duel of the fates: the role of transcriptional circuits ... · duel of the fates: the role of...
TRANSCRIPT

Duel of the fates: the role of transcriptional circuits andnoise in CD4+ cellsDaniel Hebenstreit1, Andrew Deonarine1,M Madan Babu and Sarah A Teichmann
Available online at www.sciencedirect.com
CD4+ T cells play key roles in orchestrating adaptive immune
responses, and are a popular model for mammalian cell
differentiation. While immune regulation would seem to require
exactly adjusted mRNA and protein expression levels of key
factors, there is little evidence that this is strictly the case.
Stochastic gene expression and plasticity of cell types contrast
the apparent need for precision. Recent work has provided
insight into the magnitude of molecular noise, as well as the
relationship between noise, transcriptional circuits and
epigenetic modifications in a variety of cell types. These
processes and their interplay will also govern gene expression
patterns in the different CD4+ cell types, and the determination
of their cellular fates.
Address
MRC Laboratory of Molecular Biology, Hills Road, Cambridge CB2 0QH,
UK
Corresponding author: Teichmann, Sarah A ([email protected])1 These authors contributed equally to this work.
Current Opinion in Cell Biology 2012, 24:350–358
This review comes from a themed issue on
Nucleus and gene expression
Edited by Asifa Akhtar and Karla Neugebauer
Available online 11th April 2012
0955-0674/$ – see front matter
# 2012 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.ceb.2012.03.007
IntroductionCells change phenotype when they specialize or adapt to
different environments. There are several well-known
systems that illustrate this phenomenon. Some examples
include the recently reported reprogramming of differ-
entiated cells into induced pluripotent stem cells (iPS) [1]
or the differentiation of CD4+ T cells [2]. These changes
or ‘switches’ in cellular phenotype are influenced by
changes in expression levels of key genes [3], and are
subject to molecular fluctuations [4��]. Such noise can be
classified as either intrinsic or extrinsic [5–8]. Intrinsic
noise is caused by gene-specific stochastic fluctuations in
abundances of molecules that are due to transcription,
translation, and related biochemical mechanisms, while
extrinsic noise originates from variations in the state or
concentration of other components in the cell [5]. As a
Current Opinion in Cell Biology 2012, 24:350–358
result, abundances of mRNAs and proteins of identical
genes can vary widely from cell to cell, constituting
‘noise,’ even within a homogenous cell population.
Noise propagation is strongly influenced by topologies of
regulatory networks [9,10], such as the transcriptional
circuits controlling cell fate. Examples include small
subnetworks, such as feedforward loops (FFLs) [11]
and feedback loops which can buffer or amplify noise,
depending on the parameters that influence the kinetics
of gene expression [12]. Recent theoretical studies have
tried to explore the structural requirements of network
controllability [13,14]. This question is important in the
context of ‘master regulators,’ which are essentially small
numbers or even single transcription factors (TFs) that
are able to ‘reprogram’ a cell type [15]. Besides the basic
TF–target gene interactions, other factors such as miRNA
[16], and epigenetic modifications [17,18] are part of
broader regulatory circuits and contribute to stabilization
or switching of phenotypes.
In this review we will focus on CD4+ T cells, which is a
good biological system for illustrating the relationships
between transcriptional circuits, expression level noise,
and epigenetics. CD4+ T cells form an important branch
of the adaptive immune system and function mainly by
secreting certain cytokines that regulate the behavior of
other cells, thus orchestrating the immune response [2].
Six major CD4+ T cell subtypes have been well charac-
terized to date: the naıve, T-helper 1 (Th1), Th2 and
Th17 cells, and induced and natural regulatory T cells
(iTreg and nTreg). Most of the (‘mature’-) subtypes are
derived from naıve CD4+ T cells, which enter a prolifer-
ation and differentiation process upon antigenic acti-
vation in the presence of cytokines (Figure 1a).
The phenotypic switches that CD4+ cells undergo upon
differentiation are linked to changes in epigenetic modi-
fications [19]. These switches are largely driven by TFs,
and illustrate the importance of master regulators, such as
Gata3 [20,21]. Recent studies on Th2 cells suggest that
phenotypic switches are mirrored by transcriptomic
switches [22�]. Protein expression levels of cytokines are
usually subject to large cell-to-cell variability, highlighting
the importance of noise in protein and mRNA expression.
