-
syntetaasin pesäosan pyörimään (periaate on sama kuin
vesivoimalassa). Tällöin pesän ja kaulan rajapinnassa ADP ja P
yhdistyvät jälleen uudeksi ATP:ksi.
Elektroninsiirtäjiä tarvitaan kokonainen joukko. Suurin osa niistä on
proteiineja, eivät kuitenkaan aivan kaikki. Elektroninsiirtäjiä kutsutaan
usein myös sytokromeiksi.
Elektroninsiirtäjiä voi ajatella joukoksi taskuvarkaita, joista yksi ensin
varastaa uhrin (vetyatomin) lompakon (elektronin). Lompakko
toimitetaan taskuvarkaalta seuraavalle ja taas seuraavalle jne. Uhri
päätyy nopeasti kokonaan eroon lompakostaan (protoni elektronistaan).
Edellä kuvatun kaltaisia ATP:n tuotantolinjoja viherhiukkasten ja
mitokondrioiden sisäkalvot ovat täpötäynnä. Tuotantotavan virallinen
nimi on kemiosmoosi (Chemiosmosis). Myös nimeä oksidatiivinen
fosforylaatio käytetään. Näpyttele nimitykset jollekin hakupalvelimelle,
niin saat varmasti tutkittaviksesi paljon aihetta sivuavia tiedostoja.
Viherhiukkasissa, toisin kuin mitokondrioissa, elektroninsiirtäjistä
ensimmäisenä on väriltään vihreitä klorofylli- eli lehtivihreämolekyylejä.
Ne ovat ryhmittyneet hieman lautasantennia muistuttaviksi 200 – 300
molekyylin suuruisiksi ryhmiksi.

Jokaisen yksittäisen lehtivihreämolekyylin keskellä on magnesium-
atomi. Edellä mainitun 200 – 300 molekyylin kokoisen
lehtivihreämolekyyliryhmän keskellä on yksi lehtivihreämolekyylipoloinen.
Tämän ”muiden saartamaksi joutuneen” lehtivihreämolekyylin sisältämän
magnesium-atomin yksi elektroni virittyy auringon valon vaikutuksesta.
Virittyminen tarkoittaa tapahtumaa, jossa elektroni siirtyy kauaksi
atomiytimestään. Kaukana harhaileva magnesiumin elektroni napataan
elektroninsiirtoketjuun. Lehtivihreä ei kuitenkaan tyydy tilanteeseensa,
vaan panee vahingon kiertämään ottamalla uuden elektronin
vesimolekyylin sisältämiltä vetyatomeilta, kummaltakin vuorollaan. Nämä
uudetkin elektronit joutuvat kuitenkin nopeasti elektroninsiirtoketjun
vietäviksi.
Tässä rytäkässä vesimolekyyli hajoaa happiatomiksi, kahdeksi protoniksi
ja kahdeksi elektroniksi. Elektronit päätyvät elektroninsiirtoketjuun ja
protonit ATP-syntetaasia pyörittämään. Kun vesimolekyylejä on hajonnut
kaksi, tuloksena on yksi happimolekyyli. Se vapautuu ilmaan meidän
hengitettäväksemme.
ATP:tä ei siis tehdä (sokerin tai veden sisältämistä) protoneista, mutta
protonit kylläkin pyörittävät ”konetta”, joka tekee ATP:tä. ATP:n raaka-
aineita (siis RNA-nukleotideja) me toisenvaraiset eliöt saamme syömällä.
Kasvit ovat tässäkin asiassa omavaraisia. Ne nimittäin saavat ATP:n
raaka-aineet maaperän ravinteista ja tuottamistaan sokereista.
Yhteyttäminen ja soluhengitys erikseen ja yhdessä, osina ja
kokonaisuutena, aarghh!

