distribution of neuropeptide y y1 receptors in rodent peripheral tissues
TRANSCRIPT

Distribution of Neuropeptide Y Y1Receptors in Rodent Peripheral Tissues
HIDEKI MATSUDA,1 PABLO R. BRUMOVSKY,1 JUTTA KOPP,1
THIERRY PEDRAZZINI,2AND TOMAS HOKFELT1*
1Department of Neuroscience, Karolinska Institutet, Retzius Laboratory,S-171 77 Stockholm, Sweden
2Division of Hypertension, University of Lausanne Medical School,CH-1011 Lausanne, Switzerland
ABSTRACTUsing a sensitive immunohistochemical technique, the localization of neuropeptide Y
(NPY) Y1-receptor (Y1R)-like immunoreactivity (LI) was studied in various peripheral tis-sues of rat. Wild-type (WT) and Y1R-knockout (KO) mice were also analyzed. Y1R-LI wasfound in small arteries and arterioles in many tissues, with particularly high levels in thethyroid and parathyroid glands. In the thyroid gland, Y1R-LI was seen in blood vessel wallslacking �-smooth muscle actin, i.e., perhaps in endothelial cells of capillaries. Larger arterieslacked detectable Y1R-LI. A distinct Y1R-immunoreactive (IR) reticulum was seen in the WTmouse spleen, but not in Y1R-KO mouse or rat. In the gastrointestinal tract, Y1R-positiveneurons were observed in the myenteric plexus, and a few enteroendocrine cells were Y1R-IR.Some cells in islets of Langerhans in the pancreas were Y1R-positive, and double immuno-staining showed coexistence with somatostatin in D-cells. In the urogenital tract, Y1R-LI wasobserved in the collecting tubule cells of the renal papillae and in some epithelial cells of theseminal vesicle. Some chromaffin cells of adrenal medulla were positive for Y1R. The problemof the specificity of the Y1R-LI is evaluated using adsorption tests as well as comparisonsamong rat, WT mouse, and mouse with deleted Y1R. Our findings support many earlierstudies based on other methodologies, showing that Y1Rs on smooth muscle cells of bloodvessels mediate NPY-induced vasoconstriction in various organs. In addition, Y1Rs in othercells in parenchymal tissues of several organs suggest nonvascular effects of NPY via theY1R. J. Comp. Neurol. 449:390–404, 2002. © 2002 Wiley-Liss, Inc.
Indexing terms: G-protein-coupled receptor (GPCR); immunohistochemistry; rat
(Sprague-Dawley); mouse (C57BL/6); knockout mouse
Neuropeptide Y (NPY), a 36-amino acid peptide (Tate-moto, 1982; Tatemoto et al., 1982) widely distributed inboth the central and peripheral nervous systems (CNS,PNS), has been functionally related to regulation of bloodpressure, circadian rhythms, feeding behavior, anxiety,memory processing, and cognition in the CNS, and tovasoconstriction and gastrointestinal tract motility in thePNS (Mutt et al., 1989; Allen and Koenig, 1992). Two NPYreceptors, the NPY Y1 and the NPY Y2 receptor (Y1R andY2R), were early recognized based on biochemical, bind-ing, and pharmacologic experiments (Unden et al., 1984;Michel, 1991; Dumont et al., 1992; Grundemar and Ha-kanson, 1993; Wahlestedt and Reis, 1993; Gehlert, 1994)and were classified as post- and presynaptic, respectively(Wahlestedt et al., 1986). The Y1R was cloned first (Eva etal., 1990, 1992; Herzog et al., 1992; Larhammar et al.,1992), followed by the Y2R (Gerald et al., 1995; Rose et al.,
1995); subsequently further NPY receptors were identi-fied, cloned, and characterized (Larhammar, 1996; Bala-
Grant sponsor: Marianne and Marcus Wallenberg’s Foundation; Grantsponsor: Knut and Alice Wallenberg’s Foundation; Grant sponsor: Bristol-Myers Squibb (Unrestricted Neuroscience Grant); Grant sponsor: theSwedish Medical Research Council; Grant number: 04X-2887; Grant spon-sor: the Swedish Institute.
*Correspondence to: Tomas Hokfelt, Department of Neuroscience B3:4,Karolinska Institutet, Retzius Vag 8, S-171 77 Stockholm, Sweden.E-mail: [email protected]
Received 23 October 2001; Revised 17 January 2002; Accepted 20 April2002
DOI 10.1002/cne.10303Published online the week of June 17, 2002 in Wiley InterScience (www.
interscience.wiley.com).
THE JOURNAL OF COMPARATIVE NEUROLOGY 449:390–404 (2002)
© 2002 WILEY-LISS, INC.

subramaniam, 1997; Blomqvist and Herzog, 1997; Geh-lert, 1998; Cabrele and Beck-Sickinger, 2000).
So far comparatively few studies have dealt with thelocalization of NPY receptors in the periphery. Initiallythe distribution of NPY receptors was studied with auto-radiographic ligand binding technique (Leys et al., 1987;Sheikh et al., 1991; Walsh et al., 1993), but cloning of theY1R has allowed both in situ hybridization studies andimmunohistochemical analysis. On the basis of informa-tion from the cloning studies, antibodies can be raisedagainst various peptide sequences of the receptor protein.Thus, Y1R mRNA and/or Y1R-like immunorectivity (LI)have been localized in dorsal root ganglia (Jazin et al.,1993; Zhang et al., 1994a,b, 1999; Brumovsky et al., 2002)and smooth muscle cells in cerebral blood vessels (Bao etal., 1997) and in the genital tract (Kopp et al., 1997), aswell as in neurons and enteroendocrine cells of the intes-tinal tract (Peaire et al., 1997; Jackerott and Larsson,1997).
In the present study, we have analyzed the distributionof Y1R-LI in various peripheral tissues with an antibodyraised against the C-terminal portion of the rat receptor(Kopp et al., 2002). Sensory neurons were not included,because they already have been studied in detail (for ref-erences, see above). In an attempt to evaluate the speci-ficity of this antibody, we compared these results withthose obtained in adsorption tests and in studies of wild-type (WT) and Y1R-knockout (KO) mice (Pedrazzini et al.,1998).
MATERIALS AND METHODS
Animals
A total of 22 adult male and 7 adult female Sprague-Dawley rats (body weight 250–300 g; B & K Universal,Stockholm, Sweden) were used. In addition, Y1R-KO (Ped-razzini et al., 1998) and WT (C57BL/6) mice (for bothgroups n � 6 males and 4 females) were studied. Allanimals were anesthetized with sodium pentobarbital (60mg/kg, i.p.) and perfused through the heart with calcium-free Tyrode’s solution and 4% para-formaldehyde in 0.16M phosphate buffer containing 0.2% picric acid (pH 6.9) at37°C, followed by the same ice-cold solution. The organs ofthe cardiovascular, lymphatic, integumentary, digestive,respiratory, urinary, endocrine, and reproductive systemswere dissected out, immersed in the same fixative for 90minutes at 4°C, and then transferred to phosphate-buffered saline (PBS) containing 10% sucrose (pH 7.2),0.02% bacitracin (Sigma, St. Louis, MO), and 0.01% so-dium azide (Merck, Darmstadt, Germany) for 24 hours at4°C. Frozen sections were cut at 14 �m in a cryostat(Microm, Heidelberg, Germany) and thaw-mounted ontochrome alum/gelatin-coated object slides. The local ethicscommittee approved the experiments (Stockholms norradjurforsoksetiska namnd, project N90/99), and animalswere treated according to the Swedish animal welfarelaws.
Immunohistochemistry
Sections were processed for immunohistochemistry us-ing the catalyzed receptor deposition method (Adams,1992) and a commercial kit [indirect tyramide signal am-plification (TSA); NEN Life Science Products, Boston,MA]. A rabbit polyclonal antiserum against a 13-amino
acid peptide of the C-terminal part of the rat Y1R (Kopp etal., 2002) was diluted 1:4,000 in 0.01 M PBS containing0.3% Triton X-100 and 0.5% bovine serum albumin (BSA).Briefly, the sections were rehydrated in PBS, pretreatedwith 0.03% H2O2 for 30 minutes at room temperature(RT), and incubated with diluted primary antiserum in ahumid chamber for 24 hours at 4°C. After rinsing in TNTbuffer (0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.05% Tween20) for 10 minutes, the sections were preincubated in TNBbuffer (kit; 0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.5%DuPont Blocking Reagent; NEN) for 30 minutes at RT andincubated with horseradish peroxidase-conjugated swineanti-rabbit immunoglobulin (Dako, Copenhagen, Den-mark) diluted at 1:100 in TNB buffer for 30 minutes at RT.They were washed three times for 10 minutes in TNTbuffer and incubated with biotinyl tyramide (NEN) di-luted at 1:100 in amplification diluent for 16 minutes; allsteps were at RT. After washing three times in TNTbuffer, chromogenic detection of the deposited biotin wascarried out by incubation with fluorescein-conjugatedstreptavidin (NEN), diluted at 1:100 in TNB buffer, for 30minutes at RT. Finally, the sections were rinsed in TNTbuffer.
