dna polymorphism: spectroscopic and electro-optic characterizations of z-dna and other types of...
TRANSCRIPT

DNA Polymorphism: Spectroscopic and Electro-Optic Characterizations of Z-DNA and Other Types of Left-Handed Helical
Structures Induced by Ni2+
MARIE-FRANCOISE HACQUES and CHRISTIAN MARION, Laboratoire cle Physico-Chimie Biologique, LBTM- CNRS UM 24,
Uniuersitk Claude Bernurd- Lyon I 69622, Villeurbanne Ckclex, France
Synopsis
CD and uv spectroscopy reveal that the synthetic polynucleotides poly(dG-dC) . poly(dG-dC) and poly(dG-m5dC) . poly(dG-m5dC) undergo a transition induced by small amounts of Ni++ cation from a right-handed B-form to left-handed Z-type conformations. We describe the application of steady-state and transient electric birefringence to the characterization of the transition observed at very low ionic strength (10 mM Tris HC1, pH 7.4). Dialysis experiments show that the changes in spectroscopic and electro-optic properties upon addition of Ni++ are completely reversible. The differences in shape of the inverted CD spectra suggest the existence of a family of left-handed conformations, depending on the polymer used and on the amounts of cation added. The stoichiometry required for inducing the Z-conformation of poly(dG-m5dC) is 1 cation/4 nucleotide phosphates. The transition is accompanied by a decrease in birefringence, an increase in length, and the more important contribution of a permanent or slowly induced dipole moment to the orientation mechanism. High concentrations of Ni++ promote the Z --* Z* transition. Upon increasing the Ni++ concentration, poly(dG-dC) undergoes a biphasic transition, first to one intermediate conformation that is neither B- nor Z-like and then to a left-handed form that is probably different from Z*. These conversions are accompanied by regular decreases both in birefringence and in chain length, but no transient appears in the field-reversal experiments.
INTRODUCTION
The DNA double helix has considerable structural flexibility and changes conformation with experimental conditions. The most drastic change is the transition from the right-handed B- to the left-handed Z-form. Z-conforma- tion is favored in nucleotide sequences that have alternations of purines and pyrimidines, especially alternations of cytosine and guanine residues (see Ref. 1 for a general review). The Z-DNA helix was discovered for short oligonuc- leotides d(GC), by using x-ray diffraction2 The structure explained the polymorphism observed in solution with poly(dG-dC) some years ago.3 An equilibrium exists between B-DNA and Z-DNA, and the presence of metal ions or modified bases was found to exert a strong effect on the Z-formation. One of the most important modifications, associated with gene inactivation, is the methylation of cytosine on the C5 position. 5mC appears the only modified base in eukaryotic DNA and occurs predominantly in the CpG ~equence.~ The methylated sequences are associated with decreased levels of tran~cription.~ Behe and Felsenfeld have shown that poly(dG-m5dC) adopts
Biopolymers, Vol. 25, 2281-2293 (1986) 0 1986 John Wiley & Sons, Inc. CCC OOO6-3525/86/122281- 13$04 .oO

2282 HACQUES AND MARION
the Z-conformation at moderate salt concentration, much lower than that required for the unmethylated polymer.6 The equilibrium is strongly shifted toward the Z-form with addition of small amounts of divalent cations or p~lyamines .~-~ The methylated polymer thus undergoes the B -, Z transition at very low ionic strength; this makes it possible to apply electro-optical methods for studying the equilibrium in solution and therefore protein-DNA interactions. That is of considerable interest because the differences in con- densation of chromatin by histone H1 subtypes'o'" or modified histones,12 as we have recently revealed by electric birefringence, could involve recognition of local variations in DNA conformation.
As f a r as we are aware, there are only two reports in the literature of electro-optical measurements on Z-DNA. Chen et al.13 studied length changes induced in 145 base pairs (bp) poly(dG-m5dC) by Co(NH,);+ and Wu et al.14 reported electric dichroism studies on short fragments of poly(dG-dC) in alcohol/water mixtures.
Here we present evidence that Ni'+ cation can, under appropriate condi- tions, induce a right-to-left helical transformation of the alternating co- polymers poly(dG-dC) and poly(dG-m5dC). Our evidence consists primarily of spectroscopic measurements (uv absorption and CD spectra), and we describe the application of steady-state and transient electric birefringence to the characterization of transition.
MATERIAL AND METHODS
Poly(dG-dC) * poly(dG-dC) and poly(dG-m5dC) . poly(dG-m5dC) were ob- tained from P. L. Biochemicals. The average sizes of the polymers estimated from gel electrophoresis were of about 800-850 and 900-950 bp, respectively. Because the B 3 Z transition of the poly(dG-m5dC) . poly(dG-m5dC) poly- mer is very sensitive to traces of divalent or polyvalent ions, the solutions were first exhaustively dialyzed against 10 mM Tris HC1, pH 7.4, for 24 h at 4°C. Nickel chloride was analytical grade. The equilibrium properties of poly(dG-dC) and poly(dG-m5dC) in solutions of NiC1, (0-5 mM) were determined after 1-3 h incubation at ambient temperature. The final ab- sorbances of polymers were between 1 and 1.2. The amounts of added metal ions were expressed in terms of Ni2+/nucleotide ratios, y , which varied between 0 and 10.
Absorption measurements were performed with a Beckman 25 spectropho- tometer. CD spectra were obtained at room temperature with a Jobin-Yvon Mark IV dichrograph. The molar ellipticity 181 was calculated from the measured ellipticity 8 , determined in quartz cuvettes with a light path of 1 cm. The units are degrees. cm2 - dmol-'.