Olemme tähän mennessä todenneet, että yhteyttäminen ja soluhengitys
ovat tiiviisti toisistaan riippuvaisia tapahtumia. Tarkennamme nyt
näkökulmaamme selvittämällä lähemmin, millaisista osareaktioista
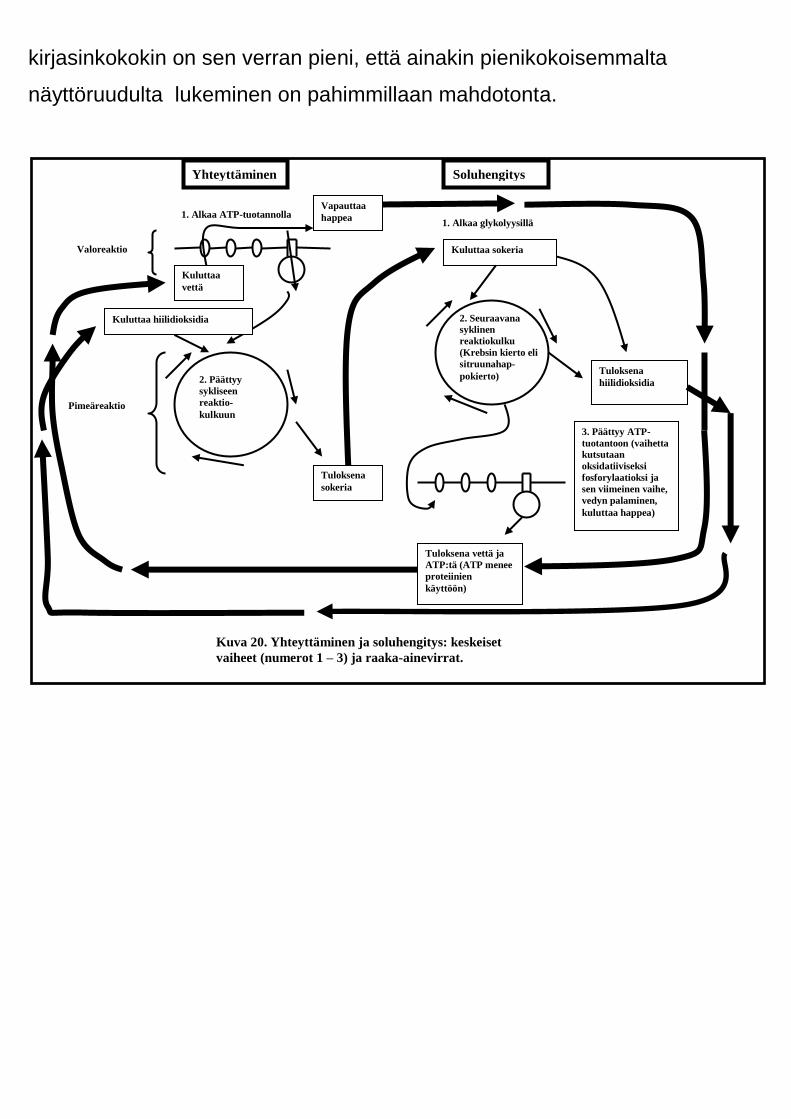
molemmat tapahtumasarjat koostuvat. Alla kuvassa 20 on nimetty
reaktiokulkujen keskeisimmät vaiheet (numerot 1 – 3) sekä osoitettu
keskeisten raaka-aineiden ja lopputuotteiden virtaussuunnat (paksut
nuolet). Vertaa aineiden virtaussuuntia edellä olleissa kuvissa 12 ja 14
esitettyihin. Pane merkille, että tämä kaavio itse asiassa on vain astetta
seikkaperäisempi katsaus kuvien 12 ja 14 esittämiin asioihin. Ennen
kaavioon uppoutumistasi kannattaa tiedostaa, että tarkasteltavissa
reaktioissa on seuraavat päävaiheet.
Yhteyttämiseen kuuluu kaksi päävaihetta:
1) valoreaktio
2) pimeärektio eli Calvinin kierto
Soluhengityksessä on kolme vaihetta:
1) glykolyysi
2) sitruunahappokierto eli Krebbsin kierto
3) ATP:n massatuotanto eli oksidatiivinen fosforylaatio
Käymme vaiheet läpi tässä järjestyksessä. Opintaipaleella karttanamme
toimivat alla olevat vielä edellistäkin tarkemmat esitykset (kuvat 22a,
22b, 22c ja 24). Kuvissa esiintyvien symbolien selitykset löydät kuvasta
21. Suosittelen, että tulostat kuvat 20, 21, 22a, 22b, 22c ja 24
paperille ennen kuin jatkat lukemistasi tämän pitemmälle. Kuvien
kirjasinkokokin on sen verran pieni, että ainakin pienikokoisemmalta
näyttöruudulta lukeminen on pahimmillaan mahdotonta.
Yhteyttäminen
1. Alkaa ATP-tuotannolla
Valoreaktio
2. Päättyy
sykliseen
reaktio-
kulkuun Pimeäreaktio
Tuloksena
sokeria
Soluhengitys
1. Alkaa glykolyysillä
2. Seuraavana
syklinen
reaktiokulku
(Krebsin kierto eli
sitruunahap-
pokierto)
3. Päättyy ATP-
tuotantoon (vaihetta
kutsutaan
oksidatiiviseksi
fosforylaatioksi ja
sen viimeinen vaihe,
vedyn palaminen,
kuluttaa happea)
Tuloksena vettä ja
ATP:tä (ATP menee
proteiinien
käyttöön)
Tuloksena
hiilidioksidia
Kuluttaa
vettä
Kuluttaa hiilidioksidia
Vapauttaa
happea
Kuluttaa sokeria
Kuva 20. Yhteyttäminen ja soluhengitys: keskeiset
vaiheet (numerot 1 – 3) ja raaka-ainevirrat.

