current knowledge about the presence of amines in wine
TRANSCRIPT

This article was downloaded by: [University of Regina]On: 22 August 2014, At: 20:53Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Critical Reviews in Food Science and NutritionPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/bfsn20
Current Knowledge about the Presence of Amines inWineCarmen Ancín-Azpilicueta a , Ana González-Marco a & Nerea Jiménez-Moreno aa Department of Applied Chemistry , Universidad Pública de Navarra , Campus de Arrosadías/n, E-31006, Pamplona, SpainPublished online: 14 Feb 2008.
To cite this article: Carmen Ancín-Azpilicueta , Ana González-Marco & Nerea Jiménez-Moreno (2008) CurrentKnowledge about the Presence of Amines in Wine, Critical Reviews in Food Science and Nutrition, 48:3, 257-275, DOI:10.1080/10408390701289441
To link to this article: http://dx.doi.org/10.1080/10408390701289441
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Critical Reviews in Food Science and Nutrition, 48:257–275 (2008)
Copyright C©© Taylor and Francis Group, LLC
ISSN: 1040-8398
DOI: 10.1080/10408390701289441
Current Knowledge about thePresence of Amines in Wine
CARMEN ANCıN-AZPILICUETA∗, ANA GONZALEZ-MARCO,and NEREA JIMENEZ-MORENO
Department of Applied Chemistry, Universidad Publica de Navarra, Campus de Arrosadıa s/n, E-31006 Pamplona, Spain
This review discusses those factors that have an influence on amine concentration in wine, as well as the vinification stageswhen these amines are formed and their evolution during the storage of the product. It also outlines the importance of thesebiogenic and volatile amines both in the sensorial aspect as well as the toxic action. Amines are nitrogenous bases of lowmolecular weight, which are especially found in food and beverages from the fermentation process, as is the case of wine.Amine concentration in wine can be very variable, ranging from trace levels up to 130 mg/L, so it is difficult to predict thecontent of these compounds in the product. At present this variability is of some concern in many countries, especially for thetoxic effect that some amines can have on people who are susceptible to these substances. Consequently, it is necessary todevelop new studies to learn more about the factors, which have an influence on their concentration in wine, and to establishlimits for these substances in the product so as to prevent any toxic effects on the consumer.
Keywords biogenic amines, volatile amines, toxic action, wines
INTRODUCTION
Amines are organic nitrogenous bases of low molecularweight which are formed during the metabolic processes of liv-ing organisms and which are found especially in foods from fer-mentative processes. Depending on their volatility, these com-pounds can be divided into non-volatile amines and volatileamines. The former virtually coincide with biogenic amines,except in the case of phenylethylamine, which is a biogenicvolatile amine. Biogenic amines mainly come from the decar-boxylation of amino acids (ten Brink et al., 1990). Figure 1shows the precursor amino acids of biogenic amines in wine.Some biogenic amines found in wine such as histamine, tyra-mine and phenylethylamine have toxic effects on the organ-ism (Radler and Fath, 1991). These substances are metabo-lized in the liver and the intestine by the action of two en-zymatic complexes, monoamine oxidase (MAO) and diamineoxidase (DAO). Ethanol and acetaldehyde, compounds foundin wine, are inhibitors of MAO and DAO. Consequently, theyincrease the toxic effect of the biogenic amines (ten Brinket al., 1990). The volatile amines have been much less stud-ied than the non-volatile ones and, with the exception ofphenylethylamine, it seems that they come from the amina-tion of non-nitrogenous compounds such as aldehydes and ke-
∗Author to whom correspondence should be addressed Carmen Ancın-Azpilicueta, Telephone: (34) 948-168909; Fax: (34) 948-169606; e-mail:[email protected]
tones found in wine (Ough et al., 1981). These compoundscould have an unfavorable effect on wine aroma, although thestudies carried out on their threshold levels have been madeon beer.
The total content of amines in wine vary from tracelevels up to 130 mg/L (Soufleros et al., 1998). Among thesenitrogenous substances putrescine is normally the most abun-dant (Soufleros et al., 1998; Lethonen, 1996). The non-volatile amines found most frequently in wine are histamine,tyramine, putrescine, cadaverine, spermine, spermidine, al-though agmatine, tryptamine and serotonine, have also beensporadically found (Lethonen, 1996; Lehtonen et al., 1992;Busto et al., 1995). The volatile amines found in wineare, among others, phenylethylamine, methylamine, dimethy-lamine, ethylamine, diethylamine, isopropylamine, isobuty-lamine, amylamine, pyrrolidine, ethanolamine, and hexy-lamine (Ough et al., 1981; Busto et al., 1995; Ough andDaudt, 1981).
Amines are formed in wine during different vinificationstages. Formation of amines has been observed during alcoholicfermentation due to the metabolism of different yeast strains(Buteau et al., 1984; Vidal-Carou and Marine-Font, 1985; Torreaand Ancın, 2002). An increase in the concentration of thesecompounds has also been observed during malolactic fermen-tation due to the proliferation of lactic acid bacteria (Aerny,1985; Bauza et al., 1995; Gerbaux and Monamy, 2000). Be-sides, a great variability in the concentration of amines existsbetween different wines as there are many factors which have
257
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

258 C. ANCıN-AZPILICUETA ET AL.
Figure 1 Precursor amino acids of biogenic amines.
an influence, to a greater or lesser extent, in the formation ofthese substances. Among these factors, the content of nitroge-nous compounds in the grape stands out which depends, amongother factors, on the level of maturation and on the nitroge-nous fertilization of the soil. Another important factor is themethod of winemaking employed, since red wines show a largerconcentration of amines compared to the rose or white wines(Radler and Fath, 1991; Zee et al., 1983). Table 1 shows theamine concentration in wines from different types and differ-ent geographical origin. The wine generally contains a greaterconcentration of amines than some foods such as fresh veg-etables and yogurt. However, the concentration of these sub-stances in wine is lower than in other fermented foods such ascheese (Table 2).
At present, the great variability in the concentration of thesesubstances in wine is a source of concern in many countries aswell as the toxic effect of the amines that could affect some
Table 1 Concentration of amines (mg/L) in different wines
Type of wine Histamine Cadaverine Putrescine Tyramine 2Phenylethylamine Spermine Ethylamine Reference
Red Spanishwines
0.00–25.00 (R) 0.00–14.00 (R) 0.00–55.00 (R) 0.00–19.00 (R) 0.00–16.26 (R) — 0.00–7.92 (R) Marcobal et al.(2006)
YoungChardonnaywine
0.67 (A) 0.51 (A) 3.04 (A) 2.09 (A) — 0.85 (A) — Gonzalez-Marcoand Ancın-Azpilicueta(2006)
Red qualityTurkish wines
nd–1.97 (R) nd–3.94 (R) nd–5.92 (R) nd–0.29 (R) nd–3.87 (R) nd–1.75 (R) — Anli et al. (2004)
Chardonnay winefrom inoculatedfermentations
0.31–0.36 (R) < 0.12 0.37–0.39 (R) < 0.18 2.48–2.89∗(R) 1.14–1.58 (R) 0.81–1.00 (R) Torrea and Ancın(2002)
Rioja young redwines
8.72 (A) 0.61 (A) 32.97 (A) 4.98 (A) — — — Vazquez-Lasa et al.(1998)
Rioja crianza redwines
6.67 (A) 1.74 (A) 31.35 (A) 5.78 (A) — — — Vazquez-Lasa et al.(1998)
Tokaj wines 0.02–0.04 (M) 0.07–0.19 (M) 2.14–2.55 (M) 0.05–0.31 (M) 0.15–3.09 (M) — — Hajos et al. (2000)Red, white, rose
French wines8.9 (A) 0.2 (A) 7.7 (A) 3.1 (A) 1.7 (A) 0.1 (A) Bauza et al. (1995)
A: average; M: means; R: range.∗ Phenylethylamine + Spermidine.
people who are particularly sensitive to the action of these com-pounds. In Europe, a lack in community legislation about thepermitted levels of amines in wine affects the import and exportof this product. The aim of this paper is to review the differ-ent factors, which influence the formation of amines in wine,and to study the formation of amines during both alcoholic andmalolactic fermentations. In addition, the evolution of these sub-stances during aging and conservation of wine is considered aswell as the enological aspects of amines, both from their toxicfeatures as well as their sensorial ones. With this informationit should be possible to have a better control of the vinificationprocess, avoiding the formation of these compounds in wineand to establish future limits with regard to the content of thesesubstances in the product.
FACTORS THAT INFLUENCE THE CONCENTRATION
OF AMINES IN WINE
As was stated earlier, the total content of amines in wineis very variable. This variability is due to the fact that numer-ous factors could influence the formation of biogenic amines.Some of these factors could have an indirect influence dueto a modification in the concentration of precursor aminoacids of the biogenic amines in the medium. Other factorscould modify the development of microorganisms with amino-genic capacity. Outlined below, the factors, which influencethe concentration of amines in wine, are described. It is worthpointing out that all these factors are interrelated and conse-quently it is difficult to know the individual action of eachone of them. Table 3 shows the variability in the contentof histamine, tyramine, putrescine, and phenylethylamine, de-pending on the different factors that affect the vinificationprocess.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4
CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 259
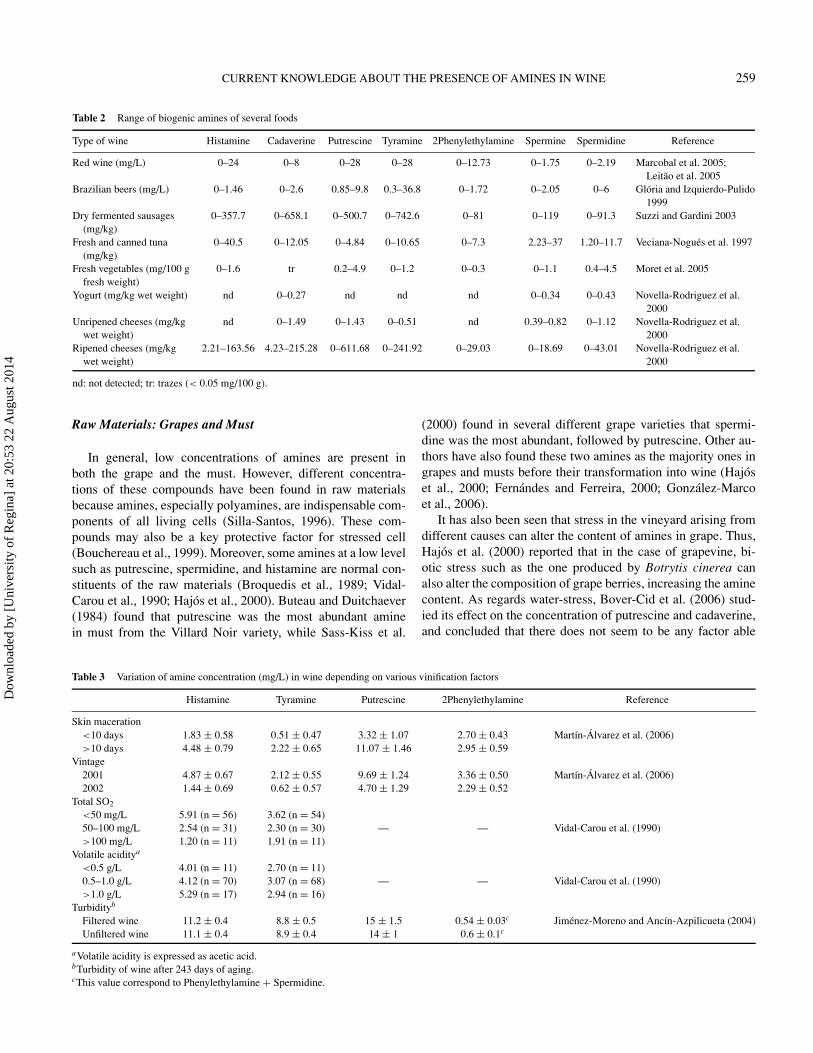
Table 2 Range of biogenic amines of several foods
Type of wine Histamine Cadaverine Putrescine Tyramine 2Phenylethylamine Spermine Spermidine Reference
Red wine (mg/L) 0–24 0–8 0–28 0–28 0–12.73 0–1.75 0–2.19 Marcobal et al. 2005;Leitao et al. 2005
Brazilian beers (mg/L) 0–1.46 0–2.6 0.85–9.8 0.3–36.8 0–1.72 0–2.05 0–6 Gloria and Izquierdo-Pulido1999
Dry fermented sausages(mg/kg)
0–357.7 0–658.1 0–500.7 0–742.6 0–81 0–119 0–91.3 Suzzi and Gardini 2003
Fresh and canned tuna(mg/kg)
0–40.5 0–12.05 0–4.84 0–10.65 0–7.3 2.23–37 1.20–11.7 Veciana-Nogues et al. 1997
Fresh vegetables (mg/100 gfresh weight)
0–1.6 tr 0.2–4.9 0–1.2 0–0.3 0–1.1 0.4–4.5 Moret et al. 2005
Yogurt (mg/kg wet weight) nd 0–0.27 nd nd nd 0–0.34 0–0.43 Novella-Rodriguez et al.2000
Unripened cheeses (mg/kgwet weight)
nd 0–1.49 0–1.43 0–0.51 nd 0.39–0.82 0–1.12 Novella-Rodriguez et al.2000
Ripened cheeses (mg/kgwet weight)
2.21–163.56 4.23–215.28 0–611.68 0–241.92 0–29.03 0–18.69 0–43.01 Novella-Rodriguez et al.2000
nd: not detected; tr: trazes (< 0.05 mg/100 g).
Raw Materials: Grapes and Must
In general, low concentrations of amines are present inboth the grape and the must. However, different concentra-tions of these compounds have been found in raw materialsbecause amines, especially polyamines, are indispensable com-ponents of all living cells (Silla-Santos, 1996). These com-pounds may also be a key protective factor for stressed cell(Bouchereau et al., 1999). Moreover, some amines at a low levelsuch as putrescine, spermidine, and histamine are normal con-stituents of the raw materials (Broquedis et al., 1989; Vidal-Carou et al., 1990; Hajos et al., 2000). Buteau and Duitchaever(1984) found that putrescine was the most abundant aminein must from the Villard Noir variety, while Sass-Kiss et al.
