determination of anti-lipid a and lipid a by enzyme immunoassay
TRANSCRIPT

Immunobiol., vol. 158, pp. 380-390 (1981)
Institute for Clinical Chemistry, Medical School, Hannover!, and Max-Planck-Institut fiir Immunbiologie2
, Freiburg/Breisgau, Federal Republic of Germany
Determination of Anti-Lipid A and Lipid A by Enzyme Immunoassay
PETER C. FINK 1 and CHRIS GALANOS2
Received December 17, 1980· Accepted January 24,1981
Abstract
Immunizing rabbits and goats with acid-treated Salmonella minnesota R595 bacterial cells, that expose lipid A on their surface, an IgG anti-lipid A antibody response could be obtained. In passive hemolysis test, using lipid-A-coated sheep red blood cells and complement, the highest IgG anti-lipid A antibody activity was found in immunized rabbits, while goats produced a relatively weak antibody response against lipid A. The passive hemolysis test showed that IgG anti-lipid A antibody activity was completely abolished by preceding preincubation with lipid A. No reactivity of IgG anti-lipid A antibodies was found with the tested intact lipopolysaccharides from one Salmonella abortus equi and one Escherichia coli strain. Quantitative analysis of free lipid A was carried out by the enzyme-linked immunosorbent assay (ELISA). In sandwich antigen type ELISA, using goat IgG anti-lipid A antibodies bound to the wells of polystyrene microassay plates, rabbit IgG anti-lipid A antibodies and peroxidase-labeled goat anti-rabbit antibodies (PGRA), the measuring range for free lipid A (0.001 to 1 ~g/ml) correlated significantly (2p < 0.01) with absorbance. In sandwich antibody type (SAB) ELISA binding lipid A to the solid phase, and using rabbit IgG anti-lipid A antibodies and PGRA insignificant correlation could be found for the same measuring range (2p > 0.1). Comparison of passive hemolysis test and SAB type ELISA to determine IgG antilipid A antibody activity showed that ELISA was more sensitive for measuring anti-lipid A antibody activity (from antibody dilution of -log: 2.43 to 3.7) than the passive hemolysis test.
Introduction
Gram-negative septicemia is known to be a clinical entity with a high
mortality rate (1-3). Many pathological alterations appearing on various
organs (e.g. lung, kidney, liver, brain) (4-9) in process of gram-negative
infections have been assigned to the deleterious effects of lipopolysaccharides (LPS, endotoxin) which are released from the cell wall during
bacteriolysis. The detection of LPS in peripheral blood may be carried out
by bioassays such as the Limulus amebocyte lysate test (10, 11), USP rabbit
Abbreviations: LPS = lipopolysaccharide; USP = United States Pharmacopeia; PBS = phosphate-buffered saline; SRBC = sheep red blood cell, VBS = veronal-buffered saline; PHT = passive hemolysis test; PHIT = passive hemolysis inhibition test; ELISA = enzymelinked immunosorbent assay; SAB = sandwich antibody; SA = sandwich antigen; PGRA = peroxidase-labeled goat anti-rabbit antibody.

Anti-Lipid A and Lipid A . 381
pyrogen test (12), histamine hypersensitization test (13), and the chick embryo cytotoxicity test (14). In these assay systems which measure the biological activity of endotoxin rather than endotoxin per se, the question of specificity often arises. The clinical value of other endotoxin-sensitive immunoassays (15, 16) may be curtailed by their limited applicability because of their restricted specificity for antigenic ally (O-antigen) defined endotoxins.
Recent studies by several authors (17) have demonstrated that the lipid component of LPS, namely lipid A, encompasses both the toxic site and the least variable antigenic part of the LPS of numerous gram-negative bacteria. In addition, common immunodeterminants in lipid A were found to elicit the production of cross-reacting highly specific anti-lipid A antibodies (18-20). While the procedure of detecting the cryptic antigenic sites of lipid A in intact LPS by cleaving the 2-keto-3-deoxy-octonate trisaccharide linkage between polysaccharide and lipid A appears at present to be unsuitable for clinical use, the determination of anti-lipid A antibodies in serum may provide an appropriate diagnostic tool since the presence of such antibodies in certain levels would reveal contact of the organism with LPS regardless of serospecificity.
