dasmahapatra et al. mechanitis barcoding title ... files/mechanitis...dasmahapatra et al. mechanitis...
TRANSCRIPT

DASMAHAPATRA ET AL. Mechanitis barcoding
1
Title: Mitochondrial DNA barcoding detects “species” that are not real
Kanchon K. Dasmahapatra 1
Marianne Elias 2
Ryan I Hill 3
Joseph I Hoffman 4
James Mallet 1,5
1
Department of Genetics, Evolution and Environment, University College London, 4
Stephenson Way, London NW1 2HE, U.K.
2
Institute of Evolutionary Biology, University of Edinburgh, West Mains Road, Edinburgh
EH9 3JT, U.K.
3
Department of Integrative Biology, University of California, Berkeley, CA 94720, U.S.A.
4
Department of Zoology, University of Cambridge, Downing Street, Cambridge CB2 3EJ,
U.K.
5
Wissenschaftskolleg zu Berlin, Wallotstraße 19, 14193 Berlin, Germany
Author for correspondence: Kanchon K. Dasmahapatra
Tel: +44 (0)207 6795100
Fax: +44 (0)207 6795052

DASMAHAPATRA ET AL. Mechanitis barcoding
2
SUMMARY
Mimicry and extensive geographic subspecies polymorphism combine to make species in the
ithomiine butterfly genus Mechanitis (Lepidoptera; Nymphalidae) difficult to determine. We
use mitochondrial (mtDNA) barcoding, nuclear sequences and amplified fragment length
polymorphism (AFLP) genotyping to investigate species limits in this genus. Although earlier
biosystematics studies based on morphology described only four species, mitochondrial
barcoding revealed eight well-differentiated haplogroups, indicating the presence of four new
putative “cryptic species”. However, using AFLP markers we find support for only one of the
four putative new “cryptic species” as biologically meaningful. We demonstrate that in this
genus deep genetic divisions expected on the basis of mtDNA are not always reflected in the
nuclear genome, and advocate the use of AFLP markers as a check when mtDNA barcoding
gives unexpected results.
Keywords: DNA barcoding, AFLP, Mechanitis, cryptic species, ithomiine

DASMAHAPATRA ET AL. Mechanitis barcoding
3
1. INTRODUCTION
DNA barcoding has recently been suggested as a quick method useful for rapid species
discovery and biodiversity assessment (Borisenko et al. 2008; Hajibabaei et al. 2006; Stoeckle
& Hebert 2008). For animal taxa the majority of these studies have used a short section of
mitochondrial DNA (mtDNA), namely the first ~650 bp of the 5’ end of the cytochrome
oxidase I gene (CoI) (Elias-Gutierrez et al. 2008; Hebert et al. 2003; Rock et al. 2008). DNA
barcoding has been argued to revolutionise taxonomy allowing rapid species identification
without need for detailed taxonomic expertise with increasing economy (Hajibabaei et al.
2007; Stoeckle & Hebert 2008). But the practice of mtDNA barcoding has received much
criticism on methodological (Will & Rubinoff 2004), theoretical (Hickerson et al. 2006) and
empirical grounds (Elias et al. 2007; Hurst & Jiggins 2005; Meyer & Paulay 2005; Wiemers
& Fiedler 2007). Despite the problems, undoubted successes for mtDNA barcoding have been
the discovery of cryptic species overlooked by more traditional taxonomic methods (Burns et
al. 2008; Smith et al. 2006).
Several studies have sought to overcome some of the problems inherent in mtDNA taxonomy
by supplementing mtDNA sequences with nuclear sequences (Elias et al. 2007; Monaghan et
al. 2005). However, the use of nuclear sequences for DNA taxonomy has been limited, largely
due to the difficulty in finding and sequencing fast-evolving nuclear loci (Dasmahapatra &
Mallet 2006). A possible alternative to nuclear sequence data is analysis based on amplified
fragment length polymorphisms (AFLPs), which are typically fast-evolving anonymous
nuclear markers, readily amplifiable in any organism (Mueller & Wolfenbarger 1999; Vos et
al. 1995). In this paper we first use mtDNA sequences to test species affiliations in the
difficult ithomiine butterfly genus Mechanitis (Lepidoptera; Nymphalidae), and then further
investigate these results utilising both nuclear gene sequences and AFLP markers.

DASMAHAPATRA ET AL. Mechanitis barcoding
4
Four Mechanitis species have been described and all are both abundant locally and widely
distributed in the neotropics (Brown 1977). Like most other ithomiines, all of the Mechanitis
species show multiple geographic subspecies variation and typically have orange, brown,
yellow and black wing colouration (Brown 1979; Lamas 2004). The many geographic forms
of Mechanitis, involved in Müllerian mimicry with various members of the ~360 species of
Ithomiinae as well as Heliconius species, had led to an extremely problematic species-level
taxonomy. However, careful biosystematic studies in the 1970s gave rise to a reasonably
stable classification, based on morphology, distribution, hybrid zones, and ecology (Brown
1977), that is still generally accepted (Lamas 2004). Here, we investigate the four putative
species comprising this genus.
2. MATERIAL AND METHODS
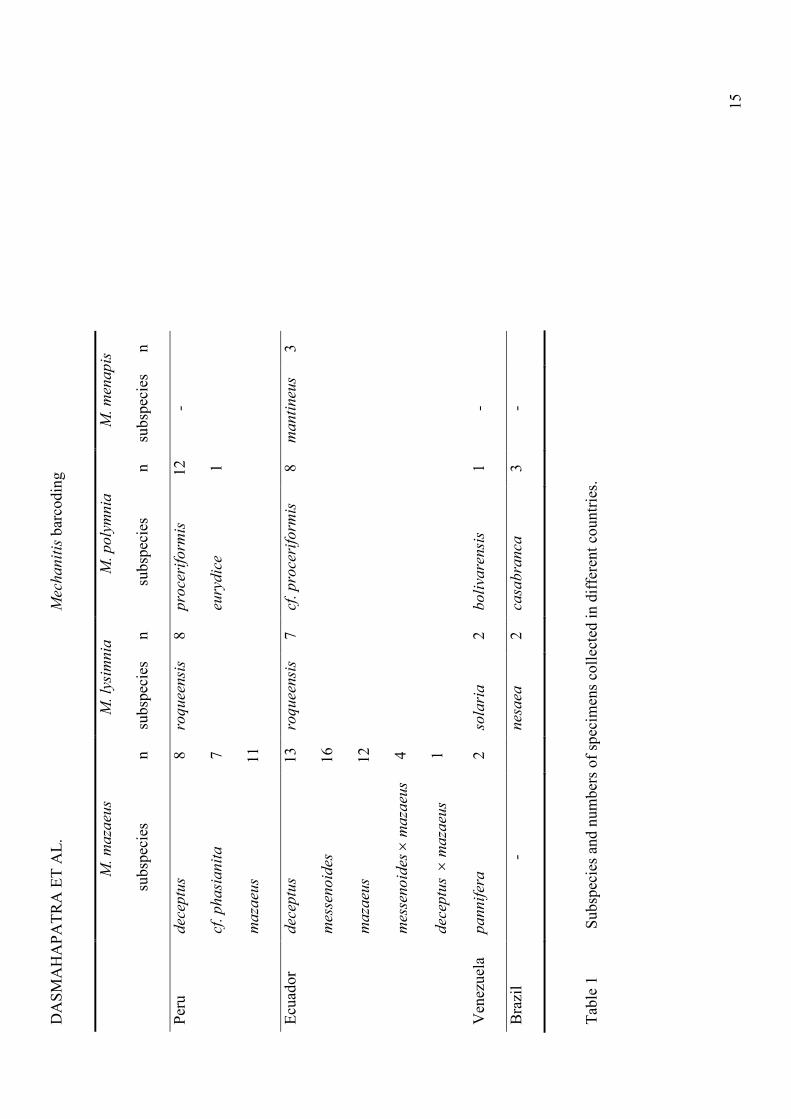
All four species of Mechanitis were used in this study: M. mazaeus (M. m. deceptus, M. m. cf.
phasianita, M. m. messenoides, M. m. pannifera, M. m. mazaeus), M. polymnia (M. p.
proceriformis, M. p. casabranca, M. p. bolivarensis, M. p. eurydice), M. lysimnia (M. l.
roqueensis, M. l. nesaea, M. l. solaria) and M. menapis (M. m. mantineus). Our samples
reflect the wide distribution of the first three species: specimens were collected from north
and central Peru (eastern San Martin and southern Loreto), Ecuador, northern Venezuela and
the Atlantic coast of Brazil (Table 1). M. menapis has a narrower distribution, replacing M.
mazaeus west of the Andes (Brown 1979) and in Central America; our M. menapis specimens
were obtained from western Ecuador. We sampled 121 specimens, mainly from Ecuador and
Peru, together with five from each of Venezuela and eastern Brazil for comparison (Table 1).
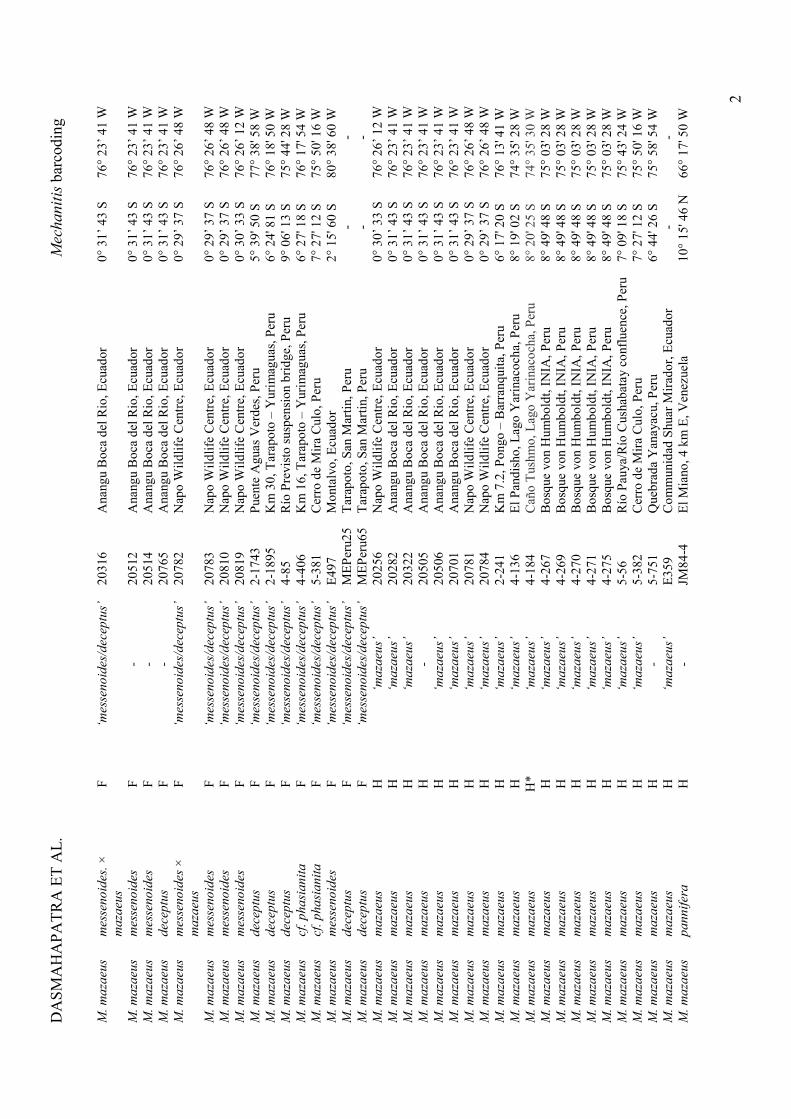
Details of sampling locations are provided in the Supplementary Materials (Table S1).

