direct cross-linking of125i-labeled glucagon to its membrane receptor by uv irradiation
TRANSCRIPT

Direct Cross-Linking of 125I-Labeled Glucagon to its Membrane Receptor by UV IrradiationAuthor(s): Victoria Iwanij and Kyu Chung HurSource: Proceedings of the National Academy of Sciences of the United States of America,Vol. 82, No. 2 (Jan. 15, 1985), pp. 325-329Published by: National Academy of SciencesStable URL: http://www.jstor.org/stable/24622 .
Accessed: 02/05/2014 09:55
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
National Academy of Sciences is collaborating with JSTOR to digitize, preserve and extend access toProceedings of the National Academy of Sciences of the United States of America.
http://www.jstor.org
This content downloaded from 130.132.123.28 on Fri, 2 May 2014 09:55:58 AMAll use subject to JSTOR Terms and Conditions

Proc. Nati. Acad. Sci. USA Vol. 82, pp. 325-329. January 1985 Biochemistry
Direct cross-linking of 125I-labeled glucagon to its membrane receptor by UV irradiation
(hormone receptors/affinity labeling/hepatocytes)
VICTORIA IWANIJ AND KYu CHUNG HUR
University of Minnesota, Department of Genetics and Cell Biology, 1445 Gortner Avenue, St. Paul, MN 55108
Communicated by Philip Siekevitz, September 17, 1984
ABSTRACT 1251-labeled glucagon was directly cross- linked to its receptor in isolated liver plasma membranes and on the surface of intact hepatocytes, by using a UV irradiation procedure. This investigation resulted in the identification of a glucagon-receptor complex of apparent Mr 62,000. The speci- ficity of labeling was shown by the interference of unlabeled hormone at physiological concentration with incorporation of radioactive glucagon into the 62,000 Mr species. The receptor behaved as a typical integral membrane protein: it was not released by extraction with lithium diiodosalicylate or at basic pH but was solubilized by digitonin treatment. Reduction of the receptor polypeptide with dithiothreitol resulted in a de- crease in its electrophoretic mobility, suggesting the presence of intramolecular disulfide bonds. Soluble glucagon-receptor complexes adsorbed to Con A-Sepharose and could be eluted with methyl a-D-mannoside, indicating that the receptor mole- cule is a glycoprotein. Treatment of glucagon-labeled liver plasma membrane with endoglycosidase F resulted in the ap- pearance of four intermediate species, indicating that gluca- gon receptor contains at least four N-linked oligosaccharide chains.
Glucagon, a 29 amino acid peptide hormone, is produced and secreted by a cells of the islets of Langerhans (1). The hor- mone acts upon its principal target site, the liver, to bring about an elevation in plasma glucose levels. To initiate a cel- lular response, peptide hormones interact with specific re- ceptor sites on the surface of target cells. Although there is a large body of work dealing with regulation of glucagon-sensi- tive adenylate cyclase, little is known about the molecular properties of the receptor itself. Isolation and purification of glucagon receptor that retains its binding properties has not yet been achieved (2, 3). The affinity cross-linking approach has resulted in conflicting information about the molecular weight and assembly of glucagon receptor. Bregman and Levy (4) labeled a 25,000 Mlr polypeptide with a biologically inactive azido derivative of glucagon. Recently, Johnson et al. (5) identified a glucagon-binding component of M, 53,000 and Demoliou-Mason and Epand (6) reported glucagon re- ceptor consisting of two polypeptides of Mr 52,000 and 70,000.
In this paper, we report affinity cross-linking studies in which UV irradiation was used to affix 125I-labeled glucagon directly to its receptor sites. We conclude that a 62,000 Mr species, which is specifically labeled both in isolated liver plasma membranes and on the surface of intact hepatocytes, represents the hepatic glucagon receptor.
MATERIALS AND METHODS
Materials. Glucagon, insulin, dithiothreitol, and bacitracin were purchased from Sigma; leupeptin and pepstatin, from
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. ?1734 solely to indicate this fact.
Vega Biochemicals; Trasylol, from FBA Pharmaceuticals; Iodine-125, from Amersham; and digitonin, from Gallard- Schlesinger (Carle Place, NY). Endoglycosidase F (endo-f3- N-acetylglucosaminidase F) was a gift from S. Rosenzweig (Yale University School of Medicine).
Animals. Male rats (100-250 g) were obtained from Bio- Laboratories (St. Paul, MN) and fed ad libitum with Purina Rat Chow.
