Download - TP Crowe Stress in Rocky Intertidal Comm

Journal of Aquatic Ecosystem Stress and Recovery7: 273–297, 2000.M.J. Attrill (ed.), Stress in Marine Communities, Part 2.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
273
Impacts of anthropogenic stress on rocky intertidal communities
T.P. Crowe1,∗, R.C. Thompson1, S. Bray2 & S.J. Hawkins1,21Biodiversity and Ecology Division, School of Biological Sciences, University of Southampton,Bassett Crescent East, Southampton SO16 7PX, U.K.2Centre for Environmental Sciences, University of Southampton, Highfield, Southampton SO17 1BJ, U.K.
Key words:anthropogenic stress, community, harvesting, human impact, introduced species, pollution,rocky shore, siltation, trampling
Abstract
Rocky shores provide a harsh environment for marine organisms and we briefly discuss natural sources of variationin community structure before considering anthropogenic impacts in detail. We review impacts caused by (a) acutedisturbances: oil spills, toxic algal blooms and (b) chronic disturbances: nutrient pollution, oil, heavy metals,pesticides, antifouling paints, collecting, trampling/habitat degradation, siltation and introduced species. Commu-nity level effects are emphasised throughout and illustrative examples are drawn from field-based case studies.Particular attention is given to the lessons learned from oil spills and the effect of chronic pollution by tributyltinon dogwhelks, the impacts of which ranged from the biochemical to community levels of organisation. Impactsare placed in a global and historical perspective and the potential for the recovery of shores under appropriatemanagement is discussed. Finally we consider the relative merits of the multivariate and univariate approaches tostudying impacted communities and suggest priorities for future research.
1. Introduction
Rocky shores occur throughout the coastlines of theworld’s oceans. They are the most extensive littoralhabitat on eroding wave-exposed coasts and are alsoa major habitat on more sheltered coastlines, such asfjords and rias. This already extensive natural habitatis further increased by the plethora of artificial hardstructures such as breakwaters, jetties, docks, groynes,dykes, sea walls which essentially function as artificialrocky shores.
Sheltered bays, fjords and rias are widely usedas harbours which range in size from small fishingports or marinas up to major industrial ports. Artificialshores generally occur in densely populated and oftenindustrialised regions, while even in remote places,rocky shores can be impacted by a variety of activitiesranging from subsistence collection of food throughto mining waste and occasional catastrophic oil spills.
* Corresponding author: Phone: 023 80594386; Fax: 02380594269; E-mail: [email protected]
Thus many rocky shores are subjected to a variety ofstresses caused by human activities. These anthropo-genic stresses are superimposed on the stress causedby natural environmental factors such as emersion inair due to the tides and wave action (see Raffaelli &Hawkins, 1996 for review).
In this review we start by briefly considering termsand definitions. We then outline natural stressors thatinfluence rocky shore communities and the problemof separating their effects from those of anthropo-genic stresses. We then consider acute and chronicanthropogenic stresses and review research into theirimpact on rocky shore communities. Special atten-tion is focussed on the acute impact of oil spillsand the chronic pollution caused by tributyltin inantifouling paints. We emphasise the need to inte-grate from individual responses up to the communitylevel. The discussion provides a global and historicalperspective. We consider the scales and relative impor-tance of different impacts and examine the potentialfor recovery of rocky shore communities. Finally

274
we highlight gaps in our knowledge and point outpriorities for future work.
We concentrate on the intertidal zone of rockyshores; coral reefs and subtidal habitats are outsidethe scope of this review but are occasionally discussed,particularly in relation to processes that have receivedlittle attention on rocky shores. Many of the ideas andexamples in this paper draw on publications in variousconference proceedings (e.g. Hawkins & Southward,1992; Hawkins et al., 1994; Hawkins, 1999a, b)and we have taken the opportunity to integrate andsynthesise these into this review.
2. Terminology
A variety of terms has emerged to describeenvironmental disturbances and their effects. Theenvironment is contaminated by input of man-madesubstances, but pollution is only said to occur whenthe contaminant actually has an effect on a biolog-ical variable (Clark et al., 1997). Similarly ‘stress’ is,broadly speaking, the response of a biological entity(individual, population, community, etc.) to a disturb-ance (or stressor). Hence anthropogenic stress is theresponse of a biological entity to an anthropogenicdisturbance. Stresses at one level of organisation (e.g.individual, population) may also have impacts on otherlevels, for example causing alterations in communitystructure. These terms are difficult to define preciselyand have been used in many different ways (see Parkeret al. (1999) for a detailed review). Inputs, disturb-ances or other sources of stress are generally describedas being either ‘chronic’ or ‘acute’. A chronic disturb-ance operates at a low level over a long period, whilean acute disturbance is a discrete, usually large event.These terms are roughly analogous to the terms ‘press’and ‘pulse’ which are used to describe environmentaldisturbances of long and short duration respectively(Bender et al., 1984). This latter terminology wasrecently refined by Glasby and Underwood (1996).Their classification distinguishes between the durationof the disturbance and the duration of the response ofthe target organisms or community (the impact). Theyrecognised that, for example, a disturbance of eithershort duration (pulse) or long duration (press) couldcause either no response, have a short term impact orcause a long term change in a population. Careful useof clearly defined terminology will aid well-designedresearch into human impacts on rocky shores. Few ofthe studies reviewed below were designed to discrim-
inate among these alternatives, however, and impactswill be discussed by reference to the acute and chronicdisturbances that caused them.
3. Separating natural and anthropogenic stress
Rocky shore organisms are subject to two majornatural gradients of physical stress (see Raffaelli &Hawkins, 1996 for review). First there is the verticalgradient of increasing stress with increasing exposureto air higher on the shore, due to the alternation ofthe tides. This gradient is essentially unidirectionalfor most marine organisms and is present on bothmicrotidal (e.g.<0.5 m the Mediterranean and Balticseas) and macrotidal shores (i.e. shores with tides ofup to 10 m or greater). Second, there is a horizontalgradient due to differences in wave action betweenexposed headlands and sheltered bays or inlets. Thisgradient is not unidirectional as some organisms arenaturally more abundant in wave swept conditions(e.g. suspension feeders such as mussels) others aremore abundant in shelter (e.g. some large algae suchasAscophyllum).
Within these two major gradients, conditionsvary under the influence of localised factors such asgeology which in turn affects the topography of theshore, resulting in complex variation in microhabitatcaused by pools, overhangs, crevices and boulders. Onmicrotidal shores these may completely override themajor gradients whilst on macrotidal shores they tendto modify the gradient locally, giving rise to patchyarrangements of physical conditions.
The other major environmental gradient is that ofsalinity in estuaries, which also has a major effect onpatterns of distribution and abundance. On a largerscale, there is an over-riding gradient of climate asso-ciated with geographic location.
Many rocky shore organisms must therefore betolerant of a wide range of stresses. Organisms usuallyoccur well within their range of tolerance of physicalconditions and are not stressed except under unusualconditions. To achieve this, they have mechanismsto deal with the stresses associated with alternatingimmersion and emersion in air (desiccation, osmoticstress, etc., see for example Newell, 1979; Davison &Pearson, 1996; Karsten et al., 1996). These mechan-isms may often be effective for withstanding anthro-pogenic stresses. Of particular note are behaviouralmechanisms to shut animals off from the environ-ment coupled with ability to withstand anaerobosis

275
which are equally effective in avoiding short pulses ofpollutant as spells of low salinity or emersion. Manyorganisms have well developed sensory mechanismsto detect and hence respond to stressors (e.g. Akberali& Davenport, 1982) and can also detect and respondto pollutants such as heavy metals (e.g. Redpath &Davenport, 1988). Many intertidal algae are equallyresilient and are able to tolerate considerable uptakeof heavy metals (Barreiro et al., 1993; Castilla, 1996)and withstand oil spills and dispersant application.Thus it is sometimes difficult to detect the effectsof anthropogenic stress at the level of the individualorganism.
Impacts are more often investigated at a popula-tion or community level. Populations and communitiesexhibit considerable variability in space and timeunder the influence of many physical and biologicalfactors (e.g. Hartnoll & Hawkins, 1985; Underwood,1985; Foster et al., 1988). On rocky shores, dramaticnatural fluctuations can be caused by factors suchas extremes in temperature (Crisp, 1964), red tides(Southgate et al., 1984) and variations in recruitmentsuccess (e.g. Bowman & Lewis, 1977; Caffey, 1985;Gaines et al., 1985; Underwood & Fairweather, 1989).Against such a background of variation, it is oftenextremely difficult to detect reliably the additionaleffects of human activities.
The difficulties in detecting the comparativelyminor effects of low-level chronic stress at thecommunity level on rocky shores have been appre-ciated for some time (Lewis, 1976, 1977; Hawkins& Hartnoll, 1983; Hiscock, 1985). In most cases,for community level effects to be expressed anddetected, the disturbance must either come from apoint source (e.g. a large effluent) or a catastrophicevent (e.g. an oil spill or the aftermath of a redtide). Recently the need for properly designed impactassessments has been emphasised (Underwood, 1992,1994). Such designs involve repeated sampling andcomparison with multiple control or reference sitesenabling natural spatial and temporal variation tobe objectively separated from putative impacts (seeUnderwood, 1992, 1994).
The majority of work on rocky shore communitieshas tended to concentrate on selected prominentspecies or functional groups analysed in a univariatemanner. Multivariate analysis of whole communities,which is common in research on soft sediment biota(e.g. Gray et al., 1990; Agard et al., 1993; Somer-field et al., 1995), has not been so widely used (butsee Hockey & Bosman, 1986; Fuji & Nomura, 1990;
Lasiak & Field, 1995; Underwood & Skilleter, 1996;Coates, 1998; Lasiak, 1998, 1999). The large amountof experimental community ecology that has beendone on rocky shores, however (Connell, 1972; Paine,1977, 1994; Underwood et al., 1983, Hawkins etal., 1992), often allows good interpretation of likelyimpacts on particular species and, in some cases,communities.
4. Acute anthropogenic disturbances
4.1. Oil spills
The best-recorded community level impacts ofanthropogenic stress are from oil spills. These havebeen well documented on rocky shores (Clark et al.,1997). Details of selected major spills are summarizedin Table 1. Most of these spills led to oil beingwashed onto rocky shores, but the major spill in theArabian/Persian Gulf during the Iraq/Kuwait warimpacted mainly depositing shorelines, althoughsome rocky areas were affected (Watt et al., 1993;Jones et al., 1998). Where oil became stranded on lowenergy depositing shores or mangroves, recovery wasgenerally slower than on more wave exposed rockyshores.
Table 1. Selected major oil spills (modified from Hawkins &Southward, 1992)
Ship or installation∗ Date Oil spilt Oil beached
(tons) (tons)
Persian Gulf∗ Jan ’91 >1million ?
Ixtoc 1∗ 3 Jun ’79 500 000 12 000
Amoco Cadiz 16 Mar ’78 233 000 80 000T
Torrey Canyon 18 Mar ’67 100 000 35 000
Braer 5 Jan ’93 85 000 ?
Sea Empress 21 Feb ’96 72 000 5–15 000
Khark 5 19 Dec ’89 70 000 ?
Metula 9 Aug ’74 51 000 42 000
Exxon Valdez 24 Mar ’89 38 000 4 500
Urquiola 2 May ’76 30 000 25 000
Aragon 30 Dec ’89 25 000 ?
Some costly early mistakes were made in cleaningup oil spills which often led to more damage beingdone during clean-up than caused by the oil itself (seeFoster, et al., 1990 for review). For example, as aresult of theTorrey Canyonoil spill in Cornwall, the

276
major damage was not caused by the 10 000 tons ofoil which came ashore, but by excessive treatmentwith over 14 000 tons of ‘first generation’ dispersants.These were later shown to be very toxic to marinelife (Corner et al., 1968). The dispersants killed thegrazerPatella vulgata, and, to a lesser extent, otherherbivores such asMonodonta lineataand Littorinaspp. This reduction led to dense growth of ephemeralgreen algae followed by an equally dense growth offucoids. This proliferation occurred on many of theexposed and moderately exposed shores of south-westCornwall (see Smith, 1968; Southward & Southward,1978; Hawkins et al., 1983; Hawkins & South-ward, 1992; Hawkins et al., 1994 for further details).Similar, but more localised, flushes of ephemeral algaeand fucoids were shown after theBraer (Newey &Seed, 1995) andSea Empressspills (Moore, 1998).Some dogwhelks survived theTorrey Canyonspilland were able to reproduce locally under the shelterof the fucoid canopy (Bryan, 1969). Barnacles wereonly affected in areas that were cleaned repeatedly.Over the next few years, however, they succumbedto smothering by algae and predation from the nowdense populations of dogwhelks. Of greatest signifi-cance was the highly favourable environment for thesettlement and early survival ofPatella vulgatacreatedby the fucoids.Patella depressaremained scarce in thearea until the mid 1980s (see Southward & Southward,1978; Hawkins & Southward, 1992).
We now have over 20 yrs of observations on therecovery of the shores affected by theTorrey Canyonspill. We also have a good background knowledge ofspatial and temporal variation of these systems to putthe results into context (see Southward & Southward,1978; Hawkins et al., 1992a). Recovery of key speciesto previously observed normal levels of spatial andtemporal variation was judged to have occurred within10–15 yrs. This recovery was through a series ofdamped oscillations (Southward & Southward, 1978;Hawkins et al., 1983; Hawkins & Southward, 1992)and a similar pattern is starting to emerge since themore recentExxon Valdezspill (Paine et al., 1996).Disruption of the community to such a vast extentresulted primarily from the widespread killing ofPatella. These limpets can be considered a ‘keystone’species on moderately exposed and exposed shoresthroughout the northeast Atlantic because of theimportance of their grazing (see Hawkins et al., 1992for review; Mills et al., 1993; Power et al., 1996;Raffaelli & Hawkins, 1996; Hurlbert, 1997 for recentcritiques of the keystone concept).
Since theTorrey Canyonspill there has been muchwork on the impacts of other spills both large (e.g.Myers et al., 1980; Conan, 1982; Shaw, 1992; DeVogelaere & Foster, 1994; van Tamelen, 1997; Joneset al., 1998) and small (e.g. Bowman, 1978; Popleet al., 1990; Lopes et al., 1997; Smith & Simpson,1998). Over the years various overviews and reviewshave been written (e.g. Southward, 1982; Lewis,1982; Hawkins & Southward, 1992; Suchanek, 1993)which summarize a diverse and often ‘grey’ literature.Paine et al. (1996) and Petersen (in press) both ablysummarize the explosion of literature following theExxon Valdezspill.
Oil spills have variable effects on the biota. Somespecies can be remarkably tolerant of oiling (e.g.barnacles, Southward & Southward, 1978; mussels,Newey & Seed, 1995), although generalization isdifficult as the degree of mortality of a particulargroup of animals varies among locations and with oiltype. For example, Smith (1968) and Southward andSouthward (1978) showed oil tolerance in barnacles insouth-west England to Kuwait oil following theTorreyCanyonspill whilst Newey and Seed (1995) demon-strated a decrease in cover due to Gullfaks Norwegiancrude oil following theBraerspill. Newly settled spathave also been shown to be much more resilient thanadults (Bonsdorff & Nelson, 1981). Other specieshave been impacted more by clean-up operations.Grazing molluscs seem to be particularly susceptibleto oil, oil/dispersant mixtures and dispersant itself (e.g.Smith, 1968; Crothers, 1983; Newey & Seed, 1995).Encouragingly, the later generations of dispersantsseem much less toxic than those used following theTorrey Canyonspill (e.g. Crothers, 1983).
Worries about the application of toxic dispers-ants have led to widespread use of physical cleaningmethods. These can also cause considerable disturb-ance and are made worse when high pressures ortemperatures are used. For example, large areas offucoid canopies (Fucus gardneri) were essentiallydestroyed by the combination of oil and clean up oper-ations using high pressure hot water following theExxon Valdez spill (De Vogelaere & Foster, 1994;Highsmith et al., 1996; Houghton et al., 1996; vanTamelen & Stekoll, 1996). Shaw (1992) ironicallypoints out that mechanical beach cleaning followingtheExxon Valdezoil spill caused destruction compar-able to that of the highly toxic dispersants followingtheTorrey Canyonspill.
Given the uncertainty about the impacts of actualoil spills it is surprising that so few replicated

