
stcechddtdVhtt
pPsa
Brain-Derived Neurotrophic Factor Val66Met AlleleImpairs Basal and Ketamine-StimulatedSynaptogenesis in Prefrontal CortexRong-Jian Liu, Francis S. Lee, Xiao-Yuan Li, Francis Bambico, Ronald S. Duman, andGeorge K. Aghajanian
Background: Knock-in mice with the common human brain-derived neurotrophic factor (BDNF) Val66Met polymorphism have impairedtrafficking of BDNF messenger RNA to dendrites. It was hypothesized, given evidence that local synapse formation is dependent on dendritictranslation of BDNF messenger RNA, that loss-of-function Met allele mice would show synaptic deficits both at baseline and in response toketamine, an N-methyl-D-aspartate antagonist that stimulates synaptogenesis in prefrontal cortex (PFC).
Methods: Whole-cell recordings from layer V medial PFC pyramidal cells in brain slices were combined with two-photon laser scanning foranalysis of wildtype, Val/Met, and Met/Met mice both at baseline and in response to a low dose of ketamine.
Results: Val/Met and Met/Met mice were found to have constitutive atrophy of distal apical dendrites and decrements in apically targetedexcitatory postsynaptic currents in layer V pyramidal cells of PFC. In addition, spine density and diameter were decreased, indicative ofimpaired synaptic formation/maturation (synaptogenesis). In Met/Met mice the synaptogenic effect of ketamine was markedly impaired,consistent with the idea that synaptogenesis is dependent on dendritic translation/release of BDNF. In parallel behavioral studies, we foundthat the antidepressant response to ketamine in the forced swim test was blocked in Met/Met mice.
Conclusions: The results demonstrate that expression of the BDNF Met allele in mice results in basal synaptic deficits and blockssynaptogenic and antidepressant actions of ketamine in PFC, suggesting that the therapeutic response to this drug might be attenuated or
blocked in depressed patients who carry the loss of function Met allele.cmoardbmlww
srTmritroddcpaBs
M
A
p
Key Words: Antidepressant, dendritic spines, EPSC, hypocretin,major depression, N-methyl-D-aspartate, serotonin
A decrease in brain-derived neurotrophic factor (BDNF) ex-pression in medial prefrontal cortex (mPFC) and other re-gions has given rise to the BDNF hypothesis of major depres-
ion (1– 4). A human polymorphism in the BDNF gene, which leadso a valine-to-methionine substitution in the proBDNF protein atodon 66 (Val66Met), is carried by approximately 30% of the gen-ral population and has been associated with mild cognitive defi-its (5–7). In addition, individuals who carry the Met polymorphismave been reported to have an increased risk of stress-related majorepression (8 –10). The Val66Met polymorphism impairs activity-ependent secretion of BDNF at synaptic sites (5,11) and reduces
ranslin-mediated trafficking of BDNF messenger RNA (mRNA) toendrites (12). Recently, Chen et al. (13) generated a BDNFal66Met knock-in mouse exhibiting phenotypic hallmarks of thisuman polymorphism, providing an animal model for analysis of
his human loss-of-function BDNF polymorphism on synaptic func-ion.
In the present study, we focused on the mPFC because of itsivotal role in neurocircuits underlying major depression (14 –16).reviously we found that rats exposed to repeated mild restrainttress or the stress hormone corticosterone exhibit atrophy of distalpical dendrites and matching deficits in excitatory postsynaptic
From the Departments of Psychiatry (R-JL, X-YL, FB, RSD, GKA) and Pharma-cology (RSD, GKA), Yale School of Medicine, New Haven, Connecticut;and the Department of Psychiatry (FSL), Weill Cornell Medical College ofCornell University, New York, New York.
Address correspondence to George Aghajanian, M.D., Ph.D., Yale School ofMedicine, Departments of Psychiatry and Pharmacology, 34 Park Street,New Haven, CT 06508; E-mail: [email protected].
aReceived Aug 11, 2011; revised Sep 27, 2011; accepted Sep 28, 2011.
0006-3223/$36.00doi:10.1016/j.biopsych.2011.09.030
urrents (EPSCs) generated in the apical dendrites of layer V pyra-idal neurons of the mPFC (17). The synaptic responses consisted
f EPSCs elicited by serotonin (5-HT) and hypocrtin/orexin, whichct via apically targeted corticocortical and thalamocortical inputs,espectively (18,19). Given the role of BDNF in modulating activity-ependent synaptic plasticity among mature neurons (20,21), it haseen suggested that reductions in EPSCs induced by chronic stressight be mediated by a stress/corticosterone-induced downregu-
ation of BDNF expression (17). It remains to be determinedhether mice with the reduced-function BDNF Val66Met knock-inould exhibit similar synaptic deficits.
Recently, it was reported that a single subanesthetic dose of thehort-acting N-methyl-D-aspartate (NMDA) antagonist ketamineapidly increases the density of functional synaptic spines in mPFC.his synaptogenic effect is preceded by a transient increase inammalian Target of Rapamycin (mTOR) in prefrontal synaptoneu-
osomes followed within approximately 2 hours by a prolongedncrease in synaptic proteins and a concurrent antidepressant ac-ion lasting 1 week or more (22). These effects are blocked byapamycin (intracerebroventricular), a drug that disrupts activationf mTOR, a serine/threonine kinase instrumental in the activation ofendritic translation of synaptic proteins (23). Studies in isolatedendrites demonstrate that puff application of BDNF increases lo-al translation of proteins in an anatomically restricted, mTOR-de-endent manner (24). The BDNF Val66Met knock-in mouse offersn opportunity to test whether this polymorphism, by impairingDNF function, interferes with the synaptogenic and antidepres-ant effects of ketamine.
ethods and Materials
nimalsThe generation of the BDNF Val66Met knock-in mice has been
reviously described (13). Heterozygous Met/Val mice were bred,
nd the 3 resulting littermate genotypes, Val/Val wildtype (WT),BIOL PSYCHIATRY 2012;71:996–1005© 2012 Society of Biological Psychiatry

mpfite
psCDmacwl
rctsvC
I
asta(Oo(m21bdwddssadd
F
tmctmJtmTmiseaii
fimSptlcMspts
R.-J. Liu et al. BIOL PSYCHIATRY 2012;71:996–1005 997
Val/Met, and Met/Met mice were used. The age of mice was 6 – 8months for the electrophysiology and spine morphology experi-ments and 3–10 months for the behavioral studies. Mice werehoused and maintained in standard conditions with a 12-hourlight/dark cycle and ad libitum access to food and water. Injec-tions of ketamine hydrochloride (10 mg/kg, IP) were made 24hours before preparation of brain slices. Animal use and proce-dures were in accordance with the National Institutes of Healthguidelines and approved by the Yale University Animal Care andUse Committees.
Brain Slice PreparationBrain slices were prepared as described (17). Briefly, mice were
anesthetized with chloral hydrate (400 mg/kg, IP), in adherencewith protocols approved by the Yale Animal Care and Use Commit-tee. After decapitation, the brains were removed rapidly and placedin ice-cold (approximately 4°C) artificial cerebrospinal fluid (ACSF)in which sucrose (252 mmol/L) was substituted for sodium chloride(sucrose-ACSF) to prevent cell swelling. A block of tissue containingPFC was dissected, and coronal slices (400 �m) were cut in sucrose-ACSF with an oscillating-blade tissue slicer (Leica VT1000S; Leica,Bannockburn, Illinois) and placed in a submerged recording cham-ber; bath temperature was raised slowly to 32°C. Known concentra-tions of drugs dissolved in ACSF, applied through a stopcock ar-rangement at a fast flow rate (approximately 4 ml/min), reached theslice within 7–10 s. The standard ACSF (pH approximately 7.35) wasequilibrated with 95% oxygen/5% carbon dioxide and contained128 mmol/L sodium chloride, 3 mmol/L potassium chloride, 2mmol/L calcium chloride, 2 mmol/L magnesium sulphate, 24mmol/L sodium bicarbonate, 1.25 mmol/L sodium dihydrog-enorthophosphate, and 10 mmol/L D-glucose. A recovery period ofapproximately 1–2 hours was allowed before commencement ofrecording.