Table 3 Variation of amine concentration (mg/L) in wine depending on various vinification factors
Histamine Tyramine Putrescine 2Phenylethylamine Reference
Skin maceration<10 days 1.83 ± 0.58 0.51 ± 0.47 3.32 ± 1.07 2.70 ± 0.43 Martın-Alvarez et al. (2006)>10 days 4.48 ± 0.79 2.22 ± 0.65 11.07 ± 1.46 2.95 ± 0.59
Vintage2001 4.87 ± 0.67 2.12 ± 0.55 9.69 ± 1.24 3.36 ± 0.50 Martın-Alvarez et al. (2006)2002 1.44 ± 0.69 0.62 ± 0.57 4.70 ± 1.29 2.29 ± 0.52
Total SO2
<50 mg/L 5.91 (n = 56) 3.62 (n = 54)50–100 mg/L 2.54 (n = 31) 2.30 (n = 30) — — Vidal-Carou et al. (1990)>100 mg/L 1.20 (n = 11) 1.91 (n = 11)
Volatile aciditya
<0.5 g/L 4.01 (n = 11) 2.70 (n = 11)0.5–1.0 g/L 4.12 (n = 70) 3.07 (n = 68) — — Vidal-Carou et al. (1990)>1.0 g/L 5.29 (n = 17) 2.94 (n = 16)
Turbidityb
Filtered wine 11.2 ± 0.4 8.8 ± 0.5 15 ± 1.5 0.54 ± 0.03c Jimenez-Moreno and Ancın-Azpilicueta (2004)Unfiltered wine 11.1 ± 0.4 8.9 ± 0.4 14 ± 1 0.6 ± 0.1c
aVolatile acidity is expressed as acetic acid.bTurbidity of wine after 243 days of aging.cThis value correspond to Phenylethylamine + Spermidine.
(2000) found in several different grape varieties that spermi-dine was the most abundant, followed by putrescine. Other au-thors have also found these two amines as the majority ones ingrapes and musts before their transformation into wine (Hajoset al., 2000; Fernandes and Ferreira, 2000; Gonzalez-Marcoet al., 2006).
It has also been seen that stress in the vineyard arising fromdifferent causes can alter the content of amines in grape. Thus,Hajos et al. (2000) reported that in the case of grapevine, bi-otic stress such as the one produced by Botrytis cinerea canalso alter the composition of grape berries, increasing the aminecontent. As regards water-stress, Bover-Cid et al. (2006) stud-ied its effect on the concentration of putrescine and cadaverine,and concluded that there does not seem to be any factor able
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

260 C. ANCıN-AZPILICUETA ET AL.
to modify these aliphatic polyamine contents, although morestudies should be made.
Likewise, volatile amines have been found both in grape andin must. Accordingly, Ough et al. (1981) found that volatileamines were present in the grapes and unlike biogenic amines,most of the volatile amines were found in California musts beforealcoholic fermentation. Gonzalez-Marco et al. (2006) just likethe above-mentioned authors, also found volatile amines (ethy-lamine, diethylamine, dimethylamine, pyrrolidine, and isobuty-lamine) in Chardonnay Spanish must.
Amino Acid Concentration
Biogenic amines and phenylethylamine, which is a volatileamine, are formed through the action of decarboxylase enzymesproduced by microorganisms (ten Brink et al., 1990) from thecorresponding amino acid precursors (Fig. 1). So, as amino acidsare the precursors of amines, the content of free amino acids inthe must could be related to the quantity of amines in wine. Theamino acids present in grape constitute a major source of yeastassimilable nitrogen and they are used by yeast as sources of ni-trogen during alcoholic fermentation (Huang and Ough, 1989;Juhasz and Torley, 1985; Tusseau et al., 1989). Furthermore,amino acids also act as nutrients for the bacteria during the sec-ondary fermentations (Soufleros et al., 2003). The concentrationof amino acids in must depends on many factors such as grape va-riety (Feuillat, 1974; Cantagrel et al., 1982; Millery et al., 1986;Etievant et al., 1988; Huang and Ough, 1991; Sauvage et al.,1993; Presa-Owens et al., 1995; Spayd and Andersen-Baggie,1996), geographical origin (Spayd and Andersen-Baggie, 1996;Ooghie, 1981; Carnevillier et al., 1999), nitrogenous fertilization(Bertrand et al., 1991; Spayd et al., 1994), degree of grape ma-turity (Millery et al., 1986; Kliewer, 1970), vintage and climaticconditions (Feuillat, 1974; Millery et al., 1986; Huang and Ough,1991; Flanzy and Poux, 1965). In addition, different winemakingtechnologies like prefermentation clarification (Ayestaran et al.,1995), crushing (Poux, 1968) and duration of maceration process(Usseglio-Tomasset and Bosia, 1990; Guitart et al., 1997) havean effect on the amino acid fraction in must. The concentrationof amino acids in must could also be modified by the additionof nutrients to the medium, to avoid fermentation problems. Inthis way, some authors have studied the possible relationshipbetween the distinct factors, which affect the concentration ofnitrogen, and amino acid precursors in the must, and the produc-tion of amines in the wine. Bertrand et al. (1991) found that thenitrogenous fertilization of vineyards of the Merlot variety pro-duced an increase in the nitrogenous compounds of the grape, aswell as in the concentration of histamine, putrescine, cadaver-ine and phenylethylamine in wine. Consequently, it would seemthat those factors that increase the concentration of total aminoacids in must means a wine production with a greater contentof amines in wine. However, in real vinifications no correlationhas been observed between the formation of biogenic amines inwine and the consumption by yeasts of their precursor amino
acids, during alcoholic fermentation. In this respect, Ancın et al.(2004) using Garnacha must have found no relationship betweenthe content of biogenic amines in wine and the utilization oftheir precursor amino acids during fermentation. In a later work,Gonzalez-Marco et al. (2006) did not find an increase in the con-centration of amines in wine during alcoholic fermentation, af-ter adding amino acids to the must from autolysate yeast. In thissame study, no correlation was found between the consumptionof amino nitrogen nor of the amino acid precursors of amineswith the formation of biogenic amines in wine after alcoholicfermentation.
As regards the relationship between the consumption ofamino acid precursors and the formation of amines during mal-olactic fermentation, Soufleros et al. (1998) found that duringmalolactic fermentation carried out by indigenous lactic acidbacteria, amino acid concentrations declined significantly whilebiogenic amines increased. In the same way, Martin-Alvarezet al. (2006) observed that during malolactic fermentation theincrease in wine of histamine, tyramine and putrescine was ac-companied by a significant decline in their amino acid precur-sors. In a more general way, Herbert et al. (2005) studied therelationship between the quantity of free amino acids in mustsof different varieties from the Alentejo region with the totalquantity of biogenic amines in red wine that had undergone bothalcoholic and malolactic fermentation. They found that the grapevariety with the highest quantity of amino acids also showed thehighest quantity of amines in the wine.
From these studies it can be concluded that higher amountsof amino acids in must produce higher amounts of biogenicamines after malolactic fermentation. However, in real condi-tions, it is difficult to establish a correlation between the con-centration of biogenic amines and the consumption of their pre-cursor amino acids during alcoholic fermentation. This couldbe due to the fact that during alcoholic fermentation, yeastsmainly use amino acids directly in biosynthesis or as a nitro-gen source and to a lesser extent for decarboxylation reactionwhere biogenic amines are formed. It is also necessary to takeinto account that the formation of amines is highly influencedby the aminogenic capacity both of yeasts as well as of bacteriathat carry out the malolactic and alcoholic fermentations. Thiswould suggest that, for a better knowledge of the nitrogenousmetabolism, including the formation of biogenic amine, it wouldbe important to carry out studies in synthetic media and with se-lected strains in order to eliminate variables from the naturalmedium.
Influence of Winemaking
In general, it has been observed in most of the studies carriedout, that red wines contain more amines than either white or rosewines. Likewise, different authors (Lethonen, 1996; Torrea andAncın, 2002; Gloria et al., 1998) have observed that putrescineis the most abundant amine in red, white, and rose wines. Zeeet al. (1983) compared biogenic amine contents in Canadian,
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 261
American, and European wines, as well as wines from differ-ent regions in France to detect variations due to geographicalfactors and possibly vinification processes. These authors foundthat red wines had significantly more histamine and putrescinethan white wines, but the concentration of cadaverine and tyra-mine were not significantly different. Among the French winesstudied the wines from Bourgogne and Champagne regions hadrelatively higher amounts of the histamine, putrescine, cadav-erine, and tyramine. Jimenez-Moreno et al. (2003) found in ared wine of Merlot variety (2001 vintage) a higher concentra-tion of biogenic amines than that found by Gonzalez-Marcoand Ancın-Azpilicueta (2006) in a white wine of a Chardon-nay variety (2003 vintage). Both vinifications had been madewith varieties from the same geographical zone and in the samecellar.
As red wines have generally a higher concentration of bio-genic amines, they have been examined in different studies.Accordingly, Gloria et al. (1998) determined the amine con-centration in 59 samples of Pinot Noir and Cabernet Sauvi-gnon wines produced in Oregon, USA, from the 1991 and 1992vintages. Pinot Noir wine contained significantly more concen-tration of putrescine, histamine, 2-phenylethylamine, serotonin,and spermine than Cabernet Sauvignon wine; however, this lastwine contained more concentration of cadaverine and spermi-dine. Landete et al. (2005) quantified histamine, tyramine, pu-trescine, and phenylethylamine in wines from three grape va-rieties (Bobal, Garnacha, and Tempranillo) produced in Utiel-Requena (Spain). In this work the authors found an appreciableinfluence of the grape variety in the concentration of amines inwine. Thus, histamine and tyramine concentrations were simi-lar in the wine from Tempranillo and Bobal varieties and higherthan in Garnacha wine. Putrescine concentration found in Bobalwine was slightly lower than those in Tempranillo and Gar-nacha wines. Phenylethylamine concentration was very similarin wines from these three grape varieties. As the soils in whichthe grape varieties were cultured are similar in composition andthey were all subjected to similar winemaking practices, differ-ences in histamine, tyramine, and phenylethylamine concentra-tion could be due to the different content in total amino acidsof these grape varieties (data not shown in the study) or be dueto the different aminogenic ability of the strains isolated in thesamples (Oenococcus oeni, Pediococcus parvulus, Lactobacil-lus hilgardii and Lactobacilus brevis).
Martın-Alvarez et al. (2006) studied the influence of certaintechnological practices on biogenic amines content in red wines.This study showed that there existed a wide variability betweendifferent wine vintages for all the amines studied as well as thefact that the addition of pectolytic enzymes did not influencethe biogenic amine concentration. It was also observed that alonger time period of skin maceration increased the formationof histamine, tyramine, and putrescine and that the wine ag-ing on lees mainly increased the concentration of putrescineand methylamine. It would seem that the fungi that attack thegrape in certain vintages could also influence the amino con-centration in wine. Thus, Kiss and Sass-Kiss (2005) found that
the concentration of biogenic amines in nonbotrytized wines ismuch poorer than in botrytized wines.
Some techniques used in winemaking can reduce the biogenicamine content in wine, such as the use of bentonite (Vidal-Carouand Marine-Font, 1985). In this work, it was found that, in orderto reduce the wine histamine in half, a dose of 1 kg/L of ben-tonita should be used. This dose is much higher than the normaldose used during the vinification process and it could affect thered wine color. Another technique proposed for elimination ofhistamine in wine is a thermal treatment of the grape after press-ing (Inigo and Bravo, 1980). Besides, Ough (1971) set out thepossibility of making refermentations with histaminolytics, butfor the moment there does not seem to have been any positiveresults.
The influence of different vinification processes on the con-centration of wine volatile amines has not been investigated asmuch as the influence that they have on the concentration ofbiogenic amines. Ough and Daudt (1981) found that most ofthe volatile amines in Pinot Noir and White Riesling winesdecreased during alcoholic fermentation. Kiss and Sass-Kiss(2005) found that some amines increased during the wine-making process in botrytized wines and Herbert et al. (2005)concluded that the vintage had a great effect on total volatileamines.
Influence of Vinification Conditions
Other important factors which influence the concentration ofamines in wine are the conditions in which both alcoholic andmalolactic fermentation are carried out. Thus, different authorshave studied factors such as pH, temperature, concentration ofSO2, turbidity, and volatile acidity.
Given that the vinification conditions can be very different,the relationship between these conditions and the concentrationof biogenic amines in wine is not very well-understood. How-ever, there are some studies which can provide some interestingconclusions in this respect. Zee et al. (1983) did not find anyrelationship between pH, alcohol content, and amine concen-trations in Canadian, American, and European wines. However,the pH of wine would seem to have an importance in the for-mation of biogenic amines during malolactic fermentation. Ac-cordingly, Aerny (1990) found that high pH values favored theproliferation of bacteria strains, which can be responsible for theformation of biogenic amines in wine. In this work it was alsofound that the addition of SO2, even in small quantities, after al-coholic fermentation slowed down the malolactic fermentationand favored the development of Pediococcus, which are bacteriawith a high aminogenic capacity. Bauza et al. (1995) found thatlees and marcs can contain high levels of biogenic amines, sothat the use of lees to facilitate malolactic fermentation has tobe avoided since that could increase the concentration of thosecompounds in wine. Gerbaux et al. (1997) found that in winewithout stabilization by lysozime, the bacterial population couldremain stable for several months at high levels, and so producean increase in biogenic amines concentration in wine.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

262 C. ANCıN-AZPILICUETA ET AL.
Vidal-Carou et al. (1990) studied the relationship between theconcentration of histamine and tyramine in wine with the level ofSO2 and volatile acidity. These authors found there existed a cor-relation (99.9%) between total sulfur dioxide level and biogenicamines in red wines, the highest amine contents were foundin wines with low total sulfur dioxide level. They also foundthat there existed a correlation (99.9%) between histamine andtyramine contents and volatile acidity in white and rose wines.However, Bauza et al. (1995) found that using SO2 (20–30 mg/L)to stabilize wine before bottling is insufficient to prevent the for-mation of biogenic amines. Besides, Vidal-Carou et al. (1991)examined changes in the histamine and the tyramine concentra-tion during wine spoilage at various temperatures. No increasein the biogenic amine content was observed during the spoilageof wines, but a decrease of histamine and tyramine content wasobserved during the spoilage of some wines. These changes wereindependent of the storage temperature of the wine.