To establish a suitable assay system for anti-lipid A antibodies, antisera were raised in rabbits and goats, and purified anti-lipid A IgG was compared in the passive hemolysis test and in an enzyme immunoassay (ELISA).
Materials and Methods
Preparation of lipid A antisera
Antisera with specificity against lipid A were raised in rabbits and goats by repeated immunizations with acid-treated Salmonella minnesota R595 bacteria coated additionally with free lipid A as described elsewhere (20). Rabbits received 100 [.Ig, 200 [.Ig, 300 [.Ig and 500 [.Ig lipid A vaccine on days 1, 4, 7 and 11, respectively, administered intravenously in 1 ml physiological phosphate-buffered saline (PBS, 0.01 molll, pH 7.2). On day 39 the animals were boostered with 500 [.Ig vaccine, administered as above, and 6 days later blood was collected from the marginal ear vein.
Goats were immunized on day 1 with 100 [.Ig lipid A vaccine. The antigen suspension (100 [.Ig in 2 ml PBS) was emulsified with 2 ml incomplete Freund's adjuvant (Behringwerke, D-3550 Marburg), and 1 ml was injected subcutaneously at four different sites. At monthly intervals 3 further booster injections were carried out with 500 [.Ig vaccine in incomplete Freund's adjuvant as described above. Twelve, twenty and twenty-five days after the last booster injection, blood was drawn from the jugular vein.
All blood samples after allowing to clot were centrifuged at 1900 g for 20 min at 4 °e, heatinactivated for 30 min at 56 °e and absorbed with packed sheep red blood cells (SRBC) (Biolog. Arbeitsgem. D-64202 Lich) (0.2 ml packed cells/5 mllipid A antiserum) for 30 min at 37 °e. The cells were pelleted at 1000 g for 10 min at 20 °e, and the serum supernatant was stored at - 60 °e until use.

382 . P. C. FINK, and CH. GALANOS
Chromatographic isolation of IgG immunoglobulins
A crude gamma-globulin fraction was obtained from rabbit and goat lipid A antisera by two precipitations with 50 % saturated ammonium sulfate. The final precipitate was dissolved in PBS (II, of the original serum volume) and dialyzed overnight against 51 PBS. The IgG fraction was prepared by elution from a DEAE-Sephadex A 50 column (2.6 X 40 cm) (Deutsche Pharmacia, D-7800 Freiburg) by PBS and concentrated by Minicon B 15 (Amicon, D-5810 Witten) (21). The protein content was determined spectrophotometrically according to LAYNE E. (22).
Immunochemical characterization of the eluted proteins
The purity of the rabbit and goat IgG preparations was tested by immunoelectrophoresis using polyspecific goat anti-rabbit plasma protein, rabbit anti-goat plasma protein (Medac, D-2000 Hamburg) and mono specific goat anti-rabbit IgG (Paesel, D-6000 Frankfurt) and rabbit anti-goat IgG (Byk-Mallinkrodt, D-6057 Dietzenbach-Steinberg).
Determination of anti-lipid A activity and specificity of the IgG preparations
Anti-lipid A antibody activity was measured by the passive hemolysis test (PHT) using lipid A sensitized SRBC as described elsewhere (20, 23). Briefly, washed SRBC (0.2 ml packed cells in 5 ml PBS) were incubated with alkali-treated lipid A (40 fll, 2 mg/ml) for 30 min at 37°C. They were washed three times with PBS and the pellet resuspended in 11 ml of veronalbuffered saline (VBS, 0.02 molll, pH 7.3 containing 15 mmolll CaCI2 and 0.5 mmolll MgCI2).
To serial dilutions (in 0.1 ml VBS) of the rabbit and goat IgG preparations, VBS (0.8 ml), lipid A sensitized SRBC (0.5 ml) and guinea pig complement (0.1 ml diluted 1 : 12 with VBS) (Behringwerke, D-3550 Marburg) were added and the samples incubated for 60 min at 37°C. Identical samples containing instead of sensitized, normal SRBC served as controls. In addition, samples from which IgG, complement or both were omitted, as well as blanks containing IgG or complement alone were included as controls. After incubation the cells were centrifuged at 1900 g for 20 min at room temperature and hemolysis in the supernatants measured at 546 nm. Anti-lipid A hemolytic activity was expressed as the negative logarithm (-lOgIO) of the dilution at which 50 % of sensitized SRBC occurred (e.g. -log ['132] = 1.51).