ElectrophysiologyPyramidal neurons in layer V were visualized by an Olympus
BX50WI microscope (�40 or �60 infrared [IR] lens) with IR differ-ential interference contrast (IR/differential interference con-trast) video-microscopy (Olympus, Hamburg, Germany), as de-scribed (19). Low-resistance patch pipettes (3–5 M�) werepulled from patch-clamp glass tubing (Warner Instruments,Hamden, Connecticut) by a Flaming-Brown Horizontal Puller(model P-97; Sutter Instruments, Novato, California). Pipetteswere filled with the following solution: 115 mmol/L potassiumgluconate, 5 mmol/L potassium chloride, 2 mmol/L magnesiumchloride, 2 mmol/L magnesium-adenosine triphosphate, 2 mmol/Ldisodium (Na2) adenosine triphosphate, 10 mmol/L Na2-phos-phocreatine, .4 mmol/L Na2 guanosine triphosphate, and 10
mol/L Hepes, pH 7.33. Neurobiotin (.3%) was added to theipette solution to mark cells for later imaging. Pipettes wererst tip-filled with regular patch solution before backfilling withhe Neurobiotin solution to avoid ejecting excess dye into thextracellular space of the slice.
Whole-cell recordings were made with an Axoclamp-2B am-lifier (Axon Instruments, Union City, California). The outputignal was low-pass-filtered at 3 KHz, amplified �100 throughyberamp, digitized at 15 kHz, and acquired with pClamp 9.2/igidata 1320 software (Axon Instruments). Series resistance,onitored throughout the experiment, was usually between 4
nd 8 M�. To minimize series resistance errors, cells were dis-arded if series resistance rose above 10 �. Postsynaptic currentsere in continuous single-electrode voltage-clamp mode (3-kHz
ow-pass filter cutoff frequency); cells were clamped near their c
esting potential (approximately 75 � 5 mV) to minimize holdingurrents. After completion of recording, slices were transferredo 4% paraformaldehyde in .1 mol/L phosphate buffer andtored overnight at 4°C. Slices were then processed with strepta-idin conjugated to Alexa 594 (1/1000; Invitrogen, Carlsbad,alifornia) for Neurobiotin visualization.
maging and Data AnalysisLabeled neurons within layer V of anterior cingulate (Cg1)
nd prelimbic mPFC (Cg3) were imaged with a two-photon lasercanning system consisting of a Ti:sapphire laser (Mai Tai; Spec-ra Physics, Santa Clara, California) tuned to wavelength 810 nmnd a direct detection Bio-Rad Radiance 2100 MP laser scannerZeiss Microimaging, Thornwood, New York) mounted on anlympus BX50WI microscope with �40 (.8 numerical aperture)r �60 (.9 numerical aperture) water-immersion objectives
Olympus). Total apical and basal dendritic branch length, 3-di-ensional Z-stacks were reconstructed from approximately 40 –
50 sequential scans at low zoom (312 � 312 �m; �40 lens) at-�m steps so as to include the entire apical tuft and basalranches for a given cell within the slice. Total branch length wasetermined within the 3-dimensional matrix of each Z-stackith Neurolucida 9 (MicroBrightField, Williston, Vermont). Spineensity, spine head diameter, and spine length analysis wereone with Neuolucida Explorer (version 9) on the raw imagetacks (2–9 optical sections, 1 �m apart). Spine density wasampled in two zones: 1) tips of apical tuft branches as theypproach the pial membrane, and 2) proximal tuft dendrites justistal to the bifurcation. Results were expressed in terms of spineensity/10 �m.
orced Swim TestThe mouse forced swim test (FST) examines the dynamics of
ransition from an active (struggling) to passive (immobility)ode of coping in an inescapable water-filled glass beaker (11
m diameter, 15.5 cm high; 10 cm water depth; 25°C wateremperature). In each run, four mice were video-recorded for 6
in with a digital video camcorder (DCR-SX20, Sony, Tokyo,apan). The locations of the animals were counter-balanced onhe basis of their experimental groups. Over the first 2 min, test
ice normally undergo habituation of struggling behavior.hereafter, the antidepressant or pro-depressive effects of phar-acological and genetic manipulations could be distinguished,
ndicated by an enhancement of struggling and immobility, re-pectively, with manual (25) or automated scoring (26). Afterach run, mice were rescued, dried with a towel, and placed near
heat source. All runs were conducted under minimal anx-ogenic conditions (27). Video clips were stored as audio videonterleaved (AVI) files for offline analyses.
For quantification of animal movement from the FST videoles, silhouette alteration method was employed with an auto-ated behavioral tracking system (Videotrack; View Point Life
cience, Montreal, Canada). Each video image is rapidly scannedoint-by-point and line-after-line under an 800 � 600 pixel set-
ing (for 40 msec, [i.e., 25 images/sec]). Each point is assigned auminosity value of 0 –255 on a gray scale. Values were thenonverted to a binary image: each pixel is white (0) or black (1).ice were distinguished from the background by adjusting the
ensitivity threshold of this conversion, associating all non-zeroixels in each frame with the silhouette of the mouse (including
he head, tail, and paws). During movement analyses, degree ofilhouette alteration (i.e., the number of pixels coded 0 or 1)
hanged between successive frames, indicates the level of strug-www.sobp.org/journal
D
ns(ts
V
998 BIOL PSYCHIATRY 2012;71:996–1005 R.-J. Liu et al.
gling or immobility displayed. With an integrated threshold al-gorithm, the threshold for immobility is fine-tuned (at a value of20) before the actual analyses so that the mouse is consideredimmobile (minimal movements) when it is visually verified assuch. This and other automated FST behavioral quantitationhave been tested to serve as reliable methods that address thelimitations of manual (visual) scoring by experimenters (26,28).
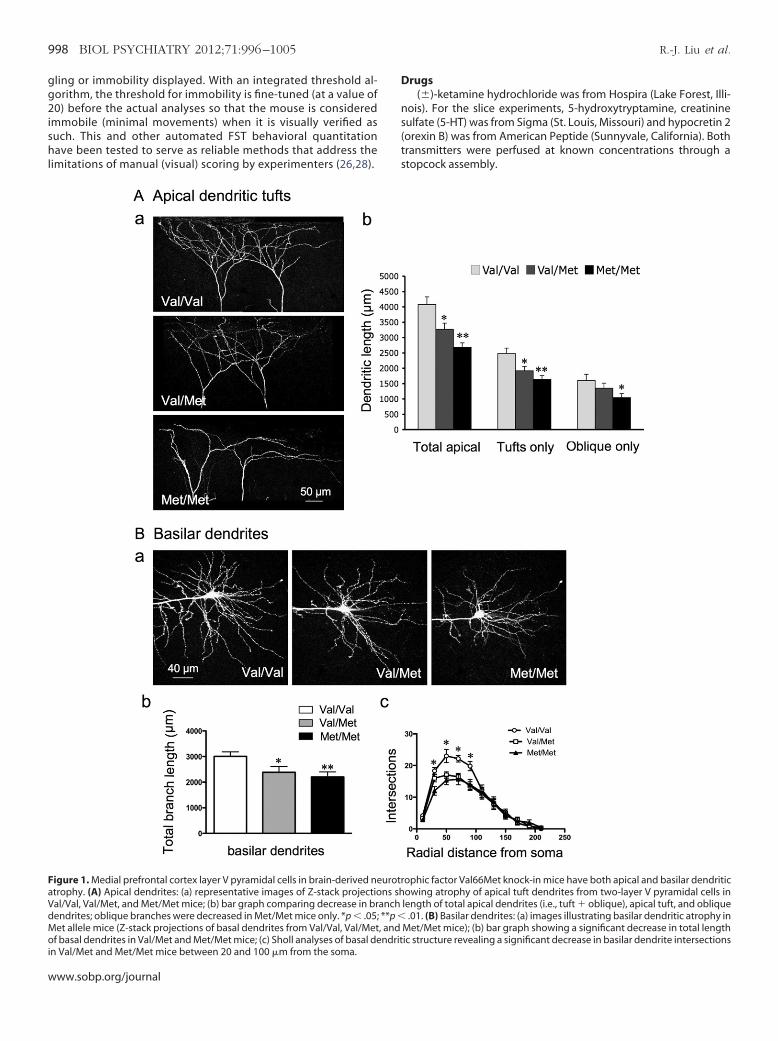
Figure 1. Medial prefrontal cortex layer V pyramidal cells in brain-derived neatrophy. (A) Apical dendrites: (a) representative images of Z-stack projectio
al/Val, Val/Met, and Met/Met mice; (b) bar graph comparing decrease in brdendrites; oblique branches were decreased in Met/Met mice only. *p � .05;Met allele mice (Z-stack projections of basal dendrites from Val/Val, Val/Met
of basal dendrites in Val/Met and Met/Met mice; (c) Sholl analyses of basal dendritin Val/Met and Met/Met mice between 20 and 100 �m from the soma.www.sobp.org/journal
rugs(�)-ketamine hydrochloride was from Hospira (Lake Forest, Illi-
ois). For the slice experiments, 5-hydroxytryptamine, creatinineulfate (5-HT) was from Sigma (St. Louis, Missouri) and hypocretin 2orexin B) was from American Peptide (Sunnyvale, California). Bothransmitters were perfused at known concentrations through atopcock assembly.