The principle of electric birefringence is to orient macromolecules by means of a rectangular electric pulse. The technique has been well described else~here. '~ Basically, the resulting birefringence A neq is proportional to the square of the electric field E at low field strength. The specific Kerr constant B is defined by B = An,(cA E 2, where c is the solute concentration and A is the wavelength of the incident light used to determine the birefringence (here, X = 632.8 nm). When the field is switched off, the birefringence decreases exponentially with a typical time dependency, An(t), given by the Benoit

DNA POLYMORPHISM 2283
where An,, and ai = An,, J h n , are the absolute and fractional steady-state birefringence values, respectively; rl is the relaxation time for process i which is related to the rotational diffusion coefficient D,, by T~ = l/6Di. Details on the polarization mechanism can be obtained by applying reverse pulses to the cell. The minimum An, appearing on the birefringence signal, allows easy calculation of the ratio r, which characterizes the electric properties of macromolecules: r = (1 - An,)/(0.1547 + Anm) = p2/Aci . kT where p is the permanent dipole moment, Aa the anisotropy of electrical polarizability, and kT the Boltzmann term.
Electric birefringence measurements were performed on an apparatus built in our laboratory and described in detail elsewhere’6*’7; here, we describe them briefly: the light source was a He-Ne laser and light beam variations were converted to electric signals by a low-noise solid-state photodetector followed by a specially designed amplifier. The signal intensity is linearly proportional to the birefringence. Rectangular and reverse electric pulses having amplitude up to loo0 and 150 V, respectively, were applied across a 0.5-cm interelectrode cell. Experiments were performed at room temperature and the birefringence results have been expressed in terms of specific birefringences (An,!A 260).
Relaxation times ( r l , “fast,” and T,, “slow”) were obtained by fitting the decay curve to an exponential curve with two parametric contributions ( a , and a,). The computer program was based on a two-step adjustment fitting.I8
RESULTS
Spectroscopic Studies
U V A bsorption
The uv spectra of poly(dG-dC) and poly(dG-m5dC) under various condi- tions are shown in Fig. 1. A t very low ionic strength (10 mM Tris HCl, pH 7.4), the polymers have the B-conformation. Although the addition of NiC1, is able to generate left-handed helical conformations of both polynucleotides, their behavior with changing salt concentration is different. Figure 3A shows the changes of their optical properties as a function of added Ni2+.
The A,,/A, ratio of poly(dG-dC) increases from 0.12 (B-form) to 0.20 for a Ni++/PO; ratio of 0.5 and becomes equal to 0.40, indicating conversion to the Z-conf~rmation~.’~ for y = 10. The titration curve reported here, however, is different from that observed by van de Sande et a1.: who describe biphasic optical transitions in the presence of NiCl, or CoCI,.
Poly(dG-m5dC) . poly(dG-m5dC) undergoes the spectral transition from B- to 2-form a t much lower Ni2+ concentrations. At a ratio of y = 0.25, the absorption ratio immediately changes from 0.21 (B-form) to 0.38. The conver- sion to the Z-form is characterized, for y 2 2, by a threefold increase in the A2g5/A260 ratio, very close to those previously r ep~r t ed .~ .~ ,~ ’
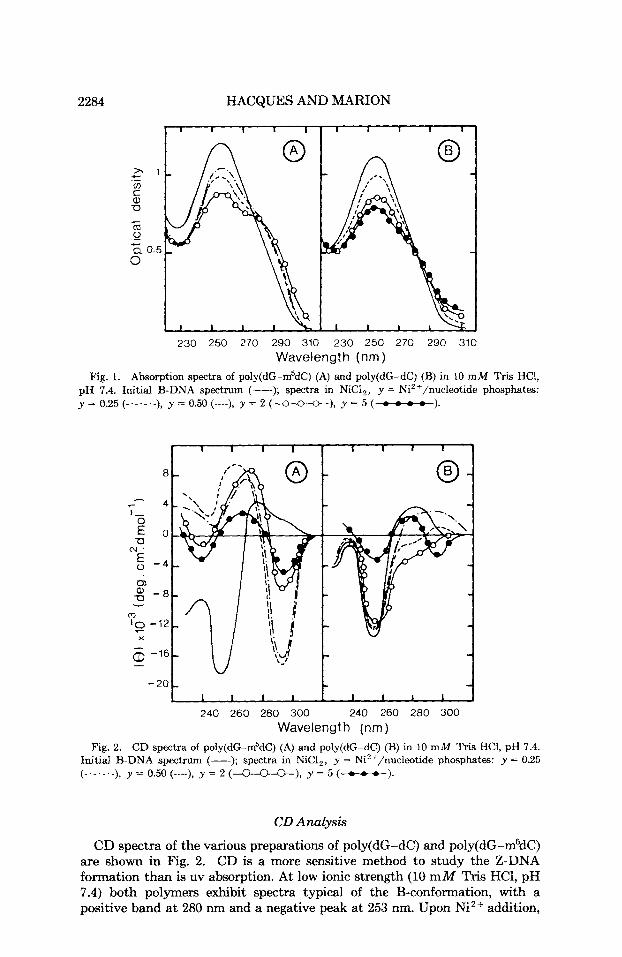
2284 HACQUES AND MARION
._ 2 ' (I) C a, -0
m 0
0. 0.5
- .- c
0
I 1 I
230 250 270 290 310 230 250 270 290 310 Wavelength (nm )
Fig. 1. Absorption spectra of poly(dG-m5dC) (A) and poly(dG-dC) (B) in 10 m M Tris HCI, pH 7.4. Initial B-DNA spectrum (-); spectra in NiCl,, y = Ni2+/nucleotide phosphates: y = 0.25 (-.-.-.-), y = 0.50 (----), y = 2 (-), y = 5 (-).
