cortes transversales en plantas monocotiledÓneas y dicotiledÓneas
TRANSCRIPT

ESTRUCTURA Y FISIOLOGÍA CELULAR
2010CORTES TRANSVERSALES EN
PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS
GUTIÉRREZ ARMIJOS LEYDI
LÓPEZ PÉREZ ANTHONY.
MARTÍNEZ BRAVO KATTYA
SANTISTEBAN FARRO ARMANDO.
VALERIANO ENRÍQUEZ GENARA
PRESEMINARIO

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
ÍNDICE
2

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
I. INTRODUCCIÓN
El reino vegetal es muy amplio ya que existen más de mil especies en todo el
planeta. Su distribución tan variada hace que se encuentren plantas casi en
cualquier lugar de la tierra. la mayoría de las plantas contienen un pigmento
llamado clorofila.
Las primeras obras botánicas importantes que se conservan datan de los siglos
IV-III anteriores a nuestra era y se deben a Teofrasto (372-287 a. C), primer
botánico que hizo una clasificación y dividió las plantas en:
Monocotiledóneas (con un cotiledón)
Dicotiledóneas (con dos cotiledones)
En la naturaleza se pueden encontrar tres tipos de plantas en función a la
fotosíntesis: plantas con fotosíntesis C3, fotosíntesis C4 y fotosíntesis CAM
(Achá et al. 1999). La mayoría de las plantas conocidas se ajustan al modelo
de fotosíntesis C3, que es el que se describe en los textos de fisiología como el
modelo general de fotosíntesis. Sin embargo las plantas C4 y CAM las
encontramos en la mayoría de los ecosistemas y se diferencian
fisiológicamente de las con fotosíntesis tipo C3 en la manera de la incorporación
del CO2.
En la actualidad con el avance de las ciencias auxiliares aplicadas a la botánica
(bioquímica, citogenética, ecología, etc.) se han incrementado notablemente los
criterios que se siguen en la sistemática. Por ello, las clasificaciones actuales
se basan en rasgos no sólo morfológicos o anatómicas, sino también
citológicos, genéticos, bioquímicos y ecológicos.
En este preseminario vamos distinguir la diferencia entre una
monocotiledóneas y una dicotiledónea y también plantas c3 y c4 esperamos
ampliar sus conocimientos y sea de mucho beneficio para ustedes.
3

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
II. OBJETIVOS.
4

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
III. PLANTAS ANGIOSPERMAS.
1. Generalidades
La sistemática de las angiospermas, el grupo de plantas más diverso y
dominante en la tierra, está basada en el reconocimiento de dos grandes
grupos. Las monocotiledóneas (o clase Liliopsida) y las dicotiledóneas (o clase
Magnoliopsida), Desde el punto de vista práctico, las características
diferenciales entre estos taxones son fáciles de observar y parecen ser
mutuamente excluyentes, generando un sistema de dos clases opuestas entre
sí y de igual rango taxonómico. Esta división ha prevalecido durante más de
dos siglos y ha sido el eje central de los sistemas de clasificación más
aceptados durante el siglo XX, entre otros los de Cronquist (1981,1988),
Dahlgren (1983), Hutchinson (1959), Takhtajan (1980, 1997) Y Thorne (1992).
Sin embargo, las relaciones filogenéticas y la diversificación inicial de las mono-
y las dicotiledóneas continúan siendo ambiguas. El estudio de nuevos
caracteres, la reevaluación de caracteres ya conocidos y el empleo de otros
métodos en sistemática, muestran que la clasificación de las angiospermas no
es tan simple y que la delimitación entre mono- y dicotiledóneas es bastante
discutible. Los datos morfológicos, paleobotánicos y moleculares disponibles en
la actualidad indican que la diversificación de las monocotiledóneas ocurrió
durante periodos muy tempranos en la evolución de las angiospermas
(Cronquist, 1988; Taylor & TayJor, 1993). El presente trabajo sintetiza las
hipótesis filogenéticas alternativas propuestas recientemente, y presenta a la
familia Aristolochiaceae como la antítesis del sistema binario que ha
prevalecido en la clasificación de las angiospermas.
2. Definición
Son plantas con flores y semillas caracterizadas por una doble
fecundación y por la existencia de frutos cerrados.
5

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
3. Características
70.000 especies, número que aumenta cada año en consonancia con
los descubrimientos de nuevas especies por parte de los botánicos
sistemáticos; éstas especies se agrupan en 300 ó 400 familias según los
criterios de los diferentes autores.
Se separan las especies de este enorme subgrupo en dos clases,
ateniéndose al número de cotiledones que poseen sus semillas. Se
distinguen así las monocotiledóneas, con un solo cotiledón en su
semilla, hojas paralelinervias y raíces fasciculadas; y las dicotiledóneas,
con dos cotiledones, hojas con nerviación articulada y raíces axomorfas.
La característica fundamental de éste subgrupo reside en la estructura
del órgano femenino: el cárpelo. Es el órgano foliáceo, cerrado, vuelto
sobre sí mismo, que se transforma en fruto después de la fecundación,
en su interior se encuentran los óvulos que evolucionan hasta ser
semillas. Como resultante de la estructura del cárpelo, el grano de polen
hasta los grupos reproductores.
La mayoría de las especies angiospermas tienen raíces, tallos y flores.
No son más que adaptaciones a ambientes extremos o a una biología
muy particular, las que provocan la atrofia de cierto órgano, traducción
del sistema foliar por parte de plantas desérticas, supremisión por parte
de los parásitos.
La morfología y la estructura de las raíces, tallos y hojas, caracterizan a
las angiospermas de manera menos rigurosa que las flores. Las
angiospermas comprenden junto a las leñosas, en considerable número
de plantas herbáceas, tienen unas hojas que ordinariamente poseen
formas de lámina más o menos recortadas y que no viven
corrientemente más allá de una estación.
Para reconocer estas plantas vamos ver sus diferentes partes de la planta
encontramos:
6