In the passive hemolysis inhibition test (PHIT) the rabbit IgG anti-lipid A preparations were preincubated with serial dilutions of lipid A (100 ng - 10 pg/ml) or LPS from Salmonella abortus equi (Novo-Pyrexal forte) (Hermal-Chemie, Kurt Hermann, D-2057 Reinbek b. Hamburg) or E. coli 055: B5 LPS (Difco laboratories, Detroit, Michigan, USA) (100 ng-10 pg/ml) respectively for 60 min at 37°C. After centrifugation, the supernatants were processed under identical conditions as in PHT, and hemolysis was determined.
Quantitation of anti-lipid A and lipid A by ELISA
Using rabbit and goat anti-lipid A antibodies of known hemolytic activity, two types of ELISA were carried out in microtitration plates (No.: M129 A, MicroElisa Reader AM 115, Compuprint 700, Dynatech, D-7310 Plochingen) as described elsewhere (24, 25) with some modifications. In sandwich antibody (SAB) type ELISA, the solid phase was sensitized with alkali-treated lipid A (1 flg/0.2 ml) dissolved in carbonate-bicarbonate buffer 0.05 molll, pH 9.7 for 60 min at 37°C. After washing three times with PBS/Tween 20R, the wells were incubated with bovine serum albumin (BSA, 5 gil) (Behringwerke, D-3550 Marburg) for 30 min at 37°C to reduce non-specific binding. Subsequent to three washings with PBSI Tween 20R the wells were incubated with serially diluted rabbit anti-lipid A antibodies (1 : 10 to 1 : 5000) (3.68 mg/0.2 ml) for 60 min at 37°C. The wells were washed again and the enzyme conjugate (0.2 ml) (an immunoadsorptive purified preparation of horse-radish peroxidaselabeled goat anti-rabbit IgG antibody, PGRA, kindly provided by Dr. Grenner, Behringwerke, D-3550 Marburg) resuspended in 0.05 molll tris(hydroxymethyl)aminomethan-HCI buffer pH 7.4 and BSA 2 mg/ml was added for 60 min at 37 cC. Continuing with the B/F separation step, the substrate, 1,2-phenylendiamin (1.5 mg/ml) (No.: 7243, Merck, D-3400 Giittingen), dissolved in 0.2 molll citrate-phosphate buffer pH 5.0 activated with hydrogen

Anti-Lipid A and Lipid A . 383
peroxide (300 g/ I) was added for 10 min at room temperature. The product of the enzymic reaction was read by photometry at 450 nm wave length.
In another series of SAB type ELISA, the solid phase was coated with varying lipid A concentrations (0.05 !J,g to 10 !J,g/0.2 ml). A constant rabbit anti-lipid A antibody dilution (-log: 1.6) was added, and the assay was performed under identical conditions as described above. The ELISA titer was defined as the negative logarithm (-logIC) of the anti-lipid A antibody dilution. To determine non-specific binding, lipid A was replaced by PBSTween 20R, and the extinction values for a serially diluted rabbit IgG fraction or PGRA were regarded as the background. In all tests a rabbit lipid A antiserum control of known antibody activity was included.
In sandwich antigen (SA) type ELISA goat anti-lipid A antibodies (3 .3 mg/0.2 ml) were bound to the solid phase. After treatment with BSA and washing, serial dilutions of alkalitreated lipid A (0.0005 Ilg to 20 Ilg/0.2 ml) were pipetted onto the presensitized solid phase and incubated for 60 min at 37 °C. The second rabbit anti-lipid A antibody (3.6 mg/0.2 ml) fixed to still exposed antigenic determinants of lipid A was indirectly labeled by measuring bound PGRA. In SA type ELISA the exstinction values for PGRA bound to the sensitized solid phase or lipid A was regarded as the background. The precision data referring to mean values of triplicate determinations for SAB and SA type ELISA were satisfactory (coefficients of variation within series ranged from 8 to 23 %).
Statistics: Spearsman's coefficient of rank correlation test was employed for comparison of methods (26).
Results
Immunochemical characterization of eluted fractions from lipid A antisera.