ATP-syntetaasi
Lehtivihreä- eli klorofyllimolekyyli
Molekyyli, jonka rungon muodostaa
viisi hiiliatomia. Mustattu pallonen
(=hiiliatomi) on liitetty molekyyliin
viimeisimpänä.
Kuva 21. Merkkien selitykset koskien kuvia 22a, 22b ja 22c.
Elektroninsiirtäjäentsyymi
NADP, NAD ja FAD Vedynsiirtäjäentsyymejä
Yhteyttämisen valoreaktio
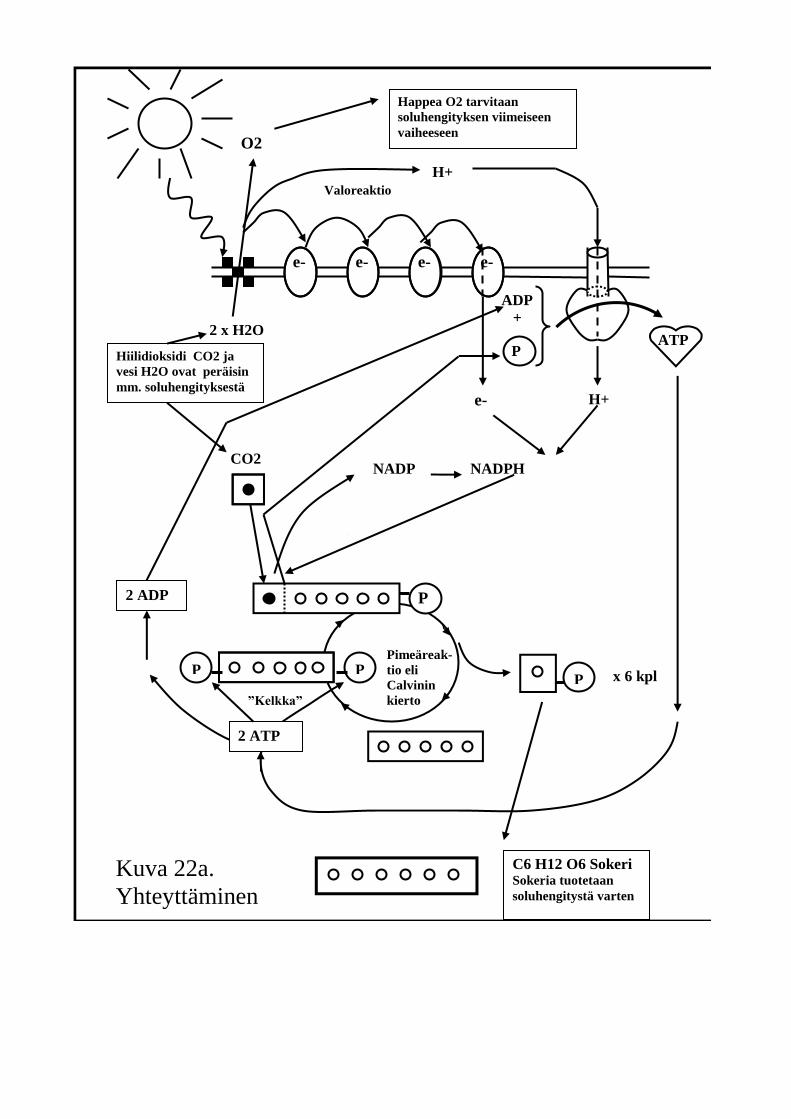
Seuraavassa kaaviossa (Kuvat 22a ja 22b) näet yhteyttämisreaktion kokonaisuudessaan. Kuvat esittävät samaa asiaa, mutta kuva 22a on hieman kuvaa 22b pelkistetympi. Kuvien yläreunassa esitetään tapahtumat, jotka toteutuvat viherhiukkasen sisäkalvon pinnalla. Tätä kutsutaan valoreaktioksi.
C6 H12 O6 Sokeri Sokeria tuotetaan
soluhengitystä varten
Kuva 22a.
Yhteyttäminen
ADP
+
P
H+ e-
NADPH
2 x H2O
O2
H+
e- e- e- e-
CO2
P P P
2 ADP
2 ATP
x 6 kpl
”Kelkka”
NADP
Valoreaktio
Pimeäreak-
tio eli
Calvinin
kierto
Happea O2 tarvitaan
soluhengityksen viimeiseen
vaiheeseen
Hiilidioksidi CO2 ja
vesi H2O ovat peräisin
mm. soluhengityksestä
P
ATP
2 kpl ATP:tä kuluu
”kelkan” fosforyloimiseen
Vettä
maa-
perästä
P
n x ATP
n x H2O
e- H+
H+ H+ H+
H+
H+ H+ n x O2
n x H
n x NADP + H
NADPH
vetyjä valoreaktiosta
Hiilidioksidia
CO2 ilmasta
P
”Kelkka” eli ribuloosi-bis-fosfaatti
P
P
Pimeäreaktio eli
Calvinin kierto
Valoreaktio
P
P
ADP
+ P
2 x ADP
P
P
2 x ADP
2 x ATP
”Kelkan” esiaste
P 5 kpl
P
P
Kuva 22 b.
Yhteyttäminen.
6 kpl = Sokeri (C6 H12 O6)
soluhengitystä varten

Valoreaktion aikana vesimolekyyli hajoaa auringon valon ja lehtivihreän
yhteisvaikutuksesta ja syntyy ATP:tä. Tapahtumasarjan kävimme läpi jo
edellä ATP:n syntytapaan tutustuessamme. Syntyvää ATP:tä
viherhiukkaset tulevat tarvitsemaan yhteyttämisen seuraavassa
vaiheessa: pimeäreaktiossa.
Valoreaktion lopussa “uudelleen syntyvät” vetyatomit ovat samoin
tärkeitä, sillä niitä tarvitaan pimeäreaktiossa sokereiden rakenneosiksi.
Vetyatomit eivät kulkeudu pimeäreaktioon itsestään, vaan ne kuljettaa
sinne molekyyli nimeltä NADP, selkokieliseltä nimeltään
vedynsiirtäjäentsyymi. Vedyllä lastattu vedynsiirtäjä on NADPH. Jos
sinulla olisi nuha, ja yrittäisit lausua sanan ”nappaa”, se kuulostaisi
suurin piirtein samalta kuin ”nadpaa”. NADP on vedynnappaaja
flunssassa!
Vilkaisepa kaaviosta valoreaktion kulku. Pane merkille, miten
vesimolekyylien sisältämät vetyatomit hajoavat protoneiksi ja
elektroneiksi lehtivihreämolekyylien kohdalla. Protoneita ja elektroneita
odottaa iloinen jälleennäkeminen valoreaktion lopussa vedynsiirtäjän
hellässä huomassa.
Sivuhuomiona mainittakoon, että vedynsiirtäjä NADP on ATP:n tapaan
rakenteeltaan nukleotidi. Asia on sikäli mielenkiintoinen, että yleensähän
entsyymit ovat proteiineja.