277
field trials of the impacts of oil and various treat-ments have been attempted (but see Nelson, 1981,1982; Bonsdorff & Nelson, 1981; Bonsdorff, 1983;Crothers, 1983). In part, this is a reluctance of investi-gators to damage the environment by field trials onthe scale necessary to simulate oil spills. Crothers(1983) simulated oil spills in winter and summeron seaweed covered and limpet/barnacle dominatedshores. The experimental treatments were oil alone,dispersants alone, oil plus dispersants and controls.This work showed minimal effects on a seaweedcovered shore. On limpet-dominated shores, plotsreceiving a combination of oil and dispersant weremost affected, followed closely by oiled plots. Theeffect of the dispersant (BP1100WD) alone wasminimal. There was also greater decrease in barnaclecover in the oiled, and oil/dispersant plots than thecontrols and dispersant only plots. Bonsdorff andNelson (1981) simulated oil spills (Ekofisk crude oil)on the Norwegian coast. Mobile crustacea such asamphipods and isopods showed avoidance responses,but some were still killed. Spat ofSemibalanusbalanoideswere shown to be very sensitive, in contrastto adults which were only affected at the highest levelsof oiling.
Turning to warmer climes, considerable effort hasgone into assessing the impact of oil resulting fromthe 1991 Gulf War and subsequent recovery (e.g. Wattet al., 1993; Jones & Richmond, 1992; Abuzinada& Krupp, 1994; Jones et al., 1994; Readman et al.,1996). The account below is based on Jones et al.,1996. By seven months after the spill, most oil hadbeen deposited high on the shore between high waterneaps and high water springs, although it had clearlyaffected lower levels on the shore. During the first2–3 yrs there were considerable fluctuations as newcolonization of species such as barnacles occurred.There was a complete absence of most major speciesin the littoral fringe, and between 80% to 100% reduc-tion in species diversity in the upper eulittoral zone.Much of the oil rapidly hardened into tar. This began todisappear in 1991–1992, a process helped by coloniza-tion by algal mats within a couple of months, whicheventually peeled off with the oil. New recruitmentbegan to occur within a year or two where the oilhad weathered and peeled away. By 1994, diversityand abundance of individual species was still reducedfrom the midshore upwards by up to 50%, but normaldiversity had returned to the lower eulittoral. Themajor high shore gastropods,Nodolittorina subnodosa
andPlenaxis sulcatusonly returned to the most badlyaffected shore studied in 1995.
Various clean up methods were tried in the Gulf,including high pressure flushing on a rocky outcrop(Watt et al., 1993). No differences in recovery ratewere apparent between treated and untreated rockand Watt et al. (1993) expressed concern that anyfurther clean up might further decrease populations ofsnails with direct development such asNodolittorinasubnodosaandPlenaxis sulcatus.
Work on several smaller oil spills (e.g. Lopes etal., 1997; Pople et al., 1990; Smith & Simpson,1998) has also been undertaken. In Brazil, a crudeoil spill caused no discernible differences after thespill when compared to before (Lopes et al., 1997). Incontrast a spill of diesel on sub-Antarctic MacquarieIsland caused widespread mortality of marine inverte-brates over 2 km of shore, with reduced densi-ties still apparent 1 yr later (Pople et al., 1998).Seven years after the spill, macrobiotic communitieshad recovered in the intertidal, but impacted kelpholdfast communities were still very different fromuncontaminated controls. They were filled with sedi-ment showing traces of diesel and the communitieswere dominated by opportunistic worms (Smith &Simpson, 1998).
Interpreting the impact of oil spills is not easywhen there is a lack of data from before the spill.This is compounded by problems in selecting adjacentcontrol areas which are similar but not affected. It isalso difficult to persuade regulatory authorities to leavesome areas untreated to act as controls for comparisonwith cleaned areas. Resources tend to be deployedin excess during the immediate post-spill phase butas public interest wanes, so does funding for longerterm studies. Sadly most oil spills provide a missedopportunity for extraction of information (see alsocomments in Foster et al., 1990; Paine et al., 1996).The initial bonanza of interest from the news media isfollowed by a phase of investigation by scientists fromgovernment agencies, industry, consultancies, non-governmental organizations and universities. Fundingis not always allocated to the most qualified teams aspeer review processes cannot operate on the timescaleneeded to respond. Not surprisingly the quality of theresearch outcomes are often disappointing and severalauthors have urged better deployment of resources forthe next spill to enable high quality research (e.g.Foster et al., 1990; Hawkins & Southward, 1992;Paine et al., 1996).

278
4.2. Toxic algal blooms
Similar dramatic disruption of communities can occurafter ‘red tides’ of toxic planktonic algae (Smayda,1997). These occur naturally, but in recent years ithas been suggested that their frequency has increasedbecause of eutrophication (Hallegraeff, 1993; Paerl &Whittall, 1999). Although their devastating effects onfish and aquaculture have been widely publicised, littlehas been published on their impacts on rocky shorecommunities. Southgate et al. (1984) documented theeffects of a dinoflagellate bloom on the coast of Irelandand its effect on the shore community. Robertson(1991) reported major toxic effects caused by a bloomof Chrysochromulina polylepsisoff the Swedish westcoast. This killed up to 99% of the dogwhelks fromsome areas of the coast and affected the reproductiveoutput of the survivors in the following year. Similarlydevastating effects were seen in several habitats inNorway (e.g. Bokn et al., 1990). Gastropods, urchinsand starfish were severely affected, but mussels andoysters were more resilient.
5. Chronic anthropogenic disturbances
Impacts caused by long-term, low level disturbancesare summarised in Table 2 and described in detailbelow.
5.1. Nutrient pollution
Chronic influx of excessive nutrients originates mainlyfrom anthropogenic sources, particularly sewageoutfalls and run-off of agricultural fertilizers (Clarket al., 1997; Raffaelli & Hawkins, 1996). It can alsooccur naturally, for example at seabird colonies (e.g.Bosman & Hockey, 1988; Wootton, 1991). On wave-exposed rocky shores, where water movements aregreat, the effects of sewage effluent are usually local-ised to within about a hundred metres of the outfall(Fairweather, 1990), but effects are more pronouncedin enclosed bays or harbours (Raffaelli & Hawkins,1996).
In California, assemblages which are usuallydominated by large brown algae were replaced in theimmediate vicinity of an outfall by cyanobacteria andomnivores and suspension feeders capable of feedingdirectly on the effluent (Littler & Murray, 1975).The diversity of the assemblage was significantlyreduced relative to controls (Littler & Murray, 1975;
see Borowitzka, 1972; Fairweather, 1990; López-Gappa et al., 1990). Fairweather (1990) found thatthe animals dominant at control shores were replacedby ephemeral green algae near sewage outfalls inSydney. Similarly, López-Gappa et al. (1990) reportedthe total absence of the otherwise dominant musselBrachidontes rodriguezifrom the immediate vicinityof an outfall in Argentina and its replacement bycyanobacteria, diatoms and green algae. Limpetsoccurred in relatively low densities at a site close to theoutfall and tended to grow markedly faster there thanat a control site (Tablado et al., 1994). This pattern issimilar to that observed at sites enriched with guano(Bosman & Hockey, 1988).
Trends observed in the Baltic suggest that largescale eutrophication has had effects similar to thosecaused by point sources of nutrient pollution, causingthe decline of diverse assemblages dominated byperennial algae (especiallyFucus vesiculosus) andtheir replacement with species-poor turfs of greenephemerals (Schramm, 1996; Worm et al., 1999;see Bonsdorff et al., 1997; Diaz & Rosenberg,1995). Large scale variations in natural nutrient levelshave also been found to correlate with variationsin the functioning of intertidal communities (e.g.Bustamante et al., 1995; Menge et al., 1997). Nutrient-rich areas of coast had greater benthic microalgalproductivity, greater biomass of grazers (Bustamanteet al., 1995) and faster growth of filter feeders(mussels, barnacles; Menge et al., 1997) than areaswith low concentrations of nutrients. These patternsremain correlative, however, and in the case of Mengeet al.’s (1996) study, evidence was derived fromcomparisons of only two sites.
Small scale experimental investigations of theeffects of nutrient enrichment on rocky shorecommunities have yielded mixed results. Wooton et al.(1996) found that the influence of molluscan grazerson community structure was far greater than thatof nutrients. Micrograzers were the only group toincrease as a result of nutrient enrichment and therewas no change in algal biomass. Worm et al. (1999),working in shallow subtidalFucusbeds, found thatnutrient enrichment overrode grazer control of algalassemblages only where propagule banks had been leftintact.
5.2. Hydrocarbons
As the most visible and newsworthy form of pollu-tion, the majority of attention to ecological effects of

279
Table 2. Summary of major impacts to rocky shore populations or communities caused by chronic disturbances. A selection of key paperspresenting or reviewing evidence from field studies is listed
Stress Impact Location Reference
Nutrient enrichmentSewage outfall Large brown algae replaced by blue-greens California Littler & Murray, 1975
Reduced diversity of assemblageSewage outfall Mussels replaced by algae Argentina L´opez-Gappa et al., 1990
Reduced density & rapid growth of limpets " Tablado et al., 1994Hydrocarbons
Oil refinery Reduced density and diversity of mollusc community Curaçao, Caribbean Nagelkerken & Debrot, 1995Heavy metals
Copper mine tailings Barnacle assemblage replaced by copper resistant green alga Chile Castilla, 1996Antifouling paints
Tributyltin (TBT) Imposex in stenoglossan gastropods World-wide Ellis & Pattisina, 1990" Extinction of dogwhelks from many parts of range Europe Gibbs et al., 1991a
Thermal pollutionPower station Mortality of macroalgae Maine, USA Vadas et al., 1978
Displacement of native barnacles by exotic spp. Swansea, UK Langford, 1990Reduced diversity of assemblages in kelp holdfasts Anglesey, UK Bamber, 1991
HarvestingMussels No effect on abundance; stable Catch Per Unit Effort KwaZulu-Natal, S. Africa Kyle et al., 1997Mussels Reduced size and abundance of target spp. Transkei, S. Africa Hockey & Bosman, 1986Mussels Replaced by bare space or assemblages dominated Transkei, S. Africa Dye et al., 1994b
by coralline algae,Gelidiumor barnaclesMolluscs Reduced size and abundance of target spp. Victoria, Australia Keough et al., 1993Grazing gastropods Growth of macroalgae Chile Branch & Moreno, 1994
" California, USA Lindberg et al., 1998Macroalgae Reduced size and abundance of target spp. Chile Castilla & Bustamente, 1989
Increased access for grazers, so poor recolonisation Chile & S. Africa Santelices & Griffiths, 1994;of algae; if no grazers, opportunistic algae invade; Vasquez, 1995loss of holdfasts and associated fauna
TramplingReduced abundance of barnacles and mussels Oregon, USA Brosnan & Cumrine, 1994Reduced abundance of macroalgae; indirect effects Northumberland, UK Fletcher & Frid, 1996;on other species & S. E. Australia Keough & Quinn, 1998Increased bare space S. E. Australia Povey & Keough, 1991Increased bare space; shift to opportunistic species Northumberland, UK Fletcher & Frid, 1996
Introduced speciesLittorina littorea Altered community structure; displaced native Eastern USA Carlton, 1982, 1989
mudsnail in placesUndaria pinnatifida Replacement of dominant macroalga; shift in France, Australia, Walker & Kendrick, 1998
community structure USA
hydrocarbons has focussed on catastrophic oil spills(see Section 4.1). In terms of total volume, the inputfrom low-level chronic sources such as urban run offand routine tanker and refinery operations is muchgreater (Foster et al., 1988; Suchanek, 1993). Despitethis, experimental knowledge of the effects of long-term chronic oil pollution on littoral systems is gener-ally lacking (Southward, 1982). An exception is themesocosm study of Bokn et al. (1993 and referencestherein) in which low doses of diesel oil were deliveredto communities in large outdoor tanks for 2 yrs andchanges compared with those of unpolluted controls.
Although detailed inference was hampered bypsuedoreplication, some clear trends were apparent.Populations of animals, particularly mussels, tendedto be more susceptible than those of algae, a pattern
which is commonly observed in more acute spills (e.g.Stirling, 1977; Pople et al., 1990). There were noimpacts on herbivores in the experiment, althoughloss of herbivores has been common after acute spills(Foster et al., 1988) and has been noted at some chron-ically affected shores (Southward, 1982). Similarly,the density and diversity of mollusc communities ontropical rubble shores impacted by oil terminals for>7 yrs were greatly reduced relative to unpollutedcontrols (Nagelkerken & Debrot, 1995). Southward(1982) suggested that affected shores are generallycharacterised by reduced abundance of herbivores andincreased standing stocks of ephemeral algae.
There is also some evidence that settlement ofbarnacles may be locally inhibited by even smallquantities of oil (Crapp, 1971; Nelson, 1981).

