effects of methylmercury on ontogeny of prey capture ability and growth in three populations of...
TRANSCRIPT

Effects of Methylmercury on Ontogeny of Prey Capture Ability and Growth inThree Populations of Larval Fundulus heteroclitus
T. Zhou, R. Scali, J. S. Weis
Department of Biological Sciences, Rutgers University, Newark, New Jersey 07102, USA
Received: 23 February 2000/Accepted: 21 January 2001
Abstract. We used three populations of mummichogs (Fundu-lus heteroclitus), one from a polluted site (Piles Creek [PC],New Jersey) and two from cleaner sites (Tuckerton [TK], NewJersey, and East Hampton [EH], New York), to study (1)whether embryonic, embryonic plus larval, or larval exposureto methylmercury (MeHg) altered larval prey capture abilityand growth; and (2) whether there were differences in toleranceto MeHg-induced behavioral changes among the three popula-tions. Eggs and sperm were obtained from mummichogs cap-tured in the field, and their embryos and larvae were kept inclean sea water or MeHg solution (5, 10mg/L). Larvae werethen tested regularly for prey capture rates and prey captureefficiencies, and their lengths were measured. Embryonic ex-posure to MeHg induced transitory and recoverable impair-ments in larval prey capture ability, whereas larval exposurealone was relatively ineffective. When both embryos and lar-vae were treated, larval prey capture ability was affected at alower concentration and a wider range of larval ages. In termsof growth and prey capture ability, response of larvae toembryonic or larval or exposure to both stages to MeHg variedwith populations. TK fish were the most tolerant with respect tobehavioral changes but were the most sensitive to MeHg inreduction of growth. EH fish were the most sensitive wheneverembryos were treated, and PC fish were the most vulnerableafter larval exposure. The population differences in response toMeHg intoxication may be due to pollution related factors ordifferences in behavioral-related genetic factors.
Many neurotoxicants, such as mercury, have been reported toinduce behavioral changes, including feeding behavior (Eisler1979; Little et al. 1990; Bryanet al. 1995). Feeding behavioris of great importance to larval fish, the most vulnerable stages
to environmental stresses. Alterations in feeding behavior mayreduce growth (Pedersen 1993), and therefore prolong larvaldevelopment. This in turn may increase predation risk, de-crease survivorship, and ultimately affect recruitment. In moststudies, feeding behavior (such as capture rate) is depressedafter exposure to various pollutants (Atchisonet al. 1987,1996; Littleet al. 1993). Every aspect of feeding behavior canbe affected by sublethal exposure to chemical stressors (Littleet al.1993). Stressors may decrease search efficiencies (Atchi-son et al. 1996), strike frequencies (Littleet al. 1990), preycapture rates (Purdy 1989; Weber 1996), capture efficiencies(Weberet al.1991), and reaction distances (Weberet al.1991)and increase handling times (Nyman 1981; Sandheinrich andAtchison 1989).
In this research, we examined larval mummichogs,Fundulusheteroclitus(family: Cyprinodontidae), from a Hg-polluted site(Piles Creek [PC], New Jersey) and two reference sites (Tuck-erton [TK], New Jersey, and East Hampton [EH], New York).Mummichogs from EH, PC, and TK represent northern, inter-mediate, and southern subspecies, respectively, in which thePC subspecies is considered as a hybrid between northern andsouthern forms (Weiset al. 1999). The PC population sharesmany features (such as morphology and mitochondrial DNA)of northern subspecies but has transitional frequency of malatedehydrogenase-A alleles (Weiset al.1999). Embryos from PCwere more tolerant to the teratogenic effects of methylmercury(MeHg) and developed faster than embryos from the cleanerareas (Weiset al. 1981). Though larval tolerance to MeHg(with mortality as the endpoint) was comparable among thesepopulations (Toppinet al.1987), PC larvae were more suscep-tible to MeHg-induced increase in prey vulnerability (Zhou andWeis 1998).
The purpose of this research is to determine to what degreeembryonic, embryonic plus larval, or larval exposure to suble-thal MeHg can alter prey capture ability and growth of larvalmummichogs from the two cleaner sites (EH and TK) and thepolluted PC and to compare the three populations in terms oftolerance to MeHg-induced behavioral changes. Embryonicplus larval exposure represents the route of exposure for mum-michogs. The other two exposure regimes were selected toseparate the effects of MeHg acquired either via maternaltransferring or larval feeding. The three populations were cho-
* Present address:Neurotoxicology Division, MD-74B, NationalHealth and Environmental Effects Research Laboratory, U.S. Envi-ronmental Protection Agency, Research Triangle Park, North Carolina27711, USA
Correspondence to:T. Zhou;email: [email protected]
Arch. Environ. Contam. Toxicol. 41, 47–54 (2001)DOI: 10.1007/s002440010219
A R C H I V E S O F
EnvironmentalContaminationa n d Toxicology© 2001 Springer-Verlag New York Inc.

sen to see if behavioral differences were genetic-based and/orcontaminant-related.
Materials and Methods
Study Sites
The three sampling sites (PC, EH, and TK) are tidalSpartinamarsheswith muddy sediments, with comparable water temperature (Santiago1997). Surrounded by many industrial sites, PC is a tributary of ArthurKill located near heavily industrialized Linden in northern New Jersey.Sediments in PC contain high levels of pollutants, including variousheavy metals and organic and organic toxicants (Eckenfelder 1991). Incontrast, EH and TK are nonindustrialized sites located south andnortheast of PC, respectively. The total Hg content in PC sedimentsaverages 11.5mg/g, more than that in EH (0.045mg/g) or TK (0.263mg/g) sediments (Zhou 1997).
