dinamika gen dalam populasi
DESCRIPTION
evolusiTRANSCRIPT

Dinamika Gen dalam Populasi
1. Ardiani Samti NA2. Yulya Fatma

Perubahan dalam Frekuensi Alel
Lokus adalah lokasi genom atau kromosom sebuah gen
Alel adalah bentuk alternatif gen pada sebuah lokus.
Populasi relatif sebuah alel disebut sebagai frekuensi alel atau frekuensi gen
Frekuensi alel A1
.
Frekuensi alel A2

lebih sederhana
frekuensi alel dalam dalam populasi dari generasi ke generasi terjadi dengan cara yang unik
dapat terprediksi
frekuensi alel muncul secara probabilistik


Seleksi alam
Reproduksi yang berbeda pada individu bergentik yang berbeda atau genotip dalam sebuah populasi
perbedaan dalam individu meliputi mortalitas, fertilitas, kesuburan, kesuksesan dalam perkawinan dan
keberlangsungan hidup keturunannya.
Reproduksi diferensial terjadi

Genotip fitness
mengukur kemampuan individu untuk bertahan dan bereproduksi
Frekueksi alel A1 = pFrekuensi alel A2 = q = 1-p

Frekuensi A2 untuk generasi berikutnya = q(t + 1)

Kodominan
Kodominan adalah seleksi gen , dua alel homozigot memiliki niai fitness yang berbeda, dimana nilai fitness alael heterozigot merupakan rata-rata dari genotip fitness alel homozigot.
frekuensi alel A2 tipe gerenarsinya adalah


Dominansi

Overdominansi dan underdominace
Perubahan frekuensi alel, untuk seleksi overdominan. Frekuensi alel dari atas ke bawah pada kurva 0.99, 0.75, 0.50 , 0.25, dan 0.01. s= 0.04 dan t = 0.02. Semenjak nilai s dan t ditolak, perubahan dari frekuensi alel semakin cepat. Sebagai catatan bahwa kesembangan stabil pada q = 0.667Perubahan frekuensi alel untuk seleksi underdominan. Frekuensi inisial dari atas ke bawah pada kurva 0.75,0.668, 0.667, 0.666, 0.50 dan 0.20, s= - 00.02 dan t = - 0.01. Sekali lagi karena nilai s dan t yang besar. Perubahan freuensi alel semakin cepat . Sebagai catatan akan mencapai keseimbangan alel q = 0.0667. Keseimbangan ini tidak stabi semenjak devisiasi sedikitpun dari alael yang dieliminasi dalam populasi

Random Genetic Drift

Perubahan frekuensi alel menjadi hanyutan genetik acak dalam populasi dengan ukuran yang
berbeda. Populasi terkecil (N= 25) mencapai kondisi tetap setelah 42 generasi.
Dua populasi lain masih polimorphic setelah 150 generasi, tetapi akan mencapai ketetatapn utimat jika eksperimen dilanjutkan secara berkelanjutkan

Setiap generasi, 25 alel yang disampelkan diganti dari generasi sebelumnya dalam populasi diwakii
oleh garis abu-abu, alel menjadi tetap pada generasi ke 27 pada populasi lain alel hilang pada
generasi 49

Probabilitas untuk menjaga frekuensi alel initial sepanjang wakt
selama dua seleksi netral apada alel.

Ukuran Efektif Populasi
Parameter dasar dalam populasi biologi adalah sensus ukuran
populasi (N)
Sudut pandang genetika populasi dan evolusi
jumlah individu yang dianggap relevan hanya terdiri dari orang-
orang yang secara aktif berpartisipasi dalam reproduksi
Ukuran populasi efektif (Ne)
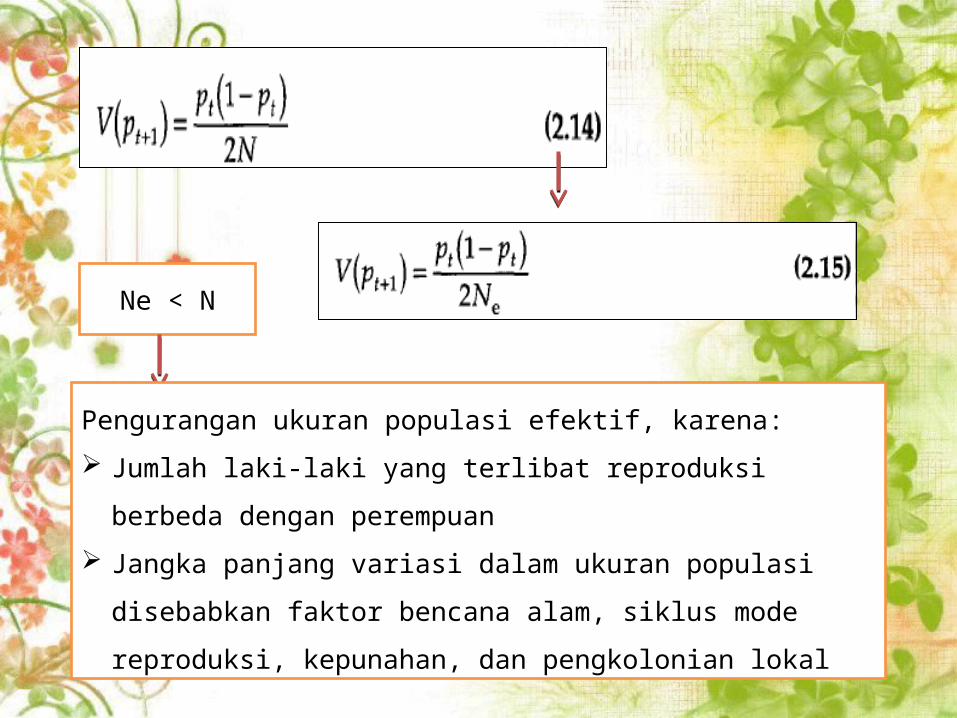
Ne < N
Pengurangan ukuran populasi efektif, karena:
Jumlah laki-laki yang terlibat reproduksi berbeda dengan
perempuan
Jangka panjang variasi dalam ukuran populasi disebabkan faktor
bencana alam, siklus mode reproduksi, kepunahan, dan
pengkolonian lokal