Preparation of '251-Labeled Glucagon. Glucagon iodination was carried out as described by Lin et al. (7). Separation of iodinated glucagon from unincorporated iodine was carried out on a Sephadex G-10 column (0.5 x 20 cm) equilibrated with 50 mM Tris Cl (pH 7.5) containing bovine serum albu- min (BSA) at 1 mg/ml. Preparation of 1251-labeled insulin was accomplished by the same procedure.
Preparation of Rat Liver Plasma Membranes. Membranes were prepared according to Neville (8) and then frozen and stored in aliquots in liquid nitrogen. In some cases, prepara- tion of plasma membranes was carried out in the presence of the following protease inhibitors: leupeptin (5 pcg/ml), pep- statin (7 ,g/ml), Trasylol (100 units/ml), and EDTA (2 mM).
Cross-Linking of '25I-Labeled Glucagon to Liver Plasna Mem- branes. Binding of glucagon to liver plasma membranes was carried out as follows: 200-400 ,ug of membrane protein was incubated with 125I-labeled glucagon (4-6 x 106 cpm) for 10- 30 min at 300, 250, or 0?C in buffer containing BSA at 2 mg/ml (or gelatin at 1 mg/ml) and a mixture of protease in- hibitors [MPI: bacitracin (0.8 mg/ml), leupeptin (5 ,tg/ml), pepstatin (7 ,g/ml), and Trasylol (100 units/ml)]. At the end of the incubation, the membranes were collected by centrifu- gation in an Eppendorf centrifuge. Pellets were washed twice with incubation buffer and once with 50 mM sodium phosphate, pH 7.5, containing MPI but not BSA. Pellets were resuspended in 350-s.l aliquots of phosphate buffer and transferred into 16-mm-diameter wells of a tissue culture plate (Costar, Cambridge, MA). The plate was then placed 12 cm from a UV lamp (115-V Black Ray) for 10-20 min. Temperature was maintained at 3-4?C by placing the plate in an ice-water bath. Membranes were then collected by cen- trifugation for 2 min. Radioactivity was measured with a y- counter. Nonspecific binding was determined routinely in the presence of 20 AM unlabeled glucagon and found to be 10% of the total binding. The membrane pellet was resus- pended by sonication in 50 ,u of phosphate buffer and proc- essed for electrophoresis. Disulfide bridges were reduced by addition of dithiothreitol. Prior to application to polyacryl- amide gels, samples were incubated for 30 min at 37?C. Boil- ing the samples in the presence of reducing agents was avoided because it resulted in extensive aggregation of the receptor molecules.
Binding and Cross-Linking of 1251-Labeled Glucagon to Cultured Hepatocytes. Preparation of rat hepatocytes was carried out according to Seglen (9). Cells were seeded on
Abbreviations: BSA, bovine serum albumin; MPI, mixture of prote- ase inhibitors.
325
This content downloaded from 130.132.123.28 on Fri, 2 May 2014 09:55:58 AMAll use subject to JSTOR Terms and Conditions

326 Biochemistry: Iwanij and Hur Proc. Natl. Acad Sci. USA 82 (1985)
collagen-coated dishes and cultured for 18-24 hr in Dulbec- co's modified Eagle's medium supplemented with 10% fetal calf serum and insulin (1 ,g/ml). Prior to the binding step, the cells were incubated for 2 hr in serum-free medium and then washed with Hanks' balanced salt solution (Hanks'). Cells were cooled to 4?C by adding ice-cold Hanks' contain- ing BSA at 3 mg/ml and bacitracin at 0.8 mg/ml (BSA/ Hanks'). Cells were incubated with 125I-labeled glucagon at 4?C for 2-12 hr. (Nonspecific binding was assayed in the presence of unlabeled glucagon.) Cells were then washed twice with BSA/Hanks' and once with protein-free Hanks'. The cross-linking step was carried out in protein-free Hanks' for 20 min at 4?C. After the cross-linking step, the cells were washed immediately with BSA/Hanks' and then lysed by the addition of 1 mM sodium bicarbonate containing MPI. The homogenate was centrifuged for 20 min at 10,000 x g, pellets were resuspended in bicarbonate/MPI, and an ali- quot of the suspension was analyzed by NaDodSO4/PAGE.
Extraction of Peripheral Membrane Proteins. Basic pH ex- traction using 0.2 M sodium carbonate was carried out ac- cording to Hubbard and Ma (10). Lithium diiodosalicylate extraction was done according to Porter and Froehner (11).