Preparation of Experimental Fish
Adult mummichogs were collected from PC, EH, and TK with Gee’sminnow traps during the breeding season of 1994 (for the first set ofexperiments) and 1995 (for the second set of experiments). Eggs werestripped from multiple ripe females into petri dishes and fertilized withthe milt from multiple males in sea water. Any uncleaved or deadembryos were discarded. The successfully fertilized eggs were thenwashed and transferred to experimental finger bowls. No more than 50embryos in each finger bowl of 50 ml solution (15 ppt salinity) wereincubated at 226 0.2°C in an incubator with a photoperiod of12L:12D. Larvae were kept in 8.5-cm diameter finger bowls at adensity of one larva per 5 ml salt water or MeHg solution, no morethan 10 larvae per bowl. Both salt water and MeHg solutions werechanged on daily basis. The desired MeHg concentrations were ob-tained by diluting stock solution of 100 mg/L methylmercuric chloride(ICN Pharmaceutical, Plainview, NY). Three days posthatch (dph) orolder larvae were fed with brine shrimp (Artemia salina) nauplii at adensity of 25Artemiaper larva once daily shortly after being testedand solution change.
Exposure Regimes
Two separate sets of experiments were conducted. In the first set ofexperiments we investigated the short-term effects of embryonic orembryonic plus larval exposure to MeHg on prey capture ability andgrowth of larval mummichogs from the three sites. The followingexposure regimes were set up: I. Both embryos and larvae were keptin synthetic salt water of 15 ppt (“Instant Ocean” Sea Salts) (controlgroup); II. Embryos were exposed to MeHg at concentrations of 5 and10 mg/L, below those which caused morphological changes in em-bryos (Zhou 1997), throughout the whole developmental period. Lar-vae were then placed in clean salt water (recovery experiment); III.Embryos were untreated but larvae were treated with 5 and 10mg/LMeHg (larval vulnerability experiment). Four replicate dishes wereused for each treatment,i.e., 12 bowls with embryos (4 as controls, 4treated with 5 or 10mg/L MeHg), 20 bowls with larvae (4 as controls,4 treated with 5 or 10mg/L MeHg from untreated embryos, 4 treatedwith 5 or 10mg/L MeHg from treated embryos).
In the second set of experiments we determined the short-termeffects of larval exposure to MeHg on prey capture ability and growthof the three populations. Exposure regimes were set up as follows: I.Both embryos and larvae were kept in synthetic salt water of 15 ppt
(control group); II. Embryos were treated with 5 and 10mg/L MeHg,and larvae were maintained in MeHg of the same concentration asembryos. Four replicate dishes were used for each treatment,i.e., 4bowls with untreated embryos, 12 bowls with larvae (4 as controls, 4treated with 5 or 10mg/L MeHg).
Growth and Behavioral Experiments
For larval growth, the total lengths of 20 larvae from each experimen-tal group at 3, 7, 10, 14, 21, and 31 dph were measured with a metricruler under a dissecting microscope. Any dead or unhealthy larvaewere discarded before the trial began,i.e., 3 dph.
At least 20 larvae were tested for each treatment between 10:00 and16:00 h. The same group of larvae were tested at different ages. Larvalprey capture ability was examined at 3, 5, 8, 11, and 14 dph. Eachindividual larva was tested only once per testing day before being fed.Five brine shrimp nauplii were introduced and randomly distributed ina 4-cm diameter dish with 3 mm of salt water. Ten seconds afterArtemia introduction, one larva was presented in the center of thefinger bowl with a transfer pipet. We recorded the time taken for alarva to eat each brine shrimp over a 10-min (for the first set ofexperiments) or 5-min (for the second set of experiments) period, aswell as the number of miscues,i.e., the number of times it attemptedbut failed to catch the prey. Prey capture efficiency was calculated bythe number of prey caught over total feeding strikes (summation of thetotal number of prey caught and miscues).
Statistical Analysis
Statistical analysis was conducted by SAS 6.09. Both prey capturerates and prey capture efficiencies were ranked before ANOVA. Totake dish effects into consideration, the nested ANOVA was used,followed by the Dunnett’s multiple comparison test to see whichtreated groups were different from the control at 0.05 level for preycapture rates, prey capture efficiency, and growth.
Results
Prey Capture Ability
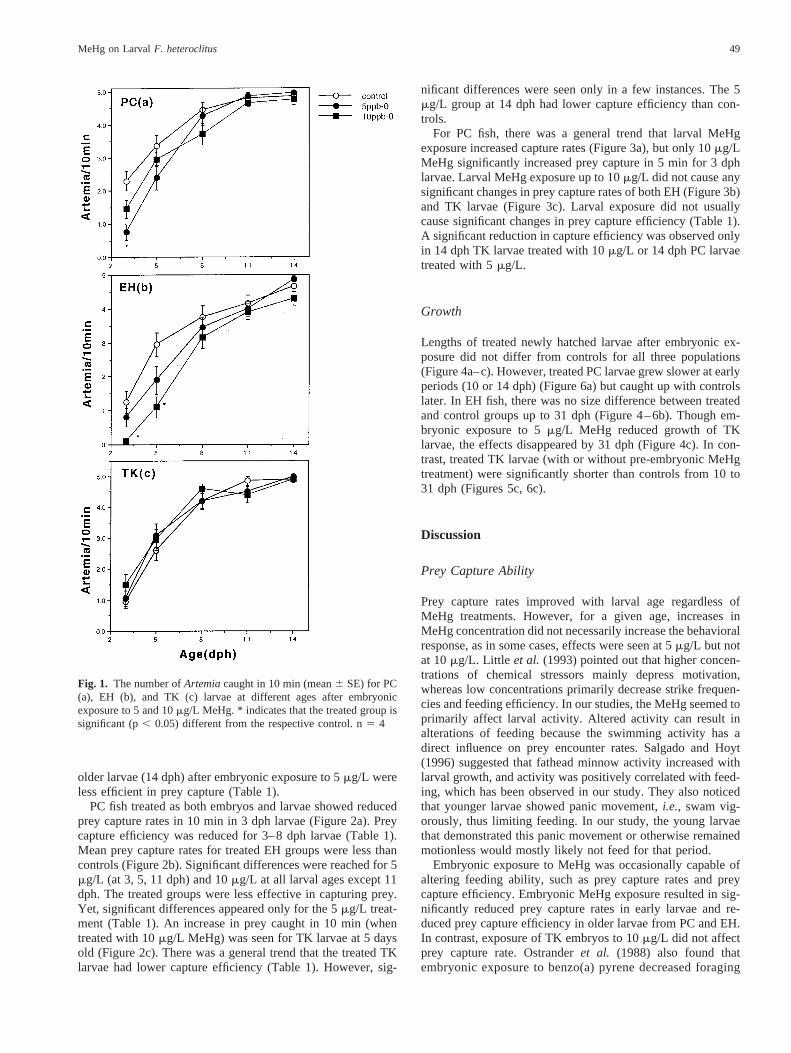
First feeding larvae encountered prey mainly as the conse-quence of random movements by shrimp rather than activesearching or pursuit, which was often seen in older or moreexperienced larvae. More than 20% of young larvae exhibited“panic” movements,i.e., swimming rapidly back and forthalong the side of the finger bowl, making turns frequently butignoring any prey item nearby. In contrast, this behavior wasseldom observed in older larvae, which showed greater moti-vation to feed. In the three populations, larval feeding strikesand prey capture rates increased with the maturity of larvae.
