
Molecular ecology of anaerobic granular sludge grown atdifferent conditions
E. Díaz, R. Amils and J.L. Sanz
Centro de Biología Molecular, Universidad Autónoma de Madrid, 28049 España (E-mail: [email protected]; [email protected]; [email protected])
Abstract Qualitative and quantitative diversity of microorganisms present in anaerobic granular sludges fedwith different substrates, as well as the structure of these granules have been studied using fluorescent 16SrRNA-targeted in situ hybridization and electron microscopy. The granules showed a multi-layered structure,in which both densely packed and loose micro-colonies, channels and holes could be observed. Only bacteriawere found in the outer shell of the granules, while both archaea and bacteria were detected in the inner core.Although high cell density was found in the granules (more than 1011 cells/gram, determined by DAPI-stain)only a low percentage of cells was able to hybridize with the rRNA-targeted probes. Significant quantitativeand qualitative differences were observed in the composition of granules fed with different substrates(formate, acetate at high and low concentrations, propionate, sucrose, starch and peptone). Bacterial cellswere mostly gram-positives. Active proteobacteria were scarce in the granules exposed to VFA.Syntrophobacteria became dominant in the propionate-grown biomass. Concerning methanogenic archaea,Methanosaeta was the predominant species using complex substrates or low acetate concentration fedgranules, while Methanosarcina and members of Methanobacteriales were predominant in the granulesgrown at high concentration of acetate or formate, respectively. Other Methanomicrobiales andMethanococcales, have been detected in the anaerobic granular sludge in the conditions used in this work.Keywords Anaerobic granular sludge; FISH; granulation; molecular ecology; molecular probes; UASB
IntroductionAround 75% of the approximately 2,000 anaerobic treatment systems presently in opera-tion world wide correspond to Upflow Anaerobic Sludge Bed (UASB) reactors or to newconfigurations based on the same principle (EGSB, IC). In all of them, microorganismsform compact aggregates up to 2–4 mm in diameter, granular sludge, with high sedimenta-tion velocity and high methanogenic activity. Due to the huge structural complexity ofgranular sludge and the many trophic interactions among the microbial populationsrequired to transform complex organic matter into biogas (CO2 and CH4), the microbialecology (taxonomy, colonization, topological distribution of microbes, etc.) of this micro-ecosystem is still not well understood. Molecular ecology techniques, such as 16S rRNAgene cloning and sequencing (Amann et al., 1995), Fluorescence In Situ Hybridization(FISH) (Amann, 1995; Amann et al., 1995), and Denaturing Gradient Gel Electrophoresis(DGGE) (Muyzer et al., 1993; Muyzer and Smalla, 1998), are nowadays the most powerfultools available to assess the diversity, abundance and distribution of microorganisms innatural and engineered ecosystems, superceding the restrictions and bias of conventionalmicrobiology techniques (isolation, plate-counting, etc.) (Amann et al., 1995; Pace, 1996).
The use of molecular ecology techniques to the study of anaerobic granular sludge isquite recent and allows the main prokaryotic populations developed in the granule and itsinteractions to be identified (Santegoeds et al., 1999; Sekiguchi et al., 1999; Chan et al.,2001; Liu et al., 2002). The importance of this approach to the study of anaerobic watertreatment processes is reflected in the special issue recently published by Water Research(2002). In this paper we present the microbial composition of anaerobic granular sludgesstudied using FISH, TEM and SEM.
Water S
cience and Technology Vo
l 48 No
6 pp
57–64 © IW
AP
ublishing 2003
57

MethodsBiomass and reactor experimental set-up
All the experiments were carried out using granular sludge from a brewery full scale UASBreactor (Mahou, Guadalajara, Spain) as inoculum and control. Because the differentbacterial populations developed in the granules are dependent on the type of pollutantspresent in the wastewater, the following substrates have been evaluated: formate, acetate atboth high and low concentrations, propionate, sucrose, starch and peptone. Assays werecarried out in 1 litre batch reactors. The reactors were fed by spike doses to keep the desiredsubstrate concentration constant. Each reactor was monitored for 3 months.