280
Although further research is needed, effects on thesettlement of intertidal species could lead to signifi-cant modifications in community structure.
5.3. Heavy metals
The majority of heavy metals (copper, tin, mercury,cadmium, etc.) enter the marine environment vianatural pathways such as the erosion of ore-bearingrocks and volcanic activity (Clark et al., 1997).Human sources include sewage (e.g. Anderlini, 1992),industrial discharges and dumping and urban run-off(Clark et al., 1997). Some trace metals are metabol-ically important but can be toxic at higher concen-trations (Bryan, 1984). The toxicity of heavy metalsand the limits of tolerance of many organisms havebeen examined in detail in laboratory studies (Mance,1987). Many researchers have documented elevatedconcentrations of heavy metals in intertidal organisms(e.g. Barreiro et al., 1993; Blackmore et al., 1998) andused them as indicators of environmental contamina-tion (e.g. Bryan, 1984; Haynes et al., 1997). Thereis, however, little published research on the effects ofheavy metals on natural communities on rocky shores.
Laboratory studies have shown that heavy metalscan have complex effects on the growth of marineorganisms such as mussels and fucoid algae such thatdifferent combinations and concentrations of metalscan either promote or inhibit growth (Strömgren,1982; Munda & Hudnik, 1986). Heavy metals can alsoaffect aspects of larval development and are usuallytoxic to larvae at far lower concentrations than toadults (Beaumont et al., 1987; Fichet et al., 1998).Hoare et al. (1995) presented evidence that musselembryos in polluted areas showed increased resistanceto copper and indicated that this adaptation may havea genetic basis (see also Russell & Morris, 1970).
Impacts on the structure of communities on rockyshores would only be likely in bays, inlets or estu-aries with high residence time to allow concentra-tion of the metals. Since heavy metals can affectthe growth, reproduction and genetics of key species,elevated concentrations could potentially have far-reaching effects, but these have yet to be examined indetail. One of the few exceptions is Castilla’s (1996)comparison of assemblages on four shores in Chilebefore and after two of them were impacted by coppermine tailings. There were no differences amongthe shores before the copper mining commenced.Within two years of the commencement of coppermining activities, however, the barnacle-dominated
assemblage at the impacted sites was replaced byEnteromorpha compressa, an alga that is resistant tocopper, with associated changes in the communityrelative to controls. With the cessation of dumping ofuntreated tailings, there was a steady increase overfour years in the diversity at the formerly impactedsites. This indicates that the changes may be reversible(see also Correa et al., 1999).
5.4. Pesticides and antifouling paints
The toxicity and sublethal effects of pesticides, such asorganochlorines and organophosphates to rocky inter-tidal organisms has been explored in the laboratory(e.g. Sarojini et al., 1989; Lundebye et al., 1997), butto date there are no published field studies of effectson populations and communities.
Antifouling paints are essentially pesticides. Theside-effects of their leachates were often ignored untilthe early 1980s. Since then many studies have high-lighted the environmental impact of pollution causedby the biocide tributyltin (TBT) in antifouling paints(Figure 1; and see Rexrode, 1987; Bryan & Gibbs,1991 for reviews). The magnitude of these effects wasfirst realised when the collapse of the French oysterfishery in Arcachon Bay was attributed to increaseduse of TBT on boats in nearby harbours (Alzieu et al.,1986). Previously it had been thought that the concen-trations of TBT present in the water posed no directthreat to marine organisms due to the large dilutionfactors involved (Bellinger & Benham, 1978).
Although a multitude of organisms is susceptibleto pollution from TBT, none is as sensitive or hassuch clearly defined responses as those which areseen in stenoglossan gastropods, such as the commondogwhelk, Nucella lapillus, which is susceptible toconcentrations as low as 0.5 ng Sn l−1 (Gibbs et al.,1988, 1991a). TBT induces the development of malesexual characteristics in female dogwhelks (Bryan etal., 1986, 1988) a phenomenon termed ‘imposex’(Smith, 1980). This response has now been reportedin up to 45 species of stenoglossan gastropods world-wide (Ellis & Pattisina, 1990).
The effects of TBT onNucella lapillusare nowwell documented at the sub-cellular, individual andpopulation levels (Figure 2). Biochemical changesresponsible for the development of imposex arethought to be a consequence of an increase in thesteroid hormone testosterone in the female (Gibbs etal., 1991b). The female develops a penis and vasdeferens which leads to the occlusion of the genital
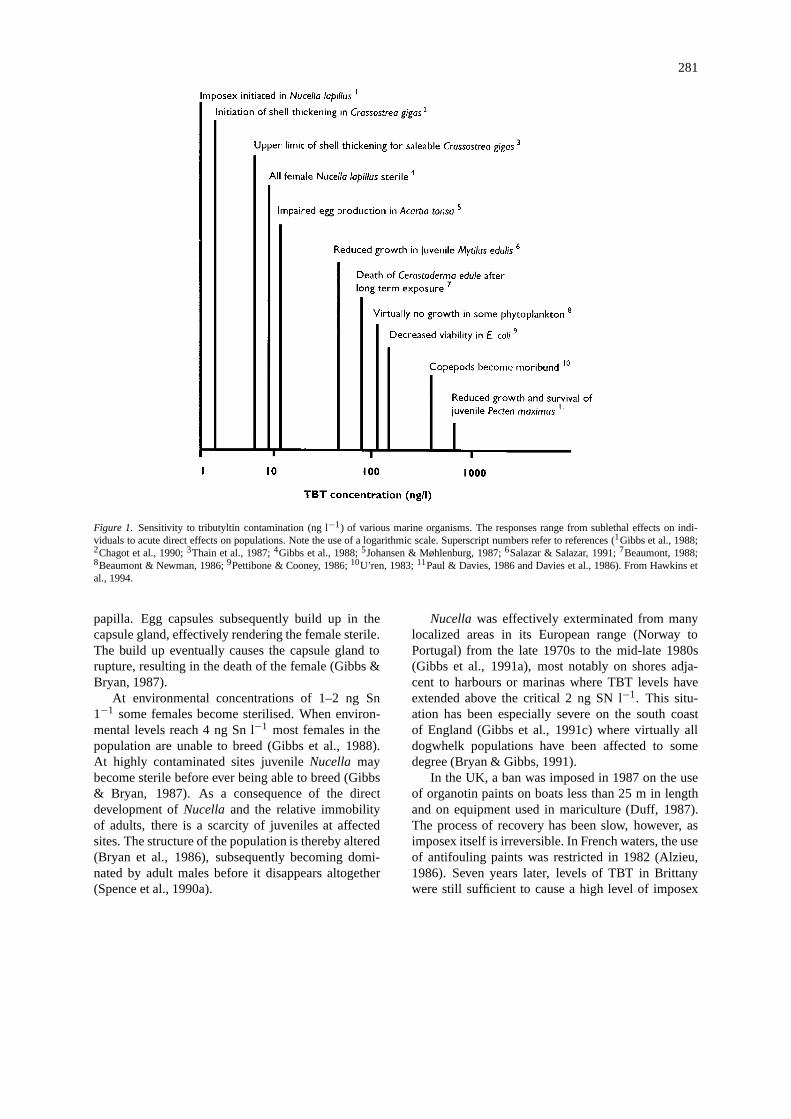
281
Figure 1. Sensitivity to tributyltin contamination (ng l−1) of various marine organisms. The responses range from sublethal effects on indi-viduals to acute direct effects on populations. Note the use of a logarithmic scale. Superscript numbers refer to references (1Gibbs et al., 1988;2Chagot et al., 1990;3Thain et al., 1987;4Gibbs et al., 1988;5Johansen & Møhlenburg, 1987;6Salazar & Salazar, 1991;7Beaumont, 1988;8Beaumont & Newman, 1986;9Pettibone & Cooney, 1986;10U’ren, 1983;11Paul & Davies, 1986 and Davies et al., 1986). From Hawkins etal., 1994.
papilla. Egg capsules subsequently build up in thecapsule gland, effectively rendering the female sterile.The build up eventually causes the capsule gland torupture, resulting in the death of the female (Gibbs &Bryan, 1987).
At environmental concentrations of 1–2 ng Sn1−1 some females become sterilised. When environ-mental levels reach 4 ng Sn l−1 most females in thepopulation are unable to breed (Gibbs et al., 1988).At highly contaminated sites juvenileNucella maybecome sterile before ever being able to breed (Gibbs& Bryan, 1987). As a consequence of the directdevelopment ofNucella and the relative immobilityof adults, there is a scarcity of juveniles at affectedsites. The structure of the population is thereby altered(Bryan et al., 1986), subsequently becoming domi-nated by adult males before it disappears altogether(Spence et al., 1990a).
Nucella was effectively exterminated from manylocalized areas in its European range (Norway toPortugal) from the late 1970s to the mid-late 1980s(Gibbs et al., 1991a), most notably on shores adja-cent to harbours or marinas where TBT levels haveextended above the critical 2 ng SN l−1. This situ-ation has been especially severe on the south coastof England (Gibbs et al., 1991c) where virtually alldogwhelk populations have been affected to somedegree (Bryan & Gibbs, 1991).
In the UK, a ban was imposed in 1987 on the useof organotin paints on boats less than 25 m in lengthand on equipment used in mariculture (Duff, 1987).The process of recovery has been slow, however, asimposex itself is irreversible. In French waters, the useof antifouling paints was restricted in 1982 (Alzieu,1986). Seven years later, levels of TBT in Brittanywere still sufficient to cause a high level of imposex

282
Figure 2. Summary diagram of the known effects of TBT on dogwhelks (Nucella lapillus) at a hierarchy of levels of organisation. FromHawkins et al., 1994.
in Nucellapopulations (Gibbs et al., 1991a). In someareas, recovery was thought likely to be inhibited bythe continued legal use of TBT on ships larger than25 m (Davies & Bailey 1991) and by the fact thatTBT stored in sediments continues to be leached intothe water column (Langston et al., 1990). Never-theless, abundance of dogwhelks is now increasingon the coasts of Britain, Norway and France, andthere is some debate about the wisdom of a total banin case alternative antifouling treatments prove to beeven more harmful (Pearce, 1999; Evans et al., 1998,2000a).
Since the initial visible stages of imposex inNucelladevelop at levels of TBT as low as 0.5 ng Snl−1, imposex can be used as a sensitive bioindicator ofTBT pollution (Gibbs et al., 1987). In addition to theirclear responses to TBT and world-wide distribution,their accessibility, commonness and ease of identifi-cation makes these gastropods ideal bioindicators forcontamination studies (Bryan et al., 1985; Bryan, 1984
but see Evans et al., 2000b). Consequently they arenow used to provide a world-wide indicator of TBTpollution (Bright & Ellis, 1990; Ellis & Pattisina,1990; Spence et al., 1990b; Smith & McVeagh, 1991).
Whelks have been shown to be important preda-tors in rocky shore communities world-wide (Connell,1961, 1970; Dayton, 1971; Underwood, 1978;Fairweather et al., 1984; Fairweather, 1987, 1988a,b). Despite speculation (Spence et al., 1990a; Bryan& Gibbs, 1991; Hughes & Burrows, 1993) no studieshave so far shown a clear community effect as aconsequence of TBT-induced dogwhelk decline.Recently, experiments on moderately exposed shoreson the Isle of Man were used to determine the likelyconsequences of their decline. On such shores, anatural cycle exists in which barnacle-dominatedassemblages alternate with fucoid-dominatedassemblages (summarised in Hawkins & Jones,1992). The alternation between these assemblagesis mediated by the grazing of limpets, which is

283
inhibited by the presence of barnacles. Algae thatsettle among barnacles are thus able to escape grazingand form mature clumps. Experimental reductions inthe density of dogwhelks increased the abundanceof barnacles relative to control plots, thus modifyingthe initiation and persistence of algal clumps andsignificantly altering the community structure anddynamics of these shores (Proud, 1994; Hawkins etal., 1994).
5.5. Thermal pollution
Thermal effluent has been a factor in the marineenvironment for a number of years and is largelyassociated with cooling water out-falls from powerstations and large industrial facilities. Studies of rockysubstrata exposed to thermal pollution are, however,scarce (reviewed by Langford, 1990). Some work hasbeen done on thermally impacted algae. For example,Vadas et al. (1978) showed thatFucus vesiculosisdeclined rapidly andAscophyllum nodosumwas alsodetrimentally affected by thermal extremes and rapiddiurnal fluctuations caused by a cooling water outfallin Maine. They pointed out, however, that althoughthis gave an insight into the response of individualorganisms, the response of the community as a wholeremained largely unknown.
In some cases exotic species colonise hardsubstrata where a cooling water out-fall is present.For example, in a thermally affected dock in Swansea(UK), communities of introduced barnaclesBalanusamphitrite and Elminius modestusshowed enhancedbreeding (Naylor, 1965a, b; reviewed by Langford,1990). The indigenous species,Balanus crenatus,did not breed in the heated water (>20 ◦C), but B.amphitritedid and consequently replacedB. crenatus.
The only study to date which has identifiedcommunity-wide effects was done at Wylfa NuclearPower Station by Bamber (1991), who founddecreasing diversity in the communities associatedwith Laminariaholdfasts with increasing proximity tothe cooling water out-fall. Interpretation is hampered,however, by the fact that no information was given onactual temperatures or on the presence of biocides inthe cooling water. This is a point highlighted by Vadas(1979): ‘There are numerous problems confoundingnearly all field studies [in this area]. Often thermalplumes produce or induce indirect effects, mostcommonly increased turbidity and sedimentation indischarge areas [reducing photosynthetically avail-able light and adversely affecting filter feeders]. Also
toxicity from chlorination to reduce fouling or fromheavy metals eroded from cooling pipes may occur.’Therefore separating the community effects of thermaldischarge from other factors associated with the out-fall is problematic.
The effects of thermal discharge at the communitylevel requires further investigation. Whether thermaleffluents can have a permanent impact on communitieson rocky substrata is unknown. In the case of theSwansea Docks, however, when the thermal dischargewas removed, the exotic species declined and indi-genous species were restored (Naylor, 1965a, b).
5.6. Collecting and gathering
Man has utilised intertidal organisms as a source offood since prehistoric times (for review see Siegfriedet al., 1994; Griffiths & Branch, 1997; Castilla, 1999).These plants and animals still provide a major resourcefor human populations in coastal regions of America,Africa (see Siegfried, 1994), Asia (Akimichi, 1995),Australasia (Underwood, 1993a) and some oceanicislands (e.g. Azores, Santos et al., 1995). Man’sexploitation of the intertidal is facultative and, unlikemost other intertidal predators, it is not directly influ-enced by density dependent predator-prey relation-ships, nor by the physical factors (such as storms,desiccation stresses) which regulate the abundance ofprey species (Dye et al., 1994a). Intertidal organismsare especially vulnerable since they are either sessileor their movement during low tide (the principal timefor collection) is confined to refuges such as pools,crevices or habitats beneath boulders. With the excep-tion of chemo-defence, via toxins, most of the anti-predator defences used by these organisms are inef-fective against man (Hockey, 1994). Consequently,there is considerable concern that certain species arebeing over-exploited and marine habitats are beingdamaged (Newton et al., 1993; Griffiths & Branch,1997).
A variety of organisms is collected (over 30 spp. ineach of South Africa and South America see Siegfriedet al., 1994). Mussels, limpets, abalone, whelks,winkles, octopus, crabs, lobsters, barnacles, echino-derms, ascidians, and algae are collected for humanconsumption, as animal feed or as fishing bait (Kings-ford et al., 1991; Keough et al., 1993; Underwood,1993a; Addessi, 1994; Siegfried, 1994). Numerousspecies of invertebrates, especially molluscs, are alsocollected for their ornamental value as marine curios(Wells, 1980; Newton et al., 1993). More recently

284
there has also been substantial increase in the collec-tion of live fish and invertebrates for aquaria (Addessi,1994). Substantial quantities of algae are collectedfrom intertidal and subtidal habitats, e.g. over 180 000t per annum in Chile and 30 000 t per annum in SouthAfrica (Siegfried et al., 1994), for production of agar,alginates, carrageenans, and to a lesser extent as afood source, or as a fertiliser (Santelices & Griffiths,1994; Vasquez, 1995). Species with low dispersalare particularly vulnerable to overexploitation, as areisolated populations such as those on oceanic islands(see Webber et al., 1998).
Harvesting can be divided into subsistence orrecreational gathering and commercial collection.Subsistence gathering has taken place, for the mostpart in a sustainable manner, for at least 100 000 yrs(Siegfried et al., 1994; Kyle et al., 1997). However,this type of collection is having a greater impactwith increasing use of non traditional equipment tofacilitate harvesting, transport and storage snorkels,SCUBA, outboard motors, freezers etc. (Underwood,1993a; Santos et al., 1995). Recreational collectorstypically gather invertebrates to use as fishing bait,as ornamental souvenirs, or for consumption (Under-wood, 1993a) and while the take per person maybe quite small (e.g. 3 to 4 crabs per person perday), cumulative effects can be considerable (Kings-ford et al., 1991; Underwood, 1993a). Commercialharvesting is a more intensive form of exploitationand provides a substantial source of export incomein some countries (e.g. US $200 million for Chileduring 1980, Siegfried et al., 1994). It is more system-atic and potentially more threatening than subsistencegathering since a few species are repeatedly targeted.
Some studies have shown little effect of collec-tion, indicating that resources are being removed ina sustainable manner (Kyle et al., 1997). However,in most cases target species are considerably lessabundant, and are of smaller average size and biomassin areas where they are exploited than in compar-able areas where collection is minimal (Hockey &Bosman, 1986; Castilla & Bustamante, 1989; Keoughet al., 1993; Underwood, 1993a). Apart from directeffects on the particular target species, collection isalso likely to have indirect effects on organisms whichare not exploited, and hence the assemblage as awhole (Figure 3). This is especially true when thetarget species has a ‘keystone’ role in structuring thecommunity (Dye, 1992; Underwood, 1993a; Branch& Moreno, 1994; Griffiths & Branch, 1997; Lasiak,1998, 1999; but see Sharpe and Keough (1998) for
an example of minimal indirect effects). Removal ofgrazers such as limpets or urchins, for example, hasbeen shown to result in proliferation of micro and thenmacroalgae on the shore (Hawkins & Hartnoll, 1983;Branch & Moreno, 1994; Dye et al., 1994b; Lind-berg et al., 1998). Conversely, removal of macroalgaeleads to loss of habitat for small invertebrates whichlive amongst the algae, and will also create spacewhich may become colonised by larger invertebrates,and opportunistic algae (Santelices & Griffiths, 1994;Vasquez, 1995). Over-exploitation of large vertebratepredators, such as sea otters, which feed in the inter-tidal at high tide has also lead to extensive changesin community structure on some shores (Estes &Palmisano, 1974; see Estes & Duggins, 1995). Collec-tion can also have indirect effects on predators, whichrely on commercially collected organisms as a foodresource. For example, over-exploitation of limpetsis believed to have contributed to extinction of theCanarian black oystercatcher which feeds on thesemolluscs (Hockey, 1987).
A range of management strategies is available tohelp reduce and regulate collection (for reviews seeUnderwood, 1993a; Dye et al., 1994a; Odendaalet al., 1994; Griffiths & Branch, 1997). There arenumerous examples where reductions in the amountof collection has led to an increase in the abund-ance of particular target species within protected areas(e.g. Castilla & Duran, 1985; Oliva & Castilla,1986). Increases in the abundance of adults may alsoenhance local recruitment of juveniles (Prince et al.,1988; Davis, 1995), however evidence of enhancedrecruitment to neighbouring areas is less apparent (seeCastilla & Bustamante, 1989; McClanahan & KaundaArara, 1996). It is difficult to develop managementstrategies which will be generally applicable becauseof the variety and complexity of ecosystems. Theseproblems are further complicated by temporal andspatial variability in recruitment to target populations(Hockey, 1994; Harris et al., 1998). Managementstrategies should therefore consider both the popula-tion and the community level of resolution (Keough &Quinn, 1991; Dye et al., 1994a; Griffiths & Branch,1997; Keough & Quinn, 1998). However, collectingand interpreting multi-species data in order to makepredictions at the community level is difficult and maytend to obscure trends for individual species (Keough& Quinn, 1991).
The complexity of resource management is illus-trated by an example from marine reserves inChile. Here a large predatory gastropod (Concholepas