To understand the interplay between these factors, sev-
eral questions need to be addressed: how do transcrip-
tional circuits mediate phenotypic switching? What are
www.sciencedirect.com
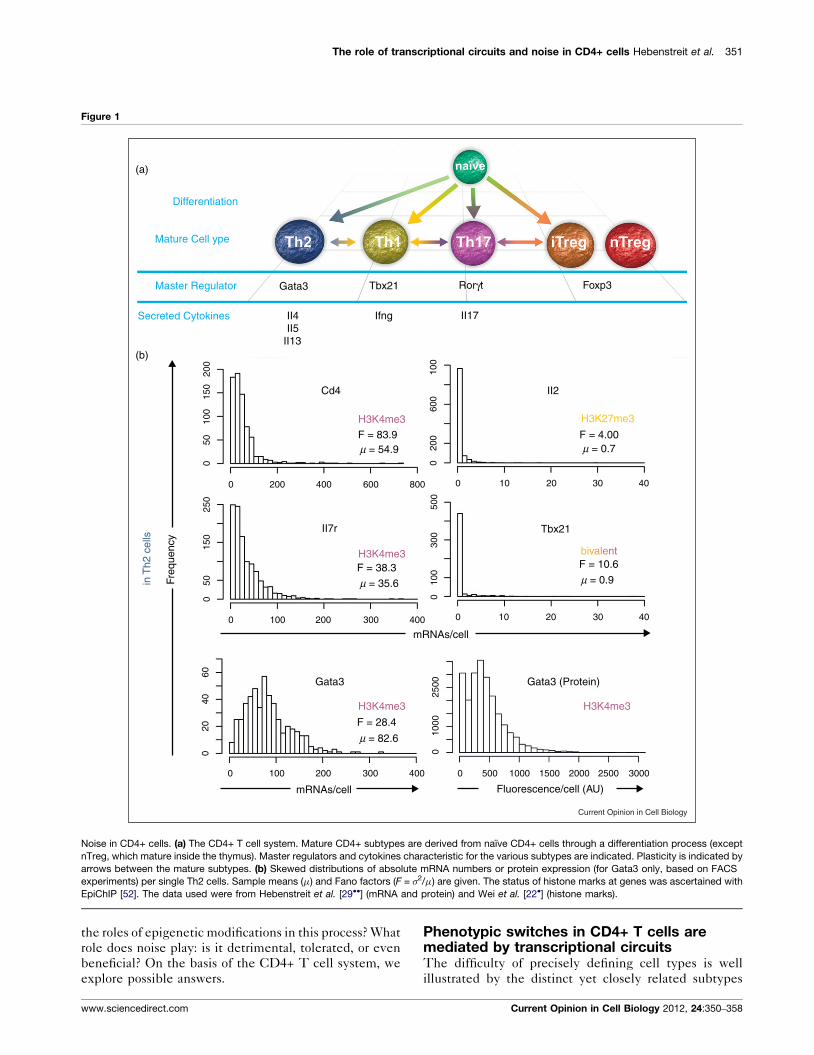
The role of transcriptional circuits and noise in CD4+ cells Hebenstreit et al. 351
Figure 1
(a)
Differentiation
Mature Cell ype
Master Regulator Gata3 Tbx21 Rorγt Foxp3
II17IfngII4II5
II13
Cd4
200
100
600
200
0
150
100
500
250
500
300
100
0
150
5060
2500
1000
0
4020
0
0 100 200
mRNAs/cell
mRNAs/cell
Fluorescence/cell (AU)
300 400 0 500 1000 1500 2000 2500 3000
0
0 100 200 300 400 0 10 20 30 40
0 200 400 600 800 0 10 20 30 40
II2
Tbx21
bivalent
II7r
Gata3
Freq
uenc
y
in T
h2 c
ells
Gata3 (Protein)
F = 83.9
F = 38.3
F = 28.4
F = 10.6
F = 4.00µ = 54.9
µ = 35.6
µ = 82.6
µ = 0.9
µ = 0.7
H3K4me3
H3K4me3
H3K4me3 H3K4me3
H3K27me3
Secreted Cytokines
(b)
Current Opinion in Cell Biology
Noise in CD4+ cells. (a) The CD4+ T cell system. Mature CD4+ subtypes are derived from naıve CD4+ cells through a differentiation process (except
nTreg, which mature inside the thymus). Master regulators and cytokines characteristic for the various subtypes are indicated. Plasticity is indicated by
arrows between the mature subtypes. (b) Skewed distributions of absolute mRNA numbers or protein expression (for Gata3 only, based on FACS
experiments) per single Th2 cells. Sample means (m) and Fano factors (F = s2/m) are given. The status of histone marks at genes was ascertained with
EpiChIP [52]. The data used were from Hebenstreit et al. [29��] (mRNA and protein) and Wei et al. [22�] (histone marks).
the roles of epigenetic modifications in this process? What
role does noise play: is it detrimental, tolerated, or even
beneficial? On the basis of the CD4+ T cell system, we
explore possible answers.
www.sciencedirect.com
Phenotypic switches in CD4+ T cells aremediated by transcriptional circuitsThe difficulty of precisely defining cell types is well
illustrated by the distinct yet closely related subtypes
Current Opinion in Cell Biology 2012, 24:350–358

352 Nucleus and gene expression
of CD4+ T cells (Figure 1a). Though each of the six cell
types illustrated in Figure 1a is associated with a particu-
lar master regulator and defined by a specific cytokine
profile, there are many reports of intermediate cell types
as well as cell-type switching after differentiation (i.e.
plasticity), and reprogramming. For instance, mature Th2
cells can be induced to give rise to a mixed Th1/Th2
phenotype, Th17 cells can be transformed into Th1 cells,
or iTreg cells can be differentiated towards Th17 cells
[23] (Figure 1a).
The cytokines that are secreted by mature CD4+ T cells
have crucial, often opposing roles in regulating the immune
response. Thus, one would expect a tight regulation in
maintaining lineage identity. The plasticity in the differ-
entiation process as a whole is due to the complexity of the
transcriptional regulatory circuits, which generally include
many more components than just a single master regulator.
In Figure 2b, a small transcriptional circuit involving the
master regulator Gata3 is illustrated. The circuit consists of
overlapping FFLs which facilitate the differentiation of a
CD4+ naıve cell to the Th2 type [24], and represents a
small subnetwork in a much larger, more complex regu-
latory network involved in phenotypic switching and in
stabilizing the cellular state.
Differentiation of CD4+ cells is strongly linked to cellular
proliferation as many studies show a percentage increase
of cytokine-secreting T cells in successive cell gener-
ations [25,26]. Notably, the percentage of differentiated
cells added after each division is small and thus suggests
the involvement of a stochastic mechanism. Where does
this randomness come from and how can we reconcile it
with the precision of immune responses?
Sources of stochasticity and their relation tocircuit topologyT cell differentiation is regulated by TFs. It has become
clear in recent years that transcriptional regulatory net-
works are subject to intrinsic noise in the expression of
their components [27]. This has been shown by tech-
niques that focus on single cells rather than population
averages, such as single molecule RNA-FISH, which
measures absolute numbers of mRNAs per cell [8].
One of the surprising findings from these assays was that
the distributions of mRNA numbers per cell are usually
non-Poissonian, meaning that the variances are higher
than the mean levels of transcripts. This gives some
mechanistic insight into transcription by RNA polymer-
ase II (RNAPII) by hinting at a burst-like production of
transcripts, where a gene is transcribed multiple times in
short periods of time [28]. Our studies on Th2 cells [29��]confirm such highly skewed, ‘noisy’ distributions of tran-
script numbers per cell for five genes classically associated
with CD4+ cell biology, including Gata3 and CD4 itself
(Figure 1b). Skewed distributions are observed for
Current Opinion in Cell Biology 2012, 24:350–358
protein expression levels too (Figure 1b), suggesting that
either the mRNA expression noise is not buffered com-
pletely or downstream processes serve as additional
sources of noise. An interesting possibility is that cell-
to-cell variation in the expression levels of genes such as
Gata3 may simply reflect the heterogeneity in terms of
their recent signaling experience. Thus some genes could
be specifically programmed to be sensitive indicators of
different levels of signaling molecules in each cell.
