development of the head and pectoral skeleton of polypterus with a note on scales (pisces:...
TRANSCRIPT

J. Zool., Lond. (1 984) 204,469-507
Development of the head and pectoral skeleton of Pofypterus with a note on scales (Pisces: Actinopter ygii)
M . J O L L I E Department of Biological Sciences, Northern Illinois University. DeKulb, Illinois 601 1.5 U.S . A .
(Accepted 13 December 1983)
(With 19 figures in the text)
Comparative study of the bony head and pectoral girdle skeletons of fishes requires the estab- lishment of sound and acceptable homologies. In this paper. the bony elements seen in a number of species of Pn/yp/cws (and Erpr/orc/irhys) are described in terms of several developmental stages and the mature condition. Variations in these bones are considered in a developmental sense. As a part of the problem of bone development. a summary account of lateral-line development is included.
A number of particular problems of homology are discussed in some detail. The most radical differences of view involve the endocranial bones. The ideas of Patterson (1975) are examined and an alternative interpretation offered. The main bone in the posterolateral angle of the endo- cranium is viewed as the pterotic-epiotic. The epiotic of the teleost is seen not as the homologue of the epioccipital of the pholidophorid but a part of the so-called “pterotic” of those fishes. Other endocranial problems appear to be of a similar (interpretative) nature. The suspensorium of Po/yp/rrus is peculiar in lacking a symplectic and an interhyal. The branchial skeleton is quite like that of a “palaeoniscoid”. The pectoral girdle presents a number of problems such as the form of the posttemporal, the lack of a sensory canal on the supracleithrum and the lack of an interclavicle.
The subject matter of this paper. and other papers by this author on the development of bones in fishes, lead to several developmental concepts. These are summarized in 18 statements. A table (Table 11) summarizes some of the anatomical information useful in taxonomy in terms of being derived, primitive or unique
Contents
. . . . . . . . . . Introduction . . . . Methods and materials . , . . . . . . . . Lateral-line system , . . . . . . . . . . . The bones of the head . , . . . . . . . . . . Pectoral girdle and fins . . . . . . Scales . . , . . , . . . . Discussion and conclusions , . . . . . . . . . References , , . . . . . . . . . . . . List of abbreviations used on figures . . , . . . . .
. . . . . . . . . .
. .
. .
. .
. .
. .
. .
. .
. .
. .
Page . . . . . . 469 . . . . , . 470 . . . . . . 471 . . . . . . 475 . . . . . . 497 . . . . . . 499 . . . . . . 499 . . . . . . 503 . . . . . . 505
Introduction
One of the problems of comparative study is to acquire an understanding of the details of structure of the organisms compared; this requires the transformation of descriptive material and two-dimensional illustrations into a three-dimensional mental image. Because of terminological problems, differences of attention, etc., one finds that no account fully
469 0022-5460/84/120469+ 39 $03.00/0 0 1984 The Zoological Society of London

470 M. JOLLIE
covers the areas with which one is concerned, nor removes all of the ambiguities. Changes in identification of bone elements increases the difficulties of understanding. The concern here is, primarily, with the origin and development of the various bony units of the head and pectoral girdle of fishes, with the end in view of a comparative account which will make more fully comprehensible the “homologies” involved. Homologies, or “proper” names for bones, have not been argued in this paper simply because the time is not yet ripe for this. However, the rationale for the system that the present author used in 1962 will eventually be defended. Commonly employed names for bones have been used throughout, but the present author has indicated in parenthesis (and Table I) his personal beliefs (Jollie, 1962, I 98 1 ). [The various papers by Allis, ( 1900, 19 19, 1922) indicate something of the evolution of bone names-as does Starks, 1901, and Harrington, 1955.1
Polypterus is central to arguments about actinopterygian fish phylogeny, so it is important that its head skeleton be reviewed and the relationships and development of the various bones clarified. Actually, there are two genera involved in this group of fishes: Po/vpterus and Erpetoichthys (Culurnoichthys-see Swinney & Heppell, 1982). Holmgren ( 1928) stated that Erpetoichthys in “all” details of the cranial osteology is like Po/jpterus and the present author agrees with the assessment with the exception of the lack of a subopercle. An early account of the bony head skeleton of Pol-vpterus is that of Agassiz (1833-1844, Poi.s.soiis Fossiles, (2): 38). J. Muller (1843, Archiv Anut. 1843: 239) expanded the coverage and in a later account (1844, Abh. K . Akud. Wiss. 1844 117-216) supplied an excellent plate. Dumeril’s (1870, Histoire Nattrrelle de Poissuns, 2: 374) can also be mentioned here. Ayres (1850) described a new species of Polypterus, but the first full account of head structure was that of Traquair (1871). Allis (1922) prepared a lengthy account which was beautifully illustrated. Other accounts dealing with details of adult anatomy included those of Sewertzoff (1925), Gregory (1933), Jarvik ( 1942, 1944, 1947, 1954), Daget ( 1950), Devillers (1958), and Patterson (1975). Erpetoichthys has not really been studied but has been commented on by Holmgren (1928) and Daget (1958).
Developmental studies (often with heavy emphasis on canal bones) began with an account by Budgett (1902). Kerr (1907) described the chondrocrania of Budgett‘s young stages as did Moy-Thomas (1933). de Beer (1937) and Pehrson (1922, 1947, 1958) reviewed the develop- ment of both the chondrocranium and dermal skeleton. Individual stages of development were described by Pollard (1892), Lehn (1918), Holmgren (1943), and Bertmar (1959). The development of the bones of the lower jaw was described by Schmiih (1934). Daget et d., (1964) and Daget & Arnault (1965) reported on raising Poljqpterus senegulus in the laboratory. Apparently, this experiment did not make available specimens for developmental studies.
Methods and materials To date, the following materials of Polypterus have been described by others in various degrees of
completeness, the number of specimens is shown in parentheses after the size (in m m total length): 6.75 (Budgett’s stage 32), 8,9.3 (2), 9.6,24,27,29,30, 32,35,37, 71,75, 76 ( 2 ) , 80,90, 125. and adult. These specimens belong to 2 species (P. senegalus and P. bichir). Most of these are in the Budgett collection or in the collection of Stockholm University.
In addition to a summary of the comments in the literature, 2 (subadult) specimens (of P . ornutipinnis) about 300 mm in length were studied in detail. One was a prepared skeleton purchased from General Biological (now Macmillan Science) and the other a preserved specimen cleared and

S K E L E T A L D E V E L O P M E N T O F POI. Y P T E R I ; S 47 I
0cc .p l
s l n. p a r a s p h e n o i d
c e r a t o b r . I
FIG. I . 8 mm stage showing sensory canal maculae (black), pit-line organs (crosshatched), endocranial cartilage (stippled) and dermal bone anlagen (white). Modified from Pehrson (1958. figs 3 , 5, b) and Bertmar (1959. fig. 79). Bertmar shows the anterior ends of the maxilla and palatoquadrate joined. (Explanation of abbreviations see p. 505).
stained with alizarin. Both were more or less totally dissected. Six osteological specimens or PoIj3ptcrir.s (senegalus, pulmus, hichir) were examined at the British Museum along with 1 prepared, but incomplete, specimen of Erpetoichthys. One preserved 300 mm specimen of Erpetoichthjls was cleared and stained with alizarin. Only I young specimen of Po/jptrrus sotiegcilus, 71 mm in length, was available for study (gift of the University of Stockholm courtesy of Ragnar Olson). This was cleared and stained with alizarin. Comparisons were made with materials of Acipenser, Amiu, Lepisosteus. salmons, and Esos. These are reported on in detail in other papers of this series.
Lateral-line system
The lateral-line system has been described a number of times beginning with Traquair’s (1871) account. An early account is that of Collinge (1893) and a more detailed study is that of Allis (1900, 1922). Jarvik (1947) added many details to our knowledge of the pit-lines, while Pehrsori (1947, 1958) gave developmental facts and Pehrson (1947) discussed, at length, details of the snout canals (as did Jollie, 1969). The earliest stage is a 6.75 mm embryo

412 M. JOLLIE
(Pehrson, 1958, fig. 1) in which the lateral-line organs are already well differentiated as surface placodes, only those of the anterior ends of the supraorbital, infraorbital and mandibular lines have yet to separate. The occipital canal and posterior pit-line are still at an undivided placode stage. The 8mm stage (Fig. I ) is much like the earlier stage. The available information on this system is summarized in Figs 1 to 6.
There are several points of special interest. Jarvik described a nasal pit-line (Fig. 3), more or less paralleling the anterior terminus of the supraorbital canal; a pit-line not seen in other fishes. In terms of the oral pit-line (horizontal and vertical portions and quadratojugal portion of the latter), the horizontal pit-line (vertical and horizontal sections), anterior pit-line, middle pit-line and posterior pit-line, this fish is much like other actinopterygians- recognizing some reduction in the pit-lines on top of the head (Allis, 1922; Gerard, 1937; Holmgren, 1942; Stensio, 1947; Jarvik, 1947) and keeping in mind that there is much variation in the horizontal and oral pit-lines. In terms of canals, there are a few points that should be made here. There is no evidence of a postnasal branch of the infraorbital canal. The preopercular canal does not connect dorsally with the temporal portion of the main lateral-line, being separated in part by the spiracle. The connection between the preopercular and mandibular canal is the last part to form (closed by 37 mm). The supraorbital canal connects at the posterior end of the dermosphenotic with the temporal canal, as in many actinopterygians, but not as in other fishes where the dermosphenotic is incorporated into the roof and the connection is at the anterior end. The infraorbital canal is of interest only in that the third pore, from the tip of the snout (there is no midline pore), lies on the ventrolateral aspect of the lip (a marginal position, Fig. 4). The main lateral-line on the

S K E L E T A L D E V E L O P M E N T O F P O L Y P T E R U S 473
FIG. 3. 24 mm stage showing the canal-related bones of the head and jaws. The sensory maculae are shown as open circles in the canal bones, pit-line organs are smaller circles joined by lines. From Pehrson (1947. fig. 16). (Explanation of abbreviations seep. 505).
e s c
FIG. 4. Lateral view of the head of a specimen of Po!)p/rru.~ ~ J r ~ i t l / i p h i k indicating exposure of dermal bones and the pores of the lateral-line system. (Explanation of abbreviations see p. 505).
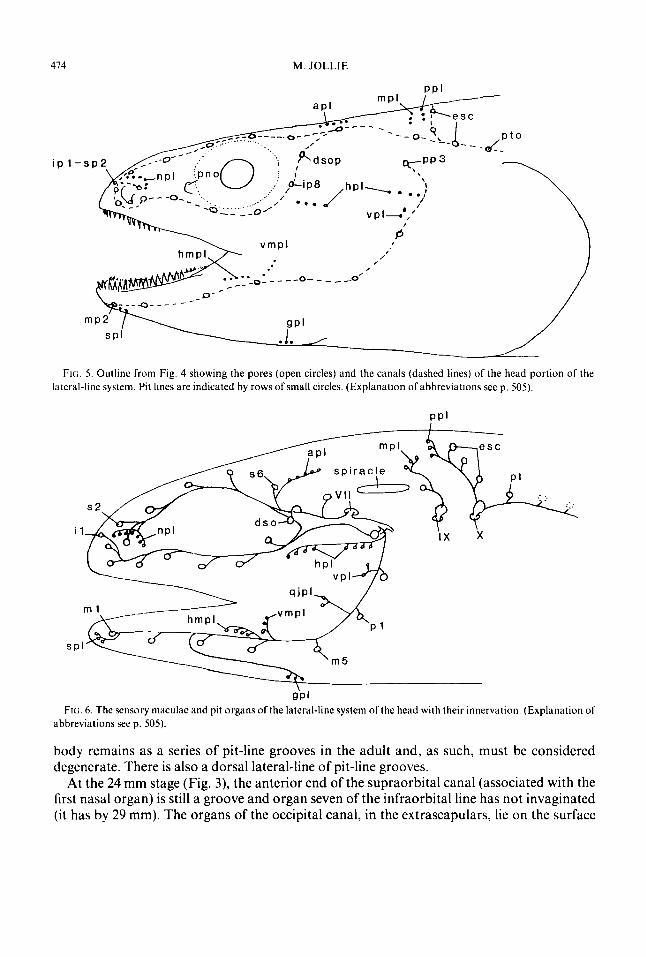
474 M. JOLLIE
i p l -
FIG. 5. Outline from Fig. 4 showing the pores (open circles) and the canals (dashed lines) of the head portion of the lateral-line system. Pit lines are indicated by rows of small circles. (Explanation ofabbreviations seep. 505) .
P P ’
QP I FIG. 6 . The sensory maculae and pit organs of the lateral-line system of the head with their innervation. (Explanation of
abbreviations seep. 505) .
body remains as a series of pit-line grooves in the adult and, as such, must be considered degenerate. There is also a dorsal lateral-line of pit-line grooves.
At the 24 mm stage (Fig. 3), the anterior end of the supraorbital canal (associated with the first nasal organ) is still a groove and organ seven of the infraorbital line has not invaginated (it has by 29 mm). The organs of the occipital canal, in the extrascapulars, lie on the surface