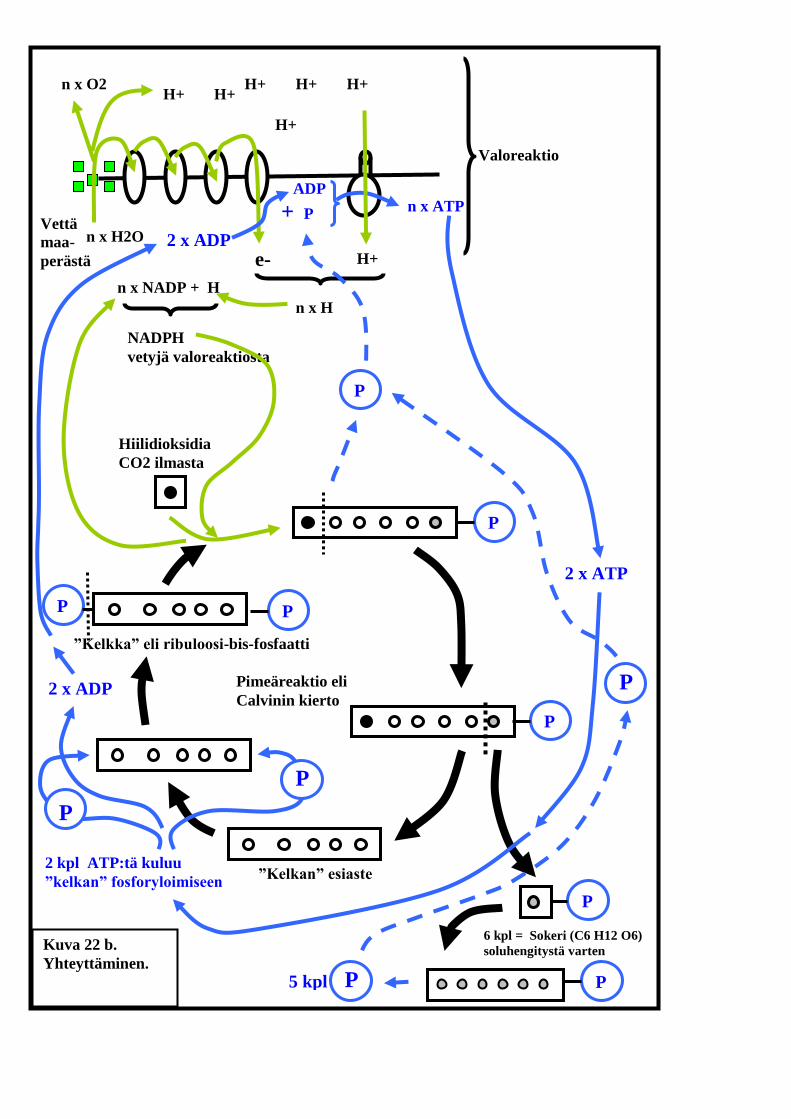
Yhteyttämisen pimeäreaktio
Edellä käsittelemämme valoreaktio tapahtuu tylakoidikalvostojen
pinnalla. Sen aikana protoneita kertyy tylakoiditilaan ja elektroneja
tylakoidikalvojen ulkopuolelle, stroomaan. Pimeäreaktio puolestaan
tapahtuu kokonaan stroomassa ja, nimestään huolimatta, valossa. Nimi
johtuu siitä, että koeolosuhteissa se saadaan tapahtumaan myös
pimeässä, jos vain ATP:tä ja NADPH:ta on tarjolla.
Pimeäreaktio on syklinen reaktiokulku. Syklin keskeltä poistuu reaktion
huipentuma: sokeri. Sokerit ovat hiilihydraatteja. Niissä on siis hiiltä ja
hydrogeniä (vedyn englanninkielinen nimi). Lisäksi sokereissa on
happea. Vety sokereihin saadaan valoreaktiosta. Hiili ja happi ovat
peräisin ilman sisältämästä hiilidioksidista.
Hiilidioksidi ei päädy sokeriin kokonaisena. Toinen hiilidioksidin
sisältämistä happiatomeista päätyy osaksi ”uudistuotantona”
muodostuvaa vesimolekyyliä (hiilidioksidista jää sokeria varten pelkkä
CO eli hiilimonoksidi, siis oikeastaan häkämolekyyli). Tämä vesimolekyyli
on merkitty kuvaan 22c valo- ja pimeäreaktion väliin kaavion oikeaan
reunaan (oikeasti vesi muodostuu vasta pimeäreaktion aikana). Kyseistä
vesimolekyyliä ei ole merkitty kuviin 22a ja b.
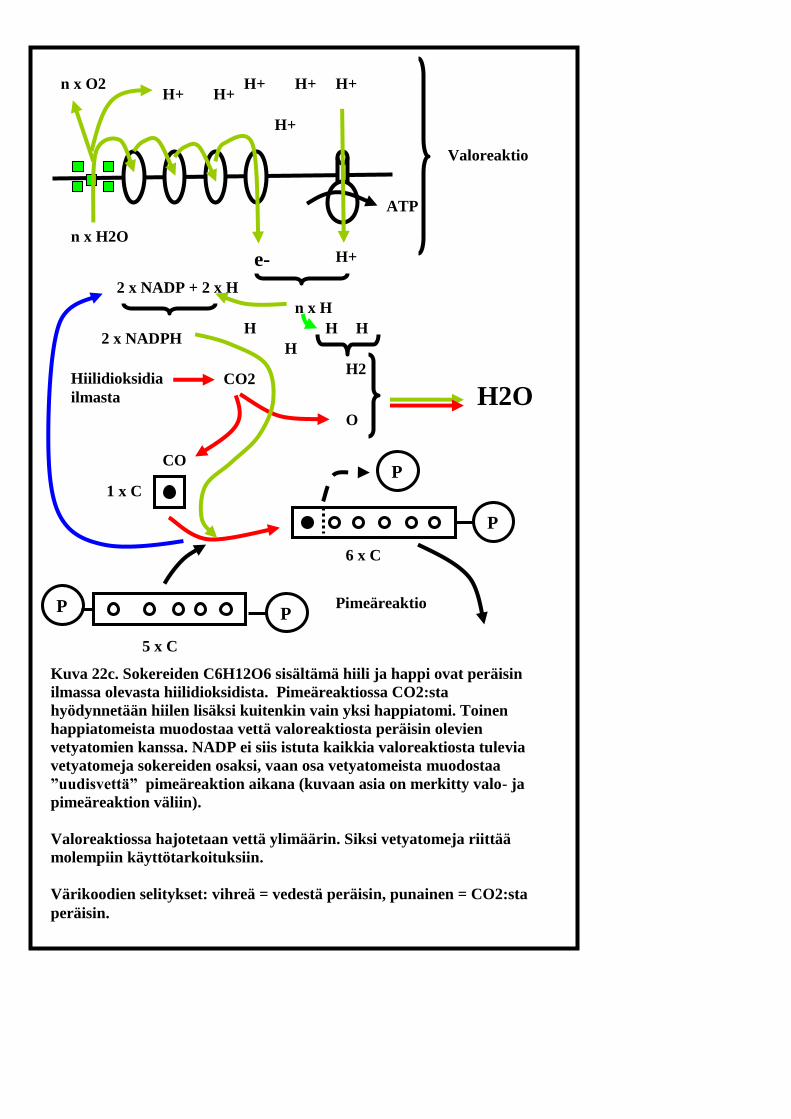
P
ATP
n x H2O
e- H+
H+ H+ H+
H+
H+ H+ n x O2
n x H
H
H
H H
2 x NADP + 2 x H
2 x NADPH
CO2
O
H2
H2O
CO
Hiilidioksidia
ilmasta
P
5 x C
6 x C
1 x C
P
P
Pimeäreaktio
Kuva 22c. Sokereiden C6H12O6 sisältämä hiili ja happi ovat peräisin
ilmassa olevasta hiilidioksidista. Pimeäreaktiossa CO2:sta
hyödynnetään hiilen lisäksi kuitenkin vain yksi happiatomi. Toinen
happiatomeista muodostaa vettä valoreaktiosta peräisin olevien
vetyatomien kanssa. NADP ei siis istuta kaikkia valoreaktiosta tulevia
vetyatomeja sokereiden osaksi, vaan osa vetyatomeista muodostaa
”uudisvettä” pimeäreaktion aikana (kuvaan asia on merkitty valo- ja
pimeäreaktion väliin).
Valoreaktiossa hajotetaan vettä ylimäärin. Siksi vetyatomeja riittää
molempiin käyttötarkoituksiin.
Värikoodien selitykset: vihreä = vedestä peräisin, punainen = CO2:sta
peräisin.
Valoreaktio