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Las Hojas:
Las hojas de las angiospermas, son en general, pecioladas. Una hoja completa
consta de vaina, pecíolo y limulo, sí falta el pecíolo, la hoja se dice sesil. La
lámina foliar no es uniforme, en ella se aprecia el nervio y su disposición
constituye su nervadura. El tipo de nervadura más común es la pinnada, con un
nervio principal, en otras no hay solo un nervio principal si no varios, ésta es la
nervadura palmeada.
Cuando el limbe foliar no presenta resalto, la hoja se llama entera, cuando los
bordes no son lisos se llama dentada, si las hendiduras tienen poca
profundidad, festronceadas, si son obtusas, sinuadas etc.
Las hojas pinnadas, pueden tener un número de folíolos impar y se llaman
imparipinnadas, y si es par, paripinnadas. El trébol es una hoja trifoliada. A
menudo en la base de la hoja ya a ambos lados aparecen dos pequeños
apéndices de forma, consistencia y tamaño muy variable, las llamadas
estípulas.
Por su disposición sobre el tallo o ramas, las hojas se denominan alternas,
cuando se insertan a distinta altura, opuestas, si nacen una enfrente de otra y
verticiladas si hay más de las situadas a un mismo nivel. A las hojas próximas a
las flores se les llama brácteas.
Las Flores:
En las angiospermas, las flores presentan una gran riqueza y extensa
diversidad de tipos, por lo que se refiere al número de sus piezas florales, la
diversidad es inmensa, el tipo más corriente es el de cinco piezas. En la
mayoría, las piezas de casa verticilo, alternan cuando las flores son isocíclicas.
Los distintos vertícilos, nacen sobre el extremo del cabillo, formando el tálamo.
A menudo es convexo y son más altos cuanto más internos. A veces, es plano
y en su parte central se asienta el gineceo y a su alrededor los estambres y el
7

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
periasto. En el primer caso se dice que la flor es hipógina, y el segundo
perígena. Si el tálamo se une al ovario se denomina epígena.
En la angiosperma, el cáliz realiza una función protectora, la corola constituye
un reclamo para los insectos, como también el perfume y el néctar de su polen.
El andróceo consta de estambre con sacos polínicos, si estos están frente al
eje de la flor son introrsos y si están hacia fuera extrorsos. En las
angiospermas el ginóseo se compone de una o varias hojas capilares cerradas.
En ella se distinguen tres partes. El ovario, el estilo y el estigma. La flor de la
angiosperma puede ser solitaria ó puede encontrarse varias flores reunidas
formando una inflorescencia.
La Inflorescencia:
La flor puede nacer aislada pero a menudo, varias flores agrupadas sobre un
eje común forman las inflorescencias. Se agrupan en dos tipos principales, la
racemosas y las cimosas. En la primera el eje común no termina en flor, en la
segunda sí, las principales inflorescencias racemosas o indefinidas son el
racimo, la espiga, el colimbo, la umbrela y el capítulo.
Las principales inflorescencias cimosas o definidas son el pleocasio, dicasio y
el monocasio.
Todas son simples pero son frecuencia las observamos compuestas. Así en las
embelíferas unos de los caracteres más importantes es poseer flores
agrupadas. En las graminias se disponen en pequeñas espígas y estas forman
espígas compuestas.
La Fecundación
En las angiospermas los rudimentos seminales están cerrados y la fecundación
es muy complicada. Se realizan mediante el tejido conductor hasta alcanzar el
ovario. A veces, la distancia es muy corta y otras relativamente grandes. Una
vez llegado al ovario el tubo polínico se emerge y se introduce en el rudimento
seminal hasta alcanzar el saco embrionario y la ovocélula, a veces, se da más
8