Embryonic exposure to 5mg/L MeHg significantly reducedthe prey capture rate in PC fish (Figure 1a) and capture effi-ciency (Table 1) for 3-day-old larvae. Older PC larvae alsoshowed decreased prey capture efficiency when exposed to 10mg/L MeHg (Table 1). Prey capture rates (Figure 1b) for 3 and5 dph EH larvae were significantly reduced following embry-onic exposure to 10mg/L. No effects on prey capture rateswere found in TK larvae (Figure 1c). For both EH and TK,
48 T. Zhouet al.
older larvae (14 dph) after embryonic exposure to 5mg/L wereless efficient in prey capture (Table 1).
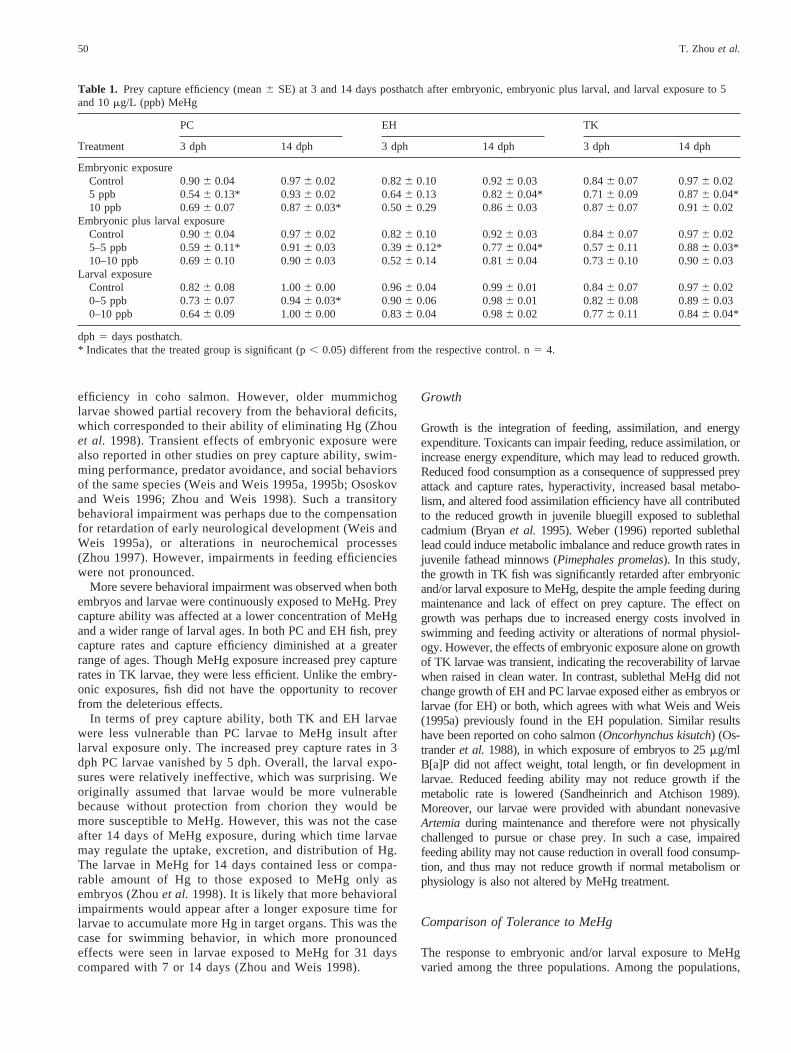
PC fish treated as both embryos and larvae showed reducedprey capture rates in 10 min in 3 dph larvae (Figure 2a). Preycapture efficiency was reduced for 3–8 dph larvae (Table 1).Mean prey capture rates for treated EH groups were less thancontrols (Figure 2b). Significant differences were reached for 5mg/L (at 3, 5, 11 dph) and 10mg/L at all larval ages except 11dph. The treated groups were less effective in capturing prey.Yet, significant differences appeared only for the 5mg/L treat-ment (Table 1). An increase in prey caught in 10 min (whentreated with 10mg/L MeHg) was seen for TK larvae at 5 daysold (Figure 2c). There was a general trend that the treated TKlarvae had lower capture efficiency (Table 1). However, sig-
nificant differences were seen only in a few instances. The 5mg/L group at 14 dph had lower capture efficiency than con-trols.