DASMAHAPATRA ET AL. Mechanitis barcoding
5
DNA was extracted from legs and thoraces using QIAamp DNA Micro and DNeasy Blood
and Tissue Kits (QIAGEN). Approximately 640 bp of mtDNA comprising the 5’-end of CoI,
the “barcoding region”, was amplified and sequenced in all specimens. To examine the effect
of using more mtDNA sequence, a further ~1500 bp, comprising the remaining 3’ portion of
CoI, the tR�A-leu gene, and the 5’ end of CoII, was also sequenced from 71 specimens
representing all four species. To examine whether the patterns revealed by mtDNA were
reflected in the nuclear genome, sequences were also obtained from three nuclear loci: Tektin
(58 sequences), Rpl5 (68 sequences) and Tpi (73 sequences) (Mallarino et al. 2005; Whinnett
et al. 2005a). For Rpl5 and Tpi these sequences included representatives of all four species,
but for Tektin no sequences were obtained for M. menapis. PCR primers and reaction
conditions have been reported previously (Dasmahapatra et al. 2007; Elias et al. 2007;
Whinnett et al. 2005a); a detailed description is also provided in Table S2. Cycle sequencing
was carried out using the Big Dye Terminator 3.1 Cycle Sequencing Kit (Applied
Biosystems). All sequences obtained for this study have been deposited in Genbank (awaiting
Accession numbers XXXXXX-XXXXXX, EU068843-EU068856, EU068966, EU068993-
EU068900, EU069070-EU069075).
Genetic divisions revealed by mitochondrial CoI may sometimes not be captured at autosomal
nuclear loci. This could be due to lack of coalescence within species given the ~4× effective
population size (�E) of autosomal loci compared to cytoplasmic mtDNA, and also because
exons typically sequenced readily across genera tend to evolve at slower rates than CoI. The
Rpl5 and Tpi loci we have used both contain fast-evolving intronic regions. In addition, Tpi is
a sex-linked locus having only ~3× �E compared to mitochondrial loci. To obtain even higher
resolution nuclear data, we genotyped 84 specimens representing all four species and all eight

DASMAHAPATRA ET AL. Mechanitis barcoding
6
mtDNA haplogroups (Figure 1a) using AFLP markers. The samples were genotyped using
four AFLP primer combinations: TaqI-CGA + EcoRI-ACA; TaqI-CAG + EcoRI-AGC; TaqI-
CAG + EcoRI-ATG; TaqI-CCA + EcoRI-ACA. AFLP primers and protocols used are
described in Madden et al. (2004). AFLP profiles were visualised by autoradiography.
Samples with aberrant AFLP profiles were discarded (Bonin et al. 2004), and to ensure
reliability, the remaining AFLP genotypes were scored by eye; 108 putative loci were
polymorphic and could be scored reliably.
Neighbour-joining (NJ) trees for each of the four sequenced loci were constructed using
MEGA 4 (Tamura et al. 2007). The optimal number of genotypic clusters indicated by the
AFLP genotypes was established with the Bayesian programme STRUCTURE 2.2 (Pritchard
et al. 2000), using standardized inference criteria (Evanno et al. 2005). Following a 100,000
step burn-in period, data were collected over 100,000 MCMC repetitions. STRUCTURE
analysis was carried out on the dataset increasing K from 1 to 10. At each value of K the
analysis was carried out three times to check between-run consistency.
3. RESULTS
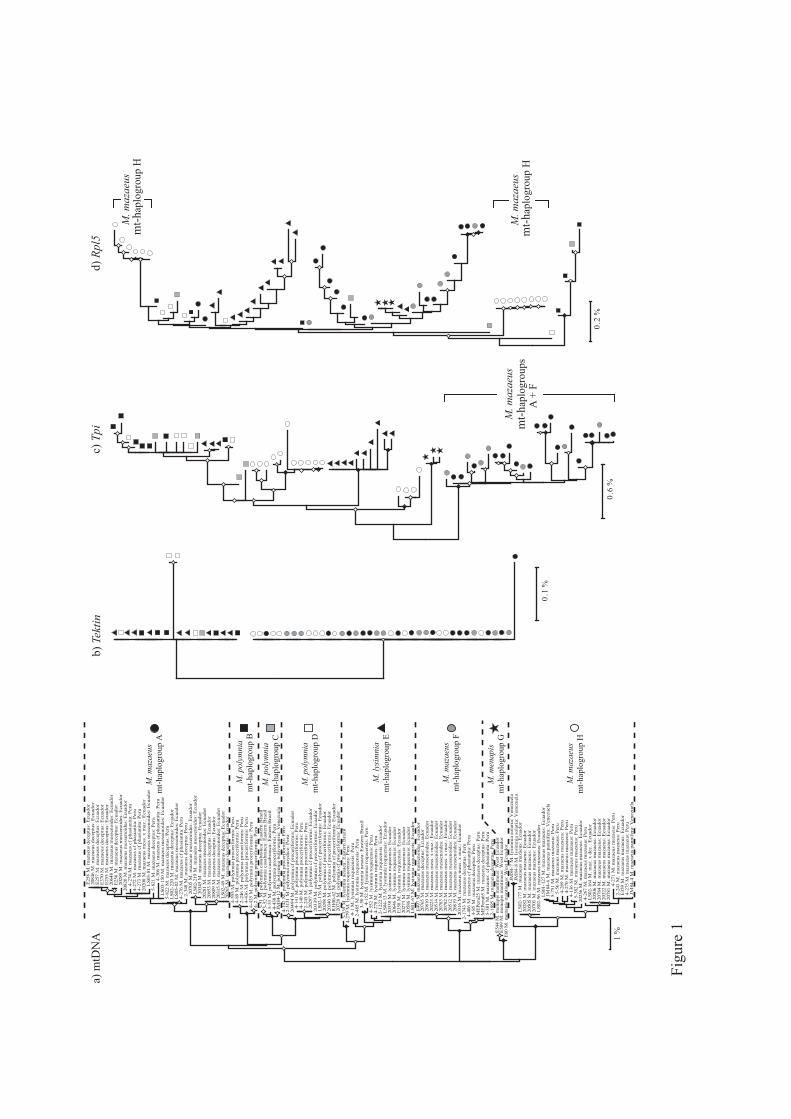
Topologies of the mtDNA NJ trees based on the 636bp CoI “barcoding” region and the full
2000bp are similar and show the same major mtDNA haplogroups. In Figure 1a we show the
former, since sequences come from a larger number of specimens. The tree shows eight major
clusters or mtDNA haplogroups: three each within M. mazaeus (mt-haplogroups A, F and H)
and M. polymnia (mt-haplogroups B, C and D), and one each corresponding to M. menapis
(mt-haplogroup G) and M. lysimnia (mt-haplogroup E). Not included in these groups are the
two specimens of M. lysimnia solaria from Venezuela that appear sister to the M. mazaeus

DASMAHAPATRA ET AL. Mechanitis barcoding
7
mt-haplogroup H, rather than lying within main M. lysimnia clade; this exception is described
further in the Discussion. The raw average pairwise distance between mt-haplogroups is
2.7%, the largest being among M. mazaeus mt-haplogroups (≥ 3.5%) and the smallest
between M. polymnia mt-haplogroups C and D (1.6%), with all other distances ≥ 2.0%. The
average intra-haplogroup variation is, in contrast, only 0.3%. Pairwise mtDNA distances
between the eight main haplogroups are shown in Table S3.
Hardly any genetic variation was present among Tektin sequences. The only clear pattern
observable is the single shared Tektin haplotype in M. lysimnia and M. polymnia, suggesting a
sister relationship (Figure 1b). In contrast, high levels of polymorphism are present at both
Tpi and Rpl5 (Figures 1c, 1d). Neither gene shows distinctions among M. polymnia mt-
haplogroups B, C and D. In the Rpl5 genealogy, there is some support for separate evolution
of M. mazaeus mt-haplogroups H from A and F, although mt-haplogroup H appears
paraphyletic. Similarly, a monophyletic M. mazaeus clade consisting of mt-haplogroups A
and F, which is distinct from M. mazaeus mt-haplogroup H, is also supported in the Tpi
genealogy, as is a separate clade corresponding to M. menapis. Overall, the nuclear gene
genealogies suggest widespread paraphyly of recognized species and mt-haplogroups.
Between-run consistency was high in the STRUCTURE analysis of AFLP genotypes:
replicate runs at each K value yielded virtually identical likelihoods. Figure 2a shows the
average likelihood from these replicates. The optimal number of groups was four (Figures 2a,
2b): two within M. mazaeus, and one each matching M. lysimnia and M. polymnia. Of the two
M. mazaeus AFLP clusters, one corresponds exactly to mt-haplogroup H (M. mazaeus
mazaeus); while the other mazaeus cluster is a mixture of mt-haplogroups A and F (M.
mazaeus deceptus, M. mazaeus cf. phasianita, and M. mazaeus messenoides), with no