240 260 280 300 240 260 280 300 Wavelength (nm)
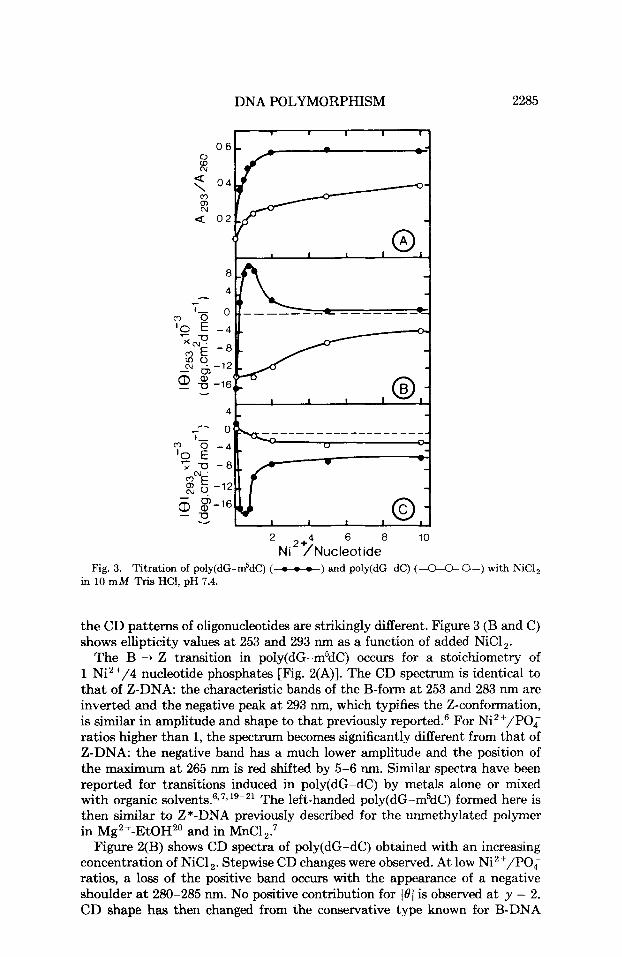
Fig. 2. CD spectra of poly(dG-m5dC) (A) and poly(dG-dC) (B) in 10 m M Tris HC1, pH 7.4. Initial B-DNA spectrum (-); spectra in NiCI,, y = Ni2+/nucleotide phosphates: y = 0.25 (-.-.-.-), y = 0.50 (----), y = 2 (-), y = 5 (-).
CD Analysis
CD spectra of the various preparations of poly(dG-dC) and poly(dG-m5dC) are shown in Fig. 2. CD is a more sensitive method to study the 2-DNA formation than is uv absorption. A t low ionic strength (10 mM Tris HCI, pH 7.4) both polymers exhibit spectra typical of the B-conformation, with a positive band at 280 nm and a negative peak at 253 nm. Upon Ni2' addition,
DNA POLYMORPHISM 2285
Fig. 3. in 10 mM
0-) with NiCl,
the CD patterns of oligonucleotides are strikingly different. Figure 3 (B and C) shows ellipticity values a t 253 and 293 nm as a function of added NiC1,.
The B + Z transition in poly(dG-m5dC) occurs for a stoichiometry of 1 Ni2'/4 nucleotide phosphates [Fig. 2(A)]. The CD spectrum is identical to that of Z-DNA: the characteristic bands of the B-form at 253 and 283 nm are inverted and the negative peak at 293 nm, which typifies the Z-conformation, is similar in amplitude and shape to that previously reported.6 For Ni2+/P0; ratios higher than 1, the spectrum becomes significantly different from that of Z-DNA: the negative band has a much lower amplitude and the position of the maximum at 265 nm is red shifted by 5-6 nm. Similar spectra have been reported for transitions induced in poly(dG-dC) by metals alone or mixed with organic s o l ~ e n t s . 6 ~ ~ * ~ ~ - ~ ~ The left-handed poly(dG-m5dC) formed here is then similar to Z*-DNA previously described for the unmethylated polymer in Mg2+-EtOH2' and in MnC1,:
Figure 2(B) shows CD spectra of poly(dG-dC) obtained with an increasing concentration of NiCl,. Stepwise CD changes were observed. A t low Ni2+/POq ratios, a loss of the positive band occurs with the appearance of a negative shoulder at 280-285 nm. No positive contribution for 101 is observed a t y = 2. CD shape has then changed from the conservative type known for B-DNA

2286 HACQUES AND MARION
helices to a completely nonconservative type. At y ratios higher than 2, the CD curves are a conservative type again, showing the inversion associated with the right- to left-handed transition. Spectra, however, differ considerably from the typical Z-spectrum, particularly for A < 260 nm.3 Moreover, the latter spectrum's features are not identical to those of the Z*-DNA ~ p e c t r u m ~ . ~ ~ observed above for poly(dG-m5dC) in that the positions of the maxima are slightly shifted [compare the curves - in Fig. 2(A) and (B)]: Z*-DNA shows negative bands at 240 and 295 nm, and a positive one a t 268 nm, while the unmethylated sample shows bands at 255, 295, and 276 nm, respectively. Therefore, we designate this latter form as Z"-DNA. In fact, such a structural form was already reported for poly(dG-dC) in the presence of CoCl or NiC17, but attributed by the authors to Z*-DNA (as they observed with MnCl,) despite significantly different CD spectra.