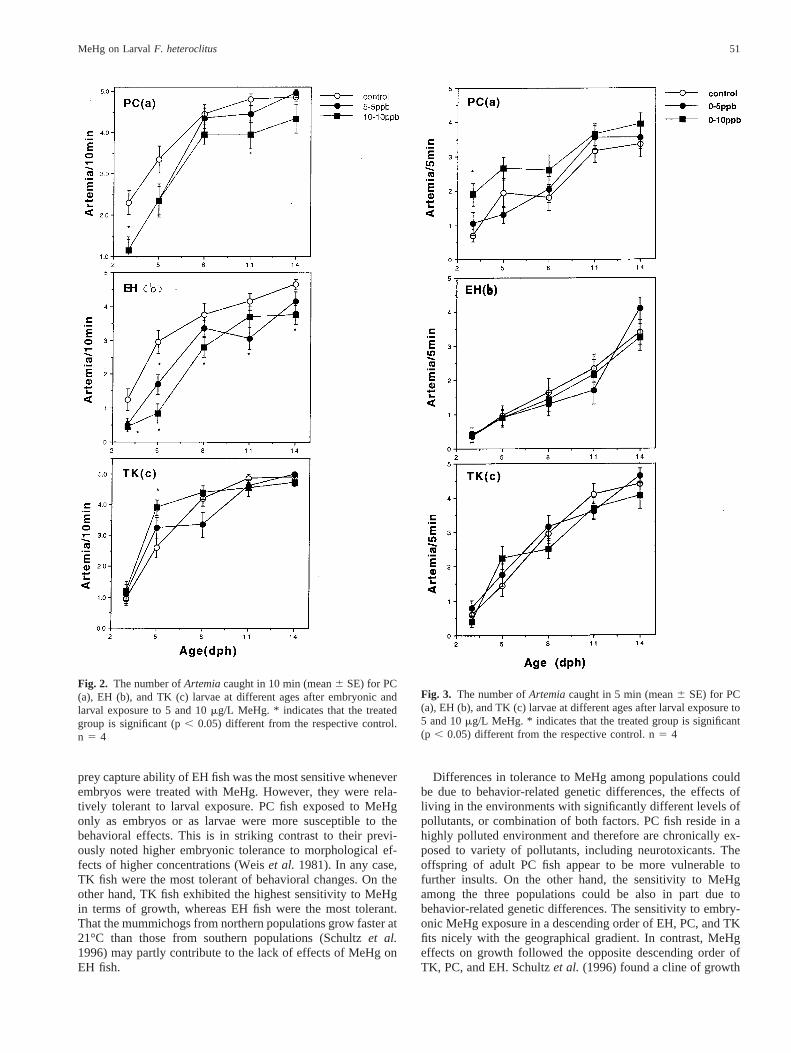
For PC fish, there was a general trend that larval MeHgexposure increased capture rates (Figure 3a), but only 10mg/LMeHg significantly increased prey capture in 5 min for 3 dphlarvae. Larval MeHg exposure up to 10mg/L did not cause anysignificant changes in prey capture rates of both EH (Figure 3b)and TK larvae (Figure 3c). Larval exposure did not usuallycause significant changes in prey capture efficiency (Table 1).A significant reduction in capture efficiency was observed onlyin 14 dph TK larvae treated with 10mg/L or 14 dph PC larvaetreated with 5mg/L.
Growth
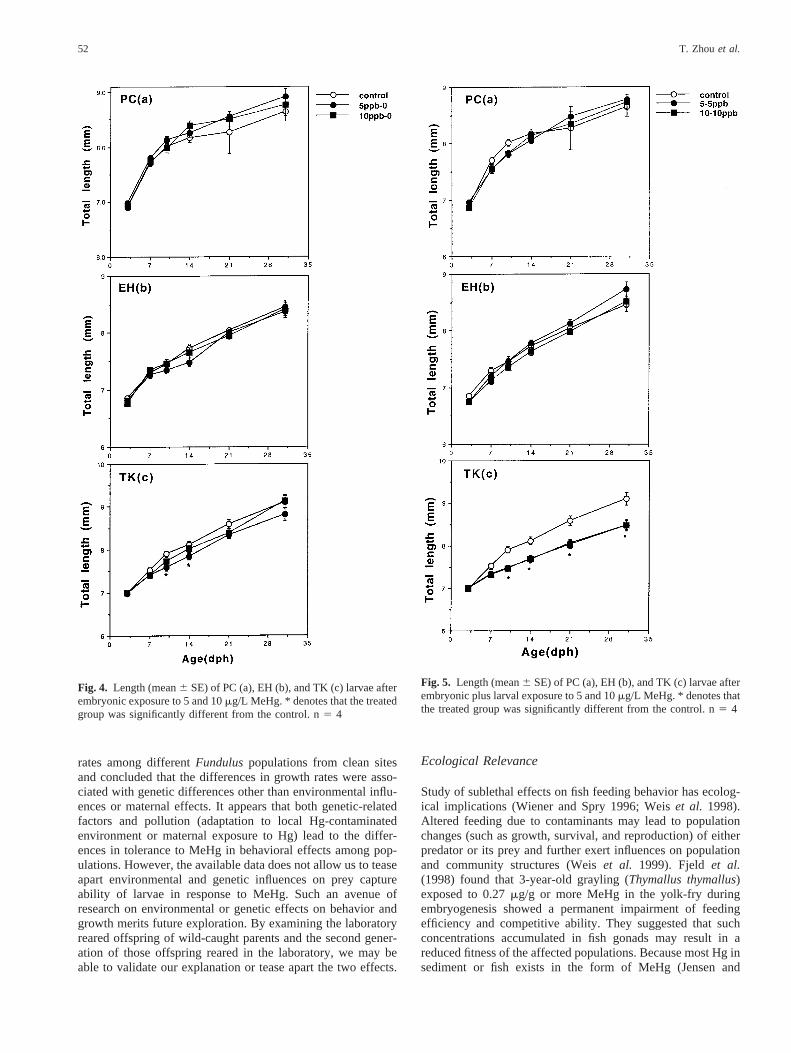
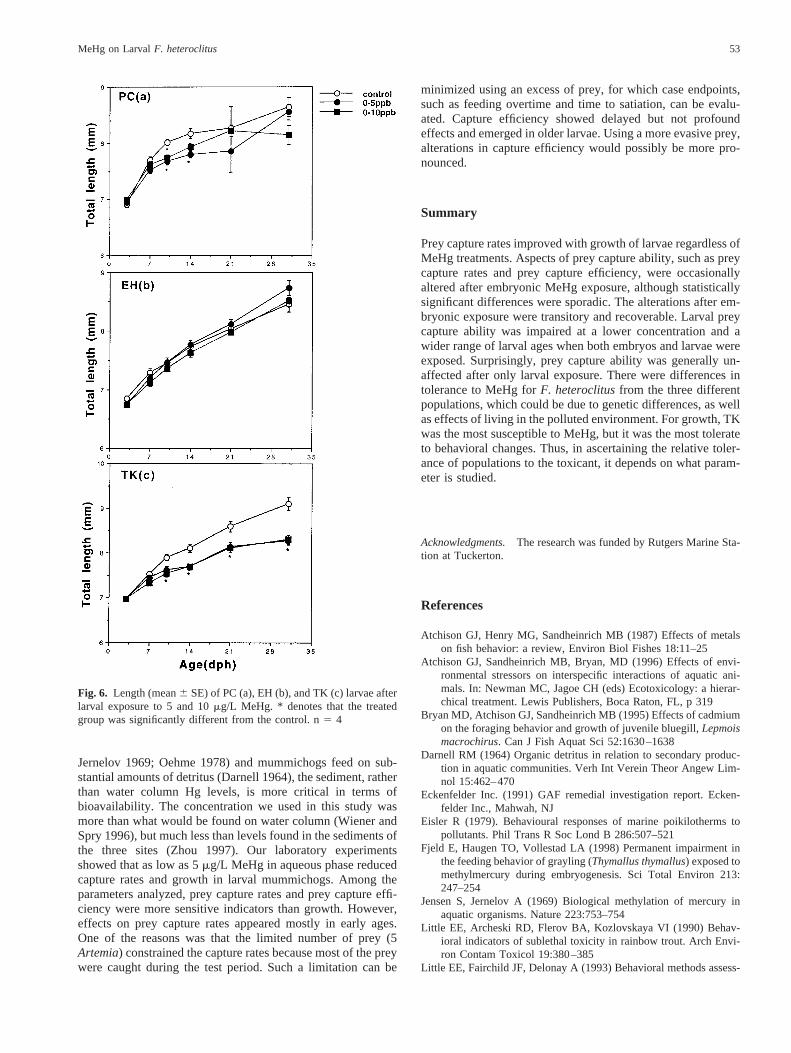
Lengths of treated newly hatched larvae after embryonic ex-posure did not differ from controls for all three populations(Figure 4a–c). However, treated PC larvae grew slower at earlyperiods (10 or 14 dph) (Figure 6a) but caught up with controlslater. In EH fish, there was no size difference between treatedand control groups up to 31 dph (Figure 4–6b). Though em-bryonic exposure to 5mg/L MeHg reduced growth of TKlarvae, the effects disappeared by 31 dph (Figure 4c). In con-trast, treated TK larvae (with or without pre-embryonic MeHgtreatment) were significantly shorter than controls from 10 to31 dph (Figures 5c, 6c).
Discussion
Prey Capture Ability
Prey capture rates improved with larval age regardless ofMeHg treatments. However, for a given age, increases inMeHg concentration did not necessarily increase the behavioralresponse, as in some cases, effects were seen at 5mg/L but notat 10mg/L. Little et al. (1993) pointed out that higher concen-trations of chemical stressors mainly depress motivation,whereas low concentrations primarily decrease strike frequen-cies and feeding efficiency. In our studies, the MeHg seemed toprimarily affect larval activity. Altered activity can result inalterations of feeding because the swimming activity has