Liver Plasma Membrane Solubilization and Fractionation on Con A-Sepharose. Liver plasma membranes were incubat- ed with 125I-labeled glucagon, then UV irradiated as de- scribed above. Cross-linked membranes were washed three times by centrifugation, then resuspended in 50 mM Tris Cl, pH 7.5, containing MPI. The concentration of membrane proteins was adjusted to 1 mg/ml, and the suspension was mixed with an equal volume of 4% (wt/vol) digitonin. The mixture was incubated for 30 min on ice with periodic mixing on a Vortex. The insoluble residue was then removed by centrifugation at 15,000 x g for 20 min. The supernatant was diluted 1:4 with column buffer (50 mM Tris Cl, pH 7.5/0.2% digitonin/1 mM CaCl2/0.5 mM MnCl2/MPI) and applied to a column of Con A-Sepharose (2-ml total volume of settled beads) previously equilibrated with column buffer. The col- umn was washed with the same buffer until there was no significant radioactivity in the eluate. Adsorbed proteins were eluted with 0.2 M methyl a-D-mannoside dissolved in column buffer and collected fractions were monitored for ra- dioactivity with a y-counter. Radioactive fractions were pooled and the proteins were precipitated with 20% tri- chloroacetic acid, with 100 jig of cytochrome c added as car- rier. The precipitate was processed for NaDodSO4/PAGE.
Endoglycosidase F Treatment of Affinity Cross-Linked Liv- er Plasma Membranes. Digestion with endoglycosidase F was carried out as described by Rosenzweig et al. (12) ex- cept that boiling of the membrane samples was omitted.
Electrophoresis and Autoradiography. Gel electrophoresis was performed according to Laemmli (13) with 5-15% acryl- amide gradient slab gels. Gels were stained with 0.25% Coo- massie blue R-250, dried, and exposed to Kodak BB-5 film with a Dupont Quanta III screen for 2-6 days. Molecular weight standards were ,B-galactosidase (115,000), phosphor- ylase b (92,000), bovine serum albumin (66,000), catalase (60,000), and tubulin monomers (53,000-55,000). The autora- diograms were scanned using a Helena Laboratories Quick Scan densitometer.
Protein Determination. Protein concentration was estimat- ed by using the fluorescamine method of Udenfriend et al. (14), with BSA as a protein standard.
RESULTS
Covalent Photoincorporation of 1251-Labeled Glucagon into Liver Plasma Membrane. The goal of our investigation was to identify and characterize hepatic glucagon receptor. To this end, we utilized an affinity-labeling approach with UV irradiation as a means of directly cross-linking glucagon to
A B Mr X 10-3
116-
66- 60- FIG. 1. UV-induced cross-linking of
44 "'25-labeled glucagon to liver plasma membranes. Shown are autoradiograms of NaDodSO4/PAGE-separated liver plasma membranes for which binding
24- was carried out in the presence (lane A) or absence (lane B) of 20 ,uM unlabeled glucagon. Positions of molecular weight standards are shown on the left. The la- beled glucagon-receptor band is indicat- ed by the arrowhead on the right.
the receptor sites. Liver plasma membranes, preincubated with 1251-labeled glucagon, washed, and irradiated with UV light, showed a single radioactive band when analyzed by NaDodSO4/PAGE and autoradiography (Fig. 1). This radio- active band migrated between BSA (66,000 Mr), and catalase (60,000 Mr), and its Mr was estimated to be 62,000. In addi- tion to this band, UV irradiation induced the appearance of high Mr radioactive species that did not enter the separating gel and that contained 25-35% of the radioactivity found in the gel. Also, occasionally one or two faint bands were ob- served that may represent dimeric and tetrameric aggregates of the receptor. After UV irradiation, there was essentially no change in the Coomassie blue-staining pattern of liver membrane proteins, which suggests that only a small number of membrane proteins were cross-linked into large aggre- gates.