Gardini et al. (2005) studied the influence of different fac-tors on the production of tyramine, spermine and spermidineof a strain of Oenococcus oeni in a model wine. These authorsfound that the arabinose concentration had an effect on tyra-mine concentration. They also observed that an increase of theSO2 concentration resulted in a reduction of spermine and sper-midine, but this relationship was more complex in the case ofthe formation of tyramine. The SO2 effect on tyramine accumu-lation depended also on other variables like pH. On the otherhand, Jimenez-Moreno and Ancın-Azpilicueta (2004) studiedthe influence of wine turbidity on the accumulation of biogenicamines during aging and found that turbidity did not affect thecontent of biogenic amines in Merlot wine.
The influence of other wine compounds such as malic acid,citric acid, ethanol and sugar has also been studied. Rollan et al.(1995) found that high ethanol (12% v/v), L-lactic acid, and citricacid concentrations reduced the histidine decarboxylase activityof cell suspension of a strain of Oenococcus oeni (Leuconostosoenos 9204). Soufleros et al. (1996) found that biogenic aminesin wine, except for phenylethylamine and putrescine, showednegative correlations with the quantity of malic acid and cit-ric acid. Lonvaud-Funel and Joyeux (1994) found that in thepoorest growth conditions (without glucose and malic acid) theproduction of histamine was enhanced while Moreno-Arribaset al. (2000) found that tyramine formation was enhanced in arich medium (with glucose), so that it would seem that the for-mation of these amines does not have the same metabolic rolefor microorganisms.
As regards the influence of different factors of vinificationon the accumulation of volatile amines, few studies have beenmade and among these the work of Ough and Daudt (1981)stands out. They studied the effect of fermentation temperatureon the concentration of volatile amines in Pinot Noir and WhiteRiesling wines. These authors found that medium fermentationtemperature ranges result in the lowest amine content in wine.
Consequently, although the factors which affect the forma-tion of amines are multiple, and in most cases dependent oneach other, it may be concluded that the greater concentration
of amines that are generally found in red wine more than rose orwhite wine, are due to the influence of different factors. Thesefactors which favor the formation of biogenic amines during redwine vinification are high fermentation temperatures, macera-tion of the solid parts of the grape, excessively non-acid pHs,high yeast biomass, development of malolactic fermentation,and low levels of free SO2. The effect of these factors will bedifferent. On the one hand, they could favor the growth of mi-croorganisms with high aminogenic activity and on the otherhand they could increase the total content of amino acids in must.It should also be pointed out that, once the amines have beenformed, their elimination would be difficult without changingthe wine composition. Accordingly, the most recommendablething would be to control, as far as possible, the factors, whichhave an influence in the formation of these substances.
FORMATION OF AMINES DURING ALCOHOLIC
AND MALOLACTIC FERMENTATION
Amines are formed during different stages of vinification asthe content of amines in wine is much higher than in must, wherethe concentration is very low (Radler and Fath, 1991). In thisway, the formation of amines has been observed during alcoholicfermentation, due to the metabolism of different yeast strains.On the other hand, malolactic fermentation is one of the stagesof vinification where the formation of these compounds could bemost important. Outlined below are the most important aspectsabout the formation of biogenic amines during the developmentof these two fermentative processes.
Formation of Amines during Alcoholic Fermentation of Wine
Transformation of must into wine is the result of a com-plex succession of different yeast species. During the firststages of fermentation, apiculated species with low toleranceto ethanol predominate (Kloeckera, Hanseniaspora, Candida,Pichia, Torulaspora), and later these are gradually replacedby Saccharomyces cerevisiae and related species, with higherethanol tolerance. While some authors have not given much im-portance to alcoholic fermentation in the formation of biogenicamines, the species of yeast that intervene in this fermentationcould be responsible for the presence of biogenic amines in wine.
Buteau et al. (1984) found increases in the concentrationof histamine, tyramine, putrescine, agmatine, and cadaverineduring alcoholic fermentation of Villard Noir musts. Likewise,Bauza et al. (1995) also found increases in the content of his-tamine, putrescine, and phenylethylamine during alcoholic fer-mentation of wine from the Rhone Valley, while the concentra-tion of tyramine did not increase until malolactic fermentation.In contrast, Vidal-Carou et al. (1990) did not detect the forma-tion of histamine during alcoholic fermentation, although theydid find tyramine formation in very low concentrations. Torrea-Goni and Ancın-Azpilicueta (2002–2001) studied the influence
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4
CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 263
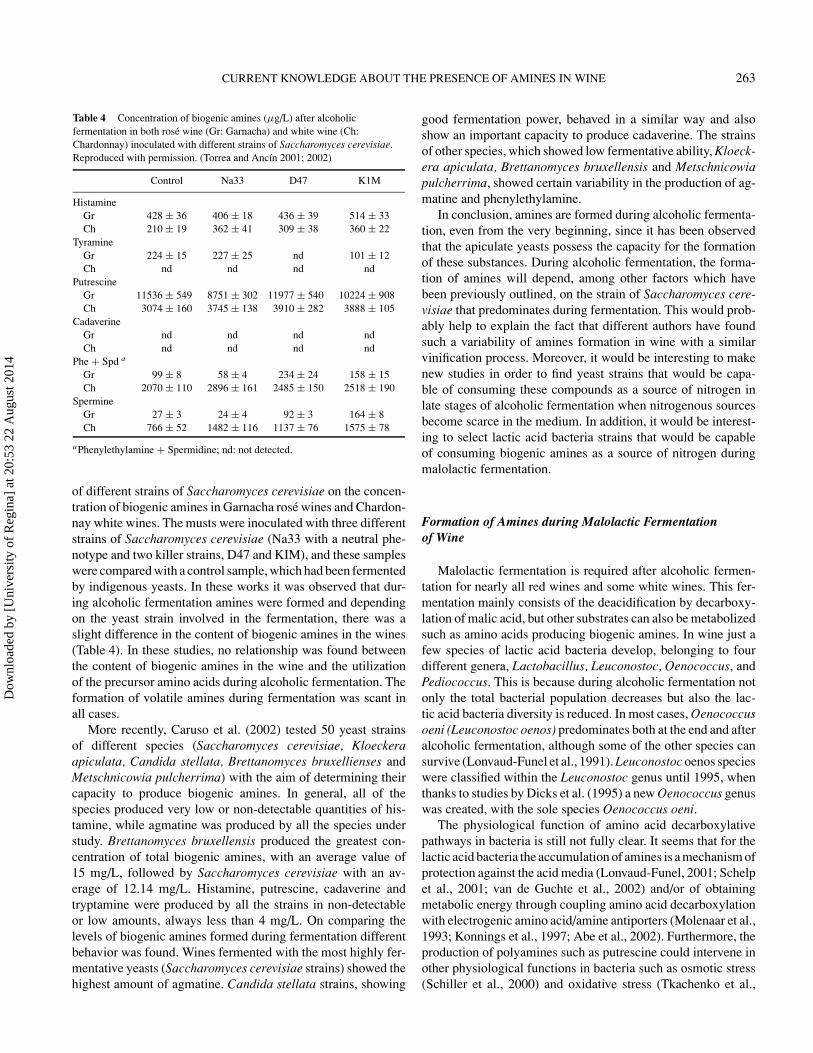
Table 4 Concentration of biogenic amines (μg/L) after alcoholicfermentation in both rose wine (Gr: Garnacha) and white wine (Ch:Chardonnay) inoculated with different strains of Saccharomyces cerevisiae.Reproduced with permission. (Torrea and Ancın 2001; 2002)
Control Na33 D47 K1M
HistamineGr 428 ± 36 406 ± 18 436 ± 39 514 ± 33Ch 210 ± 19 362 ± 41 309 ± 38 360 ± 22
TyramineGr 224 ± 15 227 ± 25 nd 101 ± 12Ch nd nd nd nd
PutrescineGr 11536 ± 549 8751 ± 302 11977 ± 540 10224 ± 908Ch 3074 ± 160 3745 ± 138 3910 ± 282 3888 ± 105
CadaverineGr nd nd nd ndCh nd nd nd nd
Phe + Spd a
Gr 99 ± 8 58 ± 4 234 ± 24 158 ± 15Ch 2070 ± 110 2896 ± 161 2485 ± 150 2518 ± 190
SpermineGr 27 ± 3 24 ± 4 92 ± 3 164 ± 8Ch 766 ± 52 1482 ± 116 1137 ± 76 1575 ± 78
aPhenylethylamine + Spermidine; nd: not detected.
of different strains of Saccharomyces cerevisiae on the concen-tration of biogenic amines in Garnacha rose wines and Chardon-nay white wines. The musts were inoculated with three differentstrains of Saccharomyces cerevisiae (Na33 with a neutral phe-notype and two killer strains, D47 and KIM), and these sampleswere compared with a control sample, which had been fermentedby indigenous yeasts. In these works it was observed that dur-ing alcoholic fermentation amines were formed and dependingon the yeast strain involved in the fermentation, there was aslight difference in the content of biogenic amines in the wines(Table 4). In these studies, no relationship was found betweenthe content of biogenic amines in the wine and the utilizationof the precursor amino acids during alcoholic fermentation. Theformation of volatile amines during fermentation was scant inall cases.
More recently, Caruso et al. (2002) tested 50 yeast strainsof different species (Saccharomyces cerevisiae, Kloeckeraapiculata, Candida stellata, Brettanomyces bruxellienses andMetschnicowia pulcherrima) with the aim of determining theircapacity to produce biogenic amines. In general, all of thespecies produced very low or non-detectable quantities of his-tamine, while agmatine was produced by all the species understudy. Brettanomyces bruxellensis produced the greatest con-centration of total biogenic amines, with an average value of15 mg/L, followed by Saccharomyces cerevisiae with an av-erage of 12.14 mg/L. Histamine, putrescine, cadaverine andtryptamine were produced by all the strains in non-detectableor low amounts, always less than 4 mg/L. On comparing thelevels of biogenic amines formed during fermentation differentbehavior was found. Wines fermented with the most highly fer-mentative yeasts (Saccharomyces cerevisiae strains) showed thehighest amount of agmatine. Candida stellata strains, showing
good fermentation power, behaved in a similar way and alsoshow an important capacity to produce cadaverine. The strainsof other species, which showed low fermentative ability, Kloeck-era apiculata, Brettanomyces bruxellensis and Metschnicowiapulcherrima, showed certain variability in the production of ag-matine and phenylethylamine.
In conclusion, amines are formed during alcoholic fermenta-tion, even from the very beginning, since it has been observedthat the apiculate yeasts possess the capacity for the formationof these substances. During alcoholic fermentation, the forma-tion of amines will depend, among other factors which havebeen previously outlined, on the strain of Saccharomyces cere-visiae that predominates during fermentation. This would prob-ably help to explain the fact that different authors have foundsuch a variability of amines formation in wine with a similarvinification process. Moreover, it would be interesting to makenew studies in order to find yeast strains that would be capa-ble of consuming these compounds as a source of nitrogen inlate stages of alcoholic fermentation when nitrogenous sourcesbecome scarce in the medium. In addition, it would be interest-ing to select lactic acid bacteria strains that would be capableof consuming biogenic amines as a source of nitrogen duringmalolactic fermentation.
Formation of Amines during Malolactic Fermentation
of Wine
Malolactic fermentation is required after alcoholic fermen-tation for nearly all red wines and some white wines. This fer-mentation mainly consists of the deacidification by decarboxy-lation of malic acid, but other substrates can also be metabolizedsuch as amino acids producing biogenic amines. In wine just afew species of lactic acid bacteria develop, belonging to fourdifferent genera, Lactobacillus, Leuconostoc, Oenococcus, andPediococcus. This is because during alcoholic fermentation notonly the total bacterial population decreases but also the lac-tic acid bacteria diversity is reduced. In most cases, Oenococcusoeni (Leuconostoc oenos) predominates both at the end and afteralcoholic fermentation, although some of the other species cansurvive (Lonvaud-Funel et al., 1991). Leuconostoc oenos specieswere classified within the Leuconostoc genus until 1995, whenthanks to studies by Dicks et al. (1995) a new Oenococcus genuswas created, with the sole species Oenococcus oeni.
The physiological function of amino acid decarboxylativepathways in bacteria is still not fully clear. It seems that for thelactic acid bacteria the accumulation of amines is a mechanism ofprotection against the acid media (Lonvaud-Funel, 2001; Schelpet al., 2001; van de Guchte et al., 2002) and/or of obtainingmetabolic energy through coupling amino acid decarboxylationwith electrogenic amino acid/amine antiporters (Molenaar et al.,1993; Konnings et al., 1997; Abe et al., 2002). Furthermore, theproduction of polyamines such as putrescine could intervene inother physiological functions in bacteria such as osmotic stress(Schiller et al., 2000) and oxidative stress (Tkachenko et al.,
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

264 C. ANCıN-AZPILICUETA ET AL.
2001) responses, and also bacterial cell cross-talk (swarming)(Sturgill and Rather, 2004).
In the formation of biogenic amines in wine an important roleis attributed to the lactic acid bacteria responsible for carryingout the malolactic fermentation. In fact, many authors feel thatlactic acid bacteria are responsible for large accumulations ofthese compounds in wine (Soufleros et al., 1998; Bauza et al.,1995; Gerbaux and Monamy, 2000). Table 5 shows the lacticacid bacteria species capable of producing different biogenicamines. Marcobal et al. (2006) found a general increase in theconcentration of amines in the first part of malolactic fermen-tation. These results along with those studies made in vitro byPessione et al. (2005) would seem to indicate that the biosynthe-sis of the amino acid decarboxylase enzymes (which intervenein the formation of amines) could take place when the bacterial
Table 5 Species of lactic acid bacteria able to produce biogenic amines.