DASMAHAPATRA ET AL. Mechanitis barcoding
8
evidence of subdivision between them. The M. lysimnia and M. polymnia AFLP clusters
correspond perfectly with morphology-based species designations except that one M. lysimnia
specimen shows genotypic evidence of being a hybrid between the two; and there is no
evidence for subdivision of M. polymnia along mt-haplogroup lines. There is no single AFLP
genotypic cluster associated with M. menapis; instead it appears to share genotypes largely
with M. mazaeus mt-haplogroups A + F, and to a lesser extent with M. mazaeus mt-
haplogroup H and M. lysimnia. Increasing the number of clusters to eight, the total number of
mt-haplogroups, did not alter the pattern and Figure 2b (K = 4) is virtually identical to Figure
2c (K = 8) with the four additional clusters making negligible contributions. This is further
evidence for the lack of nuclear divisions corresponding to mt-haplogroups within M.
polymnia and M. mazaeus (mt-haplogroups A and F).
4. DISCUSSION
Mitochondrial DNA barcoding of the four species in the genus Mechanitis revealed deep
genetic divisions corresponding to eight mt-haplogroups (Figure 1a). One mt-haplogroup each
corresponded to M. lysimnia and M. menapis. However three mt-haplogroups were present
within each of M. mazaeus and M. polymnia suggesting the existence of four new putative
cryptic species in addition to the four species already recognized in morphological and
biosystematic work. In contrast, while our nuclear sequence data give little resolution, our
AFLP data strongly indicate the existence of only four genetic clusters: two within M.
mazaeus and one each corresponding to M. lysimnia and M. polymnia (Figure 2b). Thus, we
obtain disparate results using mitochondrial and nuclear markers.

DASMAHAPATRA ET AL. Mechanitis barcoding
9
Mitochondrial DNA barcoding relies on intraspecific genetic variation being much less than
interspecific genetic variation. When this condition is met, a “barcoding gap” may exist
(Meyer & Paulay 2005), and clusters corresponding to genetically more homogenous entities
can be discerned. Based on such cases, some proponents of mtDNA barcoding have
advocated the use of a threshold of sequence divergence above which genetic clusters may be
considered species (Hebert et al. 2004b). Subsequent careful studies have demonstrated the
lack of a barcoding gap in a number of taxa when sampling of species, populations, and
individuals within the study group is thorough (Elias et al. 2007; Meyer & Paulay 2005;
Wiemers & Fiedler 2007). In the case of Mechanitis, we have sampled all four described
species in the genus. While we have not included all of the many geographic subspecies of
these species, we have used specimens collected across a wide geographic area representing
opposite ends of the species’ geographic distribution in S. America. With our current
sampling, mitochondrial data show eight major genetic clusters or haplogroups, clearly
separated from one another by a large average genetic distance of 2.7 %. We found no overlap
between intra- and inter-cluster genetic variation.
Only four of the eight mtDNA divisions are reflected in the nuclear genome. Evidence from
nuclear sequence information is weak, very likely a result of incomplete lineage sorting due to
recency of origin coupled with large effective population sizes within these very widespread
and common species. However, even multilocus AFLP genotyping fails to detect groups
corresponding to three of the four novel mt-haplogroups. The M. menapis samples do not
form their own AFLP genotypic cluster, instead appearing to share most alleles with the two
allopatric M. mazaeus clusters, and as such these samples are associated with a particular
genetic pattern (Figure 2b). However, no such patterns are associated with any of the three M.
polymnia mt-haplogroups or the M. mazaeus mt-haplogroups A and F.

DASMAHAPATRA ET AL. Mechanitis barcoding
10
Two possibilities might explain the mismatch between mt-haplogroups and AFLP genotypic
clusters. First, that some of the mt-haplogroups are not “real species,” which we take to mean
taxa that can maintain multilocus genetic, ecological, behavioural, and/or morphological
differences in sympatry. Second, the AFLP markers may not be sensitive enough to detect
small differences between real but closely related species detected by mtDNA barcoding.
Several lines of evidence strongly indicate that the former is more likely. Detailed
examination of the wing patterns of the specimens in the three M. polymnia mt-haplogroups
as well as M. mazaeus mt-haplogroups A and F failed to reveal any correlation between wing
patterning and mt-haplogroup. In contrast, every AFLP genetic cluster detected by
STRUCTURE is correlated with wing phenotype (discussed below). This suggests that while
the AFLP clusters are biologically relevant, some of the mitochondrial divisions are not.
AFLP markers provide a nuclear, multilocus, genome-wide picture of genetic divergence and
as they sample non-coding variation, they have relatively rapid rates of evolution. This
sensitivity is routinely exploited to reveal population genetic patterns within single species
(Baus et al. 2005; Chaput-Bardy et al. 2008; Takami et al. 2004). The sensitivity of AFLP
markers for detecting small genetic differences relative to mtDNA is also demonstrated by
results from a parallel study on butterfly species within the ithomiine genus Melinaea. In this
genus there are at least six clearly distinguishable morphological species (Melinaea satevis,
M. menophilus, M. marsaeus, M. idae, M. mneme, M. isocomma) which cannot be detected
using mtDNA barcoding (Elias et al. 2007; Whinnett et al. 2005b), Dasmahapatra et al. in
prep.). Yet where mtDNA barcoding failed to detect distinct groups, the same AFLP primer
combinations used in this paper were able to confirm morphology-based species divisions
(Dasmahapatra et al. in prep.). In contrast to the genome-wide picture obtained from AFLP
markers, mtDNA reflects evolution only of the maternally inherited mitochondrial genome,

DASMAHAPATRA ET AL. Mechanitis barcoding
11
which can be affected by factors such as interspecific hybridisation, and selection, such as via
maternally transmitted endosymbionts like Wolbachia (Hurst & Jiggins 2005). As such,
genome-wide genetic clustering revealed by AFLP markers is likely to be more generally
useful for discovery of “real species” than mtDNA barcoding.
Previous detailed morphological and biosystematic work (Brown 1977; Lamas 2004)
recognised four species within Mechanitis (M. lysimnia, M. polymnia, M. mazaeus and M.
menapis) each with multiple geographic subspecies. The nuclear data reported here strongly
support monophyletic M. lysimnia and M. polymnia clades. However, paraphyly of M.
lysimnia is suggested by two mtDNA sequences of Venezuelan M. lysimnia solaria (Figure
1a). Unfortunately, it was not possible to genotype these specimens using AFLPs as the DNA
came from old dried specimens and was degraded. This split within M. lysimnia may be
correlated with chromosome numbers (Brown et al. 2004), and a group of M. lysimnia
subspecies including M. l. solaria might represent a separate species from M. lysimnia sensu
stricto. Future investigation of phylogenetic relationships within Mechanitis should focus on
this apparent division within M. lysimnia.
M. mazaeus is also paraphyletic, and two genotypic clusters are supported by nuclear data,
one comprising mt-haplogroup H and the other combining mt-haplogroups A and F. M.
mazaeus mt-haplogroups A and F corresponds to melanic forms currently considered to
represent subspecies Mechanitis mazaeus messenoides (Colombia south to Ecuador) and
deceptus (Ecuador southwards), both of which are generally found at mid elevations on the
eastern slopes of the Andes. They are mimetic of melanic sympatric subspecies and races of
Melinaea marsaeus, Melinaea isocomma, Heliconius numata, and other species (Brown 1977;
Brown 1979). Although there is some evidence for intermediate colour patterns (Brown 1977

DASMAHAPATRA ET AL. Mechanitis barcoding
12
and Table 1), most specimens in M. mazaeus mt-haplogroup H correspond to paler lowland
M. mazaeus (sensu stricto) involved in mimicking the generalized lowland ithomiine and
heliconiine tiger patterns. We conclude that these “mazaeus” forms are best considered full
species, as their distributions overlap extensively in the Eastern foothills of the Andes (Brown
1977) with little evidence of hybridization from AFLP loci. Therefore, M. mazaeus mt-
haplogroups A and F should probably now regain species level designation as M. messenoides
(including M. messenoides messenoides C. & R. Felder 1865, M. messenoides cf. phasianita
and M. messenoides deceptus Butler 1873). The nuclear evidence points to the fourth species,
M. menapis, being a trans-Andean form close to M. mazaeus and M. messenoides, since it
shares genotypes with both cis-Andean M. mazaeus forms described above. M. menapis is not
sympatric with M. mazaeus, and instead replaces it west of the Andes and in Central America.
Morphologically based biosystematic work (Brown 1977) has shown itself to be more useful
in this genus than mtDNA barcoding. Brown (1977) and Lamas (2004) accepted four species
on the basis of morphology, a result largely upheld by our multilocus nuclear analysis. The
only major discrepancy is that intermediate colour patterns in Ecuador (Table 1) apparently
indicating hybridization between upland M. mazaeus deceptus/messenoides, and lowland
mazaeus (sensu stricto) led Brown (1977) to lump the highland melanic forms incorrectly as
subspecies of mazaeus. In comparison, mtDNA barcoding reveals eight clear monophyletic
mt-haplogroups within Mechanitis, three of which are not detected in our nuclear analysis,
suggesting that they do not correspond to “real species.” Mitochondrial DNA barcoding
provides a very sensitive technique to find new taxa, but a downside is that such taxa may
have no basis in biological reality.