We have tested the dependence of the covalent incorpo- ration of glucagon into the 62,000 Mr band on the length of UV irradiation. The results are shown in Fig. 2. No radioac- tive band could be detected prior to exposure to UV light, indicating that UV irradiation was necessary for the forma- tion of covalent bonds between the receptor and the ligand. A radioactive band was detected after 5 min of irradiation; its intensity increased with irradiation time up to 20 min and then plateaued with a concomitant increase in the high Mr aggregates. Therefore, we selected 20 min UV treatment for routine cross-linking experiments. Under these conditions, 0.5-1.2% of membrane-bound 125I-labeled glucagon was in- corporated into the 62,000 Mr band. A similar efficiency of glucagon cross-linking (1-2%) was reported by Johnson et al. (5). UV irradiation induced formation of covalent bonds between receptor and ligand; heat and NaDodSO4 treat- ments did not result in dissociation of the complex under the gel electrophoresis conditions.
Specificity of the Cross-Linking Procedure. The following experiments were carried out to ascertain the specificity of the covalent labeling observed by autoradiography. First, it was important to eliminate the possibility that radioactive glucagon was artifactually cross-linked to the carrier pro- teins present in the binding buffer. Second, it had to be shown that other radioactive peptides did not produce an identical pattern to the one observed for glucagon. Third, we had to ascertain whether the labeling of the 62,000 Mr band reflected high binding affinity comparable to that described for hepatic glucagon receptor. Cross-linking experiments carried out using carrier proteins of different size-BSA and gelatin-resulted in identical labeling patterns (data not shown).
In a second series of experiments, we applied UV cross- linking to membranes preincubated with '25I-labeled in-
This content downloaded from 130.132.123.28 on Fri, 2 May 2014 09:55:58 AMAll use subject to JSTOR Terms and Conditions

Biochemistry: Iwanij and Hur Proc. Natl. Acad. Sci. USA 82 (1985) 327
Min
30
20
10
5
0
FIG. 2. Time course of incorporation of 251I-labeled glucagon into the 62,000 Mr species. Densitometric scans of autoradiograms were obtained after NaDodSO4/PAGE of liver plasma membranes UV-irradiated for the indicated times.
sulin. As shown in Fig. 3, a single band of Mr 116,000, corre- sponding to that identified by others as an a subunit of the insulin receptor (15), was labeled by UV irradiation. No ra- dioactive band was observed when binding was carried out in the presence of excess unlabeled insulin.
Subsequently, we tested the specificity of the labeling of glucagon receptor in unlabeled glucagon competition experi- ments. When the binding of 125I-labeled glucagon to liver plas- ma membranes was carried out in the presence of an excess of unlabeled glucagon (20 ,uM) prior to UV irradiation, no radiolabeled band was observed (Fig. 4). Further evidence that the 62,000 Mr species represents a receptor-ligand com- plex was obtained from dose-response competition experi- ments. Increasing the concentration of unlabeled glucagon markedly reduced labeling of the 62,000 Mr band (Fig. 4). A 50% decrease in the amount of radioactive glucagon cross- linked to the 62,000 Mr band was achieved in the presence of 1.5-2.0 nM unlabeled glucagon, a range similar to that re- ported for the Kd for glucagon binding to liver plasma mem-
A B Mr X 10-3
116-
66- FIG. 3. UV-induced cross-linking 60- of 125I-labeled insulin to liver plasma
44- membranes. Shown are autoradio- grams of NaDodSO4/PAGE-separated liver plasma membranes for which
24- binding was carried out in the presence (lane A) or absence (lane B) of unla-
I beled insulin. Molecular weight mark- ers are shown to the left. Arrowhead
_ points to the a subunit of the insulin re- ;wt_ ~~ceptor.
~~~~~~~~~-9 A~~~~
7.4x 10 M
2.9x 10-8M
It9x_,7M
FIG. 4. Inhibition by unlabeled glucagon of the covalent incorpo- ration of 125I-labeled glucagon into liver plasma membranes, as shown by scanning densitometry of autoradiograms after NaDod- S04/PAGE. Concentrations of unlabeled glucagon are indicated. Arrow denotes position of the glucagon receptor. Asterisk indicates the top of the separating gel; double asterisk denotes possible recep- tor aggregates.
brane (16, 17). This observation indicates that the labeled protein has properties of the high-affinity glucagon binding sites on liver plasma membrane. We have also carried out binding and cross-linking in the presence of 1 mM GTP, which increases the dissociation rate constant of glucagon (16). The presence of GTP decreased the density of the la- beled band by 35% (data not shown).
