diel n2 fixation in an intertidal marine cyanobacterial mat community
TRANSCRIPT

Marine Chemistry, 16 (1985) 369--377 369 Elsevier Science Publishers B.V., Amsterdam -- Printed in The Netherlands
DIEL N 2 FIXATION IN AN INTERTIDAL MARINE CYANOBACTERIAL MAT COMMUNITY
MARK F. BAUTISTA
Marine Sciences Research Center, SUNY at Stony Brook, Stony Brook, NY 11794 (U.S.A.)
HANS W. PAERL
Institute of Marine Sciences, University of North Carolina, Morehead City, NC 28557 (U.S.A.)
(Received June 6, 1984; accepted September 25, 1984)
ABSTRACT
Bautista, M.F. and Paerl, H.W., 1985. Diel N2 fixation in an intertidal marine cyano- bacterial mat community. Mar. Chem., 16: 369--377.
Diurnal and diel rates of nitrogen fixation (acetylene reduction) were measured in an intertidal marine mat community consisting of Mierocoleus chthonoplastes, Lyngbya aestuarii, and an underlying layer of anaerobic bacteria (probably photosynthetic, fermentative, and sulfate reducing bacteria). The community is located on Shackleford Banks, a barrier island off the coast of North Carolina. Rates of nitrogenase activity during the diurnal period were generally highest in the morning. Rates measured during the day, but incubated in the dark, at times equalled those of samples incubated continuously in the light. Fixation also occurred at night. Mat samples deprived of light prior to the acetylene reduction assay continued to reduce acetylene during the night and into the following day, although at lower rates than samples exposed to light. The persistence of dark nitrogenase activity serves as evidence that reducing power initially generated through photosynthesis can be stored and subsequently utilized through heterotrophic metabolism. Although cyanobacteria are responsible for light-mediated nitrogenase activity, non-photosynthetic bacterial contributions to dark-mediated nitrogenase activity cannot be ruled out.
INTRODUCTION
Cyanobacterial mat communities often thrive conspicuously on intertidal sand and mud flats in tropical and temperate coastal environments. Such environments are characteristically low in biologically available nitrogen (Ryther and Dunstan, 1971); accordingly, the community 's presence and persistence may be largely due to nitrogen fixation supplying communi ty nitrogen demands.
Mat communities are consortia of autotrophic and heterotrophic bacteria, some of which possess the nitrogenase enzyme (Whitney et al., 1975; Bohlool and Wiebe, 1978; Carpenter et al., 1978; Gotto et al., 1981). The fixation of nitrogen by nitrogenase requires a reducing environment since the activity of the enzyme is sensitive to oxygen (Stewart and Pearson, 1970). Through oxygen~onsuming metabolism of the mat microbial
0304-4203/85/$03.30 © 1985 Elsevier Science Publishers B.V.

370
constituents, reducing environments are created and maintained, allowing nitrogenase activity (NA) to take place. In aerobic aquatic habitats, NA is often confined to filamentous cyanobacteria possessing heterocysts, specialized cells in which NA occurs (Stewart, 1977). However, mat communities we observed were devoid of such genera. Instead, non- heterocystous genera proved dominant throughout the entire growth season (February to October). NA has been reported in Microcoleus chthonoplastes (Pearson et al., 1979), a non-heterocystous form found as a major consti tuent in early forming mats on Shackleford Banks (Polimeni, 1976). Although Pearson et al. (1979) argue that the oxygen protect ion mechanism for NA lies within the cells regardless of filament bundling, there is evidence that reduced microzones are present in the bundled filaments (H.W. Paerl, unpublished data) similar to that postulated for the planktonic analogue Trichodesmium (Carpenter and Price, 1976).
Other potential nitrogen fixers include bacteria which are likely to be present in the lower anoxic layers of the mat (e.g., sulfate-reducing, fer- menting, and photosynthet ic bacteria). Reducing conditions are maintained through the consumption of oxygen during respiration of organic compounds. Strong hydrogen sulfide odors and darkly stained sand suggest the presence of sulfate reducers, while in more developed mats, a pink horizon just above the stained sand is visible (probably Chromatium sp.), These photosynthet ic bacteria utilize hydrogen sulfide as an electron donor during photosynthesis , thereby avoiding oxygen inhibition.