As demonstrated in Figure 1, the immunoelectrophoretic determination of eluted fractions obtained from rabbit and goat lipid A antisera showed no other plasma protein than IgG immunoglobulin.
Anti-lipid A activity and specificity testing of IgG preparations from lipid A antisera
The results show that lipid A vaccine engendered the formation of IgG anti-lipid A antibodies in rabbits and goats (Fig. 2). Using PHT to determine anti-lipid A activity in the eluted fractions, the highest IgG-antilipid A titer (-log: 2.43) at 50 % hemolysis was produced from immunized rabbits, whereas from goats IgG anti-lipid A antibodies were found in titers ranging from -log: 1.23 to -log: 2.73.
The specificity of rabbit IgG anti-lipid A antibodies was assayed by PHIT (Fig. 3). While a 100 % reduction of IgG anti-lipid A antibody activity could be obtained by preincubation with high lipid A concentrations, an increase of IgG anti-lipid A antibody activity was found in proportion to decreasing lipid A concentrations. In contrary, no loss of hemolytic activity of IgG anti-lipid A antibodies was shown by PHIT following the preceding preincubation of IgG anti-lipid A antibodies with intact Novo-Pyrexal forte LPS or E. coli 055 : B5 LPS.
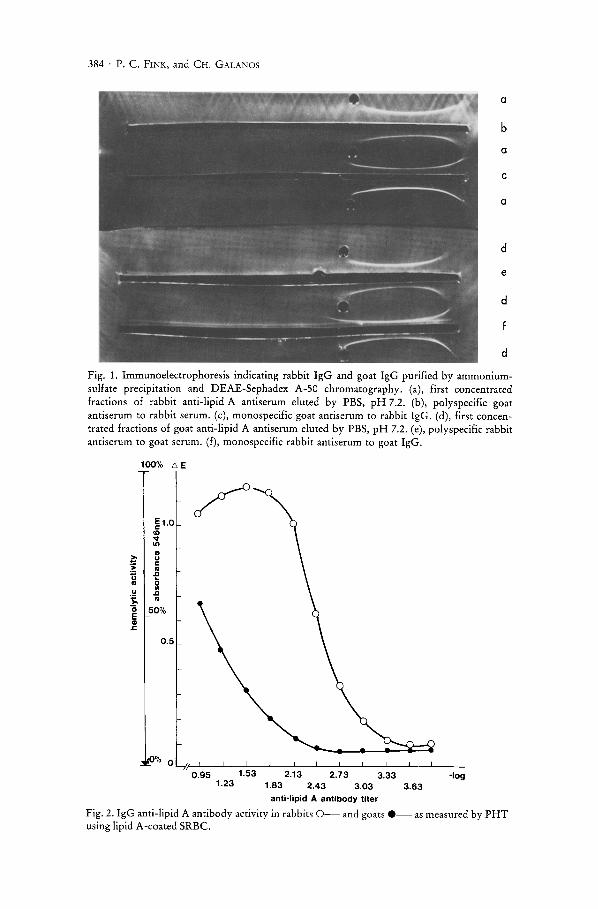
384 . P. C. FINK, and CH. GALANOS
a
b
a
c
a
d
e
d
d
Fig. 1. Immunoelectrophoresis indicating rabbit IgG and goat IgG purified by ammoniumsulfate precipitation and DEAE-Sephadex A-50 chromatography. (a), first concentrated fractions of rabbit anti-lipid A antiserum eluted by PBS, pH 7.2. (b), polyspecific goat antiserum to rabbit serum. (c), monospecific goat antiserum to rabbit IgG. (d), first concentrated fractions of goat anti-lipid A antiserum eluted by PBS, pH 7.2. (e), poly specific rabbit antiserum to goat serum. (f), monospecific rabbit antiserum to goat IgG.
~ > n II
U ; "0 E II> .t:.
100% £', E
~1.0 CD v In .. u c: II .D 0 OJ
.D II
50%
0.5
0% O~,~'-L' __ -L __ -L __ ~ __ ~ __ ~ __ ~ __ ~ __ ~ __ ~ __ ~ ____ __ 0.95 1.53
1.23 2.13
1.83 2.73
2.43 3 .33
3.03 anti-lipid A antibody titer
-109 3 .63
Fig. 2. IgG anti-lipid A antibody activity in rabbits 0- and goats .-as measured by PHT using lipid A-coated SRBC.