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
rodeo al primer modo de fecundación se designa porogamia y al segundo
aporogamia.
El Fruto:
Se entiende por fruto el ovario con semilla madura al convertirse en fruto el
ovario sufre en la angiosperma transformaciones extremas, cambio de
consistencia y la coloración y ofrece gran riqueza morfológica. La hoja u hojas
que integren el fruto junto a las semillas constituyan el pericarpo.
Atendiendo a la consistencia del pericarpo los frutos son o bien seco, si aquel
es maduro, membranoso, coriáseo ó leñoso, o bien carnosos si es blando. En
unos casos el fruto maduro se abre y suelta las semillas, en otros no se abre
jamás. Los primeros son frutos deiscentes, los segundos indeiscentes.
Entre los frutos secos indeincentes destaca la nuez, la cariopcide y el aquemio.
Los tres son monospermos. Una nuez en sentido botánico no es el fruto del
nogal sino un hueso interno, tienen el fruto en aquemio las especies como la
lechuga, alcachofa. , y en cariopcide la gramínea.
Los principales frutos secos deincentes son la cápsula, la silima, la legumbre y
el folículo.
La drupa y la balla son frutos carnosos. La drupa es fruto con hueso y la balla
es blanda como el tomate. En las flores de ovario infero, cuando el ginéceo se
suelda al tálamo aquel interviene en la formación del fruto, el que caracteriza la
tribu de las pomoídeas. Procede del tálamo escarnoso y más o menos cariáceo
en el corazón que es de origen carpelar, son pomos la manzana, la pera.etc.
Se clasifican en dos grandes grupos:
Mocotiledóneas
Dicotiledóneas
9

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
IV. PLANTAS MONOCOTILEDONEAS Y DICOTILEDONEAS
HISTORIA
La distinción entre monocotiledóneas y dicotiledóneas ha sido atribuida a
Alberto Magno (1193-1280), quien se basó en caracteres anatómicos del tallo;
sin embargo, Alberto Magno empleó el término cotyledo para referirse a
estructuras tales como el pecíolo, el pedúnculo, el pedicelo, el filamento
estaminal o el funículo, más que a la o las hojas cotiledonares contenidas en la
semilla. Es claro, sin embargo, que para el siglo XVIII ya existía una
diferenciación basada en varios caracteres. Ray (1703) resaltó las diferencias
entre estos dos grupos, incluido el rasgo fundamental en este sistema de
clasificación, es decir, la presencia de un solo cotiledón en monocotiledóneas.
Usualmente se acepta que las dicotiledóneas evolucionaron primero, una teoría
ampliamente influida por la mayor cantidad de fósiles asignables a
dicotiledóneas, y por la teoría euantial del desarrollo floral, que considera a las
flores tipo Magnolia como el prototipo de flores primitivas. Sin embargo, los
hallazgos fósiles revelan que la diversificación de las monocotiledóneas ocurrió
hacia el inicio de la evolución de las plantas con flores, los cuales datan del
Hauteriviano-Barremiano-Aptiano, en el Cretácico Inferior (aunque existen
algunos hallazgos que pueden corresponder a angiospermas fósiles del
Jurásico. Sin embargo, la teoría alternativa de que las monocotiledóneas dieron
origen a las demás angiospermas también ha sido propuesta y seriamente
argumentada.
Muchos de los caracteres tradicionalmente empleados para diferenciar entre
monocotiledóneas y dicotiledóneas son fácilmente observables. Esto ha
conllevado a una clasificación simplificada de las angiospermas, la cual se
10

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
basa en la aparente coincidencia de contrarios que muestran los caracteres de
los dos grupos de angiospermas. Sin embargo, la mayoría de los caracteres
antes mencionados están lejos de ser universales en cada grupo; ellos se
presentan con regularidad en uno u otro grupo, pero no constituyen
sinapomorfias sensu stricto.
La coincidencia de los contrarios genera una pregunta más profunda en el
sentido de si los estados de cada caracter son en realidad la expresión de un
mismo caracter, lo cual los hace homólogos, o si por el contrario son similitudes
debidas a paralelismo o convergencia, Por ejemplo, la homología del cotiledón
en rnonocotiledóneas y de los dos cotiledones en las dicotiledóneas ha sido
cuestionada por varios autores. De manera similar, surgen preguntas acerca de
la homología del profilo, o del número o la morfología de las piezas florales.
V. PLANTAS MONOCOTILEDONEAS
Las monocotiledóneas se llaman así porque sólo tienen un cotiledón, muchas
veces encargado de absorber las sustancias de reserva de endosperma. La
raíz principal de las monocotiledóneas es de corta duración y la sustituyen
otras de origen caulógeno. Sus tallos aéreos, poco ramificados, presentan los
haces conductores dispersos y carecen de cambium entre el floema y el
xilema, debido a ello, no crecen en grosor. Sus hojas tienen nerviación paralela
y, por estar desprovistas de pecíolo, se insertan en el tallo con una vaina o
base ancha.
Entre las monocotiledóneas hay herbáceas terrestres que, dotadas de bulbos o
tubérculos, pasan la temporada desfavorable dentro de la tierra, herbáceas
palustres y plantas acuáticas. Dentro de las monocotiledóneas encontramos a
las alismátidas, arécidas, lílidas.
Alismátidas.- Dentro de esta subclase se incluyen las plantas herbáceas,
acuáticas o palustres, que representan el peldaño inferior de la clase. Son, por
lo tanto, las monocotiledóneas menos evolucionadas y se sitúan en la escala
evolutiva tras el último grupo de las dicotiledóneos, el de las ninfeales.
Las flores son unisexuales o hermafroditas. Su gineceo suele consistir en
carpelos libros dispuestos helicoidalmente; en cambio, los verticilos del
11

