dose-response relationships for hepatic aflatoxin b1-dna adduct formation in the rat in vivo and in...
TRANSCRIPT

Teratogenesis, Carcinogenesis, and Mutagenesis 19:157–170 (1999)
Published 1999 Wiley-Liss, Inc.
Dose-Response Relationships for HepaticAflatoxin B1-DNA Adduct Formation in theRat In Vivo and In Vitro: The Use ofImmunoslot Blotting for Adduct Quantitation
J.C. Phillips,* S. Davies, and B.G. Lake
BIBRA International, Woodmansterne Rd, Carshalton, Surrey, United Kingdom
An immunoslot blotting (ISB) method for quantitating aflatoxin B1-DNA adduct lev-els has been developed and used to examine the relationship between dose and he-patic aflatoxin B1-DNA adduct levels in rats fed aflatoxin B1 (AFB1) in the diet atdose levels of between 0.5 and 10 µg/kg/day. The method has also been used toexamine the dose-response relationship for adduct formation in precision-cut rat liverslices incubated with AFB1 at concentrations between 0.01 and 2 µM.
For the feeding studies, groups of male Fisher F344 rats were given AFB1 in thediet for periods of 1 to 10 weeks and hepatic DNA adduct levels determined usingISB. The time for adduct levels to reach steady-state conditions was determined inanimals given approximately 10 µg of AFB1/kg/day and steady-state levels at lowerconcentrations measured.
The time course for the accumulation of AFB1-DNA adducts in rat liver slicesincubated with AFB1 at 0.5 µM has been investigated and the relationship betweenadduct formation and AFB1 concentration over a wide concentration range in liverslices has been determined. Teratogenesis Carcinog. Mutagen. 19:157–170,1999. Published 1999 Wiley-Liss, Inc.
Key words: aflatoxin; rat liver; immunoslot blotting; adduct formation
INTRODUCTION
Current methods for assessing the risk to humans of exposure to hazardous sub-stances in the environment are based on the extrapolation of results from animalexperiments. Such studies make the assumption that the mechanism of action of thecompound is essentially similar in both species and that data obtained in animals at
Contract grant sponsor: UK Ministry of Agriculture, Fisheries and Food.
*Correspondence to: J.C. Phillips, BIBRA International, Woodmansterne Rd, Carshalton, Surrey SM54DS, United Kingdom.

158 Phillips et al.
dose levels necessary to obtain reasonable statistical precision for the quantitation ofthe end-point measured are relevant to human exposure at (usually) much lower doselevels. Thus for assessing the likely carcinogenic potential of a foreign chemical, thedose-response relationship for tumour incidence over an accessible dose range maybe extrapolated to likely human exposure levels without any knowledge of the actualdose-response relationship at the lower doses [1].
Recent developments in toxicology however have the potential to allow betteruse to be made of animal bioassay data and to allow data to be generated that ismore directly relevant to likely human exposure levels. Thus the development ofsensitive methods for quantifying the interaction of an active chemical species withthe molecular target can provide both a more relevant end point for dose responsestudies and may allow measurements at substantially lower dose levels. In additionthe availability of metabolically-active human tissue, such as liver, allows data di-rectly relevant to the human situation to be obtained so that relevant cross-speciesextrapolations can be made.
One group of natural toxicant for which human risk assessment has becomeincreasingly important is the mycotoxins. Exposure to mycotoxins, particularly afla-toxins through consumption of foodstuffs contaminated with Aspergillus species, hasbeen implicated in the high incidence of liver cancer various parts of the world [2].Aflatoxin B1, the most abundant form of the aflatoxins present in contaminated food,is a known genotoxic carcinogen requiring metabolic activation to exert its biologi-cal effect. The critical event for aflatoxin B1 carcinogenesis in sensitive species isthe interaction of the proximate carcinogenic species, the highly reactive electrophileaflatoxin B1 8,9-epoxide, with DNA resulting in the formation of covalently boundadducts [3]. This epoxide, generated by the metabolic activation of AFB1 by hepaticcytochrome P-450 isoforms [4], reacts almost exclusively with DNA at the N7 posi-tion of guanine to form initially the 8,9-dihydro-8(N7-guanyl)-9-hydroxyaflatoxin B1(AFB1-N
7-Gua) adduct [5]. This adduct has been reported to constitute at least 90%of the DNA-bound material in rodent liver in vivo [6]. With time this adduct isconverted to a more stable, imidazole-ring opened form, 8,9-dihydro-8(N5-2′,5′,6′-triamino-4′-oxo-N5-pyrimidyl)9-hydroxyaflatoxin B1 (AFB1-FAPy) [7].
DNA adducts of aflatoxin B1 have been detected in the livers of a number ofspecies sensitive to aflatoxin B1 carcinogenicity both after acute treatment and insome instances after chronic exposure. In a recent analysis of the data from rodents,Choy [8] noted that for acute studies in rats, an essentially linear dose-response rela-tionship between adduct formation and aflatoxin B1 dose could be obtained overdose levels ranging from ng/kg to mg/kg. Similarly, a linear relationship could bederived between adduct levels from the acute studies and tumour incidence fromchronic feeding studies. In only one study has there been an attempt to determinedose response relationships for adduct formation following chronic aflatoxin B1 ex-posure [9]. In this study a linear relationship between adduct levels and dose wasobtained in rats given aflatoxin B1 in the drinking water for up to 10 weeks at doselevels ranging from approximately 2 ng to 2 µg/kg/day.
Estimates of likely human exposure to aflatoxin B1 vary substantially, depend-ing on the selected population at risk. The average daily intakes in the southeasternand northwestern United States have been calculated to be 0.11 and 0.0002 µg/kg/day respectively [10] and approximately 1–2 µg/kg/day for the residents of GuangxiRegion in China, an area with a high prevalence of liver cancer [11].

Hepatic Aflatoxin B1-DNA Adduct Formation 159
Relatively few chronic bioassays with a range of aflatoxin concentrations havebeen carried out [12] that would allow dose-response relationships for tumour inci-dence to be investigated. However, by using data from the rat [13,14] and the trout[15–17], an approximately linear relationship has been obtained over a dose range of1 to 65 ppb [18].
In this study we have examined the relationship between AFB1 exposure and he-patic DNA adduct formation in rodent systems over a wide dose/concentration range.