Anti-Lipid A and Lipid A . 385
HI---'---'----'----'----'------'------'----'----'-- ng/ml 0.5 0.1 0.05 0.01
lipid A Novo-Pyrexal forte lPS E.coli 055:85 LPS
Fig. 3. Rabbit IgG anti-lipid A antibody activity assayed by PHIT following the preincubation with lipid A *, Novo-Pyrexal forte LPS., and E. coli 055 : B5 O.
Use of IgG anti-lipid A antibodies in SAB type ELISA
In a first series of experiments, the ELISA titer of IgG anti-lipid A antibody activity from rabbits was examined using a constant lipid A concentration. As shown in Figure 4, solid-phase-coated lipid A was detectable by IgG anti-lipid A antibodies at dilutions of up to -log: 3.69. The results for a variation of SAB type ELISA using varying concentrations of solid-phase-coated lipid A are shown in Figure 5. The detection limit for lipid A was 0_25 J.lg/ml. Beyond the linear portion of the measuring range, concomitantly with increasing lipid A concentrations (0.5 J.lg to 25 f,lg/ml) a significant decrease of colouration occurred.
Use of IgG anti-lipid A antibodies in SA type ELISA
Performing the protocol of SA type ELISA lipid A could be detected in the measuring range of 0.001 J.lg to 10 f,lg/ml (Fig. 6). Thus a significantly lower detection limit (0.001 J.lg/ml) was attainable with SA than SAB type ELISA, although the SA method showed an upper detection limit of 10 J.lgi mllipid A of the measuring range.

386 . P. C. FINK, and CH. GALANOS
E c:
Iil
1.0
0.9
0.8
0.7
0.6
... 0.5
0.4
0.3
0.2
0.1
o L----''----'-_--'-_---'-_-'-_--'-_--'-_...L- -log IgG anti-lipid A 1.3 1.6 2 2.3 2.6 3 3.7 antibody titer
Fig. 4. Determinations of IgG anti-lipid A antibody activity in sera of rabbits immunized with lipid A-coated hydrolyzed bacteria carrying out SAB type ELISA.
Discussion
The present results confirm previous findings (20, 23) showing that lipid A, when exposed on the cell surface of hydrolysed Salmonella minnesota R595, bacteria produces an anti-lipid A antibody response in animals. In agreement with previous investigations (20), the IgG fraction from rabbit lipid A antisera revealed a similar degree of anti-lipid A activity in PHT under the conditions of immunization used. The IgG fraction from goat lipid A antiserum obtained at various intervals after the last booster injection showed a relatively weak anti-lipid A antibody titer in this case. The absence of reactivity between anti-lipid A antibodies and S. abortus equi or E. coli 055 : B5 intact LPS as measured by PHIT is in accordance with earlier findings (17). The lipid A specificity of IgG antibodies could be demonstrated in inhibition tests, where soluble lipid A could neutralize IgG anti-lipid A activity. Furthermore, the decrease in IgG anti-lipid A activity

E c o I/)
"'" tl c til .Q
i
Anti-Lipid A and Lipid A . 387
6E
0.8
i 0.5-
0.1
~~< __ ~-LI ____ -L __ L� ____ ~I __ LI __ ~I __ __ 0.25 0.5 2.5 5 25 50 100
!' g/ml lipid A
Fig. 5. Determination of varying concentrations of lipid A by SAB type ELISA.
observed with correspondingly increasing lipid A concentrations yields a specific antigen-antibody reaction.
Recently, radioimmunoassays measuring LPS from E. coli strains were applied for the study of experimental (15, 16) and clinical (27) material. The alternative method described utilized the ELISA technique thereby estimating quantitatively IgG anti-lipid A antibodies and lipid A. This procedure in comparison to radioimmunoassays had the advantage, that there was no contact with radioactive material.