Substitusi Gen
Proses dimana alel mutan sepenuhnya menggantikan alel dominan atau
jenis alel wild dalam populasi
Waktu fiksasiAlel mutan mencapai fiksasi
Tidak semua alel mutan mencapai fiksasi
Kenyataan
Perlu diperhitungkan laju substitusi gen

Fixation Probability
alel tertentu akan menjadi tetap dalam populasi tergantung pada:
1)frekuensi,
2)selektif menguntungkan atau merugikan (s),
3)ukuran populasi efektif (Ne).
Kimura (1962)
Pada alel netral, probabilitas fiksasi = frekuensi dalam
populasi.

Untuk mutasi netral, s=0, maka persamaan 2.19 menjadi
Jika ukuran populasi = ukuran populasi efektif, persamaan 2.19
mengurangi
Untuk nilai-nilai positif s dan nilai-nilai besar n, persamaan 2.22
mengurangi

Contoh
Mutan baru muncul dalam populasi adalah 1.000 orang. Bagaimana
kemungkinan bahwa alel ini akan menjadi tetap dalam populasi jika (1)
netral, (2) selektif menguntungkan 0.01, atau (3) selektif merugikan
0.001? Kita asumsikan bahwa N = Ne.
kasus netral, probabilitas fiksasi dihitung dengan menggunakan
persamaan 2.20 adalah 0,05%. Dari persamaan 2.23 dan 2.21, kita
memperoleh probabilitas 2% dan 0,004% untuk mutasi-mutasi
menguntungkan dan merugikan.
Mutasi menguntungkan tidak selalui menjadi tetap dalam populasi

kemungkinan fiksasi untuk mutasi menguntungkan tetap sama,
untuk mutasi netral menjadi lebih kecil, dan untuk alel merugikan
menjadi tak terlepaskan dari nol
jika ukuran populasi efektif diperbesar menjadi 10.000, maka
probabilitas fiksasi menjadi 0.005% (netral), 2% (mutasi
menguntungkan) , dan ~10-20 (mutasi merugikan)

Fixation Time
waktu yang dibutuhkan untuk fiksasi/hilangnya alel tergantung pada:
1)frekuensi,
2)selektif menguntungkan atau merugikan,
3)ukuran populasi.
Kimura dan Ohta (1969) merumuskan waktu fiksasi bersyarat untuk
mutasi netral diperkirakan oleh
untuk mutasi dengan selektif menguntungkan,
s, diperkirakan oleh

Contoh
Asumsikan bahwa spesies mamalia memiliki ukuran populasi efektif
sekitar 106 dan rata-rata waktu generasi 2 tahun. Di bawah kondisi ini,
dibutuhkan mutasi netral, rata-rata, 8 juta tahun menjadi tetap dalam
populasi. Sebagai perbandingan, mutasi dengan selektif
menguntungkan 1% akan menjadi tetap dalam populasi yang sama
hanya sekitar 5.800 tahun. Menariknya, waktu fiksasi bersyarat untuk
alel merugikan dengan selektif merugikan adalah sama untuk alel
menguntungkan dengan selektif menguntungkan (Maruyama dan
Kimura 1974).