285
Figure 3. (A and B) MDS ordinations based on Bray-Curtis indices of similarity derived from root-root transformed mean macrofaunalabundance and biomass estimates respectively (circles, ovals and squares represent study sites in the southern, central and northern regionof Transkei respectively; open and shaded areas represent exploited and non-exploited sites respectively. Letters/number combinations (e.g.M1, M2, etc.) refer to specific localities – see Lasiak (1999) for details. Reproduced with permission.
concholepas) is one of the main species collectedcommercially from the intertidal zone (25 000 t in1980). Creating a reserve to exclude harvesters fromthe shore led directly to a dramatic increase in theabundance ofC. concholepasand two species oflimpet which had also been exploited previously. Thiswas followed by a reduction in the abundance ofmussels, one ofC. conchelepas’ main prey items, andan increase in the abundance of algae. This was a tran-sient state as grazing by limpets substantially reducedthe abundance of algae, creating space which wassubsequently colonised by barnacles. Hence reducingcollection lead to varied and to some extent unpre-dictable changes in community structure and verticaldistribution of these organisms (Duran & Castilla,1989; Branch & Moreno, 1994). Both types ofcommunity (exploited and unexploited) are commonalong the whole coast of Chile and it is evident thathuman exploitation is affecting the shoreline at alandscape scale of resolution (Paine, 1994).
Some management strategies may actually becounter-productive. For example, introducing a size-restriction to prohibit removal of smaller individualsfrom the population may lead to a relative increasein the collection of larger more fecund individuals.For many marine invertebrates the greatest natural
mortality is on small individuals, with larger indi-viduals escaping predation and making a major contri-bution to reproductive output (Underwood, 1993a).This can be especially important in protandrousspecies such as limpets. Hence a more appropriatestrategy may be to have both lower and upper sizelimits. Lower limits would be set to allow most indi-viduals to breed once or twice before harvest whileupper limits would protect larger particularly fecundindividuals and females in protandrous species.
Mariculture may help reduce exploitation ofnatural populations. For example, in South Africa75% of the mussel fishery is now supplied from artifi-cially reared populations (Siegfried et al., 1994). Manymarine algae have also been successfully cultivated(for review see Vasquez, 1995). There are, however,considerable environmental problems associated withintensive rearing in mariculture (for review see OceanStudies Board, 1995).
5.7. Trampling and habitat degradation
Habitat degradation as a result of either collectingorganisms from the shore or by trampling on the shore,perhaps to gain access to the sea for swimming orSCUBA diving, is a major problem (Liddle & Kay,

286
1987; Hawkins & Roberts, 1992). There is a directcausal relationship between this type of human activityand damage to intertidal habitats. Damage from recre-ational visitors is more intensive during weekends andholiday periods (Kingsford et al., 1991; Vanherwerden& Griffiths, 1991) and decreases with distance fromthe nearest point of easy access (Addessi, 1994;Pombo & Escofet, 1996). Experimental studies intemperate regions have shown that the abundance ofbarnacles, mussels and macroalgae may be directlyreduced as a result of trampling with indirect changes(both increases and decreases) in the abundance ofother species (Brosnan & Crumrine, 1994; Fletcher& Frid, 1996; Keough & Quinn, 1998). This maybe accompanied by an increase in the amount ofbare space and subsequent colonisation by opportun-istic species (Povey & Keough, 1991; Fletcher &Frid, 1996). In the tropics, trampling on the reef flatcauses direct damage to both branched and massivecorals (Liddle & Kay, 1987). Apart from physicaldamage to marine organisms, human presence mayfrighten away predators, such as oystercatchers whichfeed on the shore while the tide is out. This mayhave indirect effects on the abundance of prey specieswhich are usually eaten by these birds (Lindberg et al.,1998).
Somewhat ironically, marine reserves frequentlybecome a focus for visitors to the seashore and thereis concern that the recreational carrying capacity ofshores in some reserves may be exceeded, leading tohabitat degradation. If the reserve is large enough thisproblem can be reduced by restricting public accessto a selected area, such as that adjacent to the carpark (Fletcher & Frid, 1996). Ideally, measures toreduce habitat degradation should be considered inparallel with those aimed at reducing collection inthe intertidal. This can be achieved as part of anintegrated Coastal Zone Management plan where arange of strategies offering differing levels of protec-tion are applied along a coastline with a view tomaximising the total utility of the zone. Additionalmeasures such as elevated boardwalks, fences, andvisitor education may also be effective in reducing theamount of damage caused by each individual visitor(Carlson & Godfrey, 1989; Fletcher & Frid, 1996).However, recovery after the trampling disturbance isremoved may be both variable and slow. Experimentalstudies on temperate shores have shown that somealgal and barnacle cover returned after one year (Povey& Keough, 1991), while larger invertebrates such asmussels had not completely recovered ten years after
trampling ceased (Dye, 1992; Brosnan & Crumrine,1994).
A final consideration should also be given tothe potential damage from scientific activity. Rockyshores provide a very tractable system and a greatdeal of research has been done on them. Some of themost studied shores are now riddled with remnants ofprevious research (drill holes, plastic screw anchors,screws, settlement tiles etc., pers. obs.). These activ-ities tend to be very localised and the effects if any(we are not aware of any published work assessingthis aspect) are likely to be minor (but see Brutonand Stobbs (1991) for evidence of overcollection bymarine scientists). However, if scientists hope tocredibly promote conservation strategies they mustadopt a responsible attitude to ensure minimal damage(see Hawins (1999) for further discussion).
5.8. Siltation
Rocky shore communities may potentially beimpacted by increased sediment loading in thewater column. This can result from urbanization ofcoastlines and watersheds in their hinterlands leadingto suspended material being transported by rivers orrun off into the coastal zone. It is also a problem inareas adjacent to intensive irrigated agriculture orwhere overgrazing has led to soil erosion. To datethere has been considerable attention given to theimpacts of increased silt load on coral reefs (e.g.McClanahan & Obura, 1997; Ochoa Lopez et al.,1998; Wesseling et al., 1999). In contrast this is aneglected area in rocky shore ecology, although somework, particularly in shallow subtidal habitats, hassuggested that increases in sedimentation can leadto alterations in assemblages, including proliferationof algal turfs (Thom & Widdowson, 1978; Seapy& Littler, 1982; Airoldi et al., 1995, 1996; Airoldi& Cinelli, 1997; Airoldi, 1998; Airoldi & Virgilio,1998).
5.9. Introduced species
Intertidal communities are also subject to stress fromspecies that have been introduced accidentally orintentionally by man. Most accidental introductionsare transported as fouling on the undersides of ships,or in ships’ ballast water (for reviews see Lewis,1964; Raffaelli & Hawkins, 1996).Elminius modestusbecame established in Europe following introductionby shipping during the Second World War (Lewis,1964 for review). Although widespread in occurrence,

287
its impact on local barnacle assemblages is primarilyrestricted to estuaries and bays where it can becomethe dominant barnacle. On more exposed coastlines, ithas stabilised at a lower level and appears to coexistwith the native barnacles (Southward, 1991).
Alternatively, species may be introduced as, orwith, organisms which are transplanted for aquacul-ture. For example, the edible periwinkle (Littorinalittorea) was introduced, probably accidentally, tothe east coast of America in the middle of the 19thcentury where it had a major impact on rocky shorecommunities, and in some locations displaced thenative mud snail (Ilynassa obsoleta) (Carlton, 1982,1989). Other examples include introduction to Europeof the slipper limpet and oyster drill from Americaand the marine algaSargassum muticumfrom Japan(Carlton, 1989; Critchley et al., 1990; Southward,1991; Raffaelli & Hawkins, 1996). The range ofSargassumis increasing dramatically and has beenshown to have modified macroalgal assemblages innorthern Spain and the Pacific coast of North America(Walker & Kendrick, 1998).
In many cases these introductions have not so farbeen found to have dramatic influences on marinecommunities. However, the potential for major andprobably permanent impact remains, especially ifpathogens are transferred with introduced species andfind new hosts which lack natural resistance, as hasoccurred with Salmonids transferred for aquacultureand stock enhancement (for review see Jonsson &Flemin, 1993).
6. Overview
6.1. Scale and relative importance of impacts
It is clear that rocky shores are subject to a varietyof impacts on a range of spatial and temporal scales.On a global scale, there can be few pristine shores,even in the most remote regions. Most impacts arelocalized, associated either with point sources or urbanareas with a diversity of diffuse inputs of contaminantsand elevated recreational or food collection activities.On a broader scale, intensive use of the shore for foodcollection can cause impacts at a regional level over100s to 1000s of kilometres (e.g. in Chile, reviewedby Paine, 1994). Eutrophication can also act on abroad scale (e.g. in the Baltic, Bonsdorff et al., 1997).Acute impacts are generally localized and most oilspills rarely cover more than a few 10s of kilometres
of coastline, although the Gulf spills extended to 100sof kilometres.
Clearly some stretches of rocky coastline havebeen chronically impacted for hundreds of years. Forexample, food collection in S. Africa and parts of theMediterranean dates back into pre-history (Siegfried,1994). In more developed countries this impact hasprobably lessened since medieval times. Industrial anddomestic pollution is primarily a product of the last300 yrs or so. Its impact is restricted to urbanizedcoastlines, especially embayed areas. The duration ofthese impacts in most instances is 100 yrs at most. Indeveloping countries industrialization and populationgrowth is accelerating the process (Morton, 1990; DaSilva et al., 1997; Liu & Morton, 1998). In coun-tries which are losing their traditional heavy indus-tries, reversal of degradation is occurring. Treatmentof waste and tighter environmental regulations speedthis process. Many coastal ecosystems are recovering,albeit slowly, and rocky shores in these areas are noexception (e.g. the Mersey and Liverpool Bay region,Hawkins et al. 1999a, b). Ironically, collection ofmussels is prevented in many places by public healthconcerns due to sewage contamination. Thus in someinstances pollution may actually lessen communitylevel disruption.
Disposable income, leisure time and availability ofpersonal transport have all increased dramatically overthe last 50 yrs. In some ways, the decline of subsist-ence gathering of food and fertilizer from the shore hasbeen mirrored by an increase in recreationally-relatedimpacts in the more developed world (Fletcher & Frid,1996) and in areas of the third world which are beingdeveloped for tourism (Hawkins & Roberts, 1992).Paradoxically these impacts can be particularly heavyin reserve areas where public access is encouraged topromote awareness of conservation issues (Fletcher &Frid, 1996; Section 5.7).
Table 3 is a tentative attempt to rank the impor-tance of various anthropogenic disturbances and theirimpacts at the population or community level on rockyshores. We suggest that collection of food or otherresources (e.g. algae for fertilizers, pharmaceuticals)in less developed countries probably has a greaterimpact on shores over very large scales (Section 5.6).Red tides – whether natural or prompted by eutroph-ication, can have similar broadscale effects, but areperiodic and acute (Section 4.2). Of the more chronicpollutants, TBT clearly has a marked effect on alimited suite of species (oysters and stenoglossangastropods) at the individual or population level but

288
community level effects are not widely documented(Section 5.4). Potentially one of the most seriousimpacts on shores, because it is essentially irrevers-ible is the introduction of alien species. These canmarkedly change community processes and ecosystemfunction (Section 5.9).
6.2. Recovery
Assessment of the relative severity of stresses mustconsider the ability of shore communities to recoveronce disturbances cease. As for impacts, recoveryis impossible to assess without reference to naturalspatial and temporal variation (Underwood, 1989).The degree of natural variability differs among shores.On northern European shores, for example, shelteredareas are generally dominated by a community domi-nated by algae which is stable through time. Moder-ately exposed shores, on the other hand, have beenshown to exhibit both small and larger scale fluctu-ations in the relative cover of seaweeds or barnacles(Hawkins et al., 1992; Burrows & Hawkins, 1998 forreviews), making it more difficult to assess recovery.Designing sampling programmes to assess impactsand recovery requires careful consideration of suchnatural levels of variation (Underwood, 1993b, 1994).
Our review suggests that rocky shores do notgenerally suffer persistent residual damage after aninput has been stopped. Pollutants rapidly disperse andthere are few sinks for pollutants, other than the biotathemselves and possibly sediment trapped by algalturfs or animal matrices. Exceptions include baking ofoil to form asphalt-like pavements high on the shore,which can persist for several years (Jones et al., 1998).Diesel oil has also been found trapped in kelp hold-fasts up to 7 yrs after a spill on a sub-Antarctic island(Smith & Simpson, 1998). Peterson (in press) hassuggested that oil spills are more of a press than a pulsedisturbance.
Most shore communities get back to some approx-imation of their original state(s) within 5 yrs orso after minor impacts. This process can be moreprolonged after severe impacts; for example shoresbadly impacted by dispersants following theTorreyCanyonspill probably took 10–15 yrs to recover.
Recovery after physical impacts, such as foodcollection or trampling, can also usually occur rapidlyonce the disturbance stops. Unlike seagrass bedsor coral reefs, rocky shores do not generally havemajor slow-growing biogenic structures, and canusually regenerate quite quickly. This depends on the
species involved, however. Some species of algaeand barnacles returned to experimental plots one yearafter trampling had ceased (Povey & Keough, 1991),whereas mussels had not completely recovered tenyears after trampling ceased (Dye, 1992; Brosnan& Crumrine, 1994).Ascophyllum nodosumcanopiescan also take a very long time to recover followingdisturbance (Jenkins et al., 1999).
Rapid recovery is aided by the open nature of rockyshores with recruitment via larval or algal propagulesfrom remote unimpacted sources (Hawkins et al.,1999a). Whilst this is true for most species, slowerrecovery has been noted in animals with direct devel-opment (high shore littorinids following the Gulf oilspill, Watt et al., 1993; dogwhelks following TBTpollution, Gibbs et al., 1991a). Although rafting canoccur, input from remote sources is much slower.With these species, a case could be made for re-introduction (see Hawkins et al., 1999a) but care mustbe taken that the correct genotype is re-introduced asthese animals have considerable variability generatedby local selection.
6.3. Approach
In contrast to research on soft sediment communities,which often involves the use of multivariate tech-niques to consider the assemblage as a whole (e.g.Gray et al., 1990, Agard et al., 1993; Somerfield etal., 1995), most of the papers reviewed here discussresponses of individual species or functional groups(e.g. dogwhelks, Gibbs et al., 1991; limpets, Tabladoet al., 1994), indicator communities (e.g. kelp hold-fasts, Bamber, 1991) or shifts in dominant suites ofspecies (e.g. assemblage dominated by mussels to onedominated by coralline algae, Dye et al., 1994). Ifsummaries of the entire community are discussed, itis usually in terms of diversity (e.g. Littler & Murray,1975; Bamber, 1991). This flows from the classicalapproach to the study of rocky shores, based on exper-imental manipulation of one or more conspicuousspecies or functional groups with a view to unravellingthe roles of individual species in community processes(e.g. Connell, 1961, 1972, 1974; Paine, 1974, 1977).
This largely univariate approach has the advantagethat it can characterise detailed responses of theselected dominant species, and it retains the focus ondirect links between the disturbance and the response.Although the selected fauna/flora may be the mostnoticeable or tractable, however, they may not alwaysbe the most responsive to any given impact. Responses