S K E L E T A L D E V E L O P M E N T OF P O L Y P T E R U S 415
at 24 mm. These areas have become tubes by 32 mm (also canal of posttemporal). On the basis of the pattern of development, Polypterus does not appear to differ from other actinopterygians, but does retain a midline pore in the extrascapular series.
The number of organs in the various canals in Polypterus appears to be nearly constant. Some canal variations have been reported by Jarvik (1947) but the present author observed none. Erpetoichthys has the same number of pores and the same pattern of canals. The pattern of innervation of these organs in Polypterus has been figured by Pehrson (1947, fig. 7); this is shown in Fig. 6. The pores of the canals are simple, not branched as in Amiu, Lepisosteus and some teleosts (e.g. Elops) (Fig. 4). The pores in some cases do not open at the suture between bones, but in one or the other of adjacent elements. Bone growth in this genus is like that in Amia or Lepisosteus in that the bony canal tubes first form superficially to the basal plates; later the canal comes to lie in the plate and ends up lying deeper in the bone and even bulging from the inner side of the bone. The relationship between bones and canals (and organs) is shown in Figs 3, 7-1 1.
The bones of the head Bones qf injraorhital line
Medial rostral-internasal. This is a midline dermal (Figs 7,9) bone that bears the connection between the infraorbital canals. At first, it is tubular and contains one organ on either side. At 24mm (Fig. 3) there is a bony lamella and no evidence of a bilateral origin. It is fully formed by 71 mm, extending back along the midline below and between the nasals. The internasal portion, when exposed, may show signs of partial separation or may be completely separated into (two) separate plates (Jarvik, 1947, fig. 6A). Intermediate and luteral rostral. The next three elements of the infraorbital canal (Figs 7-9) are fused in the adult to the premaxilla and are identified by Pehrson (1947: 431432) as the lateral rostral (most anterior) and the first and second antorbitals. In the system of terms used here, lateral rostral is the posterior element, the other two are intermediates. At 24 mm, Pehrson’s “lateral rostral” (first intermediate) is a tube which is not yet attached to the premaxilla (it is attached at 27 mm), while his “first antorbital” (second intermediate) is a gutter-shaped bone which is probably attached to the premaxilla. The “second antorbital” (the present author’s lateral rostral) is already a tube at this stage; i t has a medially directed lamina and is attached to the premaxilla at a couple of points (by this lamina?).
Pehrson pointed out (1947: 432) that the posterior antorbital has a pronounced dorsal process. The present author believes that this dorsal process is, in fact, comparable to the lateral rostral of Amia or Salmo (antorbital of Patterson, 1973). This homology is based on the observation that the premaxillary teeth are limited to a region of the intermediate rostrals and the fact that the palatine process of the premaxilla is limited to this same region, thus leaving the posterior rostral to form an upward and posteriorly directed process. Further, the anterior end of the maxilla reaches below the posterior intermediate rostral, thus forming a premaxillary-maxillary joint (Fig. 3).
By the present author’s definition, rostrals are limited to canal tubes which, in this case, fuse with each other and (anteriorly) the underlying premaxilla. The strong development of the preorbital process of the lateral rostral as a non-canal structure is important (it is without any canal vestige in Salmo). This ethmoid process in the mature fish (Figs 7, 8) covers the lateral ethmoid externally. The equivalence of Pehrson’s antorbital and the lateral rostral

476 M . JOLLIE
FIG. 7. (a) Anterokiterdl and a bit dorsal view of the snout of the cranium of Polypprerus showing bones, pores and canals (dashed lines). (b) Anterior ends of the supraorbital and infrdorbital canals showing pores (closed circles) and maculae (stippled). The two adjacent bone pores, upper left, actually share a common opening through the skin (see Figs 4. 5). (Explanation of abbreviations seep. 505).
bone seen in Amia (Fig. 8(a)) is probable, but as a post-nasal branch of the infraorbital canal does not occur; there is no way to establish a homology except by position. The lack of such a canal as an identification mark of the lateral rostral, and the attachment of canal bones to the premaxilla, are peculiarities of Pofypterus. Attachment of canal elements to the premaxilla is seen in some palaeoniscoid actinopterygians. Lacrimal. A triangular canal element, seen at 24mm, which margins the eye antero- ventrally and contains a single organ, can be assumed, on the basis of its position, to represent the lacrimal of the actinopterygian. This bone has a distinct preorbital process which suggests the lacrimal in terms of shape (Figs 7,8). Infraorbital. There are two infraorbital elements. The more posterior of these is just beginning to ossify (grooved disc) at the 24 mm stage (Pehrson, 1947: 43, fig. 16), while the anterior one is a tube which has not yet fused to the maxilla (Fig. 3). The posterior of these is probably roughly equivalent to ajugal.) At 27 mm, both are fused to the maxilla (Figs 7,8). Postorbitaf. There is a single postorbital element (Figs 7, 8), with one organ, which becomes fairly large in the adult fish. It is nearly tubular at 24 mm (Fig. 3). Dermosphenotic (intertemporal). This element arises as a canal element and at 24 mm is already fused with the perichondral beginning of the sphenotic. The organ of this bone is innervated by a special nerve from the root of the facialis, not by a branch of the otic nerve. In older fishes it is largely covered by the parietal and is exposed only as a restricted area surrounding the canal pore (Figs 7-8, 14). The sphenotic is also slightly exposed where the two bones fuse.

SKELETAL DEVELOPMENT O F P O L Y P T E R U S
esc ( a )
frontal, d,sr
nasal \
m. ros.-
.
417
lat . -2 qular a r t
( b )
supracleithrum
FIG. 8. (a) Lateral view of the head skeleton of a specimen of Polyprrrus. Lateral-line pores in black. (b) Same with cheek plates removed. (Explanation of abbreviations see p. 505).
Bones of supraorbital and temporal canals
Nasal. According to Pehrson (1947:426-428), at 24 mm the anterior nasal (0s terminale) is just a splint, while the second and third nasals are perforated tubes, the last with a small basal plate. At 27 mm, the antero-lateral unit is a tube and remains as such in the adult (less extensive in Erpetoichthys). In the 37 mm specimen, the medial flange of the posterior nasal is better developed, and by 45 mm the basic shape of the bone is achieved. The basal plate of the second nasal is well developed in the present author’s 71 mm specimen.

478 M. JOLLIE
Polypterus is unique among actinopterygians in having three separate nasals (Figs 7-9) resembling some crossopterygians (rhipidistians) which have an irregular number of three or more. Two of the nasals of Polypterus have distinct bony bases extending laterally, while the terminal one is a tube bone. It is quite possible that this threesome represents a fragmentation product of such a bone, as seen in Amiu (where there are three neuromasts) or in Lepisosteus (where there are even more neuromasts). In terms of the nasal, Po!,.ptrrus is more modified, in the present author’s view, than other actinopterygians where this trend of separation of the nasal into its component parts is seen but not yet achieved. Pehrson (1947: 428) has stressed the similarity in the hooking back of the supraorbital canal of the nasals of Amia and Lepisosteus (also Jollie, 1969). Frontal (parietal). The frontal (Figs 3, 7-9) is associated with three neuromasts each of which has its own primordium. That of the anterior one is separate at 24 mm (Pehrson, 1947: 430), whereas the two more posterior units can be partly distinguished and each has its own medial lamella. It can be assumed that the development of this bone begins as three canal tubes and each produces a basal lamina. The two posterior ones join first and later the anterior element joins in. Development is much like that seen in Amia, including the outward (toward the epidermis) growth of the bone lamina, so that the tube ends up deeply embedded in the mature bone. Jarvik (1947, fig. 6; 1967, fig. 6) showed a frontal divided into anterior and posterior parts and used this as evidence for anterior and posterior parts of this bone. Actually, all that is shown is that a normal bone, arising historically from several parts, may on (rare) occasions be subdivided into a few, or even many parts. Parietal (postpurietal). The parietal (Fig. 3) is evident as a blastema (membrane-like, Pehrson, 1947: 439) at 24 mm, but is soon obliterated (after 37 mm) by the meeting at the midline of the basal lamina of the intertemporosupratemporal, and (Pehrson. 1947: 440) “. . . a parietal sensu strictu does not exist in this fish, not even as a component of a complex bone.” Patterson (1977: 92-96) has discussed “fusion and loss” in some detail, but without making the problem of decision easier. “In general, the hypothesis of loss and invasion is very difficult to refute, for it has few, if any, testable consequences.” Pehrson (1947: 440) assumed an inductive relationship between the parietal and the middle pit-line (as in Acipenser), but this view is here rejected, along with much of the discussion of the “close” relationship between sensory organs of the lateral-line system and bones. There is far too much variation for such a simplistic solution. A parietal is not induced by, nor does it induce, such organs. Intertemporosupratemporal (supratemporotabulur). Two canal elements and their basal lamellae are involved in this bone (Fig. 3, 7-1 1). These fuse, as in other actinopterygians, to produce the definitive bone. The bone shows its double origin at the 24 mm stage (Pehrson, 1947: 438439, fig. 16). This evidence of double origin is lost at 27 mm. This compound bone spreads to the midline, replacing the weak parietal which now disappears by resorption. The expansion of this element (or just the anterior, intertemporal, part of it) to the midline is seen in haplolepids (and other palaeoniscoids), porolepid crossopterygians, and coelacanths. The idea that this bone is fused with the parietal, or is the parietal, (Allis, 1922; Holmgren & Stensio, 1936; Parrington, 1949, 1956; Jardine, 1969) is not supported by the developmental facts.
This bone, as in other actinopterygians, has a flange passing down over the neurocranium, medial to the spiracular duct and associated with the articular area of the hyomandibula. This pterotic process is identifiable with that of other actinopterygians (Jollie, 1975: 70, 77)

oh
FIG. 9. Dorsal view or specimen with squamosoids, cheek plates and operculum removed on right side. (Explanation of abbreviations seep. 505).

480 M . JOLLIE
FIG. 10. Dorsolateral view of osteocranium. Extrascapulars and posttemporal removed on right side. Arrow is through the ventral canal of the parasphenoid. (Explanation of abbreviations see p. 505).
but is only slightly developed, as in Amia or Lepisosteus. What is more interesting is a thick flange of bone that grows down over the posterior aspect of the endocranium in the region of the epiotic of other fishes. This epiotic process (Fig. 10) sutures closely with the pterotic- epiotic and occipital, but shows no evidence of penetration of the underlying cartilage. This epiotic-like process also has a strong process which extends posteriorly below the lateral medial (intermediate) extrascapular and the posttemporal (probably serving for the attach- ment of nape muscles). The present author prefers to identify the flanges of this bone as dermal (not membrane as suggested by Patterson, 1977), and retain the synonymy of dermal and membrane. He believes it is equally confusing to talk about membrane extensions of chondral bones (never preformed in cartilage) such as the extensions of vertebrae (which may never, in phylogeny, have been preformed in cartilage) as it is to refer to them as dermal.
One might wonder if the intertemporosupratemporal should not be identified as the intertemporosupratemporoepiotic. There is no evidence of a chondral epiotic centre here and thus no support for the assumption that the epiotic has become part of a dermal bone.
Bones of occipital canal
Extrascapulars. There are three pairs of these bones (Figs 3, 9-11), the lateral extra- scapular is displaced somewhat posteriorly (as described by Allis, 1922). There are also intermediate and medial pairs of extrascapulars. The intermediate pair of extrascapulars has appeared at 27mm and by 29mm they are tubular; the medial pair has appeared and