ophic factor Val66Met knock-in mice have both apical and basilar dendriticowing atrophy of apical tuft dendrites from two-layer V pyramidal cells in
length of total apical dendrites (i.e., tuft � oblique), apical tuft, and oblique.01. (B) Basilar dendrites: (a) images illustrating basilar dendritic atrophy in
Met/Met mice); (b) bar graph showing a significant decrease in total length
urotrns sh
anch**p �, and
ic structure revealing a significant decrease in basilar dendrite intersections
stt
l
�Ss .06 ��
R.-J. Liu et al. BIOL PSYCHIATRY 2012;71:996–1005 999
Analysis and StatisticsElectrophysiological data were displayed offline with Clampfit
software of pClamp 9.2 (Axon Instruments). Analysis of EPSCs from
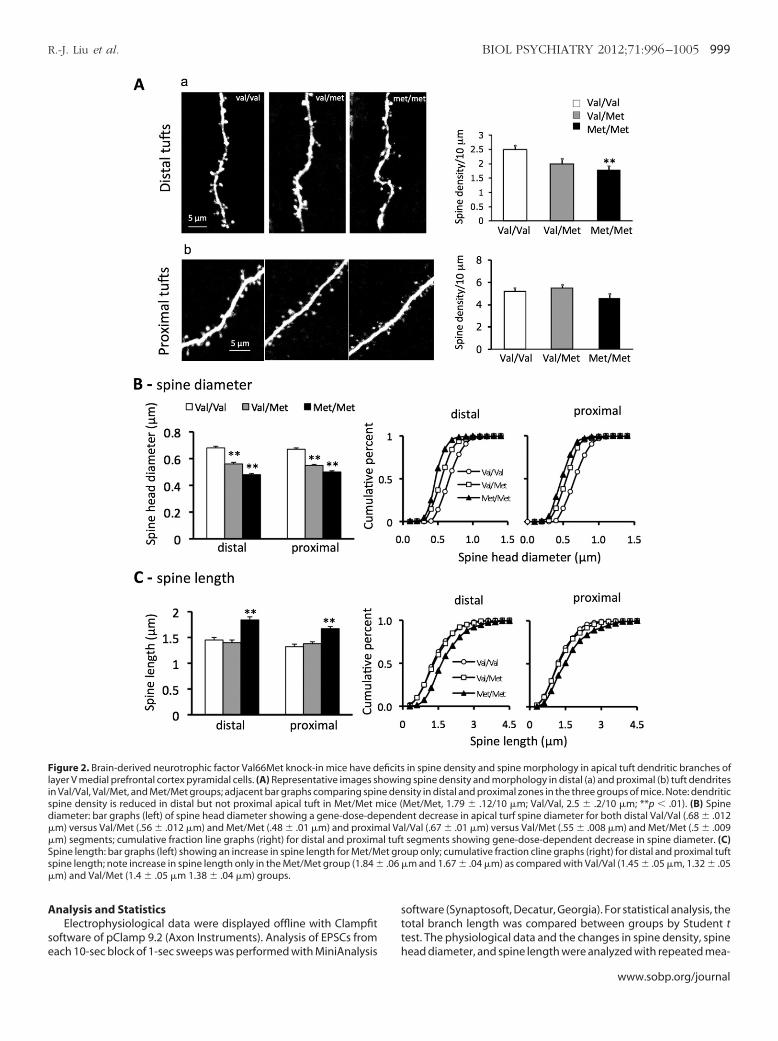
Figure 2. Brain-derived neurotrophic factor Val66Met knock-in mice have dayer V medial prefrontal cortex pyramidal cells. (A) Representative images sh
in Val/Val, Val/Met, and Met/Met groups; adjacent bar graphs comparing spinspine density is reduced in distal but not proximal apical tuft in Met/Met mdiameter: bar graphs (left) of spine head diameter showing a gene-dose-de�m) versus Val/Met (.56 � .012 �m) and Met/Met (.48 � .01 �m) and proxim
m) segments; cumulative fraction line graphs (right) for distal and proximpine length: bar graphs (left) showing an increase in spine length for Met/Mpine length; note increase in spine length only in the Met/Met group (1.84 �m) and Val/Met (1.4 � .05 �m 1.38 � .04 �m) groups.
each 10-sec block of 1-sec sweeps was performed with MiniAnalysis h
oftware (Synaptosoft, Decatur, Georgia). For statistical analysis, theotal branch length was compared between groups by Student test. The physiological data and the changes in spine density, spine
in spine density and spine morphology in apical tuft dendritic branches ofg spine density and morphology in distal (a) and proximal (b) tuft dendritessity in distal and proximal zones in the three groups of mice. Note: dendritic
Met/Met, 1.79 � .12/10 �m; Val/Val, 2.5 � .2/10 �m; **p � .01). (B) Spineent decrease in apical turf spine diameter for both distal Val/Val (.68 � .012l/Val (.67 � .01 �m) versus Val/Met (.55 � .008 �m) and Met/Met (.5 � .009segments showing gene-dose-dependent decrease in spine diameter. (C)up only; cumulative fraction cline graphs (right) for distal and proximal tuftm and 1.67 � .04 �m) as compared with Val/Val (1.45 � .05 �m, 1.32 � .05
eficitsowine denice (
pendal Va
al tuftet gro
ead diameter, and spine length were analyzed with repeated mea-
www.sobp.org/journal
wtplsa�sim1rsbmsb
AM
snmV(d.(imasrdnnt
MaFE
sih
aaMv(
7hmw
KM
rsaBeoptpd
Fnt(oVHwCaKn
1000 BIOL PSYCHIATRY 2012;71:996–1005 R.-J. Liu et al.
w
sures analysis of variance, after a post hoc comparison; values weregiven as mean � SEM. The EPSC amplitudes were analyzed with theKolmogorov-Smirnov test.
Results
BDNF Val66Met Knock-In Mice Have Both Apical and BasilarDendritic Atrophy of mPFC Layer V Pyramidal Cells
As in a previous study on stress-induced effects (17), we usedhole cell patch-clamp recording in combination with imaging by
wo-photon laser scanning to correlate morphological and electro-hysiological changes in the same cells. Compared with Val/Val WT
ittermate control mice, the Val/Met and Met/Met mice resembledtressed animals in that total branch length of apical dendrites andpical tufts decreased (WT, 4077 � 247 �m; Val/Met, 3264 � 204m; Met/Met, 2482 � 150 �m) (Figure 1A). However, in contrast to
tress, there was also a decrease in total length of basilar dendritesn both Val/Met (2382 � 225 �m) and Met/Met (2200 � 197 �m)
ice as compared with WT Val/Val mice (3006 �m � 178) (FigureB). Sholl analyses of basilar dendritic structure revealed a significanteduction in dendritic complexity between 20 �m to 100 �m from theoma in Val/Met and Met/Met mice (Figure 1C). These changes resem-le those described previously for Met/Met mice in layer II/III cells ofPFC (29). The reduction of basilar dendrites in Met allele mice corre-
ponds to the deficit in trafficking of BDNF mRNA from the soma toasilar as well as apical fields (12).
pical Tuft Dendrites of Layer V Cells of Val/Met and Met/Metice Show a Reduction in Spine Density and Maturation
As shown in Figure 2, distal but not proximal apical tuft dendriticpine density was significantly decreased in layer V mPFC pyramidaleurons in Met/Met mice compared with Val/Val littermate controlice (Met/Met, 1.79 � .12/10 �m; Val/Val, 2.5 � .2/10 �m; p � .01);
al/Met mice had a nonsignificant decrease in distal spine densityFigure 2A). In addition, spine head diameter showed a gene-dose-ependent decrease in both distal (Val/Val .68 � .012, Val/Met .56 �
012, and Met/Met 48 � .01) and proximal apical tuft dendritesFigures 2A and 2B). Spine length also showed a significant increasen both distal and proximal dendritic branches but only in Met/Met
ice (Figure 2A and 2C). Taken together, these changes resulted inshift from mushroom-like spines (�.75 �m) toward long thin
pines. These changes are similar to those found in pyramidal neu-ons of cortex and hippocampus in transgenic mice expressing aominant negative of the BDNF receptor tropomyosin-related ki-ase B (TrkB) (30). Conversely, conditional deletion of downstreamegative regulators of TrkB increases filopodial motility and synap-
ic formation (31).