Another contributor to the wide range of expression levels
between individuals in a cell population could be asym-
metric cell division. Polarized segregation of proteins has
been observed upon antigen-induced T cell division,
where activation and differentiation induced factors were
preferentially found in the daughter cell close to the
immunological synapse [30,31]. This included Tbx21
and may also affect other TFs and master regulators.
As a consequence of these sources of noise, there may not
be a simple, discrete transcriptional ‘switch.’ This may
vary from cell to cell depending on the number of copies
of a set of signaling molecules, including TFs. As a result,
statistical parameters such as the average burst size or the
average waiting time may be altered upon transcriptional
activation of RNAPII. Importantly, burst-like transcrip-
tion demonstrates that transcription is an intrinsically
noisy process per se, and in individual cells this phenom-
enon can result in large deviations in transcript numbers
from the population average. In principle, random fluctu-
ations give rise to different transcription rates in different
cells over time, even if the cells started under identical
conditions. For CD4+ cells, the large variance in the
distributions shown in Figure 1b suggests that this may
be relevant to phenotypic switching, as it means that a
certain fraction of cells will have high numbers of mRNAs
from genes that are lowly expressed, and vice versa.
The presence of large cell-to-cell variation may indicate
that noise is somewhat inevitable. This might be due to
the fact that noise suppression is a biologically expensive
process: a recent study showed that suppression only
occurs with the fourth root of the number of control
factors in a transcriptional circuit [32��]. However, many
investigations in other cell types or systems have shown
that stochastic transcriptional switches can even be
advantageous over deterministic ones. For instance, sto-
chastically switching phenotypes or noisy transcriptional
circuits provide efficient mechanisms for adapting to
fluctuating or unpredictable environments [33–35]. It
has also been shown that FFL circuits that incorporate
miRNA can dampen noise [36], while certain FFL archi-
tectures have specific noise profiles [37] and may be
selected for these profiles, thereby facilitating the
sampling of multiple phenotypes [38].
While it is uncertain whether such phenomena play a role
in T cell biology, it is clear that the transcriptional
www.sciencedirect.com
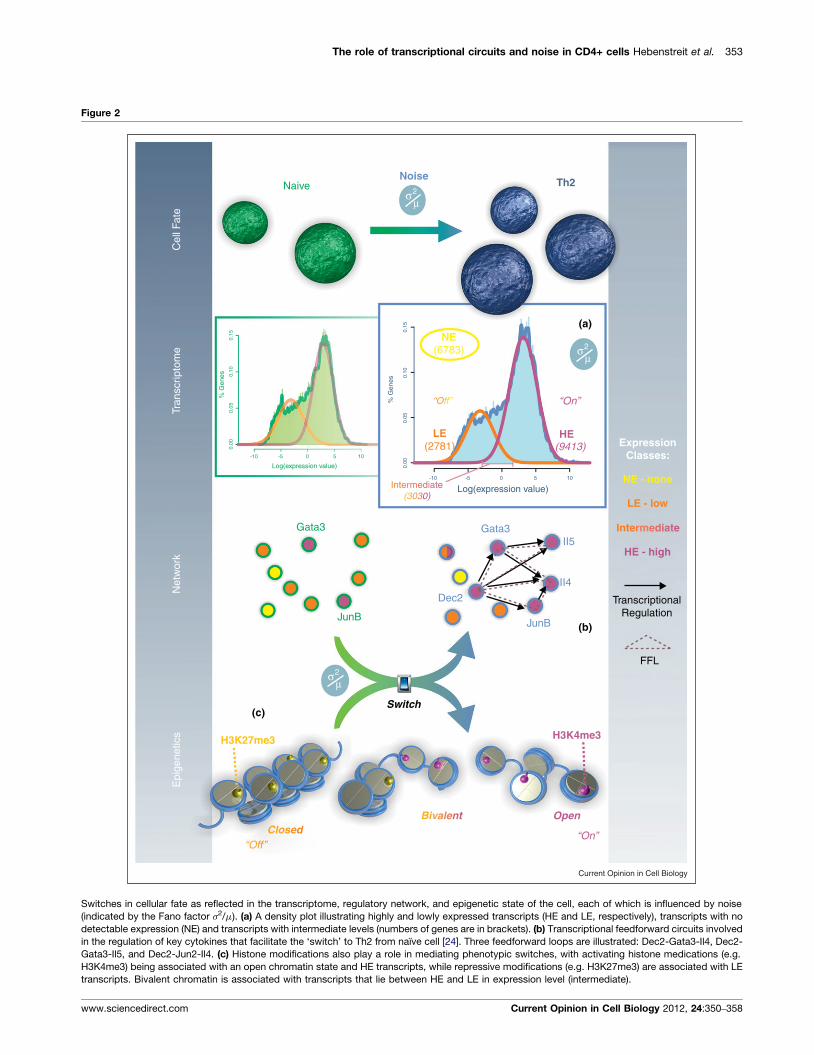
The role of transcriptional circuits and noise in CD4+ cells Hebenstreit et al. 353
Figure 2
Naive
0.15
0.10
% G
enes
0.05
0.00
-10 -5 0
Log(expression value)
5 10
Cel
l Fat
eTr
ansc
ripto
me
Net
wor
kE
pige
netic
sNoise
2σμ
Th2
ExpressionClasses:
NE - none
LE - low
HE - high
Intermediate
Intermediate(3030)
“Off”
TranscriptionalRegulation
(c)
(a)
(b)JunB
Dec2
Log(expression value)-10
0.00
0.05
% G
enes
0.10
0.15
-5 0 5 10
II4
II5Gata3
LE(2781)
NE(6783)
JunB
Gata3
Switch
H3K4me3
OpenBivalentClosed “On”
“Off”
“On”
(9413)HE
H3K27me3
FFL
2σμ
2σμ
Current Opinion in Cell Biology
Switches in cellular fate as reflected in the transcriptome, regulatory network, and epigenetic state of the cell, each of which is influenced by noise
(indicated by the Fano factor s2/m). (a) A density plot illustrating highly and lowly expressed transcripts (HE and LE, respectively), transcripts with no
detectable expression (NE) and transcripts with intermediate levels (numbers of genes are in brackets). (b) Transcriptional feedforward circuits involved
in the regulation of key cytokines that facilitate the ‘switch’ to Th2 from naıve cell [24]. Three feedforward loops are illustrated: Dec2-Gata3-Il4, Dec2-
Gata3-Il5, and Dec2-Jun2-Il4. (c) Histone modifications also play a role in mediating phenotypic switches, with activating histone medications (e.g.