adirect influence on prey encounter rates. Salgado and Hoyt(1996) suggested that fathead minnow activity increased withlarval growth, and activity was positively correlated with feed-ing, which has been observed in our study. They also noticedthat younger larvae showed panic movement,i.e., swam vig-orously, thus limiting feeding. In our study, the young larvaethat demonstrated this panic movement or otherwise remainedmotionless would mostly likely not feed for that period.
Embryonic exposure to MeHg was occasionally capable ofaltering feeding ability, such as prey capture rates and preycapture efficiency. Embryonic MeHg exposure resulted in sig-nificantly reduced prey capture rates in early larvae and re-duced prey capture efficiency in older larvae from PC and EH.In contrast, exposure of TK embryos to 10mg/L did not affectprey capture rate. Ostranderet al. (1988) also found thatembryonic exposure to benzo(a) pyrene decreased foraging
Fig. 1. The number ofArtemiacaught in 10 min (mean6 SE) for PC(a), EH (b), and TK (c) larvae at different ages after embryonicexposure to 5 and 10mg/L MeHg. * indicates that the treated group issignificant (p, 0.05) different from the respective control. n5 4
MeHg on LarvalF. heteroclitus 49
efficiency in coho salmon. However, older mummichoglarvae showed partial recovery from the behavioral deficits,which corresponded to their ability of eliminating Hg (Zhouet al. 1998). Transient effects of embryonic exposure werealso reported in other studies on prey capture ability, swim-ming performance, predator avoidance, and social behaviorsof the same species (Weis and Weis 1995a, 1995b; Ososkovand Weis 1996; Zhou and Weis 1998). Such a transitorybehavioral impairment was perhaps due to the compensationfor retardation of early neurological development (Weis andWeis 1995a), or alterations in neurochemical processes(Zhou 1997). However, impairments in feeding efficiencieswere not pronounced.