METHODSChemicals, Enzymes, and Antibodies
Aflatoxin B1, Thimerasol, calf thymus DNA, and enzymes used for the purifi-cation of adducted DNA (ribonucleases, proteinase K) and 32P post-labelling (micro-coccal nuclease, spleen and snake venom phosphodiesterases, nuclease P1, prostaticacid and calf intestine alkaline phosphatases) were obtained from Sigma ChemicalCo., Poole, UK. Anti-aflatoxin B1 primary monoclonal antibody 6E9 was obtainedfrom Rhone Poulenc Diagnostics, Glasgow, UK, and monoclonal antibody 6A10 wasthe generous gift of Dr. R.Santella, Columbia University, New York, NY. For theimmunoslotblotting (ISB) assays, biotinylated rabbit anti-mouse IgG complex (DakoA/S, Denmark) was used as the secondary antibody. Streptavidin-horseradish per-oxidase complex and ECL reagent were obtained from Amersham International,Buckinghamshire, UK and SuperSignal reagent from Pierce Chemical Co., Rock-ford, IL. All other reagents used were SLR grade or better. [γ-32P]ATP (specific ac-tivity 3,000 Ci/mmol) was obtained from DuPont NEN, Herts, UK. 32P-post labelleddigests were chromatogarphed on Machery-Nagel polygram CEL300PEI thin-layerplates supplied by Altech Associates, Lancs, UK. ISB assays were carried out usingnitrocellulose paper (0.45µm) obtained from Bio-Rad, Inc., Richmond, CA.
Adducted Standards
Aflatoxin B1-adducted DNA was synthesised using the m-chloroperbenzoic acidmethod [19,20]. The aflatoxin-adducted DNA was converted to the stable ring-openedadduct (iro-AFB1 DNA) by treatment with sodium carbonate/sodium bicarbonate.The level of adduction was determined by uv spectroscopy using the absorption at260 nm (DNA concentration) and 360 nm (aflatoxin concentration) [20].
Animals
For the in vivo studies, young male Fisher F344 rats (body weight 120–140 g),obtained from Harlan UK, Ltd., Bicester, Oxford were used. Animals were grouphoused in a positive pressure isolator and allowed free access to food (R & M No.1maintenance diet; Special Diet Services, Witham, Essex) and water.
For in vitro studies, young male Sprague-Dawley rats, obtained from Harlan UK,Ltd., were used and allowed free access to food and water. Animals were group housedin the animal unit until required. All animals were housed in air-conditioned rooms at atemperature of 21 ± 2°C and humidity 55 ± 10% with a 12 h light-dark cycle.
In Vivo Studies
Groups of four Fisher F344 rats were given aflatoxin B1 admixed with the dietto provide an intake of 0.5, 2 or 10 µg/kg/day. Untreated control groups were fed

160 Phillips et al.
diet only. Animal were killed at various times up to 10 weeks of treatment and theliver removed for extraction of adducted DNA as described below.
In Vitro Studies: Preparation and Culture of Liver Slices
Tissue slices (diameter 10 mm; thickness 200–300 µm) were prepared by usinga Krumdieck tissue slicer (Alabama Research and Development Corp, AL) and cul-tured in RPMI 1640 medium [21]. Liver slice cultures were maintained at 37oC in anatmosphere of 95% O2/ 5% CO2. After 1 h, the medium was replaced with freshmedium containing the required concentration of aflatoxin B1 dissolved in DMSO (4µl/ml medium). At the end of the incubation periods (2, 4, or 6 h), liver slices werewashed in 0.154 M KCl containing 50 mM Tris-HCl pH 7.4 and stored at –80°Cprior to extraction of DNA.
Recovery of DNA
Adducted DNA from liver samples was recovered and purified using a modi-fied phenol extraction method [22,23]. Samples of liver (0.5 g) or liver slices (12;approximately 0.3–0.4 g) were extracted and the recovered DNA dissolved in 20 mMsodium succinate/10 mM CaCl2 buffer pH 6.0. The concentration and purity was as-sessed by uv spectroscopy at 260 and 280 nm.
32P-Post Labelling
Prior to labelling with γ-32P ATP, aflatoxin-adducted calf thymus DNA was hy-drolysed by using either spleen phosphodiesterase/micrococcal nuclease [24], nu-clease P1 and prostatic acid phosphatase [25] or snake venom phosphodiesterase andcalf alkaline phosphatase [26]. Labelled digests were chromatographed on PEI-cel-lulose thin layer plates in two directions [27] and the maps visualised by autoradiog-raphy (Fuji RX Medical X-ray film).
Immunoslot Blotting
Adducted DNA samples were denatured by incubation with 1 vol 1 M NaOH and1 vol 0.5 M NaCl for 10 min at room temperature followed by 2 vol 2 M ammoniumacetate solution and then kept on ice until used. Samples were diluted to the requiredconcentration with distilled water. The membrane used, BioRad transblot nitrocellulosepaper,was soaked in 6 × SSC and mounted in a “Hybri-slot manifold” (Bethesda Re-search Laboratories, Gaithersburg, MD) supported by two similarly soaked Whatman3 MM filter pads. The membrane was carefully dehydrated under vacuum, rehydratedwith distilled water, and the DNA samples applied in a total volume of 200 µl. The slotswere washed with 2 × SSC (3 M saline: 0.3 M sodium citrate) and the membrane al-lowed to dry in air before being baked at 80°C for 2 h. Non-specific binding sites wereblocked by incubation overnight at 4°C with 2.5% casein in 0.9% NaCl/ 0.121% trisbase (w/v) solution containing Thimerasol (0.2 g/l). The membrane was developed byincubation with an appropriate dilution of primary antibody (6E9 or 6A10) for 2 h on arolling mill, washed with detergent buffer (wash buffer : 20% SDS : Triton X-100–200:1:1v/v) and wash buffer (0.5% casein in 0.9% NaCl/0.121% tris base), and incu-bated with the secondary antibody (biotinylated rabbit anti-mouse IgG) for 2 h. Themembrane was then washed extensively with wash buffer and TBS-Tween (1.15% NaCl/0.05% tris base/0.1% Tween-20) and finally incubated with streptavidin-horseradish per-oxidase complex in TBS-Tween for 20 min. The membrane was washed with TBS-Tween

Hepatic Aflatoxin B1-DNA Adduct Formation 161
and developed with ECL reagent using Hyperfilm-ECL film. Adduct levels were quanti-tated by scanning densitometry.