The NA (acetylene reduction) we determined in the mat communi ty is the sum of the activity of the constituents. Total activity would depend on nutrient availability, environmental conditions (temperature, light, salinity) and physiological state of the bacteria. In this study, NA was monitored during several diurnal and diel periods using light and dark incubations. By incubating samples under ambient light levels and in the dark, we were able to make a first-order distinction between activity due to autotrophs in the mat and activity attributable to heterotrophic metabolism. This distinction is useful in determining the quantitative importance of nitrogen fixation by the mat communi ty as an input of new nitrogen to the coastal ecosystem, and also for clarifying how reducing conditions necessary for nitrogen fixation develop and persist in the mat community.
MATERIALS AND METHODS
Description o f the study area
The cyanobacterial mat communi ty was located on an intertidal sand fiat on the western end of Shackleford Banks, N.C. (Fig. 1). The horizontal distribution of the mats was patchy and limited to the upper intertidal zone of the sand flat. The mats o f t e n appeared in the midst of short Spartina alterniflora; this grass was cropped to a height of 3--5 cm by the grazing

371
ungulates (cows, horses, sheep, goats) which inhabit the island. Mat thickness ranged from 1 to 8 mm, varying both spatially and temporally. Thinner mats were characteristic of newly developing communities consisting of Lyngbya aestuarii and Microcoleus chthonoplastes (Polimeni, 1976). New mats were observed in the spring and at the periphery of thicker, better-established mats. During summer and early fall a deeper anaerobic layer of photo- synthetic, fermentative, and sulfate-reducing bacteria appeared to develop below the cyanobacterial layer as the mats thickened. Sandy hummocks covered by short Spartina were interspersed throughout the mats. These were usually exposed at high tide, and no mats were present on them.
Acetylene reduction
On four dates during the summer of 1981, NA was measured using a modification of the acetylene reduction method (Hardy et al., 1968). In the first two studies, NA was monitored from 0600 to 2030, while in the last two studies, NA was monitored for the entire diel period. Techniques similar to those of previous investigators (Jones, 1977; Ports and Whitton, 1977; Carpenter et al., 1978; Gotto et al., 1981) were used. The mat was sectioned into 4~m 2 strips and replicate samples (3--4) were placed in 7-ml Vacutainer tubes (Becton-Dickinson). After preincubating for 30min, half the head- space of each tube was withdrawn and replaced with acetylene (Matheson, purified grade, 99.99%) through the gas-tight septum. Experiments indicated that an atmosphere of 50% acetylene: 50% air was necessary to saturate the enzyme while not being toxic. The samples were incubated in situ for 2--3 h. In a subsequent experiment investigating the linearity of acetylene reduction rates, the apparent rate of NA decreased exponentially with increasing incubation length from 1 to 3 h (Bautista, 1984). However, the variability of NA in the longer incubations decreased; therefore, 3-h incubations were used so that significant differences in NA over the diurnal and diel periods could be detected. Incubations were terminated by vigorously shaking the tubes for 30 s to liberate any ethylene produced, after which a sample of the headspace was transferred by syringe to a 4-ml evacuated Vacutainer (B-D). Sterile sand placed in Vacutainers was used as a blank, while mat samples incubated without acetylene were included in the 2--3 September study to determine the amount of naturally produced ethylene.
Two types of dark assays were used to measure the amount of hetero- trophically mediated NA present in the mat community. In the first type ('dark' assays), mat samples were incubated in aluminum foil-covered Vacutainers. In the second type ('covered' assays), an area of the mat surface (approximately 1 m 2 ) was covered with an opaque trash~an lid during the diel study. Photosynthesis was prevented, and any NA would be dependent on heterotrophically derived reducing power (i.e., the metabolism of pre- existing, reduced organic compounds) to generate ATP and NADPH. The covered area was sampled along with the light and dark assays; covered assays were incubated under the lid.