More severe behavioral impairment was observed when bothembryos and larvae were continuously exposed to MeHg. Preycapture ability was affected at a lower concentration of MeHgand a wider range of larval ages. In both PC and EH fish, preycapture rates and capture efficiency diminished at a greaterrange of ages. Though MeHg exposure increased prey capturerates in TK larvae, they were less efficient. Unlike the embry-onic exposures, fish did not have the opportunity to recoverfrom the deleterious effects.
In terms of prey capture ability, both TK and EH larvaewere less vulnerable than PC larvae to MeHg insult afterlarval exposure only. The increased prey capture rates in 3dph PC larvae vanished by 5 dph. Overall, the larval expo-sures were relatively ineffective, which was surprising. Weoriginally assumed that larvae would be more vulnerablebecause without protection from chorion they would bemore susceptible to MeHg. However, this was not the caseafter 14 days of MeHg exposure, during which time larvaemay regulate the uptake, excretion, and distribution of Hg.The larvae in MeHg for 14 days contained less or compa-rable amount of Hg to those exposed to MeHg only asembryos (Zhouet al. 1998). It is likely that more behavioralimpairments would appear after a longer exposure time forlarvae to accumulate more Hg in target organs. This was thecase for swimming behavior, in which more pronouncedeffects were seen in larvae exposed to MeHg for 31 dayscompared with 7 or 14 days (Zhou and Weis 1998).
Growth
Growth is the integration of feeding, assimilation, and energyexpenditure. Toxicants can impair feeding, reduce assimilation, orincrease energy expenditure, which may lead to reduced growth.Reduced food consumption as a consequence of suppressed preyattack and capture rates, hyperactivity, increased basal metabo-lism, and altered food assimilation efficiency have all contributedto the reduced growth in juvenile bluegill exposed to sublethalcadmium (Bryanet al. 1995). Weber (1996) reported sublethallead could induce metabolic imbalance and reduce growth rates injuvenile fathead minnows (Pimephales promelas). In this study,the growth in TK fish was significantly retarded after embryonicand/or larval exposure to MeHg, despite the ample feeding duringmaintenance and lack of effect on prey capture. The effect ongrowth was perhaps due to increased energy costs involved inswimming and feeding activity or alterations of normal physiol-ogy. However, the effects of embryonic exposure alone on growthof TK larvae was transient, indicating the recoverability of larvaewhen raised in clean water. In contrast, sublethal MeHg did notchange growth of EH and PC larvae exposed either as embryos orlarvae (for EH) or both, which agrees with what Weis and Weis(1995a) previously found in the EH population. Similar resultshave been reported on coho salmon (Oncorhynchus kisutch) (Os-tranderet al. 1988), in which exposure of embryos to 25mg/mlB[a]P did not affect weight, total length, or fin development inlarvae. Reduced feeding ability may not reduce growth if themetabolic rate is lowered (Sandheinrich and Atchison 1989).Moreover, our larvae were provided with abundant nonevasiveArtemia during maintenance and therefore were not physicallychallenged to pursue or chase prey. In such a case, impairedfeeding ability may not cause reduction in overall food consump-tion, and thus may not reduce growth if normal metabolism orphysiology is also not altered by MeHg treatment.
Comparison of Tolerance to MeHg
The response to embryonic and/or larval exposure to MeHgvaried among the three populations. Among the populations,
Table 1. Prey capture efficiency (mean6 SE) at 3 and 14 days posthatch after embryonic, embryonic plus larval, and larval exposure to 5and 10mg/L (ppb) MeHg
Treatment
PC EH TK
3 dph 14 dph 3 dph 14 dph 3 dph 14 dph
Embryonic exposureControl 0.906 0.04 0.976 0.02 0.826 0.10 0.926 0.03 0.846 0.07 0.976 0.025 ppb 0.546 0.13* 0.936 0.02 0.646 0.13 0.826 0.04* 0.716 0.09 0.876 0.04*10 ppb 0.696 0.07 0.876 0.03* 0.506 0.29 0.866 0.03 0.876 0.07 0.916 0.02
Embryonic plus larval exposureControl 0.906 0.04 0.976 0.02 0.826 0.10 0.926 0.03 0.846 0.07 0.976 0.025–5 ppb 0.596 0.11* 0.916 0.03 0.396 0.12* 0.776 0.04* 0.576 0.11 0.886 0.03*10–10 ppb 0.696 0.10 0.906 0.03 0.526 0.14 0.816 0.04 0.736 0.10 0.906 0.03
Larval exposureControl 0.826 0.08 1.006 0.00 0.966 0.04 0.996 0.01 0.846 0.07 0.976 0.020–5 ppb 0.736 0.07 0.946 0.03* 0.906 0.06 0.986 0.01 0.826 0.08 0.896 0.030–10 ppb 0.646 0.09 1.006 0.00 0.836 0.04 0.986 0.02 0.776 0.11 0.846 0.04*
dph 5 days posthatch.* Indicates that the treated group is significant (p, 0.05) different from the respective control. n5 4.
50 T. Zhouet al.
prey capture ability of EH fish was the most sensitive wheneverembryos were treated with MeHg. However, they were rela-tively tolerant to larval exposure. PC fish exposed to MeHgonly as embryos or as larvae were more susceptible to thebehavioral effects. This is in striking contrast to their previ-ously noted higher embryonic tolerance to morphological ef-fects of higher concentrations (Weiset al. 1981). In any case,TK fish were the most tolerant of behavioral changes. On theother hand, TK fish exhibited the highest sensitivity to MeHgin terms of growth, whereas EH fish were the most tolerant.That the mummichogs from northern populations grow faster at21°C than those from southern populations (Schultzet al.1996) may partly contribute to the lack of effects of MeHg onEH fish.