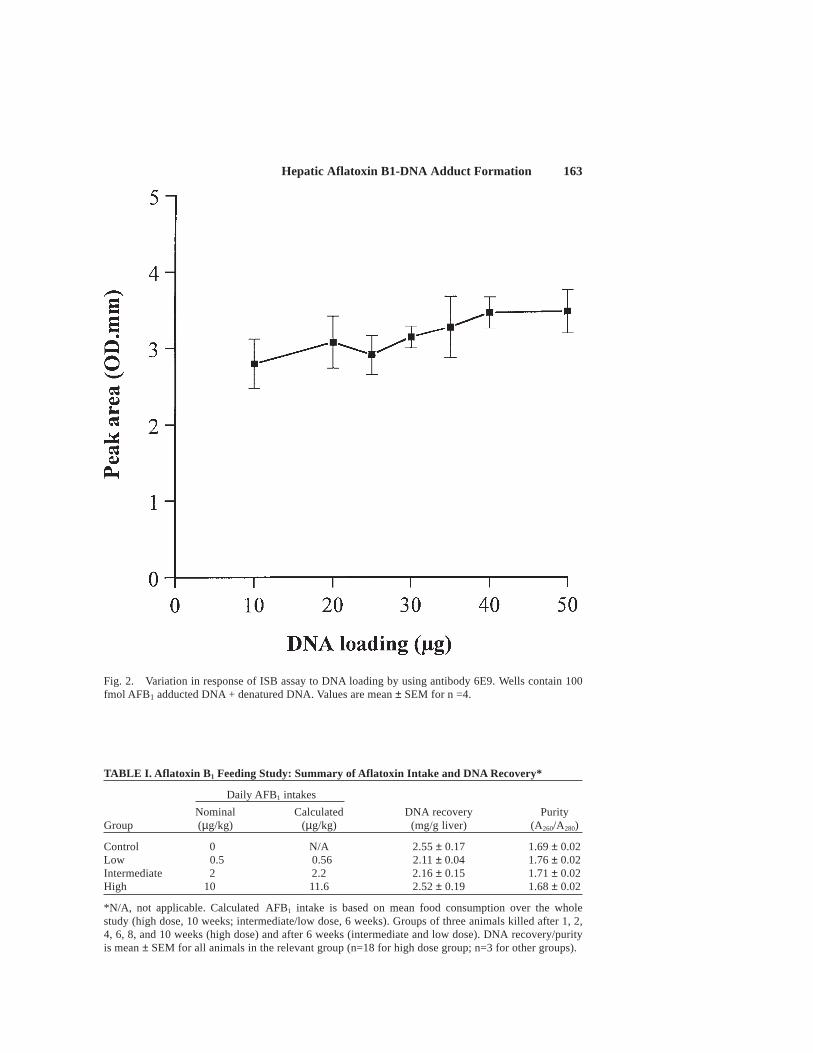
In order to establish optimum conditions for this asssay, nitrocellulose mem-branes were initially loaded with calf thymus DNA containing varying amounts ofhighly adducted DNA to provide 0, 1, 10, 100, 1000 and 10,000 fmol adduct in 5 µgtotal DNA. Having established a suitable working range of adduct concentrations(0–50 fmol/slot), the effect of loading procedure, including method of denaturing theDNA (heat denaturing or alkali), primary and secondary antibody dilution, and incu-bation times, were investigated. Studies were also undertaken to determine the maxi-mum amount of adducted DNA that could loaded onto a membrane using in vitroadducted calf thymus DNA (100 fmol/slot).
RESULTS32P-Post Labelling
No adduct spots were seen on 2-D chromatograms of adducted-DNA subjectedto post labelling under both ATP-deficient and ATP-excess conditions with or with-out nuclease P1 enhancement. Similarly, no clear evidence for adduct formation wasseen using the alternative methods for DNA hydrolysis [25,26].
Immunoslot Blotting–Method Development
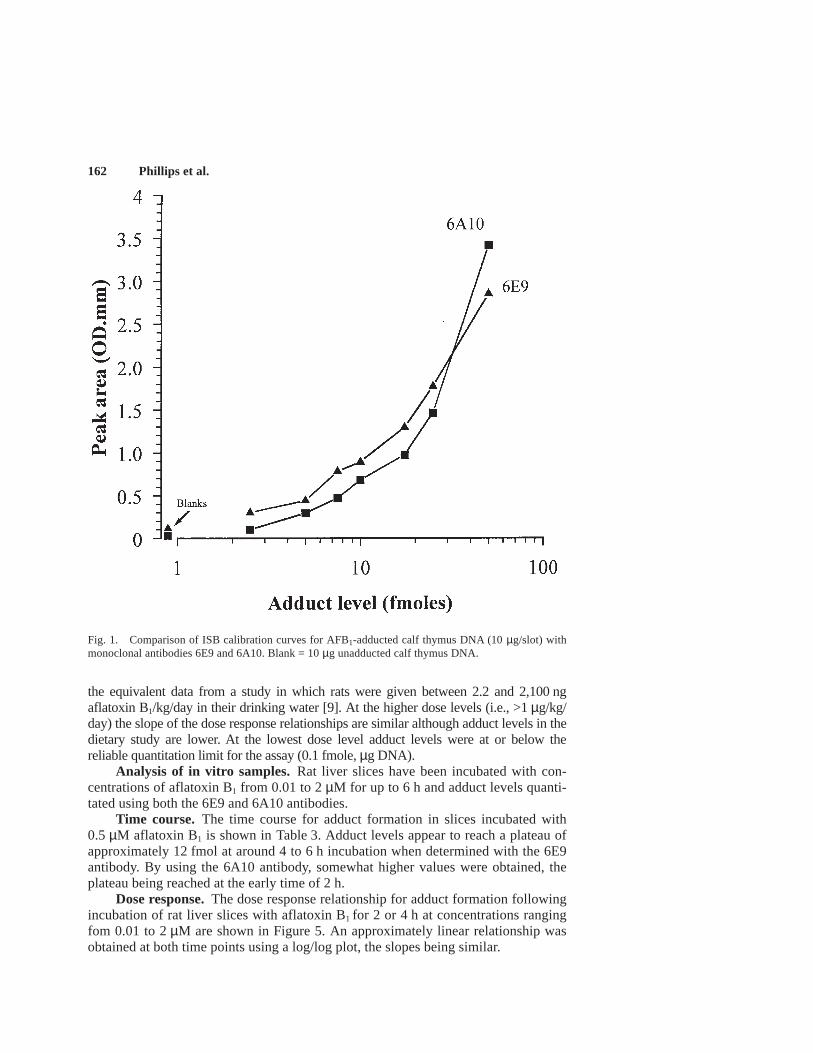
Representative calibration curves for slot blot assays using either monoclonalantibody 6E9 or 6A10 are shown in Figure 1 and DNA loading curves are shown inFigure 2. The detection limit for this assay is approximately 1–2 fmol adduct per slotwith either antibody and up to 50 µg DNA can be loaded on to each slot withoutaffecting the response in the assay. Optimum secondary antibody dilution was deter-mined to be 1:2,000, whereas that for the primary antibody (1:7,500 to 1:10,000)varied depending on the nature and batch of antibody used. Attempts to improve thedetection limit with the SuperSignal reagent system were unsuccessful.