H3K4me3) being associated with an open chromatin state and HE transcripts, while repressive modifications (e.g. H3K27me3) are associated with LE
transcripts. Bivalent chromatin is associated with transcripts that lie between HE and LE in expression level (intermediate).
www.sciencedirect.com Current Opinion in Cell Biology 2012, 24:350–358

354 Nucleus and gene expression
regulatory networks of CD4+ differentiation are subject
to intrinsic noise and the fundamental limit of noise
suppression mentioned above.
The role of epigenetic switches in CD4+ cellsOne mechanism that could have evolved in order to permit
a cost-efficient control of such transcriptional noise could
be the system of epigenetic modifications. Genes that
become differentially expressed during Th differentiation
are accompanied by changes in the chromatin landscape
[19]. For instance, the chromatin at subtype-specific loci
becomes relaxed, as illustrated by DNaseI hypersensitive
regions. Other subtype-specific genes are marked with
repressive histone modifications in cell types not expres-
sing them [39]. Mariani et al. showed that in Th2 cells,
stochastic Il4 expression patterns can be explained com-
putationally using a two-state model, where promoter
activation/inactivation states are explained in terms of
chromatin dynamics (opening and closing due to activating
or suppressing histone modifications). Here, different
populations of cells are created stochastically over gener-
ations due to chromatin dynamics [6,40�]. This work
provides support for a link between noise in gene expres-
sion levels and histone modification levels.
Similarly, our recent work relates gene expression with
histone modification levels, confirming such a two-state
model. In this study, we identified groups of lowly
expressed (LE) and highly expressed (HE) genes in
Th2 cells based on the global distribution of gene expres-
sion levels measured by RNA-sequencing (RNA-seq)
[29��], which is illustrated in Figure 2a. Here, we annotated
another subset of genes (derived from the LE group) called
not expressed (NE). NE genes are defined as genes with no
reads mapping to that particular transcript, which is de-
pendent on sequencing depth. This concept is similar to
the three expression classes of mRNA previously identified
by Hastie and Bishop [41]. While the HE genes featured
the activating histone mark H3K9/14 acetylation at their
promoters, the LE genes did not. The existence of the
lowly expressed, tissue-specific LE group of genes
suggests that the cell robustly tolerates a certain amount
of background expression, representing a type of ‘tran-
scriptional noise.’ This is often referred to as leaky expres-
sion and might be due to a tradeoff between the cost of
(nearly) completely repressed transcription and the
adverse consequences of stochastic expression [42,43].
The leaky expression reflects the inactive promoter state
in the two-state model, while HE transcripts are more
representative of the active promoter state.
In Figure 3, we analyzed the chromosomal organization of
NE, LE, and HE genes. As illustrated in Figure 3a, the
physical distances between adjacent HE and LE genes
were found to be less than between HE and NE genes.
This suggests that expression in LE genes may be due to
their proximity to HE genes. Similarly, as depicted in
Current Opinion in Cell Biology 2012, 24:350–358
Figure 3b, both HE and NE genes tend to form hom-
ogenous multigene clusters more often than expected by
chance. In contrast, LE genes usually occur alone. These
findings support a genomic organization illustrated in
Figure 3c, with LE genes close to clusters of HE genes,
and clusters of NE genes further away from HE gene
clusters. In this model, expression from neighboring HE
genes leads to leaky expression of neighboring LE genes
via some form of chromatin state spreading.
An interesting aspect of histone modifications is the
existence of ‘bivalent’ genes, which feature both activat-
ing and repressive marks. In mammalian cells, there is
increasing evidence from sequential ChIP and other
experiments that these marks are present simultaneously
at the same allele [44–48]. In Drosophila and Xenopus, it is
unclear whether this is the case [49–51]. In mouse
T-helper cells, Wei et al. observed such bivalent marks
at master regulator loci [22]. The authors argued that such
bivalent marks could poise them for expression. This
could explain the plasticity of these cell types as well
as why perfectly ‘pure’ cultures of single CD4+ subtypes
are difficult to obtain. Using CD4+ microarray expression
and H3K4me3/H3K27me3 histone modification data
(analyzed with Epichip [52]) from Wei et al., we found
that genes with bivalent marks have a higher probability
of differential expression upon maturation than genes
with the H3K27me3 mark or no modification (see
Figure 3d). They are also enriched in TFs relative to
the genomic background. Whether these bivalent genes
also have a specific noise profile is yet to be elucidated.
The functional relevance of noise intranscriptional regulatory networksThe question remains concerning how much of the
observed stochasticity in CD4+ gene expression is toler-
ated and how much is beneficial. With regard to the
beneficial aspects of stochastic expression, Guo et al.argued that probabilistic expression of the Th2 cytokine
Il4 by a fraction of cells is a more flexible mechanism of
immune regulation than fine-tuning of the exact Il4
amounts secreted by a homogenous cell population
[40�,53].