Representasi skematik dari dinamika substitusi gen untuk mutasi
(a) menguntungkan dan (b) netral

Rate of Gen Substitution
jumlah mutan mencapai fiksasi per satuan waktu.
Mutasi netral
Jika mutasi netral terjadi pada laju u per gen per generasi, maka
jumlah mutan yang timbul pada lokus pada populasi diploid ukuran N
adalah 2Nu per generasi. Karena kemungkinan fiksasi untuk
masing-masing mutasi-mutasi ini adalah 1/(2N), kita memperoleh laju
substitusi alel netral

Populasi besar, jumlah mutasi tiap generasi tinggi tapi probabilitas
fiksasi tiap mutasi rendah.
Populasi kecil jumlah mutasi tiap generasi rendah api probabilitas
fiksasi tiap mutasi tinggi.
Mutasi menguntungkan
laju substitusi untuk kasus pilihan gen tergantung pada
1)ukuran populasi (N),
2)keuntungan selektif (s),
3)kadar mutasi (u).

Polimorfisme Genetik
Monomorfik → hanya satu alel pada sebuah lokus
Polimorfik → ada dua atau lebih alel pada sebuah lokus, hanya jika frekuensi
alel paling umum kurang dari 99%
Gen Diversity
Cara mengukur polimorfisme dalam populasi:
Menghitung rata-rata proporsi polimorfik lokus (P) dengan
membagi jumlah lokus polimorfik dengan jumlah lokus sampel.

Ukuran variabilitas genetik → rata-rata heterozygosity yang diharapkan
(keragaman gen).
Keragaman gen di lokus diharapkan heterozygosity, didefinisikan
sebagai
xi = frekuensi alel i
m = jumlah total alel di lokus
h = probabilitas dua alel yang dipilih secara acak
H = rata-rata nilai h atas semua lokus dipelajari, digunakan sebagai
perkiraan tingkat variabilitas genetik dalam populasi
n = jumlah lokus

Nucleotide Diversity
Ukuran keanekaragaman gen sebagaian besar tidak cocok untuk data
sekuensing DNA karena tingkat variasi genetik di tingkat DNA di alam
cukup tinggi
Ukuran keragaman gen ini tidak akan membedakan antara lokus
berbeda atau populasi dan tidak lagi menjadi ukuran yang informatif
dari polymorphism

Setiap urutan dalam kelompok diwakili hanya sekali → nilai-nilai
ukuran keragaman lokus tunggal akan sama untuk kedua kelompok
Untuk data urutan DNA, ukuran yang lebih tepat dari polymorphism
dalam populasi adalah jumlah rata-rata perbedaan nukleotida per
situs antara setiap dua urutan yang dipilih secara acak.
Sekuens (a) kurang polimorfik dibanding sekuens (b)
tetapi nilai h dan H akan sama untuk kedua urutan

lebih bervariasi
xi = frekuensi alel i
xj = frekuensi alel j
Πij = proporsi nukleotida berbeda

Kekuatan Pendorong dalam Evolusi
3 hipotesis berkaitan pentingnya hanyutan genetik acak untuk berbagai
pilihan dalam menentukan hasil evolusi:
o Hipotesis mutasionist
Fenomena evolusi dijelaskan melalui efek masukan mutasi dan
hanyutan genetik acak
o Hipotesis neuralist
Fenomena evolusi melalui penekanan efek mutasi, hanyutan genetik
acak, memurnikan seleksi
o Hipotesis selectionist
Menekankan efek menguntungkan dan menyeimbangkan mode
seleksi sebagai pendorong utama proses evolusi

Teori neo-Darwin
Teori neo-Darwinisme
Mutasi diakui sebagai sumber utama variasi genetik, seleksi alam
(yaitu, positif) memberikan peran yang dominan dalam membentuk
genetik populasi dan dalam proses substitusi gen
Seleksi dianggap sebagai satu-satunya kekuatan yang mampu
mengemudi proses evolusi. Faktor-faktor seperti mutasi dan hanyutan
genetik acak dianggap sebagai kontributor minor
pan-selectionism

Persepsi selectionist
o Substitusi gen terjadi sebagai konsekuensi dari seleksi untuk
mutasi-mutasi yang menguntungkan.
o Polimorfisme, di sisi lain, dikelola oleh keseimbangan seleksi.
substitusi dan polimorfisme merupakan dua fenomena terpisah yang
didorong oleh kekuatan evolusi yang berbeda
Gen substitusi adalah hasil akhir dari proses adaptif yang positif
dimana alel baru mengambil alih generasi mendatang dari populasi jika
dan hanya jika itu meningkatkan kemampuan organisme, sementara
polimorfisme dipertahankan ketika dua alel atau lebih
berdampingan di lokus menguntungkan bagi organisme.