orphological Changes in Layer V Pyramidal Cells in Val/Metnd Met/Met Mice are Associated with a Reduction in therequency and Amplitude of 5-HT- and Hypocretin-InducedPSCs
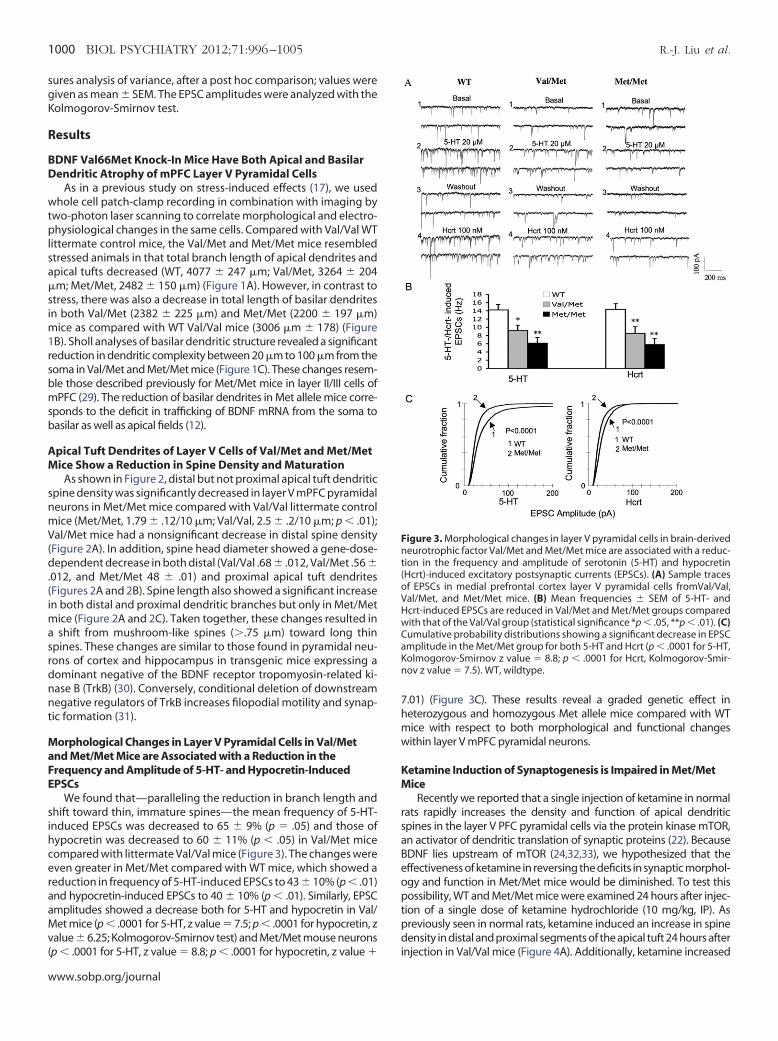
We found that—paralleling the reduction in branch length andhift toward thin, immature spines—the mean frequency of 5-HT-nduced EPSCs was decreased to 65 � 9% (p � .05) and those ofypocretin was decreased to 60 � 11% (p � .05) in Val/Met mice
compared with littermate Val/Val mice (Figure 3). The changes wereeven greater in Met/Met compared with WT mice, which showed areduction in frequency of 5-HT-induced EPSCs to 43 � 10% (p � .01)
nd hypocretin-induced EPSCs to 40 � 10% (p � .01). Similarly, EPSCmplitudes showed a decrease both for 5-HT and hypocretin in Val/et mice (p � .0001 for 5-HT, z value � 7.5; p � .0001 for hypocretin, z
alue � 6.25; Kolmogorov-Smirnov test) and Met/Met mouse neurons
p � .0001 for 5-HT, z value � 8.8; p � .0001 for hypocretin, z value � iww.sobp.org/journal
.01) (Figure 3C). These results reveal a graded genetic effect ineterozygous and homozygous Met allele mice compared with WTice with respect to both morphological and functional changesithin layer V mPFC pyramidal neurons.
etamine Induction of Synaptogenesis is Impaired in Met/Metice
Recently we reported that a single injection of ketamine in normalats rapidly increases the density and function of apical dendriticpines in the layer V PFC pyramidal cells via the protein kinase mTOR,n activator of dendritic translation of synaptic proteins (22). BecauseDNF lies upstream of mTOR (24,32,33), we hypothesized that theffectiveness of ketamine in reversing the deficits in synaptic morphol-gy and function in Met/Met mice would be diminished. To test thisossibility, WT and Met/Met mice were examined 24 hours after injec-
ion of a single dose of ketamine hydrochloride (10 mg/kg, IP). Asreviously seen in normal rats, ketamine induced an increase in spineensity in distal and proximal segments of the apical tuft 24 hours after
igure 3. Morphological changes in layer V pyramidal cells in brain-derivedeurotrophic factor Val/Met and Met/Met mice are associated with a reduc-
ion in the frequency and amplitude of serotonin (5-HT) and hypocretinHcrt)-induced excitatory postsynaptic currents (EPSCs). (A) Sample tracesf EPSCs in medial prefrontal cortex layer V pyramidal cells fromVal/Val,al/Met, and Met/Met mice. (B) Mean frequencies � SEM of 5-HT- andcrt-induced EPSCs are reduced in Val/Met and Met/Met groups comparedith that of the Val/Val group (statistical significance *p � .05, **p � .01). (C)umulative probability distributions showing a significant decrease in EPSCmplitude in the Met/Met group for both 5-HT and Hcrt (p � .0001 for 5-HT,olmogorov-Smirnov z value � 8.8; p � .0001 for Hcrt, Kolmogorov-Smir-ov z value � 7.5). WT, wildtype.
njection in Val/Val mice (Figure 4A). Additionally, ketamine increased
MdfpMiEom
AM
a
pbeodmiMmneei.