In some cases, double immunostaining was carried out.Thus, in a second sequence, the sections were processedfor the indirect immunofluorescence technique (Coons,1958). The sections were incubated with a mouse mono-clonal antibody to somatostatin (SOM; Wong et al., 1990;pancreas, 1:800), a mouse monoclonal antibody to�-smooth muscle actin (�-SMA; Sigma; thyroid gland,1:400), a mouse monoclonal antibody to NPY (Grouzmannet al., 1992; adrenal gland, 1:400), or a sheep antiserum tobovine phenylethanolamine N-methyltransferase (PNMT;Goldstein et al., 1971; adrenal gland, 1:600) for 24 hoursat 4°C followed by anti-mouse or sheep rhodamine-redX-conjugated secondary antibody (1:80; Jackson Immu-noResearch, West Grove, PA) for 40 minutes at 37°C.
All sections were coverslipped with a mixture of glyceroland PBS (9:1) containing 0.1% para-phenylenediamine(Sigma). They were examined in a Nikon (Tokyo, Japan)Microphot-FX fluorescence microscope equipped withproper filter combinations. The sections were also ana-lyzed in a Bio-Rad Radiance Plus confocal scanning mi-croscope (Bio-Rad, Hemel Hempstead, UK) installed on aNikon Eclipse E600 fluorescence microscope equippedwith 10�/0.45 N.A. and 20�/0.75 N.A. objectives. Thefluorescein labeling was excited using the 488-nm line ofthe argon ion laser and detected after passing an HQ530/60 (Bio-Rad) emission filter. For detection of therhodamine-red X signal, the 543-nm line of the greenHeNe laser was used in combination with the HQ 590/70(Bio-Rad) emission filter.
Controls
The specificity of the Y1R antiserum was tested in twodifferent ways. First, the antibody was preadsorbed withan excess (10�6 or 10�5 M) of a synthetic Y1R peptideconstituting amino acids 358–382 of the Y1R protein(kindly provided by the late Dr. J.H. Walsh and by Dr.H.C. Wong, UCLA, Los Angeles, CA) for 24 hours at 4°C,and the slides were processed for the indirect TSA methodas described above. Then tissues from Y1R-KO and WTmice processed for the indirect TSA method were ana-lyzed.
391NPY-Y1 RECEPTORS IN PERIPHERY

Digital image processing
Digital images obtained from confocal laser scanningmicroscopy were optimized for image resolution, bright-ness, and contrast using Adobe Photoshop 5.5 (Adobe Sys-tems, San Jose, CA). Images were not altered in any way,such as by removing or adding details.
RESULTS
Cardiovascular system
Y1R-LI was observed in tunica media of middle-sizedarteries, small arteries, and arterioles (diameter up to 200�m in rat). Smooth muscle cells of small arteries wereY1R-immunoreactive (IR) in the WT mouse heart (Fig.1A); however, no Y1R-LI was detected in the Y1R-KOmouse heart (Fig. 1B) and in the rat heart. Cardiac musclecells were Y1R-negative in both species. Blood cellsshowed strong fluorescence in all rat and WT mouse (Fig.1A) tissues, but this was also seen in the Y1R-KO mouse(Fig. 1B) and after incubation with preadsorbed anti-serum.
Lymphatic system
Y1R-LI was seen in a few arterioles in the rat spleen(Fig. 1C). In the WT mouse spleen, Y1R-LI was found in areticulum in the white pulp (Fig. 1D). This signal couldnot be seen in the spleen of the rat (Fig. 1C) or of theY1R-KO mouse (Fig. 1E), or in the WT mouse spleen afterincubation with control serum. There was no detectableY1R-LI in the thymus.
Integumentary system
Y1R-LI was found in small arteries and arterioles in thedermis and hypodermis of the auricle (Fig. 1F) and in thesole of the hind paw in both the rat and WT mouse.Striated muscle cells in the skin of the auricle and the soleof the hind paw of the rat were Y1R-positive (Fig. 1F); thisimmunoreactivity disappeared after adsorption (Fig. 1G).Y1R-LI was not detected in hair follicles. Sebaceousglands still showed fluorescence after incubation withpreadsorbed antiserum (Fig. 1G).
Digestive system
In salivary glands, Y1R-LI was observed in small arter-ies and arterioles (diameter between 50 and 100 �m) inrat parotid, submandibular, and sublingual glands (Fig.2A). However, no acinus or duct cells were Y1R-IR. Therewas no apparent difference in Y1R distribution pattern inthese three major salivary glands. Y1R-LI was not de-tected after incubation with preadsorbed antiserum(Fig. 2B).
In the gastrointestinal tract, Y1R-LI was observed insmall arteries and arterioles in the lamina propria, sub-mucosa, and smooth muscle layer throughout the rat gas-trointestinal tract (Fig. 2C). The diameter of these Y1R-positive vessels varied between 15 and 200 �m. Largerarteries did not have detectable Y1R-LI (Fig. 2E). A fewenteroendocrine cells in the stomach and small intestinewere Y1R-positive (Fig. 2F). Y1R-LI was found in manynerve cell bodies in the myenteric plexus of the stomachand small intestine (Fig. 2G). Nerve fibers positive for Y1Rwere also seen in the smooth muscle layer. However, nopositive cell bodies were found in the submucosal plexus.Y1R-LI observed in the gastrointestinal tract was abol-
ished by preincubation with synthetic peptide (Fig. 2D)and was seen in WT but not Y1R-KO mice.
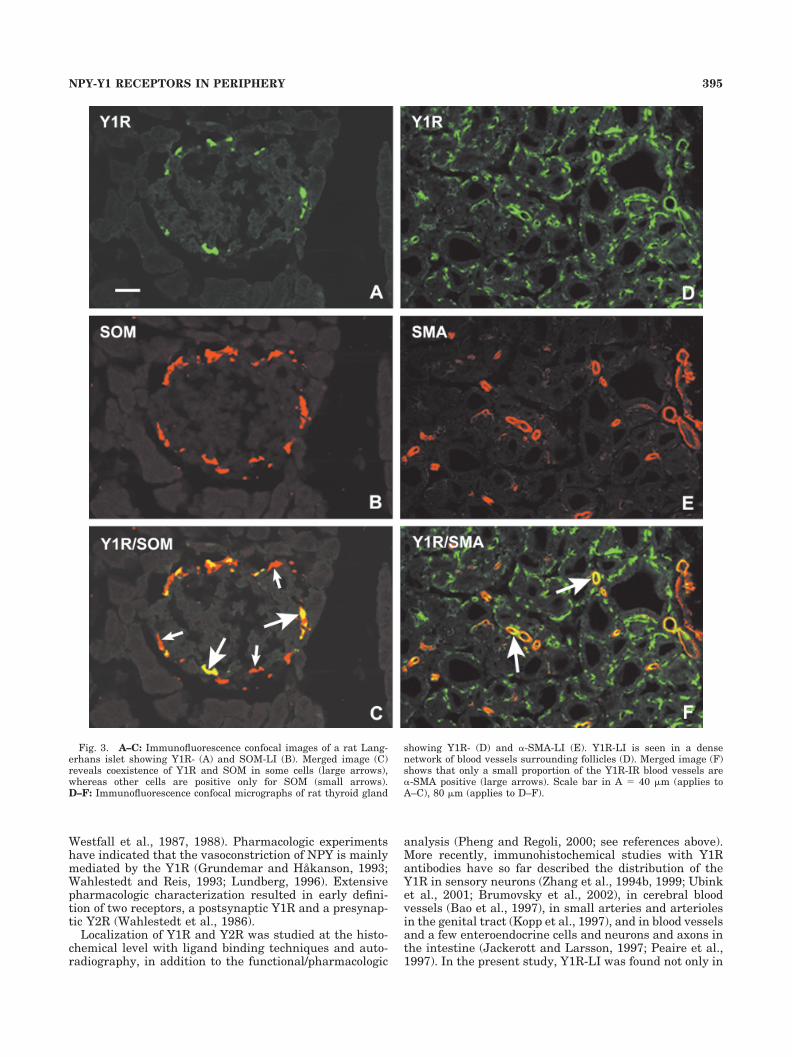
In the pancreas, Y1R-LI was seen in blood vessels in ratpancreas, but acinar cells and duct cells were negative.Some cells in islets of Langerhans of rat were Y1R-positiveand were mainly located at the periphery of the islets (Fig.3A). Double immunostaining revealed coexistence ofY1R-LI and SOM in these islets cells (Fig. 3C), althoughthe SOM-IR cells were more numerous than Y1R-IR cells(cf. Fig. 3A with B), but all Y1R-IR cells contained SOM.Approximately one-third of the SOM cells exhibitedY1R-LI.
Y1R-LI was not observed in the liver.