In Vivo Feeding Study
The calculated mean intakes of AFB1 are shown in Table I. Daily intake varied from10.8 to 11.9 µg/kg/day for the high dose animals (nominal 10 µg/kg/day), from 2.0 to2.4 µg/kg/day for the intermediate dose animals (nominal 2 µg/kg/day), and from 0.5 to0.6 µg/kg/day for the low dose animals (nominal 0.5 µg/kg/day). The growth rate oftreated animals was similar to that of the control group at all intake levels (data notshown) Overall recovery of DNA varied from approx. 1.7 to 3.7 mg/g liver wet weightand the purity was approx. 95% as assessed by the A260/A280 ratio (Table I).
Analysis of In Vivo Adducted DNA Samples
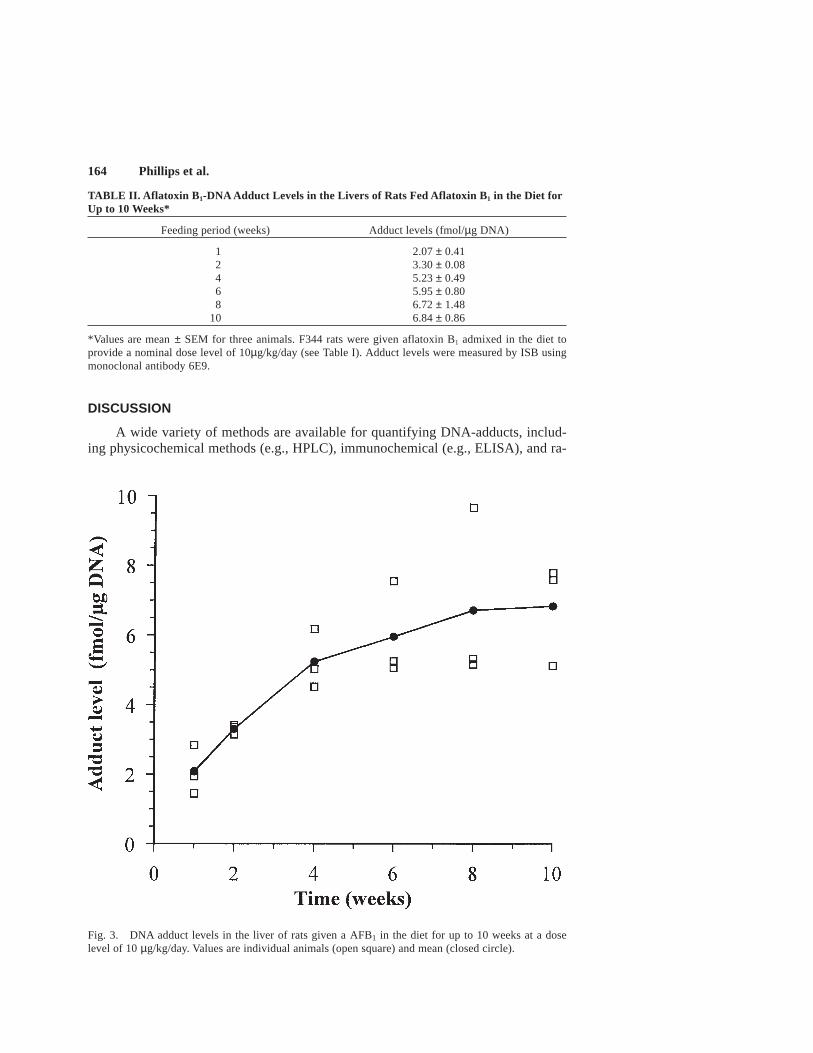
Time course. The levels of aflatoxin adducts in DNA recovered from the liversof rats given 10 µg aflatoxin B1/kg/day in the diet for 1, 2, 4, 6, 8 and 10 weeks areshown in Table II and Figure 3. Adduct levels increased from approximately 2 fmol/µg DNA at 1 week to between 5 and 8 fmol/µg DNA at 10 weeks. No marked in-crease was seen after 6 weeks however, suggesting that a steady state had been es-tablished at this time.
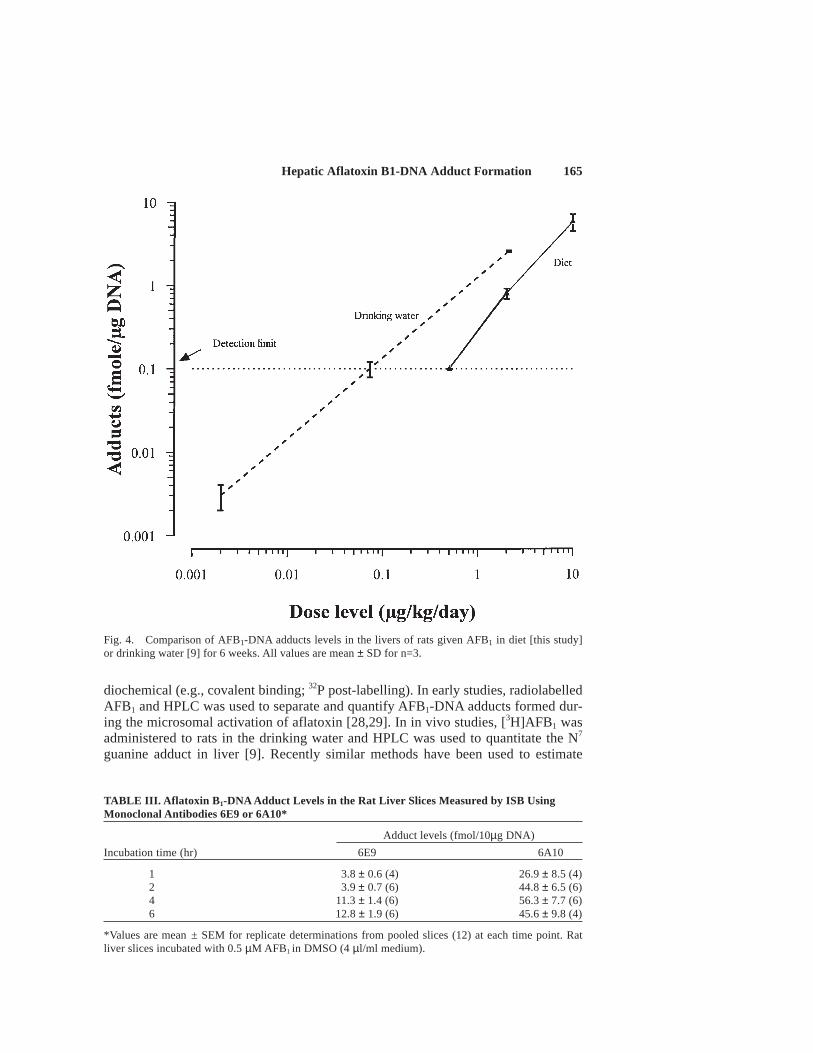
Dose response.The level of aflatoxin adducts in DNA from rats given either 0.5, 2or 10 µg aflatoxin B1/kg/day in the diet for 6 weeks are shown in Figure 4 together with