Pimeäreaktion alku- ja lopputuote on kaaviooni merkitty laatikkona, jonka
alla kuvissa 22a ja 22b lukee sana ”kelkka”. Laatikko kuvaa molekyyliä,
jonka runkona on viisi hiiliatomia (ympyrät). Palkin molemmissa päissä
on kirjain P. P kuvaa ATP:stä saatua fosfaattiryhmää.
Fosfaattiryhmät ovat tärkeitä, siksi, että ne toimivat houkuttimina
hiilidioksidille ja vetyatomeille. Kun nämä kiinnittyvät kelkan toiseen
päähän (musta ympyrä), houkuttimena toiminut fosfaattiryhmä P irtoaa.
Se siirtyy takaisin valoreaktioon odottamaan vuoroaan päästä uudelleen
ATP:n rakenneosaksi.
Kelkkaan kiinnittyneenä on vielä toinenkin fosfaattiryhmä. Sitä tarvitaan
sokereiden kokoamisessa syklisen osan jälkeen.
Kuvaamassamme mallissa sokerit kootaan yhden hiiliatomin mittaisista
pikkumolekyyleistä. Kokoamisvaiheessa pikkumolekyylit saadaan
kiinnittymään toisiinsa vain fosfaattiryhmien avulla. Siksi sokeriin
(kuusihiiliatominen C6H12O6 molekyyli) päätyvän pikkumolekyylin
kyljessä roikkuu edelleen fosfaattiryhmä P. Valmiissa sokerimolekyylissä
fosfaattiryhmiä olisi jäljellä vain yksi. Loput fosfaatit ovat päätyneet
takaisin valoreaktioon odottamaan pääsyä uusien ATP-molekyylien
rakenneosiksi.
Kelkan sisältämät fosfaattiryhmät ovat siis peräisin valoreaktion
tuottamista ATP-molekyyleistä. ATP:stä jäljelle jääneet ADP:t siirtyvät
nekin valoreaktioon ATP-syntetaasin luo heti kelkan rakennusvaiheen
(fosforyloinnin) jälkeen.

Tarkastelemamme malli seuraa hiilidioksidimolekyylin ja vetyatomien
kohtaloita pimeäreaktion aikana. Käsittelytapa havainnollistaa reaktion
yleistä periaatetta (=sokereiden sisältämä hiili on peräisin ilman
hiilidioksidista ja vety valoreaktiossa hajonneista vesimolekyyleistä).
Jos tarkastelet reaktiota muista lähteistä, huomaat, että niissä yleensä
pimeäreaktio alkaa kolmen CO2-molekyylin samanaikaisella
istahtamisella kolmeen samanlaiseen kelkkamolekyyliin. Näissä
malleissa myös kierrosta poistuvat sokerit ovat aluksi kolmen hiiliatomin
mittaisia ns. triooseja. Trioosien muodostumisen ymmärrät, jos vertaat
pimeäreaktiota sirkuksen jonglööriin, joka heittelee samanaikaisesti
kolmea (hiili)palloa. Samaten, kun hän lopettaa pallotemppunsa, hän
asettaa kerralla kaikki kolme palloa sivuun.
Alla (kuva 23) näet kelkkamolekyylin virallisen rakennekaavan. Oikealta
nimeltään kelkka on ribuloosi-bis-fosfaatti. Etsi rakennekaavasta
hiiliatomit (viisi kappaletta) ja fosfaattiryhmät (kaksi kappaletta) ja vertaa
rakennemallia omaan malliimme ribuloosi-bis-fosfaatista (= kelkka). Jos
molekyylimallit alkavat sinua kiinnostaa, löydät niitä näpyttelemällä mihin
tahansa hakupalvelimeen sanat “dark reaction” tai “Calvin-Benson
Cycle”.