Species of lacticacid bacteria References
Histamine Oenococcus oeni Lonvaud-Funel and Joyeux(1994)
Coton et al. (1998b)Guerrini et al. (2002)Landete et al. (2005a)
Lactobacillus hilgardii Farıas et al. (1993)Landete et al. (2005b)Constantini et al. (2006)
Lactobacillus 30a Moreno-Arribas et al., 2003Constantini et al., 2006
Pediococcus damnosus Aerny (1985)Delfini (1989)
Pediococcus parvulus Landete et al. (2005b)Tyramine Lactobacillus brevis Moreno-Arribas et al.
(2000)Lucas and Lonvaud-Funel
(2002)Lucas et al. (2003)Landete et al. (2005a)Constantini et al. (2006)
Lactobacillus hilgardii Moreno-Arribas et al.(2000)
Leuconostoc mesenteroides Moreno-Arribas et al.(2003)
Phenylethylamine Lactobacillus brevis Moreno-Arribas et al.(2000)
Landete et al. (2005a)Lactobacillus hilgardii Moreno-Arribas et al.
(2000)Putrescine Oenococcus oeni Coton et al. (1999)
Guerrini et al. (2002)Marcobal et al. (2004)Mangani et al. (2005)
Lactobacillus 30a Gale (1946)Tabor and Tabor (1985)
Lactobacillus hilgardii Arena and Manca de Nadra(2001)
Lactobacillus plantarum Arena and Manca de Nadra(2001)
Lactobacillus buchneri Moreno-Arribas et al.(2003)
population are at the exponential growth stage and at the endof the growth stage. Other authors have not found any relationbetween the presence of lactic acid bacteria in wine and theformation of biogenic amines. Buteau et al. (1984) have shownthat biogenic amines, especially histamine, decrease during mal-olactic fermentation. Ough et al. (1987) studied the capacity ofdifferent lactic acid bacteria (Lactobacillus, Oenococcus andPediococcus) to produce histamine from histidine under differ-ent fermentation conditions and they did not find significantamounts of histamine from decarboxylation of histidine nor inmodel solutions nor in fermented juice samples. This variabilityin the results could be explained by the fact that the lactic acidbacteria of wine have a different capacity for producing amines,and this capacity is strain dependent rather than being related tospecific species (Coton et al., 1998; Bover-Cid and Holzapfel,1999; Leitao et al., 2000). As a consequence of the variabilityfor producing biogenic amines that the lactic acid bacteria show,several studies have been carried out over the last few yearswhere strains from different lactic acid bacteria, producers ofbiogenic amines, especially histamine, tyramine and putrescine,have been isolated. These bacteria were characterized later at abiochemical and molecular scale.
From different studies (Aerny, 1985; Delfini, 1989) for a longtime it was believed that the formation of histamine and otherbiogenic amines in wine was due to spoilage bacteria, mainlyPediococcus damnosus strains and not to Oenococcus oeni, thebest adapted to carry out malolactic fermentation at the lowpH of wine. However, Lonvaud-Funel and Joyeux (1994) iso-lated a strain of Oenococcus oeni able to produce histamine viahistidine decarboxylase, from a wine from the Bordeaux area.Coton et al. (1998) purified and characterized this enzyme andthe cloning and sequencing of the corresponding gene were alsocarried out. These authors concluded that histidine decarboxy-lase enzyme of Oenococcus oeni requires pyridoxal phosphateas cofactor. Likewise, Coton et al. (1998) examined 118 winesfrom several wine producing regions and they found that prac-tically half of these wines contained lactic acid bacteria withthe hdc gene, responsible for the histidine decarboxylase activ-ity. On isolating and identifying the strains responsible for thisenzyme activity, they found that all of them belonged to theOenococcus oeni species. Landete et al. (2005) screened 136strains of lactic acid bacteria belonging to different species (Lac-tobacillus, Leuconostoc, Oenococcus, and Pediococcus) for thepresence of the hdc gene and their ability to form histamine in asynthetic medium. These authors found that the species, whichshowed the greatest frequency in the production of histaminewas Oenococcus oeni, although the concentration of histamineproduced by this species was less than the one produced bystrains belonging to the Lactobacillus and Pediococcus species.Other authors have also shown that histamine-producing strainsof Oenococcus oeni are very frequent in wine (Guerrini et al.,2002). However, in different studies no potential to form bio-genic amines was observed in different Oenococcus oeni strains(Straub et al., 1995; Moreno-Arribas et al., 2003; Constantiniet al., 2006). With regard to species of other genera different
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 265
to Oenococcus, Farıas et al. (1993) isolated a histamine pro-ducer strain of Lactobacillus hilgardii from Argentinean winesand they partially characterized the histidine decarboxylase en-zyme. Likewise, Constantini et al. (2006) after examining 133strains of lactic acid bacteria only found a strain of Lactobacil-lus hilgardii with the hdc gene responsible for the histidine de-carboxylase activity. Moreno-Arribas et al. (2003) also foundhistidine decarboxylase activity in Lactobacillus 30a. Landeteet al. (2005) found that as well as Oenococcus oeni other lac-tic acid bacteria such as Lactobacillus hilgardii, Lactobacil-lus mali, Leuconostoc mesenteroides, and Pediodoccus parvu-lus can contribute to histamine synthesis in wine, but it wouldseem that the most responsible for the high content of histaminein wine are Lactobacillus hilgardii and Pediodoccus parvulus(Landete et al., 2005).
With respect to the production of tyramine in wine, Moreno-Arribas et al. (2000) studied the formation of this amine bydifferent lactic acid bacteria isolated from different samples ofwine that had undergone malolactic fermentation. They con-cluded that Lactobacillus could be the main lactic acid bacteriaresponsible for the formation of tyramine in wine, since theirresults showed that some strains of Lactobacillus hilgardii andLactobacillus brevis were strong tyramine producers. Further-more, these authors found that Lactobacillus brevis IOEB 9809and Lactobacillus hilgardii IOEB 9649 were capable of produc-ing tyramine and phenylethylamine simultaneously. Gale (1946)had also found that some strains under specific culture condi-tions might possess more than one amino acid decarboxylaseactivity. In a later study, Moreno-Arribas and Lonvaud-Funel(2001) purified and characterized the tyrosine decarboxylaseenzyme from Lactobacillus brevis IOEB 9809. These authorsfound that this enzyme required the cofactor pyridoxal phos-phate just as was found by Coton et al. (1998) in the case of thehistidine decarboxylase enzyme from a Oenococcus oeni strain.Lucas and Lonvaud-Funel (2002) and Lucas et al. (2003) havereported the complete sequences of gene encoding the tyrosinedecarboxylase of Lactobacillus brevis IOEB 9809 and adjacentgenes found in the same locus. The sequence of the Lactobacil-lus brevis tyrosine decarboxylase locus contained four completegenes, which encode a tyrosyl-tRNA synthetase, the tyrosinedecarboxylase, a probable tyrosine permease and a Na+/H+ an-tiporter. Moreno-Arribas et al. (2003) found that Leuconostocmesenteroides may also be responsible for tyramine productionin wines. On the other hand, although Choudhury et al. (1990)reported a Oenococcus oeni strain capable of forming tyraminein a laboratory medium, it would seem that this metabolic prop-erty is not very extended within the Oenococcus oeni species(Lehtonen et al., 1992; Guerrini et al., 2002; Constantini et al.,2006).
Putrescine, the biosynthetic precursor of polyamines, isformed by the decarboxylation of either ornithine or arginineinto agmatine, which is then converted into putrescine either di-rectly or indirectly via N-carbamoylputrescine (Tabor and Tabor,1985). The ornithine decarboxylase activity in lactic acid bacte-ria has been described in Lactobacillus 30a (Gale, 1946; Tabor
and Tabor, 1985). Later, it was shown that the IOEB 8419 strainof Oenococcus oeni, isolated from a wine, was capable of pro-ducing putrescine (Coton et al., 1999), and Moreno-Arribas et al.(2003) found two strains of Lactobacillus buchneri able to formputrescine via ornithine decarboxylase. Recently, Marcobal etal. (2004) isolated a putrescine producer strain of Oenococcusoeni from the lees of a Spanish wine, and sequenced its odcgene, responsible for the ornithine decarboxylase activity. Thehigh concentration of putrescine observed in some wines af-ter malolactic fermentation cannot only come from the decar-boxylation of the free ornithine in wine since the levels of thisamino acid in this product is usually low. The explanation forthese high concentrations of putrescine in wine could be be-cause some lactic acid bacteria show a capacity for degradingarginine, one of the majority amino acids both in must as well aswine, to ornithine. Arginine can be catabolized via the arginine-deiminase (ADI) pathway, consisting of three enzymes, argininedeiminase (ADI), ornithine transcarbamoylase (OTC) and car-bamate kinase (CK) (Liu et al., 1996). Consequently, to formornithine in wine the bacteria strains should possess all the en-zymes of the ADI pathway and these should be active in wineconditions.
Arena and Manca de Nadra (2001) studied the capacity of twostrains of Lactobacillus (Lactobacillus hilgardii, Lactobacillusplantarum) to produce putrescine and agmatine from arginineand ornithine in a synthetic medium. During the growth of Lac-tobacillus hilgardii X1B, arginine concentration declined and,at the same time, citrulline and ornithine (ADI pathway) andagmatine and putrescine (by decarboxylation) were formed.However, Lactobacillus plantarum N4 was only capable ofproducing putrescine from arginine and ornithine. This lacticacid bacteria converted arginine into citrulline during whichan NH2 group was removed from arginine, and then citrullinewas converted to ornithine, which finally underwent decar-boxylation to form putrescine. Guerrini et al. (2002) showedthat the two catabolic properties (arginine degradation and or-nithine decarboxylation) are not necessarily co-existing in pu-trescine producer Oenococcus oeni strains. More recently, Man-gani et al. (2005) showed that Oenococcus oeni can produceputrescine in wine through a metabiotic association, with aninterchange of ornithine, between strains capable of metab-olizing arginine to ornithine, but unable to form putrescine,and strains capable of producing this amine from ornithine,but unable to degrade arginine. In this study, the productionof putrescine through this metabiotic association took placeonce the malolactic fermentation finished, while the conver-sion of ornithine to putrescine by a single ornithine decarboxy-lating strain took place simultaneously to the degradation ofmalic acid.
From these studies it may be concluded that the control ofmalolactic fermentation is one of the most important measuresto take in order to avoid important accumulations of biogenicamines in wine. To do so, it would be essential to do someinvestigation about the capacity of different lactic acid bacteriato produce these compounds. It would be necessary to isolate and
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

266 C. ANCıN-AZPILICUETA ET AL.
select strains with little or no biogenic amine formation ability,but it would also be necessary to check their effectiveness andimplantation in the specific conditions of each type of wine.Furthermore, it would be interesting to study the proteolyticcapacity of the strains to be used in these fermentations, as a highproteolytic enzyme activity could also potentially increase therisk for biogenic amines formation, by increasing the availabilityof free amino acids.
EVOLUTION OF AMINES DURING WINE AGING
Evolution of amines during wine storage in both bottle andbarrel has been little studied, because amines are usually relatedto food rich in proteins with microbial proteolytic activity. Wine,although not rich in proteins, contains high levels of free aminoacids that can be decarboxylated by residual microbial popula-tions to produce the corresponding amines. Wine amino acidscome from the must amino acids not used by the yeasts duringalcoholic fermentation and also from the release to the mediumof cytoplasmic content of the yeast, which is produced at theend of alcoholic fermentation.
Evolution of Amines in Wine Aged in Oak Barrels
and in Cement and Stainless Steel Tanks
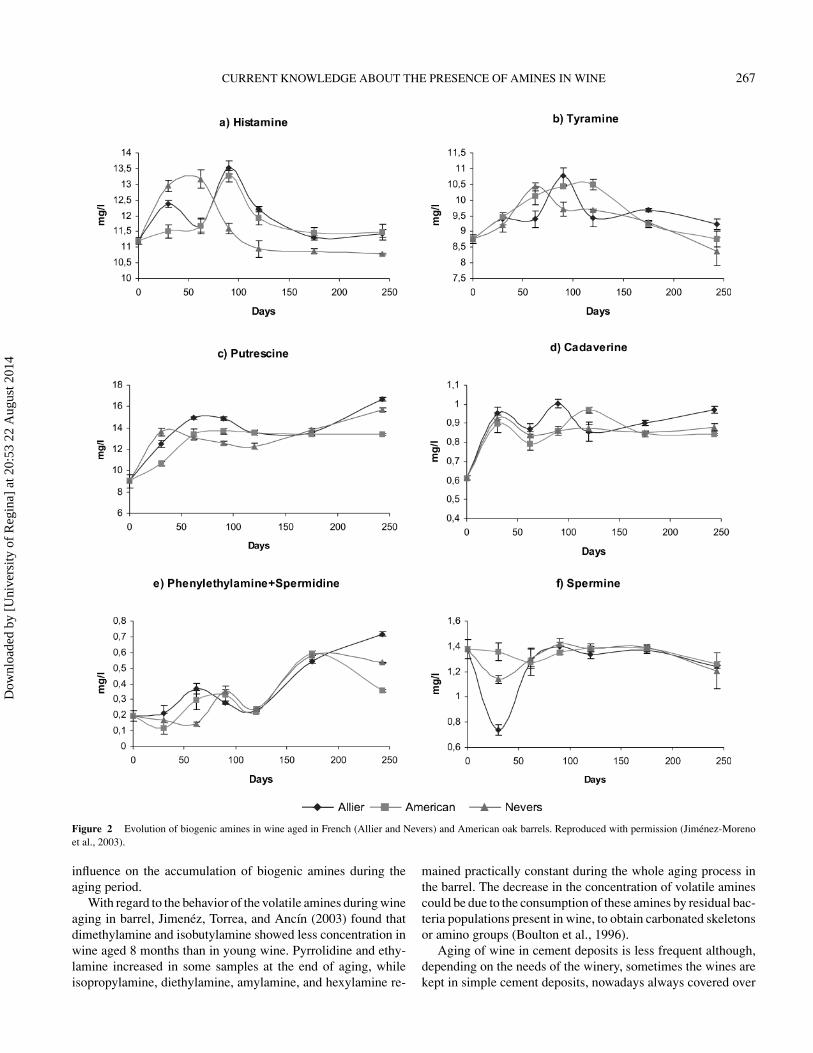
Oak has been used for many centuries to produce barrelsto age wine. During wine maturation in a barrel the productevolves since the oak releases compounds and the barrels per-mit a moderate wine oxygenation. Marcobal et al. (2006) didnot find significant changes (p < 0.05) in the content of his-tamine, tyramine, putrescine, and cadaverine with regard to theconcentration of these amines in red wine after malolactic fer-mentation to which 100 mg/L of SO2 had been added at thebeginning of alcoholic fermentation and 40 mg/L have beenadded after completion of malolactic fermentation. As a re-sult, these authors concluded that a high dose of SO2 avoidsthe formation of amines during wine aging. In this work theformation of biogenic amines in red wines elaborated in fiveSpanish winemaking cellars was studied throughout the wholeprocess of vinification. The wine was aged during six months inoak barrel. Conversely, Gerbaux and Monamy (2000) found im-portant increases in the concentrations of histamine, tyramine,and putrescine during barrel aging of Chardonnay and PinotNoir wines, especially between the fourth and eighth monthof aging. In this work the wines studied came from differentwineries of the Burgundy region and the vinification techniquesused were those of each particular winery. Jimenez, Torrea,and Ancın (2003) studied the evolution of amines during eightmonths of Merlot wine aging in oak barrels (Fig. 2). In thisstudy, the concentrations of histamine and tyramine increasedat the beginning of aging and later decreased, probably due totheir degradation. Putrescine was formed throughout the aging
period, reaching its maximum concentration at the end of theperiod under study. The concentration of cadaverine increasedduring the first month of aging and later it hardly changed. Theconcentration of phenylethylamine+spermidine in the wine in-creased about the fourth month of storage in an oak barrel andthe concentration of spermine hardly varied during the studiedperiod.