162 Phillips et al.
the equivalent data from a study in which rats were given between 2.2 and 2,100 ngaflatoxin B1/kg/day in their drinking water [9]. At the higher dose levels (i.e., >1 µg/kg/day) the slope of the dose response relationships are similar although adduct levels in thedietary study are lower. At the lowest dose level adduct levels were at or below thereliable quantitation limit for the assay (0.1 fmole, µg DNA).
Analysis of in vitro samples.Rat liver slices have been incubated with con-centrations of aflatoxin B1 from 0.01 to 2 µM for up to 6 h and adduct levels quanti-tated using both the 6E9 and 6A10 antibodies.
Time course. The time course for adduct formation in slices incubated with0.5 µM aflatoxin B1 is shown in Table 3. Adduct levels appear to reach a plateau ofapproximately 12 fmol at around 4 to 6 h incubation when determined with the 6E9antibody. By using the 6A10 antibody, somewhat higher values were obtained, theplateau being reached at the early time of 2 h.
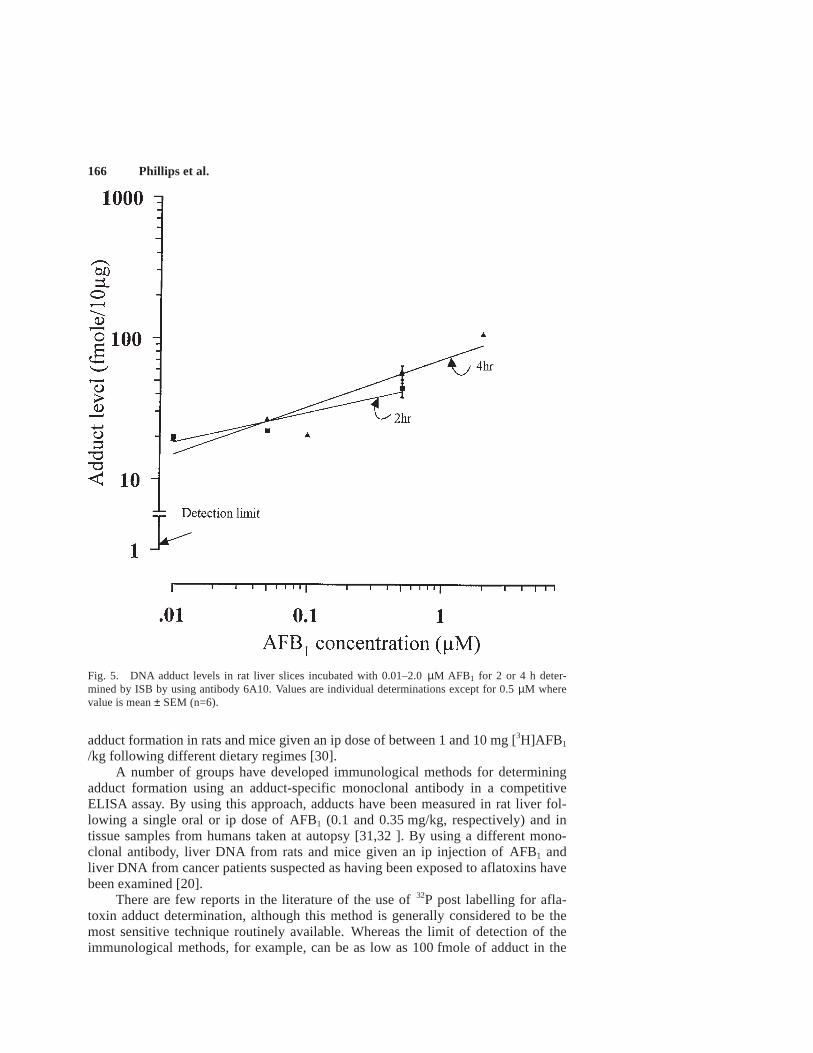
Dose response.The dose response relationship for adduct formation followingincubation of rat liver slices with aflatoxin B1 for 2 or 4 h at concentrations rangingfom 0.01 to 2 µM are shown in Figure 5. An approximately linear relationship wasobtained at both time points using a log/log plot, the slopes being similar.
Fig. 1. Comparison of ISB calibration curves for AFB1-adducted calf thymus DNA (10 µg/slot) withmonoclonal antibodies 6E9 and 6A10. Blank = 10 µg unadducted calf thymus DNA.
Hepatic Aflatoxin B1-DNA Adduct Formation 163
TABLE I. Aflatoxin B 1 Feeding Study: Summary of Aflatoxin Intake and DNA Recovery*
Daily AFB1 intakes
Nominal Calculated DNA recovery PurityGroup (µg/kg) (µg/kg) (mg/g liver) (A260/A280)
Control 0 N/A 2.55 ± 0.17 1.69 ± 0.02Low 0.5 0.56 2.11 ± 0.04 1.76 ± 0.02Intermediate 2 2.2 2.16 ± 0.15 1.71 ± 0.02High 10 11.6 2.52 ± 0.19 1.68 ± 0.02
*N/A, not applicable. Calculated AFB1 intake is based on mean food consumption over the wholestudy (high dose, 10 weeks; intermediate/low dose, 6 weeks). Groups of three animals killed after 1, 2,4, 6, 8, and 10 weeks (high dose) and after 6 weeks (intermediate and low dose). DNA recovery/purityis mean ± SEM for all animals in the relevant group (n=18 for high dose group; n=3 for other groups).
Fig. 2. Variation in response of ISB assay to DNA loading by using antibody 6E9. Wells contain 100fmol AFB1 adducted DNA + denatured DNA. Values are mean ± SEM for n =4.