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
periantio (uno o dos) y los de los estambres (con dos trímeros los más
primitivos, que luego se reducen a uno solo), tienen disposición cíclica. Los
frutos acostumbran a ser folículos o núculas, cuyas semillas carecen de
endosperma.
Arécidas.- Esta clase abarca los vegetales herbáceos o leñosos, de flores
cíclicas, por lo regular trímeras, dispuestas en inflorescencia espiciforme y
engrosada a la que rodea una bráctea vistosa, las espata. Se observa en estas
plantas la reducción del número de primordios seminales y la tendencia a
poseer gineceos de carpelos soldados. Todas dan frutos indehiscentes, como
bayas o drupas, y algunas presentan nectarios.
Lílidas.- Es la clase más compleja de las monocotiledóneas por su gran
número de especies leñosas y disponen de gineceos sincárpicos, triloculares.
Sus frutos son principalmente cápsulas, bayas y núculas, y las semillas tienen
endosperma rico en almidón, celulosa o aceites.
TALLO:
ESTRUCTURA PRIMARIA DE TALLO:
Se presentan dos tipos básicos:
1. En la mayoría de las monocotiledóneas resulta difícil distinguir un cilindro
vascular: el sistema consta de un gran número de haces repartidos
irregularmente, en varios ciclos, desde la periferia casi hasta el centro del
tallo; no es posible distinguir los límites entre córtex, cilindro vascular y
médula.
El tejido fundamental, llamado también tejido conjuntivo, puede ser
parenquimático o estar fuertemente esclerificado, o presentar numerosos
cordones de fibras, como en el tallo de los bambúes (Bambusa) y las
palmeras. El centro puede ser hueco.
12

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Transcortes de tallo primario de Monocotiledóneas
Setaria geniculata, Gramínea Scirpus sp., Cyperacea Bambusa sp.
El córtex es muy angosto en tallos aéreos, pero es muy grueso en tallos
subterráneos: rizomas (Iris, Musa), cormos (Gladiolus) y bulbos (Allium,
Tulipa). Algunos rizomas como el de Acorus presentan endodermis.
Excepcionalmente se encuentra esta organización en algunas especies de
dicotiledóneas (Podophyllum: Berberidácea).
Transcortes de rizomas (tallos primarios) de Monocotiledóneas
Convallaria majalis,
muguet
Acorus calamus, sector de rizoma y detalle de la
endodermis
13

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
2. En muchas gramíneas: Secale (centeno), Hordeum (cebada), Oryza (arroz),
Triticum (trigo), Avena y algunas dicotiledóneas (Ranunculáceas,
Ninfeáceas) los haces se disponen en dos círculos: el externo con haces
pequeños, incluidos en la capa subepidérmica de esclerénquima, y el
interno con haces mayores, incluidos en parénquima. Para diferenciar
estos cortes es necesario observar los haces vasculares: los elementos del
metafloema de los haces de gramíneas son de forma poligonal, y se
disponen en forma simétrica y regular; además presentan con frecuencia
laguna protoxilemática.
La parte central parenquimática puede ser descrita como médula, y con
cierta frecuencia, es hueca; en las Gramíneas la médula se destruye
durante el crecimiento sólo en los entrenudos, mientras los nudos la
retienen formando los diafragmas nodales.
Triticum, trigo (Monocot.),
sector de transcorte de
tallo hueco
Juncus, junco (Monocot.),
transcorte de tallo hueco
Ranunculus
(Dicotiled.), sector
de transcorte de
tallo hueco
14

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
ESTRUCTURA SECUNDARIA DEL TALLO:
La mayoría de las Pteridofitas y Monocotiledóneas conserva la estructura
primaria de tallo durante toda su vida. Muchas Pteridofitas extinguidas tenían
estructura secundaria, actualmente sólo los géneros Isoetes y Botrychium
presentan algún crecimiento secundario, bastante anómalo en el primer
género.
Entre las Monocotiledóneas la estructura secundaria es excepcional, producida
por un cámbium de naturaleza diferente.
HOJAS:
1. SISTEMA DÉRMICO
En las hojas angostas y alargadas de Monocotiledóneas las células
epidérmicas son generalmente alargadas en sentido paralelo al eje de la
hoja y los estomas están ordenados en filas longitudinales en las zonas
intercostales, es decir entre las venas.
Esto no es una regla general, las hojas de las Aráceas y Marantáceas
presentan estomas dispersos, igual que las de la "cucharita de agua"
(Limnobium spongia).
En las Gramíneas la epidermis comprende células largas, angostas, que a
menudo tienen paredes anticlinales muy onduladas. En los estomas, las
células oclusivas angostas están asociadas con células subsidiarias;
pueden haber células silícicas (cuerpos en forma de cruz o de silla de
montar), células suberosas y tricomas.
15