Kimura (1968), mendalilkan
mayoritas perubahan molekuler dalam evolusi adalah karena
fiksasi acak mutasi netral atau hampir netral
Teori evolusi melekuler netral
pada tingkat molekuler sebagian besar perubahan evolusi dan banyak
variabilitas dalam spesies disebabkan tidak dengan seleksi positif alel
menguntungkan maupun dengan keseimbangan seleksi, tapi oleh
hanyutan genetik acak dari alel mutan melalui selektif netral
nasib alel ditentukan terutama oleh hanyutan genetik acak

Teori netral
frekuensi alel ditentukan terutama oleh aturan stokastik, dan
gambar yang kita peroleh pada waktu tertentu adalah semata-mata
daerah sementara yang mewakili bingkai sementara dari proses
dinamis yang berkelanjutan
substitusi dan polimorfisme sebagai dua aspek dari
fenomena yang sama
Teori netral juga tidak menghalangi adaptasi
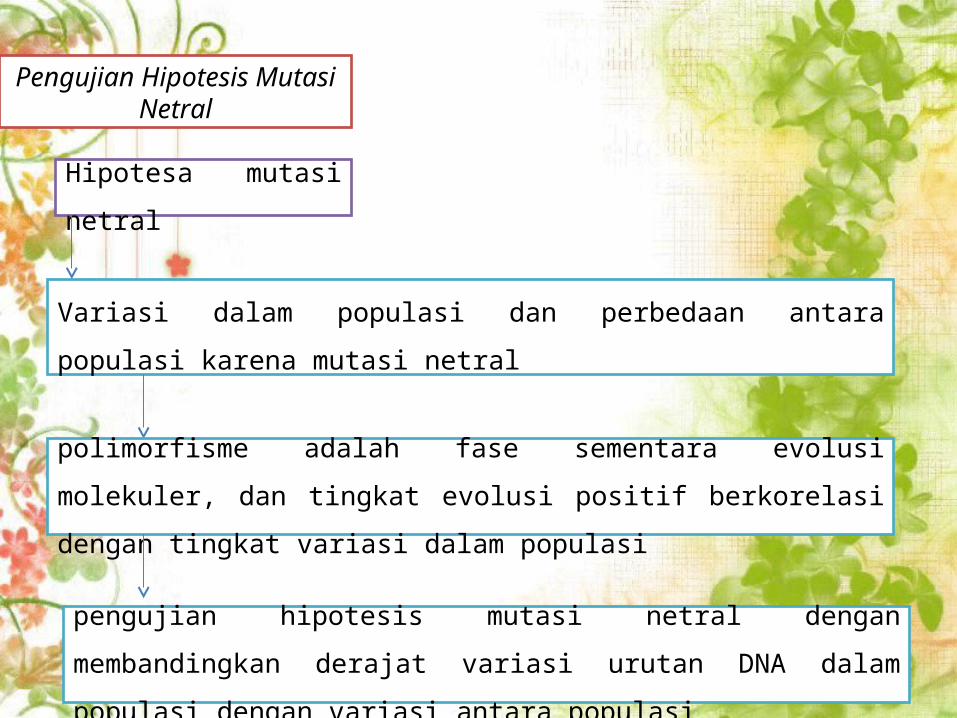
Pengujian Hipotesis Mutasi Netral
Hipotesa mutasi netral
Variasi dalam populasi dan perbedaan antara populasi karena mutasi
netral
pengujian hipotesis mutasi netral dengan membandingkan derajat
variasi urutan DNA dalam populasi dengan variasi antara populasi
polimorfisme adalah fase sementara evolusi molekuler, dan tingkat
evolusi positif berkorelasi dengan tingkat variasi dalam populasi

Metode sederhana yang diusulkan oleh McDonald dan Kreitman (1991)
situs nukleotida dalam urutan dikatakan polimorfik jika menunjukkan
setiap variasi dalam satu atau kedua spesies.
Situs dianggap mewakili perbedaan yang tetap antara dua spesies jika
itu menunjukkan variasi tidak intraspecific dalam spesies tapi berbeda
antara spesies.
Polimorfik dan perbedaan situs tetap dibagi lagi ke dalam dua
kategori, synonymous dan nonsynonymous yang didasarkan pada asumsi:
hanya mutasi nonsynonymous mungkin adaptif,
mutasi synonymous selalu netral, dan
mutasi selektif menguntungkan akan diperbaiki dalam populasi jauh
lebih cepat daripada mutasi netral

Berdasarkan hipotesis mutasi netral, harapan adalah bahwa rasio dari
perbedaan tetap nonsynonymous untuk perbedaan tetap synonymous
akan sama dengan rasio polimorfisme nonsynonymous untuk
polimorfisme synonymous

Pertanyaan:1. Hamima. Grafik b.Siklus mode reproduksi c. Bagaimana hubungan alel yang tidak mampu
bertahan dengan substitusi dengan genetik drif
d.Laju substitusi genetik2. WULAN Hukum hardy weinberg?