Differences in tolerance to MeHg among populations couldbe due to behavior-related genetic differences, the effects ofliving in the environments with significantly different levels ofpollutants, or combination of both factors. PC fish reside in ahighly polluted environment and therefore are chronically ex-posed to variety of pollutants, including neurotoxicants. Theoffspring of adult PC fish appear to be more vulnerable tofurther insults. On the other hand, the sensitivity to MeHgamong the three populations could be also in part due tobehavior-related genetic differences. The sensitivity to embry-onic MeHg exposure in a descending order of EH, PC, and TKfits nicely with the geographical gradient. In contrast, MeHgeffects on growth followed the opposite descending order ofTK, PC, and EH. Schultzet al. (1996) found a cline of growth
Fig. 2. The number ofArtemiacaught in 10 min (mean6 SE) for PC(a), EH (b), and TK (c) larvae at different ages after embryonic andlarval exposure to 5 and 10mg/L MeHg. * indicates that the treatedgroup is significant (p, 0.05) different from the respective control.n 5 4
Fig. 3. The number ofArtemiacaught in 5 min (mean6 SE) for PC(a), EH (b), and TK (c) larvae at different ages after larval exposure to5 and 10mg/L MeHg. * indicates that the treated group is significant(p , 0.05) different from the respective control. n5 4
MeHg on LarvalF. heteroclitus 51
rates among differentFunduluspopulations from clean sitesand concluded that the differences in growth rates were asso-ciated with genetic differences other than environmental influ-ences or maternal effects. It appears that both genetic-relatedfactors and pollution (adaptation to local Hg-contaminatedenvironment or maternal exposure to Hg) lead to the differ-ences in tolerance to MeHg in behavioral effects among pop-ulations. However, the available data does not allow us to teaseapart environmental and genetic influences on prey captureability of larvae in response to MeHg. Such an avenue ofresearch on environmental or genetic effects on behavior andgrowth merits future exploration. By examining the laboratoryreared offspring of wild-caught parents and the second gener-ation of those offspring reared in the laboratory, we may beable to validate our explanation or tease apart the two effects.
Ecological Relevance
Study of sublethal effects on fish feeding behavior has ecolog-ical implications (Wiener and Spry 1996; Weiset al. 1998).Altered feeding due to contaminants may lead to populationchanges (such as growth, survival, and reproduction) of eitherpredator or its prey and further exert influences on populationand community structures (Weiset al. 1999). Fjeld et al.(1998) found that 3-year-old grayling (Thymallus thymallus)exposed to 0.27mg/g or more MeHg in the yolk-fry duringembryogenesis showed a permanent impairment of feedingefficiency and competitive ability. They suggested that suchconcentrations accumulated in fish gonads may result in areduced fitness of the affected populations. Because most Hg insediment or fish exists in the form of MeHg (Jensen and
Fig. 4. Length (mean6 SE) of PC (a), EH (b), and TK (c) larvae afterembryonic exposure to 5 and 10mg/L MeHg. * denotes that the treatedgroup was significantly different from the control. n5 4
Fig. 5. Length (mean6 SE) of PC (a), EH (b), and TK (c) larvae afterembryonic plus larval exposure to 5 and 10mg/L MeHg. * denotes thatthe treated group was significantly different from the control. n5 4
52 T. Zhouet al.
Jernelov 1969; Oehme 1978) and mummichogs feed on sub-stantial amounts of detritus (Darnell 1964), the sediment, ratherthan water column Hg levels, is more critical in terms ofbioavailability. The concentration we used in this study wasmore than what would be found on water column (Wiener andSpry 1996), but much less than levels found in the sediments ofthe three sites (Zhou 1997). Our laboratory experimentsshowed that as low as 5mg/L MeHg in aqueous phase reducedcapture rates and growth in larval mummichogs. Among theparameters analyzed, prey capture rates and prey capture effi-ciency were more sensitive indicators than growth. However,effects on prey capture rates appeared mostly in early ages.One of the reasons was that the limited number of prey (5Artemia) constrained the capture rates because most of the preywere caught during the test period. Such a limitation can be
minimized using an excess of prey, for which case endpoints,such as feeding overtime and time to satiation, can be evalu-ated. Capture efficiency showed delayed but not profoundeffects and emerged in older larvae. Using a more evasive prey,alterations in capture efficiency would possibly be more pro-nounced.
Summary
Prey capture rates improved with growth of larvae regardless ofMeHg treatments. Aspects of prey capture ability, such as preycapture rates and prey capture efficiency, were occasionallyaltered after embryonic MeHg exposure, although statisticallysignificant differences were sporadic. The alterations after em-bryonic exposure were transitory and recoverable. Larval preycapture ability was impaired at a lower concentration and awider range of larval ages when both embryos and larvae wereexposed. Surprisingly, prey capture ability was generally un-affected after only larval exposure. There were differences intolerance to MeHg forF. heteroclitusfrom the three differentpopulations, which could be due to genetic differences, as wellas effects of living in the polluted environment. For growth, TKwas the most susceptible to MeHg, but it was the most tolerateto behavioral changes. Thus, in ascertaining the relative toler-ance of populations to the toxicant, it depends on what param-eter is studied.
Acknowledgments.The research was funded by Rutgers Marine Sta-tion at Tuckerton.