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Epidermis abaxial de Andropogon Epidermis adaxial de Limnobium
(Graminea) en vista superficial spongia
(Monocotiledónea)
En las Gramíneas xerófitas con numerosos estomas en la cara adaxial, la
epidermis adaxial presenta células buliformes o células motrices, que se
diferencian de las demás por su forma y su mayor tamaño. Su función es la
protección de las hojas contra la desecación, cuando el aire está seco, las
hojas se pliegan protegiendo la cara adaxial y evitando la transpiración.
La planta de arroz (Oryza sativa) vive parcialmente sumergida, sin embargo
sus hojas no se mojan, porque quedan capas de aire atrapadas entre la
superficie hidrofóbica de la epidermis foliar hoja y el agua circundante. Los
gases difunden desde la atmósfera a la parte sumergida de la planta a través
de estas capas de aire atrapadas (Moore, Clark & Stern, 1995).
Poa, transcorte de hoja Hojas de arroz (Oryza sativa),
mostrando células buliformes con superficie hidrofóbica
16

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
2. SISTEMA FUNDAMENTAL O MESÓFILO
En corte transversal, la estructura de las hojas de monocotiledóneas es
similar a la de las hojas de dicotiledóneas. En Heliconia y Musa sapientium,
con hojas horizontales, hay varias capas de empalizada y una amplia región
de parénquima esponjoso con grandes lagunas. En Lilium y algunas otras
especies se puede distinguir una capa de tejido en empalizada y el resto es
parénquima lagunoso. En monocotiledóneas de hojas erguidas el mesófilo
es generalmente homogéneo.
En las hojas tubulares de Allium el tejido en empalizada aparece cerca de la
epidermis alrededor de la circunferencia entera, y por debajo de ella hay
17
Acoelorrhaphe wrightii (Palmae), transcorte de hoja con mesófilo
homogéneo de hoja

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
parénquima esponjoso. En el centro de la hoja hay una gran cavidad. Las
hojas ensiformes presentan mesófilo homogéneo o isobilateral.
Allium (cebolla), esquema de transcorte de hoja cilíndrica
y detalle del corte
Typha (totora), transcorte de hoja con
mesófilo isobilateral
En algunas Gramíneas, especialmente Eragrostoideae y Panicoideae, el
clorénquima es radial, es decir que sus células rodean los haces vasculares
ordenadas radialmente; pueden ser algo lobadas en Bambusoideae y en
algunas especies son profundamente lobadas o ramificadas.
3. SISTEMA VASCULAR
El sistema vascular de las hojas de Monocotiledóneas está formado por
venas paralelas que convergen en el ápice, ligadas entre sí por finas venas
18

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
comisurales transversales, es decir que a nivel microscópico el sistema es
también reticulado (cerrado).
En las Gramineae el haz mediano puede ser más grande, o la parte
mediana de la lámina está engrosada sobre el lado adaxial, por la presencia
de parénquima incoloro masivo; en ese caso la costilla lleva numerosos
haces vasculares (Canna).
Las hojas unifaciales ensiformes de Iris presentan los haces vasculares en
parte en una fila, en parte en dos muy próximas, todos los haces se orientan
con el xilema hacia adentro. Las hojas tubulares de Allium son una
modificación de estas últimas, en la que todos los hacecillos se encuentran
en un círculo, con el xilema hacia adentro.
19

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Avena, venación foliar A, ápice foliar; B, venación paralela cerrada; C,
transcorte
Venación foliar en hojas de monocotiledóneas A-B, Iris, vaina y lámina
ensiforme; C-D, Canna, vaina y vena media; E, Zea mays (Gramineae)
vena media
20

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Cada hacecillo está rodeado por una vaina. En las gramíneas la vaina muestra
variaciones que son significativas taxonómicamente y son indicadoras del tipo
de fotosíntesis característico de las especies.
En muchas Gramineae se observan extensiones de la vaina: bandas de tejido
que unen los hacecillos con una (hacecillo semitrabado) o ambas epidermis
(hacecillo trabado). Las extensiones pueden estar constituídas solo por tejido
parenquimático o por tejido esclerenquimático. Cuando no hay extensiones los
hacecillos son libres.
Transcorte de hoja de Ammophyla (Gramínea) con haces trabados
21

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Poa (Gramineae), transcorte de hoja mostrando un haz semitrabado y uno libre
En muchas hojas de monocotiledóneas hay grandes cantidades de
esclerénquima; este tejido constituye en algunas especies de Agave, Yucca,
Phormium, Sanseviera, una importante fuente comercial de fibras duras.
Las fibras están asociadas con los haces vasculares o aparecen como
cordones independientes en el mesófilo.
Yucca, transcorte de lámina
22

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
La lígula de las gramíneas es membranácea, y puede estar formada
solamente por las dos capas epidérmicas. En algunas especies de Poa se
observan lígulas con tejido vascular y parénquima clorofiliano asociado
(Chaffey 1984).
VI. PLANTAS DICOTILEDONEAS
Las dicotiledóneas están incluidas dentro de la categoría de las angiospermas
y son plantas con flor. Dentro de esta clase existen al menos ciento setenta y
cinco mil.
Características.- Sus embriones poseen dos cotiledones, característica que da
nombre al grupo. A menudo son plantas compuestas, provistas de pecíolo. El
rasgo que las distingue de las monocotiledóneas es una red de nervios. En los
tallos se puede ver, en sección transversal, los haces conductores dispuestos
en círculo, que se encargan de transportar el agua y las sales minerales.
En esta clase están representados casi todos los tipos de flores. Las partes de
la flor son los sépalos, pétalos, estambres y carpelos que suelen disponerse en
verticilos (cáliz, corola, androceo y gineceo, respectivamente).
Las dicotiledóneas comprenden vegetales leñosos y herbáceos. Los tallos de
los primeros tienen porte arbustivo, si se ramifican desde la base, o arbóreo, si
las ramas aparecen a cierta distancia del suelo. A pesar de existir muchos
órdenes dentro de esta clase sólo hablaremos de algunos.
Salicales: comprende sólo la familia de las salicáceas, con árboles y arbustos
caducifolios, de hojas sencillas provistas de estípulas. Sus flores son
unisexuales. Su fruto en cápsula se abre mediante valvas y libera gran número
de semillitas, provistas de pelos largos y sedosos. Esta familia consta de dos
géneros: Salix (sauces) y Populus (álamos o chopos), cuyas especies crecen
normalmente junto al agua.
23