Stepwise CD changes similar to those shown in Fig. 2(B) were observed with transition metal^.^*'^^^^ The amount of added ions that results in the final conservative-type CD shape with inverted signs is lowered in the presence of ethylene glyco121 or after heating a t 55-65OC for 15 min.7~'~
The nonconservative CD shape observed corresponds to DNA present in an intermediate form that is neither B- nor Z-like.21 Both conformations exhibit CD spectra with positive ellipticities in the region of 265-285 nrn. The assumption that part of poly(dG-dC) is still present in the B-form and that part has adopted a left-handed conformation, then, appears unrealistic. This transition intermediate will be designated as TI-DNA. Models that accom- modate the spectroscopic transitions described above consist of sequential steps with increasing Ni2+ concentration:
poly(dG-dC) . poly(dG-dC)
poly( dG -m5dC) . pol y( dG -m5dC)
B + TI --* Z *
B + Z + Z "
These transitions are completely reversible. The B-forms of polymers can be obtained by extensively dialyzing left-handed forms against 10 m M Tris HCI, pH 7.4.
Electro-Optical Studies
We must keep in mind that aggregation is a difficulty inherent under the conditions discussed above. Therefore this problem must be considered in detail in order to obtain useful information with respect to the elec- tric birefringence of poly(dG-dC) . poly(dG-dC) and poly(dG-m5dC) . poly(dG-m5dC) in the presence of NiC1,. For y 5 5, artifacts in spectroscopic properties attributable to turbidity or differential scattering are not observed. Centrifugation at 15,000 g for 30 min does not cause a decrease in the intensity of the absorption and CD spectra. Moreover, no increase in light scattering is observed, as judged by the constant light intensity transmitted by the solutions in the absence of the electric field ("at rest") and reaching the sensitive low-noise solid-state photodetector. Aggregation is, then, not a problem in any birefringence and relaxation time measurements described below for y < 5. We only observe an increase of light-scattering intensity for Ni++/nucleotide phosphate ratios close to 10, and that corresponds to the appearance of a slight positive birefringence.
DNA POLYMORPHISM 2287
0 2 4
I I I
0 - 2 4
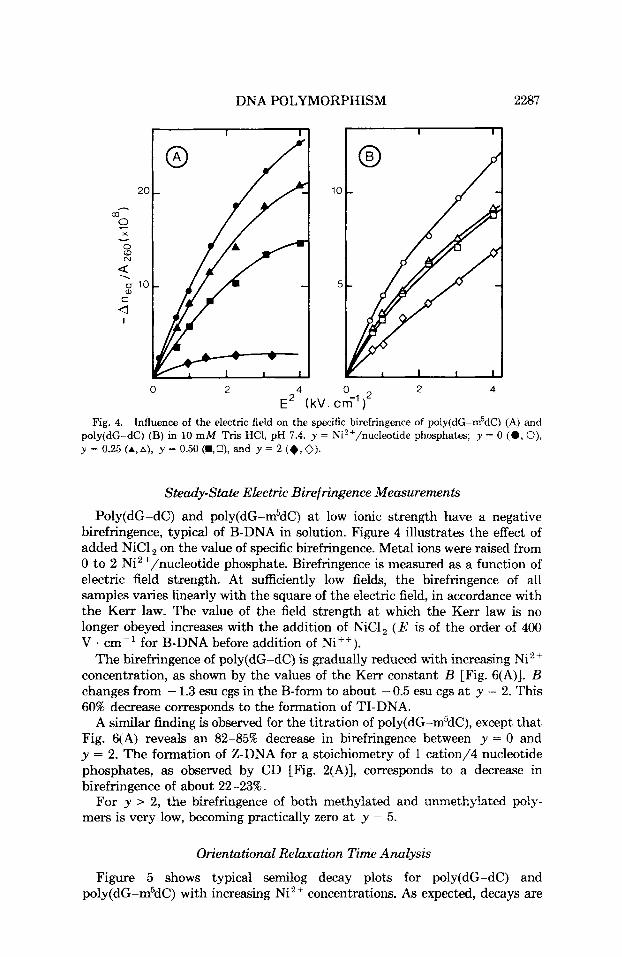
E' (kV. cm-')' Fig. 4. Influence of the electric field on the specific birefringence of poly(dG-m5dC) (A) and
poly(dG-dC) (B) in 10 mM Tris HCl, pH 7.4. y = Ni2+/nucleotide phosphates; y = 0 (0 , 0), y = 0.25 +,A), y = 0.50(a,U), and y = 2 (+,V).
Steady-State Electric Birefringence Measurements
Poly(dG-dC) and poly(dG-m5dC) at low ionic strength have a negative birefringence, typical of B-DNA in solution. Figure 4 illustrates the effect of added NiCl on the value of specific birefringence. Metal ions were raised from 0 to 2 Ni2+/nucleotide phosphate. Birefringence is measured as a function of electric field strength. At sufficiently low fields, the birefringence of all samples varies linearly with the square of the electric field, in accordance with the Kerr law. The value of the field strength at which the Kerr law is no longer obeyed increases with the addition of NiC1, ( E is of the order of 400 V . cm-I for B-DNA before addition of Ni++).
The birefringence of poly(dG-dC) is gradually reduced with increasing Ni2+ concentration, as shown by the values of the Kerr constant B [Fig. 6(A)]. B changes from - 1.3 esu cgs in the B-form to about - 0.5 esu cgs a t y = 2. This 60% decrease corresponds to the formation of TI-DNA.
A similar finding is observed for the titration of poly(dG-m5dC), except that Fig. 6(A) reveals an 82-85% decrease in birefringence between y = 0 and y = 2. The formation of Z-DNA for a stoichiometry of 1 cation/4 nucleotide phosphates, as observed by CD [Fig. 2(A)], corresponds to a decrease in birefringence of about 22-23%.
For y > 2, the birefringence of both methylated and unmethylated poly- mers is very low, becoming practically zero at y = 5.