Properties of Glucagon Receptor. To gain insight into the structure of the glucagon receptor, we subjected cross- linked membranes to electrophoretic analysis under reduc- ing and nonreducing conditions. In the absence of dithio- threitol, a single labeled band of Mr -60,000 was again pro- duced (Fig. 5). Therefore, glucagon receptor is not an oligomeric structure held together by disulfide bonds. Re- duction of NaDodSO4-solubilized membranes with various amounts of dithiothreitol (1-100 mM) indicated that reduc- tion was complete at 1 mM dithiothreitol. We observed an increase in the apparent Mr of the receptor in the presence of dithiothreitol. From this result, we infer that the receptor may contain a globular domain maintained by disulfide bonds. Similar results were obtained when 1% 2-mercapto- ethanol was used instead of dithiothreitol.
We tested the effect of protease inhibitors on the labeling pattern, since proteolytic degradation during membrane iso- lation or during the course of the experiment may significant- ly modify receptor polypeptides. When protease inhibitors such as leupeptin, pepstatin, and Trasylol were included dur- ing the membrane-isolation step, cross-linking experiments resulted in a somewhat tighter band, although there was no significant change in the apparent Mr of the receptor.
We tested whether the 62,000 Mr species behaves as an integral membrane protein by carrying out membrane ex- traction procedures known to remove peripheral proteins. Extraction of liver plasma membrane with 0.2 M sodium car- bonate, pH 11, or with 10 mM lithium diiodosalicylate before or after cross-linking of glucagon to its receptor sites did not abolish labeling of the 62,000 Mr band (data not shown), fur- ther substantiating that it represents the receptor molecule.
Since glucagon receptor can be solubilized with digitonin
This content downloaded from 130.132.123.28 on Fri, 2 May 2014 09:55:58 AMAll use subject to JSTOR Terms and Conditions
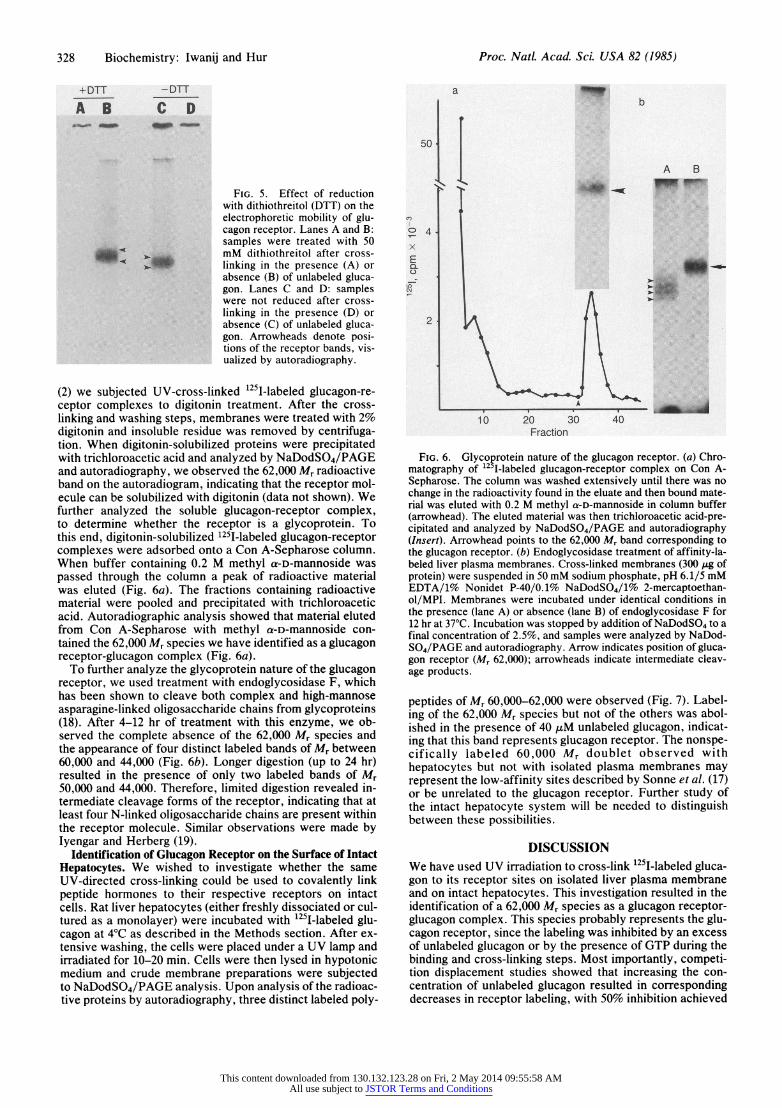
328 Biochemistry: Iwanij and Hur Proc. Natl. Acad. Sci. USA 82 (1985)
+DTT -DTT
A B C D
FIG. 5. Effect of reduction with dithiothreitol (DTT) on the electrophoretic mobility of glu- cagon receptor. Lanes A and B: samples were treated with 50 mM dithiothreitol after cross- linking in the presence (A) or absence (B) of unlabeled gluca- gon. Lanes C and D: samples were not reduced after cross- linking in the presence (D) or absence (C) of unlabeled gluca- gon. Arrowheads denote posi- tions of the receptor bands, vis- ualized by autoradiography.