Another recently published study on CD8+ cells, closely
related to CD4+ cells, demonstrates how a certain amount
of ‘controlled’ randomness in protein expression gener-
ates useful variation in the antigen responsiveness [54�] of
the cell. This system is based on a combination of positive
and negative feedback loops and demonstrates that the
architecture of the regulatory circuits can have a profound
impact on the existence of stable states and/or extent of
noise. In line with this, a recent analytical study showed
that feedback systems provided by cytokine secretion and
stimulation, including downstream transcriptional inter-
actions, are sufficient for yielding mathematically stable
states that resemble cell types among CD4+ cells. The
www.sciencedirect.com
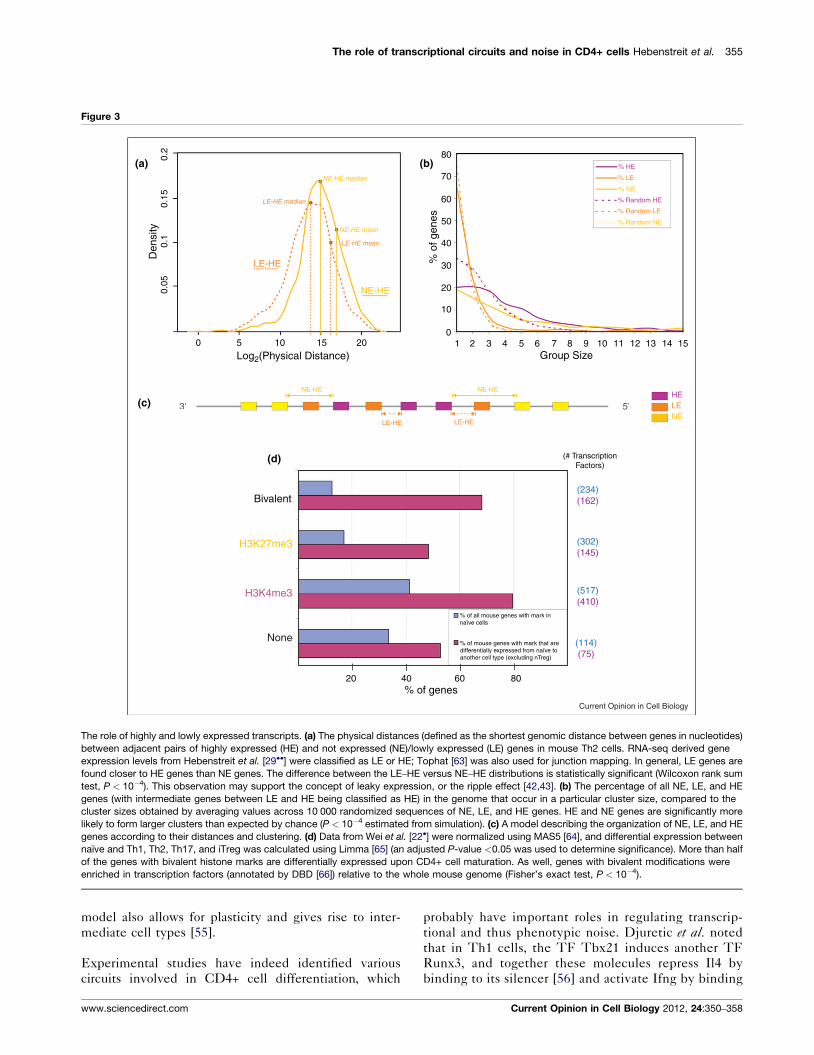
The role of transcriptional circuits and noise in CD4+ cells Hebenstreit et al. 355
Figure 3
(a) 0.2 80
70
60
50
40
% o
f gen
es
30
20
10
01 2 3 4 5 6 7 8 9
Group Size10 11 12 13 14 15
0.15
0.1
Den
sity
0.05
0
3’ 5’
5 10 15 20
20
Bivalent
HELENE
% HE
% Random HE
% Random LE
% Random NE
% LE
% NE
NE-HE
NE-HE
NE-HE
LE-HE
LE-HE
LE-HE mean
NE-HE mean
NE-HE median
LE-HE median
LE-HE
H3K27me3
H3K4me3
None
40 60% of genes
80
% of all mouse genes with mark innaïve cells
% of mouse genes with mark that aredifferentially expressed from naïve toanother cell type (excluding nTreg)
(# TranscriptionFactors)
(234)(162)
(302)(145)
(517)(410)
(114)(75)
Log2(Physical Distance)
(b)
(c)
(d)
Current Opinion in Cell Biology
The role of highly and lowly expressed transcripts. (a) The physical distances (defined as the shortest genomic distance between genes in nucleotides)
between adjacent pairs of highly expressed (HE) and not expressed (NE)/lowly expressed (LE) genes in mouse Th2 cells. RNA-seq derived gene
expression levels from Hebenstreit et al. [29��] were classified as LE or HE; Tophat [63] was also used for junction mapping. In general, LE genes are
found closer to HE genes than NE genes. The difference between the LE–HE versus NE–HE distributions is statistically significant (Wilcoxon rank sum
test, P < 10�4). This observation may support the concept of leaky expression, or the ripple effect [42,43]. (b) The percentage of all NE, LE, and HE
genes (with intermediate genes between LE and HE being classified as HE) in the genome that occur in a particular cluster size, compared to the
cluster sizes obtained by averaging values across 10 000 randomized sequences of NE, LE, and HE genes. HE and NE genes are significantly more
likely to form larger clusters than expected by chance (P < 10�4 estimated from simulation). (c) A model describing the organization of NE, LE, and HE
genes according to their distances and clustering. (d) Data from Wei et al. [22�] were normalized using MAS5 [64], and differential expression between
naıve and Th1, Th2, Th17, and iTreg was calculated using Limma [65] (an adjusted P-value <0.05 was used to determine significance). More than half
of the genes with bivalent histone marks are differentially expressed upon CD4+ cell maturation. As well, genes with bivalent modifications were
enriched in transcription factors (annotated by DBD [66]) relative to the whole mouse genome (Fisher’s exact test, P < 10�4).
model also allows for plasticity and gives rise to inter-
mediate cell types [55].
Experimental studies have indeed identified various
circuits involved in CD4+ cell differentiation, which
www.sciencedirect.com
probably have important roles in regulating transcrip-
tional and thus phenotypic noise. Djuretic et al. noted
that in Th1 cells, the TF Tbx21 induces another TF
Runx3, and together these molecules repress Il4 by
binding to its silencer [56] and activate Ifng by binding
Current Opinion in Cell Biology 2012, 24:350–358

356 Nucleus and gene expression
to its promoter. Hence, Tbx21 and Runx3 form FFLs
with Il4 and Ifng. In Treg cells, it has been shown by
Bruno et al. that the TF Runx can promote the expression
of the master regulator Foxp3 [57], and both of these TFs
regulate target cytokines such as Il2 thereby forming a
coherent FFL.