Fluorescent in situ Hybridization (FISH)
The protocols described by Amann (Amann et al., 1990, 1995) were used for the hybridiza-tion experiments. The samples were fixed, just after their collection, in 4% para-formaldehyde, subsequently washed in phosphate buffer saline (PBS) and preserved inPBS-ethanol at –20°C until use. For the hybridization procedure, the samples were fixed ona multi-dish slide at 46°C and dried in ethanol (50, 80, 100%). Formamide concentrationsused for hybridization (2 h, 46°C) and NaCl concentrations used for washing (15 min,48°C) are listed in Table 1. The probes, marked at the 5′-end with fluorescein or Cy3, werepurchased from Genotek (Barcelona, Spain).
Microscopy and cell counting
The total cells present in the samples were determined by counting 4′,6′-diamin phenylin-dol (DAPI) stained cells. Representative samples of the granules (0.03 g of wet weightgranules in 1 ml of PBS: ethanol 1:1) were sonicated 30–60 s at middle power and 1 cycle/s.To quantify the cells, the cell suspensions were diluted 1/50 for DAPI-counting and 1/5 forhybridized cell-counting. Both DAPI-stain and hybridize cells were examined with anepifluorescent microscope Zeiss Axioskop. Ten fields (corresponding to 500–1,000 DAPI-
E. D
íaz et al.
58
Table 1 DNA probes used in this work, with indications of the target site, specificity andhybridization/washing conditions
Probe Sequence (5’–3’) Formamide Specificity
(%)/NaCl (mmol l–1)
EUB338 GCTGCCTCCCGTAGGAGT 35/80 BacteriaARC915 GTGCTCCCCCGCCAATTCCT 20/225 ArchaeaNON338 ACTCCTACGGGAGGCAGC 35/80 None (negative control)ALF968 GGTAAGGTTCTGCGCGTT 20/225 Alpha-proteobacteriaBET42a GCCTTCCCACTTCGTTT 35/80 Beta-proteobacteriaGAM42a GCCTTCCCACATCGTTT 35/80 Gamma-proteobacteriaSRB385 CGGCGTCGCTGCGTCAGG 35/80 Sulfate-reducing bacteriaDSS658 TCCACTTCCCTCTCCCAT 60/15.6 Desulfosarcina, DesulfococcusDSV698 GTTCCTCCAGATATCTACGG 35/80 DesulfovibrioBAC1080 GCACTTAAGCCGACACCT 20/225 BacteroidesSYN835 GCGGGTACTCATTCCTG 35/80 Syntrophobacter sp.LGC354A TGGAAGATTCCCTACTGC 20/225 Low GC Gram-positiveHGC69A TATAGTTACCACCGCGT 25/159 High GC Gram-positiveMEB859 GGACTTAACAGCTTCCCT 25/159 Methanobacteriales
(except for Methanothermaceae)MC1109 GCAACATAGGGCACGGGTCT 35/80 MethanococcalesMG1200 CGGATAATTCGGGGCATGCTG 5/636 Methanomicrobiales
(except for Methanosarcinaceae)MSSH859 TCGCTTCACGGCTTCCCT 35/80 MethanosarcinaceaeMS1414 CTCACCCATACCTCACTCGGG 35/80 Methanosarcinaceae
(except for Methanosaeta)MX825 TCGCACCGTGGCCGACACCTAGC 20/225 Methanosaeta

stained cells) were counted to determine the average number of cells per sample. Forgranular structural studies, the specimens were microscopically examined with aRadiance2000 confocal laser scanning microscope (CLSM) equipped with He-Ne lasersand a reversed microscope Zeiss Axiovert S100. Transmission electron microscopy wasdone according to the protocol described by Grotenhuis et al. (1991) using a JEM1010microscope. Scanning electron microscopy was performed as described by Alphenaar et al.(1994) with a Philips XL30 microscope.
Results and discussionTwo different approaches were used to study the microbial diversity present in the granule:i) hybridization with 16S rRNA-targeted probes (FISH) and both transmission (TEM) aswell as scanning (SEM) electron microscopy of thin sections of the whole granule, in orderto reveal the spatial distribution of THE microorganisms, and ii) hybridization of crushedgranules to quantify the different microbial groups (FISH).