372
i I --~k-...~S H AC K L E FOR o .,./ ~,-"~-- BANKS%
T L , 4 A / ~ C Z P E / 7 r L A / V / / C ' ~ .. 0 ~ . LOOKOUT OCE~IV ~ L J ~ , "C
I km
Fig. 1. Map indicating the location of the study site on Shackleford Banks, North Carolina.
Ethylene production was measured with a Carle 311 analytical gas chro- matograph maintained at 80°C, using a flame ionization detector and a 1-m column packed with Porapak T. Nitrogen was used as the carrier gas. Rates of acetylene reduction were determined by comparing sample peak heights to peak heights of ethylene standards, subtracting the sterile sand blank, and correcting for the length of incubation, the sample-to-headspace ratio, and the sample volume injected.
RESULTS
Diurnal and diel rates o f nitrogenase activity
The first three studies showed similar diurnal patterns of light NA (Table I). Light rates were generally higher in the early morning, with a mid- morning depression in activity observed on 9 June and 15 July. A smaller peak of NA was observed in all three studies after a minimum in the after- noon. The coefficient of variation (CV) in these replicates on average was 27%.
Overall, rates during the 23--24 September study were an order of magnitude lower than those measured earlier in the summer (Table I). Replicates were also quite variable (av. CV = 60%). Highest rates were measured in the afternoon, in contrast to the first three studies, in which maximum rates were observed in the morning. ~ t rates of NA increased throughout the morning to 1230 and then decreased slightly in the after- noon.
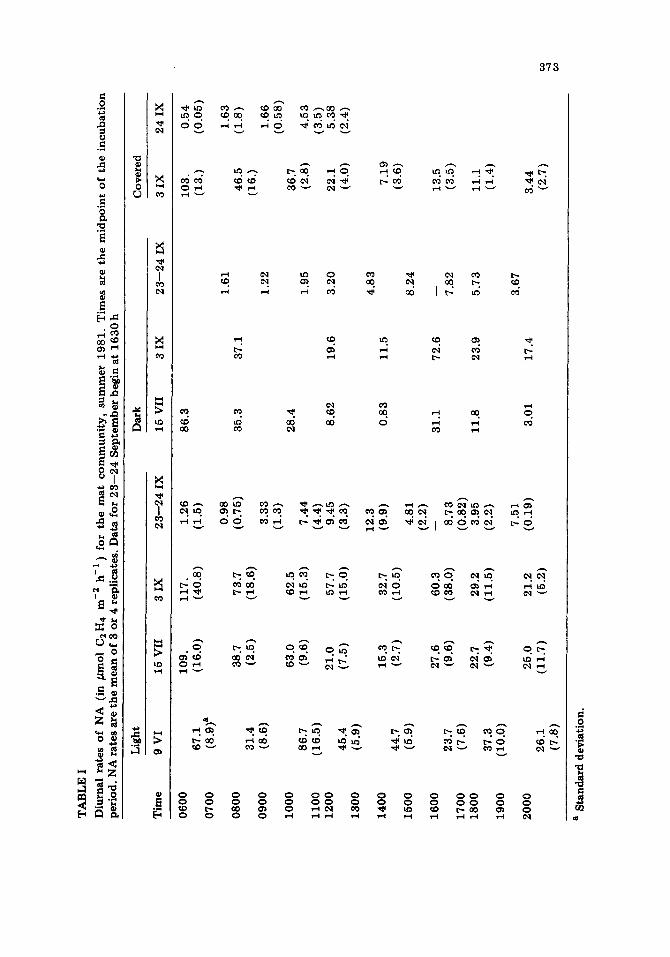
TA
BL
E I
Diu
rnal
rat
es o
f N
A (
in ~
umol
C2H
4 m
-2 h
-1 )
fo
r th
e m
at
com
mu
nit
y,
sum
mer
19
81
. T
imes
are
th
e m
idp
oin
t o
f th
e in
cub
atio
n
per
iod
. N
A r
ates
are
th
e m
ean
of
3 o
r 4
rep
lica
tes.
Dat
a fo
r 2
3--
24
Sep
tem
ber
beg
in a
t 1
63
0 h
Lig
ht
Dar
k C
over
ed
Tim
e 9
VI
15 V
H
3 IX
2
3--
24
IX
15
VII
3
IX
23
--2
4 I
X
3 IX
2
4 I
X
06
00
10
9.