(2) we subjected UV-cross-linked 125I-labeled glucagon-re- ceptor complexes to digitonin treatment. After the cross- linking and washing steps, membranes were treated with 2% digitonin and insoluble residue was removed by centrifuga- tion. When digitonin-solubilized proteins were precipitated with trichloroacetic acid and analyzed by NaDodSO4/PAGE and autoradiography, we observed the 62,000 Mr radioactive band on the autoradiogram, indicating that the receptor mol- ecule can be solubilized with digitonin (data not shown). We further analyzed the soluble glucagon-receptor complex, to determine whether the receptor is a glycoprotein. To this end, digitonin-solubilized 1251-labeled glucagon-receptor complexes were adsorbed onto a Con A-Sepharose column. When buffer containing 0.2 M methyl a-D-mannoside was passed through the column a peak of radioactive material was eluted (Fig. 6a). The fractions containing radioactive material were pooled and precipitated with trichloroacetic acid. Autoradiographic analysis showed that material eluted from Con A-Sepharose with methyl a-D-mannoside con- tained the 62,000 Mr species we have identified as a glucagon receptor-glucagon complex (Fig. 6a).
To further analyze the glycoprotein nature of the glucagon receptor, we used treatment with endoglycosidase F, which has been shown to cleave both complex and high-mannose asparagine-linked oligosaccharide chains from glycoproteins (18). After 4-12 hr of treatment with this enzyme, we ob- served the complete absence of the 62,000 Mr species and the appearance of four distinct labeled bands of Mr between 60,000 and 44,000 (Fig. 6b). Longer digestion (up to 24 hr) resulted in the presence of only two labeled bands of Mr 50,000 and 44,000. Therefore, limited digestion revealed in- termediate cleavage forms of the receptor, indicating that at least four N-linked oligosaccharide chains are present within the receptor molecule. Similar observations were made by Iyengar and Herberg (19).
Identification of Glucagon Receptor on the Surface of Intact Hepatocytes. We wished to investigate whether the same UV-directed cross-linking could be used to covalently link peptide hormones to their respective receptors on intact cells. Rat liver hepatocytes (either freshly dissociated or cul- tured as a monolayer) were incubated with 125I-labeled glu- cagon at 4?C as described in the Methods section. After ex- tensive washing, the cells were placed under a UV lamp and irradiated for 10-20 min. Cells were then lysed in hypotonic medium and crude membrane preparations were subjected to NaDodSO4/PAGE analysis. Upon analysis of the radioac- tive proteins by autoradiography, three distinct labeled poly-
a
501
Pt A B
T_ 4 : :
x E
LO
2
10 0 30 40 Fraction
FIG. 6. Glycoprotein nature of the glucagon receptor. (a) Chro- matography of "251-labeled glucagon-receptor complex on Con A- Sepharose. The column was washed extensively until there was no change in the radioactivity found in the eluate and then bound mate- rial was eluted with 0.2 M methyl a-D-mannoside in column buffer (arrowhead). The eluted material was then trichloroacetic acid-pre- cipitated and analyzed by NaDodSO4/PAGE and autoradiography (Insert). Arrowhead points to the 62,000 Mr band corresponding to the glucagon receptor. (b) Endoglycosidase treatment of affinity-la- beled liver plasma membranes. Cross-linked membranes (300 ,ug of protein) were suspended in 50 mM sodium phosphate, pH 6.1/5 mM EDTA/1% Nonidet P-40/0. 1% NaDodSO4/1% 2-mercaptoethan- ol/MPI. Membranes were incubated under identical conditions in the presence (lane A) or absence (lane B) of endoglycosidase F for 12 hr at 37?C. Incubation was stopped by addition of NaDodSO4 to a final concentration of 2.5%, and samples were analyzed by NaDod- S04/PAGE and autoradiography. Arrow indicates position of gluca- gon receptor (Mr 62,000); arrowheads indicate intermediate cleav- age products.
peptides of Mr 60,000-62,000 were observed (Fig. 7). Label- ing of the 62,000 Mr species but not of the others was abol- ished in the presence of 40 ,uM unlabeled glucagon, indicat- ing that this band represents glucagon receptor. The nonspe- cifically labeled 60,000 Mr doublet observed with hepatocytes but not with isolated plasma membranes may represent the low-affinity sites described by Sonne et al. (17) or be unrelated to the glucagon receptor. Further study of the intact hepatocyte system will be needed to distinguish between these possibilities.