S K E L E T A L D E V E L O P M E N T O F P O L Y P T E R U S 48 I
become tubular at 37 mm. The lateral extrascapulars are apparent at 29 mm at which time invagination is just beginning; i t becomes a guttered bone at 32 mm and a cylinder at 37 mm. Pehrson (1947: 443) viewed the anlagen of this bone (his “suprascapular”) as binary. The presence of a midline pore in this series has already been remarked upon, while the pattern of origin of units resembles that seen in Amiu.
Jarvik (1947, 1967: 192, fig. 6B) figures a specimen of P. senegulus in which the medial extrascapular of one side is missing, and the canal has been transferred to the expanded posterior margin of the intertemporosupratemporal. The present author observed a specimen of P. hichir (B.M. 84.6.9.11) in which an anamestic was inserted between the posterior margins of the lateral and intermediate extrascapulars. Ayres (1850) described a specimen of P. pulmas with the two medial extrascapulars fused across the midline.
Bones yf’cheek und operculum
Preopercle. The fully developed preopercle (Figs 3, 7, 8(a)) is peculiar in that it is a superficial plate covering the entire cheek (in living actinopterygians the cheek is largely without a bony cover or is scaled). Anteriorly, this plate has an overlapping suture with the maxilla. A ventroanterior prong passes over the medial surface of the maxilla while a prong of the maxilla, lying just dorsal to the latter, laps inside the preopercle. Whereas the preopercle of actinopterygians (except Acipenser which lacks an organized preopercle) has a flange of bone extending antero-medially from the line of the canal, from which muscle fibres of the jaw take origin, this is lacking here. Rather, there is a thickened region lying along the course of the sensory canal, below the level of the opercular process of the hyomandibula (to which the hyomandibula is tied by connective tissue). The lack of a ridge dorsally, in the region of the most dorsal sensory organ, is of interest.
The preopercle has three sensory organs in i t . The anterior cheek process of the preopercle is the first part to develop (Pehrson, 1958, fig. 8); at 8 mm (Fig. 1 ) a small ossicle lies close to the posterior end of the maxilla. From here it extends posteriorly until, at 9.6mm, it is associated with the two dorsal preopercular organs or projects between them. Pehrson (1958: 251) has identified this anterior part as a supramaxilla, but this view is rejected here on the grounds that functional incorporation of such a bone into the preopercle seems most unlikely, and that this splint, whatever its origin, gradually expands posteriorly to form the canal part of the (real) preopercle. (Also, the supramaxilla is a bone of higher actinopts.) At 24 mm the preopercle is well developed, but the ventral preopercular organ is associated with a separate (binary) ossification centre. At 27mm this centre is a tube which at 29mm becomes attached to the rest of the preopercle. Pehrson’s (1958: 252) view was that this bone arises from four centres, three laterosensory centres connected with neuromasts and the “supramaxilla”; it is interpreted here as arising basically as a coordinated unit, even though one centre, associated with the ventral neuromast, appears separately.
There is little question that the preopercle of this group includes more than that of other living actinopterygians (or extends into regions not occupied by that bone). I t must include those parts separated as “suborbitals” (Gardiner’s 1960 terminology) in palaeoniscoids and many fossil forms. The suggestion (Siive-Soderbergh, 1933; Holmgren & Stensio, 1936: 395-396; Pehrson, 1947) that the preopercle includes a squamosal component seems reasonable if one equates the squamosal-preopercle of some crossopterygians (certainly osteolepiforms and porolepiforms) with the actinopterygian preopercle. The use of a

482 M. JOLLIE
compound name is rejected on the grounds that each of the cheek systems seen in osteichthyans seems to have been independently achieved, and thus are only partially homologous (in terms of developmental controls-Jollie, 198 1). Thus the interpretation suggested here is that the preopercle of Polypterus is unique in form and origin within actinopterygians. It is, in effect, another argument for the recognition of the isolation of this group.
A quadratojugal is lacking in Polypterus. There is a fleshy gap in the ventral margin of the cheek between the tooth-bearing maxilla and the ventral end of the preopercle bearing the canal. The anterior half of this gap is filled with an ovoid plate. Behind this is often a smaller ovoid plate (or two). The anterior plate is seen (by others) as part of the maxilla, while the posterior one(s) are described as the quadratojugal. The late appearance of the anterior plate-present at 71 mm-suggests it is not a part of the maxilla. The even later, or non-appearance, of the posterior plate(s), eliminates effectively the quadratojugal as an explanation. The more posterior plate is associated with the quadratojugal pit-line of Jarvik (1947). Nothing is to be gained by debating the accuracy of this identification, the best that can be said is that it might be true. As the present author’s stained specimen of a 300 mm Erpetoichthys lacked these plates, it is preferred to identify them as secondary. One, however, is surprised that this group of fishes, assumed to be primitive, should lack an identifiable quadratojugal. Accessory plutes (squamosoids). Polypterus (Figs 7, 9, 1 1) is unique among actinoptery- gians in that an irregular line (and number) of accessory plates overlies the “joint” between roof, cheek and operculum. This series also extends down behind the postorbital and lateral to the extrascapular. The slit-like spiracle opens between two rather consistently larger plates and the intertemporosupratemporal. Traquair (1871: 180) spoke of their valve function. These plates are of interest as a superficial (secondary) layer of elements overlying the deeper original bones. They thus correspond to the scales on the cheek of the perch or lungfish and, as such, are not really anamestics but squamosoids (i.e. scale-related plates like those on the cheek of Lepisosteus). It seems probable that these plates were not yet started at 37mm (Pehrson’s account) but they are well developed in the present author’s 71 mm specimen. Dermohyul. A triangular piece of bone is associated with the posterodorsal angle of the head of the hyomandibula (Figs 8(b), 9, 11,14). It is already present at 24 mm but expands in relative size and, in the adult, is closely sutured with the hyomandibula. This bone is exposed (and attached) in the cranial roof at the posterior end of the spiracle.
Holmgren (1943: 95, fig. 45) and Pehrson (1947: 406, fig. 6) speak of the “perichondral nature” of part of this bone, but also imply that it extends extrachondrally (dermally) outward and upward. The assumption that this bone arose by fusion of tooth plates in the wall of the spiracle (Nielsen, 1936) is possibly valid, but rules out a perichondral origin. Comparisons with the dermohyal of palaeoniscoids (term introduced by Nielsen, 1942) are not of the best. Patterson (1973: 245) remarked that the dermohyal of palaeoniscoids is “applied” to the hyomandibula (see also Gardiner, 1973). It lies posteriorly to the incipient connection between the preopercular canal and the temporal part of the lateral-line canal. (This connection is lacking in Polypterus-because of the large size of the spiracle?) In the present author’s view, it has no apparent, fixed relationship to the spiracle; lying behind that opening and somewhat lateral to it in many of the palaeoniscoids (Pteronisculus). Opercle. The opercle (Figs 4, 8(a), 12) is a large plate which is joined to the preopercle along a slightly irregular line. The preopercle overlaps a curved process of the opercle which

S K E L E T A L D E V E L O P M E N T O F P O L Y P T E R U S 483
FIG. 1 1 . Dorsal view of posterior end of the osteocranium. Medial. intermediate and lateral extrascapulars (right side) have been removed and the processes of the posttemporal are shown dashed (on right). Left side undisturbed (inter- mediate extrascapular removed to show anteromedial process of posttemporal). Sensory canal dashed on right; pores, black circles. (Explanation of abbreviations see p. 505).
lies anterodorsally to the facet for articulation with the hyomandibula. Daget e l al., (1964: fig. 19) reported a small opercular cartilage. Patterson’s (1977: 90) discussion suggests a cartilaginous origin for the opercular series, whereas the present author is inclined to see this series as of dermal origin and the appearance of cartilage as secondary. Patterson (1977: 91) discussed “adventitious” cartilage but did not apply the idea here. Subopercle. This is a small triangular plate (Figs 6, 8(a)) wedged between the opercle and preopercle ventrally. It is slightly overlapped by the preopercle and has a broad area of contact with that bone. suggesting rather limited movement of the whole branchial chamber cover. Pehrson (1947, 1958) does not comment on the time of appearance of the opercle and subopercle but they are certainly present at 24 mm. A subopercle is lacking in Erpetoichthys. Lateral gular. A pair of large lateral gulars (Fig. 8(a)) cover the throat region from near the mandibular symphysis to the posterior end of the mandible. Pehrson (1958: 254) was of the opinion that the gular, as suggested by Berg (1940), was of branchiostegal origin. His fig. 12 of this bone in the 24 mm specimen does not support this contention, nor does his account of younger stages. Like so many dermal bones (such as the gular in Amirc), this one arises as a fenestrated plate. There is no indication of its forming by the fusion of branchiostegal

Fici. 12. Ventral view of head skeleton, lower jaw removed. Teeth of premaxilla and anterior end o f maxilla of right side removed. Sacculolith seen in vestibular fontanelle of left side. Baudelot’s ligament of right side cut off. (Explanation of abbreviations see p. 505).

S K E L E T A L D E V E L O P M E N T OF POL Y P T E R U S 485
rudiments, which would run roughly across the gular rather than anteroposteriorly as the fibres of this bone do. There is little variation in these plates. Jarvik (1947, fig. 6D) showed a separate posterior portion on one side of a P. senegulus while the present author’s specimen of Erpetoichthys has both gulars weakly ossified transversely, anterior to their middles. This gular is tied by stout ligaments (posteriorly and midway laterally) to the ceratohyals.
Jarvik (1947, fig. 1; also Lehman, 1966, fig. 2) illustrated a branchiostegal ray between the margin of the lateral gular and the mandible about midway along the length of the gular. Jarvik (1947: footnote p. 74) stated “In one specimen of Po1.vptrru.F (SMPz, No. 3) stained in alizarin, there is a thin film of dermal bone (Rbrv, fig. lC), which is so situated that it very likely represents a vestige of the branchiostegal rays.” I t should be pointed out that Jarvik saw Polypterus as more like a crossopterygian than an actinopterygian, and this detail would support such a view. (The branchiostegal rays of actinopts and crossopts are quite different in appearance.) The present author has not seen such a bone, nor have others reported it, nor does he believe it could be identified, if present, as anything except an anamestic.
Bones of the mouth Dernzuls qfupper.juw Premuxillu-rostrul. The premaxilla is present at 9.6 mm (Fig. 2) with two teeth attached (Pehrson, 1958, fig. 8). The development of this bone (Figs 3,7-9, 12) is like that of Amiu and Lepisosteus. It spreads out over the palatal aspect of the cartilage of the ethmoid region to form distinct processes figured by Jarvik (1942, fig. 5B). The palatal process is overlapped (viewed ventrally) by a process of the maxilla. Dorsally, the premaxilla extends upward to suture with the medial rostral; i t fuses with three rostral canal elements which extend below the nasal cavity. The premaxilla (plus rostrals) of Pol-vpterus differs from that in Amiu and Lepisosteus (and many teleost fishes-Sulnio) in that no lamina extends upward over the backwall of the nasal capsule.
The mechanics of growth seem to be different here as compared with the “holosteans” and the end result is certainly unique to this group. The suggestion by Westoll (1936; Moy-Thomas, 1934) that the premaxillary teeth, which arise separately, have transferred their attachment to the lateral rostrals is rejected on the grounds of the general form and the significant palatal extent of this bone. Among palaeoniscoids, there is the suggestion of such “transference” of teeth and, in the case of the lacrimals of Lepisosteus, it is assumed (Jollie, 1984) that teeth arise in the tissue overlying these bones, but in a sense are not transferred from the maxilla. Rather, it is a case of teeth being produced by jaw margin tissue and attaching to the most convenient base. In Polypterus a premaxilla is present, and develops in a way comparable to many other fishes. To this tooth-bearing bone. rostral canal tubes fuse. The premaxilla is, therefore, identified as the premaxilla-rostral, since there is no doubt of its compound origin in this group (and similarly in rhipidistian crossopts?); to do so, however, is confusing, so that thinking of the premaxilla separately from its attached rostrals is not out of place. The present author has described these rostrals separately. Maxillu-infruorbitul. The maxilla (Fig. l), as a thin splint without teeth, has appeared by 8 mm (Pehrson, 1958, fig. 5); i t is long and curved with its posterior end up behind the eye. Tips of teeth are probably present in the overlying tissue but not reported. By 9.6 mm (Fig. 2) this bone has several (4) teeth attached. It is important to note that the separate