The synthesis of biogenic amines during wine aging in oakbarrels would be due to the presence of residual populations oflactic acid bacteria in the wine which, in case of a scarcity ofnutrients, obtain energy through the decarboxylation of aminoacids (Konnings et al., 1997). Therefore, it would seem that thestrains of lactic acid bacteria with decarboxylase activity sur-vive more time in wine than those that do not have this activity(Lonvaud-Funel, 2001). Coton et al. (1998) found that the his-tidine descarboxilase activity and the capacity to form coloniesof Leuconostoc oenos 9204 during wine aging decreased withtime. They also observed that after 2 months of aging bacteriapopulation had completely disappeared while there still existeda notable enzyme activity that lasted even after 4 months. Thiswould suggest that the biogenic amines could be formed in theabsence of viable lactic acid bacteria in wine. Another factor,which could favor the production of amines during wine barrelaging, is the release of amino acids after fermentation due to theautolysis of some yeasts (Slaughter et al., 1987; Kruger et al.,1992) and the alteration of the plasmic membrane of the livingyeasts (Ayestaran et al., 1995; Monteiro and Bisson, 1991). Inmany fermented foods the degradation of histamine and tyra-mine has been observed due to the action of histamine and tyra-mine oxidase enzymes present in some bacteria. These enzymesact by degrading the histamine and tyramine respectively (Voigtand Eitenmiller, 1978; Leuschner et al., 1998; Enes-Dapkeviciuset al., 2000). The results of Jimenez, Torrea, and Ancın (2003)suggest that in wine this enzyme activity also exists and catalyzesthe oxidation of histamine and tyramine so that the concentrationof these amines decreased after the first months of wine aging inbarrel. In wine, the action of these enzymes would be hinderedby acid pH as the greatest activity of these enzymes takes placeat neutral or basic pH. It is not likely the oxygen requirementof these enzymes would be a limiting factor for their activity, asthe barrels permit the micro-oxygenation of the wine.
Wine aging in barrels may be carried out either retaining wineturbidity or after filtering the wine, thus avoiding racking andsaving on costs. Jimenez-Moreno and Ancın-Azpilicueta (2004)studied the accumulation of biogenic amines both in red winefrom Merlot variety aged after diatomaceous earth filtration andin the same wine aged without filtration. The wine remained for243 days in American oak and French oak barrels. Diatoma-ceous earth has a relatively large negative surface charge andso nitrogenous compounds from must such as cationic aminoacids and proteins can be expected to be partially adsorbed onthe surface (Boulton et al., 1996). Consequently, filtration usingdiatomaceous earth could influence the evolution of biogenicamines during wine aging. The results of this work showedthat the different degree of wine turbidity did not have any
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4
CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 267
Figure 2 Evolution of biogenic amines in wine aged in French (Allier and Nevers) and American oak barrels. Reproduced with permission (Jimenez-Morenoet al., 2003).
influence on the accumulation of biogenic amines during theaging period.
With regard to the behavior of the volatile amines during wineaging in barrel, Jimenez, Torrea, and Ancın (2003) found thatdimethylamine and isobutylamine showed less concentration inwine aged 8 months than in young wine. Pyrrolidine and ethy-lamine increased in some samples at the end of aging, whileisopropylamine, diethylamine, amylamine, and hexylamine re-
mained practically constant during the whole aging process inthe barrel. The decrease in the concentration of volatile aminescould be due to the consumption of these amines by residual bac-teria populations present in wine, to obtain carbonated skeletonsor amino groups (Boulton et al., 1996).
Aging of wine in cement deposits is less frequent although,depending on the needs of the winery, sometimes the wines arekept in simple cement deposits, nowadays always covered over
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

268 C. ANCıN-AZPILICUETA ET AL.
(alicated, glass-covered, covered in epoxy resin). Other times,well-protected iron, PVC or stainless steel tanks are used. Bauzaet al. (1995) studied the evolution of biogenic amines duringthe vinification process of different wines from the Rhone Val-ley, including 3 months of wine aging in cement and steel de-posits. These authors found important increases in histamine,putrescine, and tyramine during wine aging while the concen-tration of phenylethylamine hardly evolved.
From these studies it may be concluded that at the begin-ning of wine aging the concentration of most of the biogenicamines increases. Later, the concentration of some amines suchas histamine and tyramine decreases. In this respect, it would beimportant to carry out further studies to get a better knowledgeabout the evolution of biogenic amines as well as to determinewhether these amines could bind to the barrel wood or whethertheir concentration diminished by oxidative degradation or fromreactions with other substances present in wine. On the otherhand, some authors have suggested using high concentrationsof SO2 in order to control the formation of biogenic amines dur-ing aging as this prevents the proliferation of microorganisms.However, the efficiency of this vinification practice is uncertainas viable decarboxylase activity could remain in the medium.
Evolution of Amines in Wine Aged in a Bottle
Sooner or later wine is bottled and this becomes its final anddefinitive storage container. In this form the wine will stay fordifferent lengths of time depending on whether they are youngwines or vintage wines. These latter reach their full maturityin the bottle and the former also improve their quality. Vidal-Carou, Codony-Salcedo, and Marine-Font (1991) studied theevolution of four commercial wines, with different contents ofhistamine and tyramine, during their storage in bottle at differ-ent temperatures. These authors did not find any increase in theconcentration of these amines at any of the temperatures studied(4◦C to 6◦C, 20◦C to 24◦C, 30◦C to 35◦C). However, they founda decrease in the content of histamine in one of the bottles keptat between 20–24◦C after 78 days of wine aging, when it beganto show clear signs of spoilage (volatile acidity higher than 1 g/Lacetic acid). Likewise, they found that the concentration of tyra-mine decreased after 80 days of aging in one of the bottles kept atbetween 4–6◦C. More recently, Landete et al. (2005) studied theinfluence of aging in a bottle on the content of biogenic aminesin young wines from Utiel-Requena and they found an increasein the concentration of histamine during the first 6 months ofwine storage in a bottle. Later, the concentration of this aminedecreased. Tyramine, putrescine and phenylethylamine did notshow any important changes during 12 months of wine storagein a bottle.
Gonzalez-Marco and Ancın-Azpilicueta (2006) studied theevolution of amines from a Chardonnay wine aged in a bottle atdifferent temperatures (4◦C, 20◦C, and 35◦C) over 105 days. Inthis study the concentration of histamine increased during thefirst 45 days in all the samples, independently of the temperature.
At the end of the studied period, the wine kept at 20◦C showeda greater concentration of this amine than the wines stored atmore extreme temperatures. This difference is probably due tothe fact that decarboxylase activity was more favored at roomtemperature than at extreme temperatures of 4◦C and 35◦C. Inaddition, tyramine, putrescine, and cadaverine were formed atthe beginning of aging in all samples and later their concentrationhardly varied, except in the case of tyramine, whose concentra-tion decreased in all samples after 75 storage days in a bottle.The evolution of spermine was completely different to the restof the amines, as this compound progressively disappeared fromthe wine in all the samples, although it did so quicker in the winekept at 4◦C and 35◦C than in the wine at room temperature. So,it would seem that the utilization of this amine by residual pop-ulations of microorganisms is not important to obtain energy.In this study, the evolution of volatile amines was also studied.The concentrations of ethylamine, pyrrolidine, and hexylamineincreased slightly at the beginning of aging, while the concen-tration of dimethylamine decreased during the first 45 days andlater remained constant. The concentrations of isobutylamineand isopropylamine did not show important variations duringthe studied period and the content of diethylamine decreasedslightly in the wine kept at 35◦C.
Consequently, it would seem that, just as was observed in awine aging in a barrel, the amines are also formed at the begin-ning of wine storage in a bottle. On the other hand, the evolutionof these compounds in a bottle scarcely depends on the wineconservation temperature.
ENOLOGICAL ASPECTS OF AMINES
Some polyamines have an important biological function as asource of nitrogen for cells and as precursors in the synthesis ofhormones, alkaloids, nucleic acids, and proteins (Silla-Santos,1996). Furthermore, some amines play an important role in theregulation of body temperature, as well as in the volume and pHof the stomach and in brain activity (Silla-Santos, 1996). How-ever, besides their biological function, amines are important inother aspects. Some biogenic amines, if they are consumed incertain concentrations, can act on the neurotransmitters of thecentral nervous system or on the vascular system, producing,among other symptoms, important modifications in blood pres-sure (Bardocz et al., 1995). Besides, some amines can be precur-sors of nitrosamines and finally, the volatile amines can affectwine aroma.
Toxic Action
Of the amines found in wine, not all of them present toxiceffects for consumers. The main biogenic amines implicated infood poisoning are: histamine, tyramine, putrescine, cadaver-ine, phenylethylamine and tryptamine. Table 6 shows the maineffects that these amines produce in human beings.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 269
Table 6 Biogenic amines and their pharmacological effects (Shalaby, 1996)
Amine Precursor Pharmacological effects
Histamine Histidine Liberates adrenaline and noradrenalineExcites the smooth muscles of uterus, the intestine and the respiratory tractStimulates both sensory and motor neuronsControls gastric acid secretion
Tyramine Tyrosine Peripheral vasoconstrictionIncreases the cardiac outputCauses lacrimation and salivationIncreases respirationIncreases blood sugar levelRelease noradrenaline from the sympathetic nervous systemCauses migraine
Putrescine and cadaverine Ornithine and lysine HypotensionBradycardiaLockjaw Paresis of the extremitiesPotentiate the toxicity of other amines
Phenylethylamine Phenylalanine Releases noradrenaline from the sympathetic nervous systemIncreases the blood pressureCauses migraine
Tryptamine Tryptophane Increase the blood pressure
Histamine is found in a natural way in blood and the concen-tration of this amine in the blood flow is normally at between25 to 130 mg/L (Cardona-Galvez and Gonzalez-Domınguez,2005). Histamine is the most studied biogenic amine and it isusually taken as an indicator of freshness and quality in manyfoods. The effects produced by histamine are the most well-known as they are implicated in most cases of food poisoning(Taylor, 1983). This amine is freed in certain tissues as a resultof allergic hyper-sensitivity or of inflammations and has a potentvasodilatory action which could cause important drops in bloodpressure (Rice et al., 1976). Other symptoms of intoxicationby histamine are vomiting, diarrhea, headaches, facial flushes,itching, and throat inflammation (Silla-Santos, 1996). Histaminehas traditionally been the amine most related to the so-calledwine intolerance that some sensitive people have to this product.However, at present, a certain controversy exists as to whetherhistamine in wine is the cause of all cases of toxic effects pro-duced by the product. In this respect, Kanny et al. (2001) usinghistamine-poor or histamine-rich wines demonstrated that thereis no correlation between the histamine content of wine and hu-man wine tolerance. Consequently, these authors conclude thatthe presence of other compounds in wine, such as acetaldehyde,could explain the fact that there are some people who are in-tolerant to this beverage as these compounds could provoke theliberation of endogenous histamine as the cause of the intol-erance. Histamine consumed in food and drinks is detoxifiedin the intestine gut by two enzymes, diamine oxidase (DAO)and histamine N-methyl transferase (HMT) (Shalaby, 1996).This protection mechanism can become inhibited if the intakeof histamine and/or other biogenic amines is very high, or ifthe enzymes are blocked by other compounds (Hayashi, 1954;Bjedanes et al., 1978). Figure 3 shows the difference betweennormal conditions where the MAO and DAO enzymes act sothat innocuous oxidation products reach the bloodstream and
conditions where there is an absence of enzyme activity wherethere is an entry of amines with a toxic activity in the blood-stream. Among the possible compounds which increase the his-tamine toxicity are trimethylamine, oxide of trimethylamine,acetaldehyde and other biogenic amines such as putrescine, ca-daverine, spermine, spermidine (Cardona-Galvez and Gonzalez-Domınguez, 2005). The action of these enzymes is also reducedwith the consumption of ethanol and by the secondary effects ofsome drugs (Gafner, 2003). The presence of histamine in food
Figure 3 Representation of the biogenic amines absorption through the in-testine gut (normal conditions and in absence of enzyme activity).
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

270 C. ANCıN-AZPILICUETA ET AL.
and drinks does not necessarily imply danger. Many foods con-tain small doses of histamine, which can be easily metabolizedby the human organism. Histamine intolerance seems to be re-lated to a reduced gastrointestinal DAO activity (Wantke et al.,1993; Wantke et al., 1994; Jarisch and Wantke, 1996; Wantkeet al., 1996). Few rigorous studies exist about the different lev-els of activity of the gastrointestinal DAO in both tolerant andintolerant individuals, nor about the differences that could existbetween the sexes or between people of different ages or evenof people with different physical conditions. In addition, geneticpolymorphisms or differences for DAO activity have been iden-tified between tolerant and intolerant individuals (Petersen et al.,2003; Schewelberger et al., 2003). Accordingly, further studiesare needed to determine the mechanism of intolerance to wine.