FFL circuits also play an important role in phenotype
determination in biological systems other than CD4+
cells. Foxp3 is known to suppress the oncogene Satb1.
Additionally, it was recently found that Foxp3 also forms
coherent FFLs involving miRNAs that bind to the 30-UTR of Satb1 (miR-7 and miR-155) [58] in breast tissue.
Satb1 also plays an important role in CD4+ cell biology,
particularly cytokine regulation. As noted previously, the
Th2-specific cytokines Il4 and Il5 are regulated by FFL
circuits (see Figure 2) [24]. At the epigenetic level, Satb1
remodels chromatin to create a 200 kb transcriptionally
active region associated with Stat6 and Gata3. These TFs
physically bridge this region with the Th2 ‘cytokine
locus,’ which contains the genes Il4, Il5, and Il13. Cai
et al. showed that this Satb1 mediated remodeling, in
addition to the presence of Stat6 and Gata3, is essential to
cytokine induction [59].
Finally, the regulation of cytokine receptors plays an
important role in the circuits that stabilize Th lineages.
This is supported by the aforementioned modeling
approach [55]. A well-known biological example includes
the IL-4 receptor a-chain (Il4ra), which is upregulated by
IL-4 via Stat6 [60,61] and thus constitutes a Th2-reinfor-
cing positive feedback loop.
ConclusionsStudies on CD4+ cells and other systems have high-
lighted the importance of transcriptional circuits and
other mechanisms in regulating noise and mediating
phenotypic switches. The strongly skewed mRNA
distributions suggest that the sharpness of lineage iden-
tities is subject to fundamental limitations. Some
examples indicate that there can be benefits from tol-
erating a certain amount of stochasticity, but in general
the cell employs multiple mechanisms to control noise.
These include the topologies of transcriptional networks
and circuits, and epigenetic mechanisms, as exemplified
by the discovery of the LE/HE transcript expression
classes. Future studies will provide insight into the role
of transcriptional circuits and epigenetics in broader
regulatory networks, how these networks affect noise,
and the probabilistic nature of phenotypic switches
and lineage stabilities. This may eventually lead to
more accurate, probabilistic definitions of these switches
and stabilities, paving the way for new pharmaceutical
strategies [62], and improving how we confront the
‘dueling fates’ of pathological versus normal cellular
phenotypes.
Current Opinion in Cell Biology 2012, 24:350–358
AcknowledgementsWe thank Guilhem Chalancon, Jesper Nissen and Valentina Proserpio forhelpful suggestions concerning this manuscript. The authors acknowledgethe Lister Institute (UK), the Medical Research Council (UK) grantreference nos. U105161047 and U105185859, and the Clinician InvestigatorProgram at the University of British Columbia (Canada) for their support.
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
� of special interest
�� of outstanding interest
1. Takahashi K, Yamanaka S: Induction of pluripotent stem cellsfrom mouse embryonic and adult fibroblast cultures bydefined factors. Cell 2006, 126:663-676.
2. Zhu J, Yamane H, Paul WE: Differentiation of effector CD4 T cellpopulations (*). Annu Rev Immunol 2010, 28:445-489.
3. Kalmar T, Lim C, Hayward P, Munoz-Descalzo S, Nichols J,Garcia-Ojalvo J, Martinez Arias A: Regulated fluctuations innanog expression mediate cell fate decisions in embryonicstem cells. PLoS Biol 2009, 7:e1000149.
4.��
Trott J, Surani A, Babu MM, Martinez-Arias A: Dissectingensemble networks in ES cell populations revealsmicro-heterogeneity underlying pluripotency. Mol BioSyst2012, 8:744-752.
In this work, it was found that individual mouse ES cells maintainedpluripotency by utilizing different transcription factor subnetworks. Thiswas discovered using single-cell gene expression profiling.
5. Swain PS, Elowitz MB, Siggia ED: Intrinsic and extrinsiccontributions to stochasticity in gene expression. Proc NatlAcad Sci U S A 2002, 99:12795-12800.
6. Miller-Jensen K, Dey SS, Schaffer DV, Arkin AP: Varyingvirulence: epigenetic control of expression noise and diseaseprocesses. Trends Biotechnol 2011, 29:517-525.
7. Silva-Rocha R, de Lorenzo V: Noise and robustness inprokaryotic regulatory networks. Annu Rev Microbiol 2010,64:257-275.
8. Raj A, van Oudenaarden A: Single-molecule approaches tostochastic gene expression. Annu Rev Biophys 2009,38:255-270.
9. Bruggeman FJ, Bluthgen N, Westerhoff HV: Noise managementby molecular networks. PLoS Comput Biol 2009, 5:e1000506.
10. Chalancon G, Ravarani C, Balaji S, Martinez Arias A, Aravind L,Jothi R, Babu MM: Interplay between gene expression noiseand regulatory network architecture. Trends Genet 2012 doi:10.1016/j.tig.2012.01.006.
11. Mangan S, Alon U: Structure and function of the feed-forwardloop network motif. Proc Natl Acad Sci U S A 2003,100:11980-11985.
12. Rodrigo G, Elena SF: Structural discrimination of robustness intranscriptional feedforward loops for pattern formation. PLoSOne 2011, 6:e16904.
13. Liu YY, Slotine JJ, Barabasi AL: Controllability of complexnetworks. Nature 2011, 473:167-173.
14. Muller FJ, Schuppert A: Few inputs can reprogram biologicalnetworks. Nature 2011, 478:E4 (discussion E4–5).
15. Mullen AC, Orlando DA, Newman JJ, Loven J, Kumar RM,Bilodeau S, Reddy J, Guenther MG, Dekoter RP, Young RA:Master transcription factors determine cell-type-specificresponses to TGF-beta signaling. Cell 2011, 147:565-576.