486 M . JOLLIE
preopercular plate, often identified as part of the maxilla, has not yet appeared (Pehrson, 1947: 436).
The maxilla (Figs 3, 7-9, 12) develops a small palatal process anteriorly, which indicates its fairly rigid incorporation into the jaw margin. The maxilla lacks the characteristic medial articular process seen in Amia and Salmo and suggested early in the development of Lepisosreus. This bone also fuses to two canal elements, a condition not seen in palaeoniscoids or other actinopterygians. This fusion warrants recognition by the use of the name maxilla-infraorbital but again confusion results. It is perhaps best to speak of the two elements separately-as has the present author. It is noteworthy that the maxilla does not lie in the labial fold, as in other actinopts (Allis, 1919).
In the ventral cheek, behind the tooth row, usually two separate ovoid plates of bone develop (see above). The anterior plate is just beginning to form in the present author’s 71 mm specimen, at a time when the head skeleton is essentially fully laid out. This suggests that it is not a part of the maxilla (or a supramaxilla) but actually a secondary scale-related plate which the present author would prefer to call a squamosoid. Comparison of the anterior part of the preopercle, by Pehrson, with the supramaxilla is less likely than comparison of the dorsoposterior process of the maxilla with that bone.
Pehrson (1958: 248-249) discussed at length his views of the homology of the maxilla. The present author rejects the idea that any part of the maxilla is incorporated into the preopercle, and rejects the identification of the free superficial plates, just behind the toothed margin of the maxilla, as part of it. Comparisons with Lehman’s (1966) account of a speci- men of Prrronisculus broughi and Pehrson’s fig. 9 do not shed light on this problem. It is quite true that the area in question is occupied by the maxilla in the palaeoniscoid but once that bone withdraws (fails to develop) it can be replaced by squamosoids, just as deep-lying bones can become overlain with scales. (Erpetoichfhys lacks these cheek plates.) It is evident that the condition of the maxilla, both in terms of shape and association with canal bones, is unique, The maxilla cannot be compared with the same bone of the palaeoniscoid but must be viewed as much modified [Allis’s (1900) homologizing of the maxilla of Polypferus with the “dermopalatine” of Amia or the teleost is a fine example of confusion of interpretation.] Palatine. The palatine, (dermopalatine-Pehrson, 1958, fig. lo), with attached teeth, is present at 9.6 mm (Fig. 12). Pehrson, 1958 (fig. 6) figures a palatine in the 8 mm stage, but the position of that bone and its relationship to the palatoquadrate cartilage suggests that it may be the ectopterygoid. In this genus, the palatine (Fig. 12) extends forward nearly to the midline, close behind the premaxillary teeth and overlapped (dorsally) by the palatine process of the premaxilla. In Amia the palatine lies below the ossified anterior end of the palatoquadrate cartilage (autopalatine) where that cartilage connects with the lateral ethmoid area. Lepisosteus is different from most actinopterygian fishes in that the palatoquadrate extends far forward in relation to the lateral ethmoid region (just anterior to the eye) as a result of the development of the snout. A similar extension may have been involved in Poiypterus but it is not evident in the 76 mm specimen (Pehrson, 1947, fig. 4), in which the autopalatine bone lies posterior to the posterior end of the palatine, which in turn lies below the palatine processes of the premaxilla and maxilla, a unique situation. Thus the palatine is not closely related to this cartilaginous process, and is, probably, displaced. Ecropterygoid. The ectopterygoid has appeared, with teeth, at 9.6 mm and may be present at 8 mm (see comment above under palatine). It becomes sutured (Fig. 12) to the palatine anteriorly, the pterygoid medially, and the metapterygoid and quadrate posteriorly. The

S K E L E T A L D E V E L O P M E N T OF P O L Y P T E R U S
palatal surface of this bone is covered with a shagreen of small teeth. The anterior end of this bone is associated with the autopalatine. Pterygoid. The pterygoid (endopterygoid of some) arises sometime after the 9.6 mm stage. It forms a large lamina of bone covered by a shagreen of small teeth on its palatal surface (Fig. 12). The palatoquadrate cartilage lies on its dorsal surface. Vomer. A midline vomer is present at 9.6 mm (Pehrson, 1958, fig. 10). Teeth appear very late, at about 32 mm. In the present author’s 71 mm specimen there were four attached teeth and two tips of teeth; in the larger specimen (Fig. 12) there were nine irregularly arranged teeth (no regular rows suggested by Pehrson). The bone could not be seen in any of the British Museum (Nat. Hist.) specimens but this is due to its small size and close suturing (fusion) with the palatines. It is literally lost between the anterior ends of the palatines. The highly reduced and modified (no evidence of bilateral origin) state of this bone is another peculiarity of this group. Purusphenoid. The parasphenoid is present at 8 mm but as a midline bone, not as paired bilateral plates as shown by Pehrson (1958, fig. 6). By 9.6 mm this bone is well outlined with its ascending processes and bifurcate posterior end.
The form of the adult bone of Poljipterus (Figs 10, 12, 15) was a subject of concern to Jarvik (1954: 60-64), suffice to say that its complexity is not matched in any other group. The peculiar ascending process forks, one branch suturing with the sphenotic (as in Amia) and two parallel posterior processes suturing with the pterotic-epiotic. These two posterior processes form a canal for the lateral head vein. Below this is a second canal (longitudinal canal of Jarvik). In addition to this last, there is a palatal process paralleling, on the palatal surface, the posterior margins of the spiracular cleft (see Jarvik, 1954: fig. 27B). The pharyngobranchial of the first branchial arch attaches to the posterior margin of the tip of this process, and its palatal surface bears a shagreen of teeth. In Amia, this same(?) process is represented only by some tooth plates in the mouth lining.
487
Puluto yuudru te und suspensorium
Autopulutine. A small autopalatine (Fig. 8) lies below the lateral ethmoid. This ossicle supports the anterior end of the ectopterygoid and the posterior angle of the palatine process of the maxilla. In Aniiu or the teleost (Sulmo, Esos), it is associated with the anterior end of the palatine. Van Wijhe (1882: 253) reported it fused to the anterior end of the ectopterygoid but this is definitely not the case. The time of origin is not known but is late as in other fish. Quudrute. The quadrate was first observed in the 71 mm specimen as a perichondral ossification in the area of the mandibular articulation (Pehrson, 1947, a t 76 mm). The form of the quadrate (Figs 8(b), 12) in this group is unique because of a posterior process which hooks ventrally. This process (quadratojugal?) is attached to the end of the hyomandibula, whereas the main posterolateral margin of the quadrate is connected to the anterior margin of the ventral process of the preopercle. Metapterygoid. This bone (Figs 8(b), 12) is present at 24mm (Pehrson, 1947) and is of particular interest since it is entirely dermal (and perichondral?) in this group (also Patterson, 1977: 87). (Apparently its original chondral origin is simply bypassed.) I t covers the palatal surface of the posterior end of the palatoquadrate cartilage and bears a shagreen of teeth on its’ surface. The posterior truncated end is braced to the “hyomandibula” and laterally this end contacts the quadrate. Posteriorly, the dorso-medial margin is wrapped

488 M. JOLLlE
around the cartilage but shows no invasion of i t (similar observations are recorded by Allis, 1922; Pehrson, 1947: 448449).
The condition of this bone is like that seen in palaeoniscoids (e.g. Pteronisculus, Nielsen, 1942). Its position, relationships, and dermal character clearly indicate that it is an intrusion into the toothed palatal area between the pterygoid and the quadrate by what was originally a chondral ossification. In the crossopterygian (including Actinistia), dipnoan or tetrapod there is a separate ossified epipterygoid, but no dermal palatal component. In Amia, the metapterygoid is composed of an upward projecting ossified “epipterygoid” part and a tooth plate between pterygoid and quadrate. In Salmo or Lepisosteus, there is no toothed part, only cartilage bone which projects upward or inward; Acipenser lacks a metapterygoid either as dermal plate or ossified epipterygoid. Polypterus is thus like the palaeoniscoid, probably a derived condition, and quite unlike most other actinopterygians. The metaptery- goid of actinopts thus shows all of the variations of relationship of an original chondral bone which develops a dermal (tooth bearing) extension and then becomes totally dermal.
Dermals of lower jaw
Dentosplenial. The dentary (Figs 1,8, 13) is present at 8 mm and teeth are probably present in the overlying tissue. By 9.6mm about a dozen teeth are attached to the bone but the splenial portion has not appeared (Pehrson, 1958: 253). By 24 mm three canal elements are present, the anterior one not yet attached to the dentary. By 29 mm the anterior splenial is also fused to the dentary and the main shape of the bone is achieved. This bone appears to be entirely comparable to that of other actinopterygians. Angular. This bone is already present in the 8 mm specimen and well developed by 9.6 mm (Fig. 1). In the adult, this is a large plate (Figs 3,8) , which lies on the outside of the mandible just anterior to the articular area, with two canal units attached to it. Pehrson (1958: 254) and others have identified the canal tubes attached to the angular as splenials . If the osteolepid or porolepid crossopterygian lower jaw is to act as the source of our terms, then i t should be pointed out that the angular bears a canal (i.e. has canal elements fused to it) but those canal elements are not “splenials”. Thus a compound name is not called for.
Pehrson (1958, fig. 8) showed a small “supraangular” behind the angular, but this is not
d e n t o s p l e n i a l
FIG. 13. Internal view of right mandibular ramus. Groove of Meckel’s cartilage stippled

S K E L E T A L D E V E L O P M E N T OF POL Y P T E R U S 489
seen in the 24 mm specimen (Fig. 3) nor is i t in the proper position for a supraangular. I t can be concluded that it is a temporarily detached part of the angular bearing the posterior end of the mandibular canal. A supraangular does not occur in this group but is seen in some palaeoniscoids, e.g. Amiu and Lepisosteus. (There is the question of the validity of this name for the bone of these actinopterygians but that is discussed elsewhere in the present author’s series of papers.) Prearticulur. The prearticular is present at 8 mm and has attached teeth by 9.6 mm (Fig. 1). This is a large bone (Fig. 13) extending three-quarters of the length of the ramus and forming part of the internal wall of the mandibular fossa. It has a large “coronoid” process which forms the anterior wall of that fossa; it sutures laterally with the dentosplenial. Anteriorly, this bone tapers to a point underlying the posterior coronoid. Its dorsal margin and inner surface are largely covered with small teeth. The bone anteriorly lies above the groove for Meckel’s cartilage, but posteriorly overlaps that groove and the articular bone.
There is a strong connective tissue connection between the coronoid process of thc prearticular and the angle of the mouth. In this connection is a rectangular plate of cartilage (Fig. 4) which has been identified as a “labial” cartilage (Allis, 1919, pl. 3). There is no evident morphological, or functional, correlation with the labial cartilages of sharks. A more appropriate name for this cartilage is needed.
Pehrson (1947: 445, 450) suggested that the prearticular is just a posterior coronoid. Certainly, this bone does carry the posterior continuation of the “coronoid” teeth but, in addition to this, has an investing function+overing the inner surface of the jaw and thus extending anteriorly below the true coronoids. I t does not extend so far forward in other actinopterygians. In crossopterygians this bone extends clear to the anterior end of the ramus below the coronoids proper. Coronoids. There are two short coronoids (Fig. 13) which occupy the anterior quarter of the mandible; the teeth bend strongly posteriorly. The first coronoid is present at 24 mm and the second at 37 mm. Apparently, the number is constant within the group, at least no more have been seen or reported in the literature. Holmgren & Stensio (1936: 327) observed two small tooth-bearing ossicles at the symphysis of the mandible of Po/j.pterus which are probably the anterior pair of coronoids. Jarvik (1972: 114-115) identified these as parasymphysial dental plates. That these anterior plates are coronoids, comparable to those of Amiu, cannot be doubted; these plates of Pol-vpterus do not compare well with the “sym- physial plates” of Holoprychius (or other fossil forms in which such plates occur). The late appearance of the coronoids, in contrast to Amiu or Lepisosteus, is of interest.
Curtiluge hones of’lo,cer juw
Arficulur. The articular (Fig. 13) is present at 71 mm in Polypfrrus as a separate (perichondral) entity (free of angular). It becomes a massive element with an articular groove and a retroarticular process. The retroarticular process in other actinopterygian fishes tends to ossify separately, perhaps because of its relationship to tendons from the interopercle and the posterior end of the ceratohyal. The retroarticular process of the articular has a dermal exposure between the angular, laterally, and the prearticular, medially. A similar dermal exposure (which can arise as a distinct dermal element) is seen in Lepisosreus or Esos. Berg (1958: 160) speaks of the retroarticular being missing in Poljypterus. In Anziu. the articular ossifies in three parts (four parts if one includes the coronomeckelian), one of which is a