DASMAHAPATRA ET AL. Mechanitis barcoding
13
Focussing on the butterfly genus Mechanitis we have used nuclear sequences and sensitive
AFLP genotyping to demonstrate how deep genetic divisions in mtDNA are not always
reflected by corresponding divisions in the nuclear genome. Such “cryptic species” clusters
may instead represent locally divergent populations that have undergone a bottleneck, rather
than having speciated in the sense of producing divergent populations that can coexist.
Putative cryptic species detected by mtDNA barcoding merit closer investigation via analysis
of nuclear genetic data, or more in-depth examination of ecology and taxonomy e.g. (Hebert
et al. 2004a; Smith et al. 2006). Owing to their ease of amplification across taxa, sensitivity to
small genetic differences and genome-wide coverage, we advocate the use of AFLP markers
in cases where mtDNA barcoding reveals unexpected results, such as a failure to recover
known taxonomic divisions or the presence of additional “cryptic” taxa. Traditional mtDNA
sequencing was always a useful way of investigating possible cryptic biological groupings
long before “DNA barcoding” was first promoted (Hebert et al. 2003). The simple argument
that mtDNA barcoding may be used in accurate species discovery, or even in identification of
known species, ignores the crucial differences between mtDNA and supermarket barcodes. In
a supermarket, each product type has a unique barcode, shared by all other examples of that
type. In organisms, there is no such unique barcode, and a one-to-one correspondence
between mtDNA sequence and species does not exist.
Acknowledgements
We are grateful to Gerardo Lamas of the Museo de Historia Natural, UNMSM, Lima, Peru,
and Keith Willmott from the Florida Museum of Natural History, Gainesville, USA, for
tireless lepidopteran expertise. Fraser Simpson assisted with many aspects of this project.
Collecting permits and support were provided by INRENA (Peru) and the Ministerio del
Ambiente and the Museo Ecuatoriano de Ciencias Naturales (Ecuador). Logistic support in

DASMAHAPATRA ET AL. Mechanitis barcoding
14
Ecuador was given by the Napo Wildlife Centre and La Selva Jungle Lodge. We thank
NERC, DEFRA–Darwin, the Leverhulme Trust and the Margaret C. Walker fund for teaching
and research in systematic entomology for funding different aspects of this project.
DA
SM
AH
AP
AT
RA
ET
AL
.
Mec
hanit
is b
arco
din
g
15
M
. m
aza
eus
M. ly
sim
nia
M
. poly
mnia
M
. m
enapis
su
bsp
ecie
s n
subsp
ecie
s n
subsp
ecie
s n
subsp
ecie
s n
Per
u
dec
eptu
s
cf. phasi
anit
a
maza
eus
8
7
11
roquee
nsi
s 8
pro
ceri
form
is
eury
dic
e
12
1
-
Ecu
ador
dec
eptu
s
mes
senoid
es
maza
eus
mes
senoid
es ×
maza
eus
dec
eptu
s ×
maza
eus
13
16
12
4
1
roquee
nsi
s 7
cf. pro
ceri
form
is
8
manti
neu
s 3
Ven
ezuel
a pannif
era
2
sola
ria
2
boli
vare
nsi
s 1
-
Bra
zil
-
nes
aea
2
casa
bra
nca
3
-
Tab
le 1
S
ubsp
ecie
s an
d n
um
ber
s of
spec
imen
s co
llec
ted i
n d
iffe
rent
countr
ies.

DASMAHAPATRA ET AL. Mechanitis barcoding
16
REFERENCES
Baus, E., Darrock, D. J., & Bruford, M. W. 2005 Gene-flow patterns in Atlantic and
Mediterranean populations of the Lusitanian sea star Asterina gibbosa. Mol. Ecol. 14,
3373-3382.
Bonin, A., Bellemain, E., Eidesen, P. B., Pomparon, F., Brochmann, C., & Taberlet, P. 2004
How to track and assess genotyping errors in population genetics studies. Mol. Ecol.
13, 3261-3273.
Borisenko, A. V., Lim, B. K., Ivanova, N. V., Hanner, R. H., & Hebert, P. D. N. 2008 DNA
barcoding in surveys of small mammal communities: a field study in Suriname.
Molecular Ecology Resources 8, 471-479.
Brown, K. S. 1977 Geographical patterns of evolution in Neotropical Lepidoptera:
differentiation of the species of Melinaea and Mechanitis (Nymphalidae, Ithomiinae).
Syst. Entomol. 2, 161-197.
Brown, K. S. 1979 Ecologia Geográfica e Evolução nas Florestas �eotropicais. Campinas,
Brazil: Universidade Estadual de Campinas.
Brown, K. S., von Schoultz, B., & Suomalainen, E. 2004 Chromosome evolution in
neotropical Danainae and Ithomiinae (Lepidoptera). Hereditas 141, 216-236.
Chaput-Bardy, A., Lemaire, C., Picard, D., & Secondi, J. 2008 In-stream and overland
dispersal across a river network influences gene flow in a freshwater insect,
Calopteryx splendens. Mol. Ecol. 17, 3496-3505.
Dasmahapatra, K. K. & Mallet, J. 2006 DNA barcodes: recent successes and future prospects.
Heredity 97, 254-255.

DASMAHAPATRA ET AL. Mechanitis barcoding
17
Dasmahapatra, K. K., Silva, A., Chung, J.-W., & Mallet, J. 2007 Genetic analysis of a wild-
caught hybrid between non-sister Heliconius butterfly species. Biol. Lett. 3, 360-363.
Elias, M., Hill, R. I., Willmott, K. R., Dasmahapatra, K. K., Brower, A. V. Z., Mallet, J., &
Jiggins, C. D. 2007 Limited performance of DNA barcoding in a diverse community
of tropical butterflies. Proc. Roy. Soc. Lond. B 274, 2881-2889.
Elias-Gutierrez, M., Jeronimo, F. M., Ivanova, N. V., Valdez-Moreno, M., & Hebert, P. D. N.
2008 DNA barcodes for Cladocera and Copepoda from Mexico and Guatemala,
highlights and new discoveries. Zootaxa, 1-42.
Evanno, G., Regnaut, S., & Goudet, J. 2005 Detecting the number of clusters of individuals
using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611-2620.
Hajibabaei, M., Janzen, D. H., Burns, J. M., Hallwachs, W., & Hebert, P. D. N. 2006 DNA
barcodes distinguish species of tropical Lepidoptera. Proc. �atl. Acad. Sci. U. S. A.
103, 968-971.
Hajibabaei, M., Singer, G. A. C., Hebert, P. D. N., & Hickey, D. A. 2007 DNA barcoding:
how it complements taxonomy, molecular phylogenetics and population genetics.
Trends Genet. 23, 167-172.
Hebert, P. D. N., Cywinska, A., Ball, S. L., & deWaard, J. R. 2003 Biological identifications
through DNA barcodes. Proc. Roy. Soc. Lond. B 270, 313-321.
Hebert, P. D. N., Penton, E. H., Burns, J. M., Janzen, D. H., & Hallwachs, W. 2004a Ten
species in one: DNA barcoding reveals cryptic species in the neotropical skipper
butterfly Astraptes fulgerator. Proc. �atl. Acad. Sci. U. S. A. 101, 14812-14817.

DASMAHAPATRA ET AL. Mechanitis barcoding
18
Hebert, P. D. N., Stoeckle, M. Y., Zemlak, T. S., & Francis, C. M. 2004b Identification of
birds through DNA barcodes. PLOS Biology 2, 10.
Hickerson, M., Meyer, C. P., & Moritz, C. 2006 DNA barcoding will often fail to discover
new animal species over broad parameter space. Syst. Biol. 55, 729-739.
Hurst, G. D. D. & Jiggins, F. M. 2005 Problems with mitochondrial DNA as a marker in
population, phylogeographic, and phylogenetic studies: the effects of inherited
symbionts. Proc. Roy. Soc. Lond. B 272, 1525-1534.
Lamas, G. 2004 Ithomiinae. In Atlas of �eotropical Butterflies. Checklist: 4A. Hesperioidea -
Papilionoidea (ed. G. Lamas), pp. 172-191. Gainesville: Scientific Publishers.
Madden, J. R., Lowe, T. J., Fuller, H. V., Coe, R. L., Dasmahapatra, K. K., Amos, W., &
Jury, F. 2004 Neighbouring male spotted bowerbirds are not related, but do maraud
each other. Anim. Behav. 68, 751-758.
Mallarino, R., Bermingham, E., Willmott, K. R., Whinnett, A., & Jiggins, C. D. 2005
Molecular systematics of the butterfly genus Ithomia (Lepidoptera: Ithomiinae): a
composite phylogenetic hypothesis based on seven genes. Mol. Phylogenet. Evol. 34,
625-644.
Meyer, C. P. & Paulay, G. 2005 DNA barcoding: error rates based on comprehensive
sampling. PLOS Biology 3.
Monaghan, M. T., Balke, M., Ryan Gregory, T., & Vogler, A. P. 2005 DNA-based species
delineation in tropical beetles using mitochondrial and nuclear markers. Phil. Trans.
Roy. Soc. Lond. B 360, 1925-1933.

DASMAHAPATRA ET AL. Mechanitis barcoding
19
Mueller, U. G. & Wolfenbarger, L. L. 1999 AFLP genotyping and fingerprinting. Trends
Ecol. Evol. 14, 389-394.
Pritchard, J. K., Stephens, M., & Donnelly, P. 2000 Inference of population structure using
multilocus genotype data. Genetics 155, 945-959.
Rock, J., Costa, F. O., Walker, D. D., North, A. W., Hutchinson, W. F., & Carvalho, G. R.
2008 DNA barcodes of fish of the Scotia Sea, Antarctica indicate priority groups for
taxonomic and systematics focus. Antarctic Science 20, 253-22.
Smith, M. A., Woodley, N. E., Janzen, D. H., Hallwachs, W., & Hebert, P. D. N. 2006 DNA
barcodes reveal cryptic host-specificity within the presumed polyphagous members of
a genus of parasitoid flies (Diptera: Tachinidae). Proc. �atl. Acad. Sci. U. S. A. 103,
3657-3662.
Stoeckle, M. Y. & Hebert, P. D. N. 2008 Bar code of life: DNA tags help classify life. Sci.
Amer.
Takami, Y., Koshio, C., Ishii, M., Fujii, H., Hidaka, T., & Shimizu, I. 2004 Genetic diversity
and structure of urban populations of Pieris butterflies assessed using amplified
fragment length polymorphism. Mol. Ecol. 13, 245-258.
Tamura, K., Dudley, J., Nei, M., & Kumar, S. 2007 MEGA4: Molecular Evolutionary
Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24, 1596-1599.
Vos, P., Hogers, R., Bleeker, M., Reijans, M., van de Lee, T., Hornes, M., Frijters, A., Pot, J.,
Peleman, J., Kuiper, M., & Zabeau, M. 1995 AFLP: a new technique for DNA
fingerprinting. �ucl. Acids Res. 23, 4407-4414.