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Fagales: dentro de esta categoría encontramos a muchos vegetales leñosos,
arbóreos y arbustivos. Sus flores son unisexuales y se reúnen por lo regular en
amentos, erectos o péndulos. En algunos casos, las femeninas son solitarias o
forman grupos reducidos. El cáliz tiene poco volumen o bien, puede no existir.
Las flores de las especies europeas de hoja caduca se abren a veces antes de
que broten las hojas. Dentro de las fagales encontramos tres tipos: fagáceas
(árboles de hojas perennes o caducas, enteras o lobuladas: haya, castaño, el
roble...); betuláceas (incluye árboles o arbustos, cuyas hojas caducas tienen los
bordes dentados como el abedul); las coriláceas (árboles y arbustos
caducifolios como el avellano).
Ericales: se compone en su mayor parte de arbustos, aunque hayan especies
herbáceas y de árboles. Todas son típicas de regiones templadas y
montañosas tropicales. Sus flores completas acostumbran a tener los pétalos
soldados y generalmente la corola parece un tubo. A este orden pertenecen la
familia de las ericáceas, de arbustos de diversos tamaños. El cáliz de sus flores
actinoformas se compone de cuatro o seis sépalos que, normalmente, no están
soldados a la corola. Dentro de esta variante encontramos al madroño y el
rododendro.
TALLO:
ESTRUCTURA PRIMARIA DE TALLO:
En corte transversal de tallo de Gimnospermas y Dicotiledóneas, los haces
vasculares aparecen formando un círculo que delimita una región externa y una
interna de tejido fundamental: córtex y médula respectivamente. Los haces
vasculares están separados entre sí por paneles de parénquima interfascicular
(eustela).
24

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Transcorte de tallo primario de Trifolium, trébol, Dicotiledónea
Los haces vasculares pueden estar muy próximos unos de otros, como
sucede en Tilia o Pelargonium, semejando un anillo continuo, o pueden estar
separados por anchas áreas interfasciculares como sucede en Aristolochia y
otras especies trepadoras.
Transcortes de tallo primario de Tilia (tilo, Dicotiledónea): vista
general y detalle
25

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
El córtex incluye los tejidos situados entre la epidermis y el sistema vascular.
Generalmente es delgado, excepto en las plantas en roseta como Apium,
Plantago y Taraxacum y en las Cycadales. Está constituido básicamente por
parénquima, pero frecuentemente hay también tejidos de sostén (colénquima y
esclerénquima en Dicotiledóneas) y/o estructuras glandulares como los
conductos resiníferos en las Coníferas, y los laticíferos o los conductos
mucilaginosos en Dicotiledóneas.
26

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Aristolochia. A: planta. B: transcorte de tallo primario. C: detalle del transcorte
En los tallos de plantas superiores generalmente no se observa endodermis,
excepto en algunos ejes florales, tallos subterráneos o acuáticos. Puede haber
en cambio una vaina amilífera.
La epidermis puede retener la actividad mitótica para compensar el
crecimiento en espesor del tallo, especialmente cuando la peridermis, el tejido
secundario de protección, se forma tardíamente.
En tallos de hidrófitas sumergidas la epidermis carece de estomas, la cutícula
es muy reducida o falta, el córtex es amplio, constituido por aerénquima. El
cilindro vascular (protostela) es reducido, el xilema está constituido por unos
pocos elementos dispersos en la parte central (Myriophyllum aquaticum), a
veces parcialmente reemplazado por una laguna (Ceratophyllum demersum,
Egeria najas), siendo el floema el principal tejido conductor. El xilema puede
carecer de elementos traqueales, está formado por parénquima xilemático.
En algunas especies se desarrollan hidropotes, estructuras glandulares que
funcionan como "captadores de iones", tomando del agua los minerales
necesarios.
27

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Myriophyllum aquaticum, helechito de agua
Transcorte de tallo Detalle del cilindro central
Ceratophyllum demersum
Transcorte del cilindro central
La médula es la porción que queda encerrada por el sistema vascular. Es
básicamente parenquimática, a veces puede tener función de almacenamiento,
o presentar idioblastos diversos y estructuras glandulares. El contorno de la
médula (región perimedular) puede esclerificarse. En algunas especies la
médula se destruye, resultando en la formación de un tallo hueco; a veces la
28