DISCUSSION We have used UV irradiation to cross-link 125I-labeled gluca- gon to its receptor sites on isolated liver plasma membrane and on intact hepatocytes. This investigation resulted in the identification of a 62,000 Mr species as a glucagon receptor- glucagon complex. This species probably represents the glu- cagon receptor, since the labeling was inhibited by an excess of unlabeled glucagon or by the presence of GTP during the binding and cross-linking steps. Most importantly, competi- tion displacement studies showed that increasing the con- centration of unlabeled glucagon resulted in corresponding decreases in receptor labeling, with 50% inhibition achieved
This content downloaded from 130.132.123.28 on Fri, 2 May 2014 09:55:58 AMAll use subject to JSTOR Terms and Conditions

Biochemistry: Iwanij and Hur Proc. Nati Acad. Sci. USA 82 (1985) 329
A B
*4~ C FIG. 7. UV-mediated cross-linking of 125[
labeled glucagon to the receptor sites on in- tact hepatocytes, as visualized by autoradiog- raphy after NaDodSO4/PAGE of membrane preparations from cells labeled with 125I-la- beled glucagon in the presence (lane A) or ab- sence (lane B) of excess unlabeled glucagon. Arrow points to the glucagon receptor (Mr 62,000). Asterisk indicates the nonspecifically labeled bands.
at physiological concentration of the peptide. The receptor behaves as an integral membrane protein: it
is not removed from the lipid bilayer by extraction at basic pH or with lithium diiodosalicylate. Our estimate of an Mr of 62,000 for the glucagon receptor is higher than that reported by Johnson et al. (5) but closer to the size determined by Rodbell (16), who used target-size analysis. Demoliou-Ma- son and Epand (6) reported that glucagon receptor may con- sist of two polypeptides of Mr 53,000 and 70,000. The recep- tor band observed in the present study appears sometimes as a closely spaced doublet, but the spacing is not as broad as that described by Demoliou-Mason and Epand (6). We have investigated the organization of the glucagon receptor by carrying out NaDodSO4/PAGE analysis in the presence and absence of reducing agents. A single band was observed regardless of the conditions used, indicating that the recep- tor is not present in the membrane as an oligomer linked to disulfide bonds. However, we cannot rule out the possibility that complexes may exist in the form of noncovalently inter- acting subunits.
Reduction of disulfide bonds resulted in an increase of the receptor's apparent Mr, indicating that the receptor contains globular regions maintained by disulfide bridges. Similar ob- servations were made by Demoliou-Mason and Epand (6). Further characterization of the cross-linked glucagon-recep- tor complexes showed that the receptor is a glycoprotein, since it could be adsorbed onto and subsequently released with competing sugar from an immobilized Con A column. Limited digestion of the receptor with endoglycosidase F re- sulted in the formation of four intermediate species of lower apparent Mr indicating that the receptor possesses at least four N-glycosydic linkages.
Bregman and Levy (4) reported the labeling of a 25,000 Mr polypeptide. Johnson et al. (5) and Demoliou-Mason and Epand (6) also reported the presence of a low Mr component in addition to a 55,000 Mr polypeptide. We have never ob- served a 25,000 Mr component when intact hepatocytes were labeled. This component was observed in membrane prepa- rations that were repeatedly thawed or stored at -20?C in- stead of in liquid nitrogen. Our interpretation is that the 25,000 Mr polypeptide represents a degraded receptor mole- cule.
Bifunctional reagents, such as disuccinimidyl suberate, were extremely useful in the identification of insulin recep- tor (14) and of other hormone receptors (20, 21). Johnson et al. (5) reported that disuccinimidyl suberate and other homo- bifunctional reagents were not able to cross-link glucagon to
its receptor. Since disuccinimidyl suberate requires func- tional amino groups in close proximity to the binding site to effect cross-linking, it may be that the glucagon-receptor complex does not possess amino groups in the right config- uration. UV irradiation has been used to achieve direct cova- lent attachment with a variety of nonpeptide ligands (22, 23) and with protein-RNA complexes (24) but has been used only to a limited extent to elucidate peptide-protein interac- tion (25, 26).