Respiratory system
Strong Y1R-LI was observed in blood vessels in thelamina propria of the rat trachea (Fig. 4A) and WT mousetrachea but not in the lung.
Urinary system
Many tubular cells in the renal papilla of rat and WTmouse (Fig. 4B,C) were Y1R-positive, with a stronger flu-orescence in mouse. This Y1R-LI could not be observedafter incubation with control serum or in Y1R-KO mouse(Fig. 4D). In WT (Fig. 4E) but not Y1R-KO (Fig. 4F) mice,a small number of cells in the wall of arterioles near theglomeruli were Y1R-IR, probably corresponding to juxta-glomerular cells. However, Y1R-LI could not be detectedin the juxtaglomerular apparatus of rat.
Y1R-LI was seen in arterioles in lamina propria andmuscle layers of the bladder of rat (Fig. 4G) and WTmouse, but bladder smooth muscle cells were Y1R-negative. Y1R-LI was not detected after incubation withpreadsorbed antiserum (Fig. 4H) or in Y1R-KO mouse.
Endocrine organs and autonomic ganglia
Very strong Y1R-LI was found in dense networks ofcapillaries surrounding follicles in the thyroid gland of rat(Figs. 3D, 5A) and WT mouse, as well as in capillaries inthe parathyroid gland (Fig. 5C). Y1R-LI in both the thy-roid (Fig. 5B) and parathyroid gland (Fig. 5D) was abol-ished by preincubation with synthetic Y1R peptide, and noY1R-LI was seen in these glands of Y1R-KO mouse. Dou-ble immunostaining revealed that only a small proportionof the Y1R-IR blood vessels were �-SMA-positive. (Fig.3D–F).
Y1R-LI was seen in both the cortex and medulla of therat adrenal gland. In the cortex, it was found in capsulararterioles and capillaries (Fig. 6A) but not in any of thesteroid hormone-producing endocrine cells of the threecortical zones (Fig. 6A,C). In the medulla, several cells,mostly forming small groups, expressed Y1R-LI (Fig. 6C).Double immunostaining revealed both PNMT-positiveand -negative Y1R-IR chromaffin cells (Fig. 6E,F), andsome chromaffin cells were positive for both Y1R and NPY(Fig. 6G,H). All fluorescent signals disappeared after ad-sorbing Y1R antibody with synthetic peptide (Fig. 6B,D).
There was no detectable Y1R-LI in the pineal gland orin the superior cervical or sphenopalatine ganglia of ratand mouse.
Reproductive system
Y1R-LI was observed mainly in the wall of many bloodvessels in the male and female reproductive system, inboth rat and WT mouse, with in general a stronger signal
392 H. MATSUDA ET AL.

in rat than in WT mouse. In the testis, Y1R-LI was ob-served in the small arteries and arterioles around theseminiferous tubules (Fig. 7A), as well as in the connectivetissues around the ductus epididymidis (Fig. 7C). In thevas deferens, the small arteries and arterioles in the lam-ina propria and smooth muscle layer were Y1R-IR, but
larger arteries with a diameter above 200 �m exhibited nosignal (Fig. 7E). Y1R-LI was seen in blood vessels betweenthe tubuloacinar units of the prostate gland (Fig. 7G,H).Some epithelial cells of the seminal vesicle were Y1R-positive, but only in rat (Fig. 7J,K). In the female repro-ductive system, Y1R-LI was found in the small arteries
Fig. 1. Immunofluorescence micrographs of the mouse heart (A,B),the spleen of rat (C) and mouse (D,E), and the rat auricle (F,G).A,B: Y1R-LI is found in small arteries in the WT mouse heart (A);however, no Y1R-LI is seen in the Y1R-KO mouse heart (B). Cardiacmuscle cells are Y1R-negative. Arrows indicate blood vessels.C: Y1R-LI is seen in arterioles in the white pulp (arrows). No Y1R-LIis found in the red pulp (asterisk). D,E: Y1R-LI is found in a reticulumin the white pulp of the WT mouse spleen (D) but not in the Y1R-KO
mouse spleen (E). F,G: Y1R-LI is found in arterioles in dermis andhypodermis of the rat auricle (F). Striated muscle cells (smc) areY1R-positive. Y1R-LI in blood vessels and striated muscles disappearafter incubation with preadsorbed antiserum (G). Asterisks indicatecartilage. Sebaceous glands (arrows) show nonspecific fluorescence(F,G) as well as blood cells (arrowheads; A,B,C,E). Scale bar � 80 �min A (applies to A–G).
393NPY-Y1 RECEPTORS IN PERIPHERY

and arterioles in the uterus (Fig. 8A), the ovary (Fig. 8C),and the oviduct (Fig. 8E). None of these signals were seenafter incubation with preadsorbed antiserum (Figs. 7B,D,F,I,L, 8B,D,F).
Controls
Results from control experiments (adsorption test,Y1R-KO mouse) have in many cases already been reportedabove. However, if this was not done, the described immu-noreactivity is specific in the sense that it is abolished bypreadsorption, alternatively not present in the KO mouse.
DISCUSSION
Early on, it was found that NPY is closely associatedwith the sympathoadrenal system. Thus, NPY is colocal-
ized with noradrenaline in a considerable proportion ofneurons in sympathetic ganglia and their nerve terminals(Lundberg et al., 1982, 1983; Ekblad et al., 1984), as wellas in the adrenal medulla, where it is present both inchromaffin cells (Allen et al., 1983a; Lundberg et al., 1983)and in some ganglion cells (Schalling et al., 1988). Subse-quently it was observed that NPY is not confined to sym-pathetic neurons but could also be found in parasympa-thetic ganglia (Leblanc et al., 1987; Kuwayama et al.,1988). NPY exerts three effects at sympathetic neuroef-fector junctions: 1) a direct postjunctional response (vaso-constriction); 2) a postjunctional potentiation of the re-sponse to vasoconstrictors, such as noradrenaline; and 3) aprejunctional suppression of stimulated noradrenaline re-lease (Lundberg and Tatemoto, 1982; Edvinsson et al.,1984; Lundberg et al., 1985; Wahlestedt et al., 1985, 1987;
Fig. 2. Immunofluorescence micrographs of the rat digestive sys-tem. A,B: Y1R-LI is seen in arterioles in the parotid gland (A). Acinusand duct cells are Y1R-negative. Y1R-LI is abolished after incubationwith preadsorbed antiserum (B). Arrows indicate blood vessels.C–E: Y1R-LI is found in small arteries and arterioles (diameter up to200 �m) in the lamina propria, submucosa, and muscle layer in thestomach (C). Larger arteries lack detectable Y1R-LI (asterisks; C,E),
but a branch with a diameter of less than 200 �m shows Y1R-LI(arrowheads; E). Y1R-LI in vessels is abolished by preincubation withsynthetic peptide (D). F: Y1R-LI is seen in an enteroendocrine cell inthe jejunum. G: A cell body in the myenteric plexus of the stomach isY1R-positive. Scale bar � 80 �m in A (applies to A–D), 40 �m (appliesto E–G).
394 H. MATSUDA ET AL.
Westfall et al., 1987, 1988). Pharmacologic experimentshave indicated that the vasoconstriction of NPY is mainlymediated by the Y1R (Grundemar and Hakanson, 1993;Wahlestedt and Reis, 1993; Lundberg, 1996). Extensivepharmacologic characterization resulted in early defini-tion of two receptors, a postsynaptic Y1R and a presynap-tic Y2R (Wahlestedt et al., 1986).
Localization of Y1R and Y2R was studied at the histo-chemical level with ligand binding techniques and auto-radiography, in addition to the functional/pharmacologic
analysis (Pheng and Regoli, 2000; see references above).More recently, immunohistochemical studies with Y1Rantibodies have so far described the distribution of theY1R in sensory neurons (Zhang et al., 1994b, 1999; Ubinket al., 2001; Brumovsky et al., 2002), in cerebral bloodvessels (Bao et al., 1997), in small arteries and arteriolesin the genital tract (Kopp et al., 1997), and in blood vesselsand a few enteroendocrine cells and neurons and axons inthe intestine (Jackerott and Larsson, 1997; Peaire et al.,1997). In the present study, Y1R-LI was found not only in
Fig. 3. A–C: Immunofluorescence confocal images of a rat Lang-erhans islet showing Y1R- (A) and SOM-LI (B). Merged image (C)reveals coexistence of Y1R and SOM in some cells (large arrows),whereas other cells are positive only for SOM (small arrows).D–F: Immunofluorescence confocal micrographs of rat thyroid gland
showing Y1R- (D) and �-SMA-LI (E). Y1R-LI is seen in a densenetwork of blood vessels surrounding follicles (D). Merged image (F)shows that only a small proportion of the Y1R-IR blood vessels are�-SMA positive (large arrows). Scale bar in A � 40 �m (applies toA–C), 80 �m (applies to D–F).