Orientational Relaxation Time Analysis
Figure 5 shows typical semilog decay plots for poly(dG-dC) and poly(dG-m5dC) with increasing Ni2+ concentrations. As expected, decays are
2288 HACQUES AND MARION
c. (J a, c
> a c. v
8 a U
8 -I
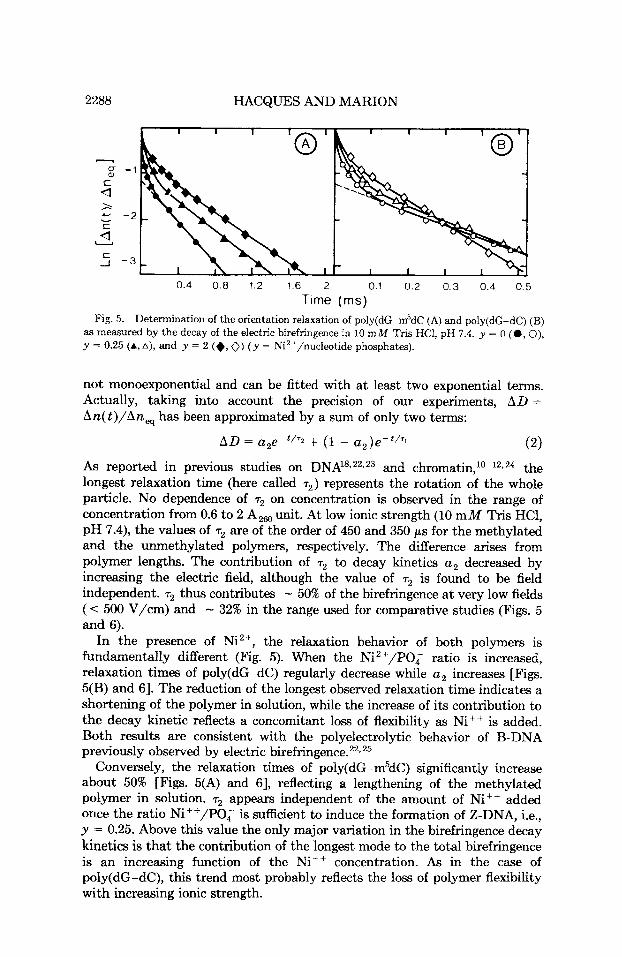
0.4 0.8 1.2 1.6 2 0.1 0.2 0.3 0.4 0.5 Time (rns)
Fig. 5. Determination of the orientation relaxation of poly(dG-m5dC (A) and poly(dG-dC) (B) as measured by the decay of the electric birefringence in 10 m M Tris HCl, pH 7.4. y = 0 (0 , 0), y = 0.25 (A, A), and y = 2 (+, 0) ( y = Ni2+/nucleotide phosphates).
not monoexponential and can be fitted with a t least two exponential terms. Actually, taking into account the precision of our experiments, AD = An(t)/An, has been approximated by a sum of only two terms:
AD = a2eP'/'2 + (1 - u2)e-'/'1 (2)
As reported in previous studies on DNA18s22r23 and c h r ~ m a t i n , ~ ~ - ~ ~ . ~ ~ the longest relaxation time (here called r2) represents the rotation of the whole particle. No dependence of r2 on concentration is observed in the range of concentration from 0.6 to 2 A,, unit. At low ionic strength (10 m M Tris HC1, pH 7.4), the values of r2 are of the order of 450 and 350 p s for the methylated and the unmethylated polymers, respectively. The difference arises from polymer lengths. The contribution of r2 to decay kinetics u2 decreased by increasing the electric field, although the value of r2 is found to be field independent. r2 thus contributes - 50% of the birefringence a t very low fields (< 500 V/cm) and - 32% in the range used for comparative studies (Figs. 5 and 6).
In the presence of Ni2+, the relaxation behavior of both polymers is fundamentally different (Fig. 5). When the Ni2+/POq ratio is increased, relaxation times of poly(dG-dC) regularly decrease while u2 increases [Figs. 5(B) and 61. The reduction of the longest observed relaxation time indicates a shortening of the polymer in solution, while the increase of its contribution to the decay kinetic reflects a concomitant loss of flexibility as Ni++ is added. Both results are consistent with the polyelectrolytic behavior of B-DNA previously observed by electric birefringence.22T25
Conversely, the relaxation times of poly(dG-m5dC) significantly increase about 50% [Figs. 5(A) and 61, reflecting a lengthening of the methylated polymer in solution. r2 appears independent of the amount of Nit+ added once the ratio Ni++/PO, is sufficient to induce the formation of Z-DNA, i.e., y = 0.25. Above this value the only major variation in the birefringence decay kinetics is that the contribution of the longest mode to the total birefringence is an increasing function of the Ni++ concentration. As in the case of poly(dG-dC), this trend most probably reflects the loss of polymer flexibility with increasing ionic strength.