16. Bartel DP: MicroRNAs: target recognition and regulatoryfunctions. Cell 2009, 136:215-233.
17. Goldberg AD, Allis CD, Bernstein E: Epigenetics: a landscapetakes shape. Cell 2007, 128:635-638.
www.sciencedirect.com

The role of transcriptional circuits and noise in CD4+ cells Hebenstreit et al. 357
18. Natoli G: Maintaining cell identity through global control ofgenomic organization. Immunity 2010, 33:12-24.
19. Wilson CB, Rowell E, Sekimata M: Epigenetic control ofT-helper-cell differentiation. Nat Rev Immunol 2009,9:91-105.
20. Zhang DH, Cohn L, Ray P, Bottomly K, Ray A: Transcriptionfactor GATA-3 is differentially expressed in murine Th1 andTh2 cells and controls Th2-specific expression of theinterleukin-5 gene. J Biol Chem 1997, 272:21597-21603.
21. Zheng W, Flavell RA: The transcription factor GATA-3 isnecessary and sufficient for Th2 cytokine gene expression inCD4 T cells. Cell 1997, 89:587-596.
22.�
Wei G, Wei L, Zhu J, Zang C, Hu-Li J, Yao Z, Cui K, Kanno Y,Roh TY, Watford WT et al.: Global mapping of H3K4me3 andH3K27me3 reveals specificity and plasticity in lineage fatedetermination of differentiating CD4+ T cells. Immunity 2009,30:155-167.
This article provides ChIP-sequencing data for two histone modificationsin all major CD4+ cell types.
23. Murphy KM, Stockinger B: Effector T cell plasticity: flexibility inthe face of changing circumstances. Nat Immunol 2010,11:674-680.
24. Yang XO, Angkasekwinai P, Zhu J, Peng J, Liu Z, Nurieva R, Liu X,Chung Y, Chang SH, Sun B et al.: Requirement for the basichelix-loop-helix transcription factor Dec2 in initial TH2 lineagecommitment. Nat Immunol 2009, 10:1260-1266.
25. Bird JJ, Brown DR, Mullen AC, Moskowitz NH, Mahowald MA,Sider JR, Gajewski TF, Wang CR, Reiner SL: Helper T celldifferentiation is controlled by the cell cycle. Immunity 1998,9:229-237.
26. Gett AV, Hodgkin PD: Cell division regulates the T cell cytokinerepertoire, revealing a mechanism underlying immune classregulation. Proc Natl Acad Sci U S A 1998, 95:9488-9493.
27. Balazsi G, van Oudenaarden A, Collins JJ: Cellular decisionmaking and biological noise: from microbes to mammals. Cell2011, 144:910-925.
28. Suter DM, Molina N, Gatfield D, Schneider K, Schibler U, Naef F:Mammalian genes are transcribed with widely differentbursting kinetics. Science 2011, 332:472-474.
29.��
Hebenstreit D, Fang M, Gu M, Charoensawan V, vanOudenaarden A, Teichmann SA: RNA sequencing reveals twomajor classes of gene expression levels in metazoan cells. MolSyst Biol 2011, 7:497.
This article is the first to contain RNA-seq and single molecule RNA-FISHdata for mouse Th2 cells. It describes two classes of mRNA expressionlevels which are distinguished by histone marks at the promoters of thegenes.
30. Chang J, Palanivel V, Kinjyo I, Schambach F, Intlekofer A,Banerjee A, Longworth S, Vinup K, Mrass P, Oliaro J et al.:Asymmetric T lymphocyte division in the initiation of adaptiveimmune responses. Science 2007, 315:1687-1691.
31. Chang J, Ciocca M, Kinjyo I, Palanivel V, McClurkin C, Dejong C,Mooney E, Kim J, Steinel N, Oliaro J et al.: Asymmetricproteasome segregation as a mechanism for unequalpartitioning of the transcription factor T-bet during Tlymphocyte division. Immunity 2011, 34:492-504.
32.��
Lestas I, Vinnicombe G, Paulsson J: Fundamental limits onthe suppression of molecular fluctuations. Nature 2010,467:174-178.
This is an elegant theoretical study providing mathematical proofs for theextremely high costs of limiting expression noise in the cell.
33. Acar M, Mettetal JT, van Oudenaarden A: Stochastic switchingas a survival strategy in fluctuating environments. Nat Genet2008, 40:471-475.
34. Cagatay T, Turcotte M, Elowitz MB, Garcia-Ojalvo J, Suel GM:Architecture-dependent noise discriminates functionallyanalogous differentiation circuits. Cell 2009, 139:512-522.
35. Jothi R, Balaji S, Wuster A, Grochow JA, Gsponer J, Przytycka TM,Aravind L, Babu MM: Genomic analysis reveals a tight link
www.sciencedirect.com
between transcription factor dynamics and regulatorynetwork architecture. Mol Syst Biol 2009, 5:294.
36. Osella M, Bosia C, Cora D, Caselle M: The role of incoherentmicroRNA-mediated feedforward loops in noise buffering.PLoS Comput Biol 2011, 7:e1001101.
37. Ghosh B, Karmakar R, Bose I: Noise characteristics of feedforward loops. Phys Biol 2005, 2:36-45.
38. Kittisopikul M, Suel GM: Biological role of noise encoded in agenetic network motif. Proc Natl Acad Sci U S A 2010,107:13300-13305.
39. Hirahara K, Vahedi G, Ghoreschi K, Yang XP, Nakayamada S,Kanno Y, O’Shea JJ, Laurence A: Helper T-cell differentiationand plasticity: insights from epigenetics. Immunology 2011,134:235-245.
40.�
Mariani L, Schulz EG, Lexberg MH, Helmstetter C, Radbruch A,Lohning M, Hofer T: Short-term memory in gene inductionreveals the regulatory principle behind stochastic IL-4expression. Mol Syst Biol 2010, 6:359.
In this study, a combination of experimental and theoretical analysesreveals how stochastic mechanisms acting at the chromatin level can giverise to population heterogeneities of cytokine expression.