Structure and microbial distribution of a granular sludge
TEM of granular sludge showed different structural features: densely packed micro-colonies, loose colonies, channels and holes. Clearly defined micro-colonies of a singlemicrobial species appear in the granules, together with overlapping heterogeneous colonies(Figure 1: A, B), probably as a result of distinct metabolic specificities and interactions(hydrogen or other metabolic products) of different microbial populations. The holes andchannels might be related to the growth of the granule and the required diffusion of sub-strates and products. The presence of low electron dense cells (ghost microorganisms) isnoteworthy (Figure 1: C). These forms could be resting or dead cells and their presence mayexplain the low percentage of active microorganisms detected by FISH.
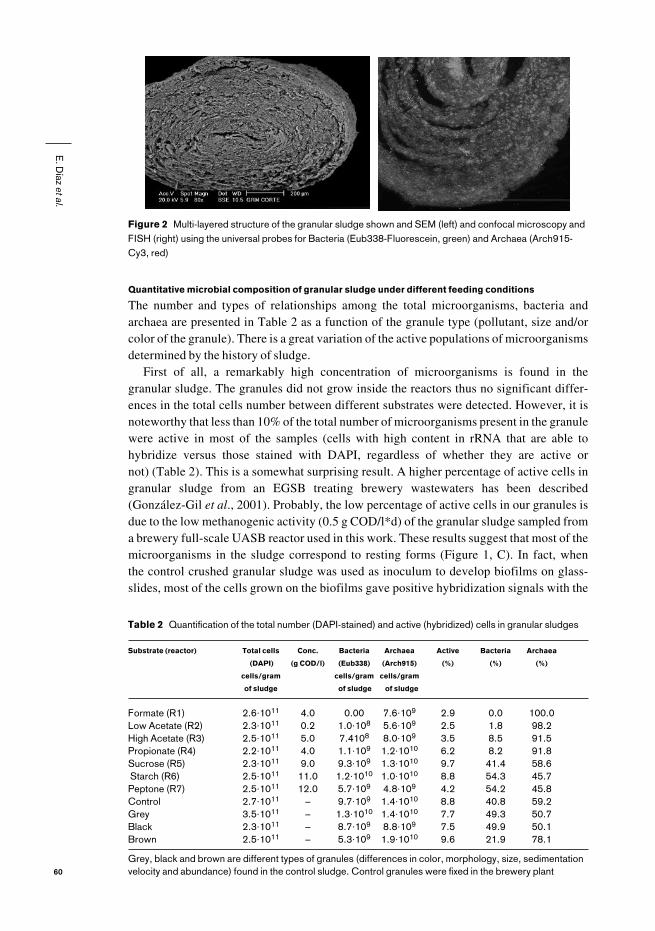
FISH, TEM and SEM microscopy of the granular sludge revealed a multi-layer structure(Figure 2). A primary spatial distribution of archaea and bacteria could be observed byFISH. Bacteria always appear in the outer shell of the granule, while archaea are alwayslocated in its interior (Figure 2b), as was also described by Sekiguchi et al. (1999). Thisfeature reflects the distribution of well characterized facultative aerobic bacteria in theouter layer which consume oxygen (higher metabolic activity) and prevent its diffusioninto the granule, thus protecting the strict anaerobes (especially methanogenic archaea andsulfate-reducing bacteria) from oxygen toxicity. Inside the granule, bacteria and archaeaare arranged in micro-colonies (Figures 1 and 2).
E. D
íaz et al.
59
Figure 1 TEM of granular sludge. A: colony of a single microorganism. B: heterogeneous colonies made ofseveral microorganisms, with a channel between them. C: low electron density cells. Amplifications: A andB: 1,500×; C: 4,000×
Quantitative microbial composition of granular sludge under different feeding conditions
The number and types of relationships among the total microorganisms, bacteria andarchaea are presented in Table 2 as a function of the granule type (pollutant, size and/orcolor of the granule). There is a great variation of the active populations of microorganismsdetermined by the history of sludge.