395NPY-Y1 RECEPTORS IN PERIPHERY

smooth muscle cells of blood vessels but also in capillarywalls, in a reticulum in the mouse spleen, and in someepithelial and endocrine cells, as will be discussed below.The positive blood vessels were of various sizes and werepresent in many peripheral organs. However, large arter-ies with a diameter over 200 �m lacked detectable Y1R-LI.This is in agreement with the situation in rat cerebralblood vessels (Bao et al., 1997), as well as in the rat genitaltract (Kopp et al., 1997).
Negative immunohistochemical results should, how-ever, be interpreted with caution. Large vessels may havetoo low numbers of Y1Rs to be detected with our tech-nique, but these receptors may still be of functional sig-nificance. Taken together, it is highly likely that in manyinstances discussed below, in which discrepancies existbetween physiological/pharmacologic and our immunohis-
tochemical results, functional Y1Rs in fact are present butcannot be detected with our technique.
Because an electron microscopic study of cerebral bloodvessels has shown presence of Y1R on the peripheral aswell as endothelial side of the smooth muscle cell (Bao etal., 1997), vasoconstrictory Y1Rs appear to be accessibleby NPY originating from both surrounding nerves andblood-borne NPY.
Aspects of specificity
There is a considerable risk that antisera raised againstpeptide fragments of G protein-coupled receptors lackspecificity. A simple and necessary control is preadsorp-tion of the primary antiserum with an excess of the im-munogenic peptide. Even if this control is successful, spec-ificity cannot be guaranteed. A second approach is
Fig. 4. Immunofluorescence micrographs of the rat trachea (A),mouse kidney (B–F), and rat urinary bladder (G,H). A: Y1R-LI is seenin blood vessels (arrowheads) in lamina propria. Asterisk indicatescartilage. B: Almost all tubular cells in the renal papilla of WT mouseare Y1R-positive. C: Higher magnification of the WT mouse renalpapilla. D,F: Y1R-LI is not found in the Y1R KO mouse kidney. E: In
WT mouse Y1R-LI can be seen in cells (arrow) in the wall of arterioleclose to a glomerulus (g). G,H: Y1R-LI is seen in blood vessels (arrow-heads) of the rat urinary bladder (G), and it is abolished by preincu-bation with Y1R peptide (H). Scale bar in A � 40 �m (applies toA,C,E,F); 80 �m in A (applies to G,H), 200 �m in A (applies to B,D).
396 H. MATSUDA ET AL.

comparison with in situ hybridization, i.e., mRNA distri-bution. This has not been performed in the present study,but we will compare our results with such published datain the literature. A third approach is to study mice withthe deleted receptor (cf. Kopp et al., 2002). Here we havecompared a WT mouse with a Y1R-KO mouse created byPedrazzini et al. (1998). This approach requires that ourY1R antiserum raised against the C-terminal tridecapep-tide of the rat receptor protein cross-reacts with the mouseY1R and that the Y1R in principle has the same distribu-tion in rat and mouse. This is fortunately the case inseveral instances, but the overlap is not complete. Oneproblem is the possibility of truncated receptor, as hasbeen shown to be present in mouse (Nakamura et al.,1995). Such a truncated receptor may not cross-react withour antibody.
Another problem is the fact that the C-terminal aminoacid of the Y1R is not exactly identical in rat and mouse,and this difference may lead to a lower sensitivity of theantibody for mouse tissues, because this amino acid isincluded in the immunogenic peptide. As is evident fromthe present results, the Y1R immunostaining does notshow a total overlap between mouse and rat, i.e., in somecases staining is seen in mouse but not in rat and viceversa. For all these reasons, the Y1R-KO mouse does notrepresent a perfect specificity control for Y1R staining inrat. A similar situation has been encountered in a studyon rat brain, in which the same type of WT and Y1R-KOmice were used as specificity control (Kopp et al., 2002).Finally, Western blot analysis of receptor antisera is a
valuable test. However, in spite of several attempts in ourand another laboratory, we were not able to obtain con-vincing blots.
Cardiovascular system and muscle cells
Much interest has been focused on the role of NPYand NPY receptors in the cardiovascular system (Grun-demar and Hakanson, 1993; McDermott et al., 1993;Lundberg, 1996; Malmstrom, 1997; Franco-Cerecedaand Liska, 1998). In agreement, we observed Y1Rs inmuscle cells in many vascular beds. NPY inhibits thecontractile force and reduces the coronary flow in theisolated, perfused heart (Allen et al., 1983b), and NPYbinding sites have been demonstrated over the myocar-dium (Balasubramaniam, 1997). However, in our studymyocardial cells were not immunoreactive for Y1R, norcould Y1R-LI be detected even around coronary arteriesof rat. It may therefore be possible that NPY bindingsites in the heart of rat are not Y1-Rs but other receptorsubtypes. Actually, pharmacologic studies have indi-cated the existence of the NPY Y3 receptor and anothernovel receptor in the heart (Balasubramaniam, 1997).Recently, expression of the NPY Y5 receptor but notY1R mRNA was detected in primary cultured cardiomy-ocytes of mouse by reverse transcriptase-polymerasechain reaction (RT-PCR; Pellieux et al., 2000).
Using RT-PCR, Y1R mRNA has been demonstrated insmooth muscle cells cultivated from human subcutaneousarteries (Erlinge, 1994), and physiologic and pharmaco-logic experiments have shown NPY-induced vasomotion in
Fig. 5. Immunofluorescence micrographs of rat thyroid (A,B) and parathyroid (C,D) glands. A: Highermagnification of rat thyroid gland showing Y1R-LI in vessels. B,D: Y1R-LI is not found after incubationwith preadsorbed antiserum. C: Y1R-LI is observed in blood vessels in the parathyroid gland. Scale barin A � 20 �m in A, 80 �m in A (applies to B–D).
397NPY-Y1 RECEPTORS IN PERIPHERY

Fig. 6. Immunofluorescence micrographs of the rat adrenalgland. A: Y1R-LI is found in capsular arterioles and capillaries.C: Some chromaffin cells in the adrenal medulla are Y1R-positive.Note lack of signal in cortical cells (asterisk). B,D: Y1R-LI isabolished by preincubation with synthetic peptide. E,F: Y1R- (E)and PNMT-LI (F) in some chromaffin cells. Both Y1R-positive,
PNMT-positive chromaffin cells (arrows), Y1R-positive, PNMT-negative cells (doublearrows), and Y1R-negative, PNMT-positivecells (arrowheads) are found. G,H: Large arrows point to Y1R-positive, NPY-positive chromaffin cells. Scale bar in A � 80 �m(applies to A–D), 40 �m in A (applies to E–H).
398 H. MATSUDA ET AL.

Fig. 7. Immunofluorescence micrographs of sections of the ratmale genital tract incubated with antiserum to Y1R (A,C,E,G,H,J,K)or with Y1R antiserum adsorbed with synthetic peptide (B,D,F,I,L).A,C,E,G,H: Y1R-LI is seen in small arteries and arterioles in thetestis (A), ductus epididymidis (C), vas deferens (E), and prostate
gland (G,H). J,K: Some of the epithelial cells of the seminal vesicle areY1R-IR. B,D,F,I,L: Y1R-LI is not found after incubation with pread-sorbed antiserum. Scale bar in A � 80 �m (applies to A–G,I,J,L), 40�m in A (applies to H,K).

subcutaneous arteries mediated by Y1Rs (Hashim andTadepalli, 1995; Nilsson et al., 1996), in agreement withour observation of Y1R-LI in subcutaneous arteries indermis and hypodermis.
Y1R-LI was also seen in the striated muscle cells of theauricle and paw of rat. Although neither pharmacologicnor physiologic studies have shown effects of NPY on suchmuscle cells, Grassi and colleagues (1996) have reported adirect action of NPY on skeletal muscle fibers of rabbitjaw, where NPY induces a long-lasting and dose-dependent increase in peak tension and duration of max-imal twitches. It could therefore be interesting to test theeffect of NPY and Y1R agonists on the subcutaneous stri-ated muscle cells in rat auricle and paw.
Exocrine glands
NPY-IR fibers have in many studies been reported to existaround acini, ducts, and blood vessels of salivary glands(Kusakabe et al., 1997; Matsuda et al., 1997), with fibersaround ducts and blood vessels in the parotid gland beingsympathetic and periacinar NPY fibers having parasympa-
thetic origin (Ekstrom et al., 1996), suggesting a role in thecontrol of protein secretion. However, we only demonstratedY1Rs in blood vessels, suggesting that sympathetic NPYmodulates blood flow by controlling vascular tone via Y1Rs.Parasympathetic NPY, on the other hand, may affect proteinsecretion via another NPY receptor subtype.
In addition, in the pancreas, we found Y1R-LI only inblood vessels, i.e., neither in acinar cells nor in exocrine ductcells. Pharmacologic experiments have revealed that NPYinhibits pancreatic exocrine secretion (Sheikh, 1991; Grandtet al., 1995). Sheikh (1991) reported an indirect inhibitoryeffect of NPY on pancreatic secretion and speculated thatNPY inhibits secretion through reduction in pancreatic bloodflow, in agreement with the present results. In contrast,Grandt et al. (1995) reported direct inhibitory effects of NPYon pancreatic exocrine secretion via Y1Rs, perhaps presentat such low levels that we cannot detect them.