164 Phillips et al.
TABLE II. Aflatoxin B 1-DNA Adduct Levels in the Livers of Rats Fed Aflatoxin B1 in the Diet forUp to 10 Weeks*
Feeding period (weeks) Adduct levels (fmol/µg DNA)
1 2.07 ± 0.412 3.30 ± 0.084 5.23 ± 0.496 5.95 ± 0.808 6.72 ± 1.48
10 6.84 ± 0.86
*Values are mean ± SEM for three animals. F344 rats were given aflatoxin B1 admixed in the diet toprovide a nominal dose level of 10µg/kg/day (see Table I). Adduct levels were measured by ISB usingmonoclonal antibody 6E9.
Fig. 3. DNA adduct levels in the liver of rats given a AFB1 in the diet for up to 10 weeks at a doselevel of 10 µg/kg/day. Values are individual animals (open square) and mean (closed circle).
DISCUSSION
A wide variety of methods are available for quantifying DNA-adducts, includ-ing physicochemical methods (e.g., HPLC), immunochemical (e.g., ELISA), and ra-
Hepatic Aflatoxin B1-DNA Adduct Formation 165
TABLE III. Aflatoxin B 1-DNA Adduct Levels in the Rat Liver Slices Measured by ISB UsingMonoclonal Antibodies 6E9 or 6A10*
Adduct levels (fmol/10µg DNA)
Incubation time (hr) 6E9 6A10
1 3.8 ± 0.6 (4) 26.9 ± 8.5 (4)2 3.9 ± 0.7 (6) 44.8 ± 6.5 (6)4 11.3 ± 1.4 (6) 56.3 ± 7.7 (6)6 12.8 ± 1.9 (6) 45.6 ± 9.8 (4)
*Values are mean ± SEM for replicate determinations from pooled slices (12) at each time point. Ratliver slices incubated with 0.5 µM AFB1 in DMSO (4 µl/ml medium).
Fig. 4. Comparison of AFB1-DNA adducts levels in the livers of rats given AFB1 in diet [this study]or drinking water [9] for 6 weeks. All values are mean ± SD for n=3.
diochemical (e.g., covalent binding; 32P post-labelling). In early studies, radiolabelledAFB1 and HPLC was used to separate and quantify AFB1-DNA adducts formed dur-ing the microsomal activation of aflatoxin [28,29]. In in vivo studies, [3H]AFB1 wasadministered to rats in the drinking water and HPLC was used to quantitate the N7
guanine adduct in liver [9]. Recently similar methods have been used to estimate
166 Phillips et al.
adduct formation in rats and mice given an ip dose of between 1 and 10 mg [3H]AFB1
/kg following different dietary regimes [30].A number of groups have developed immunological methods for determining
adduct formation using an adduct-specific monoclonal antibody in a competitiveELISA assay. By using this approach, adducts have been measured in rat liver fol-lowing a single oral or ip dose of AFB1 (0.1 and 0.35 mg/kg, respectively) and intissue samples from humans taken at autopsy [31,32 ]. By using a different mono-clonal antibody, liver DNA from rats and mice given an ip injection of AFB1 andliver DNA from cancer patients suspected as having been exposed to aflatoxins havebeen examined [20].
There are few reports in the literature of the use of 32P post labelling for afla-toxin adduct determination, although this method is generally considered to be themost sensitive technique routinely available. Whereas the limit of detection of theimmunological methods, for example, can be as low as 100 fmole of adduct in the
Fig. 5. DNA adduct levels in rat liver slices incubated with 0.01–2.0 µM AFB1 for 2 or 4 h deter-mined by ISB by using antibody 6A10. Values are individual determinations except for 0.5 µM wherevalue is mean ± SEM (n=6).

Hepatic Aflatoxin B1-DNA Adduct Formation 167
analysed DNA sample, 32P post-labelling can detect attomoles of adduct and requiresless adducted DNA per assay. This technique has been reported to detect AFB1-DNAadducts from both in vivo and in vitro adducted DNA [27] and has been used toshow that the major adduct detected in DNA reacted with AFB1-8,9-epoxide was thesame as that obtained from the reaction of this epoxide with guanine monophophate(GMP) [33 ]. No indication of the sensitivity of the method was presented.
Attempts to detect AFB1-DNA adducts from in vivo adducted samples in thepresent study using 32P post-labelling have been unsuccessful. The expected adductmaps were not obtained when highly-adducted DNA was treated with spleenphosphdiesterase and microccocal nuclease [24]. Subsequent experiments using dif-ferent techniques for degrading the adducted DNA [25,26] have suggested that theadduct is a poor substrate for the 32P phosphorylation reaction, a suggestion sup-ported by others (Harrison JC, personal communication).
The sensitive immunological technique, immunoslot blotting (ISB), developedfor the detection of O6-ethyldeoxyguanosine [34] has been used recently to quanti-tate methyleugenol [35] and O6-carboxymethyl deoxyguanosine adducts [36]. In ourstudy, the approach was developed to detect aflatoxin B1-DNA adducts, by usingeither of the available anti-aflatoxin-DNA monoclonal antibodies, 6E9 [31] or 6A10[20]. With both antibodies a detection limit of approx. 1-2 fmoles in the assay wasachieved with in vitro adducted DNA as standard, equivalent to 1 adduct in approxi-mately 107 normal bases.
By using this approach, the time to reach steady-state levels for DNA adductsin the liver of rats fed aflatoxin B1 in the diet at a level of 10 µg/kg/day was approxi-mately 6–8 weeks, a period similar to that reported in animal given aflatoxin B1 inthe drinking water at dose levels of between 2 ng and 2 µg/kg/day [9]. In the presentstudy the steady-state adduct level at 10 µg/kg/day was approximately 6 fmole/µgDNA or 2,000 adducts in 109 nucleotides and at 2 µg/kg/day was 0.8 fmole/µg DNAor approximately 250 adducts in 109 nucleotides. These values compare with 850and 200 adducts in 109 nucleotides found at dose levels of 2 and 0.5 µg/kg/day,respectively [9]. Adduct levels in liver DNA from animals at the next lower doselevel (0.5 µg/kg/day) in our study were too low for reliable quantitation. Thus toextend the dose response curve to levels representative of likely human exposure, byusing adduct levels as the biomarker of exposure, requires a substantially more sen-sitive assay method. Such a method may be the recently developed AMS technique[37] which can detect adducts at a level of 1 in 1012 normal bases. This technique hasbeen used to quantitate adducts from low levels of cooked food mutagens and toinvestigate adduct formation from tamoxifen [38].