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
destrucción ocurre sólo en los entrenudos, mientras los nudos retienen la
médula formando los diafragmas nodales como en Phytolacca americana. En
Cecropia las cámaras de los entrenudos están habitadas por hormigas del
género Azteca, que a su vez protegen a la planta contra otros herbívoros.
Hedera helix: médula
rasgada en transcorte
Delphinium ajacis:
transcorte, médula
hueca
Cecropia: tallo,
diafragmas nodales y
cámaras en corte
longitudinal
ESTRUCTURA SECUNDARIA DEL TALLO:
Las Gimnospermas y Dicotiledóneas leñosas desarrollan estructura secundaria,
es decir que se agregan tejidos vasculares y protectores adicionales en tallo y
raíz. En las hojas, el crecimiento secundario se reduce al pecíolo y a la vena
media.
Los tejidos secundarios se producen en dos meristemas laterales de forma
cilíndrica:
1. Cámbium que origina tejidos vasculares secundarios.
2. Felógeno que origina el tejido de protección secundario.
29

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Algunas Dicotiledóneas herbáceas (Ranunculaceae, Nelumbonaceae,
Nymphaeaceae) no producen tejidos secundarios, y su sistema vascular se
parece al de las Monocotiledóneas, ya que tienen haces vasculares cerrados
dispuestos en dos o más ciclos.
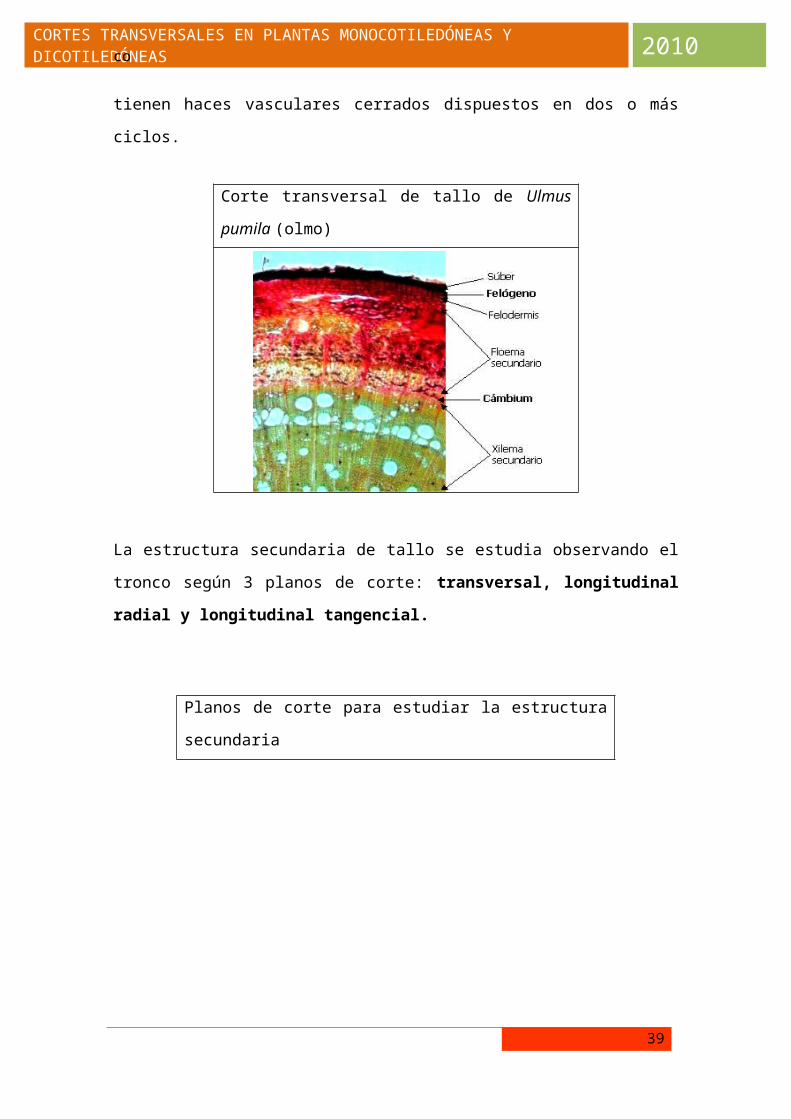
La estructura secundaria de tallo se estudia observando el tronco según 3
planos de corte: transversal, longitudinal radial y longitudinal tangencial.
Planos de corte para estudiar la estructura secundaria
30
Corte transversal de tallo de Ulmus pumila
(olmo)

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
CÁMBIUM
ORIGEN: Una vez terminado el crecimiento primario de la planta, parte del
procámbium permanece en estado meristemático en los haces conductores, y
luego se convierte en cámbium. Este cámbium se denomina fascicular porque
se encuentra en los haces vasculares primarios. Los arcos de cámbium
fascicular son luego conectados por el cámbium interfascicular, que se forma
por desdiferenciación de células del parénquima interfascicular. Esta
desdiferenciación comienza cerca de los haces conductores y enseguida se
extiende lateralmente hacia el parénquima interfascicular.
Origen del cámbium en el tallo (esquemas de transcortes y detalles)
FORMA Y LOCALIZACION: El cámbium se halla localizado entre el floema y el
xilema secundarios; en tallos y raíces tiene la forma de un cilindro, en las hojas
queda reducido a una o varias bandas de células.
31