Gastrointestinal tract
Autoradiographic studies have shown the presence ofY1R in the submucosal and myenteric ganglia in the in-
Fig. 8. Immunofluorescence micrographs of the rat female genitaltract incubated with Y1R antiserum (A,C,E) or Y1R antiserum ad-sorbed with Y1R peptide (B,D,F). A,C,E: Y1R-LI is observed in blood
vessels in the uterus (A), ovary (C), and oviduct (E). B,D,F: Y1R-LI isabolished by preincubation with Y1R peptide. Scale bar in A � 80 �m(applies to A–F).
400 H. MATSUDA ET AL.

testine (Walsh et al., 1993; Mao et al., 1996), and recentlyY1R-LI has been studied with immunohistochemical tech-niques in the gastrointestinal tract of rat (Jackerott andLarsson, 1997) and human (Peaire et al., 1997). Thus,Y1R-LI was seen in nerve cell bodies in the myenteric andsubmucosal plexuses, in the nerve fibers in lamina propriaand muscle layers, in a few endocrine-like cells, in endo-thelial cells of blood vessels, and in cells in the mucosa andcrypts. In the present study we observed a somewhat morelimited distribution of Y1R-LI, i.e., in nerve cell bodies inthe myenteric plexus, in a few enteroendocrine cells, insmall arteries and arterioles (diameter up to 200 �m inrat), and in some nerve fibers in muscle layers. Thesedifferences may reflect different characteristics of antibod-ies as well as sensitivity of techniques used in the studies.
Biologic effects of NPY in the gastrointestinal tract in-clude inhibition of motility and secretion (Sheikh, 1991).NPY may act on the gastrointestinal tract via Y1Rs onneurons in the myenteric plexus, and presumably in thesubmucosal plexus, although we have been unable to de-tect Y1R-LI in the latter location. Interestingly, in allthree studies Y1R-LI was observed in fibers, suggesting apresynaptic localization. Although Y1Rs have been consid-ered to be primarily postsynaptic, as originally proposedby Wahlestedt and his colleagues (1986), there is nowevidence for a presynaptic localization (Peaire et al., 1997;Pickel et al., 1998; St-Pierre et al., 2000; Kopp et al., 2002;Ubink et al., 2001; Brumovsky et al., 2002). The fact thata few enteroendocrine cells were positive for the Y1Rsuggests that secretion of a peptide hormone and/or bio-genic amine (Polak and Bloom, 1988; Sundler and Hakan-son, 1988) in single instances may be regulated by NPY.
Endocrine system
The present study shows Y1R-IR endocrine cells in is-lets of Langerhans of rat. In agreement with these results,Y1R mRNA has been detected in islets of Langerhans ofmouse by RT-PCR (Burcelin et al., 2001). However,Y1R-LI was not found in this location in the mouse withour technique. Double immunostaining revealed that allY1R-positive cells contained SOM, but only around one-third of the SOM cells showed Y1R-LI. The presence ofSOM in the pancreatic D-cells was early reported (Hokfeltet al., 1975), and SOM inhibits both glucose- andcarbachol-induced insulin secretion (Ahren, 1981; Ho etal., 1987). NPY has been reported to have a biphasic effecton insulin secretion in the mouse: it enhances basalplasma insulin levels at high doses and inhibits glucose-induced insulin secretion (Pettersson et al., 1987). Al-though the inhibitory pattern of NPY is different fromthat of SOM, both NPY and SOM are functional inhibitorsof insulin secretion and could work in a cooperative man-ner. Because the Y1R in general is considered to be inhib-itory, the presence of this receptor in SOM cells is surpris-ing. Also, species differences seem to exist, as we observedY1R-LI only in rat D-cells.
Y1R-LI was found at high levels in a dense plexus ofblood vessels in the thyroid gland. In agreement withthese results, this gland receives a dense NPY innervation(Grunditz et al., 1988); a physiologic study has shown thatendogenous NPY regulates thyroid blood flow in normalrats (Michalkiewicz et al., 1993). NPY also potentiates thesecretory response to thyroid-stimulating hormone in thethyroid gland (Ahren, 1986); however, our study revealedno Y1R-LI in the follicular cells. Therefore, it is possible
that a main action of NPY is to regulate thyroid blood flowvia Y1Rs in blood vessels and that modulation of thyroidhormone secretion occurs via other receptor subtypes. In-terestingly, most of the Y1R-IR blood vessels in the thy-roid gland were �-SMA-negative, and Y1R-LI in this or-gan therefore seems to be located mainly in capillaries.However, to classify fully the identity of the Y1R-positivecells (i.e., whether they are endothelial cells or pericytes),electron microscopic studies are needed.
The parathyroid gland is richly innervated by NPY-IRnerve fibers (Luts and Sundler, 1994), and the presentinvestigation showed Y1R-LI around blood vessels also inthis gland, suggesting that NPY can influence parathyroidfunction through effects on blood flow, although no phys-iologic studies have been reported so far.
Some chromaffin cells in the adrenal medulla were Y1R-positive, but only a subpopulation was PNMT-positive,i.e., represents adrenaline cells. Thus, Y1Rs are present inboth types of chromaffin cells. It is well known that NPYis expressed in many adrenaline chromaffin cells (Lund-berg et al., 1986). Therefore, part of the Y1Rs in theadrenal medulla may be autoreceptors. In agreement,some Y1R-positive chromaffin cells contained NPY. Phar-macologic experiments have indicated that NPY positivelyregulates tyrosine hydroxylase mRNA expression viaY1Rs in the rat adrenal medulla (Hong et al., 1995) andalso enhances catecholamine synthesis (Nussdorfer andGottardo, 1998). The lack of Y1R-LI in the superior cervi-cal and sphenopalatine ganglia is in agreement with an insitu hybridization study on the rat superior cervical gan-glion (Zhang et al., 1997).
It is well known that the pineal gland of rat is denselyinnervated by NPY-containing nerve fibers, most of whichoriginate from the superior cervical ganglion (Zhang et al.,1991). NPY plays a modulatory role in noradrenergic neu-rotransmission in the rat pineal gland (Mikkelsen et al.,1999), and RT-PCR analysis has revealed expression ofY1R mRNA. However, we could not detect Y1R-LI in thepineal gland, perhaps again due to a low sensitivity of ourtechnique.
Respiratory tract
Pharmacologic investigation indicates the presence andfunction of Y1R in the guinea pig bronchi (Grundemar,1988). NPY suppresses sensory C-fiber-mediated contrac-tions of bronchi via Y1R. However, we could not findY1R-LI in the rat lung, not even around blood vessels.Parasympathetically mediated contractions in the guineapig trachea are suppressed by NPY, mainly via Y2Rs(Grundemar, 1997). NPY also inhibits cholinergic para-sympathetic vasodilation in the bronchial circulation ofdog by an action on Y2Rs (Mahns et al., 1998). Thus,considerable species differences may exist, and it is pos-sible that NPY mainly acts on Y2Rs in the rat lung, inagreement with our failure to detect Y1Rs in this tissue.
Urogenital tract
An autoradiographic binding study has revealed NPYbinding sites in the proximal convoluted tubules and thevascular smooth muscle in the rabbit kidney, but not inthe rat kidney (Leys et al., 1987). In situ hybridizationexperiments have shown Y1R mRNA expression in thevascular smooth muscle and glomeruli in the rat kidney(Modin et al., 1999) and in the juxtaglomerular apparatus,collecting ducts, and Henle’s loop in the human kidney
401NPY-Y1 RECEPTORS IN PERIPHERY

(Wharton et al., 1993). Pharmacologic studies have dem-onstrated the existence of Y1R in the pig renal vascula-ture (Modin et al., 1991). In the present study, littleY1R-LI was seen in blood vessels, but an impressive find-ing was the strong Y1R-LI in the renal papilla of both ratand WT, but not Y1R-KO, mouse. Thus, many cells ofcollecting tubules were Y1R-positive.
This result suggests that NPY is related to readsorptionof water in the collecting tubules via Y1Rs, although nei-ther pharmacologic nor physiologic data have so far beenreported on such an effect. However, NPY influences glo-merular filtration and natriuresis (Echtenkamp and Dan-dridge, 1989; Persson et al., 1991) and regulates, togetherwith noradrenaline, renal Na�, K�-ATPase activity, butthis occurs at the tubular level and is considered to beexerted via Y2Rs (Ohtomo et al., 1994). The Y1R-positivecells in arterioles of WT mouse are thought to correspondto the juxtaglomerular cells. This finding indicates thatNPY may modulate renin release, at least in mouse. Ac-tually, administration of NPY as well as Y1R-agonistcauses increases in mean arterial blood pressure in pig(Modin et al., 1999), perhaps involving release of renin.