Hepatic aflatoxin B1-DNA adduct levels have also been measured in in vitrosystems [39–42]. In one study in which hepatocytes were incubated with 0.2 µM 3H-labelled aflatoxin B1 [40], adduct levels at 24 hr (25–69 pmol/mg) were similar inhepatocytes isolated from female Sprague-Dawley rats and from three humans sub-jects. However, in hepatocytes from male rats, the adduct level (203 pmol/mg) wassubstantially greater. Similar adduct levels were also found in male rat hepatocytesincubated with approximately 0.1 µM aflatoxin B1 for 6 h [41,42].
In the only report in the literature on the level of adducts in precision-cut liverslices incubated with aflatoxin B1 [39], a value of 53.6 pmol/mg (fmol/µg) DNA wasreported in slices incubated with 0.5µM 3H-labelled aflatoxin B1 for 2 h in a dy-namic organ culture system. As with cultured hepatocytes, markedly lower adduct

168 Phillips et al.
levels were found in human liver slice DNA (1.6–14 pmol/mg) compared with thatin male rat liver slices. In the present work, the level of adducts in slices incubatedwith 0.5µM aflatoxin B1 for 2 h was between 0.4 and 4.4 fmol/µg DNA, dependingon the antibody. The reason for the marked difference between the values in the twostudies is not clear, although it has been noted that aflatoxin adduct levels measuredusing 3H-labelled compound tend to be higher than when measured by immunochemi-cal methods [20].
Since adduct levels represent the balance between activation and detoxificationpathways, the generally lower level of adduct formation in human systems in vitrocompared with the male rat [39,40] and the relative ineffectiveness of constitutivelyexpressed human glutathione transferases to detoxify AFB1-epoxide has been takento suggest that there are significant differences in the activation/detoxification path-ways of aflatoxin B1 in rats and humans [43,44]. Further studies to examine dose-response relationships for adduct formation in rat and human liver slices shouldprovide valuable data to assess the value of rat data for the extrapolation of risk tohumans of exposure to low levels of aflatoxin B1.
ACKNOWLEDGMENTS
The results of this research are the property of the Ministry of Agriculture,Fisheries and Food and are Crown Copyright.
REFERENCES
1. Kodell RL, Gaylor DW. Uncertainty of estimates of cancer risk derived by extrapolation from highto low doses and from animals to humans. Int J Toxicol 1997;16:449–460.
2. International Agency for Research on Cancer. Whelan SO, Parkin DM, Masuyer E, editors. 1990.Patterns of cancer in five continents, Vol VI; Lyon: IARC.
3. Eaton DL, Gallagher EP. Mechanisms of aflatoxin carcinogenesis. Annu Rev Pharmacol Toxicol1994;34:135–174.
4. Gonzalez FJ, Crespi CL, Gelboin HV. H.V. c-DNA expressed human cytochrome P-450’s : a newage of molecular toxicology and human risk assessment. Mutation Res 1991;247:113–127.
5. Essigmann JM, RG Croy RG, Nadzan AN, Busby WH, Reinhold VN, Buchi G, Wogan GN. Struc-tural identification of the major DNA adduct formed by aflatoxin B1 in vitro. Proc Natl Acad SciUSA 1977;74:1870–1874.
6. Croy RG, Essigmann JM, Reinhold DV, Wogan GN. Identification of the principal aflatoxin B1 -DNA adduct formed in vivo in rat liver. Proc Natl Acad Sci USA 1978;75:1745–1749.
7. Croy RG, Wogan GN. Temporal patterns of covalent DNA adducts in rat liver after single andmultiple doses of aflatoxin B1. Cancer Res 1981;41:197–203.
8. Choy WN. A review of dose response induction of DNA adducts by aflatoxin B1 and its implica-tions to quantitative cancer-risk assessment. Mutation Res 1993;296:181–198.
9. Buss P, Caviezel M, Lutz WK. Linear dose-response relationship for DNA adducts in rat liverfrom chronic exposure to aflatoxin B1. Carcinogenesis 1990;11:2133–2135.
10. Stoloff L. Aflatoxin as a cause of primary liver cancer in the United States: a probability study.Nutr Cancer 1983;5:165–185.
11. Groopman JD, Jiaqui Z, Donahue PR, Pikul A, Zhang L, Chen J-S, Wogan GN. Molecular dosim-etry of urinary aflatoxin-DNA adducts in people living in Guangxi Autonomous Region, People’sRepublic of China. Cancer Res 1992;52, 45–52.
12. International Agency for Research on Cancer. IARC Monograph on the Evaluation of Carcino-genic Risk to Humans: some naturally occurring substances : food items and constituents, hetero-cyclic aromatic amines and mycotoxins, Vol 56. Lyons: IARC; 1993. p 245–395.
13. Wogan GN, Paglialunga S, Newberne PN. Carcinogenic effect of low dietary levels of aflatoxin B1
in rats. Fd Cosmet Toxicol 1974;12:681–685.

Hepatic Aflatoxin B1-DNA Adduct Formation 169
14. Nixon JE, RO Sinnhuber RO, Lee DJ, Landers MK, Harr JR. Effect of cyclopropenoid compoundson the carcinogenic activity of diethylnitrosamine and aflatoxin B1 in rats. J Natn Cancer Inst1974;53:453–458.
15. Dashwood RH, Arbogast DN, Fong AT, Pereira C, Hendricks JD, Bailey GS. Quantitative interre-lationships between aflatoxin B1 carcinogen dose, indole-3-carbinol anticarcinogenic dose, targetorgan DNA adduction and final tumour response, Carcinogenesis 1988;10:175–181.
16. Dashwood RH, Arbogast DN, Fong AT, Hendricks JD, Bailey GS. Mechanism of anticarcinogenesisby indole-3-carbinol: Detailed in vivo DNA binding dose-ressponse studies after dietary adminis-tration with aflatoxin B1. Carcinogenesis 1988;9:427–432.
17. Lee DJ, Wales JH, Sinnhuber RO. Promotion of aflatoxin-induced hepatoma growth in trout bymethyl malvalate and sterculate. Cancer Res 1971;31:960–963.
18. Bechtel DH. Molecular dosimetry of hepatic aflatoxin B1 -DNA adducts: linear correlation withhepatic cancer risk, Regul Tox Pharmacol 1989;10:74–81.
19. Martin CN, Garner RC. Aflatoxin B1-oxide generated by chemical or enzymic oxidation of afla-toxin B1 causes guanine substitution in nucleic acids. Nature 1977;267:863–865.