M . n.s.,
R.-J. Liu et al. BIOL PSYCHIATRY 2012;71:996–1005 1001
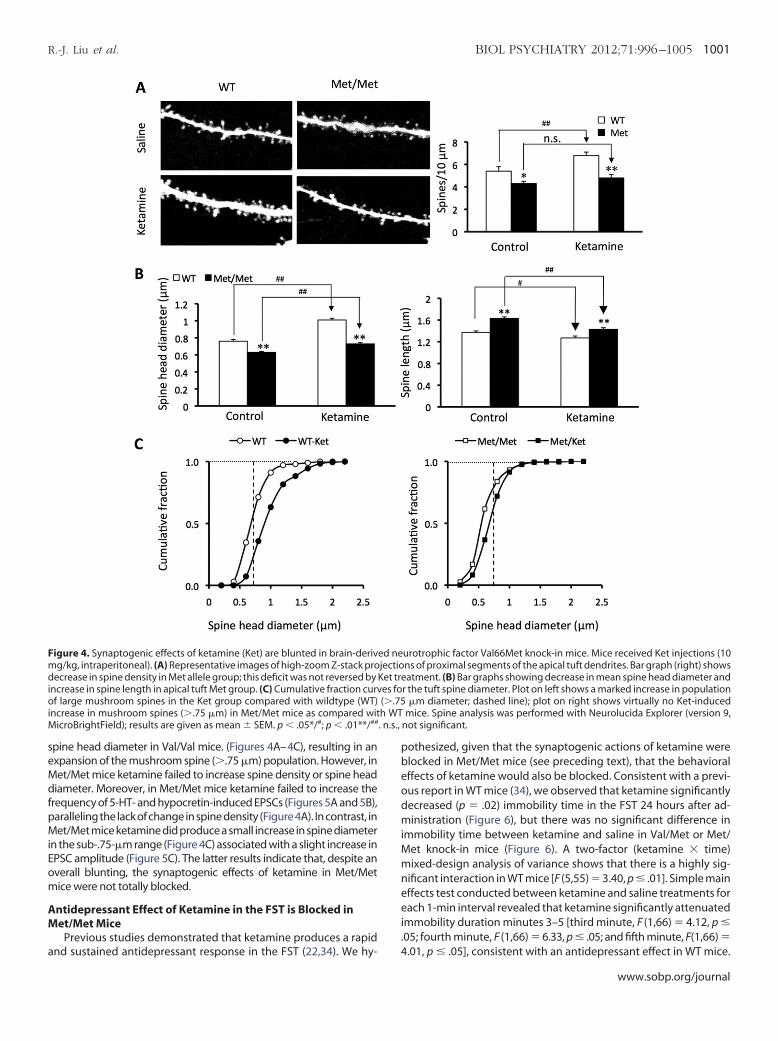
spine head diameter in Val/Val mice. (Figures 4A–4C), resulting in anexpansion of the mushroom spine (�.75 �m) population. However, in
et/Met mice ketamine failed to increase spine density or spine headiameter. Moreover, in Met/Met mice ketamine failed to increase the
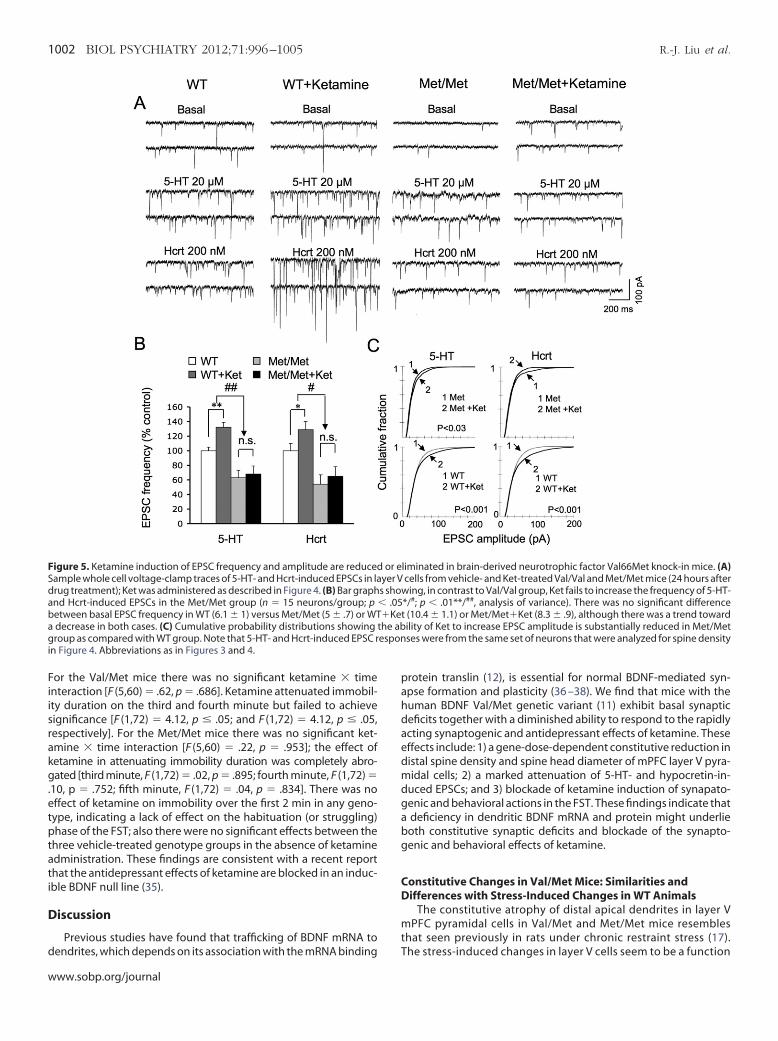
requency of 5-HT- and hypocretin-induced EPSCs (Figures 5A and 5B),aralleling the lack of change in spine density (Figure 4A). In contrast, inet/Met mice ketamine did produce a small increase in spine diameter
n the sub-.75-�m range (Figure 4C) associated with a slight increase inPSC amplitude (Figure 5C). The latter results indicate that, despite anverall blunting, the synaptogenic effects of ketamine in Met/Metice were not totally blocked.
ntidepressant Effect of Ketamine in the FST is Blocked inet/Met Mice
Previous studies demonstrated that ketamine produces a rapid
Figure 4. Synaptogenic effects of ketamine (Ket) are blunted in brain-derivmg/kg, intraperitoneal). (A) Representative images of high-zoom Z-stack prodecrease in spine density in Met allele group; this deficit was not reversed byincrease in spine length in apical tuft Met group. (C) Cumulative fraction curvof large mushroom spines in the Ket group compared with wildtype (WT)increase in mushroom spines (�.75 �m) in Met/Met mice as compared wit
icroBrightField); results are given as mean � SEM. p � .05*/#; p � .01**/##
nd sustained antidepressant response in the FST (22,34). We hy- 4
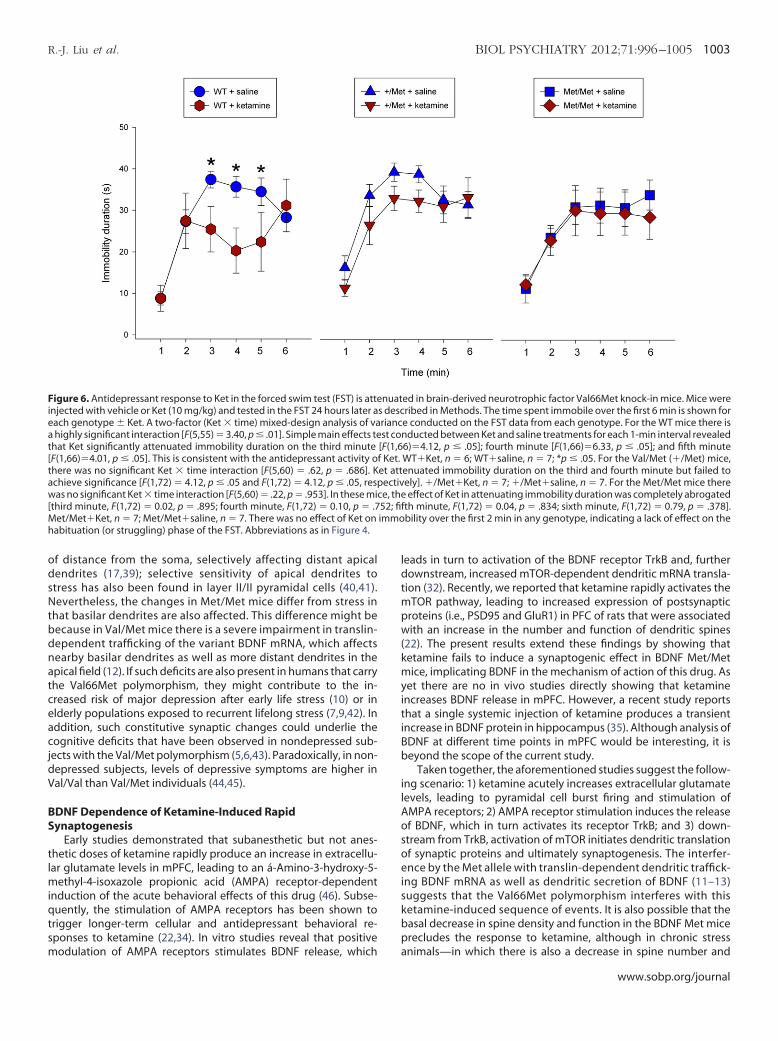
othesized, given that the synaptogenic actions of ketamine werelocked in Met/Met mice (see preceding text), that the behavioralffects of ketamine would also be blocked. Consistent with a previ-us report in WT mice (34), we observed that ketamine significantlyecreased (p � .02) immobility time in the FST 24 hours after ad-inistration (Figure 6), but there was no significant difference in
mmobility time between ketamine and saline in Val/Met or Met/et knock-in mice (Figure 6). A two-factor (ketamine � time)ixed-design analysis of variance shows that there is a highly sig-
ificant interaction in WT mice [F (5,55) � 3.40, p � .01]. Simple mainffects test conducted between ketamine and saline treatments forach 1-min interval revealed that ketamine significantly attenuated
mmobility duration minutes 3–5 [third minute, F (1,66) � 4.12, p �05; fourth minute, F (1,66) � 6.33, p � .05; and fifth minute, F(1,66) �
urotrophic factor Val66Met knock-in mice. Mice received Ket injections (10ns of proximal segments of the apical tuft dendrites. Bar graph (right) showsatment. (B) Bar graphs showing decrease in mean spine head diameter and
r the tuft spine diameter. Plot on left shows a marked increase in population�m diameter; dashed line); plot on right shows virtually no Ket-induced
mice. Spine analysis was performed with Neurolucida Explorer (version 9,not significant.
ed nejectioKet trees fo(�.75h WT
.01, p � .05], consistent with an antidepressant effect in WT mice.
www.sobp.org/journal
i
rakg.etptati
pahdaedmdgabg
CD
mt
Sda
agi
1002 BIOL PSYCHIATRY 2012;71:996–1005 R.-J. Liu et al.
w
For the Val/Met mice there was no significant ketamine � timenteraction [F (5,60) � .62, p � .686]. Ketamine attenuated immobil-
ity duration on the third and fourth minute but failed to achievesignificance [F (1,72) � 4.12, p � .05; and F (1,72) � 4.12, p � .05,espectively]. For the Met/Met mice there was no significant ket-mine � time interaction [F (5,60) � .22, p � .953]; the effect ofetamine in attenuating immobility duration was completely abro-ated [third minute, F (1,72) � .02, p � .895; fourth minute, F (1,72) �
10, p � .752; fifth minute, F (1,72) � .04, p � .834]. There was noffect of ketamine on immobility over the first 2 min in any geno-ype, indicating a lack of effect on the habituation (or struggling)hase of the FST; also there were no significant effects between the
hree vehicle-treated genotype groups in the absence of ketaminedministration. These findings are consistent with a recent reporthat the antidepressant effects of ketamine are blocked in an induc-ble BDNF null line (35).