DASMAHAPATRA ET AL. Mechanitis barcoding
20
Whinnett, A., Brower, A. V. Z., Lee, M. M., Willmott, K. R., & Mallet, J. 2005a Phylogenetic
utility of Tektin, a novel region for inferring systematic relationships amongst
Lepidoptera. Ann. Entomol. Soc. Amer. 98, 873-886.
Whinnett, A., Zimmermann, M., Willmott, K. R., Herrera, N., Mallarino, R., Simpson, F.,
Joron, M., Lamas, G., & Mallet, J. 2005b Strikingly variable divergence times inferred
across an Amazonian butterfly 'suture zone'. Proc. Roy. Soc. B 272, 2525-2533.
Wiemers, M. & Fiedler, K. 2007 Does the DNA barcoding gap exist? - a case study in blue
butterflies (Lepidoptera: Lycaenidae). Frontiers in Zoology 4.
Will, K. W. & Rubinoff, D. 2004 Myth of the molecule: DNA barcodes for species cannot
replace morphology for identification and classification. Cladistics 20, 47-55.

DASMAHAPATRA ET AL. Mechanitis barcoding
21
FIGURE CAPTIONS
Figure 1
Neighbour-joining trees constructed from (a) mitochondrial sequences and (b, c and d)
nuclear sequences, showing relationships between the eight Mechanitis mitochondrial
haplogroups. Nodes marked with filled and open symbols have > 90 % and > 50 % (and < 90
%) bootstrap support respectively. Scale bars represent raw percentage sequence divergence.
(dec. = deceptus; maz. = mazaeus; mess. = messenoides)
Figure 2
STRUCTURE analysis of AFLP genotypes. a) Log likelihood of data as a function of K, the
number of clusters. Likelihoods from each of the three replicate runs at each K are
indistinguishable and the average likelihood is shown. Highest likelihood is achieved with
four clusters, even though eight mt-haplogroups are present. STRUCTURE results for b) the
optimal number of genotypic clusters, K = 4, and c) the number of mt-haplogroups, K = 8.
Each of the 85 individuals is represented by a vertical bar broken into K coloured segments.
The length of each colour in the bar indicates the posterior mean proportion of ancestry from
each genetic clusters. b) and c) are virtually identical, indicating the absence of any significant
additional genetic structure for more than four clusters.
SHORT TITLE: Mechanitis barcoding
0.1
%
b)
Tek
tin
0.2
%
d)
Rp
l5
M.
ma
zaeu
sm
t-h
aplo
gro
up
H
M.
ma
zaeu
sm
t-h
aplo
gro
up
H
0.6
%
c) T
pi
M.
ma
zaeu
s
mt-
hap
log
rou
ps
A +
F
E258 M
. m
azaeus
deceptu
s: E
cuador
20846 M
. m
azaeus
deceptu
s: E
cuador
E257 M
. m
azaeus
deceptu
s: E
cuador
E370 M
. m
azaeus
deceptu
s: E
cuador
E255 M
. m
azaeus
deceptu
s: E
cuador
20648 M
. m
azaeus
mess
enoid
es:
Ecuador
E256 M
. m
azaeus
deceptu
s: E
cuador
20289 M
. m
azaeus
mess
enoid
es:
Ecuador
20820 M
. m
azaeus
mess
enoid
es:
Ecuador
4-2
74 M
. m
azaeus
cf
phasi
anit
a:
Peru
4-2
72 M
. m
azaeus
cf
phasi
anit
a:
Peru
4-2
73 M
. m
azaeus
cf
phasi
anit
a:
Peru
20208 M
. m
azaeus
mess
. x m
az.:
Ecuador
LS
03-8
1 M
. m
azaeus
mess
enoid
es:
Ecuador
5-8
21 M
. m
azaeus
deceptu
s: P
eru
4-3
06 M
. m
azaeus
cf
phasi
anit
a:
Peru
LS
03-1
50 M
. m
azaeus
mess
enoid
es:
Ecuador
5-8
22 M
. m
azaeus
deceptu
s: P
eru
LS
02-2
20 M
. m
azaeus
deceptu
s: E
cuador
LS
05-0
2 M
. m
azaeus
mess
enoid
es:
Ecuador
4-2
76 M
. m
azaeus
cf
phasi
anit
a:
Peru
2-3
48 M
. m
azaeus
deceptu
s: P
eru
20705 M
. m
azaeus
mess
enoid
es:
Ecuador
LS
02-2
19 M
. m
azaeus
mess
enoid
es:
Ecuador
20345 M
. m
azaeus
deceptu
s: E
cuador
20281 M
. m
azaeus
mess
enoid
es:
Ecuador
20853 M
. m
azaeus
deceptu
s: E
cuador
20809 M
. m
azaeus
deceptu
s: E
cuador
20315 M
. m
azaeus
mess
enoid
es:
Ecuador
LS
02-4
9 M
. m
azaeus
deceptu
s: E
cuador
20265 M
. m
azaeus
deceptu
s: E
cuador
4-5
55 M
. poly
mnia
pro
ceri
form
is:
Peru
4-8
9 M
. poly
mnia
pro
ceri
form
is:
Peru
2-2
46 M
. poly
mnia
pro
ceri
form
is:
Peru
4-4
36 M
. poly
mnia
pro
ceri
form
is:
Peru
4-4
35 M
. poly
mnia
pro
ceri
form
is:
Peru
4-2
M. poly
mnia
pro
ceri
form
is:
Peru
3-5
1 M
. poly
mnia
casa
bra
nca:
East
ern
Bra
zil
3-5
2 M
. poly
mnia
casa
bra
nca:
East
ern
Bra
zil
3-5
3 M
. poly
mnia
casa
bra
nca:
East
ern
Bra
zil
4-4
38 M
. poly
mnia
pro
ceri
form
is:
Peru
JM
84-1
2 M
. poly
mnia
boli
vare
nsi
s: V
enezuela
4-1
42 M
. poly
mnia
pro
ceri
form
is:
Peru
2-3
131 M
. poly
mnia
eury
dic
e:
Peru
4-1
41 M
. poly
mnia
pro
ceri
form
is:
Peru
4-1
40 M
. poly
mnia
pro
ceri
form
is:
Peru
2-2
45 M
. poly
mnia
pro
ceri
form
is:
Peru
20297 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
LS
02-1
76 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
20298 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
20340 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
RH
00-6
2 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
20728 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
3-1
39 M
. ly
sim
nia
nesa
ea:
East
ern
Bra
zil
4-2
79 M
. ly
sim
nia
roqueensi
s: P
eru
4-1
M. ly
sim
nia
roqueensi
s: P
eru
2-4
85 M
. ly
sim
nia
roqueensi
s: P
eru
3-5
0 M
. ly
sim
nia
nesa
ea:
East
ern
Bra
zil
4-1
52 M
. ly
sim
nia
roqueensi
s: P
eru
4-5
52 M
. ly
sim
nia
roqueensi
s: P
eru
4-2
78 M
. ly
sim
nia
roqueensi
s: P
eru
2-1
222 M
. ly
sim
nia
roqueensi
s: E
cuador
LS
04-1
3 M
. ly
sim
nia
roqueensi
s: E
cuador
20338 M
. ly
sim
nia
roqueensi
s: E
cuador
20646 M
. ly
sim
nia
roqueensi
s: E
cuador
E358 M
. ly
sim
nia
roqueensi
s: E
cuador
20273 M
. ly
sim
nia
roqueensi
s: E
cuador
20586 M
. ly
sim
nia
roqueensi
s: E
cuador
LS
01-6
7 M
. ly
sim
nia
roqueensi
s: E
cuador
E497 M
. m
azaeus
mess
enoid
es:
Ecuador
20765 M
. m
azaeus
deceptu
s: E
cuador
20810 M
. m
azaeus
mess
enoid
es:
Ecuador
20223 M
. m
azaeus
mess
. x m
az.:
Ecuador
20514 M
. m
azaeus
mess
enoid
es:
Ecuador
20783 M
. m
azaeus
mess
enoid
es:
Ecuador
20782 M
. m
azaeus
mess
. x m
az.:
Ecuador
20512 M
. m
azaeus
mess
enoid
es:
Ecuador
20819 M
. m
azaeus
mess
enoid
es:
Ecuador
20316 M
. m
azaeus
mess
. x m
az.:
Ecuador
2-1
743 M
. m
azaeus
deceptu
s: P
eru
4-4
06 M
. m
azaeus
cf
phasi
anit
a:
Peru
4-8
5 M
. m
azaeus
deceptu
s: P
eru
ME
Per
u25 M
. m
azaeus
deceptu
s: P
eru
ME
Per
u65 M
. m
azaeus
deceptu
s: P
eru
5-3
81 M
. m
azaeus
cf
phasi
anit
a:
Peru
2-1
895 M
. m
azaeus
deceptu
s: P
eru
E544 M
. m
enapis
manti
neus:
West
Ecuador
E589 M
. m
enapis
manti
neus:
West
Ecuador
E60 M
. m
enapis
manti
neus:
West
Ecuador
JM
84-7
M. ly
sim
nia
sola
ria:
Venezuela
JM
84-8
M. ly
sim
nia
sola
ria:
Venezuela
LS
02-1
77 M
. m
azaeus
mazaeus:
Ecuador
20282 M
. m
azaeus
mazaeus:
Ecuador
20505 M
. m
azaeus
mazaeus:
Ecuador
20701 M
. m
azaeus
mazaeus:
Ecuador
LS
02-9
6 M
. m
azaeus
mazaeus:
Ecuador
LS
01-1
27 M
. m
azaeus
mazaeus:
Ecuador
JM
84-6
M. m
azaeus
pannif
era
: V
enezuela
5-7
51 M
. m
azaeus
mazaeus:
Peru
5-5
6 M
. m
azaeus
mazaeus:
Peru
4-2
69 M
. m
azaeus
mazaeus:
Peru
4-2
70 M
. m
azaeus
mazaeus:
Peru
4-1
36 M
. m
azaeus
mazaeus:
Peru
5-3
82 M
. m
azaeus
mazaeus:
Peru
20256 M
. m
azaeus
mazaeus:
Ecuador
4-2
67 M
. m
azaeus
mazaeus:
Peru
LS
02-1
64 M
. m
az. x d
ec.:
Ecuador
20506 M
. m
azaeus
mazaeus:
Ecuador
20784 M
. m
azaeus
mazaeus:
Ecuador
20322 M
. m
azaeus
mazaeus:
Ecuador
20781 M
. m
azaeus
mazaeus:
Ecuador
4-2
71 M
. m
azaeus
mazaeus:
Peru
2-2
41 M
. m
azaeus
mazaeus:
Peru
E359 M
. m
azaeus
mazaeus:
Ecuador
4-2
75 M
. m
azaeus
mazaeus:
Peru
JM
84-4
M. m
azaeus
pannif
era
: V
enezuela
1 %
M. m
aza
eus
mt-
hap
logro
up A
M. poly
mnia
mt-
hap
logro
up B
M. poly
mnia
mt-
hap
logro
up C
M. m
aza
eus
mt-
hap
logro
up H
M. m
enapis
M. m
aza
eus
mt-
hap
logro
up F
M. ly
sim
nia
mt-
hap
logro
up E
mt-
hap
logro
up G
a) m
tDN
A
20339 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
20494 M
. poly
mnia
cf
pro
ceri
form
is:
Ecuador
M. poly
mnia
mt-
hap
logro
up D
Fig
ure
1
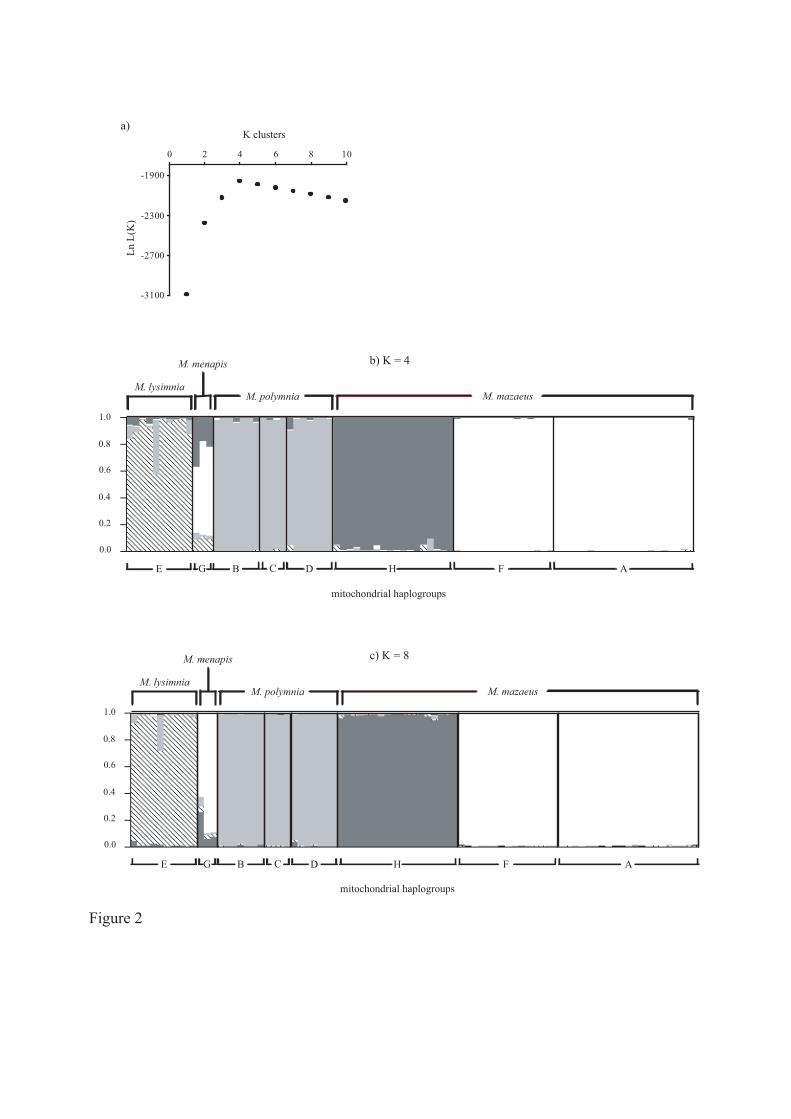
Figure 2
M. lysimnia
M. menapis
M. polymnia M. mazaeus
B AFHD
0.0
1.0
0.2
0.4
0.6
0.8
CE G
mitochondrial haplogroups
b) K = 4
0.0
1.0
0.2
0.4
0.6
0.8
mitochondrial haplogroups
c) K = 8
-3100
-2700
-2300
-1900