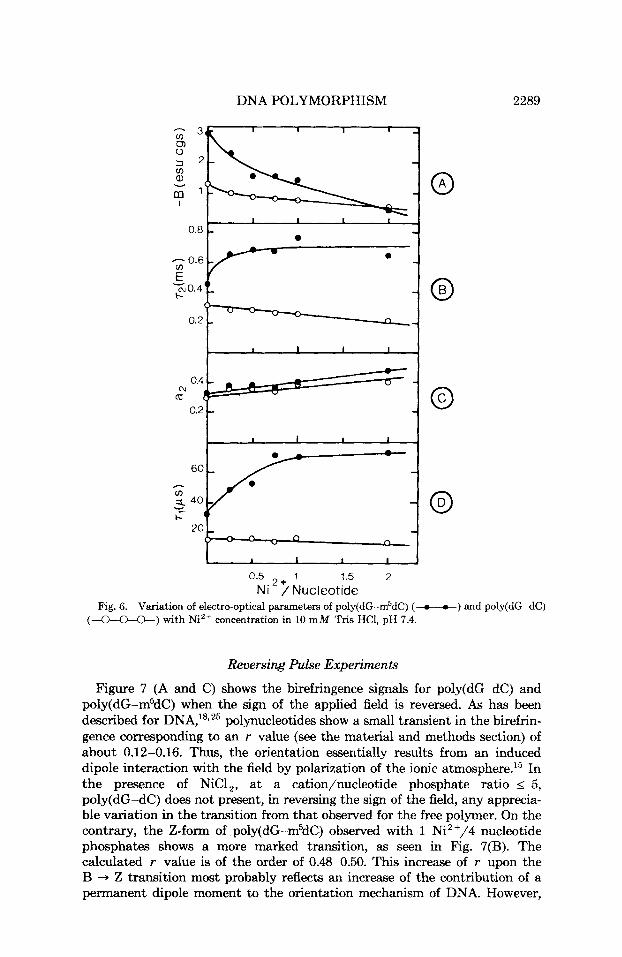
DNA POLYMORPHISM 2289
0.5 + 1 1.5 2 Ni / Nucleotide
Fig. 6. Variation of electro-optical parameters of poly(dG-m5dC) (-o-o-) and poly(dG-dC) (a*) with Ni2+ concentration in 10 m M Tris HCI, pH 7.4.
Reuersing Pulse Experiments
Figure 7 (A and C) shows the birefringence signals for poly(dG-dC) and poly(dG-m5dC) when the sign of the applied field is reversed. A s has been described for DNA,'8*25 polynucleotides show a small transient in the birefrin- gence corresponding to an r value (see the material and methods section) of about 0.12-0.16. Thus, the orientation essentially results from an induced dipole interaction with the field by polarization of the ionic atm0~phere.l~ In the presence of NiCI,, at a cation/nucleotide phosphate ratio I 5, poly(dG-dC) does not present, in reversing the sign of the field, any apprecia- ble variation in the transition from that observed for the free polymer. On the contrary, the Z-form of poly(dG-m5dC) observed with 1 Ni2+/4 nucleotide phosphates shows a more marked transition, as seen in Fig. 7(B). The calculated r value is of the order of 0.48-0.50. This increase of r upon the B + Z transition most probably reflects an increase of the contribution of a permanent dipole moment to the orientation mechanism of DNA. However,
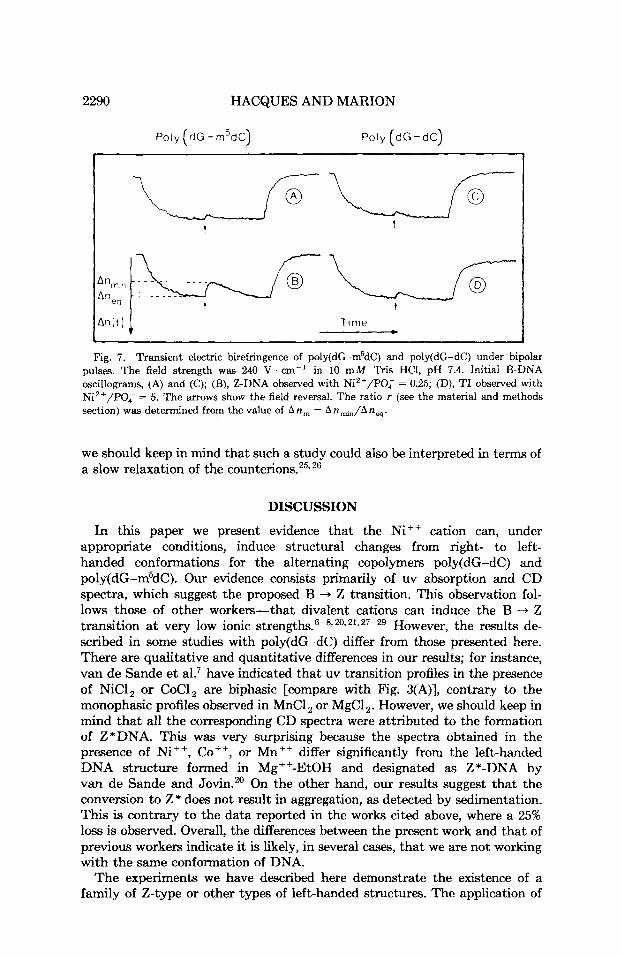
2290 HACQUES AND MARION
Poly (dG - m5dC) Poly (dG-dC) ~
u L fi- t t
Fig. 7. Transient electric birefringence of poly(dG-m5dC) and poly(dG-dC) under bipolar pulses. The field strength was 240 V . cm-' in 10 mM ' k i s HCI, pH 7.4. Initial B-DNA oscillograms, (A) and (C); (B), Z-DNA observed with Ni2+/P0; = 0.25; (D), TI observed with Ni2+/PO; = 5. The mows show the field reversal. The ratio r (see the material and methods section) was determined from the value of An,,, = An~JAn, .