Discussion
Previous studies have found that trafficking of BDNF mRNA to
Figure 5. Ketamine induction of EPSC frequency and amplitude are reducedample whole cell voltage-clamp traces of 5-HT- and Hcrt-induced EPSCs in larug treatment); Ket was administered as described in Figure 4. (B) Bar graphnd Hcrt-induced EPSCs in the Met/Met group (n � 15 neurons/group; p
between basal EPSC frequency in WT (6.1 � 1) versus Met/Met (5 � .7) or WTdecrease in both cases. (C) Cumulative probability distributions showing troup as compared with WT group. Note that 5-HT- and Hcrt-induced EPSC r
n Figure 4. Abbreviations as in Figures 3 and 4.
dendrites, which depends on its association with the mRNA binding T
ww.sobp.org/journal
rotein translin (12), is essential for normal BDNF-mediated syn-pse formation and plasticity (36 –38). We find that mice with theuman BDNF Val/Met genetic variant (11) exhibit basal synapticeficits together with a diminished ability to respond to the rapidlycting synaptogenic and antidepressant effects of ketamine. Theseffects include: 1) a gene-dose-dependent constitutive reduction inistal spine density and spine head diameter of mPFC layer V pyra-idal cells; 2) a marked attenuation of 5-HT- and hypocretin-in-
uced EPSCs; and 3) blockade of ketamine induction of synapato-enic and behavioral actions in the FST. These findings indicate thatdeficiency in dendritic BDNF mRNA and protein might underlieoth constitutive synaptic deficits and blockade of the synapto-enic and behavioral effects of ketamine.
onstitutive Changes in Val/Met Mice: Similarities andifferences with Stress-Induced Changes in WT Animals
The constitutive atrophy of distal apical dendrites in layer VPFC pyramidal cells in Val/Met and Met/Met mice resembles
hat seen previously in rats under chronic restraint stress (17).
liminated in brain-derived neurotrophic factor Val66Met knock-in mice. (A)cells from vehicle- and Ket-treated Val/Val and Met/Met mice (24 hours after
ing, in contrast to Val/Val group, Ket fails to increase the frequency of 5-HT-*/#; p � .01**/##, analysis of variance). There was no significant difference(10.4 � 1.1) or Met/Met�Ket (8.3 � .9), although there was a trend towardility of Ket to increase EPSC amplitude is substantially reduced in Met/Metses were from the same set of neurons that were analyzed for spine density
or eyer Vs show� .05�Kethe ab
espon
he stress-induced changes in layer V cells seem to be a function
Ntbdnatc
ldtmpw(kmyitiBb
ilAosoeiskbp
t[
a
[immo
h
R.-J. Liu et al. BIOL PSYCHIATRY 2012;71:996–1005 1003
of distance from the soma, selectively affecting distant apicaldendrites (17,39); selective sensitivity of apical dendrites tostress has also been found in layer II/II pyramidal cells (40,41).
evertheless, the changes in Met/Met mice differ from stress inhat basilar dendrites are also affected. This difference might beecause in Val/Met mice there is a severe impairment in translin-ependent trafficking of the variant BDNF mRNA, which affectsearby basilar dendrites as well as more distant dendrites in thepical field (12). If such deficits are also present in humans that carryhe Val66Met polymorphism, they might contribute to the in-reased risk of major depression after early life stress (10) or in
elderly populations exposed to recurrent lifelong stress (7,9,42). Inaddition, such constitutive synaptic changes could underlie thecognitive deficits that have been observed in nondepressed sub-jects with the Val/Met polymorphism (5,6,43). Paradoxically, in non-depressed subjects, levels of depressive symptoms are higher inVal/Val than Val/Met individuals (44,45).
BDNF Dependence of Ketamine-Induced RapidSynaptogenesis
Early studies demonstrated that subanesthetic but not anes-thetic doses of ketamine rapidly produce an increase in extracellu-lar glutamate levels in mPFC, leading to an á-Amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptor-dependentinduction of the acute behavioral effects of this drug (46). Subse-quently, the stimulation of AMPA receptors has been shown totrigger longer-term cellular and antidepressant behavioral re-sponses to ketamine (22,34). In vitro studies reveal that positive
Figure 6. Antidepressant response to Ket in the forced swim test (FST) is atteinjected with vehicle or Ket (10 mg/kg) and tested in the FST 24 hours later aeach genotype � Ket. A two-factor (Ket � time) mixed-design analysis of vaa highly significant interaction [F(5,55) � 3.40, p � .01]. Simple main effects tehat Ket significantly attenuated immobility duration on the third minuteF(1,66)�4.01, p � .05]. This is consistent with the antidepressant activity o
there was no significant Ket � time interaction [F(5,60) � .62, p � .686]. Kchieve significance [F(1,72) � 4.12, p � .05 and F(1,72) � 4.12, p � .05, resp
was no significant Ket � time interaction [F(5,60) � .22, p � .953]. In these micthird minute, F(1,72) � 0.02, p � .895; fourth minute, F(1,72) � 0.10, p � .7
Met/Met�Ket, n � 7; Met/Met�saline, n � 7. There was no effect of Ket onabituation (or struggling) phase of the FST. Abbreviations as in Figure 4.
modulation of AMPA receptors stimulates BDNF release, which a
eads in turn to activation of the BDNF receptor TrkB and, furtherownstream, increased mTOR-dependent dendritic mRNA transla-
ion (32). Recently, we reported that ketamine rapidly activates theTOR pathway, leading to increased expression of postsynaptic
roteins (i.e., PSD95 and GluR1) in PFC of rats that were associatedith an increase in the number and function of dendritic spines
22). The present results extend these findings by showing thatetamine fails to induce a synaptogenic effect in BDNF Met/Metice, implicating BDNF in the mechanism of action of this drug. As
et there are no in vivo studies directly showing that ketaminencreases BDNF release in mPFC. However, a recent study reportshat a single systemic injection of ketamine produces a transientncrease in BDNF protein in hippocampus (35). Although analysis ofDNF at different time points in mPFC would be interesting, it iseyond the scope of the current study.
Taken together, the aforementioned studies suggest the follow-ng scenario: 1) ketamine acutely increases extracellular glutamateevels, leading to pyramidal cell burst firing and stimulation ofMPA receptors; 2) AMPA receptor stimulation induces the releasef BDNF, which in turn activates its receptor TrkB; and 3) down-tream from TrkB, activation of mTOR initiates dendritic translationf synaptic proteins and ultimately synaptogenesis. The interfer-nce by the Met allele with translin-dependent dendritic traffick-
ng BDNF mRNA as well as dendritic secretion of BDNF (11–13)uggests that the Val66Met polymorphism interferes with thisetamine-induced sequence of events. It is also possible that theasal decrease in spine density and function in the BDNF Met micerecludes the response to ketamine, although in chronic stress
ed in brain-derived neurotrophic factor Val66Met knock-in mice. Mice wereribed in Methods. The time spent immobile over the first 6 min is shown fore conducted on the FST data from each genotype. For the WT mice there is
ducted between Ket and saline treatments for each 1-min interval revealed6)�4.12, p � .05]; fourth minute [F(1,66)�6.33, p � .05]; and fifth minuteWT�Ket, n � 6; WT�saline, n � 7; *p � .05. For the Val/Met (�/Met) mice,enuated immobility duration on the third and fourth minute but failed toely]. �/Met�Ket, n � 7; �/Met�saline, n � 7. For the Met/Met mice thereeffect of Ket in attenuating immobility duration was completely abrogated
th minute, F(1,72) � 0.04, p � .834; sixth minute, F(1,72) � 0.79, p � .378].bility over the first 2 min in any genotype, indicating a lack of effect on the
nuats descriancst con
[F(1,6f Ket.et attective, the52; fif
nimals—in which there is also a decrease in spine number and
www.sobp.org/journal

C
tdtu(cpjcfBipclcc
miaom
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
1004 BIOL PSYCHIATRY 2012;71:996–1005 R.-J. Liu et al.
w
function— ketamine still induces synaptogenic and antidepressantbehavioral responses (Li et al. [22]).