289
Table 3. Tentative summary of relative impacts of different sources of stress on rocky shore communitites. To enable comparisonamong the different sources of stress, each has been given a score from 1 to 5 points, based on the literature reviewed
Source of stress Extent of impact Severity of impact Time to recovery Amount of field[local/rare (•) – broadscale/ [minor (•, sublethal) – [short (•, 1-2 yr) – research donecommon (• • • • •)] severe (• • • • •, mass long (• • ••, 10–15 yr) –Developing world Developed world mortality of most species)] permanent (• • • • •)]
Toxic algal blooms ? • • • • • •• spp. dependent ••Oil – acute • •• • • •• • • •• • • •Oil – chronic • • • • •• •• ••Nutrient pollution • •• • • • • ••Heavy metals •? ••? • • •∗ • • •∗ •Antifouling paints •• • • • • • ••# • • ••# • • ••Thermal pollution • • •• spp. dependent •Harvesting • • • • • • • spp. dependent • • •Trampling (recreational) •• • • • • • •• spp. dependent • • •Introduced species ? • • •• • • • • • • • • • • •
? not known;∗ only example available: copper mine tailings (Castilla, 1996);# for dogwhelk populations.
of meiofauna on rocky shores, for example, havereceived very little attention (Gibbons & Griffiths,1986) and may in some cases be more sensitive ata community level than the macrofauna (Warwick,1993). It may be informative to compare the responsesof different sections of the community, or individualindicator species with responses of the community asa whole (see Somerfield et al., 1995). This would serveas ‘ground-truthing’ for existing approaches againstthe broader, multivariate background, and improve ourcapacity for early detection of impacts.
6.4. Way forward
Predicting the likely impacts of disturbances dependson an understanding of the natural dynamics ofthe community (Suchanek, 1994). Although wehave considerable knowledge of community-levelprocesses that operate on exposed rocky shores (e.g.Underwood et al., 1983; Hawkins et al., 1992),particularly at certain well-studied locations, we havelittle understanding of communities on sheltered orestuarine rocky shores and artificial substrata. Thesehabitats are among those most likely to be impactedby human activities due to their proximity to urbanareas and because contaminants are more concentratedin sheltered coastal waters than on exposed coasts.Research on such habitats should therefore be anurgent priority and models of community processes inother rocky habitats should be further refined and theirspatial and temporal generality assessed (see Under-wood & Denley, 1984; Foster et al., 1990; Underwood& Petraitis, 1993).
Research into anthropogenic impacts is needed atthe basic level of identifying patterns of impact bycomparing communities at putatively impacted rockyshores with those at unimpacted controls. Responsesto many sources of stress, e.g. heavy metals, thermalpollution, have simply not been assessed in field-basedstudies or at a community level (see Sections 5.3, 5.5).Experimental research to understand the mechanismsinvolved in impacts is also scarce, even for such high-profile events as oil-spills (see Section 4.1). We shouldaim to be better prepared to extract more thorough,high-quality information from acute events, e.g. oilspills (Section 4.1). Wherever different managementtechniques are applied, the opportunity should betaken to determine how effective they have been,to compare different approaches and refine them forfuture application – ‘adaptive management’ (Walters,1986, 1993). For example, comparisons of differentmethods for dispersing oil can help reduce the damagedone during future clean-up operations (e.g. Watt etal., 1993).
There is a particular need for long term studies.Long term effects of chronic sources of stress areparticularly hard to distinguish from natural changes(Section 3; GESAMP, 1990). Carefully designedmonitoring programs are required to identify suchimpacts and should also be accompanied by experi-mental research to enable identification of causativelinks (GESAMP, 1990). Recovery after acute eventsmay also take several years and can only be studiedproperly over long periods of time (e.g.Torrey Canyonspill, Hawkins & Southward, 1992). Apparent short-term recovery may simply be part of continued period

290
of increased variability or temporary convergence ofpopulations or communities at impacted and controlsites (Underwood, 1989). Such studies should includeproper controls to enable analysis using Beyond BACI(Before After Control Impact) techniques (see Under-wood, 1992, 1994).
6.5. Conclusions
It is clear that the communities of chronicallyimpacted shores can continue to function, albeit ina different state. On such shores, a proliferation ofephemeral, opportunistic species tends to occur anddifferent communities converge towards a commonaltered (degraded) state. Although easily damaged,rocky shore communities are, on the whole, quiteresilient and are able to recover if sources of stressare removed (see Grimm and Wissel (1997) fordetailed definition of resilience and related terms).Communities on most impacted shores recover within10–15 yrs. This is due largely to the fact that themajority of species have a planktonic larval phase andcan recruit to recovering areas from nearby popula-tions. Thus, as long as some healthy populationspersist, for example in marine reserves, managementto allow recovery of impacted shores has a reasonablechance of being effective.
The biggest problem in identifying anthropo-genic impacts on rocky shore communities is theconsiderable spatial and temporal variation of thecommunities. This also means that recovery can bedifficult to define and measure. The ease of non-destructive sampling and relatively easy access does,however, allow the research required to describespatial and temporal variation, understand its causesand to predict and identify community level responsesto anthropogenic stress.
Acknowledgements
The authors would like to thank Enid Scapaticcifor text processing SJH’s early scribblings, SarahAnderson for chasing up references, Nick Smith fordrawing the figures, an anonymous referee for valu-able comments and Martin Attrill for his patienceand input. TPC was funded by NERC FellowshipGR5/98/7/MS, RCT by Leverhulme Grant F180AOand SB by the Faculty of Science, University ofSouthampton. SJH has been funded by NERC formany studies relevant to this paper over the years
and more recently by EU grant MAS3-CT95-0012(EUROROCK).
References
Abuzinada, A. H. & F. Krupp, 1994. The status of coastal andmarine habitats two years after the Gulf War oil spill. Cour.Forschungsinst. Senck. 166: 1–80.
Addessi, L., 1994. Human disturbance and long term changes on arocky intertidal community. Ecol. Appl. 4: 786–797.
Agard, J. B. R., J. Gobin & R. M. Warwick, 1993. Analysis ofmarine macrobenthic community structure in relation to pollu-tion, natural seepage and seasonal disturbance in a tropicalenvironment (Trinidad, West Indies). Mar. Ecol. Prog. Ser. 92:233–243.
Airoldi, L., 1998. Roles of disturbance, sediment stress, andsubstratum retention on spatial dominance in algal turf. Ecology79(8): 2759–2770.
Airoldi, L. & F. Cinelli, 1997. Effect of sedimentation on subtidalmacroalgal assemblages: An experimental study from a Mediter-ranean rocky shore. J. Exp. Mar. Biol. Ecol. 215: 271–290.
Airoldi, L. & M. Virgilio, 1998. Responses of turf-forming algae tospace variations in the deposition of sediments. Mar. Ecol. Prog.Ser. 165: 271–282.
Airoldi, L., F. Rindi & F. Cinelli, 1995. Structure, seasonaldynamics and reproductive phenology of a filamentous turfassemblage on a sediment influenced, rocky subtidal shore. Bot.Mar. 38: 227–237.
Airoldi, L., M. Fabiano & F. Cinelli, 1996. Sediment deposition andmovement over a turf assemblage in a shallow rocky coastal areaof the Ligurian Sea. Mar. Ecol. Prog. Ser. 133: 241–251.
Akberali, H. B. & J. Davenport, 1982. The detection of salinitychanges by the marine bivalve mollusksScrobicularia plana(Dacosta) andMytilus edulis L. J. Exp. Mar. Biol. Ecol. 58:59–71.
Akimichi, T., 1995. Indigenous resource management and sustain-able development: Case studies from Papua New Guinea andIndonesia. Anthropol. Sci. 103: 321–327.
Alzieu, C., 1986. TBT detrimental effects on oyster culture inFrance – evolution since antifouling paint regulation. In: Oceans’86 Proceedings. Volume 4: International Organotin Symposium.Institute of Electrical and Electronic Engineers, New York,pp. 1130–1134.
Alzieu, C., J. Sanjuan, J. P. Deltreil & M. Borel, 1986. Tin contam-ination in Arcachon Bay: effects on oyster shell anomalies. Mar.Pollut. Bull. 17: 494–498.
Anderlini, V. C., 1992. The effect of sewage on trace metal concen-trations and scope for growth inMytilus edulis aoteanusandPerna canaliculusfrom Wellington Harbour, New Zealand. Sci.Total Environ. 125: 263–288.
Bamber R. N., 1991. TheLaminaria holdfast community of WylfaPower Station cooling water discharges, National Power, NuclearElectric Research Report, TEC/L/0321/R90., CMP/L/0001/R91.
barreiro, R., C. Real & A. Carballeira, 1993. Heavy-metal accu-mulation byFucus ceranoidesin a small estuary in north-westSpain. Marine Environ. Res. 36: 39–61.
Beaumont, A. R., 1988. Some ecological consequences of the use ofanti-fouling paints containing tributyltin. Adv. Underwat. Tech.,Ocean Sci. Offshore Eng. 16: 291–298.
Beaumont, A. R. & P. B. Newman, 1986. Low levels of tributyltinreduce growth of marine microalgae. Mar. Pollut. Bull. 17: 457–461.

291
Beaumont, A. R., G. Tserpes & M. D. Budd, 1987. Some effects ofcopper on the veliger larvae of the musselMytilus edulisand thescallopPecten maximus(Mollusca, Bivalvia). Marine Environ.Res. 21: 299–309.
Bellinger, E. G. & B. R. Benham, 1978. The levels of metals indockyard sediments with particular reference to the contributionsfrom ship-bottom paints. Environ. Pollut. 15: 71–81.
Bender, E. A., T. J. Case & M. E. Gilpin, 1984. Perturbation exper-iments in community ecology: Theory and practice. Ecology 65:1–13.
Blackmore, G., B. Morton & Z. G. Huang, 1998. Heavy metalsin Balanus amphitriteandTetrclita squamosa(Crustacea: Cirre-pedia) collected from the coastal waters of Xiamen, China. Mar.Pollut. Bull. 36: 32–40.
Bokn, T., J. A. Berge, N. Green & B. Rygg, 1990. Inva-sion of the planktonic algaeChrysochromulina polylepisalongsouth Norway in May–June 1988. Acute effects on biologicalcommunities along the coast. In: Lancelot, C., G. Billen &H. Barth (eds),Eutrophication and Algal Blooms in North SeaCoastal Zones, the Baltic and Adjacent Areas: Prediction andAssessment of Preventative Actions. Water Pollution ResearchReport 12. Commission of the European Communities, Brussels:183–193.
Bokn, T. L., F. E. Moy & S. N. Murray, 1993. Long term effects ofthe water-accommodated fraction (WAF) of diesel oil on rockyshore populations maintained in experimental mesocosms. Bot.Mar. 36: 313–319.
Bonsdorff, E., 1983. Effects of experimental oil exposure on thefauna associated withCorallina officinalis in intertidal rockpools. Sarsia 68: 149–155.
Bonsdorff, E. & W. G. Nelson, 1981. Fate and effects of Ekofiskcrude oil in the littoral of a Norwegianfjord. Sarsia 66(3): 231–240.
Bonsdorff, E., E. M. Blomqvist, J. Mattila & A. Norkko, 1997.Coastal eutrophication: Causes, consequences and perspectivesin the archipelago areas of the northern Baltic Sea. Est. CoastalShelf Sci. 44: 63–72.
Borowitzka, M. A., 1972. Intertidal algal species diversity and theeffect of pollution. Aust. J. Mar. Freshwat. Res. 23: 73–84.
Bosman, A. L. & P. A. R. Hockey, 1988. The influence of seabirdguano on the biological structure of rocky intertidal communitieson islands off the west coast of Southern Africa. S. Afr. J. Mar.Sci. 7: 61–68.
Bowman, R. S.,1978. Dounreay oil spill: Major implications of aminor incident. Mar. Pollut. Bull. 9: 269–273.
Bowman, R. S. & J. R. Lewis, 1977. Annual fluctuations in therecruitment ofPatella vulgataL. J. Mar. Biol. Assoc. (U. K.)57: 793–815.
Branch, G. M. & C. A. Moreno, 1994. Intertidal and subtidalgrazers. In: Siegfried, R. S. (ed.),Rocky Shores: Exploitationin Chile and South Africa. Springer-Verlag, Berlin.
Bright, D. A. & D. V. Ellis, 1990. A comparative survey ofimposex in NE Pacific neogastropods (Prosobranchia) related totributyltin contamination and choice of a suitable bioindicator.Can. J. Zool. 68: 1915–1924.
Brosnan, D. B. & L. L. Crumrine, 1994. Effects of human tramplingon marine rocky shore communities. J. Exp. Mar. Biol. Ecol. 177:79–97.
Bruton, M. N. & R. E. Stobbs, 1991. The ecology and conservationof the coelocanthLatimeria chaluumnae. Env. Biol. Fish. 32:313–319.
Bryan, G. W. 1984. Pollution due to heavy metals and theircompounds. In: Kinne, O. (ed.),Marine Ecology. John Wileyand Sons, Chichester: 1289–1431.
Bryan, G. W. & P. E. Gibbs, 1991. Impacts of low concentrations oftributyltin (TBT) on marine organisms: A review. In: Newman,M. C. & A. W. McIntosh (eds),Metal Ecotoxicology: Conceptsand Applications. Lewis Publishers, Boston: 323–361.
Bryan, G. W., W. J. Langston, L. G. Hummerstone & G. R. Burt,1985. A guide to the assessment of heavy metal contamination inestuaries using biological indicators. J. Mar. Biol. Assoc. (U.K.)occasional publication 4: 1–92.
Bryan, G. W., P. E. Gibbs, L. G. Hummerstone & G. R. Burt, 1986.The decline of the gastropodNucella Lapillusaround south-westEngland: Evidence for the effect of tributyltin from antifoulingpaints. J. Mar. Biol. Assoc. (U.K.) 66: 611–640.
Bryan, G. W., P. E. Gibbs, G. R. Burt & L. G. Hummerstone,1987. The effects of tributyltin (TBT) accumulation on adultdogwhelks Nucella lapillus: Long-term field and laboratorystudies. J. Mar. Biol. Assoc. UK. 67: 525–544.
Bryan, G. W., P. E. Gibbs & G. R. Burt, 1988. A comparison ofthe effectiveness of tri-n-butyltin chloride and five other organ-otin compounds in promoting the development of imposex inthe dogwhelk,Nucella Lapillus. J. Mar. Biol. Assoc. (U.K.) 68:733–744.
Burrows, M. T. & S. J. Hawkins, 1998. Modelling patch dynamicson rocky shores using deterministic cellular automata. Mar. Ecol.Prog. Ser. 167: 1–13.
Bustamante, R. H., G. M. Branch, S. Eekhout, B. Robertson,P. Zoutendyk, M. Schleyer, A. Dye, N. Hanekom, D. Keats,M. Jurd & C. McQuaid, 1995. Gradients of intertidal primaryproductivity around the coast of South Africa and relationshipswith consumer biomass. Oecologia 102: 189–201.
Caffey, H. M., 1985. Spatial and temporal variation in settlementand recruitment of intertidal barnacles. Ecol. Monogr. 55: 313–332.
Carlson, L. H., & P. J. Godfrey, 1989. Human impact managementin coastal recreation and natural area. Biol. Conserv. 49: 141–156.
Carlton, J. T., 1982. The historical biogeography ofLittorina littoreaon the Atlantic coast of North America and implications forthe interpretation of ther structure of New England intertidalcommunities. Malacol. Rev. 15: 146.
Carlton J. D., 1989. The introduced marine and estuarine mollusksof North America. An end of century perspective on fourcenturies of human mediated introductions. J. Shellfish Res. 8:465.
Castilla, J. C., 1996. Copper mine tailing disposal in northernChile rocky shores:Enteromorpha compressa(Chlorophyta) asa sentinel species. Environ. Monit. Assess. 40: 171–184.
Castilla, J. C., 1999. Costal marine communities: trends andperspectives from human-exclusion experiments. TREE 14:280–283.
Castilla, J. C., & R. H. Bustamante, 1989. Human exclusion fromrocky intertidal of Las-Cruces, Central Chile – effects onDurvil-laea antarctica(Phaeophyta, Durvilleales). Mar. Ecol. Prog. Ser.50: 203–214.
Castilla, J. C. & L. R. Duran, 1985. Human exclusion from therocky intertidal zone of Central Chile the effects onConcholepasconcholepas(Gastropoda). Oikos 45: 391–399.
Chagot, D., J. Alzieu, J. Sanjuan & H. Grizel, 1990. Sublethal andhistopathological effects of trace levels of tributyltin fluoride onadult oystersCrassostrea gigas. Aquat. Liv. Resour. 3: 121–130.
Clark, R. B., C. Frid & M. Attrill, 1992.Marine Pollution, 4th edn.Clarendon Press, Oxford.
Coates, M., 1998. A comparison of intertidal assemblages onexposed and sheltered tropical and temperate rocky shores. Glob.Ecol. Biogeog. Letters 7: 115–124.