we should keep in mind that such a study could also be interpreted in terms of a slow relaxation of the counter ion^.'^.^^
DISCUSSION
In this paper we present evidence that the Ni++ cation can, under appropriate conditions, induce structural changes from right- to left- handed conformations for the alternating copolymers poly(dG-dC) and poly(dG-m5dC). Our evidence consists primarily of uv absorption and CD spectra, which suggest the proposed B -+ Z transition. This observation fol- lows those of other workers-that divalent cations can induce the B -+ Z transition a t very low ionic strengths.6-8,20,21,27-29 However, the results de- scribed in some studies with poly(dG-dC) differ from those presented here. There are qualitative and quantitative differences in our results; for instance, van de Sande et al? have indicated that uv transition profiles in the presence of NiC1, or CoCl, are biphasic [compare with Fig. 3(A)], contrary to the monophasic profiles observed in MnCl , or MgCl 2. However, we should keep in mind that all the corresponding CD spectra were attributed to the formation of Z*DNA. This was very surprising because the spectra obtained in the presence of Ni++, CO++, or Mn++ differ significantly from the left-handed DNA structure formed in Mg++-EtOH and designated as Z*-DNA by van de Sande and Jovin." On the other hand, our results suggest that the conversion to Z* does not result in aggregation, as detected by sedimentation. This is contrary to the data reported in the works cited above, where a 25% loss is observed. Overall, the differences between the present work and that of previous workers indicate it is likely, in several cases, that we are not working with the Same conformation of DNA.
The experiments we have described here demonstrate the existence of a family of Z-type or other types of left-handed structures. The application of

DNA POLYMORPHISM 2291
steady-state and transient electric birefringence to the characterization of the right-to-left-handed transition provides information on the different electro- optical properties exhibited by DNA in solution. Such a study has never been reported before.
The B -+ Z transition in poly(dG-m5dC) occurs at a stoichiometry of Ni++ of - 1 cation/4 nucleotide phosphates. The transition results in an increase in the longest relaxation time of about 45%, corresponding to a lengthening of the polymer in solution. This findinp of changes in length is in very *good agreement with helical pitches of 34 A (10.5 bp) in the B-form and 44.6 A (12 bp) in the Z-form, respectively,3o and with resul? showing an increase in the rise per base pair of 9% going from B-form (3.4 A) to Z-form (3.7 A) DNA.I3 The increase in length is accompanied by an increase of rigidity since the contribution of r2 to total birefringence increases. This finding is consistent with theoretical data,31*32 and static and dynamic laser light-scattering stud-
A particularly interesting result, never reported before is that obtained when the sign of the applied field is reversed. However, at this stage we cannot give sufficient evidence to explain the large transient observed in the presence of Z-DNA through a permanent dipole orientation mechanism. Theories26*34 are useful for rigid cylindrical molecules but treatment of flexible molecules has not been considered in the literature, and we should recall that the transient may also be ascribed to a slowly induced dipole moment caused by the relaxation of the counterions. Determination of the role played by the different dipole moments in governing the orientation of Z-DNA requires much additional data.
The left-handed DNA formed in the presence of higher amounts of Ni++ has spectroscopic characteristics reported for Z*.20 The tendency for Z to aggregate has been reported and the formation of Z*-form appeared an indication of condensation and intermolecular aggregation?.20 We should recall that the large negative anisotropy of DNA reflects the arrangement of its base pairs perpendicular to the helix axis.15 It is admitted that a positive anisotropy may arise either from superhelical arrangements of DNA in com- pact structure or from irregular aggregati~n.~~ We present here evidence of the existence in the concentration range of 0.25 to 5 Ni2+/PO; of a Z*-type exhibiting a negative birefringence without the appearance of turbidity effects. The Z -+ Z* transition is not accompanied by a change in the longest relaxation time. r2 remains constant and equal to that of Z-DNA. The only variation in birefringence kinetics in this salt range is a regular increase of a2 as Ni '+ is added, indicating a loss of flexibility. A t the largest Ni++ concentra- tions a positive birefringence is observed that likely corresponds to the formation of supramolecular forms of Z-DNA, as discussed recently by Castleman et ai.I9
In the presence of NiCI,, the unmethylated polymer poly(dG-dC) is also found to undergo optical changes consistent with transitions to left-handed helical forms. This transition shows four different features from the case of poly(dG-m5dC) discussed above: First, the critical salt concentration neces- sary for the formation of a Z-like structure is much higher than those needed for the methylated polymer. Second, none of the left-handed forming CD spectra is a canonical Z-form spectrum. Third, left-handed forms of

2292 HACQUES AND MARION
poly(dG-dC) are accompanied by a regular decrease in orientational relaxa- tion time, indicating a shortening of the polymer. Finally, the field-reversal experiments show no modification in the orientation mechanism of DNA upon addition of Ni++.
CD data suggest two left-handed DNA conformations different from Z. The one that we designate as a transition intermediate (TI) is neither B- nor Z-like, and corresponds to a completely nonconservative shape of CD, already observed in the presence of Co++.'l The formation of TI results in a decrease in r2 by about 38-40%. As for its methylated analog a2 increases, indicating an increase in the chain stiffness of the TI-form compared to the right-handed B-form. The other structure that we designate as Z"-DNA is observed in the presence of higher concentrations of Ni++ ( y 2 5). Poly(dG-dC) exhibits a CD spectrum resembling but not totally identical to that of Z* formed in Mg2'-EtOH.20. In the Z"-DNA spectrum the positions of the maxima (except for the negative band at 295 nm) are slightly shifted (compare -tt in Fig. 2A and B). This difference makes the assertion that both spectra arise from the same left-handed structure questionable. The field-free decay results also favor the existence of two different conformations. We observe a shortening of poly(dG-dC) with added Ni++. If the end result of titration is the formation of Z*, why should it have such a different behavior from the Z*-form of poly(dG-m5dC) where the relaxation time increases, indicating a lengthening of polymer?