41. Hastie ND, Bishop JO: The expression of three abundanceclasses of messenger RNA in mouse tissues. Cell 1976,9:761-774.
42. Ebisuya M, Yamamoto T, Nakajima M, Nishida E: Ripples fromneighbouring transcription. Nat Cell Biol 2008, 10:1106-1113.
43. De S, Babu MM: Genomic neighbourhood and the regulation ofgene expression. Curr Opin Cell Biol 2010, 22:326-333.
44. Bernstein B, Mikkelsen T, Xie X, Kamal M, Huebert D, Cuff J, Fry B,Meissner A, Wernig M, Plath K et al.: A bivalent chromatinstructure marks key developmental genes in embryonic stemcells. Cell 2006, 125:315-326.
45. Roh T, Cuddapah S, Cui K, Zhao K: The genomic landscape ofhistone modifications in human T cells. Proc Natl Acad Sci U S A2006, 103:15782-15787.
46. Pan G, Tian S, Nie J, Yang C, Ruotti V, Wei H, Jonsdottir GA,Stewart R, Thomson JA: Whole-genome analysis of histone H3lysine 4 and lysine 27 methylation in human embryonic stemcells. Cell Stem Cell 2007, 1:299-312.
47. Stock J, Giadrossi S, Casanova M, Brookes E, Vidal M, Koseki H,Brockdorff N, Fisher A, Pombo A: Ring1-mediated ubiquitinationof H2A restrains poised RNA polymerase II at bivalent genes inmouse ES cells. Nat Cell Biol 2007, 9:1428-1435.
48. Brookes E, de Santiago I, Hebenstreit D, Morris K, Carroll T, Xie S,Stock J, Heidemann M, Eick D, Nozaki N et al.: Polycombassociates genome-wide with a specific RNA polymerase IIvariant, and regulates metabolic genes in ESCs. Cell Stem Cell2012, 10:157-170.
49. Schuettengruber B, Ganapathi M, Leblanc B, Portoso M,Jaschek R, Tolhuis B, van Lohuizen M, Tanay AGC: Functionalanatomy of polycomb and trithorax chromatin landscapes inDrosophila embryos. PLoS Biol 2009, 7:e13.
50. Gan Q, Schones D, Ho Eun S, Wei G, Cui K, Zhao K, Chen X:Monovalent and unpoised status of most genes inundifferentiated cell-enriched Drosophila testis. Genome Biol2010, 11:R42.
51. Akkers R, van Herringen S, Jacobi U, Janssen-Megens E,Francoijs K, Stunnenberg H, Vennstra G: A hierarchy of H3K4me3and H3K27me3 acquisition in spatial gene regulation inXenopus embryos. Dev Cell 2009, 17:425-434.
52. Hebenstreit D, Gu M, Haider S, Turner DJ, Lio P, Teichmann SA:EpiChIP: gene-by-gene quantification of epigeneticmodification levels. Nucleic Acids Res 2011, 39:e27.
53. Guo L, Hu-Li J, Paul WE: Probabilistic regulation of IL-4production in Th2 cells: accessibility at the Il4 locus. Immunity2004, 20:193-203.
54.�
Feinerman O, Veiga J, Dorfman JR, Germain RN, Altan-Bonnet G:Variability and robustness in T cell activation from
Current Opinion in Cell Biology 2012, 24:350–358

358 Nucleus and gene expression
regulated heterogeneity in protein levels. Science 2008,321:1081-1084.
This article shows how ‘controlled stochasticity’ can benefit the cell bygenerating useful variation during T cell activation.
55. Naldi A, Carneiro J, Chaouiya C, Thieffry D: Diversity andplasticity of Th cell types predicted from regulatory networkmodelling. PLoS Comput Biol 2010, 6:e1000912.
56. Djuretic IM, Levanon D, Negreanu V, Groner Y, Rao A, Ansel KM:Transcription factors T-bet and Runx3 cooperate to activateIfng and silence Il4 in T helper type 1 cells. Nat Immunol 2007,8:145-153.
57. Bruno L, Mazzarella L, Hoogenkamp M, Hertweck A, Cobb BS,Sauer S, Hadjur S, Leleu M, Naoe Y, Telfer JC et al.: Runx proteinsregulate Foxp3 expression. J Exp Med 2009, 206:2329-2337.
58. McInnes N, Sadlon TJ, Brown CY, Pederson S, Beyer M,Schultze JL, McColl S, Goodall GJ, Barry SC: FOXP3 and FOXP3-regulated microRNAs suppress SATB1 in breast cancer cells.Oncogene 2011:1-10.
59. Cai S, Lee CC, Kohwi-Shigematsu T: SATB1 packages denselylooped, transcriptionally active chromatin for coordinatedexpression of cytokine genes. Nat Genet 2006, 38:1278-1288.
Current Opinion in Cell Biology 2012, 24:350–358
60. Wei L, Vahedi G, Sun H, Watford W, Takatori H, Ramos H,Takahashi H, Liang J, Gutierrez-Cruz G, Zang C et al.: Discreteroles of STAT4 and STAT6 transcription factors in tuningepigenetic modifications and transcription during T helper celldifferentiation. Immunity 2010, 32:840-851.
61. Kotanides H, Reich N: Interleukin-4-induced STAT6 recognizesand activates a target site in the promoter of the interleukin-4receptor gene. J Biol Chem 1996, 271:25555-25561.
62. Amit I, Regev A, Hacohen N: Strategies to discover regulatorycircuits of the mammalian immune system. Nat Rev Immunol2011, 11:873-880.
63. Trapnell C, Pachter L, Salzberg SL: TopHat: discovering splicejunctions with RNA-Seq. Bioinformatics 2009, 25:1105-1111.
64. Hubbell E, Liu WM, Mei R: Robust estimators for expressionanalysis. Bioinformatics 2002, 18:1585-1592.
65. Smyth GK: Linear models and empirical Bayes methods forassessing differential expression in microarray experiments.Stat Appl Genet Mol Biol 2004, 3 Article3.
66. Kummerfeld SK, Teichmann SA: DBD: a transcription factorprediction database. Nucleic Acids Res 2006, 34:D74-D81.
www.sciencedirect.com