292
Conan, G., 1982. The long-term effects of theAmoco Cadizoil spill.Phil. Trans. R. Soc. Lond. B. 297: 323–333.
Connell, J. H., 1961. The influence of inter-specific competitionand other factors on the distribution of the barnacleCthamalusstellatus. Ecology 42: 710–723.
Connell, J. H., 1972. Community interactions on marine rockyintertidal shores. Ann. Rev. Ecol. Syst. 31: 196–192.
Connell, J. H., 1974. Ecology: field experiments in marineecology. In: Mariscal, R. N. (ed.),Experimental Marine Biology.Academic Press, New York: 131–138.
Corner, A. D. S., A. J. Southward & E. C. Southward, 1968. Toxicityof oil–spill removers (“detergents”) to marine life: An assess-ment using the intertidal barnacleElminius modestus. J. Mar.Biol. Assoc. (U.K.) 48: 29–47.
Correa, J. A., J. C. Castilla, M. Ramirez, M. Varas, N. Lagos, S.Vergara, A. Moenne, D. Roman & M. T. Brown, 1999. Copper,copper mine tailings and their effect on marine algae in northernChile. J. Appl. Phycol. 11: 57–67.
Crapp, G. B., 1971. Chronic oil pollution. In: Cowell, E. B. (ed.),The Ecological Effects of Oil Pollution on Littoral Communities.Institute of Petroleum, London: 187–203.
Crisp D. J., 1964. The effects of the severe winter of 1962–1963 onmarine life in Britain. J. Anim. Ecol. 33: 165–210.
Critchley, A. T., W. F. Farnham, T.Yoshida & T. A. Norton, 1990.A bibliography of the invasive algaSargassum muticum(Yendo)Fensholt (Fucales, Sargassaceae). Bot. Mar. 33: 5551–562.
Crothers, J. H., 1983. Field experiments on the effects of crudeoil and dispersant on the common animals and plants of rockyseashores. Marine Environ. Res. 8: 215–239.
Cullinane, J. P., P. McCarthy & A. Fletcher, 1975. The effect of oilpollution in Bantry Bay. Mar. Pollut. Bull. 6: 173–176.
Da Silva, E. M., M. C. Peso-Aguiar, M. d. F. T. Navarro & C.de Barros e Azevedo Chastinet, 1997. Impact of petroleum onaquatic coastal ecosystems in Brazil. Environ. Toxicol. & Chem.16: 112–118.
Davis A. R., 1995. Over-exploitation ofPyura chilensis(Ascidi-acea) in Southern Chile – the urgent need to establish marinereserves. Rev. Chil. Hist. Nat. 68: 107–116.
Davies, I. M. & S. K. Bailey, 1991. The impact of tributyltin fromlarge vessels on the dogwhelk (Nucella lapillus) populationsaround Scottish oil ports. Marine Environ. Res. 32: 201–211.
Davies, I. M., J. C. McKie & J. D. Paul, 1986. Accumulationof tin and tributyltin from antifouling paint by cultivated scal-lops (Pecten maximus) and Pacific oysters (Crassostrea gigas).Aquaculture 55: 103–144.
Davison, I. R. & G. A. Pearson, 1996. Stress tolerance in intertidalseaweeds. J. Phycol. 32: 197–211.
De Vogelaere, A. P. & M. S. Foster, 1994. Damage and recoveryin intertidal Fucus gardneriassemblages following the “ExxonValdez” oil spill. Mar. Ecol. Prog. Ser. 106: 263–271.
Duran, L. R., & J. C. Castilla, 1989. Variation and persistence ofthe middle rocky intertidal community of central Chile, with andwithout human harvesting. Mar. Biol. 103: 555–562.
Diaz, R. J. & R. Rosenberg, 1995. Marine benthic hypoxia: Areview of its ecological effects and the behavioural responses ofbenthic macrofauna. Oceanogr. Mar. Biol. 33: 245–303.
Dye, A. H., 1992. Experimental studies of succession and stabilityin rocky intertidal communities subject to artisanal shellfishgathering. Neth. J. Sea Res. 30: 209–217.
Dye, A. H., G. M. Branch, J. C. Castilla & B. A. Bennett, 1994a.Biological options for the management of the exploitation ofintertidal and subtidal resources In: Siegfried, R. S. (ed.),RockyShores: Exploitation in Chile and South Africa. Springer-Verlag,Berlin.
Dye, A. H., G. L. Schleyer & T. A. Lasiak, 1994b. Intertidal andsubtidal filter feeders in southern Africa. In: Siegfried, R.S. (ed.),Rocky Shores: Exploitation in Chile and South Africa. Springer-Verlag, Berlin: 57–73.
Dyrynda, E. A., R. J. Law, P. E. J. Dyrynda, C. A. Kelly, R.K. Pipe, K. L. Graham & N. A. Ratcliffe, 1997 Modulationsin cell-mediated immunity ofMytilus edulisfollowing the ‘SeaEmpress’ oil spill. J. Mar. Biol. Assoc. (U.K.) 77: 281–284.
Ellis, D. V. & L. A. Pattisina, 1990. Widespread neogastropodimposex: A biological indicator of global TBT pollution. Mar.Pollut. Bull. 21: 248–253.
Estes, J. A. & J. F. Palmisano, 1974. Sea otters: Their role instructuring nearshore communities. Science 185: 1058–1060.
Estes, J. A. & D. O. Duggins, 1995. Sea otters and kelp forestsin Alaska: Generality and variation in a community ecologicalparadigm. Ecol. Monogr. 65: 75–100.
Evans, S. M., A. Hutton, M. A. Kendall & A. M. Samosir, 1991.Recovery in populations of dogwhelksNucella lapillus (L.)suffering from imposex. Mar. Pollut. Bull. 22: 331–333.
Evans, S. M., G. J. Nicholson, C. Browning, E. Hardman, O.Seligman & R. Smith, 1998. An assessment of tributyltincontamination in the North Atlantic using imposa in thedogwhelkNucella lapillus(L.) as a biological indicator of TBTpollution. Invertebr. Reprod. Dev. 34: 277–287.
Evans, S. M., A. C. Birchenough & M. S. Brancato, 2000a. TheTBT Ban: Out of the frying pan into the fire? Mar. Pollut. Bull.40: 204–211.
Evans, S. M., E. Kerrigan & N. Palmer, 2000b. Causes of imposexin the dogwhelkNucella lapillus(L.) and its use as a biologicalindicator of tributyltin contamination. Mar. Pollut. Bull. 40: 212–219.
Fairweather, P. G., 1990. Sewage and the biota on seashores:Assessment of impact in relation to natural variability. Environ.Monit. Assess. 14: 197–210.
Fichet, D., G. Radenac & P. Miramand, 1998. Experimental studiesof impacts of harbour sediments resuspension to marine inverte-brates larvae: Bioavailability of Cd, Cu, Pb and Zn and toxicity.Mar. Pollut. Bull. 36: 7–12.
Fioroni, P., J. Oehlmann & E. Stroben, 1991. The pseudoherm-aphroditism of prosobranchs; morphological aspects. Zool. Anz.226: 1–26.
Fletcher, H. & C. L. J. Frid, 1996. Impact and management of visitorpressure on rocky intertidal algal communities. Aquatic Conserv:Mar. Freshw. Ecosyst. 6: 287–297.
Foster, M. S., 1990. Organization of macroalgal assemblages inthe Northeast Pacific: The assumption of homogeneity and theillusion of generality. Hydrobiol 192: 21–33.
Foster, M. S., A. P. DeVogelaere, C. Harrold, J. S. Pearse & A. B.Thum, 1988. Causes of spatial and temporal patterns in rockyintertidal communities in central and northern California. Mem.Calif. Acad. Sci. 9: 1–45.
Foster, M. S., J. A. Tarpley & S. L. Dearn, 1990. To clean or not toclean: the rationale, methods, and consequences of removing oilfrom temperate shores. Northwest Environ. J. 6: 105–120.
Fuji, A. & H. Nomura, 1990. Community structure of the rockyshore macrobenthos in southern Hokkaido, Japan. Mar. Biol.107: 471–477.
Gaines, S., S. Brown & J. Roughgarden, 1985. Spatial variationin larval concentrations as a cause of spatial variation in settle-ment for the barnacleBalanus glandula. Oecologia 67: 267–272.
GESAMP, 1990.The State of the Marine Environment. BlackwellScientific Publications, Oxford.

293
Gibbons, M. J. & C.L. Griffiths, 1986. A comparison of macro-faunal and meiofaunal standing stock across a rocky shore, withan estimate of their productivities. Mar. Biol. 93: 181–188.
Gibbs P. E., 1993. A male genital defect in the dogwhelk,Nucellalapillus (Neogastropoda) favoring survival in a TBT pollutedarea. J. Mar. Biol. Assoc. (U.K.) 72: 667–678.
Gibbs, P. E. & G. W. Bryan, 1987. TBT paints and the demiseof the dogwhelk, Nucella lapillus (gastropoda). In: Oceans’87 proceedings. Volume 4: International organotin symposium.Institute of Electrical and Electronic Engineers, New York:1482–1487.
Gibbs, P. E., G. W. Bryan, P. L. Pascoe & G. R. Burt, 1987. Theuse of the dogwhelkNucella lapillus as an indicator of TBTcontamination. J. Mar. Biol. Assoc. (U.K.) 67: 507–523.
Gibbs, P. E., P. L. Pascoe & G. R. Burt, 1988. Sex change in thefemale dogwhelkNucella lapillus, induced by TBT from anti-fouling paints. J. Mar. Biol. Assoc. (U.K.) 68: 715–732.
Gibbs, P. E., G. W. Bryan & P. L. Pascoe, 1991a. TBT-induced imposex in the dogwhelk,Nucella lapillusgeographicaluniformity of the response and effects. Marine Environ. Res. 32:79–87.
Gibbs, P. E., P. L. Pascoe & G. W. Bryan, 1991b. Tributyltin inducedimposex in stenoglossan gastropods: Pathological effects on thefemale reproductive system. Comp. Biochem. Physiol. 100C:231–235.
Gibbs, P. E., G. W. Bryan & S. K. Spence, 1991c. The impact oftributyltin (TBT) pollution on theNucella lapillus(Gastropoda)populations around the coast of south-east England. Ocea. Acta,Proceedings of the International Colloquium on the Environ-ment of Epicontinental Seas, Lille, 20–22 March, 1990; Specialvolume 11, pp. 257–261.
Gilfillan, E. S., D. S. Page, E. J. Harner & P. D. Boehm,1995. Shoreline ecology program for Prince William Sound,Alaska, following the Exxon Valdez oil spill. Part 3. Biology.Philadelphia PA: Amer. Soc. for Testing and Materials: 398–443.
Glasby, T. M. & A. J. Underwood, 1996. Sampling to differentiatebetween pulse and press perturbations. Environ. Monit. Assess.42: 241–252.
Glegg, G. A., L. Hickman & S. J. Rowland, 1999 Contaminationof limpets (Patella vulgata) following the Sea Empress oil spill.Mar. Pollut. Bull. 38(2): 119–225.
Gray, J. S., K. R. Clarke, R. M. Warwick & G. Hobbs, 1990.Detection of initial effects of pollution on marine benthos: Anexample from the Ekofisk and Eldfisk oilfields, North Sea. Mar.Ecol. Prog. Ser. 66: 285–299.
Griffiths, C. L. & G. M. Branch, 1997. The exploitation of coastalinvertebrates and seaweeds in South Africa: Historical trends,ecological impacts and implications for management. Tran. R.Soc. S. Afr. 52: 121–148.
Grimm, V. & C. Wissel, 1997. Babel, or the ecological stabilitydiscussions: An inventory and analysis of terminology and aguide for avoiding confusion. Oecologia 109: 323–334.
Hallegraeff, G. M., 1993. A review of harmful algal blooms andtheir apparent global increase. Phycologia 32: 79–99.
Harris, J. M., G. M. Branch, B. L. Elliott, B. Currie, A. H. Dye, C.D. McQuaid, B. J. Tomalin & C. Velasquez, 1998. Spatial andtemporal variability in recruitment of intertidal mussels aroundthe coast of southern Africa. S. Afr. J. Zool. 33: 1–11.
Hartnoll, R. G. & S. J. Hawkins, 1985. Patchiness and fluctuationson moderately exposed rocky shores. Ophelia 24: 53–63.
Haven, S. B., 1971. Effects of land-level changes on intertidal inver-tebrates, with discussion of postearthquake ecological sucession.The Great Alaska Earthquake of 1964, Washington DC: Natl.Res. Coun: 82–126.
Hawkins, J. P. & C. M. Roberts, 1992. Effects of recreationalSCUBA diving on fore-reef slope communities of coral reefs.Biol. Conserv. 62: 171–178.
Hawkins, S. J., 1999. Experimental ecology and coastal conserva-tion: Conflicts on rocky shores. Aquatic Conserv: Mar. Freshw.Ecosyst. 9: 565–572.
Hawkins, S. J. & R. G. Hartnoll, 1983. Grazing of intertidal algae bymarine invertebrates. Oceanogr. Mar. Biol. Ann. Rev., 21: 195–285.
Hawkins, S. J., A. J. Southward & R. L. Barrett, 1983. Populationstructure ofPatella vulgataduring sucession on rocky shoresin south-west England. In: Oceanologica Acta, Proc. 17th Eur.Mar. Biol. Symp., Brest, France, 27 September–1 October 1982,Special Volume, pp. 103–107.
Hawkins, S. J. & A. J. Southward, 1992. The Torrey Canyon oilspill: Recovery of rocky shore communities. In: Thayer, G. W.(ed.),Restoring the Nation’s Environment. Maryland Sea GrandColl., Maryland: 583–631.
Hawkins, S. J., R. G. Hartnoll, J. M. Kain & T. A. Norton, 1992.Plant-animal interactions on hard substrata in the north-eastAtlantic. In: John, D. M., S. J. Hawkins & J. H. Price (eds),Plant–Animal Interactions in the Marine Benthos. ClarendonPress, Oxford: 1–32.
Hawkins, S. J., S. V. Proud, S. K. Spence & A. J. Southward,1994. From the individual to the community and beyond: Waterquality, stress indicators and key species in coastal ecosystems.In: Sutcliffe, D. W. (ed.), Water Quality and Stress Indi-cators in Marine and Freshwater Ecosystems: Linking Levelsof Organisation (Individuals, Populations, Communities). Fresh-water Biological Association Publishers, Ambleside: 35–62.
Hawkins, S. J., J. R. Allen & S. Bray, 1999a. Restoration oftemperate marine and coastal ecosystems: Nudging nature.Aquatic Conserv: Mar. Freshw. Ecosyst. 9: 23–46.
Hawkins, S. J., J. R. Allen, N. J. Fielding S. B. Wilkinson & I.D. Wallace, 1999b. Liverpool Bay and the estuaries: humanimpact, recent recovery and restoration. In: Greenwood, E. F.(ed.), Ecology and Landscape Development: A History of theMersey Basin. Conf. Proc: 155–165.
Haynes, D., J. Leeder & P. R. Rayment, 1997. A comparison of thebivalve speciesDonax deltoidesandMytilus edulisas monitorsof metal exposure from effluent discharges along the Ninety MileBeach, Victoria, Australia. Mar. Pollut. Bull. 34: 326–311.
Highsmith, R., M. S. Stekoll, P. G. van Tamelen, A. J. Hooten, S.M. Saupe, L. Deysher et al, 1995. Herring Bay monitoring andrestoration studies. Monitoring and restoration project no. 93039.Inst. Mar. Sci., Univ. Alaska, Fairbanks.
Highsmith, R., T. L. Rutger, M. S. Stekoll, S. M. Saupe, M. R.Lindeberg, R. N. Jenne & W. P. Erickson, 1996. Impact of theExxon Valdezoil spill on intertidal biota. In: S. D. Rice, R. B.Spies, D. A. Wolfe & B. A. Wright (eds),Exxon Valdez OilSpill Symposium Proceedings. Am. Fish. Soc. Symp. 18: 212–237.
Hiscock, K., 1985.Rocky Shore Survey and Monitoring Workshop.May 1–4, 1984. British Petroleum International Ltd., London.
Hoare, K., A. R. Beaumont & J. Davenport, 1995. Variationamong populations in the resistance ofMytilus edulisembryosto copper: Adaptation to pollution? Mar. Ecol. Prog. Ser. 120:155–161.
Hockey, P. A. R., 1987. The influence of coastal utilization by manon the presumed extinction of the Canarian black oystercatcherHaematopus meadewaldoiBannerman. Biol. Conserv. 39: 49–62.
Hockey, P. A. R., 1994. Man as a component of the littoral predatorspectrum: A conceptual overview. In: Siegfried, R. S. (ed),Rocky