PO3H-
PO3H-
O
O CH2
CH2
C O
C
C
OH
OH
H
H
Kuva 23. Ribuloosi – bis - fosfaatin
rakennekaava
Esityksessämme ei näy, että pimeäreaktiossa kiertävää molekyyliä
muokataan moneen otteeseen reaktiokulun aikana. Muokkauksesta
vastaa joukko proteiineja. Jokainen pieni muutos kohdemolekyylin
rakenteessa on jonkin yksittäisen proteiinin toiminnan tulosta. Koska
kyseessä ovat proteiinit, niiden rakenneohjeet on tallennettu solun
sisältämään DNA:han geeneinä. Nämäkin proteiinit tuotetaan
läpikäymällä kaikki tavanomaiset proteiinisynteesin vaiheet.
Soluhengityksen yleispiirteitä (I)
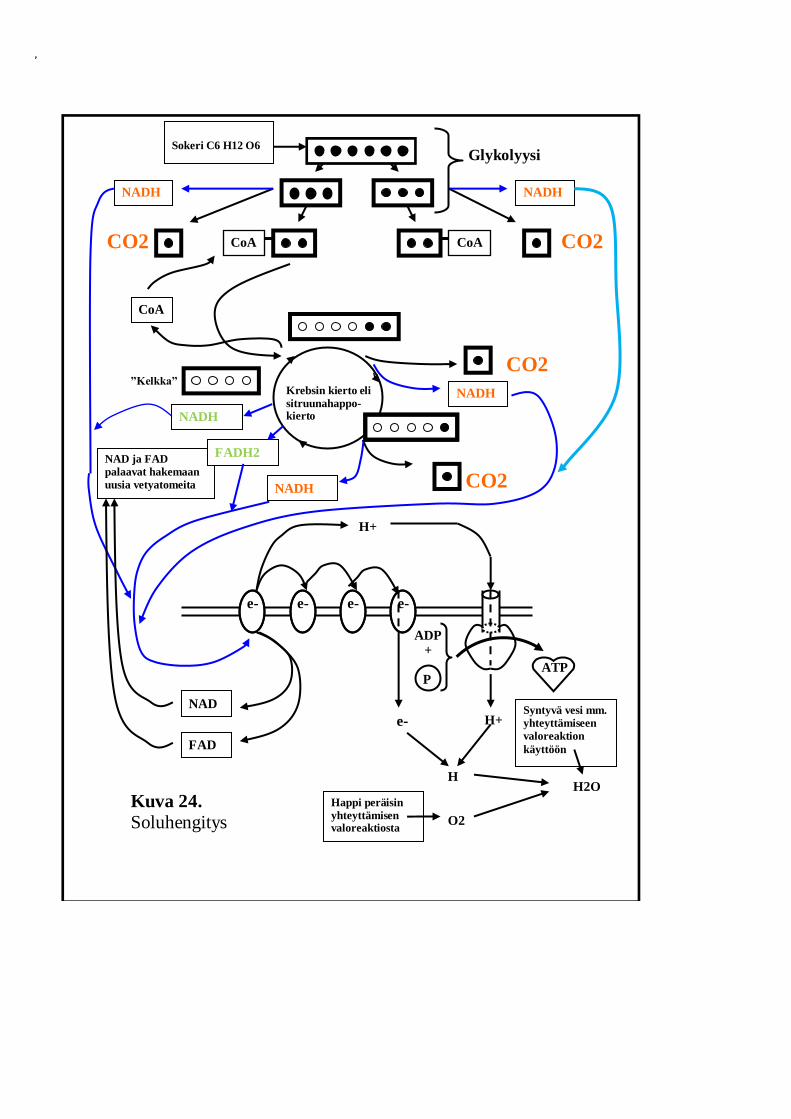
P
Sokeri C6 H12 O6
ADP +
P
ATP
H+ e-
H
H+
e- e- e- e-
Kuva 24. Soluhengitys
”Kelkka”
CoA CoA
CoA
CO2
NADH
NADH
Krebsin kierto eli
sitruunahappo-kierto
NADH NADH
NAD
FAD
O2
Happi peräisin
yhteyttämisen valoreaktiosta
Syntyvä vesi mm.
yhteyttämiseen
valoreaktion
käyttöön
NAD ja FAD
palaavat hakemaan
uusia vetyatomeita
H2O
Glykolyysi
FADH2
NADH
CO2
CO2 CO2