0 2 4 6 8 10
K clustersL
n L
(K)
a)
B AFHDCE G
M. lysimnia
M. menapis
M. polymnia M. mazaeus
DA
SM
AH
AP
AT
RA
ET
AL
.
M
echanit
is b
arco
din
g
1
SU
PP
LE
ME
NT
AR
Y M
AT
ER
IAL
Sp
ecie
s S
ub
spec
ies
(by w
ing p
atte
rn)
mt-
hap
logro
up
A
FL
P g
eno
typ
ic c
lust
er
Sp
ecim
en
ID
Co
llec
tio
n l
oca
lity
L
atit
ud
e L
on
git
ud
e
M. m
aza
eus
mes
senoid
es ×
maza
eus
A
‘mes
senoid
es/d
ecep
tus’
2
02
08
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
31
' 45
S
76
° 2
5' 0
1 W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
2
02
65
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
30
' 33
S
76
° 2
6’
12
W
M. m
aza
eus
mes
senoid
es
A
‘mes
senoid
es/d
ecep
tus’
2
02
81
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
mes
senoid
es
A
‘mes
senoid
es/d
ecep
tus’
2
02
89
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
mes
senoid
es
A
‘mes
senoid
es/d
ecep
tus’
2
03
15
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
2
03
45
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
mes
senoid
es
A
‘mes
senoid
es/d
ecep
tus’
2
06
48
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
30
’ 3
3 S
7
6°
26
’ 1
2 W
M. m
aza
eus
mes
senoid
es
A
‘mes
senoid
es/d
ecep
tus’
2
07
05
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
2
08
09
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
29
’ 3
7 S
7
6°
26
’ 4
8 W
M. m
aza
eus
mes
senoid
es
A
‘mes
senoid
es/d
ecep
tus’
2
08
20
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
30
’ 3
3 S
7
6°
26
’ 1
2 W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
2
08
46
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
2
08
53
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
2
-34
8
Po
ngo
de
Caynar
achi,
Per
u
6°
20
' 7 S
7
6°
15
' 15
W
M. m
aza
eus
cf. phasianita
A
‘mes
senoid
es/d
ecep
tus’
4
-27
2
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
cf. phasianita
A
‘mes
senoid
es/d
ecep
tus’
4
-27
3
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
cf. phasianita
A
‘mes
senoid
es/d
ecep
tus’
4
-27
4
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
cf. phasianita
A
‘mes
senoid
es/d
ecep
tus’
4
-27
6
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
cf. phasianita
A
‘mes
senoid
es/d
ecep
tus’
4
-30
6
Km
7.2
, P
ongo
– B
arra
nq
uit
a, P
eru
6°
17
' 20
S
76
° 1
3' 4
1 W
M. m
aza
eus
dec
eptu
s A
-
5-8
21
Q
ueb
rad
a Y
anayac
u,
Per
u
6°
44
' 26
S
75
° 5
8' 5
4 W
M. m
aza
eus
dec
eptu
s A
-
5-8
22
Q
ueb
rad
a Y
anayac
u,
Per
u
6°
44
' 26
S
75
° 5
8' 5
4 W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
E
25
5
Po
do
carp
us
Nat
ional
Rese
rve,
Ecu
ado
r 4
° 0
6' 8
0 S
7
8°
57
' 90
W
M. m
aza
eus
dec
eptu
s A
-
E2
56
P
odo
carp
us
Nat
ional
Rese
rve,
Ecu
ado
r 4
° 0
6' 8
0 S
7
8°
57
' 90
W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
E
25
7
Po
do
carp
us
Nat
ional
Rese
rve,
Ecu
ado
r 4
° 0
6' 8
0 S
7
8°
57
' 90
W
M. m
aza
eus
dec
eptu
s A
‘m
esse
noid
es/d
ecep
tus’
E
25
8
Po
do
carp
us
Nat
ional
Rese
rve,
Ecu
ado
r 4
° 0
6' 8
0 S
7
8°
57
' 90
W
M. m
aza
eus
dec
eptu
s A
-
E3
70
S
ucua,
Ecu
ado
r 2
° 2
7' 3
5 S
7
8°
10
' 11
W
M. m
aza
eus
dec
eptu
s A
-
LS
02
-49
G
arza
Co
cha,
Sucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
aza
eus
mes
senoid
es
A
- L
S0
2-2
19
G
arza
Co
cha,
Sucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
aza
eus
dec
eptu
s A
-
LS
02
-22
0
Gar
za C
och
a, S
ucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
aza
eus
mes
senoid
es
A
- L
S0
3-8
1
Gar
za C
och
a, S
ucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
aza
eus
mes
senoid
es
A
- L
S0
3-1
50
G
arza
Co
cha,
Sucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
aza
eus
mes
senoid
es
A
- L
S0
5-0
2
San
i Is
la,
Ecuad
or
0°
30
’ 4
1 S
7
6°
21
’ 0
4 W
M. m
aza
eus
mes
senoid
es
× m
aza
eus
F
‘mes
senoid
es/d
ecep
tus’
2
02
23
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
30
’ 3
3 S
7
6°
26
’ 1
2 W
DA
SM
AH
AP
AT
RA
ET
AL
.
M
echanitis
bar
codin
g
2
M. m
aza
eus
mes
senoid
es. ×
maza
eus
F
‘mes
senoid
es/d
ecep
tus’
2
03
16
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
mes
senoid
es
F
- 2
05
12
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
mes
senoid
es
F
- 2
05
14
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
dec
eptu
s F
-
20
765
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
mes
senoid
es ×
maza
eus
F
‘mes
senoid
es/d
ecep
tus’
2
07
82
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
29
’ 3
7 S
7
6°
26
’ 4
8 W
M. m
aza
eus
mes
senoid
es
F
‘mes
senoid
es/d
ecep
tus’
2
07
83
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
29
’ 3
7 S
7
6°
26
’ 4
8 W
M. m
aza
eus
mes
senoid
es
F
‘mes
senoid
es/d
ecep
tus’
2
08
10
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
29
’ 3
7 S
7
6°
26
’ 4
8 W
M. m
aza
eus
mes
senoid
es
F
‘mes
senoid
es/d
ecep
tus’
2
08
19
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
30
’ 3
3 S
7
6°
26
’ 1
2 W
M. m
aza
eus
dec
eptu
s
F
‘mes
senoid
es/d
ecep
tus’
2
-17
43
P
uen
te A
guas
Ver
des
, P
eru
5°
39
' 50
S
77
° 3
8' 5
8 W
M. m
aza
eus
dec
eptu
s
F
‘mes
senoid
es/d
ecep
tus’
2
-18
95
K
m 3
0,
Tar
apo
to –
Yuri
mag
uas
, P
eru
6°
24
' 81
S
76
° 1
8' 5
0 W
M. m
aza
eus
dec
eptu
s
F
‘mes
senoid
es/d
ecep
tus’
4
-85
R
ío P
revis
to s
usp
ensi
on b
rid
ge,
Per
u
9°
06
' 13
S
75
° 4
4' 2
8 W
M. m
aza
eus
cf. phasianita
F
‘mes
senoid
es/d
ecep
tus’
4
-40
6
Km
16
, T
arap
oto
– Y
uri
mag
uas
, P
eru
6°
27
' 18
S
76
° 1
7' 5
4 W
M. m
aza
eus
cf. phasianita
F
‘mes
senoid
es/d
ecep
tus’
5
-38
1
Cer
ro d
e M
ira
Culo
, P
eru
7°
27
' 12
S
75
° 5
0' 1
6 W
M. m
aza
eus
mes
senoid
es
F
‘mes
senoid
es/d
ecep
tus’
E
49
7
Mo
nta
lvo
, E
cuad
or
2°
15
' 60
S
80
° 3
8' 6
0 W
M. m
aza
eus
dec
eptu
s F
‘m
esse
noid
es/d
ecep
tus’
M
EP
eru2
5
Tar
apo
to, S
an M
arti
n,
Per
u
- -
M. m
aza
eus
dec
eptu
s F
‘m
esse
noid
es/d
ecep
tus’
M
EP
eru6
5
Tar
apo
to, S
an M
arti
n,
Per
u
- -
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
02
56
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
30
’ 3
3 S
7
6°
26
’ 1
2 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
02
82
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
03
22
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
maza
eus
H
- 2
05
05
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
05
06
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
07
01
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
07
81
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
29
’ 3
7 S
7
6°
26
’ 4
8 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
07
84
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
29
’ 3
7 S
7
6°
26
’ 4
8 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
2
-24
1
Km
7.2
, P
ongo
– B
arra
nq
uit
a, P
eru
6°
17
' 20
S
76
° 1
3' 4
1 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
4
-13
6
El
Pan
dis
ho
, L
ago
Yar
inac
och
a, P
eru
8°
19
' 02
S
74
° 3
5' 2
8 W
M. m
aza
eus
maza
eus
H*
‘maza
eus’
4
-18
4
Cañ
o T
ush
mo
, L
ago
Yar
inac
och
a, P
eru
8°
20
' 25
S
74
° 3
5' 3
0 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
4
-26
7
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
4
-26
9
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
4
-27
0
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
4
-27
1
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
4
-27
5
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
5
-56
R
ío P
auya/
Río
Cush
abat
ay c
onfl
uence
, P
eru
7°
09
' 18
S
75
° 4
3' 2
4 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
5
-38
2
Cer
ro d
e M
ira
Culo
, P
eru
7°
27
' 12
S
75
° 5
0' 1
6 W
M. m
aza
eus
maza
eus
H
- 5
-75
1
Queb
rad
a Y
anayac
u,
Per
u
6°
44
' 26
S
75
° 5
8' 5
4 W
M. m
aza
eus
maza
eus
H
‘maza
eus’
E
35
9
Co
mm
unid
ad S
huar
Mir
ado
r, E
cuad
or
- -
M. m
aza
eus
pannifer
a
H
- JM
84
-4
El
Mia
no
, 4
km
E,
Ven
ezuel
a
10
° 1
5' 4
6 N
6
6°
17
' 50
W