294
Shores: Exploitation in Chile and South Africa. Springer-Verlag,Berlin: 17–26.
Hockey, P. A. R. & A. L. Bosman, 1986. Man as an intertidalpredator in Transkei: disturbance, community convergence andmanagement of a natural food resource. Oikos 46: 3–14.
Houghton, J. P., A. K. Fukuyama, W. B. Driskell, D. C. Lees, G.Shigenaka & A. J. Mearns, 1993. Recovery of Prince WilliamSound intertidal epibiota fromExxon Valdezspill and treatments,1990–1992.Exxon Valdezoil spill symposium abstract book.Anchorage, AK: February 2–5, pp. 79–82.
Houghton, J. P., A. K. Fukuyama, D. C. Lees, P. J. Hague, H.L. Cumberland, P. M. Harper et al., 1993. Evaluation of thecondition of the Prince William Sound shorelines following theExxon Valdezoil spill and subsequent shoreline treatment. Vol.2. 1992 Biological monitoring survey. NOAA technical memor-andum NOS ORCA 73. Natl. Oceanic and Atmos. Admin., USDept. Commerce.
Houghton, J. P., D. C. Lees, W. B. Driskell, S. C. Lindstrom &A. J. Mearns, 1996. Recovery of Prince William Sound intertidalepibiota fromExxon Valdezoiling and shoreline treatments, 1989through 1992. In: Rice, S. D., R. B. Spies, D. A. Wolfe & B. A.Wright (eds),Exxon Valdez Oil Spill Symposium Proceedings,Vol 18. Amer. Fish. Soc. Symp.: 379–411
Hurlbert, S. H., 1997. Functional importance vs keystoneness: refor-mulating some questions in theoretical biocenology. Aust. J.Ecol. 22: 369–382.
Jenkins, S. R., T. A. Norton & S. J. Hawkins, 1999. Direct andindirect effects of a macroalgal canopy and limpet grazing instructuring a sheltered intertidal community. Mar. Ecol. Prog.Ser. 188: 81–92.
Johansen, K. & F. Møhlenberg, 1987. Impairment of egg productionin Acartia tonsaexposed to tributyltin oxide. Ophelia. 27: 137–141.
Jones, D. A. & M. D. Richmond, 1992. Intertidal and subtidalmarine habitat surveys. Establishment of a marine habitat andwildlife sanctuary for the Gulf region. Final report for Phase 1.CEC/NCWCD, Jubail, Frankfurt. Available from SenckenbergResearch Institute, Frankfurt, Germany, pp. 134–160.
Jones, D. A., I. Watt, T. D. Woodhouse & M. D. Richmond, 1994.Intertidal rcovery in the Dawhat ad Dafi and Dawhat al Mussal-lamiya region (Saudi Arabia) after the Gulf War oil spill. Cour.Forschungsinst. Senck. 166: 27–33.
Jones, D. A., I. Watt, J. Plaza & T. D. Woodhouse, 1996. Naturalrecovery of the intertidal biota within the proposed marinehabitat and wildlife sanctuary for the Gulf (Saudi Arabia) afterthe 1991 Gulf War oil spill. In: Krupp, F., A. H. Abuzinada &I. A. Nader (eds),A Marine Wildlife Sanctuary for the ArabianGulf. Environmental Research and Conservation Following the1991 Gulf War Oil Spill. NCWCD, Riyadh and SenckenbergResearch Institute, Frankfurt, Germany: 138–158.
Jones, D. A., J. Plaza, I. Watt & M. A. Sanei, 1998. Long-term(1991–1995) monitoring of the intertidal biota of Saudi Arabiaafter the 1991 Gulf War oil spill. Mar. Pollut. Bull. 36(6): 472–489.
Jonsson, B. & A. Flemin, 1993. Enhancement of wild salmonpopulations. In: Sundnes, G. (ed.),Human Impact on Self-Recruiting Populations. Tapir, Trondheim: 209–242.
Karsten, U., S. Koch, J. A. West & G. O. Kirst, 1996.Physiological responses of the eulittoral macroalgaStictosi-phonia hookeri(Rhodomelaceae, Rhodophyta) from Argentinaand Chile: Salinity, light and temperature acclimation. Europ. J.Phycol. 31: 361–368.
Keough, M. J. & G. P. Quinn, 1991. Causality and the choice ofmeasurements for detecting human impacts in marine environ-ments. Aust. J. Mar. and Freshwat. Res. 42: 539–554.
Keough, M. J. & G. P. Quinn, 1998. Effects of periodic disturb-ances from trampling on rocky intertidal algal beds. Ecol. Appl.8: 141–161.
Keough, M. J., G. P. Quinn & A. King, 1993. Correlations betweenhuman collecting and intertidal mollusk populations on rockyshores. Conserv. Biol. 7: 378–390.
Kingsford, M. J., A. J. Underwood & S. J. Kennelly, 1991. Humansas predators on rocky reefs in New South Wales, Australia. Mar.Ecol. Prog. Ser. 72: 1–2.
Kyle, R., B. Pearson, P. J. Fielding, W. D. Robertson & S. L. Birnie,1997. Subsistence shellfish harvesting in the Maputaland MarineReserve in northern KwaZulu-Natal, South Africa: Rocky shoreorganisms. Biol. Conserv. 82: 183–192.
Langford, T. E., 1990.Ecological Effects of Thermal Discharges.Elsevier Science, Barking, Essex, England.
Langston, W. J., G. W. Bryan, G. R. Burt & P. E. Gibbs, 1990.Assessing the impact of tin and TBT in estuaries and coastalregions. Funct. Ecol. 4: 433–443.
Lasiak, T., 1998. Multivariate comparisons of rocky infratidalmacrofaunal assemblages from replicate exploited and non-exploited localities on the Transkei coast of South Africa. Mar.Ecol. Prog. Ser. 167: 15–23.
Lasiak, T., 1999. The putative impact of exploitation on rockyinfratidal macrofaunal assemblages: A multiple area comparison.J. Mar. Biol. Assoc. (U.K.) 79: 23–34.
Lasiak, T. A. & J. G. Field, 1995. Community-level attributesof exploited and non-exploited rocky infratidal macrofaunalassemblages in Transkei. J. Exp. Mar. Biol. Ecol. 185: 33–53.
Lewis, J. R., 1964.The Ecology of Rocky Shores. English Univer-sities Press, London.
Lewis, J. R., 1976. Long-term ecological surveillance: Practicalrealities in the rocky littoral. Oceanogr. Mar. Biol. Ann. Rev. 14:505–515.
Lewis, J. R., 1977. The role of physical and biological factors inthe distribution and stability of rocky shore communities. In:Keegan, B. F., P. O. Ceidigh & P. J. S. Boaden (eds),Biologyof Benthic Organisms. Pergamon Press, Oxford: 417–424.
Lewis, J. R., 1982. The composition and functioning of benthicecosystems in relation to the assessment of long-term effects ofoil pollution. Phil. Trans. R. Soc. Lond. B. 297: 257–267.
Liddle, M. J. & A. M. Kay, 1987. Resistance, survival and recoveryof trampled corals on the Great Barrier Reef. Biol. Conserv. 42:1–18.
Lindberg, D. R., J. A. Estes & K. I. Warheit, 1998. Human influ-ences on trophic cascades along rocky shores. Ecol. Appl. 8:880–890.
Littler, M. M. & S. N. Murray, 1975. Impact of sewage on the distri-bution, abundance and community structure of rocky intertidalmacro organisms. Mar. Biol. 30: 277–291.
Liu, J. H. & B. Morton, 1998. The impacts of pollution on thegrowth, reproduction and population structure of Hong Konglimpets. Mar. Pollut. Bull. 36: 152–158.
Lopes, C. F., J. C. C. Milanelli, V. A. Prosperi, E. Zanardi & A.C. Truzzi, 1997. Coastal monitoring program of Sao SebastiaoChannel: assessing the effects of “Tebar V” oil spill on rockyshore populations. Mar. Pollut. Bull. 34(11): 923–927.
López-Gappa, J. J., A. Tablado & N. H. Magaldi, 1990. Influenceof sewage pollution on a rocky intertidal community dominated

295
by the mytilidBrachidontes rodriguezi. Mar. Ecol. Prog. Ser. 63:163–175.
Lundebye, A.-K., T. M. Curtis, C. J. Braven & M. H. Depledge,1997. Effects of the organophosphorous pesticide, dimethoate,on cardiac and acetylcholinesterase (AChE) activity in the shorecrabCarcinus maenas. Aquat. Toxicol. 40: 23–36.
Mance, G., 1987.Pollution Threat of Heavy Metals in AquaticEnvironments. Elsevier Applied Science, London: 372.
McClanahan, T. R. & B. Kaunda Arara, 1996. Fishery recovery ina coral-reef marine park and its effect on the adjacent fishery.Conserv. Biol. 10: 1187–1199.
McClanahan, T. R. & D. Obura, 1997. Sedimentation effects onshallow coral communities in Kenya. J. Exp. Mar. Biol. Ecol.209: 103–122.
Mearns, A. J., 1993. Recovery of shoreline ecosystems followingthe Exxon Valdez oil spill and subsequent treatment. CoastalZone ’93: Proc. 8th Symp. On Coastal and Ocean Manage. NewOrleans LA, pp. 466–479.
Menge, B. A., B. A. Daley, P. A. Wheeler, E. Dahlhoff, E. Sanford& P. T. Strub, 1997. Benthic-pelagic links and rocky intertidalcommunities: Bottom-up effects on top-down control? Proc. Nat.Acad. Sci. 94: 14530–14535.
Meyers, A. A., T. Southgate & T. F. Cross, 1980. Distinguishing theeffects of oil pollution from natural cyclical phenomena on thebiota of Bantry Bay, Ireland. Mar. Pollut. Bull. 11: 204–207.
Mills, L. S., M. E. Soulé & D. F. Doak, 1993. The keystone-speciesconcept in ecology and conservation. Bioscience 43: 219–234.
Moore, J. J., 1998.Sea Empressoil spill: Impacts on rocky andsedimentary shores. In: Edwards, R. & H. Sime (eds),The SeaEmpress Oil Spill: Proceedings of the International Conference,Cardiff, Feb. Terence Dalton: 173–187.
Morton, B., 1990. Pollution and the sub-tropical inshore hydro-graphic environment of Hong Kong. In: Morton, B. (ed.),TheMarine Fauna and Flora of Hong Kong and Southern China II.Hong Kong University Press, Hong Kong: 3–30.
Munda, I. M. & V. Hudnik, 1986. Growth response ofFucus vesicu-losusto heavy metals, singly and in dual combinations, as relatedto accumulation. Bot. Mar. 29: 410–412.
Nagelkerken, I. A. & A. O. Debrot, 1995. Mollusc communities oftropical rubble shores of Curaçao: Long-term (7+ years) impactsof oil pollution. Mar. Pollut. Bull. 30: 592–598.
Naylor, E., 1965a. Biological effects of heated effluents in docks atSwansea S. Wales. Proc. Zool. Lond. 144: 253–268.
Naylor, E., 1965b. Effects of heated effluents on marine andestuarine organisms. Adv. Mar. Biol 3: 63–103.
Nelson, W. G., 1981. Inhibition of barnacle settlement by Ekofiskcrude oil. Mar. Ecol. Prog. Ser. 5: 41–43.
Nelson, W. G., 1982. Experimental studies of oil pollution on therocky intertidal community of a Norwegian Fjord. J. Exp. Mar.Biol. Ecol. 65: 121–138.
Newell, R. C., 1979.Biology of Intertidal Animals. Marine Ecolog-ical Surveys, Faversham.
Newey, S. & R. Seed, 1995. The effects of theBraer oil spill onrocky intertidal communities in South Shetland, Scotland. Mar.Pollut. Bull. 30(4): 274–280.
Newton, L. C., E. V. H. Parkes & R. C. Thompson, 1993. The effectsof shell collecting on the abundance of gastropods on Tanzanianshores. Biol. Conserv. 63: 241–245.
Ocean Studies Board, 1995.Understanding Marine Biodiversitya Research Agenda for the Nation. National Academy Press,Washington.
Ochoa Lopez, E., H. Reyes Bonilla & J. Ketchum Mejia, 1998.Effects of sedimentation on coral communities of southern
Socorro Island, Revellagigedo Archipelago, Mexico. CienciasMarinas 2: 233–240.
Odendaal, F. J., M. O. Bergh & G. M. Branch, 1994. Socio-economic options for the management of the exploitation ofintertidal and subtidal resources In: Siegfried, R. S. (ed.),RockyShores: Exploitation in Chile and South Africa. Springer-Verlag,Berlin.
Oliva, D. & J. C. Castilla, 1986. The effect of human exclusionon the population structure of key-hole limpetsFissurella crassaand Fissurella-limbataon the coast of central Chile. MarineEcology-Pubbl. Stn. Zool. Napoli. I. 7: 201–217.
Paine, R. T., 1974. Intertidal community structure. Experimentalstudies on the relationship between a dominant competitor andits principal predator. Oecologia 15: 93–120.
Paine, R. T., 1977. Controlled manipulations in the marine intertidalzone, and their contributions to ecological theory. In: Goulden,C. E. (ed.),Changing Scenes in Natural Sciences, 1776–1976.Academy of Natural Sciences, Philadelphia: 245–270.
Paine, R. T., 1994.Marine Rocky Shores and Community Ecology:An Experimentalist’s Perspective. Ecology Institute, Oldendorf/Luhe.
Paine, R. T., J. L. Ruesink, A. Sun, E. L. Soulanille, M. J. Wonham,C. D. G. Harley, D. R. Brumbaugh & D. L. Secord, 1996. Troubleon oiled waters: Lessons from theExxon Valdezoil spill. Ann.Rev. Ecol. Syst. 27: 197–235.
Parker, E. D., V. E. Forbes, S. L. Nielsen, C. Ritter, C. Barata, D.J. Baird, W. Admiraal, L. Levin, V. Loeschke, P. Lyytikäinen-Saarenmaa, H. Høgh-Jensen, P. Calow & B. J. Ripley, 1999.Stress in ecological systems. Oikos 86: 179–184.
Patarnello, T., R. Guinez & B. Battaglia, 1991. Effects of pollutionon heterozygosity in the barnacleBalanus amphitrite(Cirripedia,Thoracica), Mar. Ecol. Prog. Ser. 70(3): 237–243.
Paul, J. D. & I. M. Davies, 1986. Effects of copper and tinbased antifouling compounds on the growth of scallops (Pectenmaximus) and oysters (Crassostrea gigas). Aquaculture 54: 191–203.
Pearce, F., 1999. Choose your poison: Ban one pollutant and anothermay soon take its place. New Sci. 162: 23.
Peterson, C. H., in press. The web of ecosystem interconnectionsto shoreline habitats as revealed by the Exxon Valdez oil spillperturbation: A synthesis of acute direct vs indirect and chroniceffects. Adv. Mar. Biol.
Pettibone, G. W. & J. J. Cooney, 1986. Effects of organotins onfecal pollution indicator organisms. Appl. Environ. Microbiol.52: 562–566.
Pombo, O. A. & A. Escofet, 1996. Effect of exploitation of thelimpetLottia gigantea: A field study in Baja California (Mexico)and California (USA). Pacific Science 50: 393–403.
Pople, A., R. D. Simpson & S. C. Cairns, 1990. An incident ofsouthern oil pollution: Effects of a spillage of diesel fuel on therocky shore of Macquarie Island (sub-Antarctic). Aust. J. Mar.Freshwat. Res. 41: 603–620.
Povey, A. & M. J. Keough, 1991. Effects of trampling on plant andanimal populations on rocky shores. Oikos 61: 355–368.
Power, M. E., D. Tilman, J. A. Estes, B. A. Menge, W. J. Bond, L. S.Mills, G. Daily, J. C. Castilla, J. Lubchenco & R. T. Paine, 1996.Challenges in the quest for keystones. Bioscience 46: 609–620.
Prince, J. D., T. L. Sellers, W. B. Ford & S. R. Talbot, 1988.Confirmation of a relationship between the localized abund-ance of breeding stock and recruitment forHaliotis rubra Leach(Mollusca, Gastropoda). J. Exp. Mar. Biol. Ecol., 122: 91–104.
Proud, S. V., 1994. Tributyltin pollution and the bioindicatorNucella lapillus: population recovery and community levelresponses. Ph.D. thesis, University of Liverpool.