The advantage of direct cross-linking by UV irradiation is its simplicity; this procedure eliminates the need for chemi- cally modified peptides or the addition of cross-linking arms. Direct cross-linking by UV irradiation can be used with iso- lated plasma membranes or with live cells and thus may prove to be a powerful analytical tool in identification and characterization of peptide hormone receptors.
We thank Dr. Steve Rosenzweig for the gift of endoglycosidase F, Drs. Dana Giulian and Sara E. Barsel for critical reading of this manuscript, and Ms. Karen Backlund for help in manuscript prepa- ration. This work was supported by National Science Foundation Grant PCM 8118859H01 and by University of Minnesota Graduate School funds.
1. Unger, R. M., Dobbs, R. E. & Orci, L. (1978) Annu. Rev. Physiol. 40, 307-343.
2. Iwanij, V. (1978) Dissertation (The Rockefeller Univ., New York).
3. Welton, A. F., Lad, P. M., Newby, A. C., Yamamuka, H., Nicosia, S. & Rodbell, M. (1977) J. Biol. Chem. 252, 5947- 5950.
4. Bregman, M. D. & Levy, D. (1977) Biochem. Biophys. Res. Commun. 78, 584-590.
5. Johnson, G. L., MacAndrew, V. I., Jr., & Pilch, P. F. (1981) Proc. Natl. Acad. Sci. USA 78, 875-878.
6. Demoliou-Mason, C. & Epand, R. M. (1982) Biochemistry 21, 1996-2004.
7. Lin, M. C., Wright, D. E., Hruby, V. J. & Rodbell, M. (1975) Biochemistry 14, 1559-1563.
8. Neville, D. M. (1968) Biochim. Biophys. Acta 154, 540-552. 9. Seglen, P. 0. (1973) Exp. Cell. Res. 82, 391-398.
10. Hubbard, A. L. & Ma, A. (1983) J. Cell Biol. 96, 230-239. 11. Porter, S. & Froehner, S. C. (1983) J. Biol. Chem. 258, 10034-
10040. 12. Rosenzweig, S. A., Madison, L. D. & Jamieson, J. D. (1984)
J. Cell Biol. 99, 1110-1116. 13. Laemmli, U. K. (1970) Nature (London) 227. 680-685. 14. Udenfriend, S., Stein, S., Bohlen, P., Dairman, W., Lein-
gruber, W. & Weigele, M. (1972) Science 178, 871-873. 15. Pilch, P. F. & Czech, M. P. (1979) J. Biol. Chem. 254, 3375-
3381. 16. Rodbell, M. (1980) Nature (London) 284, 17-22. 17. Sonne, O., Berg, T. & Christoffersen, T. (1978) J. Biol. Chem.
253, 3203-3210. 18. Elder, J. H. & Alexander, S. (1982) Proc. Natl. Acad. Sci.
USA 79, 4540-4544. 19. Iyengar, R. & Herberg, J. T. (1984) J. Biol. Chem. 259, 5222-
5229. 20. Paglin, S. & Jameson, J. D. (1982) Proc. Natl. Acad. Sci. USA
79, 3739-3743. 21. Bonifacino, J. S. & Dufau, M. L. (1984) J. Biol. Chem. 259,
4542-4549. 22. Pressin, J. E., Tilloston, L. G., Yamada, K., Gitomer, W.,
Carter-Su, C., Mora, R., Isselbacher, K. J. & Czech, M. P. (1982) Proc. Natl. Acad. Sci. USA 79, 2286-2290.
23. Akera, T. & Brody, T. M. (1969) Mol. Pharmacol. 5, 605-614. 24. Brimacombe, R., Stoffler, G. & Wittman, H. G. (1978) Annu.
Rev. Biochem. 47, 217-249. 25. Oswald, R. E. & Changeux, J-P. (1982) FEBS Lett. 139, 225-
229. 26. Svoboda, M., Lambert, M., Furnell, J. & Christophe, J. (1982)
Reg. Pept. 4, 164-172.
This content downloaded from 130.132.123.28 on Fri, 2 May 2014 09:55:58 AMAll use subject to JSTOR Terms and Conditions