Tyramine, tryptamine, and phenylethylamine form part ofthe group of vaso-constrictor amines. Tyramine has a vaso-constricting action due to the fact that it provokes a release ofnoradrenaline (Forsythe and Redmond, 1974), which increasesarterial pressure. The importance of tyramine in foods exists bothin its toxic effect and that it also reacts with drugs that inhibitthe (MAOI) giving place to hypertensive crisis (Marine-Font,1978). The use of drugs that inhibit the MAO for the treatmentof mental depression interferes in the detoxification process ofthese amines and can produce hypertensive crisis in patients thataccumulate tyramine in their blood (Blackwell, 1963; Lejonicet al., 1979; Smith and Durack, 1978). Phenylethylamine, liketyramine, provokes increases in blood pressure but in this caseit is due to the liberation of norefedrine (Radler and Fath, 1991).Another symptom of poisoning by these amines could be theappearance of string migraine attacks (Sandler et al., 1974).
Polyamines (putrescine, cadaverine, spermidine, and sper-mine) are indispensable compounds for the living cells. Thesecompounds are necessary for the tissues in growth, given the di-versity of functions attributed to them in the metabolism and thecell growth (Bardocz, 1989). Although cells are able to synthe-size these compounds, diet would seem to be their main source.Besides, spermine and spermidine were seen to be implicated inthe evolution of the intestine tissue. These compounds, althoughthey do not have any toxic activity, can inhibit the action of theMAO and DAO enzymes which are responsible for the degrada-tion of histamine, tyramine, and phenylethylamine and so, theycan reinforce the toxic activity of these amines. In experimentswith animals, it has been proved that toxicity of histamine isincreased by a factor of 10 when this amine is administered 40minutes after the ingestion of putrescine (Parrot and Nicot, 1966)and cadaverine (Hui and Taylor, 1985; Bjedanes et al., 1978).
On the other hand, those amines which contain secondaryamino groups (spermine, spermidine, dimethylamine, pyrroli-dine. . . ) can react with the nitrous acid and its salts to formnitrosamines (Scheme 1) which are compounds with carcino-genic, mutagenic, and theratogenic activity (Scanlan et al., 1982;Gloria et al., 1997). Although wine hardly has any nitrites (Oughand Crowell, 1980) these compounds could reach the intestinegut due to the intake of other types of food and could react withthe secondary amines present in wine in a reaction catalyzed by
SCHEME 1 General reaction of the nitrosamines formation.
enzymes of the intestine flora (Allison and McFarlane, 1989).Maduagwu and Uhaerghu (1986) found that the concentrationof nitrosamines in Nigerian brewed beers and palm wine wasnegligible even though no amount of a chemical carcinogen inhuman foodstuff could be regarded as a safe level.
The toxic limits of biogenic amines have still not been estab-lished and depend on the type of amine, on the food simultane-ously being taken which could interact with them, and on theefficiency of the detoxification mechanisms of different people(Taylor, 1985), although some authors have established differentranges. The different food and drinks and even the different vari-eties of the same food can have different toxic limits. In general,8–40 mg of histamine can cause slight poisoning, over 40 mgmoderate, while over 100 mg severe poisoning. However, itwould seem logical to think that the human tolerance level to thisamine would depend on the sensitivity of each consumer to thiscompound. At present, there are few studies that determine thehistamine concentrations that produce toxic effects on sensitivepeople. The study made by Wankle et al. (1994) stands out. Thiswork was carried out with 28 patients who showed symptoms ofbeing intolerant to wine. These patients, after oral ingestion of125 mL of red wine (equivalent to 50 μg of histamine), showedallergy problems at these levels, and which are easily toleratedby healthy people. Tyramine doses of 10–80 mg could causetoxic swelling and over 100 mg may cause migraine (Askar andTerptow, 1986). The dose of phenylethylamine able to producesymptoms is much lower than that of histamine or tyramine.Sandler et al. (1974) reported that 3 mg of phenylethylaminecauses migraine. Spanjer and van Roode (1971) suggested thatthe sum total of tyramine, histamine, putrescine, and cadaverinein cheese should not exceed 900 mg/kg. Most countries have notestablished limits of concentration for these substances for winealthough there are some pioneer states whose laws recommendlimits for the content of histamine, which as was stated earlier,is one of the most dangerous amines for human health. Thus,Switzerland will not accept wines whose content of histamine ishigher than 10 mg/L, and the quantities recommended in othercountries is even lower than that Germany (2 mg/L), Belgium(from 5 to 6 mg/L), France (8 mg/L) (Bauza et al., 1995; Landeteet al., 2005).
In conclusion, limits in the amine content in wine are difficultto establish. As was mentioned earlier, the appearance of symp-toms after the ingestion of a determined quantity of biogenicamines varies widely from individual to individual as it wouldseem to be due to a reduced gastrointestinal DAO activity. There-fore, before beginning to legislate about recommended or per-mitted levels, it will be necessary to find out what concentrations
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 271
of biogenic amines can provoke toxic effects in both tolerant andintolerant individuals, and the level of intolerance that can ex-ist as well as the fundamental causes for these. Consequently,it would be convenient to study a wide range of individuals,which would include healthy people and those with differentintolerance levels, as well as including members of both sexesand people from a wide range of ages.
Sensorial Effect
The non-proteic nitrogenous compounds such as amines andpeptides contribute to the flavor and taste of food (Maga, 1978).Volatile amines have intense and characteristic odors and thistype of compound could affect wine flavor. Few studies havebeen made on the influence of the concentration of volatileamines in wine and their sensorial effects. Lehtonen et al. (1992)described how the volatile amines, at the pH of wine, are odor-less, but once in contact with the mouth they become par-tially liberated and their undesirable odor comes out. Palamandet al. (1969) studied the effect of these amines on beer aromaand they found that the threshold perception for ethylamine,methylamine, and dimethylamine was at 2 mg/L, 40 μg/L and50 μg/L, respectively. These authors along with Slaughter andUvgart (1971) found that important concentrations of aminesare associated with a decrease in aroma intensity and with theappearance of a vaguely harsh or unpleasant taste in beer. Theyare the only researchers who have attempted to assess the ef-fect of amines on beer aroma while none have been made todetermine the role played by amines in wine flavor. However,it is important to bear in mind that it generally takes a greaterquantity of a compound to cause sensory effects in wine than inbeer because of the greater levels of the alcohol and the flavorcomponents in the first product (Ough and Daudt, 1981).
The majority volatile amines in both white and red winesare ethylamine, dimethylamine, isopropylamine, and pyrroli-dine. Although there are few studies on the content of theseamines in wine Ough and Daudt, (1981) Torrea and Ancın(2002), Jimenez et al. (2003), and Gonzalez-Marco and Ancın-Azpilicueta (2006) found that dimethylamine was the onlyvolatile amine in wine which exceeded the limits establishedby Palamand et al. (1969) affecting beer aroma.
CONCLUSIONS
A wide variability in amine content exists in wine, althoughit can be generally stated that red wine shows a higher con-centration of these substances than rose or white wine. In gen-eral, putrescine is the most abundant amine in most wines. Nu-merous factors can influence the formation of amines, amongthem the maceration of must with the solid parts of the grape,higher fermentation temperatures, not-very-acid pH values, ahigh biomass of yeasts, and lactic acid bacteria. The biogenicamines are mainly formed during malolactic fermentation, al-
though yeasts also synthesize them during alcoholic fermenta-tion. Formation of these substances during alcoholic fermen-tation depends, among other things, on the type of yeast thatpredominates in the fermentation. Although it would seem that,the higher the content of nitrogenous compounds in must, thehigher the content of biogenic amines, no direct correlation hasbeen found between the formation of biogenic amines by theyeasts and the consumption of their precursor amino acids bythese microorganisms. Amines also evolve during wine aging insuch a way that histamine and tyramine are produced at the be-ginning of the aging process, although they do not accumulate inwine, due to their degradation. The concentration of cadaverineincrease slightly at the first stage of aging and, like putrescine,does not degrade at all. In order to better establish the permittedlevels of amines in wine, it would be necessary to know moreabout their toxic effect as well as the influence they have onwine aroma. It is also necessary to do further studies to learnmore about the factors that determine the formation of thesesubstances in wine.
In summary, there are some practices that might permit wine-makers to reduce amine accumulation. Among these, the mostimportant are healthy grape, less excessive nitrogenous fertiliza-tion, yeast and, especially, lactic acid bacteria selected with lowaminogenic capacity, and also a maceration process as short aspossible. Given that, at present, the concentration of amines,which produce toxic effects among the different populationgroups, is not known or indeed, whether there are other winecompounds which release endogenous histamine, it is very dif-ficult at the moment to recommend concrete limits for these sub-stances. In any case, it might be proper to indicate the histaminecontent on the bottle labels as this is the indicator amine for thepresence of other amines and an indicator of the hygienic-healthconditions in the elaboration of the product.
ACKNOWLEDGEMENT
This work was supported by the Ministerio de Ciencia y Tec-nologıa through the project AGL 2006–02414.
REFERENCES
Abe, K., Ohnishi, F., Yagi, K., Nakajima, T., Higuchi, T., Sano, M., Machida, M.,Sarder, R. I., and Maloney, P. C. (2002). Plasmid-encoded as operon confersa proton motive metabolic cycle catalyzed by an aspartate-alanine exchangereaction. J. Bacteriol., 184:2906–2913.
Aerny, J. (1985). Origene de l’histamine dans les vins. Connaissances actuales.Bull. O.I.V., 656–657:1016–1019.
Aerny, J. (1990). Presence d’histamine et d’autres amines biogenes dans lesvins. Off. Int. Vigne Vin, 656–657:1017–1020.
Allison, C., and McFarlane, G. T. (1989). Influence of pH, nutrient availability,and growth rate on amine production by Bacteroides fragilis and Clostridiumperfringens. Appl. Environ. Microbiol., 55:2894–2898.
Ancın, C., Torrea, D., Fraile, P., and Garde, T. (2004). Amino acids and volatilescompounds in the fermentation of inoculated musts: biogenic amines in thewines. In: Shahidi, F., and Weerasinghe, D. K. Eds. Nutraceutical Beverages.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

272 C. ANCıN-AZPILICUETA ET AL.
Chemical, Nutrition and Health effects. Washington: ACS Symposium series871. 302–313.
Anli, R. E., Vural, N., Yilmaz, S., and Vural, Y. H. (2004). The determinationof biogenic amines in Turkish red wines. J. Food Comp. Anal., 17:53–62.
Arena, M. E., and Manca de Nadra, M. C. (2001). Biogenic amine productionby Lactobacillus. J. Appl. Microbiol., 90:158–162.
Askar, A., and Terptow, H. (1986). Biogene amine in Lebensmitteln. Vorkorn-men, Bedeutung und Bestimmung. Stuttgart, Germany: Eugen Ulmer GmbHand Co.
Ayestaran, B. M., Ancın, M. C., Garcıa, A. M., Gonzalez, A., and Garrido, J. J.(1995). Influence of prefermentation clarification on nitrogenous contents ofmusts and wines. J. Agric. Food Chem., 43:476–482.
Bardocz, S. (1989). Polyamines in tissue regeneration. In: Barhrach, U., andHeimer, Y. M. Eds. Physiology of Polyamines. Vol I. CRC Press, Boca Raton,FL, USA, 96–106.
Bardocz, S., Grant, G., Brow, D. S., Ralph, A., and Pusztai, A. (1995).Polyamines in food-implications for growth and health. J. Nutr. Biochem.,4:66–71.
Bauza, T., Blaise, A., Teissedre, P. L., Mestres, J. P., Daumas, F., and Cabanis,J. C. (1995). Evolution des teneurs en amines biogenes des mouts et des vinsau cours de la vinification. Sci. Aliments., 15:559–570.
Bertrand, A., Ingargiola, M. C., and Delas, J. (1991). Effects of nitrogen fertil-ization and grafting on the composition of must and wine from merlot grapes,particularly on the presence of ethyl carbamate. Internat. Symp. Nitrogen inGrapes and Wine, 215–220.
Bjedanes, L. F., Schutz, D. E., and Morris, M. M. (1978). On the aetiologyof scromboid poisoning: cadaverine potentiation of histamine toxicity in theguinea pig. Food Cosmet. Toxicol., 167:157–159.
Blackwell, B. (1963). Hypertensive crises due to monoamine oxidase inhibitors.Lancet, 2:849–851.
Bouchereau, A., Aziz, A., Larher, F., and Martin-Tanguy, J. (1999). Polyaminesand environmental challenges: recent development. Plant Sci., 140:103–125.
Boulton, R. B., Singleton, V. L., Bisson, L. F., and Kunkee, R. E. (1996). Thefining and clarification of wines. In: Boulton, R. B., Singleton, V. L., Bisson,L. F., and Kunkee, R. E. Eds. Principles and Practices of Winemaking. NewYork: Chapman & Hall, 279–319.
Bover-Cid, S., and Holzapfel, W. H. (1999). Improved screening procedure forbiogenic amine production by lactic acid bacteria. Int. J. Food Microbiol.,53:33–41.
Bover-Cid, S., Izquierdo-Pulido, M., Marine-Font, A., and Vidal-Carou, M. C.(2006). Biogenic mono-, di- and polyamine contents in Spanish wines andinfluence of a limited irrigation. Food Chem., 96:43–47.
Broquedis, M., Dumery, B., and Boucard, J. (1989). Miss en evidence depolyamines (putrescine, cadaverine, nor-spermidine, spermidine et spermine)dans les feuilles et les grappes de Vitis vinifera L. Conn. Vigne Vin, 23:1–6.
Busto, O., Guasch, J., and Borrull, F. (1995). Improvement of solid-phase extrac-tion applied to the determination of biogenic amines in wines. J. Chromatogr.,718:307–317.
Buteau, C., and Duitschaever, C. L. (1984). High-performance liquid chromato-graphic detection and quantitation of amines in must and wine. J. Chromatogr.,284:201–210.