11
7.
1.26
8
6.3
67
.1
(16
.0)
(40
.8)
(1.5
) 0
70
0
(8.9
) a
0.9
8
1.61
0
80
0
38.7
73
.7
(0.7
5)
35.3
37
.1
31.4
(2
.5)
(18
.6)
09
00
(8
.6)
3.33
1.
22
(1.3
) 1
00
0
63
.0
62.5
2
8.4
86
.7
(9.6
) (1
5.3
) 7
.44
1.
95
11
00
(1
6.5
) (4
.4)
12
00
2
1.0
57
.7
9.45
8
.62
1
9.6
3
.20
4
5.4
(7
.5)
(15
.0)
(3.3
) 13
00
(5.9
) 1
2.3
4
.83
1
40
0
15.3
32
.7
(9.9
) 0
.83
11
.5
44.7
(2
.7)
(10
.5)
15
00
(5
.9)
4.81
8
.24
(2
.2)
16
00
2
7.6
6
0.3
--
31
.1
72
.6
--
23.7
(9
.6)
(38
.0)
8.7
3
7.8
2
17
00
(7
.6)
(0.8
2)
18
00
22
.7
29
.2
3.95
11
.8
23
.9
5.7
3
37.3
(9
.4)
(11
.5)
(2.2
) 1
90
0
(10
.0)
7.5
1
3.67
2
00
0
25
.0
21
.2
(0.1
9)
3.01
1
7.4
26
.1
(11
.7)
(5.2
) (7
.8)
103.
0
.54
(1
3.)
(0
.05
)
46.5
(1
6.)
36.7
(2
.8)
22.1
(4
.0)
7.1
9
(3.6
)
13.5
(3
.5)
11.1
(1
.4)
3.44
(2
.7)
1.6
3
(1.8)
1.6
6
(0.5
8)
4.5
3
(3.5)
5.3
8
(2.4
)
¢0
O~
a St
anda
rd deviation.

374
TABLE II
Ratio of dark to light assays for the mat community. Data for 23--24 September begin a~ 1630h
Dark/light Covered/light
Time 15 VII 3 IX 23--24 IX 3 IX 24 IX
0600 0.80 0.88 0700
1.64 0800 0.91 0.50 0.63 0900 0.37 1000 0.45 0.59
0.26 1100 1200 0.41 0.34 0.34 0.38 1300
0.39 1400 0.05 0.35 0.22 1500 1.71 1600 1.13 1.20 -- 0.22
0.90 1700 1800 0.52 0.82 1.45 0.38 1900
0.49 2000 0.12 0.82 0.16
0.43
1.66
0.50
0.61
0.57
In each study, rates of NA assayed wi thout light paralleled and at times equalled rates measured in the corresponding light incubations. Night-time rates during the 2--3 September s tudy were substantial, with an average rate of 96 #mol C2 H4 m- 2 h- 1, while on 23--24 September, night rates were low (~ 4 #mol C2 H4 m - 2 h - i ).
In order to determine the relative contr ibut ion of dark NA during the day, the NA rates o f both types of dark incubations were divided by their respective light rates (Table If). During the daytime, dark NA was responsible for a substantial p ropor t ion of the activity in the early morning and late af ternoon. At midday, dark NA const i tuted less than half o f the total activity measured.
On 2--3 September, the activity in the covered assays was greater than 50% of the total light activity during the momkng and decreased steadily from 0600 to 1400 hours. At every point af ter 1000 h, NA in covered assays was less than half of the total activity, while NA in the dark assays increased to over 100% of the total activity.