DA
SM
AH
AP
AT
RA
ET
AL
.
M
echanitis
bar
codin
g
3
M. m
aza
eus
pannifer
a
H
- JM
84
-6
El
Mia
no
, 4
km
E,
Ven
ezuel
a
10
° 1
5' 4
6 N
6
6°
17
' 50
W
M. m
aza
eus
maza
eus
H
- L
S0
1-1
27
G
arza
Co
cha,
Sucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
aza
eus
maza
eus
H
- L
S0
2-9
6
San
i Is
la,
Ecuad
or
0°
30
’ 4
1 S
7
6°
21
’ 0
4 W
M. m
aza
eus
maza
eus ×
dec
eptu
s
H
- L
S0
2-1
64
G
arza
Co
cha,
Sucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
aza
eus
maza
eus
H
- L
S0
2-1
77
G
arza
Co
cha,
Sucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. ly
sim
nia
so
laria
(H)
- JM
84
-7
Cu
manac
oa
- C
oco
kar
, K
m 1
7,
Ven
ezuel
a 1
0°
14
' 20
N
63
° 5
5' 1
5 W
M. ly
sim
nia
so
laria
(H)
- JM
84
-8
Cu
manac
oa
- C
oco
kar
, K
m 1
7,
Ven
ezuel
a 1
0°
14
' 20
N
63
° 5
5' 1
5 W
M. poly
mnia
pro
cerifo
rmis
B
‘poly
mnia
’ 2
-24
6
Km
7.2
, P
ongo
– B
arra
nq
uit
a, P
eru
6°
17
' 20
S
76
° 1
3' 4
1 W
M. poly
mnia
pro
cerifo
rmis
B
‘poly
mnia
’ 4
-2
Cas
till
o G
rand
e, T
ingo
Mar
ia,
Per
u
9°
16
' 11
S
76
° 0
0' 2
9 W
M. poly
mnia
pro
cerifo
rmis
B
‘poly
mnia
’ 4
-89
R
ío P
revis
to s
usp
ensi
on b
rid
ge,
Per
u
9°
06
' 13
S
75
° 4
4' 2
8 W
M. poly
mnia
pro
cerifo
rmis
B*
‘poly
mnia
’ 4
-21
2
Km
59
, P
uca
llp
a -
Tin
go
Mar
ia,
Per
u
8°
38
' 28
S
74
° 5
7' 1
3 W
M. poly
mnia
pro
cerifo
rmis
B
‘poly
mnia
’ 4
-43
5
Km
22
, N
uev
o L
ima
- S
elva
And
ina,
Per
u
7°
11
' 36
S
76
° 2
0' 0
6 W
M. poly
mnia
pro
cerifo
rmis
B
‘poly
mnia
’ 4
-43
6
Km
22
, N
uev
o L
ima
- S
elva
And
ina,
Per
u
7°
11
' 36
S
76
° 2
0' 0
6 W
M. poly
mnia
pro
cerifo
rmis
B
‘poly
mnia
’ 4
-55
5
Km
22
.5,
Nuev
o L
ima
- L
a P
erla
del
Po
nac
illo
, P
eru
7°
10
' 56
S
76
° 1
0' 5
2 W
M. poly
mnia
ca
sabra
nca
C
‘p
oly
mnia
’ 3
-51
C
am
pin
as,
Bra
zil
22
° 4
9' 3
4 S
4
7°
4' 4
W
M. poly
mnia
ca
sabra
nca
C
‘p
oly
mnia
’ 3
-52
C
am
pin
as,
Bra
zil
22
° 4
9' 3
4 S
4
7°
4' 4
W
M. poly
mnia
ca
sabra
nca
C
‘p
oly
mnia
’ 3
-53
C
am
pin
as,
Bra
zil
22
° 4
9' 3
4 S
4
7°
4' 4
W
M. poly
mnia
pro
cerifo
rmis
C
‘poly
mnia
’ 4
-43
8
Km
22
, N
uev
o L
ima
- S
elva
And
ina,
Per
u
7°
11
' 36
S
76
° 2
0' 0
6 W
M. poly
mnia
boliva
rensis
C
- JM
84
-12
E
l M
iano
, 4
km
E,
Ven
ezuel
a
10
° 1
5' 4
6 N
6
6°
17
' 50
W
M. poly
mnia
cf
. pro
cerifo
rmis
D
‘poly
mnia
’ 2
03
40
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. poly
mnia
cf
. pro
cerifo
rmis
D
‘poly
mnia
’ 2
07
28
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. poly
mnia
cf
. pro
cerifo
rmis
D
- 2
02
97
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. poly
mnia
cf
. pro
cerifo
rmis
D
- 2
02
98
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. poly
mnia
cf
. pro
cerifo
rmis
D
- 2
03
39
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. poly
mnia
cf
. pro
cerifo
rmis
D
- 2
04
94
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. poly
mnia
pro
cerifo
rmis
D
- 2
-24
5
Km
7.2
, P
ongo
– B
arra
nq
uit
a, P
eru
6°
17
' 20
S
76
° 1
3' 4
1 W
M. poly
mnia
eu
rydic
e D
‘p
oly
mnia
’ 2
-31
31
P
alm
a R
eal,
Per
u
12
° 3
7' 1
8 S
7
2°
41
' 25
W
M. poly
mnia
pro
cerifo
rmis
D
‘poly
mnia
’ 4
-14
0
El
Pan
dis
ho
, L
ago
Yar
inac
och
a, P
eru
8°
19
' 02
S
74
° 3
5' 2
8 W
M. poly
mnia
pro
cerifo
rmis
D
‘poly
mnia
’ 4
-14
1
El
Pan
dis
ho
, L
ago
Yar
inac
och
a, P
eru
8°
19
' 02
S
74
° 3
5' 2
8 W
M. poly
mnia
pro
cerifo
rmis
D
‘poly
mnia
’ 4
-14
2
El
Pan
dis
ho
, L
ago
Yar
inac
och
a, P
eru
8°
19
' 02
S
74
° 3
5' 2
8 W
M. poly
mnia
cf
. pro
cerifo
rmis
D
- L
S0
2-1
76
G
arza
Co
cha,
Sucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. poly
mnia
cf
. pro
cerifo
rmis
D
- R
H0
0-6
2
Gar
za C
och
a, S
ucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. ly
sim
nia
ro
quee
nsis
E
- 2
02
73
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. ly
sim
nia
ro
quee
nsis
E
‘lys
imnia
’ 2
05
86
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. ly
sim
nia
ro
quee
nsis
E
- 2
06
46
N
apo
Wil
dli
fe C
entr
e, E
cuad
or
0°
30
’ 3
3 S
7
6°
26
’ 1
2 W
M. ly
sim
nia
ro
quee
nsis
E
‘lys
imnia
’ 2
03
38
A
nang
u B
oca
del
Rio
, E
cuad
or
0°
31
’ 4
3 S
7
6°
23
’ 4
1 W
M. ly
sim
nia
ro
quee
nsis
E
- 2
-48
5
Km
7.2
, P
ongo
– B
arra
nq
uit
a, P
eru
6°
17
' 20
S
76
° 1
3' 4
1 W
DA
SM
AH
AP
AT
RA
ET
AL
.
M
echanitis
bar
codin
g
4
M. ly
sim
nia
ro
quee
nsis
E
- 2
-12
22
K
m 8
, T
arap
oto
– Y
uri
maguas,
Per
u
6°
27
' 48
S
76
° 1
9' 2
0 W
M. ly
sim
nia
nes
aea
E
‘lys
imnia
’ 3
-50
C
am
pin
as,
Bra
zil
22
° 4
9' 3
4 S
4
7°
04
' 04
W
M. ly
sim
nia
nes
aea
E
‘lys
imnia
’ 3
-13
9
Ser
ra B
onit
a, B
razi
l 1
5°
25
' 18
S
39
° 3
0' 1
2 W
M. ly
sim
nia
ro
quee
nsis
E
‘lys
imnia
’ 4
-1
Cas
till
o G
rand
e, T
ingo
Mar
ia,
Per
u
9°
16
' 11
S
76
° 0
0' 2
9 W
M. ly
sim
nia
ro
quee
nsis
E*
‘lys
imnia
’ 4
-12
8
Río
Pre
vis
to s
usp
ensi
on b
rid
ge,
Per
u
9°
06
' 13
S
75
° 4
4' 2
8 W
M. ly
sim
nia
ro
quee
nsis
E
- 4
-15
2
Cañ
o T
ush
mo
, L
ago
Yar
inac
och
a, P
eru
8°
20
' 25
S
74
° 3
5' 3
0 W
M. ly
sim
nia
ro
quee
nsis
E
‘lys
imnia
’ 4
-27
8
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. ly
sim
nia
ro
quee
nsis
E
‘lys
imnia
’ 4
-27
9
Bo
sque
vo
n H
um
bo
ldt,
IN
IA,
Per
u
8°
49
' 48
S
75
° 0
3' 2
8 W
M. ly
sim
nia
ro
quee
nsis
E
‘lys
imnia
’ 4
-55
2
Km
22
.5,
Nuev
o L
ima
- L
a P
erla
del
Po
nac
illo
, P
eru
7°
10
' 56
S
76
° 1
0' 5
2 W
M. ly
sim
nia
ro
quee
nsis
E
‘lys
imnia
’ E
35
8
Co
mm
unid
ad S
huar
Mir
ado
r, E
cuad
or
M. ly
sim
nia
ro
quee
nsis
E
- L
S0
1-6
7
Gar
za C
och
a, S
ucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. ly
sim
nia
ro
quee
nsis
E
- L
S0
4-1
3
Gar
za C
och
a, S
ucu
mb
ios,
Ecu
ado
r 0
° 2
9’
47
S
76
° 2
2’
25
W
M. m
enapis
m
antineu
s G
m
ixed
E
60
M
ind
o,
Ecu
ado
r 0
° 1
' 60
S
78
° 4
7' 6
0 W
M. m
enapis
m
antineu
s G
m
ixed
E
54
4
Lo
s C
edro
s, E
cuad
or
0°
19
' 48
N
78
° 4
8' 3
6 W
M. m
enapis
mantineu
s G
m
ixed
E
58
9
Lo
s C
edro
s, E
cuad
or
0°
19
' 48
N
78
° 4
8' 3
6 W
Tab
le S
1
Det
ails
of
sam
ple
s an
d s
ampli
ng l
oca
liti
es
* O
nly
a s
hort
er l
ength
of
mtD
NA
seq
uen
ce (
~560
bp)
could
be
obta
ined
fo
r th
ese
indiv
idual
s. T
his
was
suff
icie
nt
for
mt-
hap
logro
up
clas
sifi
cati
on, but
thes
e sp
ecim
ens
are
not
rep
rese
nte
d i
n t
he
mtD
NA
NJ
tree
(F
igu
re 1
a).