490 M. JOLLIE
retroarticular. Nelson (1973a b) and Patterson (1977: 93-95) have discussed some of the variations of the articular in fossil and living forms and the present author (Jollie, 1975: 77) has considered the replacement of the articular joint area (pars mandibuloarticularis), but not the pars retroarticularis, by invasion from the angular. The separate, one-piece articular of Polypterus is primitive. Lack of ossification in Acipenser or subdivision of the articular into parts is derived.
Daget (1958: 2505) mentioned that the mandibulo-hyoidean ligament is lacking in Polypterus, but this is not the case. The ligament extends from the posterior end of the ceratohyal to the ventrolateral margin of the angular anterior to the quadrate articulation. Mentomrckeliun. A small mentomeckelian is present at 76mm (not seen in the author’s 71 mm specimen). It was described by Allis (1922) from a grown specimen and was quite evident in the present author’s alizarin-stained specimen (300 mm). The two perichondral elements, one on either side, closely approach the symphysis and are quite distinct from the dentosplenial (though firmly grasped by it). There is no coronomeckelian in Polypterus (Nelson, 1973b).
Endocrunium
The bones of the endocranium are fewer in number than in actinopterygians generally. The endocranial ossifications are so different, as compared with other actinopterygians, as to make the identification of the one large otic bone, the “opisthotic” of Patterson, seemingly impossible.
Patterson (1975: 415-473) has a lengthy account of variations of the bones of the brain case of actinopterygian fishes and has hypothesized a number of new views. No account of the endocranium would be complete without reference to his ideas. He has both confused and clarified problems. We shall consider only those clarifications which involved the identification of the so-called opisthotic in Polypterus. From his account, conclusions as to the homologies of bones can only be reached if one is willing to concede the possibility of error when viewing an incomplete series of facts.
Patterson’s chief contribution deals with the origin of the supraoccipital in teleosts and also, importantly, with the homology of bones in the otic region. The primitive condition is suggested by a developing specimen of Perleidus (Patterson, 1975: figs 115-1 16) in which the occipital segment is separated from the endocranium by the cranial fissure (otico-occipital fissure). This is the case for the braincase among palaeoniscoids (Nielsen, 1942,1949; Gardiner & Bartram, 1977) and was an original feature of the actinopterygian as compared with other osteichthyans (Schaeffer, 1968). Patterson has demonstrated that, in pholidophorids, the cranial fissure remains open dorsally and that the occipital segment has produced paired epioccipital bones and a midline supraoccipital element. [The supraoccipital has always been assumed to be of occipital origin so this is not new in itself, but some of his derivative ideas are surprising. Jollie ( 1975: 78) assumed an extrascapular origin for the supraoccipital, but this needs to be retracted in view of Patterson’s account] Patterson showed that, in pholidophorids, there are massive paired “pterotics” in the posterior part of the endocranium that meet at the midline, above the foramen magnum; these form part of the lateral wall of the cranium above an opisthotic. Patterson located the centre of development of the “pterotic” as over the posterior semi-circular canal. He assumed that, in the course of the phylogeny of early teleosts, the cranial fissure closed dorsally and that the supraoccipital

S K E L E T A L D E V E L O P M E N T OF P O L Y P T E R U S 49 I
and epioccipitals were incorporated into the rear wall of the cranium, the latter becoming, at least in part, the epiotic (fused with part of the pterotic?).
Patterson has discussed a number of alternative hypotheses but, in this case, proposed only that the epiotic and epioccipital are associated in the teleost line, and that the epiotic of some holostean fishes is the homologue of the pholidophorid pterotic. Using this hypothesis, the confusion that has reigned regarding the endocranial bones of actinoptery- gians continues, although some names are shifted. His review of the subject suggests to the present author a more parsimonious and useful hypothesis, which is that the supraoccipital is derived from the occipital arch and that its primitive condition was paired preepioccipitals (Schaeffer, 1971, identified paired supraoccipitals in Mucrepistius). In the course of phylo- geny, development went in two directions. In the first, the preepioccipitals fused across the midline to form a supraoccipital, as in living teleosts. In the second, the pholidophorids, probably because of the strong attachment of nape muscles laterally, and the fused preepi- occipitals became subdivided into a midline “supraoccipital” and lateral epioccipitals. The original condition of paired preepioccipitals is seen in early stages of “Aspidorhynchus”, a caturid, in paraseminonotids, and Perleidus (Patterson, 1975, figs 97, 100, 115). Thus, in the teleost line, except for pholidophorids and Zchthyokentemu, only a supraoccipital is present. Paired preepioccipitals, or a supraoccipital, are not seen in amiids, semionotoids, chon- drosteans or polypterids. This last suggests that the preepioccipital developed only in some lines of actinopterygian fishes, not all. (Or alternatively, the preepioccipital was present in the ancestral actinopterygian fish but disappeared in some lines.)
From this approach (hypothesis) it follows that the pholidophorid epioccipital and teleost epiotic are not homologues. One is a superficial bone of the occipital arch, the other a bone of the cranial wall housing part of the inner ear. The question then results, “What is the epiotic?” First one should discuss names. Epiotic and pterotic are historically derived from studies of teleost fishes. As such these names are occupied and cannot be used for anything else. If the teleost epiotic is not derived from the pholidophorid epioccipital, from where did it come? The answer seems simple, but is dependent upon the idea of the “bone” as a multigenic field response, a field that can be cut in two or modified in a variety of ways. In the teleost, the “pterotic” of the pholidophorid, which for clarity can be identified as the pterotic-epiotic, appears to have been subdivided, probably by the posttemporal fossa, into a medial epiotic component with its centre overlying the posterior semicircular canal (Patterson’s centre for the pterotic) and a lateral component, the pterotic of classical ichthyology, with its centre lying lateral to the horizontal semicircular canal. The teleost epiotic is thus a fragment of an older bone with a centre of ossification associated with the posterior semicircular canal. These two components (epiotic, pterotic) and a supraoccipital are seen only in the teleost line. (An epioccipital is known only in pholidophorids.)
In living teleosts, the pterotic is ossified from the intertemporosupratemporal (Patterson’s dermopterotic) and does not appear as a separate element. In leptolepids and the caturid “Aspidorhynchus” (Patterson, 1975, figs 99, loo), both an epiotic (identified by him as pterotic) and a, seemingly chondral, pterotic (identified by Patterson in this organism as the opisthotic) are present. In parasemionotids (Patterson, 1975, fig. 97) only the medial (original) part is present. If one accepts these ideas of homology, the bone of the pholidophorid is a pterotic-epiotic and one can easily identify the bone seen in living non- teleostean actinopterygians as the pterotic-epiotic. (The teleostean pterotic is thus not the complete homologue of the pholidophorid “pterotic”.)
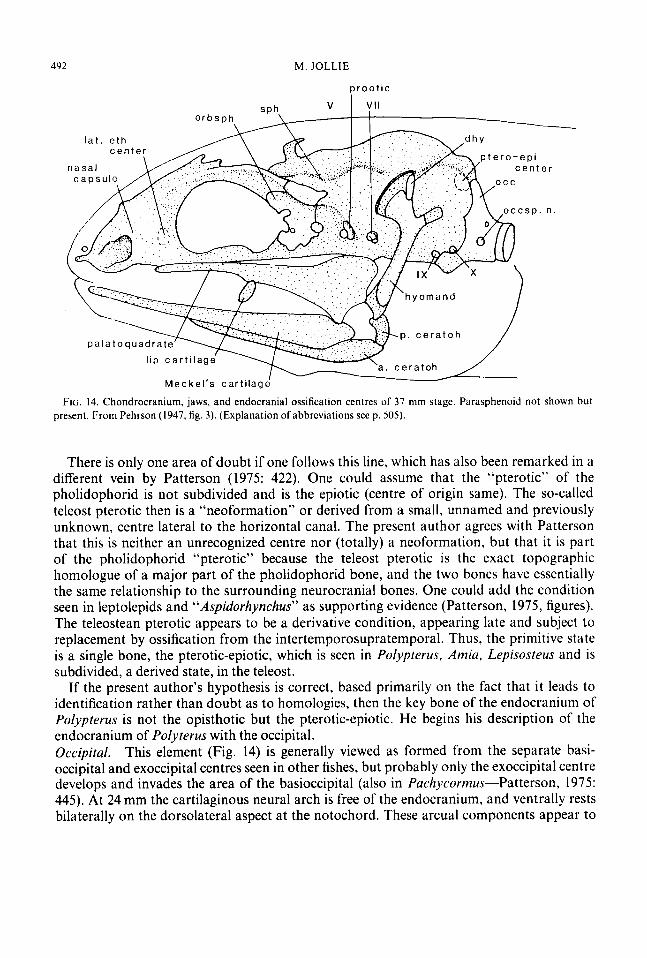
492 M . JOLLIE
FIG. 14. Chondrocranium, jaws, and endocranial ossification centres of 37 mm stage. Parasphenoid not shown but present. From Pehrson (1947, fig. 3). (Explanation of abbreviations see p. 305).
There is only one area of doubt if one follows this line, which has also been remarked in a different vein by Patterson (1975: 422). One could assume that the “pterotic” of the pholidophorid is not subdivided and is the epiotic (centre of origin same). The so-called teleost pterotic then is a “neoformation” or derived from a small, unnamed and previously unknown, centre lateral to the horizontal canal. The present author agrees with Patterson that this is neither an unrecognized centre nor (totally) a neoformation, but that it is part of the pholidophorid “pterotic” because the teleost pterotic is the exact topographic homologue of a major part of the pholidophorid bone, and the two bones have essentially the same relationship to the surrounding neurocranial bones. One could add the condition seen in leptolepids and “Aspidorhynchus” as supporting evidence (Patterson, 1975, figures). The teleostean pterotic appears to be a derivative condition, appearing late and subject to replacement by ossification from the intertemporosupratemporal. Thus, the primitive state is a single bone, the pterotic-epiotic, which is seen in Polypterus. Amia, Lepisosteus and is subdivided, a derived state, in the teleost.
If the present author’s hypothesis is correct, based primarily on the fact that i t leads to identification rather than doubt as to homologies, then the key bone of the endocranium of Polypterus is not the opisthotic but the pterotic-epiotic. He begins his description of the endocranium of Polyterus with the occipital. Occipital. This element (Fig. 14) is generally viewed as formed from the separate basi- occipital and exoccipital centres seen in other fishes, but probably only the exoccipital centre develops and invades the area of the basioccipital (also in Pachycormus-Patterson, 1975: 445). At 24 mm the cartilaginous neural arch is free of the endocranium, and ventrally rests bilaterally on the dorsolateral aspect at the notochord. These arcual components appear to

S K E L E T A L D E V E L O P M E N T O F P O L Y P T E R U S 493
FIG. 15. Endocranium of half-grown Po/vppreru.s with dermal bones of roof and snout still attached. (Explanation of abbreviations seep. 505).
have ossified and expanded around the notochord (the basioccipital component) by 37 mm. By 76mm the bone is essentially fully developed, perforated above the notochord by the spino-occipital nerves and fused dorsally to the foramen magnum. There is one vertebral body fused to the occipital. Its neural arch is separate and bears a rib and a neural spine. This occipital bone “does not extend beyond the vagus canal, which notches its anterior margin, and does not reach any part of the labyrinth cavity. Dorsally, it does not reach the roofing bones, leaving a quadrangular area of the cartilage” (Patterson, 1975: 463). This opening is seen in Fig. 10. Pterotic-epiotic. The endocranial part of this bone has appeared at 76 mm (Fig. 14) and the dermal process, associated with the ventral (lateral) prong of the posttemporal, appears later. The adult bone (Figs 8, 10, 12, 15) is massive and encloses the posterior and lateral aspects of the inner ear. The centre of origin is above the posterior vertical semicircular canal.
The present author has already indicated why he has chosen to call this the pterotic- epiotic, but will add some specific points to the argument. The homology of this bone has been questioned (Pollard, 1892; Allis, 1922; Patterson, 1973, 1975) with the result that there has been much confusion. The present author rejects Traquair’s (1 87 1) assumption that it is an epiotic-opisthotic because of lack of any evident appearance of the latter component in development. Certainly, this bone has extended into the area of the opisthotic. Allis (1922: 218) pointed out that the pterotic-epiotic (his opisthotic) houses the utriculus and the posterior ends of the horizontal and posterior vertical semicircular canals (Fig. 16).
Patterson (1975: 463-464) identified this bone as the opisthotic and noted that: “The opisthotic ossifies late, at more than 37 mm (Pehrson, 1947), from a centre.. . over the middle part of the posterior semicircular canal. In the adult, the bone becomes large and has extensive membrane bone outgrowths. The opisthotic meets the basi-exoccipital above and below the vagus canal, which is incised in its posterior edge. The opisthotic also forms the posterior margin of the facial canal, contains the glossopharyngeal canal, and lodges parts of the posterior and external semicircular canals. This bone could be interpreted as an opisthotic or a pterotic, or as representing both these bones. The centre of ossification of the bone is intermediate between that of a pterotic (above the posterior semicircular canal) and an opisthotic (lateral to the external semicircular canal) but its relations to the ninth and

494 M. JOLLIE
f ronta! p o s t . s. c .
/ t - s t \ h o r i z . s . C .
FIG. 16. Median view of posterior end of right half of the endocranium of unnumbered specimen in British Museum. Arrow through right posterior opening of aortic canal. (Explanation of abbreviations see p. 505).
tenth cranial nerves are those of an opisthotic, not a pterotic. . . the fact that the opisthotic is the dominant ossification in the otic region of palaeonisciforms [as in Perleidus], while the pterotic only becomes large in relatively advanced forms like Pachycorrnus and pholidophorids, favours interpretation of the bone in Polypterus as an opisthotic.”
The above can be criticized as misplacing the centre of origin of the opisthotic (the present author believes the opisthotic ossifies in the vicinity of the vagus nerve-anterior to it) and attempting to homologize the “pterotic” of the pholidophorid with a particular bone of other fishes, without realizing its equivalence to the epiotic and pterotic of the teleost. These acts can only result in confusion. Patterson’s description, from the location of the developmental centre to the secondary relation with the vagus, clearly identifies this bone as the pterotic-epiotic and indicates that Polypterus lacks an opisthotic. Lack of a prootic in Pulypterus would seem to go along with the same suppression of bone formation seen in the lack of an opisthotic.
A separate intercalar is lacking in Pol-vprerus but a strong intercalar process of the pterotic-epiotic serves as attachment of the ligament from the pectoral girdle. Prootic. According to Pehrson (1947), the prootic (Fig. 14) begins to ossify around the facial foramen at 37 mm, but by 45 mm it has disappeared and is not present thereafter.
There is a slight downgrowth of pterotic flange (process) from the inter- temporosupratemporal in Polypterus just as in Amia, Lepisosteus or E.sus (Figs 8(b), 10). This rudimentary development has been largely ignored by other writers. Sphenotic. The sphenotic (Fig. 14) has appeared at 27 mm and is already continuous with the dermosphenotic. This chondral bone is not distinct until 37mm. Fusion with the dermosphenotic is relatively distinct among actinopterygians (Figs 8(b), 10-also fused in catfishes, scorpaenids, beryciforms and Chauna; Patterson, 1977: 97). Orhifosphenoid, The orbitosphenoid (Fig. 14) is present at 24 mm in the orbit region and it comes to form the lateral and anterior walls of the brain cavity. Anteriorly, this plate lies laterally to the olfactory nerve, but in older fish it has extended medially to enclose the olfactory nerve and meet its opposite, and suture, at the midline (Jarvik, 1942, fig. 5A).