DISCUSSION
In order to estimate and compare daffy rates of nitrogen fixation for the Shackleford Banks mat communi ty , the average o f the hour ly NA rates f rom

375
the first three studies (54#mol C2H 4 m -2 h -1 ) was used in calculating the diel average. Assuming the theoretical ratio of 3 C2 H2 reduced: 1 N2 fixed, a square meter of mat fixed 12.1 mg of nitrogen during a diel period. Maximum day and night rates for the mat were 1.09 and 1.04 mg N m -2 h -1 , respectively. The average diel rate was comparable to activity measured in other coastal intertidal environments (20mg N m -2 d -1 , Bohlool and Wiebe, 1978; l l . 0 m g N m -2 d -1, Gotto et al., 1981), salt marshes (10-- 20 mg N m -2 d - l , Carpenter et al., 1978; 4.6 mg N m -2 d -1, Whitney et al., 1975) and a coral reef flat (1 .86--8 .38mgNm -2 d -1, Burris, 1976). Nitrogen fixation rates of this magnitude may be of biological significance to this mat community and may be important to community biomass maintenance.
The mat community was capable of significant dark NA. High rates were detected at night (2--3 September), and dark NA made up a substantial proportion of the activity during the day, except at midday when activity in samples exposed to light and dark assays both reached a minimum (Table I). Bacteria located in the reduced zone of the mat (probably fermentative and sulfate-reducing bacteria) may have been responsible for the majority of this activity, using reducing power and ATP generated from the metabolism of pre-existing reduced carbon compounds. The diurnal and diel studies suggest that the major contribution of NA by this dark, anaerobic component was at periods of low light intensity (early morning, evening and night).
An important difference between the two dark assays further demonstrates the heterotrophic nature of the NA observed. The difference in diurnal and diel patterns of dark NA appears to be due to the length of time the samples were preincubated in the dark. In the dark assays, the mat was darkened only for the period of the assay (in general, 3 h). Previous to the assay, the mat was exposed to ambient light conditions. In the covered assays, a given area of the mat community was deprived of light for the entire photoperiod prior to NA assays. No reducing power for NA could have been generated through photosynthesis in these assays, so only the heterotrophic break- down of pre-existing reduced carbon compounds could have provided the required reducing power.
On the morning of the 2--3 September study, the proportion of dark NA decreased in both types of dark assays (Table II). The proportion of 'covered' activity, after a small increase, continued to decline into the afternoon; however, the 'dark' activity increased to the same level as in light samples by 1600 h. The steady decrease of the proportion of NA in the covered assays may reflect the depletion of reduced carbon compounds in darkness due to energy-requiring processes (e.g., respiration and NA). In the dark assays, photosynthetic generation of reducing power could have continued until the samples were darkened, and this may account for the increased rates of NA in the afternoon.

376
In general, highest rates of NA in the Shackleford mat communi ty were observed in early morning. Other investigators have reported similar patterns of NA in mat communities. In a communi ty of heterocystous cyanobacteria (Scytonema sp.), Potts (1979) observed light NA which increased from 0500 h and peaked at 1100 h. Jones (1977) saw a similar pattern with a peak at 1100 h in a terrestrial mat of Nostoc. Both authors recorded an afternoon peak around 1600h. Carpenter et al. (1978) observed low, variable NA throughout the diel period, with a peak between 1000 and 1200 h. Potts (1979) and Carpenter et al. (1978) cite Horne (1975), who suggested that the drop in activity during the afternoon may be due to competi t ion for reducing power generated during photosynthesis by other biological processes.
The decrease in the afternoon in NA was a consistent feature of diurnal and diel patterns of NA measured in the Shackleford mat community. A similar situation, as described by Home (1975), may exist, where the decrease in NA in the afternoon is due to competi t ion between biological processes requiring reducing power (e.g., NA, photosynthesis, photorespiration). Indeed, diurnal studies of NA and photosynthesis conducted by one of the authors in order to detect a temporal separation between photosynthesis and NA in the mat communi ty suggest that lower rates of NA in the after- noon coincide with higher rates of photosynthesis (Bautista, 1984). How- ever, it was observed that the presence of this temporal separation also depends on the season and the degree of mat maturity.
CONCLUSIONS
Substantial rates of nitrogen fixation were measured in the Shackleford Banks cyanobacterial mat communi ty . A significant proport ion of NA appears to be heterotrophically mediated, particularly at periods of low light intensities. This heterotrophic NA is probably due to the anaerobic bacteria located in the dark, lower layers of the mat communi ty and becomes increasingly important as the mat develops in time and space. The use of dark incubations and light-depriving treatments helps to elucidate the presence and significance of the dark activity and should be considered when studying such communities. In addition, determination of night-time NA rates is also important, since they can comprise a significant proport ion of total diel NA.