DA
SM
AH
AP
AT
RA
ET
AL
.
M
echanitis
bar
codin
g
5
Am
pli
con
Pri
mer
s P
rim
er s
equen
ces
(5’-
3’)
T
a
/ ºC
MgC
l 2
(mM
)
dN
TP
s
(mM
)
mtD
NA
bar
code
Lep
3.1
f a
Lep
3.1
r a
TC
TA
CA
AA
TC
AT
AA
AG
AT
AT
TG
GA
AC
AT
T
AA
AT
TT
TA
AT
TC
CT
GT
TG
GT
AC
AG
C
55
2.3
0.2
mtD
NA
CoI
Jerr
y b
Pat
lep a
CA
AC
AT
TT
AT
TT
TG
AT
TT
TT
TG
G
TC
CA
TT
AC
AT
AT
AA
TC
TG
CC
AT
AT
TA
57
2.3
0.2
mtD
NA
CoII
Geo
rgei
th *
Evai
th *
TA
GG
WT
TA
GC
WG
GA
AT
AC
C
GA
GA
CC
AA
TA
CT
TG
CT
TT
CA
GA
CA
TC
T
58
2.3
0.2
Tpi
Jess
*
Nab
*
AG
CA
AN
CA
GA
TW
AA
TG
AA
AT
CR
TY
A
TA
GC
CA
YT
TA
CG
NA
GA
GA
NG
C
63
– 5
6
3.0
0.8
Tek
tin c
tek3
teka
CG
CA
GT
TT
YT
GA
TR
CT
YT
AC
CA
GT
GG
RG
AY
AT
YC
TW
GG
57
2.7
5
0.6
8
Rpl5
d
Rpl5
-F44
Rpl5
-R441
TC
CG
AC
TT
TC
AA
AC
AA
GG
AT
G
CA
TA
TC
CT
GG
GA
AT
CT
CT
TG
AT
G
55
2.3
0.2
Tab
le S
2.
PC
R c
ondit
ions
for
the
ampli
cons
use
d i
n t
his
stu
dy.
All
rea
ctio
ns
wer
e ca
rrie
d o
ut
in 1
0µ
l volu
mes
usi
ng 1
0×
bu
ffer
(S
igm
a), 0.5
µM
eac
h o
f fo
rwar
d a
nd r
ever
se p
rim
ers
and 0
.25U
Taq p
oly
mer
ase
(Sig
ma)
. F
or Tpi
0.1
6 µ
g/m
l B
SA
was
als
o a
dded
. P
CR
cycl
ing c
ondit
ions
wer
e: i
nit
ial
den
aturi
ng a
t 94ºC
for
2 m
inute
s; f
oll
ow
ed b
y 3
5 c
ycl
es
of
94ºC
for
45s,
annea
ling t
emper
ature
(T
a) f
or
45
s, 7
2ºC
for
60s;
fin
al e
xte
nsi
on a
t 72ºC
for
5 m
inute
s. F
or Tpi,
PC
R c
ycl
ing c
ondit
ions
wer
e
the
sam
e ex
cept
that
a t
ouch
dow
n p
roce
dure
was
use
d w
ith T
a dro
ppin
g f
rom
63ºC
to 5
6ºC
over
14 c
ycl
es, fo
llow
ed b
y a
furt
her
23 c
ycl
es a
t
56ºC
. * T
hes
e pri
mer
s w
ere
dev
eloped
for
this
stu
dy. Tpi
pri
mer
s Je
ss a
nd N
ab, w
ere
dev
eloped
by A
lain
e W
hin
net
t.
a (D
asm
ahap
atra
et
al. 20
07);
b (
Sim
on e
t al
. 1994);
c (W
hin
net
t et
al.
2005a)
; d (
Mal
lari
no e
t al
. 2005)

DA
SM
AH
AP
AT
RA
ET
AL
.
M
echanitis
bar
codin
g
6
Sp
ecie
s
M. ly
sim
nia
M
. m
enapis
M. poly
mnia
M
. m
aza
eus
mt-
hap
logro
up
E
G
B
C
D
A
F
H
M. ly
sim
nia
E
0.6
2
.3
2.4
2
.3
2.0
2
.8
2.8
3
.1
M. m
enapis
G
0.1
2
.5
2.5
2
.1
2.9
2
.1
2.6
M. poly
mnia
B
0.2
2
.3
2.2
2
.8
3.0
3
.3
C
0.2
1
.6
2.7
3
.3
3.3
D
0.2
2
.7
2.9
3
.2
M. m
aza
eus
A
0.6
3
.6
4.1
F
0.4
3
.5
H
0.5
Tab
le S
3. R
aw p
erce
nta
ge
pai
rwis
e dis
tance
s bet
wee
n m
tDN
A h
aplo
gro
ups.
Intr
a-hap
logro
up d
ista
nce
s ar
e it
alic
ised
.