In the present study, Y1R-LI was observed in the smallarteries and arterioles in the testis, ductus epididymidis,vas deferens, and prostate gland. In our previous report,Y1R-LI could not be detected in the vas deferens (Kopp etal., 1997), probably due to a less “sensitive” antiserumthan used here, but in the testis good agreement wasobtained, suggesting high Y1R levels. The vas deferenshas been studied as a model system for characterization ofthe prejunctional action of NPY, revealing that NPY in-hibits release of NPY and noradrenaline via presynapticY2Rs (Wahlestedt et al., 1986; Pheng and Regoli, 1998).Our findings that there are no Y1R-positive nerve termi-nals, but that Y1R-LI only exists in blood vessels, indi-rectly support these pharmacologic data.
The seminal vesicle of rodents is innervated by abun-dant NPY-positive fibers located in the smooth musclelayer, in the subepithelial region, and around blood ves-sels (Pinho et al., 1997). However, pharmacologic studiesin rat indicate that NPY has no direct motor effect on theseminal vesicle and that the primary effect of NPY ismodulation of adrenergic motor transmission by prejunc-tional inhibition of noradrenaline release (Iravani andZar, 1994). However, NPY fibers beneath the epitheliummay participate in the regulation of epithelial secretoryfunction, possibly via the putative Y1Rs observed here tobe present on epithelial cells. It should be noted that theY1R-positive epithelial cells were only found in rat, sospecificity could not be monitored with the Y1R-KOmouse.
We observed Y1R-LI in blood vessels in the female re-productive system not detected by Kopp and her col-leagues (1997), again probably related to the antiseraused. Injection of NPY close to the ovarian artery hasshown a dose-dependent decrease in the ovarian bloodflow rate, and the selective Y1R agonist [Leu31,Pro34]NPY potentiates the noradrenaline response (Jør-gensen, 1994), suggesting the presence of Y1R in the ovar-ian artery. Y1Rs in the female reproductive system maymediate vasomotion by direct effects or by potentiation ofnoradrenaline.
CONCLUSIONS
The present study demonstrates the distribution ofY1Rs in various peripheral tissues of rat and of WT andY1R-KO mice. They were predominantly found in vascularsmooth muscle cells of vessels, with a diameter up to 200�m in rat, but also in several cell types not related tovessels. Future immunohistochemical studies on othersubtypes of NPY receptors, as well as the use of moresensitive histochemical approaches including in situ hy-bridization, may shed further light on the role of NPY andits receptors in peripheral tissues.
ACKNOWLEDGMENTS
We thank the late Dr. J.H. Walsh and H. Wong [Gas-troenteric Biology Center (CURE), Department of Medi-cine, University of California, Los Angeles, CA], for thegenerous donation of Y1R and somatostatin antisera, andDrs. E. Grouzmann and C. Bohuon (University of Lau-sanne, Lausanne, Switzerland) for NPY antibodies. Wethank Katarina Aman for excellent technical assistance.Hideki Matsuda was supported by a guest scholarshipfrom the Swedish Institute.
LITERATURE CITED
Adams JC. 1992. Biotin amplification of biotin and horseradish peroxidesignals in histochemical stains. J Histochem Cytochem 40:1457–1463.
Ahren B. 1981. On the peptidergic and aminergic regulation of insulinsecretion. M.D. Thesis. University of Lund, Lund, Sweden.
Ahren B. 1986. Neuropeptide Y and pancreatic polypeptide: effects onthyroid hormone secretion in the mouse. Eur J Pharmacol 126:97–102.
Allen JJ, Koenig JI, editors. 1992. Central and peripheral significance ofneuropeptide Y and its related peptides. Ann NY Acad Sci 611.
Allen JM, Adrian T, Polak J, Bloom SR. 1983a. Neuropeptide in theadrenal gland. J Auton Nerv Syst 9:559–563.
Allen JM, Bircham PMM, Edwards AV, Tatemoto K, Bloom SR. 1983b.Neuropeptide Y (NPY) reduces myocardial perfusion and inhibits theforce of contraction of the isolated perfused rabbit heart. Regul Pept6:247–253.
Balasubramaniam A. 1997. Neuropeptide Y family of hormones: receptorsubtypes and antagonists. Peptides 18:445–457.
Bao L, Kopp J, Zhang X, Xu Z-QD, Zhang L-F, Wong H, Walsh J, HokfeltT. 1997. Localization of neuropeptide Y Y1 receptors in cerebral bloodvessels. Proc Natl Acad Sci U S A 94:12661–12666.
Blomqvist AG, Herzog H. 1997. Y-receptor subtype—how many more?Trends Neurosci 20:294–298.
Brumovsky PR, Shi TJ, Matsuda H, Kopp J, Villar MJ, Hokfelt T. 2002.NPY Y1 receptors are present in axonal processes of DRG neurons. ExpNeurol 174:1–10.
Burcelin R, Brunner HR, Seydoux J, Thorens B, Pedrazzini T. 2001.Increased insulin concentrations and glucose storage in neuropeptide YY1 receptor-deficient mice. Peptides 22:421–427.
Cabrele C, Beck-Sickinger AG. 2000. Molecular characterization of theligand-receptor interaction of the neuropeptide Y family. J Pept Sci6:97–122.
Coons AH. 1958. Fluorescent antibody methods. In: Danielli JF, editor.General cytochemical methods. New York: Academic Press. p 399–422.
Dumont Y, Martel JC, Fournier A, St-Pierre S, Quirion R. 1992. Neuropep-tide Y and neuropeptide Y receptor subtypes in brain and peripheraltissues. Prog Neurobiol 38:125–167.
Echtenkamp SF, Dandridge PF. 1989. Renal actions of neuropeptide Y inthe primate. Am J Physiol 256:F524–F531.
Edvinsson L, Ekblad E, Hakanson R, Wahlestedt C. 1984. Neuropeptide Ypotentiates the effect of various vasoconstrictor agents on rabbit bloodvessels. Br J Pharmacol 83:519–525.
Ekblad E, Edvinsson L, Wahlestedt C, Uddman R, Hakanson R, Sundler F.1984. Neuropeptide Y co-exists and co-operates with noradrenaline inperivascular nerve fibers. Regul Pept 8:225–235.
402 H. MATSUDA ET AL.

Ekstrom J, Ekman R, Luts A, Sundler F, Tobin G. 1996. Neuropeptide Y insalivary glands of the rat: origin, release and secretory effects. RegulPept 61:125–134.
Erlinge D. 1994. The sympathetic cotransmitters neuropeptide Y and ATPin the regulation of the vascular smooth muscle cell. M.D. Thesis.University of Lund, Lund, Sweden.
Eva C, Keinanen K, Monyer H, Seeburg P, Sprengel R. 1990. Molecularcloning of a novel G protein-coupled receptor that may belong to theneuropeptide receptor family. FEBS Lett 271:81–84.
Eva C, Oberto A, Sprengel R, Genazzani E. 1992. The murine NPY-1receptor gene. Structure and delineation of tissue-specific expression.FEBS Lett 314:285–288.
Franco-Cereceda A, Liska J. 1998. Neuropeptide Y Y1 receptors in vascularpharmacology. Eur J Parmacol 349:1–14.
Gehlert DR. 1994. Subtypes of receptors for neuropeptide Y: implicationsfor the targeting of therapeutics. Life Sci 55:551–562.
Gehlert DR. 1998. Multiple receptors for the pancreatic polypeptide (PP-fold) family: physiological implications. Proc Soc Exp Biol Med 218:7–22.
Gerald C, Walker MW, Vaysse PJ, He C, Branchek TA, Weinshank RL.1995. Expression cloning and pharmacological characterization of ahuman hippocampal neuropeptide Y/peptide YY Y2 receptor subtype.J Biol Chem 270:26758–26761.
Goldstein M, Fuxe K, Hokfelt T, Joh TH. 1971. Immunohistochemicalstudies on phenylethanolamine-N-methyltransferase, dopa-decarbox-ylase and dopamine-�-hydroxylase. Experientia 27:951–952.
Grandt D, Siewert J, Sieburg B, Al Tai, O, Schimiczek M, Goebell H, LayerP, Eysselein VE, Reeve Jr, JR, Muller MK. 1995. Peptide YY inhibitsexocrine pancreatic secretion in isolated perfused rat pancreas by Y1receptor. Pancreas 10:180–186.
Grassi C, Deriu F, Roatta S, Santarelli R, Azzena GB, Passatore M. 1996.Sympathetic control of skeletal muscle function: possible co-operationbetween noradrenaline and neuropeptide Y in rabbit jaw muscles.Neurosci Lett 212:204–208.
Grouzmann E, Comoy E, Walker P, Burinier M, Bohuon C, Waeber B,Brunner H. 1992. Production and characterization of four anti-neuropeptide Y monoclonal antibodies. Hybridoma 11:409–424.