While reducing power for NA is ultimately derived through the photo- reduction reaction during photosynthesis, the byproduct , oxygen, inhibits NA. The two processes must therefore be separated, either temporally or spatially. There is evidence that a temporal separation of photosynthesis and NA exists in the cyanobacteria found in the upper layers of the mat (Bautista, 1984). Deeper in the mat, the two processes become spatially separated as oxygen is consumed through biological and chemical processes. The mat structure therefore provides an environment in which biochemical

377
processes necessary for the growth and maintenance of the mat community can take place contemporaneously.
ACKNOWLEDGMENTS
Financial support for this research was provided in part by NSF grant DEB 8104689 and North Carolina Sea Grant Project R/MP-9 to H.W. Paerl.
REFERENCES
Bautista, M.F., 1984. Nitrogen fixation in a marine cyanobacterial mat community. Masters Thesis, University of North Carolina at Chapel Hill, Chapel Hill, NC.
Bohlool, B.B. and Wiebe, W.J., 1978. Nitrogen-fixing communities in an intertidal eco- system. Can. J. Microbiol., 24: 932--938.
Burris, R.H., 1976. Nitrogen fixation by blue green algae of the Lizard Island area of the Great Barrier Reef. Aust. J. Plant Physiol. 3: 41--51.
Carpenter, E.J. and Price, C.C., IV, 1976. Marine Oscillatoria (Trichodesmium): explanation for aerobic nitrogen fixation without heterocysts, Science, 191: 1278-- 1280.
Carpenter, E.J., Van Raalte, C.D. and Valiela, I., 1978. Nitrogen fixation by algae in a Massachusetts salt marsh. Limnol. Oceanogr., 23: 318--327.
Gotto, J.W., Tabita, F.R. and Van Baalen, C., 1981. Nitrogen fixation in intertidal environments of the Texas Gulf Coast. Estuarine Coastal Shelf Sci., 12: 231--235.
Hardy, R.W.F., Holsten, R.D., Jackson, E.K. and Burns, R.C., 1968. The acetylene-- ethylene assay for N: fixation: laboratory and field evaluation. Plant Physiol., 43: 1185--1207.
Horne, A.J., 1975. Algal nitrogen fixation in California streams: diel cycles and nocturnal fixation. Freshwater Biol., 5: 471--477.
Jones, K., 1977. Acetylene reduction by blue-green algae in subtropical grassland. New Phytol., 78: 421--426.
Pearson, H.W., Howsley, R., Kjeldsen, C.K. and Walsby, A.E., 1979. Aerobic nitrogenase activity associated with a non-heterocystous filamentous cyanobacterium. FEMS Lett., 5 : 163--167.
Polimeni, C., 1976. Seasonality and life history of an algal mat community. Masters Thesis, University of North Carolina at Chapel Hill, Chapel Hill, NC.
Potts, M., 1979. Nitrogen fixation (acetylene reduction) associated with communities of heterocystous and non-heterocystous blue-green algae in mangrove forests of Sinai. Oecologia (Berlin), 39:359--373.
Ports, M. and Whitton, B.A., 1977. Nitrogen fixation by blue-green algal communities in the intertidal zone of the lagoon of Aldabra Atoll. Oecologia (Berlin), 27: 275-- 283.
Ryther, J.H. and Dunstan, W.M., 1971. Nitrogen, phosphorus, and eutrophicatioll in the coastal marine environment. Science, 171: 1008--1013.
Stewart, W.D.P., 1977. A botanical ramble among the blue-green algae. Br. Phycol. J., 12: 89--115.
Stewart, W.D.P. and Pearson, H.W., 1970. Effects of aerobic and anaerobic conditions on growth and metabolism of blue-green algae. Proc. R. Soe. London, Ser. B, 175: 293--311.
Whitney, D.E., Woodwell, G.M. and Howarth, R.W., 1975. Nitrogen fixation in Flax Pond: a Long Island salt marsh. Limnol. Oceanogr., 20: 640--643.