Soluhengityksen tavoitteena on käsitellä sokerimolekyylejä tavalla, jonka
tuloksena voidaan tuottaa mahdollisimman paljon ATP:tä. Koska ATP:n
tuottamiseen tarvitaan vetyatomeita, kohdistuu vaivannäkö reaktioissa
nimenomaan sokerimolekyylin sisältämien vetyatomien irrottamiseen
(siniset nuolet).
Koska vetyatomien irrottaminen tapahtuu eri paikassa (mitokondrion
matriksissa) kuin ATP:n tuottaminen (mitokondrion sisäkalvolla),
tarvitaan täälläkin vedyn kuljettamiseen erikoistuneita
vedynsiirtäjäentsyymeitä. Mitokondriossa niitä on kaksi eri nimistä: FAD
ja NAD (leikillisesti fatsit ja muut natsit, hii!). FAD kuljettaa mukanaan
peräti kahta vetyatomia: vedyillä lastattuna se on siis FADH2. Molemmat
mainitut vedynsiirtäjät ovat rakenteeltaan nukleotideja, aivan kuten oli
yhteyttämisessä esiintyvä vedynsiirtäjä NADP:kin.
Soluhengityksessä etenevää sokerimolekyyliä muokataan reaktiokulun
aikana siten, että molekyylin toiseen päähän syntyy useampaan
otteeseen karboksyyliryhmänä tunnettu molekyylirakenne COOH.
Rakenne tuli aiemminkin tutuksi myös aminohappojen osana.
Karboksyyliryhmää erikseen tuijottamalla on helppoa havaita, että siinä
on selvästi kuin tarjottimella ainekset hiilidioksidimolekyyliin (CO2 eli
COO) ja yhteen vetyatomiin (H). Karboksyyliryhmästä napattuja
vetyatomeja kuljettavat vedynsiirtäjät on merkitty kuvaan 24 punaisella,
samoin karboksyyliryhmästä peräisin olevat hiilidioksidimolekyylit CO2.
Soluhengityksestä vapautuva hiilidioksidi ei synny palamisen tuloksena,
vaan se irtoaa karboksyyliryhmästä sellaisenaan. Emme siis

hengittäessämme puhalla ulos palanutta hiiltä, vaan karboksyyliryhmien
rääseitä, jehh! Tätä asiaapa ei kovin moni tässä maailmassa tiedäkään.
Karboksyyliryhmiä ei omaan kaavioomme ole merkitty. On kuitenkin
hyvä tiedostaa niiden olemassaolo ja merkitys.
Kuvassa 24, suoraan kelkan alapuolella olevat NADH ja FADH2, on
korostettu vihreällä värillä. Nämä viimeistelevät sitruunahappokiertoon
tarvittavan kelkkamolekyylin (oksaloasetaatin) valmiiksi. Näissä kohdissa
ei muodostu enää CO2:ta.
Soluhengitys alkaa glykolyysistä (I)
Soluhengitykseen saapuva sokerimolekyyli on yleensä rypälesokeri eli
glukoosi. Glukoosi on rungoltaan kuuden hiiliatomin (laatikossa olevat
mustat ympyrät) mittainen molekyyli. Glukoosi hajotetaan ensin kahdeksi
kolmen hiiliatomin mittaiseksi molekyyliksi, palorypälehapoksi. Tätä
glukoosin “lysähtämistä” kahdeksi kappaleeksi kutsutaan glykolyysiksi.
Jo glykolyysin aikana glukoosimolekyylistä saadaan talteen kaksi
vetyatomia, jotka NAD kuljettaa mitokondrion sisäkalvolle ATP-
syntetaasin käyttövoimaksi.
Glykolyysi tapahtuu, vaikka solu kärsisi hapen puutteesta. Tällöin tosin
palorypälehappomolekyylit muuttuvat maitohapoksi. Tämäkin on
kolmen hiiliatomin mittainen molekyyli. Glukoosin hyödyntäminen

pysähtyy hapen puuttuessa tähän. Maitohappo jäykistää ja kipeyttää
lihakset.
Glykolyysi tapahtuu solulimassa, ei siis vielä mitokondrioiden sisällä.
Sitruunahappokierto eli Krebbsin kierto (I)
Jos happea on tarjolla, palorypälehappomolekyyleistä irrotetaan yksi
hiilidioksidimolekyyli kummastakin (molekyylin päässä olevasta
karboksyyliryhmästä tietenkin, hää). Jäljelle jäävä kahden hiiliatomin
mittainen molekyyli liitetään jatkoksi molekyyliin nimeltä koentsyymi-A.
Kirjallisuudessa koentsyymi-A lyhennetään yleensä CoA.
Sitruunahappokierto on soluhengityksen syklinen osa. Se tapahtuu
mitokondrioiden matriksissa eli sisimmässä ontelotilassa.
Sitruunahappokierron kelkkamolekyylinä on oksaloasetaatti. Se on
rungoltaan neljän hiiliatomin mittainen (valkoiset ympyrät). Yhdestä
glukoosimolekyylistä riittää lastia kahteen kelkkamolekyyliin (kaksi
hiiliatomia kummastakin palorypälehappomolekyylistä). Kaaviossamme
sitruunahappokierto on esitetty vain yhden kelkkamolekyylin osalta.
Sitruunahappokierrossa kulkevasta molekyylistä irroitetaan vetyatomeja.
Samalla vapautuu hiilidioksidia. NADit ja FAD toimittavat vedyt
soluhengityksen viimeiseen vaiheeseen ATP:n tuotantolinjalle.
Jos vertaat kelkkailutapahtumaa yhteyttämisen pimeäreaktion ja
sitruunahappokierron kesken, havaitset, että sitruunahappokierrossa ei