Grundemar L. 1988. Suppression of sensory C fiber-mediated contractionsby neuropeptide Y Y1 receptors in the guinea pig bronchi. Regul Pept75–76:181–184.
Grundemar L. 1997. Characterization of the receptor response for theneuropeptide Y-evoked suppression of parasympathetically-mediatedcontractions in the guinea pig trachea. Regul Pept 71:97–101.
Grundemar L, Hakanson R. 1993. Multiple neuropeptide Y receptors areinvolved in cardiovascular regulation. Peripheral and central mecha-nisms. Gen Pharmacol 24:785–796.
Grunditz T, Ekman R, Hakanson R, Sundler F, Uddman R. 1988. Neu-ropeptide Y and vasoactive intestinal peptide coexist in rat thyroidnerve fibers emanating from the thyroid ganglion. Regul Pept 23:193–208.
Hashim MA, Tadepalli AS. 1995. Cutaneous vasomotor effects of neuropep-tide Y. Neuropeptides 29:263–271.
Herzog H, Hort YJ, Ball HJ, Hayes G, Shine J, Selbie LA. 1992. Clonedhuman neuropeptide Y receptor couples to two different second mes-senger systems. Proc Natl Acad Sci U S A 89:5794–5798.
Ho LT, Pu HF, Sheu WJ, Wang WC, Wang PS. 1987. Inhibition of soma-tostatin on glucose-induced release of gastric inhibitory polypeptide inrat. Chin J Physiol 30:45–53.
Hokfelt T, Efendic S, Hellerstrom C, Johansson O, Luft A, Arimura A.1975. Cellular localization of somatostatin in endocrine-like cells andneurons of the rat with special references to the A1-cells of the pan-creatic islets and to the hypothalamus. Acta Endocrinol Suppl 200:5–41.
Hong M, Li S, Fournier A, St-Pierre S, Pelletier G. 1995. Role of neuropep-tide Y in the regulation of tyrosine hydroxylase gene expression in ratadrenal glands. Neuroendocrinology 61:85–88.
Iravani MM, Zar MA. 1994. Presence of neuropeptide Y in the rat seminalvesicle and its effects on noradrenaline- and nerve-induced contraction.Br J Pharmacol 113:877–882.
Jackerott M, Larsson L-I. 1997. Immunocytochemical localization of theNPY/PYY Y1 receptor in enteric neurons, endothelial cells, andendocrine-like cells of the rat intestinal tract. J Histochem Cytochem45:1643–1650.
Jazin EE, Zhang X, Soderstrom S, Williams R, Hokfelt T. 1993. Expressionof peptide YY and mRNA for the NPY/PYY receptor of the Y1 subtype
in dorsal root ganglia during rat embryogenesis. Dev Brain Res 76:105–113.
Jørgensen JC. 1994. Neuropeptide Y in mammalian genital tract: localiza-tion and biological action. Dan Med Bull 41:294–305.
Kopp J, Zhang X, Hokfelt T. 1997. Neuropeptide Y1 receptors in the ratgenital tract. Regul Pept 70:149–160.
Kopp J, Xu Z-Q, Zhang X, Pedrazzini T, Herzog H, Kresse A, Wong H,Walsh JH, Hokfelt T. 2001. Expression of the neuropeptide Y Y1receptor in the CNS of rat, wild type and Y1 receptor knock-out mouse.Focus on immunohistochemical localization. Neuroscience 111:443–532.
Kusakabe T, Matsuda H, Kawakami T, Syoui N, Kurihara K, Tsukuda M,Takenaka T. 1997. Distribution of neuropeptide-containing nerve fi-bers in the human submandibular gland, with special reference to thedifference between serous and mucous acini. Cell Tissue Res 288:25–31.
Kuwayama Y, Emson PC, Stone RA. 1988. Pterygopalatine ganglion cellscontain neuropeptide Y. Brain Res 446:219–224.
Larhammar D. 1996. Structural diversity of receptors for neuropeptide Y,peptide YY and pancreatic polypeptide. Regul Pept 65:165–174.
Larhammar D, Blomqvist AG, Yee F, Jazin E, Yoo H, Wahlested C. 1992.Cloning and functional expression of a human neuropeptide Y/peptideYY receptor of the Y1 type. J Biol Chem 267:10935–10938.
Leblanc GG, Trimmer BA, Landis SC. 1987. Neuropeptide Y-like immu-noreactivity in rat cranial parasympathetic neurons: coexistence withvasoactive intestinal peptide and choline acetyltransferase. Proc NatlAcad Sci U S A 84:3511–3515.
Leys K, Schachter M, Sever P. 1987. Autoradiographic localisation of NPYreceptors in rabbit kidney: comparison with rat, guinea-pig and hu-man. Eur J Pharmacol 134:233–237.
Lundberg JM. 1996. Pharmacology of cotransmission in the autonomicnervous system: integrative aspects of amines, neuropeptides, adeno-sine triphosphate, amino acids and nitric oxide. Pharmacol Rev 48:113–178.
Lundberg JM, Tatemoto K. 1982. Pancreatic polypeptide family (APP,BPP, NPY and PYY) in relation to sympathetic vasoconstriction resis-tant to alpha-adrenoceptor blockade. Acta Physiol Scand 116:393–402.
Lundberg JM, Terenius L, Hokfelt T, Martling CR, Tatemoto K, Mutt V,Polak J, Bloom SR, Goldstein M. 1982. Neuropeptide Y (NPY)-likeimmunoreactivity in peripheral noradrenergic neurons and effects ofNPY on sympathetic function. Acta Physiol Scand 116:477–480.
Lundberg JM, Terenius L, Hokfelt T, Goldstein M. 1983. High levels ofneuropeptide Y in peripheral noradrenergic neurons in various mam-mals including man. Neurosci Lett 42:167–172.
Lundberg JM, Pernow J, Tatemoto K, Dahlof C. 1985. Pre- and postjunc-tional effects of NPY on sympathetic control of rat femoral artery. ActaPhysiol Scand 123:511–513.
Lundberg JM, Hokfelt T, Hemsen A, Theodorsson-Norheim E, Pernow J,Hamberger B, Goldstein M. 1986. Neuropeptide Y-like immunoreactiv-ity in adrenaline cells of adrenal medulla and in tumors and plasma ofpheochromocytoma patients. Regul Pept 13:169–182.
Luts L, Sundler F. 1994. Peptide-containing nerve fibers in the parathyroidglands of different species. Regul Pept 50:147–158.
Mahns DA, Lacroix JS, Potter EK. 1998. Inhibition of vagal vasodilation bya selective neuropeptide Y Y2 receptor agonist in the bronchial circu-lation of anaesthetised dogs. J Auton Nerv Syst 73:80–85.
Malmstrom RE. 1997. Neuropeptide Y Y1 receptor mechanisms in sympa-thetic vascular control. Acta Physiol Scand Suppl 636:1–55.
Mao YK, Wang YF, Warg G, Clipris S, Daniel EE, McDonald TJ. 1996.Peptide YY receptor in submucosal and myenteric plexus synapto-somes of canine small intestine. Am J Physiol 271:G36–G41.
Matsuda H, Kusakabe T, Kawakami T, Nagahara T, Takenaka T, TsukudaM. 1997. Neuropeptide-containing nerve fibers in the human parotidgland: a semiquantitative analysis using an antibody against proteingene product 9.5. Histochem J 29:539–544.
McDermott BJ, Millar BC, Piper HM. 1993. Cardiovascular effects ofneuropeptide Y: receptor interactions and cellular mechanisms. Car-diovasc Res 27:893–905.
Michalkiewicz M, Huffman LJ, Dey M, Hedge GA. 1993. Endogenousneuropeptide Y regulates thyroid blood flow. Am J Physiol 264:E699–E705.
Michel MC. 1991. Receptors for neuropeptide Y: multiple subtypes andmultiple second messengers. Trends Pharmacol Sci 12:389–394.
Mikkelsen JD, Hauser F, Olcese J. 1999. Neuropeptide Y (NPY) and NPYreceptors in the rat pineal gland. Adv Exp Med Biol 460:95–107.
403NPY-Y1 RECEPTORS IN PERIPHERY

Modin A, Pernow J, Lundberg JM. 1991. Evidence for two neuropeptide Yreceptors mediating vasoconstriction. Eur J Pharmacol 203:165–171.
Modin A, Malmstrom RE, Meister B. 1999. Vascular neuropeptide Y Y1-receptors in the rat kidney: vasoconstrictor effects and expression ofY1-receptor mRNA. Neuropeptides 33:253–259.
Mutt V, Fuxe K, Hokfelt T, Lundberg JM, editors. 1989. Neuropeptide Y.Karolinska Institute Nobel Conference Series. New York: Raven Press.
Nakamura M, Sakanaka C, Aoki Y, Ogasawara H, Tsuji T, Kodama H,Matsumoto T, Shimizu T, Noma M. 1995. Identification of two isoformsof mouse neuropeptide Y-Y1 receptor generated by alternative splicing.J Biol Chem 270:30102–30110.