S K E L E T A L D E V E L O P M E N T OF P O L Y P T E R U S 495
n n hypoh YA 3 0. cerotoh
copula
ig. bone
cerobobr. JI!
porosph
( C ) ( b ) FIG. 17. (a) Ventral view of branchial arches and ends of hyoid arches. Ligament attachments of ligament bone and
urohyal shown to right. (b) Dorsal view of branchial and hyoid elements of mature fish from Allis (1922. pl. VIII. fig. 17). ( c ) Lateral view ofattachment ofdorsal end of the first branchial arch to the parasphenoid and otic capsule. (Explanation of abbreviations see p. 505).
Internal processes (Fig. 16) meet at the midline to form a bridge covering the pituitary similar to that formed by the prootic of other actinopterygians. The form and extent of this bone is unique. Patterson (1975: 464,469) has discussed some of the confusion relative to the homology of this bone; the present author has opted for the view, based on amphibians, of a single bone in this area which often breaks up into anterior and posterior parts (Jollie, 1962). Luferuletlimoid. This bone appears in the 71 mm stage; a thin lamina in the anterior wall of the orbit. It later forms the anterior wall of the orbit and the lateroposterior wall of the nasal cavity. Dorsally, it contacts the parietal, laterally the lateral rostral, ventrally the autopalatine, and medially the orbitosphenoid (see Jarvik, 1942, fig. 5A). Allis (1922: 230) described and figured three bones in the anterior orbit wall. Traquair, and others, observed only one pair. The preorbital process of the lateral rostral overlies i t laterally. N o ossifications occur in the wall of the eye.
Brunch id skr 1. ton Pehrson has not supplied data on the appearance of the branchial skeleton (Fig. 17), but
the anterior and posterior ceratohyals are present at 71 mm, as is the beginning of the hypohyal (a tubular perichondral ossification). The urohyal is present (anteriorly to the edge of the clavicles) as are paired ligamental ossifications. Four ceratobranchials, three hypobranchials and the copula are beginning to ossify.

496 M . JOLLIE
Hyoid arch
Epihyal (hyomandibula). The epihyal (upper half of hyoid arch) of this group is peculiar in that there is no symplectic cut from its distal end. It is not perforated by a facial nerve foramen and there is an accessory bone associated with its dorsal end. It is, therefore, a real epihyal. The “hyomandibula” is thus actually a hyomandibulosymplectic (Edgeworth, 1926, 1935). Bertmar (1959: 331-332) had doubts as to what the symplectic actually is, but reported that it was the distal end of the epihyal. This was assumed to be the case in Polypterus (see Moy-Thomas, 1933).
Comparison of the 8 mm stage of Polypterus (de Beer, 1937; Bertmar, 1959, fig. 79) with a similar stage of Esox, Salmo, Amia or Lepisosteus reveals that, unlike these other fishes, the entire epihyal in Polypterus ossifies as the “hyomandibula” and there is no tapered distal part. It is laid down in cartilage by the 8 mm stage (Fig. 1) but lacks the tapered distal end, associated with the quadrate, which would give rise to the symplectic. Ossification begins at 24 mm, at which stage it has already developed its characteristic shape-with a strong angle between proximal and distal limbs and a distinct opercular process (Figs 8(b), 14). Com- pared with most actinopterygians, this bone has a long distal (below the opercular process) portion which articulates with the posterior ceratohyal. As palaeoniscoids possibly had a symplectic, and the present author believes the primitive teleostome (defined morphologi- cally by Jarvik, 1977: 200), or osteichthyan, had at least the beginnings of one (i.e. slightly perichondrally ossified), this is a case of regression (a modified, derived state). (Loss of the symplectic in a catfish is a much later and separate event). The hyomandibula of the actinopts is said to include at its dorsal angle a “laterohyal” component. Bertmar (1959: 121-122), in a confusing account, equates the laterohyal with transformed gill rays. The present author has not seen a laterohyal in his materials so assumes it may not exist. Ceratohyal. The ceratohyal, like that of other actinopterygians, is in two pieces; the two parts are joined by ligaments, not by an overlapping suture. In most advanced actinoptery- gians there is a distinct cartilaginous (or bony) interhyal. This is lacking in Po/ypterus where a ligament suspends the posterior end of the ceratohyal. Hypohyal. There is a single (tubular) ossification centre for the hypohyal, as in Amiu or Lepisosteus (not two as in Salmo or Esox). A separate ossified basihyal is lacking. Urohyul. There is a small urohyal in the “coracoarcualis” muscles below the copula. Its anterior end is bifurcate and joined by connecting tissue, with ossifications in the tendons of the coracohyoideus muscles just before their attachments to the hypohyals (Jarvik, 1954, fig. lOC). Patterson (1977: 104-1 10) has discussed the urohyal at some length and presumed a relationship (p. 109) with the interclavicle. The present author’s own view is that the urohyal has nothing (directly) to do with the interclavicle (see pectoral girdle).
Branchial arche5
There are only four branchial arches in this group, each with a well-developed ceratobran- chial (Allis, 1922, pl. 8). The first three have hypobranchials which articulate with an ossified copula in older specimens. (The copula ossifies from front to back so that in the present author’s young adult specimens only the first two hypobranchials articulate with bone; hypobranchial 111 and ceratobranchial IV articulate with cartilage). This copula remains cartilaginous at its posterior end even in older fish. Ceratobranchial 1V bears two elongated toothplates on its pharyngeal surface and has a kink in the middle of its length, below

S K E L E T A L DEVEL.OPMENT O F P O L Y P 7 E R U . Y 491
(medial to) the margin of the slit (which marks a ligamentous cross connection with the other ceratobranchials).
The epibranchials are feebly developed as cartilages in this group. The first arch has an ossified suspensory piece (suprapharyngobranchial) which attaches to the underside of the bulge of the otic area of the cranium. There is also an infrapharyngobranchial (ossified in one of the present author's specimens, unossified in the other) which runs forward to attach to the end of the palatal process of the parasphenoid. This attachment is double; the cartilage or bone attaches medially and a broad ligament laterally (Fig. 17(c)). The second epibranchial is shown as having an ossified area by Allis (1922, pl. VIII, fig. 17).
The branchial system differs from that seen in other actinopterygians generally in the weak development of epibranchials and in that dorsal pharyngeal tooth plates are missing. The (ventral) tooth plates of the fourth ceratobranchials are weak. Each arch does have rows of small plates to either side, as in other fishes. The difference in the number of arches, the lack of ossified epibranchials, restriction of the slit to the ceratal region, and the copular structure suggests a quite distinctive fish. The copular structure is well matched by that seen in some palaeoniscoids (Gardiner, 1973).
Pectoral girdle and fins Dermal hones of 'girde
Jarvik (1944) and Jessen (1972) have described and figured the pectoral girdle of Polyp- terus, but no one has commented on its development. Pehrson commented only on the posttemporal, but i t can be assumed that all dermal elements had begun to ossify in the 24 mm stage. Posttemporal. The posttemporal is already present at 24 mm as a surface splint of bone. Its further development probably resembles that seen in Amia and Lepisosteus. There is one sensory organ in the canal developed on this bone.
The posttemporal (Figs 3, 4, 8-11, 18) is a distinctive, triangular, superficial plate fitted into the pattern of plates and scales of the nape region of the skull. It is sutured into the roof on two sides and as such is immovable. It is complexly shaped below the superficial plate. There is a posteriorly tapered prong for muscle attachment, and anteriorly a large, flat medial and a smaller lateral process for attachment to the cranium. The last of these processes does not extend beyond the margin of the plate and connects by a thick ligament with the intercalar process of the pterotic-epiotic. (There is no separate intercalar). The anteriorly directed processes suggest those seen in other actinopterygians (already present in ancestor), but the posterior prong is unique.
Teleosts do not have the heavy superficial plate seen in Polj.pterus, rather the body of the posttemporal is overlapped by scales. in Amiu there is a small superficial exposure of the posttemporal, while in Lepisosteus this bone is reduced to a small rectangular surface plate with only a slight indication of the deep (lateral) process. Supracleithrum. The supracleithrum (Figs 8 ,9 , 18) is large but without a canal. I t is tied by a strong ligament running medially to the parasphenoid (Baudelot's ligament). Cleithrum. The dorsal end of the cleithrum (Fig. 18) has a large anterodorsal prong like that seen in other actinopterygians. This is the major bone of the girdle and extends by a tapered process nearly to the ventral midline. Postcleithrum. There is a small postcleithral scale (Fig. 18) in this group but it is not known

49 x M. JOLLIE
FIG. 18. (a) Lateral view of pectoral girdle and adjacent scales. (b) Same with upper part of girdle removed and outline of fin lobe added. (Explanation of abbreviations seep. 505).
whether it appears early, as in other actinopterygians (where it occurs), or at the time of the other scales on the margin of the pectoral girdle. (The 71 mm specimen is fully scaled except on the bases of pectoral fins.) There is a second scale below the postcleithral scale which is sometimes included under this name but the present author would prefer to limit the term to that enlarged scale or plate below the overlap of the cleithrum and supracleithrum. (When the postcleithrum is reduced to a scale, no different from that of the body, then the bone is said to be missing.) Cluvicle. A large triangular clavicle (Fig. 18) overlaps the ventroanterior end of the cleithrum and meets along the midline with its opposite. There is no interclavicle in this group, although one is found in the sturgeons (Acipenser, Scaphirhynchus, but not Polyodon, Jollie, 1962, 1980; Patterson, 1977). The lack of this bone indicates modification from the ancestral condition.
Endochondral bones of girdle
Scapula. A small ossified scapula is present in this group. Unlike other actinopterygians (Jessen, 1972), there is no dorsal muscle canal formed by a mesocoracoid. The simple scapula extends anterolaterally from the glenoid articulation and is anchored on the cleith- rum. Jessen (1972: 83) noted that the shoulder girdle is unlike that of other actinopterygians, but did not recognize the basic difference which involves, primarily, the form of the scapula, and lack of the dorsal muscle canal.

S K E L ETA L D E V E I. OPM E N T 0 F PO L Y P T E R 1 ’ s 499
Coracoid. A well-developed coracoid, initially a perichondral ossification, extends ventro- laterally from the fin base to the cleithrum. The abbreviatcd nature of the coracoid is a part of the uniqueness of the Polyprevus girdle.
Prctoru1,fin
The fin structure is quite unique in its narrow base (lack of middle piece-Jessen, 1972) and this is related to its use in the group. Comparison of the pectoral of Polj~ptrrws with that in other actinopterygians is not necessary, but the differences indicate the ability to undergo considerable adaptive change. The anterior and posterior basal radials are present at 71 mm, but the middle plate is not. The distal radials are also ossifying at this stage.
Scales
N o information is available on the dcvelopment of scales in this group. Professor T. Pehrson in a letter to Miss A. E. Miller stated that there were no scales in Polvptrrus larvae shorter than 40 mm (Smith, 1956: 11). Sewertzoff (1932: 392), speaking of Po/i.plrrus, merely states, “Die Knockenschuppen bilden sich zuerst an den lateralen FlBchen des Korpers: die Schuppen der dorsalen und ventralen Region legen sie etwas spiiter an.” In the 71 mm specimen the body is fully covered and there are some scales on the base of the pectoral tin lobe. All that remains then is the covering of this lobe. The scales show a webby structure different from that seen in Lepisosteus, Anziu, or the teleost (see Meinke, 1982).
Discussion and conclusions
This review of the structure of the head skeleton and pectoral girdle of Po1vpterii.s suggests that the bones of fishes can be homologized, that the structure of Po!,,ptrrus is quite different from that of other actinopterygians, and that a number of features are valuable in the discussion of systematics of fishes.
The problems of homology have been discussed at great length by a number of prominent writers yet, oddly enough, remain. The system of names used here appears to be practical and presents no discouraging problems. The homologies indicated in Table I and in the account of the endocranium seem self-evident-all that can be expected in a comparative study. The only test of a system of names is its applicability. The alternative is a variety of names, some of which apply to the same structure as seen in different species, or the application of the same term to two or more non-homologous structures. The system of terms used in this account, with the substitution of those indicated in Table I, is applicable to all actinopterygians and, with some exceptions, to all osteichthyans (Jollie. I98 I ) . The exceptions are not “refuting” kinds of problems but can easily be dealt with in terms of known bone developmental (ontogenetic and assumed phylogenetic) dynamics. These dynamics are complex but can be summarized in a number of statements (others are given by Hall, 1975): (1) Skeletal elements are products of multigenic developmental mechanisms which
(2) The primitive skeletal unit is conceived of as tooth-like and larger elements were produce a field response.
formed by coalescence.