tarvita fosfaattiryhmiä kiinnittämään sokerista tulevaa pikkumolekyyliä
kelkkamolekyyliin. Istuttamisesta huolehtii koentsyymi-A. Se selviytyy
tehtävästä ilman fosfaattiryhmiäkin.
ATP:n tuotanto eli oksidatiivinen fosforylaatio (I)
Soluhengityksen päätteeksi syntyy ATP:tä meille jo tutuksi tulleella
tavalla. ATP-tuotannosta vapautuvat vetyatomit poltetaan. Tuloksena
syntyy vettä. Samalla vapautuu lämpöä, joka on eliöille hyödyllistä.
On jokseenkin kiinnostavaa, että ainoa aine, jota elimistössämme
poltetaan onkin vety. Se ei siis ole hiili, jota hiilidioksidimolekyyleinä
puhallamme keuhkoistamme uloshengityksemme aikana. Elimistömme
energiahuolto toimii sokereista saatavien vetyatomien voimalla.
Vedynsiirtäjäentsyymit kiertävät jatkuvasti hakemassa
sitruunahappokierrosta uusia vetyatomeja mitokondrion sisäkalvon ATP-
tuotantolinjan käyttöön.
Soluhengityksen jokaista vaihetta katalysoi jokin proteiini. Jo yksin
glykolyysiin sisältyy ainakin kaksitoista eri vaihetta.
Vertaa lopuksi vielä kerran CO2:n, H2O:n, sokerin ja hapen kohtaloita
molekyylitason esityksessä ja yksilötason esityksessä (kuvan 12 kukka
ja hämähäkki). Pidä mielessäsi, että kyseessä ovat samat ilmiöt.
Esityksiä erottaa vain tarkkuus, jolla asioihin on paneuduttu.
Yhteenveto hiili-, vety- ja happiatomien lukumääristä ja kohtaloista

Yhteyttämisessä…
Yhteyttäminen tuottaa sokereita C6 H12 O6. Hiiltä ja happea saadaan
yhtä paljon, kun jokaisesta hiilidioksidimolekyylistä ”lempataan” pois
toinen sen sisältämistä happiatomeista: CO2 → CO + O. Kuusi
hajonnutta CO2-molekyyliä tarjoaa aina tarpeeksi hiiltä ja happea yhtä
sokerimolekyyliä varten. Vapautuvat kuusi happiatomia yhtyvät
valoreaktiosta peräisin oleviin vetyatomeihin muodostaen kuusi
molekyyliä vettä 6 x H2O (Kuva 22c).
Yhteyttämisessä NADP-vedynsiirtäjät kuljettavat kukin vain yhtä
vetyatomia mukanaan. NADPH:ta tarvitaankin 12 kappaletta yhden
sokerimolekyylin, C6 H12 O6, tekemiseen.
… ja soluhengityksessä…
Huomaa, että kuvassa 24 glykolyysi on esitetty kummankin
palorypälehapon (kolmen hiiliatomin mittaiset molekyylit) osalta, mutta
sitruunahappokierto näkyy kuvassa vain kerran. Siksi
sitruunahappokierrosta poistuvien CO2-molekyylien ja vedynsiirtäjien
määrä tulee kertoa kahdella, jotta alkuaineatomien lukumäärät saadaan
täsmäämään.
Glykolyysissä muodostuu kaksi CO2-molekyyliä ja kaksi NADH:ta.
Yhteensä siis 2 hiili-, 2 vety- ja 4 happiatomia.
Sitruunahappokierrossa muodostuu yhden kierroksen aikana 2 kpl
CO2:ta, 3 kpl NADH:ta, yksi FADH2 (jokainen FAD siis kuljettaa
mukanaan aina kahta vetyatomia). Koska sitruunahappokierto tapahtuu

kummallekin sokerinpuolikkaalle erikseen, on nämä määrät kerrottava
kahdella. Lopputulos sitruunahappokierrosta siis on 4 hiili-, 10 vety- 8
happiatomia.
Kun lasketaan yhteen glykolyysistä ja sitruunahappokierrosta poistuvat
alkuainetatomit, saadaan 6 hiili-, 12 vety- ja kuusi happiatomia, siis C6
H12 O6 eli sokerimolekyyli. Ja sokerithan ne ovat soluhengityksen
lähtöaine!
.
Energia-aineenvaihdunnan yhteydessä kannattaa muistella myös
haimaa (V)
Haima säätelee elimistömme sokeritasapainoa. Säätely perustuu
haimasolujen tuottamiin kahteen proteiinirakenteiseen hormoniin:
insuliiniin ja glukagoniin. Sokeritasapainon säätelystä saat hyvän
käsityksen tutkimalla vaikkapa WSOY:n biologian oppikirjasarjan osaa
Ihminen.
Insuliini vaikuttaa kohdesoluihinsa siten, että solujen sisällä olevat
sokerikanavaproteiinit siirtyvät solukelmulle. Tällöin sokeri pääsee
siirtymään solun sisälle, missä se jatkaa matkaansa soluhengityksen
raaka-aineeksi mitokondrioihin.
Sokeritaudissa eli diabeteksessa insuliinia erittyy liian vähän tai se ei
vaikuta. Tällöin sokeri jää ajelehtimaan solujen ulkopuolelle eikä
soluhengitys pääse tapahtumaan täydellä teholla. Elimistön proteiinit
alkavat kärsiä ATP:n puutteesta ja muuttuvat toimintakyvyttömiksi. Ei
ihme, että sokeritautipotilas tuntee olonsa heikoksi.

Nuoruustyypin sokeritauti on autoimmuunisairaus (III)
Sokeritaudista tunnetaan kaksi päämuotoa: nuoruus- ja aikuistyypin
sokeritauti. Nuoruustyypin sokeritauti on autoimmuunisairaus.
Autoimmuunisairauksissa elimistön oma puolustusjärjestelmä alkaa
tuhota elimistön omia kudoksia. Muita autoimmuunisairauksia ovat
nivelreuma, astma ja ms-tauti. Jos aihepiiri tuntuu kiinnostavalta, sinun
kannattaa käydä maistelemassa immunologiaa käsittelevää
tiedostoamme biologian neloskurssista.
Ja tähän vihdoin päättyykin vähemmän pintapuolinen katsauksemme elämän energiatalouden salaisuuksista!