Buteau, C., Duitschaever, C. L., and Ashton, G. C. (1984). High performanceliquid chromatographic detection and quantitation of amines in must and wine.J. Chromatogr., 284:201–210.
Cantagrel, R., Symonds, P., and Carles, J. (1982). Composition en acides aminesdu mout en fonction du cepage et de la technologie et son influence sur laqualite du vin. Sci. Aliments, 2:109–142.
Cardona-Galvez, M., and Gonzalez-Domınguez, A. (2005). Intoxicacion his-tamınica o escombroidosis en pescados. Alimentaria, 365:68–73.
Carnevillier, V., Schlich, P., Guerreau, J., Charpentier, C., and Feuillat, M.(1999). Characterization of the production regions of Chardonnay wines byanalysis of free amino acids. Vitis, 38:37–42.
Caruso, M., Fiore, C., Contursi, M., Salzano, G., Paparella, A., and Romano,P. (2002). Formation of biogenic amines as criteria for the selection of wineyeasts. World J. Microbiol. Biotechnol., 18:159–163.
Choudhury, N., Hansen, W., Engesser, D., Hammes, H. P., and Holzapfel, W.H. (1990). Formation of histamine and tyramine by lactic acid bacteria indecarboxylase assay medium. Lett. Appl. Microbiol., 11:278–281.
Constantini, A., Cersosimo, M., del Prete, V., and Garcıa-Moruno, E. (2006).Production of biogenic amines by lactic acid bacteria: screening by PCR,thin-layer chromatography, and High-Performance Liquid Chromatographyof strains isolated from wine and must. J. Food Prot., 69:391–396.
Coton, E., Rollan, G. C., and Lonvaud-Funel, A. (1998). Histidine decarboxy-lase of Leuconostoc oenos 9204: purification, kinetic properties, cloning andnucleotide sequence of the hdc gene. J. Appl. Microbiol., 84:143–151.
Coton, E., Rollan, G., Bertrand, A., and Lonvaud-Funel, A. (1998). Histamine-producing lactic acid bacteria in wines: early detection, frequency, and distri-bution. Am. J. Enol. Vitic., 49:199–204.
Coton, E., Torlois, S., Bertrand, A., and Lonvaud-Funel, A. (1999). Biogenicamines and wine lactic acid bacteria. Bull. O.I.V., 72:22–34.
Delfini, C. (1989). Ability of wine malolactic bacteria to produce histamine. Sci.Aliments, 9:413–416.
Dicks, L. M., Dellaglio, F., and Collins, M. G. (1995). Proposal to reclassifyLeuconostoc oenos as Oenococcus oeni gen. nov., comb. nov. Int. J. Syst.Bacteriol., 45:395–397.
Enes-Dapkevicius, M. L. N., Nout, M. J. R., Rombouts, F. M., Houben, J. H.,and Wymenga, W. (2000). Biogenic amine formation and degradation bypotential fish silage starter microorganisms. Int. J. Food Microbiol., 57:107–114.
Etievant, P., Schlich, P., Bouvier, J. C., Symonds, P., and Bertrand, A. (1988). Va-rietal and geographic classification of French red wines in terms of elements,amino acids and aromatic alcohols. J. Sci. Food Agricult., 45:25–41.
Farıas, M. E., Manca de Nadra, M. C., Rollan, G. C., and Strasser de Saad, A.M. (1993). Histidine decarboxylase activity in lactic acid bacteria from wine.J. Int. Sci. Vigne Vin, 27:191–199.
Fernandes, J. O., and Ferreira, M. A. (2000). Combined ion-pair extraction andgas chromatography-mass spectrometry for the simultaneous determinationof diamines, polyamines and aromatic amines in Port wine and grape juice.J. Chromatogr. A, 886:183–195.
Feuillat, M. (1974). Contribution a l’etude des composes azotes du mout deraisin et du vin, These Univ. Dijon, France.
Flanzy, C., and Poux, C. (1965). Note sur la teneur en acides amines du moutde raisin et du vin en fonction des conditions de l’annee (maturation et fer-mentation). Ann. Technol. Agric., 14:87–91.
Forsythe, W. I., and Redmond, A. (1974). Two controlled trials of tyramine inchildren with migraine. Develop. Med. Child Neurol., 16:794–799.
Gafner, J. (2003). Biological stability of wine and biogenic amines. Proceedingsof the 32nd Annual New York Wine Industry Workshop, Geneva: New YorkState Agricultural Experiment Station. Dept. of Food Science and Technology,74–80.
Gale, E. F. (1946). The bacterial amino acid decarboxylases. Adv. Enzym., 6:1–32.
Gardini, F., Zaccarelli, A., Belletti, N., Faustini, F., Cavazza, A., Martuscelli,M., Mastracola, D., and Suzzi, G. (2005). Factors influencing biogenic amineproduction by a strain of Oenococcus oeni in a model system. Food Control,16:609–616.
Gerbaux, V., and Monamy, C. (2000). Biogenic amines in Burgundy wines.Contents and origin in wines. Rev. Fr. Oenol., 183:25–28.
Gerbaux, V., Villa, A., Monamy, C., and Bertrand, A. (1997). Use of lysoz-ime to inhibit malolactic fermentation and to stabilize wine after malolacticfermentation. Am. J. Enol. Vitic., 48:49–54.
Gloria, M. B. A., Watson, B. T., Simon-Sarkardi, L., and Daeschel, M. A. (1998).A survey of biogenic amines in Oregon Pinot noir and Cabernet sauvignonwines. Am. J. Enol. Vitic., 49:279–282.
Gloria, M. B. A., and Izquierdo-Pulido, M. (1999). Levels and significance ofbiogenic amines in Brazilian beers. J. Food Comp. Anal., 12:129–136.
Gloria, M. B. A., Barbour, J. F., and Scanlan, R. A. (1997). N-Nitrosodimehtylamine in Brazilian, U.S. domestic and U.S. imported beers.J. Agric. Food Chem., 45:814–816.
Gonzalez-Marco, A., and Ancın-Azpilicueta, C. (2006). Amine concentrationsin wine stored in bottles at different temperatures. Food Chem., 99:680–685.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 273
Gonzalez-Marco, A., Jimenez-Moreno, N., and Ancın-Azpilicueta, C. (2006).Influence of addition of yeast autolysate on the formation of amines in wine.J. Sci. Food Agric., 86:2221–2227.
Guerrini, S., Mangani, S., Granchi, L., and Vincenzini, M. (2002). Biogenicamine production by Oenococcus oeni. Curr. Microbiol., 44:374–378.
Guitart, A., Hernandez-Orte, P., and Cacho, J. (1997). Effects of maceration onthe amino acid content of Chardonnay musts and wines. Vitis, 36:43–47.
Hajos, G., Sass-Kiss, A., Szerdahelyi, E., and Bardocz, S. (2000). Changes inbiogenic amine content of Tokaj grapes, wines and Aszu-wines. J. Food Sci.,65:1142–1144.
Hayashi, M. (1954). Investigations on food poisoning caused by ordinary pu-trefaction. IV. Synergism between histamine and several biogenic amines. J.Pharm. Soc. Jpn., 74:1148–1151.
Herbert, P., Cabrita, M. J., Ratola, N., Laureano, O., and Alves, A. (2005).Free amino acids and biogenic amines in wines and musts from the Alen-tejo region. Evolution of amines during alcoholic fermentation and re-lationship with variety, sub-region and vintage. J. Food Eng., 66:315–322.
Huang, Z., and Ough, C. S. (1989). Effect of vineyard locations, varieties androotstocks on the juice amino acid composition of several cultivars. Am. J.Enol. Vitic., 40:135–139.
Huang, Z., and Ough, C. S. (1991). Amino acid profiles of commercial grapejuices and wines. Am. J. Enol. Vitic., 42:261–267.
Hui, J. Y., and Taylor, S. L. (1985). Inhibition in vivo histamine metabolismin rats by foodborne and pharmacologic inhibitors of diamine oxidases, N-methyl transferase, and monoamine oxidase. Food Add. Contam., 8:531–542.
Inigo, B., and Bravo, F. (1980). Histaminogenesis en vinos. I. Estudio de vinosde diversas regiones espanolas. Aliment., 117:57–63.
Jarisch, R., and Wantke, F. (1996). Wine and headache. Int. Arch. Allergy Im-munol., 110:7–12.
Jimenez-Moreno, N., and Ancın-Azpilicueta, C. (2004). Influence of wine tur-bidity on the accumulation of biogenic amines during aging. J. Sci. FoodAgric., 84:1571–1576.
Jimenez-Moreno, N., Torrea-Goni, D., and Ancın-Azpilicueta, C. (2003).Changes in amine concentrations during aging of red wine in oak barrels.J. Agric. Food Chem., 51:5732–5737.
Juhasz, O., and Torley, D. (1985). Effect of free amino acids of the grape on thedevelopment of organoleptic properties of wine. Acta Aliment., 14:101–112.
Kanny, G., Gerbaux, V., Olszewski, A., Fremont, S., Empereur, F., Nabet, F.,Cabanis, J. C., and Moneret-Vautrin, D. A. (2001). No correlation betweenwine intolerance and histamine content of wine. J. Allergy Clin. Immunol.,107:375–378.
Kiss, J., and Sass-Kiss, A. (2005). Protection of originality of Tokaji Aszu:amines and organic acids in botrytized wines by high-performance liquidchromatography. J. Agric. Food Chem., 53:10042–10050.
Kliewer, W. H. (1970). Free amino acids and other nitrogenous substances inwine grapes varieties. J. Food Sci., 35:17–21.
Konnings, W. N., Lolkema, J. S., Bolhuis, H., van Veen, H. W., Poolman, B.,and Driessen, A. J. M. (1997). The role of transport processes in survival oflactic acid bacteria. Antonie van Leeuwenhoek, 71:117–128.
Kruger, L., Pickerell, A. T. W., and Axcell, B. (1992). The sensitivity of dif-ferent brewing yeast strain to carbon dioxide inhibition: Fermentation andproduction of flavour-active volatile compounds. J. Inst. Brew., 98:133–138.
Landete, J. M., Ferrer, S., and Pardo, I. (2005). Which lactic acid bacteria areresponsible for histamine production in wine? J. Appl. Microbiol., 99:580–586.
Landete, J. M., Ferrer, S., Polo, L., and Pardo, I. (2005). Biogenic amines inwines from three Spanish regions. J. Agric. Food Chem., 53:1119–1124.
Lehtonen, P., Saarinen, M., Vesanto, M., and Riekkola, M. L. (1992). Determina-tion of wine amines by HPLC using automated precolumn derivatisation witho-phtahaldehyde and fluorescence detection. Z. Lebensm. Unters. Forsch.,194:434–437.
Leitao, M. C., Marques, A. P., and San Romao, M. V. (2005). A survey of biogenicamines in commercial Portuguese wines. Food Control, 16:199–204.
Leitao, M. C.; Teixeira, H. C., Barreto Crespo, M. T., and San Romao, M. V.(2000). Biogenic amines occurrence in wine. Amino acid decarboxylase and
proteolytic activities expression by Oenococcus oeni. J. Agric. Food Chem.,48:2780–2784.
Lehtonen, P. (1996). Determination of amines and amino acids in wine. A review.Am. J. Enol. Vitic., 47:127–133.
Lejonic, J. L., Gusmini, D., and Brochard, P. (1979). Isoniazid and reaction tocheese. Ann. Int. Med., 91:793.
Leuschner, R. G., Heidel, M., and Hammes, W. P. (1998). Histamine and tyra-mine degradation by food fermenting microorganisms. Int. J. Food Microbiol.,39:1–10.
Liu, S. Q., Pritchard, G. G., Hardman, M. J., and Pilone, G. J. (1996). Argininecatabolism in wine lactic acid bacteria: is it via the arginine deiminase pathwayor the arginase-urease pathway? J. Appl. Bacteriol., 81:486–492.
Lonvaud-Funel, A. (2001). Biogenic amines in wines: role of lactic acid bacteria.FEMS Microbiol. Lett., 199:9–13.
Lonvaud-Funel, A., and Joyeux, A. (1994). Histamine production by wine lacticacid bacteria. Isolation of a histamine-producing strain of Leuconostoc oenos.J. Appl. Bacteriol., 77:401–407.
Lonvaud-Funel, A., Joyeux, A., and Ledoux, O. (1991). Specific enumeration oflactic acid bacteria in fermenting grape must and wine by colony hybridizationwith non-isotopic DNA probes. J. Appl. Bacteriol., 71:501–508.
Lucas, P., and Lonvaud-Funel, A. (2002). Purification and partial gene sequenceof the tyrosine decarboxylase of Lactobacillus brevis IOEB 9809. FEMSMicrobiol. Lett., 211:85–89.
Lucas, P., Landete, J., Coton, M., Coton, E., and Lonvaud-Funel, A. (2003). Thetyrosine decarboxylase operon of Lactobacillus brevis IOEB 9809: charac-terization and conservation in tyramine-producing bacteria. FEMS Microbiol.Lett., 229:65–71.
Maduagwu, E. N., and Uhegbu, F. O. (1986). N-Nitrosamines and Nigerianhabitual drinks, and cancer. Carcinogenesis, 7:149–151.
Maga, J. A. (1978). Amines in foods. Crit. Rev. Food Sci. Nutri., 12:373–403.Mangani, S., Guerrini, S., Granchi, L., and Vincenzini, M. (2005). Putrescine
accumulation in wine: role of Oenococcus oeni. Curr. Microbiol., 51:6–10.Marcobal, A., de las Rivas, B., Moreno-Arribas, M. V., and Munoz, R. (2004).
Identification of the ornithine decarboxylase gene in the putrescine-producerOenococcus oeni BIFI-83. FEMS Microbiol. Lett., 239:213–220.
Marcobal, A., Martın-Alvarez, P. J., Polo, M. C., Munoz, R., and Moreno-Arribas, M. V. (2006). Formation of biogenic amines throughout the industrialmanufacture of red wine. J. Food Prot., 69:397–404.
Marcobal, A., Polo, M. C., Martin-Alvarez, P. J., and Moreno-Arribas, M. V.(2005). Biogenic amine content of red Spanish wines: comparison of a directELISA and an HPLC mehtod for the determination of histamine in wines.Food Res. Int., 38:387–394.