500 M. J O L L I E
Usual term used by ichthyologists Present author’s term
Dermosphenotic intertemporal Frontal parietal Parietal postparietal lntertemporosupratemporal supratemporotabular Nasal part of nasofrontal series
(3) Living forms lack any indication in their embryology of this early coalescence stage, only cellular blastematic masses suggesting the structure to be developed appear. i.e. the “field” of the structure.
(4) The skeletal blastema can give rise to any specific type of skeletal tissue: dentine, bone, cartilage, etc.
( 5 ) The blastemas generally involve interaction between mesodermal cells and ectomeso- dermal tissue-neural crest, delaminated ectodermal cells, or induction from the neural tube or notochord; the ectodermal “source” of much of the head skeleton is stressed.
(6) Separation into dermal (membral) and chondral bones is a secondary and evolved situation, but it is not clear at this time what the evolutionary relationship between these two types is. Certainly they are firmly established states in living species.
(7) Endochondral bones can have dermal extensions and dermal bones can have cartilagi- nous inclusions, or extend into and ossify chondral elements. A chondral bone can be converted into an apparent dermal bone (metapterygoid of this account).
(8) Blastemas appear to have a segmental correlation as well as relationship with active metabolic areas (and degrees of vascularization).
(9) As an active metabolic centre, the sensory organs have a strong, but not necessarily complete, control over bone formation; where the laterosensory system is strongly developed there is a direct, but not constant, relationship of organs and bones; where the laterosensory system is weak or lacking a similar basic pattern of bones results. Canal elements may be transferred from one bone to another.
(10) Bones, as “irregular” plates, not scales, have been derived by fusions of areas of scales with secondary decrease in number of centres with phylogeny-the trend is to a single centre for each bone.
(1 1) The field of a bone responds to adjacent field changes in its size and shape and may even be eliminated.
(12) As a result of changes in head morphology, bone centres are not fixed (topographically) and are not necessarily single.
( 13) Well established “bones” (named and fairly regular entities) can fuse (ontogenetically) with an adjacent bone or bones, whether dermal or cartilage. The mechanism of main- tenance of sutures is not known but it can be lost.
(14) When bones fuse ontogenetically, the tendency is for the developmental centre of one to take precedence over that of the other, and for ossification to extend from this

S K E L E T A L D E V E L O P M E N T OF POL Y P T E R U S 50 1
Halecostomes caturids , parasemionotids pholidophorids leptolepids teleosts amioids, semionotoids Perleidus
fused preepioccipitals divided into supra-
occipital and lateral epiocci pi to I
lass of preepioccipitols \ retained \ I fused preepioccipitals
form supraoccipital only
2 pterotic-epioiic sub- divided into separate
chondrosteans Polypterus
metotic fissure closed
not fused , f u s e ~ e p i o c c i p i t a l s \
loss of preepioccipitol operculum, as before
interopercle developed
I metotic fissure open 2 preepioccipitals paired 3 sphenotics and pterotic- epiotic present 4 symplectic well developed 5 dentosDlenial in lower law
Other osteichthyans (Sarcopterygii 1
I operculum \/ of opercle and subopercle
2 symplectic indicated
FIG. 19. Phylogenetic scheme derived from part of the information from this study.
centre into the area of the adjacent bone, thus replacing it. Thus bones appear to be competitive entities. The centre of one bone cannot move into the region of another until that bone has been eliminated. As a generality, there is a limit to the topographic shift of centres. The bone, as a field response, can be subdivided as the result of gross morphological changes in proportions, shapes or functional needs into two (or more) parts. Sub- division is not common and, in a sense, does not contradict the generalization that the phylogenetic course on Osteichthyes, and their derived tetrapods, is toward fewer head bones. A cartilage bone may be converted to a dermal one. (The reverse has been assumed but there is no good evidence of it.) Occasionally, new ossifications arise, but they develop late, tend to be even more irregular in shape than standard bones and, as such, can be viewed as secondary.
These statements can be viewed as ud hoc solutions to otherwise refuting observations, or they can be accepted as indicative of the plasticity of morphogenetic systems; an epistemological base.
The contribution of some of these anatomical data to the systematics of fishes is outlined in Table 11. Lists of such features in some accounts involve complexities of viewpoint that

TABLE I 1 Defining / e o w e s of Polypterus-Erpetoichthys ronipured with living.fislic~.s. D = dc.riverl. P =priniii iw, U = unique
D P PU P U
U
D D DU D DU
DU DU D D
DU D DU D D DU DU DU DU D D DU D DU D DU
DU P P D P
U P P DU D D D D
U U D DU
Lateral-line canal reduced to pits on body. Lip fold does not enclose rear of maxilla (Allis, 1919).
Medial rostral-internasal extends back under or between nasals to contact frontal. N o separate lateral rostral. but an element is appressed to the lateral aspect of the lateral ethmoid which passes upward medial to the posterior nasal opening. This element is fused with the premaxilla. Premaxilla. fused to three canal elements (see no. 5 ) . has triangular palatal process which “contacts” the anterior tip of the parasphenoid. Maxilla. fused with three canal elements, has posterior dorsal cheek process. Maxilla lacks anterior process articulating with ethmoid region. Preopercle with canal and cheek parts; latter forms broad cheek plate. Dermosphenotic fused to sphenotic. Intertemporosupratemporal composed of two units; e.ytends 10 midline I U repluce parietuls; has pterotic Range. Parietals present at 24 mm, gone by 37 mm. Squamosoids along cheek-roof suture. No quadratojugal. Boney plates of lower cheek are squamosoids (no contact with quadrate). Vomer much reduced and seemingly missing (fused to palatines) in adult fish. Midline rather than bilateral origin. Palatine not associated with autopalatine. Metapterygoid dermal (no “epipterygoid” process). Complex form of parasphenoid in ascending process Ned. Posterior hook of quadrate contacts ”hyomandibula”. Ectopterygoid contacts autopalatine. Orhitosphenoid single and with shelf over pituitary and saccus vasculous (replacing prootic). Prootic appears at 37 mm. lost by 45 mm. Exoccipital forms rear of cranium; no basioccipital. Pterotic-epiotic with a large intercalar process for the posttemporal. No sclerotic bones. N o opisthotic. “Hyomandibula” an epihyal, not perforated. N o symplectic. Dermohyal present. N o supraangular. Prearticular with large coronoid process (attachment of add. mandibulae); no coronoid process of Meckel’s cartilage. Lip cartilage at angle of mouth attached dorsally to tip ofcoronoid process of prearticular. Articular not subdivided into “articular” and retroarticular. Articular not fused with other jaw bones. Retroarticular process has dermal component. Meckel’s cartilage complete and lies in groove on inner surface of mandible; small free mentomeckelian ossification at symphysis. Large lateral gulars; no branchiostegal rays. Two-piece ceratohyal. Hypohyal a single piece. Four branchial arches, slits limited to ceratal region. Epibranchials small, no dorsal pharyngeal tooth plates. Large external gill in larva. No interclavicle. Posttemporal medial to lateral (extrascapular). firmly sutured into skull roof (like LqJi.so.s/eu.s): basically a roof plate. Posttemporal with posterior muscle process. N o canal element associated with supracleithrum; process for Baudelot’s ligament. Postcleithral scale reduced. Unlike primitive actinopterygian. scapulocoracoid cartilage has no dorsal canal Tor dorsal fin muscles. Scapula a single piece rather than with a separate mesocoracoid ossification or process.
Three SCpardte WdSdl hones.

S K E L E T A L D E V E L O P M E N T O F POL Y P T E R U S 503
are difficult to understand. Those used here are discussed in the text, with a couple of exceptions. Many of the details of this table suggest that Po1yptcm.s (and Erperoichrhys) could be separated as the Brachiopterygii, a sister group of the rest of the living Actinop- terygii. Such a viewpoint, which the present author has supported in the past, simply does not stand up to analysis (Fig. 19). It does speak loudly of the differences, but loses sight of the phylogenetic necessities.
I would like to thank Dr Ragnar Olsson for specimens from the University of Stockholm collection, and Dr Tor 0 v i g and Dr Colin Patterson for their many kindnesses in making available ideas, materials, and facilities at the Riksmuseet and British Museum. The published works of the late Dr Torsten Pehrson have been most valuable, as have the developmental studies of Dr Gunnar Bertmar. These have largely shaped my thinking. I want to thank Dr Colin Patterson for his criticisms of the first draft of this paper. Last of all, 1 wish to thank Northern Illinois and the people of Illinois for the time and support to do this research.
REFERENCES
Allis. E. P., Jr. (1900). The lateral sensory canals of Po/yp/rrus hichir. Anut. An-. 17: 433451. Allis. E. P., Jr. (1919). The homologies of the maxillary and vomer bones of Po/i~/Jtrric.s. Atu. J . Ancrt. 25: 349 3XX f 6 . Allis. E. P.. Jr. (1922). The cranial anatomy of Po/vp/(w.s with special reference to Po/j,p/wr.s hichir. J . A m / . , Lorid. 56:
Ayres, w. P. (1850). Description of a new species of Po/j.p/cru.s from West Africa. Boslori J . nu/. Hbsr. 6 241 246. Berg, L. S. (1940). Classification of fishes. both recent and fossil. Trirc/j, zoo/ . / t i .s / . , Lcwingr. 5: X7 517. Berg, L. S. (1958). S ~ ~ . s / c n i der rc'zen/cw cine/ /iJ.\,si/cn Fi.sc/iur/igcw und F/.sc/ie,. (Translated f r o m the 2nd Russian edition).
Bertmar. G. (1959). On the ontogeny of the chondral skull in Characidae. with a discussion on the chondrocranial base
Budgett. J . S. (1902). On the structure of the larval Po/j,prcw.s. Truns. ZOO/. SO(.. Lond. 1 6 3 I 5 340. Collinge, W. E. (1893). Note on the lateral canal system ofPo/).p/cw.s. Pro(. . Bir/n. phi/ . SOC.. 8: 255 262. Daget, J . (1950). Revision des affinites phylogenetique des Polypterides. Mini. h i s t . 1;. A f i . noirc No. 1 I : I 17X. Daget, J . (1958). Sous-Classe des Brachiopterygicns (Brachiopterygii). In TruirC dc Z O O / O ~ ~ C V A ~ / o t i i i ( , . Sj..srCniu/iyirc,.
Daget. J. & Arnault, J . (1965). Etude de la croissance chez Po/),prcw.s .sc~nc~gcr/u.s Cuvicr. A c / u ZOO/.. Stockh. 46: 2Y7 309. Daget. J., Bauchot. M. -L., Bauchot. R. & Arnault. J . (1964). Developpcinent du chondrocrinc et dcs arcs aortiques Che7
de Beer, G. R. ( 1937). The, dridopnicwi o / / k i w t c h i r c , .skir//. Oxford: Oxford Univ. Press. Devillers, C. ( 1958). Le Crine des poissons. In Trui/C de Zoohgie: Anulonlie. Sy.s/lniu/iyuc~, Biologic,: 1 3 55 I ~687, Grasse,
Edgeworth, F. H. (1926). On the hyomandibula of Selachii, Teleostomi and Ccratodus. J . Antrr. Lord. 60: 173 I Y3. Edgeworth, F. H. (1935). T/ie cruniu/musc/es ofver/rhru/e.v. Cambridge: Cambridge University Press. Gardiner, B. G . (1960). A revision of certain actinopterygian and coelacanth fishes. chiefly from the lower lias. Bull. Br.
Gardiner, B. G. (1973). Interrelationships of tekostomes. Zoo/. J . f h i . Sot,. 53 (suppl. I ): 105 135. Gardiner, B. G. & Bartram. A. W. H. (1977). The homologies of ventral cranial fissures i n osteichthyans. Linn. SW.
Gerard, P. ( 1937). Les appareils sensoriels de I'epiderrnc chez Po/j~p/cric.s t i d i . s i . Anncr/.s So( , . ZOO/. Br/gc 67: 33 40. Gregory, W. K . (1933). Fish skulls. a study of the evolution of natural mechanisms. Trtrri.\. ,4nl. phi/ SOC. 2 3 75 48 I . Hall, B. K . (1975). Evolutionary consequences of skeletal differentiation. A m . Zoo/. 15: 329 -350. Harrington, R. W. ( 1955). The osteocranium of the American cyprinid lish. No/ropis / ir/rcnu/u.\, with a n annotated
Holmgren, N. (1928). Notes on the skull of ~u/uf~ifJ;c' / i / / i .~' .s . Ac,/tr Zoo/. . S / O ~ ~ / I . 9: 327 329. Holmgren, N. (1942). General morphology of the lateral sensory line system of the head i n fishes. K. .\i~on.sLu
Holmgren. N. (1943). Studies on the head of fishes. Pt. IV (General morphology of the head in fishes.) A(,rtr ZOO/.. Srocklr.
189-294.
Berlin: Deutschen Verlag der Wissensch.
and the visceral chondrocranium in fishes. Ac,rrr ZOO/.. S/(~c,kli. 40: 203 364.
Biologic. 13: 2501- 2521. Grasse, P. -P. (Ed.). Paris: Maison et Cie.
Pdypterus senegulus Cuvier. Ac/u ZOO/. , S / ~ J ( , ~ / I . 4 6 20 I -244.
P.-P. (Ed.). Paris: Masson et Cie.
Mus. nu/. H i s / . (Geology) 4 239-384.
Synip. Sc,ric,.s No. 4: 227 245.
synonymy of teleost skull bones. CopcJiu 1955: 267 290.
Ve/en.skAkud. Hund . (3) 2 0 I 46.
2 4 1-188.