First of all, a remarkably high concentration of microorganisms is found in the granular sludge. The granules did not grow inside the reactors thus no significant differ-ences in the total cells number between different substrates were detected. However, it isnoteworthy that less than 10% of the total number of microorganisms present in the granulewere active in most of the samples (cells with high content in rRNA that are able tohybridize versus those stained with DAPI, regardless of whether they are active or not) (Table 2). This is a somewhat surprising result. A higher percentage of active cells ingranular sludge from an EGSB treating brewery wastewaters has been described(González-Gil et al., 2001). Probably, the low percentage of active cells in our granules isdue to the low methanogenic activity (0.5 g COD/l*d) of the granular sludge sampled froma brewery full-scale UASB reactor used in this work. These results suggest that most of themicroorganisms in the sludge correspond to resting forms (Figure 1, C). In fact, when the control crushed granular sludge was used as inoculum to develop biofilms on glass-slides, most of the cells grown on the biofilms gave positive hybridization signals with the
E. D
íaz et al.
60
Figure 2 Multi-layered structure of the granular sludge shown and SEM (left) and confocal microscopy andFISH (right) using the universal probes for Bacteria (Eub338-Fluorescein, green) and Archaea (Arch915-Cy3, red)
Table 2 Quantification of the total number (DAPI-stained) and active (hybridized) cells in granular sludges
Substrate (reactor) Total cells Conc. Bacteria Archaea Active Bacteria Archaea
(DAPI) (g COD/l) (Eub338) (Arch915) (%) (%) (%)
cells/gram cells/gram cells/gram
of sludge of sludge of sludge
Formate (R1) 2.6·1011 4.0 0.00 7.6·109 2.9 0.0 100.0Low Acetate (R2) 2.3·1011 0.2 1.0·108 5.6·109 2.5 1.8 98.2High Acetate (R3) 2.5·1011 5.0 7.4108 8.0·109 3.5 8.5 91.5Propionate (R4) 2.2·1011 4.0 1.1·109 1.2·1010 6.2 8.2 91.8Sucrose (R5) 2.3·1011 9.0 9.3·109 1.3·1010 9.7 41.4 58.6Starch (R6) 2.5·1011 11.0 1.2·1010 1.0·1010 8.8 54.3 45.7Peptone (R7) 2.5·1011 12.0 5.7·109 4.8·109 4.2 54.2 45.8Control 2.7·1011 – 9.7·109 1.4·1010 8.8 40.8 59.2Grey 3.5·1011 – 1.3·1010 1.4·1010 7.7 49.3 50.7Black 2.3·1011 – 8.7·109 8.8·109 7.5 49.9 50.1Brown 2.5·1011 – 5.3·109 1.9·1010 9.6 21.9 78.1
Grey, black and brown are different types of granules (differences in color, morphology, size, sedimentationvelocity and abundance) found in the control sludge. Control granules were fixed in the brewery plant
fluorescent probes, indicating that the majority of the cells were metabolically active (datanot shown).