The SAB and SA type ELISA utilizing various concentrations of lipid A showed that increase of absorbance due to the enzymic activity was proportional to only high concentration differences of lipid A. In SAB type ELISA the decrease of colouration (Fig. 5) might indicate that intermolecular aggregation forces between lipid A molecules hindered the sensitization of solid phase with lipid A. Concerning the plateau of the reaction observed with SA type ELISA (Fig. 6), it is presumed that either the amount of solid

388· P. C. FINK, and CH. GALANOS
0.8
E 0 .5 I:
~ ~
I ... •
0.1
0.001 0.01 0 .1 10 100 .ug/ml lipid A
Fig. 6. Determination of varying concentrations of lipid A by SA type ELISA.
phase sensitizing goat IgG anti-lipid A antibody or of the second rabbit IgG anti-lipid A was limited. Therefore these findings imply that further studies on lipid A quantitation are necessary.
In contrast, the SAB type ELISA proved to be a suitable method for the measurement of anti-lipid A antibody activity. It also showed to be more sensitive for measuring anti-lipid A antibody activity than PHT. The clinical application of this type of ELISA in determining anti-lipid A antibody activity might be of value in the future, since a possible role of these antibodies in protecting against bacterial infection (30) and in neutralizing endotoxic activities has been shown as inhibition of the local Shwartzman reaction and inhibition of endotoxin-induced fever (28). In addition, anti-lipid A antibodies protected pregnant mice challenged with lipid A or LPS from endotoxin-induced abortion (29). Anti-lipid A antibodies have also been found in patients with gram-negative bacterial urinary tract infections and in experimental work with dogs, where lipid A and anti-lipid A antibodies were suggested to participate among other cellular immune mechanisms on kidney damage (19). Therefore, antilipid A antibody determination by means of ELISA might be of clinical interest in the future.

Anti-Lipid A and Lipid A . 389
Acknowledgements
The skillful technical assistance of SABINE SCHUMACHER and CORDULA SERR is very much appreciated. The work was supported in part by grant Fi 281/1 from the Deutsche Forschungsgemeinschaft.
References
1. LEVIN, J., T. E. POORE, S. E. YOUNG, M. MARGOLIS, P. ZAUBA, A. S. TOWNES, and W. R. BELL. 1972. Gram-negative sepsis. Detection of endotoxemia with the Limulus test. With studies of associated changes in blood coagulation, serum lipids, and complement. Ann. Int. Med. 76: 1-7.
2. WElL, M. H., H. SCHUBIN, and M. BIDDLE. 1964. Shock caused by gram-negative microorganisms. Analysis of 169 cases. Ann. Int. Med. 60: 384-400.
3. CHRISTY, J. H. 1971. Pathophysiology of gram-negative shock. Amer. Heart J. 81: 694-701.
4. FISCHER, P., J. E. MILLEN, and F. L. GLAUSER. 1977. Endotoxin-induced increased alveolar capillary membrane permeability. Circulatory Shock 4: 387-395.
5. URBASCHEK, B., and R. M. URBASCHEK. 1979. The inflammatory response to endotoxins. In: Microcirculation in Inflammation, Ed.: Hauck, G., J. W. Irwin. Bibliotheca anat. Karger, Basel 17: pp. 74-104.
6. WARDLE, E. N. 1975. Endotoxemia and the pathogenesis of acute renal failure. Quarterly J. Med. New Series XLIV 174: 389-398.
7. NOLAN, J. P. 1975. The Role of endotoxin in liver injury. Gastroenterology 69: 1346-1356.
8. WILKINSON, S. P., H. MOODIE, J. D. STAMATAKIS, V. V. KAKKAR, and R. WILLIAMS. 1976. Endotoxemia and renal failure in cirrhosis and obstructive jaundice. Brit. Med. J. 2: 1415-1418.
9. COOPERSTOCK, M. S., R. P. TUCKER, and J. V. BAUBLIS. 1975. Possible pathogenic role of endotoxin in Reye's syndrom. Lancet 7: 1272-274.
10. FINK, P. C., L. LEHR, R. M. URBASCHEK, and J. KOZAK. 1980. Limulus amebocyte lysate test for endotoxemia: Investigations with a femtogram sensitive spectrophotometric assay. Klin. Wschr. In Press.
11. ELIN, R. J., R. A. ROBINSON, A. S. LEVINE, and S. M. WOLFF. 1975. Lack of clinical usefulness of the Limulus test in the diagnosis of endotoxemia. N. Eng!. J. Med. 293: 521-524.