In conclusion, the existence of left-handed DNA sequences that possibly play important roles in the control of both chromatin structure and gene transcriptional activity% may be promoted by small amounts of divalent cations such as Ni++. However, the observed differences in CD spectra and electro-optical properties indicate that the corresponding conformations are probably not identical but form a part of the highly polymorphic family of left-handed helices. All structural transitions are found to be reversible. We are currently examining how these different forms may significantly affect the interactions of DNA with various histone H1 subtypes. HI is essential for the formation of the 30-nm chromatin fiber from the extended 10-nm chain of nucleosomes, and it seems possible that higher order structures may be responsible, a t least in part, for tissue-specific gene upp press ion.^^
We wish to thank B. Roux, with whom this work was initiated. We also acknowledge the careful examination of the manuscript by the referees of Bwpolymers, as well as their fruitful suggestions. This work was supported by the Centre National de la Recherche Scientifique (UM 24).
References 1. Rich, A., Nordheim, A. & Wang, A. H. 3. (1984) Ann. Rm. Biochem. 53, 791-846. 2. Wang, A. H. J., Quigley, G. J., Kolpak, F. J., Crawford, J. L., van Boom, J. H.,
van der Marel, G. & Rich, A. (1979) Nature 282, 680-686. 3. Pohl, F. M. & Jovin, T. M. (1972) J. Mol. Bwl. 67, 375-396. 4. Doerfler, W. (1983) Ann. Rev. Bbchem. 52,93-124. 5. Razin, A. & Riggs, A. D. (1980) Science 210, 604-610. 6. Behe, M. & Felsenfeld, G. (1981) Proc. Nutl. Acud. Sci. USA 78, 1619-1623. 7. van de Sande, J. H., McIntosh, L. P. & Jovin, T. M. (1982) EMBO J . 1, 777-782. 8. Chen, H. H., Behe, M. J. & Rau, D. C. (1984) Nucleic Aclds Res. 12, 2381-2389.

DNA POLYMORPHISM
9. Thomas, T. J., Bloomfield, V. A & Canellakis, Z. N. (1985) Biopolymrs 24, 725-729. 10. Roche, J., Marion, C., Gorka, C., Roux, B. & Lawrence, J. J. (1984) Biochem. Bwphys. Res.
11. Marion, C., Roche, J., RQUX, B. & Gorka, C. (1985) Biochemistry 24, 6328-6335. 12. Marion, C., Martinage, A., Tirard, A., Roux, B., Dame, M. & Mazen, A. (1985) J . Mol.
13. Chen, H. H., Charney, E. & Rau, D. C. (1982) Nucleic Aclds Res. 10,3561-3571. 14. Wu, H. M., Dattagupta, N. & Crothers, D. M. (1981) Proc. Natl. Acad. Sci. USA 78,
15. Fredericq, E. & Houssier, C. (1973) Electric Dichroism and Electric Birefringence, Claren-
16. Bemengo, J. C., Roux, B. & Hans, M. (1973) Reo. Sci. Znstrum. 44, 1083-1086. 17. Marion, C., RQUX, B. & Bemengo, J. C. (1984) Makromol. Chem. 185, 1649-1664. 18. Roux, B., Bemengo, J. C., Marion, C. & Hans, M. (1978) J . Colloid Interface Sci. 66,
19. Castleman, H., Specthrie, L., Makowski, L. & Erlanger, B. F. (1984) J. Bwmol. Struct.
20. van de Sande, J. H. & Jovin, T. M. (1982) EMBO J. 1, 115-120. 21. Zacharias, W., Larson, J. E., Klysik, J., Stirdivant, S. M. & Wells, R. D. (1982) J . Biol.
22. Marion, C., Roux, B. & Hans, M. (1983) Bwpolymrs 22, 2353-2366. 23. Rau, D. C. & Bloomfield, V. A. (1979) Biopolymrs 18, 2783-2805. 24. Chauvin, F., Roux, B. & Marion, C. (1985) J . Biomol. Struct. Dyn. 2,805-819. 25. Marion, C., Roux, B., Bemengo, J. C. & Hans,. M. (1980) Znt. J. Biol. Macromol. 2,
26. Takezoe, H. & Yu, H. (1981) Biochemistry 20, 5275-5281. 27. Taillandier, E., Taboury, J. A., Adam, S. & Liquier, J. (1984) Biochemistry 23, 5703-5706. 28. Behe, M. J., Felsenfeld, G., Szu, S. C. & Charney, E. (1985) Bwpolymrs 24, 289-300. 29. Woisard, A., Fazakerley, V. & Guschlbauer, W. (1985) J . Bbmol. Struct. Dynam. 2,
30. Zimmerman, S. B. (1982) Ann. Rev. Bwchem. 51, 395-427. 31. Mitra, C. K., Sanna, M. H. & Sarma, R. H. (1981) J. Am. Chem. SOC. 103,6727-6737. 32. Kollman, P., Weiner, P., Quigley, G. & Wang, A. (1982) Biopolymrs 21, 1945-1969. 33. Thomas, T. J. & Bloomfield, V. A. (1983) Nucleic Acids Res. 11, 1919-1930. 34. Tinoco, I. Jr. & Yamaoka, K. (1959) J . Phys. Chem. 63,423-427. 35. Houssier, C., Lasters, I., Muyldermans, S. & Wyns, L. (1981) Nucleic Ac& Res. 9,
36. Reeves, R. (1984) Biochim. Biophys. Acta 782, 343-393. 37. Weintraub, H. (1984) Cell 38, 17-27.
Commun. 121, 530-537.
Bid . 186, 367-379.
6808- 681 1.
don Press, Oxford, England.
421-427.
Dynam. 2,271-283.
Chem. 257,2775-2782.
235-240.
1205- 1220.
5763-5784.
Received November 7, 1985 Accepted June 16, 1986