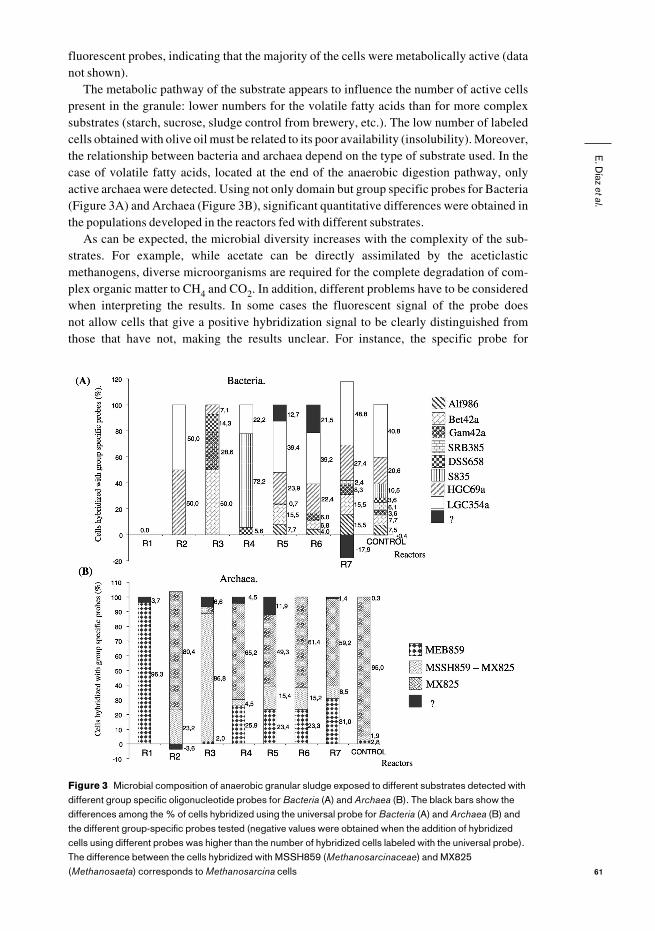
The metabolic pathway of the substrate appears to influence the number of active cellspresent in the granule: lower numbers for the volatile fatty acids than for more complexsubstrates (starch, sucrose, sludge control from brewery, etc.). The low number of labeledcells obtained with olive oil must be related to its poor availability (insolubility). Moreover,the relationship between bacteria and archaea depend on the type of substrate used. In thecase of volatile fatty acids, located at the end of the anaerobic digestion pathway, onlyactive archaea were detected. Using not only domain but group specific probes for Bacteria(Figure 3A) and Archaea (Figure 3B), significant quantitative differences were obtained inthe populations developed in the reactors fed with different substrates.
As can be expected, the microbial diversity increases with the complexity of the sub-strates. For example, while acetate can be directly assimilated by the aceticlasticmethanogens, diverse microorganisms are required for the complete degradation of com-plex organic matter to CH4 and CO2. In addition, different problems have to be consideredwhen interpreting the results. In some cases the fluorescent signal of the probe does not allow cells that give a positive hybridization signal to be clearly distinguished fromthose that have not, making the results unclear. For instance, the specific probe for
E. D
íaz et al.
61
Figure 3 Microbial composition of anaerobic granular sludge exposed to different substrates detected withdifferent group specific oligonucleotide probes for Bacteria (A) and Archaea (B). The black bars show thedifferences among the % of cells hybridized using the universal probe for Bacteria (A) and Archaea (B) andthe different group-specific probes tested (negative values were obtained when the addition of hybridizedcells using different probes was higher than the number of hybridized cells labeled with the universal probe).The difference between the cells hybridized with MSSH859 (Methanosarcinaceae) and MX825(Methanosaeta) corresponds to Methanosarcina cells

gram-positives did not uniformly label whole cells, or the brightness of probe MA1414 wasvery intense when used to hybridize Methanosarcina but also other rod-shapes cells gaveless intense positive hybridization, and as a consequence the cell count can be overestimat-ed. In other cases, mainly for VFA and bacteria, the percentage of active cells was very low(i.e. 1.1% of active bacteria in the granules grown at low acetate concentration), thus spe-cial care must be taken in the interpretation of the results. The control granule is the onlysystem that presents all the bacterial groups tested in this work. Diversity decreases whenthe granules are exposed to a variety of substrates ranging from macromolecules (starch,peptone) to VFA, in which typical proteobacteria (α-, β-, γ-proteobacteria) are scarce (withthe only exception in the reactor being fed at high acetate concentration). The formate-grown granules are an extreme case, since active bacteria are absent. Interestingly enough,syntrophobacteria (propionate-oxidizing bacteria) become dominant in the propionate-grown biomass. On the other hand, gram-positive bacteria accounted for a high percentageof the total hybridized bacteria. A similar result was found by Liu (Liu et al., 2002). 50% ofthe bacterial clones obtained from granular sludge were affiliated to the low G+C gram-positive group of bacteria.