12. The U.S. Pharmacopeia. 1970. Pyrogen test. Mack Publishing, Easton, PA, 18: pp. 886-887.
13. BERGMAN, R. K., K. C. MILNER, and J. J. MUNOZ. 1977. New test for endotoxin potency based upon histamin sensitization in mice. Infect. Immun. 18: 352-355.
14. FINKELSTEIN, R. A. 1964. Observations on mode of action of endotoxin in chick embryos. Proc. Soc. Exp. Bio!. Med. 115: 702-707.
15. GUTOWSKI, J. A., and D. M. JACOBS. 1979. A solid-phase radioimmunoassay for bacterial lipopolysaccharide. Immuno!. Commun. 8: 347-364.
16. KIMURA, H. 1976. Measurement of endotoxin. I. Fundamental studies on radioimmunoassay of endotoxin. Acta Med. Okayama 30: 245-255.
17. GALANOS, C., O. LODERITZ, E. T. RIETSCHEL, and O. WESTPHAL. 1977. Newer aspects of the chemistry and biology of bacteriallipopolysaccharides with special reference to their lipid A component. Int. Rev. Biochemistry of Lipids II, 14: 239-335.
18. GALANOS, c., M. A. FREUDENBERG, O. LODERITZ, E. T. RIETSCHEL, and O. WESTPHAL. 1979. Chemical, physicochemical and biological properties of bacterial lipopolysaccharides. Progr. Clin. BioI. Res. 29: 321-332.
19. WESTENFELDER, M., C. GALANOS, P. O. MADSEN, and W. MARGET. 1977. Pathological activities of lipid A: Experimental studies in relation to chronic pyelonephritis. In:

390 . P. C. FINK, and CH. GALANOS
Microbiology. Ed.: Schlessinger, D., Washington DC. Amer. Soc. Microbiol. pp. 2-7-279.
20. GALANOS, c., O. LUDERITZ, and O. WESTPHAL. 1971. Preparation and properties of antisera against the lipid A component of bacteriallipopolysaccharides. Eur. J. Biochem. 24: 116-122.
21. FINK, P. C., I. SCHEDEL, and H. DEICHER. 1976. Estimation of IgG antiglobulins in sera of patients with rheumatoid arthritis and chronic liver disorders. Klin. Wschr. 54: 601-606.
22. LAYNE, E. 1957. Spectrophotometric and turbidimetric methods for measuring proteins. In: Methods in Enzymology III, Ed.: Colowick, S. P., N. o. Kaplan. Academic Press, New York-London, pp. 447-454.
23. GALANOS, C., J. ROPPEL, J. WECKESSER, E. T. RIETSCHEL, and H. MAYER. 1977. Biological activities of lipopolysaccharides and lipid A from rhodospirillaceae. Infect. Immun. 16: 407-412.
24. OELLERICH, M. 1980. Enzyme immunoassays in clinical chemistry: Present status and trends. J. Clin. Chern. Clin. Biochem. 18: 197-208.
25. SCHUURS, A. H. W. M., and B. K. VAN WEEMAN. 1977. Enzyme immunoassay. Clin. Chim. Acta 81: 1-40.
26. KENDALL, M. G. 1955. Rank correlation methods, 2nd ed. Charles Griffin, London. 27. TAl, C. 1974. Clinical and experimental endotoxemia related to the intestinal,ischemia,
detection of endotoxin by means of radioimmunoassay. Jap. J. Med. Sci. BioI. 27: 111-114.
28. RIETSCHEL, E. T., and C. GALANOS. 1977. Lipid A antiserum-mediated protection against lipopolysaccharide and lipid A induced fever and skin necrosis. Infect. Immun. 8: 173-177.
29. RIoux-DARRIEULAT, F., M. PARANT, and L. CHEDID. 1978. Prevention of endotoxininduced abortion by treatment of mice with antisera. J. Infect. Dis. 137: 7-13.
30. GALANOS, c., M. A. FREUDENBERG, S. HASE, F. JAY, and E. RUSCHMANN. 1977. Biological activities and immunological properties of lipid A. In: Microbiology. Ed.: Schlessinger, D. Amer. Soc. Microbiol., Washington DC, pp. 269-276.
Dr. med. PETER C. FINK, Medizinische Hochschule Hannover, Institut fUr Klinische Chemie, Karl-Wiechert-Allee 9,3000 Hannover 61, Federal Republic of Germany