504 M . J O L L I E
Holmgren. N. & Stensio, E. A. (1936). Kranium und Visceralsketett der Akranier. Cyclostomen und Fische. Hundh. ivrKl.
Jardine, N. (1969). The observational and theoretical components of homology: a study based on the morphology of the
Jarvik, E. (1942). On the structure of the snout ofcrossopterygians and lower gnathostomes in general. Zoo/. Birlr. Upps.
Jarvik, E. ( 1944). On the exoskeletal shoulder-girdle of the teleostomian fishes. with special reference to Elts/hroprc,r(~,i
Jarvik, E. (1947). Notes on the pit-lines and dermal bones of the head in Poljp'lerus. Zoo/. Eiclr. U p / n 2 5 6&78. Jarvik, E. (1954). On the visceral skeleton in Eusrhencperon with a discussion of the parasphenoid and palatoquadratc in
fishes. K . svensku VelenskAkud. Hand/. (4) 5: 1-104. Jarvik, E. (1967). The homologies of frontal and parietal bones in fishes and tetrapods. Co//oyuc~.s / n / . Cmr. nuin. Rcch.
Scient. No. 163: 181-213. Jarvik, E. (1972). Middle and Upper Devonian porolepiformes from East Greenland with special reference to G/ypro/epis
groenlundicu n.sp. and a discussion on the structure of the head in the Porolepiformes. Meclll. Gronluncl 187: 1-307.
Anat. Wirhelriere 4 233-500.
dermal skull-roofs of rhipidistian fishes. B i d . J . Linn. Soc. I : 327-361.
21: 235-675.
Jbordi Whiteaves. K . svensku VetenskAkud. Hundl. (3) 21: I - 32.
Jarvik, E. (1977). The systematic position of acanthodian fishes. Linn. Soc. Symp. Ser. No. 4: 199 225. Jessen, H. (1972). Schultergiirtel und Pectoralflosse bei Actinopterygiern. Fossils Srruru, O.do I : I 10 I . Jollie. M . (1962). Chordate morphology. New York: Reinholt. Jollie, M. (1969). Sensory canals of the snout of actinopterygian fishes. Truns. ///. S/. Acud. Sci. 62: 61 69. Jollie. M. (1975). Development of the head skeleton and pectoral girdle in Ews. J . Morph. 147: 61-88. Jollie, M. (1980). Development of head and pectoral girdle skeleton and scales in Acipenser. Cupcirr 1980 226249. Jollie, M. (1981). Segment theory and the homologizing ofcranial bones. Am. Nu/. 1 1 8 785 802. Jollie, M. (1984). Development of cranial and pectoral girdle bones of Lepisosreus with a note on scales. Cop& 1984:
Kerr. J. G. (1907). The development of Polyprerus senegalus Cuv. Budgerr Mem. Volume: 195-284. Lehman, J . -P. (1966). Brachiopterygii. In TruirP de Puleuniologie, 4(3): 413-420. Piveteau. J . (Ed.). Paris: Masson et Cie. Lehn. C. (1918). Beitrag zur Kenntnis des Primordialschadels von Poljpprerus. Z. ungew. Anur. Konsrir Lrhre 2: 349~408. Meinke, D. K. (1982). A light and scanning electron microscope study of microstructure. growth and development of the
Moy-Thomas, J. A. (1933). Notes on the development of the chondrocranium of Polypprerus .senegulu.v. Q . J l micro.sc. Sc,i.
Moy-Thomas, J. A. (1934). On the teeth of the larval Eelonr vulgaris and the attachment of teeth in fishes. Q. Jlmicrosc.
Nelson. G . (1973~) . Notes on the structure and relationships of certain Cretaceous and Eocene teleostean fishes. Am.
Nelson. G . (19736). Relationships of clupeomorphs, with remarks on the structure of the lower jaw in fishes. Zool. J .
Nielsen, E. (1936). Some few preliminary remarks on the Triassic fishes from East Greenland. Mecldl. Gronlund I 1 2 1-55. Nielsen. E. (1942). Studies on triassic fishes from East Greenland. I . Gluucolrpis and Eforeosomus. Pulueomo. Groenlund.
Nielsen, E. (1949). Studies on triassic fishes from East Greenland. Pu1eo:oo. Groenlund. 3: I 309. Parrington. F. R. (1949). A theory of the relations of lateral lines to dermal bones. Proc. :oo/. Soc. Lond. 119: 65-78. Parrington. F. R. (1956). The patterns of dermal bones in primitive vertebrates. Proc. :oo/. SOC. Lond. 127: 3 8 9 4 1 I , Patterson, C. (1973). Interrelationships of holosteans. ZOO/. J . Linn. Soc. 53 (Suppl. No. I ) : 233-305. Patterson. C. (1975). The braincase of pholidophorid and leptolepid fishes, with a review of the actinopterygian
Patterson, C . ( I 977). Cartilage bones, dermal bones and membrane bones, or the exoskeleton versus the endoskeleton.
Pehrson. T. (1922). Some points in the cranial development of teleostomian fishes. Acru Zoo/ . , Srockh. 3 1-63. Pehrson. T. (1947). Some new interpretations of the skull in Polypferus. Actu Zool., Srockl7. 28: 399455.
476-502.
dermal skeleton of Polypterus (Pisces: Actinopterygii). J . Zool., Lond. 197: 355-382.
(N.S.) 7 6 209-229.
Sc,i. (N.S.) 7 6 481498.
MUS. Novir. NO. 2524: 1-3 I ,
Linn. Soc. 53 (Suppl. No. I ) : 333- 349.
1: 1403.
braincase. Phil. Truns. R . Soc. Lond. (B.) 269: 275-579.
Linn. Soc. S m p . Ser. No. 4: 77-121.
Pehrson. T. (1958). The early ontogeny of the sensory lines and the dermal skull in Polyprerus. Acru Zool., Srockh. 39: 241-258.
Pollard, H. B. (1892). On the anatomy and phylogenetic position of Polyppterus. Zool. Jb. (Anat.) 5: 525-550. SCve-Soderbergh. G . (1933). The dermal bones of the head and the lateral line system in Osreolepis macrolepidorus. Nova
Acru Reg. Soc. Scienr.. Llpsul. (4) 9: I - 129.

S K E L E T A L D E V E L O P M E N T O F P O L Y P T E R U S 505
Schaeffer. B. (1968). The origin and basic radiation of the Osteichthyes. In Currenr prohlerns of' lower vertehrura
SchaefTer. B. (1971). The braincase of the holostean fish Muc,rc,pi.stiu.v with comments on ncurocranial ossification in the
Schmah. R. (1934). Die Entwicklung der Unterkisferknochen Bei Po1j.preru.r. Morph. Jh. 7 4 364 379. Sewertzoff. A. N. (1925). Zur Morphologie des Schadels von Po/j,p/rru.v ddliezi. Anur. Anz. 59: 271 278. Sewertzoff, A. N. (1932). Die Entwicklung der Knochenschuppen von Poljp/rrrrs delhc~i. Jontr Z . Nururw. 67: 387-392. Smith. I . C. (1956). The structure of the skin and dermal scales in the tail of Aciptwser ruthrnus L. Truns. R . Soc. Edith.
Starks, E. C. (1901). Synonymy of the fish skeleton. Pro(,. Wusli. Acud. Sci. 3: 507-539. Stensio. E. A. (1947). The sensory lines and dermal bones of the cheek in fishes and amphibians. K . .vwnsku Verensk Akud.
Swinney, G. N. & Heppell, D . (19x2). Erperoichrhj~.~ or Culm~oiclr/l~j~s.. the correct name for the African Reed-fish. J . nut.
Traquair, R. H. (1871). On thecranial anatomy of Po/jptrru.r. J . Anur. Physiol. (2)7: 166183. van Wijhe, J. W. (1882). Ueber das Visceralskelett und die Nerven des Kopfes der Ganoiden und von Ceru/odus. Nietkcvl.
Westoll. T. S. (1936). On the structure of the dermal ethmoid shield of 0sreolepi.r. Grol. Mug. 7 3 157 171.
phjhgeriy; 207-222. Orvig, T. (Ed.). (Nobel Symp. 4). Stockholm: Almqvist & Wiksell.
Actinopterygii. Am. Mus. Nevi/. No. 2459: I 34.
6 3 1-14.
Hundl. (3) 24 (3): 1-195.
His/. 1 6 95-100.
Arch. Zool. 5: 207-320.
ang ano aPl art autopal a.ceratoh ceratobr I-IV dhY dso dsop dsph dspl epter esc f gPl hmpl horiz. s. c hP1 h yomand hypobr I hYP0h . . '1-'6 intercal. pr
it-st lac
iP1-iP6
List of abbreviations used on figures
an g u 1 a r anterior nasal opening anterior pit-line articular autopalatine anterior ceratohyal ceratobranchial I-IV dermohyal dermosphenotic organ dermosphenotic pore dermosphenotic dentosplenial ectopterygoid extrascapular organ or bone frontal gular pit-line horizontal mandibular pit-line horizontal semicircular canal horizontal pit-line h yomandibula hypobranchial I hypohyal infraorbital organs intercalar process of pterotic-epiotic infraorbital pores intertemporosupratemporal lacrimal

506
lat.esc lat.eth lat. gular lat. ros lig lig. bone
m. esc
mP1 mPt m. ros-intern mx n3 n. op nPl n. sp
occ. a occ. pl occsp. n OP orbsph
ml-m5
mp2-mp6
occ
Pl-P3 Pal pal. pr. pmx Par parasph part p. ceratoh pharyngobr I Pmx pmx-ros Pno POP porb p0st.s.c. PtO PP1-PP3 PPl Pt pter ptero-epi sip1 quad
iPs6
M. J O L L I E
lateral extrascapular lateral ethmoid lateral gular lateral rostral ligament bone in ligament mandibular organs medial extrascapular mandibular pores middle pit-line metapterygoid medial rostral-internasal maxilla nasal 3 nasal opening nasal pit-line neural spine occipital occipital arch occipital placodes occipi tospinal foramen opercle o rbi tosphenoid preopercular organs palatine palatine process premaxilla parietal parasphenoid prearticular posterior ceratohyal pharyngobranchial I premaxilla premaxilla plus rostral posterior nasal opening preopercle pos torbi tal posterior vertical semicircular canal posttemporal pore preopercular pores posterior pit-line posttemporal pterygoid pterotic-epiotic q uadratoj ugal pi t-line quadrate supraorbital organs

SOP SPl, SP2 SPh SPl squam vmpl VPl I-x
S K E L E T A L D E V E L O P M E N T OF P O L Y P T E R U S
subopercle supraorbital pore sphenotic symphyseal pit-line squamosoid(s) vertical mandibular pit-line vertical pit-line cranial nerves
501