In the case of methanogenic archaea, we did not find positive hybridization signals witheither MC1109 or with MG1200 probes. Araujo (Araujo et al., 2000) has also reportednegative hybridization results using both probes in biofilms developed using granular sludgefrom a domestic sewage treatment plant as inoculum, and Liu (Liu et al., 2002) found only 1% of Methanococcales. MC1109 is a specific probe for Methanococcales, which have,until now, only been isolated from marine environments. MG1200 is specific formethanogens belonging to the families Methanomicrobiaceae, Methanocorpusculaceae andMethanoplanaceae (included in the order Methanomicrobiales) limited to the use of H2 andformate as substrates. Most of the members of this group require a relatively high sodium ionconcentration. Thus, the absence of this type of methanobacteria in the granular sludgewould not be surprising. On the other hand, due to the apparent low specificity of the MS1414probe, probes MSSH859 and MX825 were used to test all the Methanosarcinaceae species,and to discriminate between Methanosarcina and Methanosaeta.
The granules fed with complex substrates presented higher diversity of methanogenicarchaea, similar to what was observed with Bacteria. In this case, Methanosaeta was thepredominant species. It is important to underscore that the reactor fed with formate present-ed a total prevalence of microorganisms hybridizing with the Methanobacteriales probe(MEB859), whose morphology resembled that of Methanobrevibacter spp. orMethanobacterium spp., with almost total absence of active bacteria. Both observations areconsistent considering that formic acid is directly metabolized by methanogenic archaea.
Other meaningful examples are the characteristics of the biomass obtained in the reac-tors fed with acetate, in which 90–100% of the active microorganisms were archaea. In thiscase all the cells stained with the universal archaeal probe also hybridized with theMethanosarcinaceae probe (MSSH859). The sludge exposed to high concentrations ofacetate presented a high proportion of Methanosarcina (probe MS1414, Figures 3B and4A), while members of this genera were scarcely detectable in the reactor fed with low con-centrations of acetate. However, in this case a high level of hybridization was obtained withthe specific probe for Methanosaeta (MX825, Figures 3B and 5B). These results agree withthe kinetic parameters for both species of aceticlastic methanobacteria. Methanosarcinahas a higher growth rate for acetate that Methanosaeta (µmax 0.21 d–1 versus 0.11 d–1),although its affinity for acetate is lower (Ks 4.02 mmol l–1 versus 0.44 mmol l–1). Thus, thepredominance of Methanosaeta at low concentrations of acetate can be linked to its highaffinity for this substrate. Methanosarcina became dominant if the acetate concentrationincreased to several grams per litre.
E. D
íaz et al.
62

ConclusionsMolecular techniques (FISH) and electron microscopy (TEM and SEM) are very usefultools to study complex micro-cosmos such as anaerobic granular sludge. Using these tech-niques the following conclusions can be drawn.1. Granules have a multi-layer structure. In the outer layers only bacteria are present.
Densely packed micro-colonies, loose colonies (both pure and heterogeneous), chan-nels and holes can be observed in the inner layers.
2. FISH using domain and group-specific probes, showed significant differences in themicrobial populations developed using different substrates, although only a low per-centage of cells were active. Microbial diversity (both bacteria and methanogenicarchaea) increased with the complexity of the substrates.
3. Gram-positive bacteria accounted for a high percentage of the total hybridized bacteria.α-, β-, and γ-proteobacteria were scarce in VFA-cultivated granules. Active bacteriawere absent in formate-grown granules. Syntrophobacteria became dominant in thepropionate-grown biomass.
4. Methanosaeta was the predominant archaeal species using granules exposed to com-plex substrates or low acetate concentration. Methanosarcina and members ofMethanobacteriales were the majority in the granules cultivated with a high concentra-tion of acetate and formate respectively. Methanococcales and Methanomicrobiales,with the exception of members of the family Methanomicrobiaceae, were neverobserved in the different incubation conditions used in this work.
AcknowledgementsThis work was partially supported by a grant from the Ministerio español de Ciencia yTecnología to José L. Sanz, project number REN2001-2980-C02-02/HID. E. Díaz is a fellowof the Agencia Española de Cooperación Internacional (Ministerio de Asuntos Exteriores).
ReferencesAlphenaar, P.A., Groeneveld, N. and van Aelst, A.C. (1994). Scanning electron microscopical method for
internal structure analysis of granular sludge. Micron, 25(2), 129–133.