linical ImplicationsThe BDNF Val66Met polymorphism represents the first altera-
ion in a neurotrophin gene linked to clinical pathology in affectiveisorders. Thus, the cognitive impairments in individuals carrying
he Met polymorphism (5) are associated with a reduction in vol-me of various brain regions including the PFC and hippocampus
47–50). The reductions in PFC gray matter volumes were signifi-antly greater in the Met/Met compared with the Val/Met bipolaratients, correlating with relative memory deficits in the same sub-
ects (50). A graded reduction in gray matter volume in humansorresponds to the gene-dose-response relationship we observedor loss of dendritic branches and spine density in layer V cells of theDNF Val/Met gene knock-in mice. These synaptic deficits have
mplications for both intracortical and subcortical connectivity. Inarticular, the layer V pyramidal cells of mPFC give rise to majororticolimbic projection pathways innervating regions such as the
ocus coeruleus, dorsal raphe nucleus, ventral tegmental area, nu-leus accumbens, and amygdala (51) that are within circuits impli-ated in mood disorders (15,16,52).
The therapeutic implications of the failure of BDNF Met alleleice to respond to the synaptogenic effect of ketamine are also of
nterest. A single subanesthetic dose of ketamine produces a rapidnd relatively long-lasting antidepressant response in a majority oftherwise treatment-resistant patients (approximately 70%) withajor depression or bipolar depression (53–55). Moreover, a single
dose of ketamine is effective in rapidly resolving suicidal ideation inemergency room situations (56,57). However, the reason for thelack of response in the remaining approximately 30% of patients isnot clear. Recently, it was reported that mice with an inducibleknockout of the BDNF gene fail to show the expected antidepres-sant-like behavioral responses to ketamine (35). The present studyconfirms that a similar resistance to ketamine occurs in BDNF Met/Met mice. As yet it is not known whether patients homologous forthe Met polymorphism are also resistant to ketamine. Not only doesa meta-analysis of clinical studies with “standard” antidepressants(e.g., selective serotonin reuptake inhibitors) not show an overallnegative influence of the Met allele on outcome—there are somestudies reporting an enhanced treatment response (58). Perhapsthis can be explained by the finding in mouse models that thesedrugs transactivate TrkB independently of BDNF (59). It remains tobe determined whether the approximately 30% of patients that areresistant to the rapid antidepressant effect of ketamine includedisproportionally those who carry the loss-of-function BDNF Metallele. If so, it might be possible to bypass the BDNF deficit bytreating nonresponders with BDNF-mimetic compounds that canact directly on TrkB to activate the mTOR synaptogenic pathway.
This work was supported by National Institutes of Health GrantsMH17871 and MH093897 and the State of Connecticut.
The authors have no biomedical financial interests or potentialconflicts pertaining to this work.
1. Duman RS, Monteggia LM (2006): A neurotrophic model for stress-related mood disorders. Biol Psychiatry 59:1116 –1127.
2. Castren E, Voikar V, Rantamaki (2007): Role of neurotrophic factors indepression. Curr Opin Pharmacol 7:18 –21.
3. Martinowich K, Manji H, Lu B (2007): New insights into BDNF function indepression and anxiety. Nat Neurosci 10:1089 –1093.
4. Dwivedi Y (2009): Brain-derived neurotrophic factor: Role in depressionand suicide. Ann Med 42:87–96.
ww.sobp.org/journal
5. Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS, Bertolino A,et al. (2003): The BDNF val66met polymorphism affects activity-depen-dent secretion of BDNF and human memory and hippocampal function.Cell 112:257–269.
6. Hariri AR, Goldberg TE, Mattay VS, Kolachana BS, Callicott JH, Egan MF,Weinberger DR (2003): Brain-derived neurotrophic factor val66metpolymorphism affects human memory-related hippocampal activityand predicts memory performance. J Neurosci 23:6690 – 6694.
7. Harris SE, Fox H, Wright AF, Hayward C, Starr JM, Whalley LJ, Deary IJ(2006): The brain-derived neurotrophic factor Val66Met polymor-phism is associated with age-related change in reasoning skills. MolPsychiatry 11:505–513.
8. Kim J-M, Stewart R, Kim S-W, Yang S-J, Shin I-S, Kim Y-H, Yoon J-S (2007):Interactions between life stressors and susceptibility genes (5-HTTLPRand BDNF) on depression in Korean elders. Biol Psychiatry 62:423– 428.
9. Miyajima F, Ollier W, Mayes A, Jackson A, Thacker N, Rabbitt P, et al.(2008): Brain-derived neurotrophic factor polymorphism Val66Met in-fluences cognitive abilities in the elderly. Genes Brain Behav 7:411– 417.
0. Gatt JM, Nemeroff CB, Dobson-Stone C, Paul RH, Bryant RA, Schofield PR,et al. (2009): Interactions between BDNF Val66Met polymorphism andearly life stress predict brain and arousal pathways to syndromal de-pression and anxiety. Mol Psychiatry 14:681– 695.
1. Chen ZY, Patel PD, Sant G, Meng CX, Teng KK, Hempstead BL, Lee FS (2004):Variant brain-derived neurotrophic factor (BDNF) (Met66) alters the intra-cellular trafficking and activity-dependent secretion of WT BDNF in neuro-secretory cells and cortical neurons. J Neurosci 24:4401–4411.
2. Chiaruttini C, Vicario A, Li Z, Baj G, Braiuca P, Wu Y, et al. (2009): Dendritictrafficking of BDNF mRNA is mediated by translin and blocked by theG196A (Val66Met) mutation. Proc Natl Acad Sci U S A 106:16481–16486.
3. Chen ZY, Jing D, Bath KG, Ieraci A, Khan T, Siao CJ, et al. (2006): Geneticvariant BDNF (Val66Met) polymorphism alters anxiety-related behavior.Science 314:140 –143.
4. Drevets WC, Price JL, Furey ML (2008): Brain structural and functionalabnormalities in mood disorders: Implications for neurocircuitry mod-els of depression. Brain Struct Funct 213:93–118.
5. Mayberg HS (2009): Targeted electrode-based modulation of neuralcircuits for depression. J Clin Invest 119:717–725.
6. Krishnan V, Nestler EJ (2010): Linking molecules to mood: New insightinto the biology of depression. Am J Psychiatry 167:1305–1320.
7. Liu RJ, Aghajanian GK (2008): Stress blunts serotonin- and hypocretin-evoked EPSCs in prefrontal cortex: Role of corticosterone-mediatedapical dendritic atrophy. Proc Natl Acad Sci U S A 105:359 –364.
8. Aghajanian GK, Marek GJ (1997): Serotonin induces excitatory postsyn-aptic potentials in apical dendrites of neocortical pyramidal cells. Neu-ropharmacology 36:589 –599.
9. Lambe EK, Aghajanian GK (2003): Hypocretin (orexin) induces calciumtransients in single spines postsynaptic to identified thalamocorticalboutons in prefrontal slice. Neuron 40:139 –150.
0. Poo MM (2001): Neurotrophins as synaptic modulators. Nat Rev Neurosci2:24 –32.
1. Lu B (2004): Acute and long-term synaptic modulation by neurotro-phins. Prog Brain Res 146:137–150.
2. Li N, Lee B, Liu RJ, Banasr M, Dwyer JM, Iwata M, et al. (2010): mTOR-dependent synapse formation underlies the rapid antidepressant ef-fects of NMDA antagonists. Science 329:959 –964.
3. Hoeffer CA, Klann (2009): mTOR signaling: At the crossroads of plasticity,memory and disease. Trends Neurosci 33:67–75.
4. Takei N, Inamura N, Kawamura M, Namba H, Hara K, Yonezawa K, NawaH (2004): Brain-derived neurotrophic factor induces mammalian targetof rapamycin-dependent local activation of translation machinery andprotein synthesis in neuronal dendrites. J Neurosci 24:9760 –9769.
5. Porsolt RD, Bertin A, Jalfre M (1977): Behavioral despair in mice: A pri-mary screening test for antidepressants. Arch Int Pharmacodyn Ther229:327–336.