296
Raffaelli, D. & S. Hawkins, 1996.Intertidal Ecology. Chapman andHall, London.
Readman, J. W., J. Bartocci, I. Tolosa, S. W. Fowler, B. Oregioni& M.Y. Abdulraheem, 1996. Recovery of the coastal marineenvironment in the Gulf following the 1991 war related oil spills.Mar. Pollut. Bull. 32: 493–498.
Redpath, K. J. & J. Davenport, 1988. The effect of copper, zincand cadmium on the pumping rate ofMytilus edulisL. Aquat.Toxicol. 13: 217–226.
Robertson, A., 1991. Effects of a toxic bloom ofChrysomulinapolylepis on the common dog-whelk,Nucella lapillus, on theSwedish west-coast. J. Mar. Biol. Assoc. (U.K.) 71: 569–578.
Russell, G. & O. P. Morris, 1970. Copper tolerance in the marinefouling algaeEctocarpus siliculosus. Nature 228: 288–289.
Salazar, M. H. & S. M. Salazar, 1991. Assessing the effects of TBTcontamination with mussel growth rates. Marine Environ. Res.32: 131–150.
Santelices, B. & C. L. Griffiths, 1994. Seaweeds as resources. In:Siegfried, R. S. (ed.),Rocky Shores: Exploitation in Chile andSouth Africa. Springer-Verlag, Berlin: 33–53.
Santos, S. R., S. J. Hawkins, L. R. Montieiro, M. Alves & E. J.Isidro, 1995. Marine research, resources and conservation in theAzores. Aqua. Conserv: Mar. Freshw. Ecosyst. 5: 311–354.
Sarojini, R., A. K. Khan & R. Nagabhushanam, 1989. Oxygenconsumption in the marine portunid crabOzius rugulosusexposed to organophosphate (nuvan) and organochlorine (DDT)pesticides. Indian J. Fish. 36: 82–84.
Schramm, W., 1996. The Baltic Sea and its transition zones. In:Schramm, W. & P. Nienhuis (eds),Marine Benthic Vegetation:Recent Changes and the Effects of Eutrophication. Springer,Berlin: 131–164.
Seapy, R. P. P. & M. M. Littler, 1982. Population and speciesdiversity fluctuations in a rocky intertidal community relative tosevere aerial exposure and sediment burial. Mar. Biol. 71: 87–96.
Sharpe, A. K. & M. J. Keough, 1998. An investigation of the indirecteffects of intertidal shellfish collection. J. Exp. Mar. Biol. Ecol.223: 19–38.
Shaw, D. G.,1992. TheExxon Valdezoil-spill: Ecological and socialconsequences. Environ. Cons. 19(3): 253–258.
Siegfried, W. R. (ed.), 1994.Rocky Shores: Exploitation in Chileand South Africa. Springer-Verlag, Berlin.
Siegfried, W. R., P. A. R. Hockey & G. M. Branch, 1994. Theexploitation of intertidal and subtidal biotic resources of rockyshores in Chile and South Africa – an overview. In: Siegfried, R.S. (ed.),Rocky Shores: Exploitation in Chile and South Africa.Springer-Verlag, Berlin: 1–13.
Smayda, T. J., 1997. Harmful algal blooms: Their ecophysiologyand general relevance to phytoplankton blooms in the sea.Limnol. Oceanogr. 42: 1137–1153.
Smith, J. E., 1968.Torrey Canyon Pollution and Marine Life.Cambridge University Press, London.
Smith, P. J. & M. McVeagh, 1991. Widespread organotin pollu-tion in New Zealand coastal waters as indicated by imposex indogwhelks. Mar. Pollut. Bull. 22: 409–413.
Smith, S. D. A. & R. D. Simpson, 1998. Recovery of benthiccommunities at Macquarie Island (sub-Antarctic) following asmall oil spill. Mar. Biol. 131: 567–581.
Somerfield, P. J., H. L. Rees & R. M. Warwick, 1995. Interrela-tionships in community structure between shallow-water marinemeiofauna and macrofauna in relation to dredgings disposal.Mar. Ecol. Prog. Ser. 127: 103–112.
Southgate, T., K. Wilson, T. F. Cross & A. A. Myers, 1984.Recolonization of a rocky shore in S. W. Ireland following a toxic
bloom of the dinoflagellateGyrodinium aureolum. J. Mar. Biol.Assoc. (U.K.) 64: 485–492.
Southward, A. J., 1982. An ecologists view of the implications ofthe observed physiological and biochemical effects of petroleumcompounds on marine organisms and ecosystems. Phil. Trans. R.Soc. B. 297: 241–255.
Southward, A. J., 1991. Forty years of changes in species composi-tion and population density of barnacles on a rocky shore nearPlymouth. J. Mar. Biol. Assoc. (U.K.) 71: 495–513.
Southward, A. J. & E. C. Southward, 1978. Recolonisation of rockyshores in Cornwall after use of toxic dispersants to clean-up theTorrey Canyonspill. J. Fish. Res. Board Can. 35: 682–706.
Stekoll, M. S. & L. Deysher, 1996. Recolonization and restoration ofupper intertidalFucus gardneri(Fucales, Phaeophyta) followingtheExxon Valdezoil spill. In: Lindstrom, S. C. & D. J. Chapman(eds), Fifteenth International Seaweed Symposium. Develop-ments in Hydrobiology 116. Kluwer Academic Publishers,Dordrecht: 311–316. Reprinted from Hydrobiologia 326/327.
Stirling, H. P., 1977. Effects of a spill of marine diesel oil on therocky shore fauna of Lama Island, Hong Kong. Environ. Pollut.12: 93–117.
Strömgren, T., 1982. Effect of heavy metals (Zn, Hg, Cu, Cd, Pb,Ni) on the length growth ofMytilus edulis. Mar. Biol. 72: 69–72.
Suchanek, T. H., 1993. Oil impacts on marine invertebrate popula-tions and communities. Amer. Zool. 33: 510–523.
Suchanek, T. H., 1994. Temperate coastal marine communities:Biodiversity and threats. Amer. Zool. 34: 100–114.
Tablado, A., J. J. López Gappa & N.H. Magaldi, 1994. Growth ofthe pulmonate limpetSiphonaria lessoni(Blainville) in a rockyintertidal area affected by sewage pollution. J. Exp. Mar. Biol.Ecol. 175: 211–226.
Thain, J., M. J. Waldcock & M. E. Waite, 1987. Toxicity anddegradation studies of tributyltin (TBT) and dibutyltin (DBT)in the aquatic environment. Oceans ’87 Proceedings, Vol. 4:International Organotin Symposium. Institute of Electrical andElectronic Engineers, New York, pp. 1398–1404.
Thom, R. M. & T. B. Widdowson, 1978. A resurvey of a YaleDawson’s 42 intertidal transects on the southern Californiamainland after 15 years. Bull. S. Calif. Acad. Sci. 77:1–13.
Underwood, A. J., 1985. Physical factors and biological interac-tions: The necessity and nature of ecological experiments. In:Moore, P. G. & R. S. Seed (eds),The Ecology of Rocky Coasts.Hodder and Stoughton, London: 372–390.
Underwood, A. J., 1989. The analysis of stress in natural popula-tions. Biol. J. Linn. Soc. 37: 51–78.
Underwood, A. J., 1992. Beyond BACI: The detection of environ-mental impacts on populations in the real, but variable world. J.Exp. Mar. Biol. Ecol. 161: 145–178.
Underwood A. J., 1993a. Exploitation of species on the rocky coastof New South Wales (Australia) and options for its management.Ocean & Coast. Manage. 20: 41–62.
Underwood, A. J., 1993b. The mechanics of spatially replicatedsampling programmes to detect environmental impacts in avariable world. Aust. J. Ecol. 18: 99–116.
Underwood, A. J., 1994. On beyond BACI: Sampling designs thatmight reliably detect environmental disturbances. Ecol. Appl. 4:3–15.
Underwood, A. J. & E. J. Denley, 1984. Paradigms, explana-tions and generalizations in models for the structure of intertidalcommunities on rocky shores. In: Strong, D. R., D. Simberloff,L. G. Abele & A. B. Thistle (eds),Ecological Communities:Conceptual Issues and the Evidence. Princeton University Press,Princeton: 151–180.

297
Underwood, A. J. & P. G. Fairweather, 1989. Supply-side ecologyand benthic marine assemblages. TREE 4: 16–20.
Underwood, A. J. & P. S. Petraitis, 1993. Structure of intertidalassemblages in different locations: How can local processesbe compared? In: Ricklefs, R. & D. Schluter (eds),SpeciesDiversity in Ecological Communities. University of ChicagoPress, Chicago: 38–51.
Underwood, A. J. & G. A. Skilleter, 1996. Effects of patch-size onthe structure of assemblages in rock pools. J. Exp. Mar. Biol.Ecol. 197: 63–90.
Underwood, A. J., E. J. Denley & M. J. Moran, 1983. Exper-imental analyses of the structure and dynamics of mid-shorerocky intertidal communities in New South Wales. Oecologia 56:207–219.
U’ren, S. C., 1983. Acute toxicity of bis(tributyltin) oxide to amarine copepod. Mar. Pollut. Bull. 14: 303–306.
Vadas, R. L., 1979 Abiotic diseases in seaweeds: Thermal effluentsas causal agents, Experientia, 35: 435–437.
Vadas, R. L., M. Keser & B. Larson, 1978. Effects of reducedtemperatures on previously stressed populations of an inter-tidal alga. In: Thorp, J. H. & J. W. Gibbons (eds),Energyand Environmental Stress in Aquatic Systems, DOE SymposiumSeries. (Conf-771114) National Technical Information Service,Springfield, Virginia, USA.
Vanherwerden, L. & C. L. Griffiths, 1991. Human recreationalactivity along the North-Western shores of False Bay. Tran. R.Soc. S. Afr. 47: 737–748.
van Tamelen, P. G. & M. S. Stekoll, 1995. Recovery mechanisms ofthe brown algaFucus gardneri, following catastrophic disturb-ance: Lessons from the Exxon Valdez oil spill. In: Engstrom, D.R. (ed.),Proc. 3rd Glacier Bay Science Symp. US Departmentof the Interior, National Park Service, Alaska Regional Office,Anchorage: 221–228.
van Tamelen, P. G. & M. S. Stekoll, 1996. Population response ofthe brown alga,Fucus gardneri, and other algae in Herring Bay,Prince William sound, to theExxon Valdezoil spill. In: Rice,S. D., R. B. Spies, D. A. Wolfe & B. A. Wright (eds),ExxonValdez Oil Spill Symposium Proceedings, Vol. 18. Amer. Fish.Soc. Symp.: 193–211.
van Tamelen, P. G., M. S. Stekoll & L. Deysher, 1997. Recoveryprocesses of the brown algaFucus gardneri following the“Exxon Valdez” oil spill: settlement and recruitment. Mar. Ecol.Prog. Ser. 160: 265–277.
Vasquez, J. A., 1995. Ecological effects of brown seaweedharvesting. Bot. Mar. 38: 251–257.
Walker, D. I. & G. A. Kendrick, 1998. Threats to macroalgaldiversity: Marine habitat destruction and fragmentation, pollu-tion and introduced species. Bot. Mar. 41: 105–112.
Walters, C. J., 1986.Adaptive Management of RenewableResources. Macmillan, New York.
Walters, C. J., 1993. Dynamic models and large scale field exper-iments in environmental impact assessment and management.Aust. J. Ecol. 18: 53–61.
Warwick, R. M., 1993. Environmental impact studies on marinecommunities: Pragmatical considerations. Aust. J. Ecol. 18:63–80.
Watt, I., T. Woodhouse & D. A. Jones, 1993. Intertidal clean-upactivities and natural regeneration on the Gulf Coast of SaudiArabia from 1991 to 1992 after the 1991 Gulf oil spill. Mar.Pollut. Bull. 27: 325–331.
Webber, L. I., J. P. Thorpe, R. S. Santos & S. J. Hawkins, 1998.Identification of stocks of the exploited limpetsPatella asperaand P. candei at Madeira Archipelego by allozyme electro-phoresis. J. Shellfish Res. 17: 945–953.
Wells, R. A., 1980. Activity patterns as a mechanism of predatoravoidance in two species of Acmaeid limpet. J. Exp. Mar. Biol.Ecol. 48: 151–168.
Wesseling, I., A. J. Uychiaoco, P. M. Alino, T. Aurin & J. E.Vermaat, 1999. Damage and recovery of four Philippine coralsfrom short-term sediment burial. Mar. Ecol. Prog. Ser. 176:11–15.
Wooton, J. T., 1991. Direct and indirect effects of nutrients onintertidal community structure: Variable consequences of seabirdguano. J. Exp. Mar. Biol. Ecol. 151: 139–153.
Worm, B., H. K. Lotze, C. Bostrom, R. Engkvist, V. Labanauskas& U. Sommer, 1999. Marine diversity shift linked to interactionsamong grazers, nutrients and propagule banks. Mar. Ecol. Prog.Ser. 185: 309–314.