Nilsson T, Erlinge D, Cantera L, Edvinsson L. 1996. Contractile effects ofneuropeptide Y in human subcutaneous resistance arteries are medi-ated by Y1 receptors. J Cardiovasc Pharmacol 28:764–768.
Nussdorfer GC, Gottardo G. 1998. Neuropeptide-Y family of peptides in theautocrine-paracrine regulation of adrenocortical function. Horm MetabRes 30:368–373.
Ohtomo Y, Meister B, Hokfelt T, Aperia A. 1994. Coexisting NPY and NEsynergistically regulate renal tubular Na�, K�-ATPase activity. Kid-ney Int 45:1606–1613.
Peaire AE, Krantis A, Staines WA. 1997. Distribution of the NPY receptorsubtype Y1 within human colon: evidence for NPY targeting a subpopu-lation of nitergic neurons. J Auton Nerv Syst 67:168–175.
Pedrazzini T, Seydoux J, Kuenstner P, Aubert JF, Grouzmann E, BeermanF, Brunner HR. 1998. Cardiovascular response, feeding behavior andlocomotor activity in mice lacking the NPY Y1 receptor. Nat Med4:722–726.
Pellieux C, Sauthier T, Domenighetti A, Marsh DJ, Palmiter RD, BrunnerHR, Pedrazzini T. 2000. Neuropeptide Y (NPY) potentiatesphenylephrine-induced mitogen-activated protein kinase activation inprimary cardiomyocytes via NPY Y5 receptors. Proc Natl Acad Sci U SA 97:1595–1600.
Persson PB, Ehmke H, Nafz B, Lang R, Hackenthal E, Nobiling R, DietrichMS, Kirchheim HR. 1991. Effects of neuropeptide-Y on renal functionand its interaction with sympathetic stimulation in conscious dogs.J Physiol 444:289–302.
Pettersson M, Ahren B, Lundquist I, Bottcher G, Sundler F. 1987. Neu-ropeptide Y: intrapancreatic neuronal localization and effects on insu-lin secretion in the mouse. Cell Tissue Res 248:43–48.
Pheng LH, Regoli D. 1998. Bioassays for NPY receptors: old and new.Regul Pept 75–76:79–87.
Pheng LH, Regoli D. 2000. Receptors for NPY in peripheral tissues bioas-says. Life Sci 67:847–862.
Pickel VM, Beck-Sickinger AG, Chan J, Weiland HA. 1998. Y1 receptors inthe nucleus accumbens: ultrastructural localization and associationwith neuropeptide Y. J Neurosci Res 52:54–68.
Pinho MS, Afonso F, Rodrigues G, Gulbenkian S, Mata LR. 1997. Neu-ropeptides in the seminal vesicles: locations, binding sites and func-tional implications. Histol Histopathol 12:503–512.
Polak JM, Bloom SR. 1988. Review: the enterochromaffin-like cell, intra-gastric acidity and the trophic effect of plasma gastrin. Aliment Phar-macol Ther 2:291–296.
Rose PM, Fernandes P, Lynch JS, Frazier ST, Fisher SM, Kodukula K,Kienzle B, Seethala R. 1995. Cloning and functional expression of acDNA encoding a human type 2 neuropeptide Y receptor. J Biol Chem270:22661–22664.
Schalling M, Seroogy K, Hokfelt T, Chai SY, Hallman H, Persson H,Larhammar D, Ericsson A, Terenius L, Graffi J, Massoulie J, GoldsteinM. 1988. Neuropeptide tyrosine in the rat adrenal gland—immunohistochemical and in situ hybridization studies. Neuroscience24:337–349.
Sheikh SP. 1991. Neuropeptide Y and peptide YY: major modulators ofgastrointestinal blood flow and function. Am J Physiol 261:G701–G715.
Sheikh SP, Roach E, Fuhlendorff J, Williams JA. 1991. Localization of Y1receptors for NPY and PYY on vascular smooth muscle cells in ratpancreas. Am J Physiol 260:G250–G257.
St-Pierre JA, Nouel D, Dumont Y, Beaudet A, Quirion R. 2000. Associationof neuropeptide Y Y1 receptors with glutamate-positive and NPY-positive neurons in rat hippocampal cultures. Eur J Neurosci 12:1319–1330.
Sundler F, Hakanson R. 1988. Peptide hormone-producing endocrine/paracrine cells in the gastro-entero-pancreatic region. In: Bjorklund A,Hokfelt T, Owman C, editors. Handbook of chemical neuroanatomy.The peripheral nervous system. Amsterdam: Elsevier. p 219–295.
Tatemoto K. 1982. Neuropeptide Y. Complete amino acid sequence of thebrain peptide. Proc Natl Acad Sci U S A 79:5485–5489.
Tatemoto K, Carlquist M, Mutt V. 1982. Neuropeptide Y—a novel brainpeptide with structural similarities for peptide YY and pancreaticpolypeptide. Nature 296:659–660.
Ubink R, Kopp J, Wong H, Walsh JH, Pedrazzini T, Hokfelt T. 2001.Transient prenatal expression of NPY-Y1 receptor in trigeminal axonsinnervating the mystacial vibrissae. J Comp Neurol 429:183–191.
Unden A, Tatemoto K, Mutt V, Bartfai T. 1984. Neuropeptide Y receptor inthe rat brain. Eur J Biochem 145:525–530.
Wahlestedt C, Reis DJ. 1993. Neuropeptide Y-related peptides and theirreceptors—are the receptors potential therapeutic drug targets? AnnuRev Pharmacol Toxicol 32:309–352.
Wahlestedt C, Edvinsson L, Ekblad E, Hakanson R. 1985. Neuropeptide Ypotentiates noradrenaline-evoked vasoconstriction: mode of action.J Pharmacol Exp Ther 234:735–741.
Wahlestedt C, Yanaihara N, Hakanson R. 1986. Evidence for different pre-and post-junctional receptors for NPY and related peptides. Regul Pept13:307–318.
Wahlestedt C, Wohlfart B, Hakanson R. 1987. Effects of neuropeptide Y(NPY) on isolated guinea-pig heart. Acta Physiol Scand 129:459–463.
Walsh DA, Wharton J, Blake DR, Polak JM. 1993. Localization and char-acterization of neuropeptide Y binding sites in porcine and humancolon. Br J Pharmacol 108:304–311.
Westfall TC, Carpentier S, Chen X, Beinfeld MC, Naes L, Meldrum MJ.1987. Prejunctional and postjunctional effects on neuropeptide Y at thenoradrenergic neuroeffector junction of the perfused mesenteric arte-rial bed of the rat. J Cardiovasc Pharmacol 10:716–722.
Westfall TC, Martin J, Chen XL, Ciarleglio A, Carpentier S, Henderson K,Knuepfer M, Beinfeld M, Naes L. 1988. Cardiovascular effects andmodulation of noradrenergic neurotransmission following central andperipheral administration of neuropeptide Y. Synapse 2:299–307.
Wharton J, Gordon L, Byrne J, Herzog H, Selbie LA, Moore K, SullivanMHF, Elder MG, Moscoso G, Taylor KM, Shine J, Polak JM. 1993.Expression of the human neuropeptide tyrosine Y1 receptor. Proc NatlAcad Sci U S A 90:687–691.
Wong HC, Walsh JH, Yang H, Tache Y, Buchan AM. 1990. A monoclonalantibody to somatostatin with potent in vivo immunoneutralizing ac-tivity. Peptides 11:707–712.
Zhang ET, Mikkelsen JD, Møller M. 1991. Tyrosine hydroxylase- andneuropeptide Y-immunoreactive nerve fibers in the pineal complex ofuntreated rats and rats following removal of the superior cervicalganglia. Cell Tissue Res 265:63–71.
Zhang X, Bao L, Xu Z-Q, Kopp J, Arvidsson U, Elde R, Hokfelt T. 1994a.Localization of neuropeptide Y Y1 receptors in the rat nervous systemwith special reference to somatic receptors on small dorsal root gan-glion neurons. Proc Natl Acad Sci U S A 91:11738–11742.
Zhang X, Wiesenfeld-Hallin Z, Hokfelt T. 1994b. Effect of peripheral axo-tomy on expression of neuropeptide Y receptor mRNA in rat lumbardorsal root ganglia. Eur J Neurosci 6:43–57.
Zhang X, Shi T, Holmberg K, Landry M, Huang W, Xiao HS, Ju G, HokfeltT. 1997. Expression and regulation of the neuropeptide Y Y2 receptorin sensory and autonomic ganglia. Proc Natl Acad Sci U S A 94:729–734.
Zhang X, Tong YG, Bao L, Hokfelt T. 1999. The neuropeptide Y Y1 receptoris a somatic receptor on dorsal root ganglion neurons and a postsyn-aptic receptor on somatostatin dorsal horn neurons. Eur J Neurosci11:2211–2225.
404 H. MATSUDA ET AL.