6. Hayashi E, Shimamura M, Kuratani K, Kinoshita M, Hara H (2011): Auto-mated experimental system capturing three behavioral componentsduring murine forced swim test. Life Sci 88:411– 417.
7. Kelliher P, Connor TJ, Harkin A, Sanchez C, Kelly JP, Leonard BE (2000):Varying responses to the rat forced-swim test under diurnal and noctur-nal conditions. Physiol Behav 69:531–539.
8. Kulikov AV, Morozova MV, Kulikov VA, Kirichuk VS, Popova NK (2010):
Automated analysis of antidepressants’ effect in the forced swim test.J Neurosci Methods 191:26 –31.
4
4
4
4
4
5
5
5
5
5
5
5
5
5
5
R.-J. Liu et al. BIOL PSYCHIATRY 2012;71:996–1005 1005
29. Yu H, Wang Y, Pattwell S, Jing D, Liu T, Zhang Y, et al. (2009): VariantBDNF Val66Met polymorphism affects extinction of conditioned aver-sive memory. Neurosci 29: 4056 – 4064.
30. Chakravarthy S, Saiepour MH, Bence M, Perry S, Hartman R, Couey JJ, etal. (2006): Postsynaptic TrkB signaling has distinct roles in spine main-tenance in adult visual cortex and hippocampus. Proc Natl Acad Sci U S A103:1071–1076.
31. Luikart BW, Zhang W, Wayman GA, Kwon CH, Westbrook GL, Parada LF(2008): Neurotrophin-dependent dendritic filopodial motility: a conver-gence on PI3K signaling. J Neurosci 28:7006 –7012.
32. Jourdi H, Hsu YT, Zhou M, Qin Q, Bi X, Baudry M (2009): Positive AMPAreceptor modulation rapidly stimulates BDNF release and increasesdendritic mRNA translation. J Neurosci 29:8688 – 8697.
33. Slipczuk L, Bekinschtein P, Katche C, Cammarota M, Izquierdo I, MedinaJH (2009): BDNF activates mTOR to regulate GluR1 expression requiredfor memory formation. PLoS One 4:e6007.
34. Maeng S, Zarate CA Jr, Du J, Schloesser RJ, McCammon J, Chen G, ManjiHK (2008): Cellular mechanisms underlying the antidepressant effectsof ketamine: role of alpha-amino-3-hydroxy-5-methylisoxazole-4-pro-pionic acid receptors. Biol Psychiatry 63:349 –352.
35. Autry AE, Adachi M, Nosyreva E, Na ES, Los MF, Cheng PF, et al. (2011):NMDA receptor blockade at rest triggers rapid behavioural antidepres-sant responses. Nature 475:91–95.
36. Tongiorgi E, Baj G (2008): Functions and mechanisms of BDNF mRNAtrafficking. Novartis Found Symp 289:136 –147.
37. An JJ, Gharami K, Liao GY, Woo NH, Lau AG, Vanevski F, et al. (2008):Distinct role of long 3’ UTR BDNF mRNA in spine morphology andsynaptic plasticity in hippocampal neurons. Cell 134:175–187.
38. Soulé J, Messaoudi E, Bramham CR (2006): Brain-derived neurotrophicfactor and control of synaptic consolidation in the adult brain. BiochemSoc Trans 34:600 – 604.
39. Goldwater DS, Pavlides C, Hunter RG, Bloss EB, Hof PR, McEwen BS,Morrison JH (2009): Structural and functional alterations to rat medialprefrontal cortex following chronic restraint stress and recovery. Neuro-science 164:798 – 808.
40. Brown SM, Henning S, Wellman CL (2005): Mild, short-term stress altersdendritic morphology in rat medial prefrontal cortex. Cereb Cortex 15:1714 –1722.
41. Radley JJ, Rocher AB, Miller M, Janssen WG, Liston C, Hof PR, et al. (2006):Repeated stress induces dendritic spine loss in the rat medial prefrontalcortex. Cereb Cortex16:313–320.
42. Nagel IE, Chicherio C, Li SC, von Oertzen T, Sander T, Villringer A, et al.(2008): Human aging magnifies genetic effects on executive function-ing and working memory. Front Hum Neurosci 2:1– 8.
43. Soliman F, Glatt CE, Bath KG, Levita L, Jones RM, Pattwell SS, et al. (2010):A genetic variant BDNF polymorphism alters extinction learning in bothmouse and human. Science 327:863– 866.
44. Hilt LM, Sander LC, Nolen-Hoeksema S, Simen AA (2007): The BDNFVal66Met polymorphism predicts rumination and depression differently in
young adolescent girls and their mothers. Neurosci Lett 429:12–16.5. Duncan LE, Hutchison KE, Carey G, Craighead WE (2009): Variation inbrain-derived neurotrophic factor (BDNF) gene is associated with symp-toms of depression. J Affect Disord 115:215–219.
6. Moghaddam B, Adams B, Verma A, Daly D (1997): Activation of glutama-tergic neurotransmission by ketamine: a novel step in the pathway fromNMDA receptor blockade to dopaminergic and cognitive disruptionsassociated with the prefrontal cortex. J Neurosci 17:2921–2927.
7. Drevets WC, Price JL, Simpson JR Jr, Todd RD, Reich T, Vannier M, RaichleME (1997): Subgenual prefrontal cortex abnormalities in mood disor-ders. Nature 386:824 – 827.
8. Pezawas L, Verchinski BA, Mattay VS, Callicott JH, Kolachana BS, StraubRE, et al. (2004): The brain-derived neurotrophic factor val66met poly-morphism and variation in human cortical morphology. J Neurosci 24:10099 –10102.
9. Frodl, Schule C, Schmitt G, Born C, Baghai T, Zill P, et al. (2007): Associa-tion of the brain-derived neurotrophic factor Val66Met polymorphismwith reduced hippocampal volumes in major depression. Arch Gen Psy-chiatry 64:410 – 416.
0. Matsuo K, Walss-Bass C, Nery FG, Nicoletti MA, Hatch JP, Frey BN, et al.(2009): Neuronal correlates of brain-derived neurotrophic factorVal66Met polymorphism and morphometric abnormalities in bipolardisorder. Neuropsychopharmacology 34:1904 –1913.
1. Gabbott PL, Warner TA, Jays PR, Salway P, Busby SJ (2005): Prefrontalcortex in the rat: Projections to subcortical autonomic, motor, and lim-bic centers. J Comp Neurol 492:145–177.
2. Savitz J, Drevets WC (2009): Bipolar and major depressive disorder:Neuroimaging the developmental-degenerative divide. Neurosci Biobe-hav Rev 33:699 –771.
3. Berman RM, Cappiello A, Anand A, Oren DA, Heninger GR, Charney DS,Krystal JH (2000): Antidepressant effects of ketamine in depressed pa-tients. Biol Psychiatry 47:351–354.
4. Zarate CA Jr, Singh JB, Carlson PJ, Brutsche NE, Ameli R, Luckenbaugh DA, etal. (2006): A randomized trial of an N-methyl-D-aspartate antagonist intreatment-resistant major depression. Arch Gen Psychiatry 63:856–864.
5. Diazgranados N, Ibrahim L, Brutsche NE, Newberg A, Kronstein P, KhalifeS, et al. (2010a): A randomized add-on trial of an N-methyl-D-aspartateantagonist in treatment-resistant bipolar depression. Arch Gen Psychia-try 67:793– 802.
6. Price RB, Nock MK, Charney DS, Mathew SJ (2009): Effects of intravenousketamine on explicit and implicit measures of suicidality in treatment-resistant depression. Biol Psychiatry 66:522–526.
7. Diazgranados N, Ibrahim LA, Brutsche NE, Ameli R, Henter ID, LuckenbaughDA, et al. (2010): Rapid resolution of suicidal ideation after a single infusionof an N-methyl-D-aspartate antagonist in patients with treatment-resistantmajor depressive disorder. J Clin Psychiatry 71:1605–1611.
8. Tsai1 S-J, Hong C-J, Liou Y-J (2010): Effects of BDNF polymorphisms onantidepressant action. Psychiatry Investig 7:236 –242.
9. Rantamäki T, Vesa L, Antila H, Di Lieto A, Tammela P, Schmitt A, et al.(2011): Antidepressant drugs transactivate TrkB neurotrophin receptors
in the adult rodent brain independently of BDNF and monoamine trans-porter blockade. PLoS One 6:e20567.www.sobp.org/journal