20. Hsieh L-L, Hsu S-W, Chen D-S, Santella R. Immunological detection of aflatoxin B1 -DNA ad-ducts formed in vivo. Cancer Res 1988;48:6328–6331.
21. Price RJ, Ball SE, Renwick AB, Barton PT, Beamand JA, Lake BG. Use of precision-cut liverslices for studies of xenobiotic metabolism and toxicity : comparison of the Krumdieck and Brendeltissue slicers. Xenobiotica 1998;28:361–371.
22. Gupta RC. Non-random binding of the carcinogen N-hydroxy-2-acetylaminofluorene to repetitivesequences of rat liver DNA in vivo. Proc Natl Acad Sci USA 1984;81:6943–6947.
23. Ingram AJ, Phillips JC, Lee R. DNA adduct formation by mineral oils and their fractions as indi-cated by 32P-postlabelling: is adduct formation truly indicative of carcinogenic potential, J AppliedToxicol 1995;15:275–283.
24. Reddy MV, Randerath K. Nuclease P1-mediated enhancement of the sensitivity of 32P postlabelingtest for structurally diverse DNA adducts. Carcinogenesis 1986;7:1543–1551.
25. Randerath K, Randerath E, TF Danna TF, van Golen KL, Putman KL. A new sensitive 32 postlabelingassay based on the specific enzymic conversion of bulky DNA lesions to radiolabeled dinucle-otides and nucleoside-5′-monophosphates. Carcinogenesis 1989;10:1231–1239.
26. Weinfeld M, Soderlind K-JM. 32 Postlabeling detection of radiation- induced DNA damage: identi-fication and estimation of thymine glycols and phosphoglycolate termini, Biochemistry1991;30:1091–1097.
27. Randerath K, Randerath E, Agrawal HP, Reddy MV. Biochemical (postlabelling) methods for analysisof carcinogen-DNA adducts. In: A Berlin A, Draper M, Hemminki K, Vainio H, editors. Monitor-ing human exposure to carcinogenic and mutagenic agents. IARC Scientific Publication No. 59.Lyon: IARC; 1985. p 217–231.
28. Lin J-K, Miller JA, Miller EC. 2,3-Dihydro-2-(guan-7-yl)-3-hydroxy-aflatoxin B1 , a major acidhydrolysis product of aflatoxin B1 -DNA or -ribosomal RNA adducts formed in hepatic microsome-mediated reactions and in rat liver in vivo, Cancer Res 1977;37:4430–4438.
29. Groopman JD, Croy RG, Wogan GN. In vitro reactions of aflatoxin B1-adducted DNA. Proc NatlAcad Sci USA 1981;78:5445–5449.
30. Chou MW, Chen W. Food restriction reduces aflatoxin B1 (AFB1)-DNA adduct formation, AFB1-glutathione conjugation and DNA damage in AFB1-treated male F344 rats and B6C3F1 mice. JNutr 1997;122:210–217.
31. Harrison JC, Carvajal M, Garner RC. Immunological detection of aflatoxin B1-DNA and AFB1-serum albumin adducts in rat and human tissues. In: Garner RC, PB Farmer PB, Steel GT, WrightAS, editors. Human carcinogen exposure. Biomonitoring and risk assessment. Oxford: Oxford Uni-versity Press: 1991. Ch 20, p 255–265.
32. Harrison JC, Carvajal M, Garner RC. Does aflatoxin exposure in the United Kingdom constitute acancer risk. Environ Hlth Perspect 1993;99:99–105.
33. Routledge MN, Garner RC, Harrison JC. 32P post-labelling assay for the detection of aflatoxin B1-DNA adducts. Mutagenesis 1991;6:428.
34. Nehls P, Adamkiewicz J, Rajewsky MF. Immuno-slot-Blot: a highly sensitive immunoassay for thequantitation of carcinogen-modified nucleosides in DNA. Cancer Res Clin Oncol 1984;108:23–29.
35. Gardner I, Bergin P, Stening JG, Caldwell J. Immunochemical detection of covalently modifiedprotein adducts in livers of rats treated with methyleugenol. Chem Res Toxicol 1996;9:713–721.
36. Harrison KL, Fairhurst N, Challis BC, Shuker DEG. Synthesis, characterisation and immunochemical

170 Phillips et al.
detection of O6-carboxymethyl-2′- deoxyguanosine: a major DNA adduct formed by nitrosatedglycine derivatives. Chem Res Toxicol 1997;10:652–659.
37. Turtletaub KW, Vogel JS, Frantz CE, Fultz E. Studies on DNA adduction with heterocyclic aminesusing accelerator mass spectrometry: a new technique for tracing isotope-labelled DNA adducts.In: Phillips DH, Castegnaro M, Bartsch H, editors. Post labelling methods for detection of DNAadducts. IARC Scientific Publications No 124. Lyons: IARC; 1993. p 293–301.
38. Shuker DEG, Benford DJ. DNA modification in carcinogen risk assessment in relation to diet:recent advances and some perspectives from a Workshop. Biomarkers 1997;2:265–278.
39. Heinonen JT, Fisher R, Brendel K, Eaton DL. Determination of aflatoxin B1 biotransformation andbinding to hepatic macromolecules in human precision liver slices. Toxic Appl Pharmacol1996;136:1–7.
40. Cole KE, Jones TW, Lipsky MM, Trump BF, Hsu I-C. In vitro binding of aflatoxin B1 and 2-acetylaminofluorene to rat, mouse and human hepatocyte DNA: the relationship of DNA bindingto carcinogenicity. Carcinogenesis, 1988;9:711–716.
41. Loury DJ, Hseih DPH, Byard DL. The effect of phenobarbital pretreatment on the metabolism,covalent binding and cytotoxicity of aflatoxin B1 in primary cultures of rat hepatocytes. J ToxicolEnviron Hlth 1984;13:145–159.
42. Salocks CB, Hseih DPH, Byard JL. Effect of butylated hydroxytoluene pretreatment on the me-tabolism and genotoxicity of aflatoxin B1 in primary cultures of adult rat hepatocytes: selectivereduction of nucleic acid binding, Toxic Appl Pharmacol 1984;76:498–509.
43. Moss EJ, Neal GE. The metabolism of aflatoxin B1 by human liver. Biochem Pharmacol1985;34:3193–3197.
44. Raney KD, Meyer DJ, Ketterer B, Harris TM, Guengerich FP. Glutathione conjugation of aflatoxinB1 exo-epoxides and endo-epoxides by rat and human glutathione S-transferases. Chem Res Toxicol1992;5:470–478.