Amann, R.I., Binder, B.J., Olson, R.J., Chisholm, S.W., Devereux, R. and Stahl, D.A. (1990). Combination
of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial
populations. Appl. Environ. Microbiol., 56(6), 1919–1925.
Amann, R.I. (1995). In situ identification of microorganisms by whole cell hybridization with rRNA-
targeted nucleic acid probes. In: Molecular Microbial Ecology Manual, D.L. Akkermans, J.D. van Elsas
and F.J. de Bruijn (eds.). Kluwer Academic Publishers, Dordrecht, 3.3.6, 1–15.
Amann, R.I., Ludwig, W. and Schleifer, K.H. (1995). Phylogenetic identification and in situ detection of
individual microbial cells without cultivation. Microbiol. Rev., 59, 143–169.
E. D
íaz et al.
63
A B
Figure 4 Microbial diversity of an anaerobic granular sludge fed exclusively with acetate. 4A: high concen-tration of acetate. Methanosarcina (FISH: MS1414-Cy3). 4B: low concentration of acetate. Prevalence ofMethanosaeta-like microorganisms (SEM)

Araujo, J.C., Brucha, G., Campos, J.R. and Vazoller, R.F. (2000). Monitoring the development of anaerobic
biofilms using fluorescent in situ hybridization and confocal laser scanning microscopy. Wat. Sci. Tech.,
41(12), 69–77.
Chan, O.-C., Liu, W-T. and Fang, H.H.P. (2001). Study of microbial community of brewery-treating
granular sludge by denaturing gradient gel electroforesis of 16S rRNA gene. Wat. Sci. Tech., 43(1),
77–82.
González-Gil, G., Lens, P.N.L., van Aelst, A., Versprille, A.I. and Lettinga, G. (2001). Cluster structure of
anaerobic aggregates of an expanded granular sludge bed reactor. Appl. Microbiol. Biotechnol., 67,
3683–3692.
Grotenhuis, J.T.C., Smit, M., van Lammeren, A.A.M., Stams, A.J.M. and Zehnder, A.J.B. (1991).
Localization and quantification of extracellular polymers in methanogenic granular sludge. Appl.
Microbiol. Biotechnol., 36, 115–119.
Liu, W.-T., Chan, O.-C. and Fang, H.H.P. (2002). Characterization of microbial community in granular
sludge treating brewery wastewater. Wat. Res., 36, 1767–1775.
Muyzer, G. and Smalla, K. (1998). Application of denaturing gradient gel electrophoresis (DGGE) and
temperature gradient gel electrophoresis (TGGE) in microbial ecology. Mini Review. Antonie van
Leeuwnhoek, 73, 127–141.
Muyzer, G., de Waal, E.C. and Uitterlinden, A.G. (1993). Profiling of complex microbial populations by
denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding
for 16S rRNA. App. Environ. Microbiol., 59(3), 695–700.
Pace, N.R. (1996). New perspective on the natural microbial world: molecular microbial ecology. ASM
News, 62(9), 463–470.
Raskin, L., Stromley, J.M., Rittmann, B.E. and Stahl, D.A. (1994). Group specific 16S rRNA hybridation
probes to describe natural communities of methanogens. App. Environ. Microbiol., 60(4), 1232–1240.
Santegoeds, C.M., Damgaard, L.R., Hesselink, G., Zopfi, J., Lens, P., Muyzer, G. and de Beer, D. (1999).
Distribution of sulfate-reducing and methanogenic bacteria in anaerobic aggregates determined by
microsensor and molecular analyses. Appl. Environ. Microbiol., 65(10), 4618–4629.
Sekiguchi, Y., Kamagata, Y., Nakamura, K., Ohashi, A. and Harada, H. (1999). Fluorescence in situ
hybridization using 16S rRNA-targeted oligonucleotides reveals localization of methanogens and
selected uncultured bacteria in mesophilic and thermophilic sludge granules. Appl. Environ. Microbiol.,
65(3), 1280–1288.
Water Research, 36(2), (2002). Special Issue: Modern Scientific Tools in Bioprocessing.
E. D
íaz et al.
64