References
Atchison GJ, Henry MG, Sandheinrich MB (1987) Effects of metalson fish behavior: a review, Environ Biol Fishes 18:11–25
Atchison GJ, Sandheinrich MB, Bryan, MD (1996) Effects of envi-ronmental stressors on interspecific interactions of aquatic ani-mals. In: Newman MC, Jagoe CH (eds) Ecotoxicology: a hierar-chical treatment. Lewis Publishers, Boca Raton, FL, p 319
Bryan MD, Atchison GJ, Sandheinrich MB (1995) Effects of cadmiumon the foraging behavior and growth of juvenile bluegill,Lepmoismacrochirus. Can J Fish Aquat Sci 52:1630–1638
Darnell RM (1964) Organic detritus in relation to secondary produc-tion in aquatic communities. Verh Int Verein Theor Angew Lim-nol 15:462–470
Eckenfelder Inc. (1991) GAF remedial investigation report. Ecken-felder Inc., Mahwah, NJ
Eisler R (1979). Behavioural responses of marine poikilotherms topollutants. Phil Trans R Soc Lond B 286:507–521
Fjeld E, Haugen TO, Vollestad LA (1998) Permanent impairment inthe feeding behavior of grayling (Thymallus thymallus) exposed tomethylmercury during embryogenesis. Sci Total Environ 213:247–254
Jensen S, Jernelov A (1969) Biological methylation of mercury inaquatic organisms. Nature 223:753–754
Little EE, Archeski RD, Flerov BA, Kozlovskaya VI (1990) Behav-ioral indicators of sublethal toxicity in rainbow trout. Arch Envi-ron Contam Toxicol 19:380–385
Little EE, Fairchild JF, Delonay A (1993) Behavioral methods assess-
Fig. 6. Length (mean6 SE) of PC (a), EH (b), and TK (c) larvae afterlarval exposure to 5 and 10mg/L MeHg. * denotes that the treatedgroup was significantly different from the control. n5 4
MeHg on LarvalF. heteroclitus 53

ing impacts of contaminants on early life stage fishes. Am FishSoc Sym 14:67–76
Nyman HG (1981) Sublethal effects of lead (Pb) on size selectivepredation by fish-applications on the ecosystem level. Verh Inter-nat Verein Limnol 21:1126–1130
Oehme FW (1978) Toxicity of heavy metals in the environment, part1. Marcel Dekker, New York, p 515
Ososkov I, Weis JS (1996) Development of social behavior in larvalmummichogs after embryonic exposure to methylmercury. TransAm Fish Soc 125:983–987
Ostrander GK, Marsha LL, Kocan RM (1988) The ontogeny of cohosalmon (Oncorhynchus kisutch) behavior following embryonicexposure to benzo[a]pyrene. Aquat Tox 13:325–346
Pedersen BH (1993) Growth and mortality in young larval herring(Clupea harengus): effects of repetitive changes in food availabil-ity. Mar Biol 117:547–550
Purdy JE (1989) The effects of brief exposure to aromatic hydrocar-bons on feeding and avoidance behaviour in coho salmon,On-corhynchus kisutch. J Fish Biol 34:621–629
Salgado SD, Hoyt RD (1996) Early behavior formation in fatheadminnow larvae,Pimephales promelas, implications for sensoryfunction. Mar Fresh Behav Physiol 28:91–106
Sandheinrich MB, Atchison GJ (1989) Sublethal toxicant effects onfish foraging behavior: empirical vs. mechanistic approaches. En-viron Toxicol Chem 9:107–119
Santiago C (1997) Size-frequency distribution ofPalaemonetes pugioin two New Jersey estuaries and predator-prey interactions withFundulus heteroclitusMS thesis, New Jersey Institute of Tech-nology, Newark, NJ
Schultz ET, Reynolds KE, Conover DO (1996) Countergradient vari-ation in growth among newly hatchedFundulus heteroclitus:geographic differences revealed by common-environment exper-iments. Functional Ecol 10:366–374
Toppin SRV, Heber M, Weis JS, Weis P (1987) Changes in repro-ductive biology and life history ofFundulus heteroclitusin apolluted environment. In: Vernberg WB, Calabrese A, ThurbergFP, Vernberg FJ (eds) Pollution physiology of estuarine organ-isms. University of South Carolina Press, Columbia, SC, p.171
Weber DN (1996) Lead-induced metabolic imbalance and feeding
alterations in juvenile fathead minnows (Pimephales promelas).Environ Toxicol Water Quality 11:45–51
Weber DN, Russo A, Seale DB, Spieler RE (1991) Waterborne leadaffects feeding abilities and neurotransmitter levels of juvenilefathead minnow (Pimephales promelas). Aquat Toxicol 21:71–80
Weis JS, Weis P (1995a) Effects of embryonic exposure to methyl-mercury on larval prey-capture ability in the mummichog,Fun-dulus heteroclitus. Environ Toxicol Chem 14:153–156
Weis JS, Weis P (1995b) Swimming performance and predator avoid-ance by mummichog (Fundulus heteroclitus) larvae after embry-onic or larval exposure to methylmercury. Can J Fish Aquat Sci52:2168–2173
Weis JS, Weis P, Heber M, Vaidya S (1981) Methylmercury toleranceof killifish (Fundulus heteroclitus) embryos from a polluted vs.non-polluted environment. Mar Biol 65:283–287
Weis JS, Smith GM, Zhou T (1999) Altered predator/prey behavior inpolluted environments: implications for fish conservation. EnvironBiol Fishes 55:43–51
Weis JS, Mugue N, Weis P (1999) Mercury tolerance, populationeffects, and population genetics in the mummichog,Fundulusheteroclitus. In: Forbes VE (ed) Genetics and ecotoxicology.Taylor & Francis, New York, pp 31–54
Wiener JG, Spry DJ (1996) Toxicological significance of mercury infreshwater fish. In: Beyer WN, Heinz GH, Redmon-Norwood AW(eds) Environmental contaminants in wildlife: interpreting tissueconcentrations. Special Publication of the Society of Environmen-tal Toxicology and Chemistry, Lewis Publishers, Boca Raton, FL,pp 297–339
Zhou T (1997) Behavioral development and neurobehavioral effects ofmethylmercury on larval mummichogs,Fundulus heteroclitus,from polluted and reference environments. PhD thesis. RutgersUniversity, Newark, NJ
Zhou T, Weis JS (1998) Swimming behavior and predator avoidancein three populations ofFundulus heteroclituslarvae after embry-onic and/or larval exposure to methylmercury. Aquat Toxicol43:131–148
Zhou T, Weis P, Weis JS (1998) Mercury burden in two populationsof Fundulus heteroclitusafter sublethal methylmercury exposure.Aquat Toxicol 42:37–47
54 T. Zhouet al.