Marine-Font, A. (1978). Alimentos y medicamentos: Interacciones (3a parte).Circ. Farm., 258:43–45.
Martın-Alvarez, P. J., Marcobal, A., Polo. C., and Moreno-Arribas, M. V. (2006).Influence of technological practices on biogenic amine contents in red wines.Eur. Food Res. Technol., 222:420–424.
Millery, A., Duteurtre, B., Boudaille, J. P., and Maujean, A. (1986).Differenciation des trois cepages champenois a partir de l’analyse des acidesamines libres des mouts des recoltes 1983 et 1984. Rev. Fr. Oenol., 103:32–50.
Molenaar, D., Bosscher, J. S., ten Brink, B., Driessen, A. J. M., and Konings, W.N. (1993). Generation of a proton motive force by histidine decarboxylationand electrogenic histidine/histamine antiport in Lactobacillus buchneri. J.Bacteriol., 175:2864–2870.
Monteiro, F., and Bisson, L. F. (1991). Amino acid utilization and urea formationduring vinification fermentations. Am. J. Enol. Vitic., 42:199–208.
Moreno-Arribas, M. V., Polo, M. C., Jorganes, F., and Munoz, R. (2003). Screen-ing of biogenic amine production by lactic acid bacteria isolated from grapemust and wine. Int. J. Food Microbiol., 84:117–123.
Moreno-Arribas, V., and Lonvaud-Funel, A. (2001). Purification and character-ization of tyrosine decarboxylase of Lactobacillus brevis IOEB 9809 isolatedfrom wine. FEMS Microbiol. Lett., 195:103–107.
Moreno-Arribas, V., Torlois, S., Joyeux, A., Bertrand, A., and Lonvaud-Funel,A. (2000). Isolation, properties and behaviour of tyramine-producing lacticacid bacteria from wine. J. Appl. Microbiol., 88:584–593.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

274 C. ANCıN-AZPILICUETA ET AL.
Moret, S., Smela, D., Populin, T., and Conte, L. S. (2005). A survey of biogenicamine content of fresh and preserved vegetables. Food Chem., 89:355–361.
Novella-Rodriguez, S., Veciana-Nogues, M. T., and Vidal-Carou, M. C.(2000). Biogenic amines and polyamines in miles and cheeses by ironhigh performance liquid chromatography. J. Agric. Food Chem., 48:5117–5123.
Ooghie, W., Kastelijn, H., De Waele, A. (1981). Determination de l’origined’un vin rouge a l’aide du spectre des acides amines. Ann. Fals. Exp. Chim.,789:381–408.
Ough, C. S. (1971). Measurement of histamine in California wines. J. Agric.Food Chem., 19:241–244.
Ough, C. S., and Crowell, E. A. (1980). Nitrate determination in California mustsand wines. Am. J. Enol. Vitic., 31:344:346.
Ough, C. S., and Daudt, C. E. (1981). Quantitative determination of volatileamines in grapes and wines. I. Effect of fermentation and storage temperatureon amine concentrations. Am. J. Enol. Vitic., 32:185–188.
Ough, C. S., Crowell, E. A., Kunkee, R. E., Vilas, M. R., and Lagier, S. (1987).A study of histamine production by various wine bacteria in model solutionsand in wine. J. Food Proc. Pres., 12:63–70.
Ough, C. S., Daudt, C. E., and Crowell, E. A. (1981). Identification of newvolatile amines in grapes and wines. J. Agric. Food Chem., 29:938–941.
Palamand, S. R., Hardwick, W. A., and Markl, K. S. (1969). Volatile amines inbeer and their influence on beer flavour.Proceedings of the American Society ofBrewing Chemists. Baltimore, MD: American Society of Brewing Chemists.
Parrot, J., and Nicot, G. (1966). Pharmacology of histamine. In: Roche e Silva,M., Ed. Handbuch der Experimentellen Pharmakologie. Vol XVIII, New York:Springer-Verlag, 148.
Pessione, E., Mazzoli, R., Giuffrida, M. G., Lamberti, C., Garcia-Moruno, E.,Barello, C., Conti, A., and Giunta, C. (2005). A proteomic approach to study-ing biogenic amine producing lactic acid bacteria. Proteomics, 5:687–698.
Petersen, J., Drasche, A., Raithel, M., and Schwelberger, H. G. (2003). Analysisof genetic polymorphisms of enzymes involved in histamine metabolism.Inflamm. Res., 52:S69–70.
Poux, C. (1968). Composes azotes dans les vins de goutte vins de presse et vinsde lies. Ann. Technol. Agric., 17:315–332.
Presa-Owens, C., Lamuela-Raventos, R., Buxaderas, S., and Torre-Boronat, C.(1995). Characterization of Macabeo, Xarel.lo and Parrellada white winesfrom the Penedes region. II. Am. J. Enol. Vitic., 46:529–541.
Radler, F., and Fath, K. P. (1991). Histamine and others amines in wines. In:Rantz, J. Ed. Proceedings of the international symposium on nitrogen ingrapes and wine. Davis, CA: American Society for Enology and Viticulture,185–195.
Rice, S. L., Eitenmiller, R. R., and Koehler, P. E. (1976). Biologically activeamines in food. A review. J. Milk Food Technol., 39:353–358.
Rollan, G. C., Coton, E., and Lonvaud-Funel, A. (1995). Histidine decarboxylaseactivity of Leuconostos oenos 9204. Food Microbiol., 12:455–461.
Sandler, M., Youdin, M. B. H., and Hanington, E. (1974). A phenylethylamineoxidizing defect in migraine. Nature, 250:335–336.
Sass-Kiss, A., Szerdahelyi, E., and Hajos, G. (2000). Study of biologically activeamines in grapes and wines by HPLC. Chromatrogr. Suppl., 51: 316–320.
Sauvage, F. X., Nicol, M. Z., Verries, C., Sarris, J., Pradal, M., and Robin, J. P.(1993). Acides amines libres et quelques activites enzymatiques de mouts deraisins murs. Analyses statistiques de l’effect varietal. Sci. Aliments, 13:443–462.
Scanlan, R. A., Barbour, J. F., Castegnaro, M., Clark, T., Fazio. T., Feit, M.,Fiddler, W., Griffith, A., Herwig, W., and Koski, P. (1982). Nitrosamines inmalt and beer. J. Am. Soc. Brew. Chem., 92–95.
Schelp, E., Worley, S., Monzingo, A. F., Ernst, S., and Robertus, J. D. (2001).pH-induced structural changes regulate histidine decarboxylase activity inLactobacillus 30a. J. Mol. Biol., 306: 727–732.
Schewelberger, H. G., Drasche, A., Petersen, J., and Raithel, M. (2003). Geneticpolymorphisms of histamine degrading enzymes: from small-scale screeningto high-throughput routine testing. Inflamm. Res., 52:S71–73.
Schiller, D., Kruse, D., Kneifel, H., Kramer, R., and Burkovski, A. (2000).Polyamine transport and role of potE in response to osmotic stress in Es-cherichia coli. J. Bacteriol., 182:6247–6249.
Shalaby, A. R. (1996). Significance of biogenic amines to food safety and humanhealth. Food Res. Int., 29:675–690.
Silla-Santos, M. H. (1996). Biogenic amines: their importance in foods. Int. J.Food Microbiol., 29:213–231.
Slaughter, J. C., and Uvgard, A. R. (1971). Volatile amines of malt and beer. J.Inst. Brew., 77:446–450.
Slaughter, J. C., Flint, P. W. N., and Kular, K. S. (1987). The effect of CO2 onthe absorption of amino acids from malt extract medium by Saccharomycescerevisiae. FEMS Microbiol. Lett. 40:239–243.
Smith, C. K., and Durack, D. T. (1978). Isoniazid and reaction to cheese. Ann.Int. Med., 88:520–521.
Soufleros, E. H., Bouloumpasi, E., Tsarchopoulos, C., and Biliaderis, C. G.(2003). Primary amino acid profiles of Greek white wines and their use inclassification according to variety, origin and vintage. Food Chem., 80:261–273.
Soufleros, E., Barrios, M. L., and Bertrand, A. (1996). Correlations en-tre les amines biogenes et d’autres constituants du vin adaptation d’unemethode HPLC au dosage des acides amines. Oenologie 95: 5e Sym-posium International d’Oenologie/ coord par Aline Lonvaud-Funel. 665–669.
Soufleros, E., Barrios, M. L., and Bertrand, A. (1998). Correlation between thecontent of biogenic amines and other wine compounds. Am. J. Enol. Vitic.,49:266–278.
Spanjer, M. C., and Van Roode, B. A. S. W. (1991). Towards a regulatory limitfor biogenic amines in fish, cheese and sauerkraut. De Ware (n)-Chemicus,21:139–167.
Spayd, S. E., and Andersen-Baggie, J. (1996). Free amino acid composition ofgrape juice from 12 Vitis vinifera cultivars in Washington. Am. J. Enol. Vitic.,47:389–402.
Spayd, S. E., Wample, R. L., Evans, R. G., Stevens, R. G., Seymour, B. J.,and Nagel, C. W. (1994). Nitrogen fertilization of white Riesling grapesin Washington. Must and wine composition. Am. J. Enol. Vitic., 45:34–42.
Straub, B. W., Kicherer, M., Schilcher, S. M., and Hammes, W. P. (1995). Theformation of biogenic amines by fermentation organisms. Z. Lebensm. Unters.Forsch., 201:79–82.
Sturgill, G., and Rather, P. N. (2004). Evidence that putrescine acts as an extra-cellular signal required for swarming in Proteus mirabilis. Mol. Microbiol.,51:437–446.
Suzzi, G., and Gardini, F. (2003). Biogenic amines in dry fermented sausages:a review. Int. J. Food Microbiol., 88:41–54.
Tabor, C. W., and Tabor, H. (1985). Polyamines in microorganisms. Microbiol.Rev., 49:81–99.
Taylor, S. L. (1983). Histamine poisoning associated with fish, cheese and otherfoods. FAO/WHO monograph CX/PH 83/11.
Taylor, S. L. (1985). Histamine poisoning associated with fish, cheeseand other food. Monograph VPH/FOS/85.1. World Health Organization,Geneva.
ten Brink, B., Damink, C., Joosten, H. M. L. J., and Huis in ’t Veld, J. H. J.(1990). Occurrence and formation of biologically active amines in foods. Int.J. Food. Microbiol., 11:73–84.
Tkachenko, A., Nesterova, L., and Pshenichnov, M. (2001). The role of thenatural polyamine putrescine in defense against oxidative stress in Escherichiacoli. Arch. Microbiol., 176:155–157.
Torrea, D., and Ancın C. (2002). Content of biogenic amines in a Chardonnaywine obtained through spontaneous and inoculated fermentations. J. Agric.Food Chem., 50:4895–4899.
Torrea-Goni, D., and Ancın-Azpilicueta, C. (2001). Influence of yeaststrain on biogenic amines content in wines: relationship with the uti-lization of amino acids during fermentation. Am. J. Enol. Vitic., 52:185–190.
Tusseau, D., Benoit, C., and Valade, M. (1989). Etude de l’evolution des acidesamines au cours de la maturation. Actual. Oenol., 20–24.
Usseglio-Tomasset, L., and Bosia, P. D. (1990). Evolution des acidesamines et des oligopeptides du mout au vin. Bull. O.I.V., 63:21–46.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4

CURRENT KNOWLEDGE ABOUT THE PRESENCE OF AMINES IN WINE 275
van de Guchte, M., Serror, P., Chervaux, C., Smokvina, T., Ehrlich, S. D.,and Maguin, E. (2002). Stress responses in lactic acid bacteria. Antonie vanLeewenhoek., 82:187–216.
Vazquez-Lasa, M. B., Iniguez-Crespo. M., Gonzalez-Larraina, M., andGonzalez-Guerrero, A. (1998). Biogenic amines in Rioja wines. Am. J. Enol.Vitic., 49:229.
Veciana-Nogues, M. T., Marine-Font, A., and Vidal-Carou, M. C. (1997). Bio-genic amines in fresh and canned tuna. Effects of canning on biogenic aminescontents. J. Agric. Food Chem., 45:4324–4328.
Vidal-Carou, M. C., Ambatle-Espunyes, A., Ulla-Ulla, M. C., and Marine-Font,A. (1990). Histamine and tyramine in Spanish wines: their formation duringthe winemaking process. Am. J. Enol. Vitic., 41:160–167.
Vidal-Carou, M. C., and Marine-Font, A. (1985). Histamina en vinos. Rev. Agro-quim. Tecnol. Aliment., 25:58–78.
Vidal-Carou, M. C., Codony-Salcedo, R., and Marine-Font, A. (1990). His-tamine and tyramine in Spanish wines relationships with total sulfur diox-ide level, volatile acidity and malolactic fermentation intensity. Food Chem.,35:217–227.
Vidal-Carou, M. C., Codony-Salcedo, R., and Marine-Font, A. (1991). Changesin the concentration of histamine and tyramine during wine spoilage at varioustemperatures. Am. J. Enol. Vitic., 42:145–149.
Voigt, M. N., and Eitenmiller, R. R. (1978). Role of histidine and tyrosinedecarboxylases and mono- and diamine oxidases in amine build-up in cheese.J. Food Prot., 41:182–186.
Wantke, F., Gotz, M., and Jarisch R. (1993). Histamine-free diet: treatment ofchoice for histamin-induced food intolerance and supporting treatment forchronic headaches. Clin. Exp. Allergy, 12:982–985.
Wantke, F., Hemmer, W., Haglmuller, T., Gotz, M., and Jarisch, R. (1996).Histamine in wine. Bronchoconstriction after a double-blind placebo-controlled red wine provocation test. Int. Arch. Allergy Immunol., 110:397–400.
Wantke, F., Manfred, G., and Jarisch, R. (1994). The red wine provocationtest: intolerance to histamine as a model for food intolerance. Allergy Proc.,15:27–32.
Zee, J. A., Simard, R. E., Heureux, L. L., and Tremblay, J. (1983). Biogenicamines in wines. Am. J. Enol. Vitic., 34:6–9.
Dow
nloa
ded
by [
Uni
vers
ity o
f R
egin
a] a
t 20:
53 2
2 A
ugus
t 201
4