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Localización del cámbium y los tejidos vasculares en un tronco
HOJAS:
PECÍOLO
El pecíolo frecuentemente muestra forma redondeada en transcorte; cuando la
base foliar es amplia, puede tener forma semilunar (Mahonia). La epidermis
presenta estomas. El córtex generalmente exhibe un anillo completo de
colénquima periférico, o cordones de colénquima si el pecíolo presenta
costillas. El parénquima fundamental puede contener estructuras glandulares, y
células con cristales. A veces hay fibras alrededor del tejido vascular.
Se observan tres tipos básicos de organización vascular:
a) Un haz vascular con el xilema orientado hacia la cara adaxial y el floema
hacia la cara abaxial.
Este tipo de organización es muy frecuente (Camellia, Ilex, Ligustrum,
Nerium).
b) Varios hacecillos dispuestos en un semicírculo (Aristolochia, Mahonia,
Sanguisorba) o en un círculo completo (Platanus, Pelargonium,
32

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Liriodendron, Vitis, Cucurbita, Hedera helix).
c) Los tejidos vasculares pueden formar un anillo continuo, donde el xilema es
interno y el floema periférico (Citrus, Catalpa).
Pecíolos de hojas de Dicotiledóneas en transcorte: organización vascular
Los pulvinos y pulvínulos, situados en los pecíolos y peciólulos
respectivamente, son hinchados, con superficie arrugada. El sistema vascular
está en el centro, rodeado por parénquima con paredes delgadas con espacios
intercelulares. La epidermis presenta estomas escasos o ausentes, puede
haber tricomas. Los movimientos se producen por cambios rápidos de
turgencia, es decir incorporación o pérdida de agua en las células.
Arachis hypogaea
Transcortes de pulvino y pecíolo
33

CORTES TRANSVERSALES EN PLANTAS MONOCOTILEDÓNEAS Y DICOTILEDÓNEAS 2010co
Robinia pseudoacacia
A, transcorte de raquis y corte longitudinal de pulvínulo
situado en el peciólulo. B, transcorte de pulvínulo
34

LÁMINA FOLIAR
1. SISTEMA DÉRMICO
Normalmente la epidermis es el tejido protector de la hoja, aunque
excepcionalmente las pérulas de algunas Dicotiledóneas leñosas pueden
desarrollar una peridermis.
Las células epidérmicas fundamentales de la mayoría de las especies
carecen de cloroplastos; sin embargo, en las hidrófitas sumergidas y en las
higrófitas las células tienen cloroplastos.
Se distingue la epidermis adaxial y la epidermis abaxial. La cutícula es
generalmente más gruesa en la epidermis adaxial. En plantas de ambientes
húmedos la cutícula es delgada, en cambio en las xerófitas, puede ser muy
gruesa.
Maclura tinctoria, esquema de transcorte de lámina foliar y detalle

La epidermis es generalmente simple, pero puede ser múltiple
(pluriestratificada) como en Nerium y Ficus. En Peperomia la epidermis
múltiple puede tener 15 capas de células, siendo más gruesa que el
mesófilo.
En algunas especies, como en Mesembryanthemum cristallinum, hay
células epidérmicas modificadas que cumplen la función de almacenar
agua; son células muy voluminosas, de paredes delgadas, que otorgan un
aspecto particular a la especie, conocida como "planta de hielo".
Simple en Camellia Epidermis foliar
Múltiple en Nerium oleander
Múltiple en Peperomia

La repartición de los
estomas es variable.
Comúnmente las hojas
son
hipostomática s, con
estomas sólo en la
epidermis abaxial (Ej. Maclura tinctoria, Patagonula americana, Camellia).
Las hojas anfistomáticas los presentan en ambas caras, generalmente con
más estomas en la cara abaxial (Ej. Dianthus, clavel). Las hojas epistomáticas
presentan estomas sólo en la cara
adaxial (Ejs. Nymphaea, Victoria
cruziana, irupé).
Dianthus: hoja anfistomática
isobilateral
Nymphaea: hoja epistomática dorsiventral

En las hojas amplias de las dicotiledóneas los estomas están esparcidos. En
plantas mesófitas los estomas generalmente están al mismo nivel que las
demás células. En plantas higrófitas los estomas están elevados sobre las
demás células. Los vegetales de ambientes secos (xerófitos), tienen
dispositivos que evitan la pérdida de agua y al mismo tiempo protegen contra la
radiación luminosa y calor demasiado intensos: la epidermis con pared externa
muy gruesa, cutícula gruesa, revestimientos céreos, resinosos o calcáreos;
estomas estrechos y hundidos. El número de estomas es elevado en las
xerófitas para favorecer el intercambio gaseoso cuando el suministro hídrico es
favorable, pero pueden estar cerrados permanentemente durante la estación
desfavorable.
Transcorte de hoja de Ruellia, planta higrófita
Epidermis de Gasteria, planta xerófita
A veces los estomas se encuentran en surcos o en
criptas para evitar la evaporación (Ej.: Nerium oleander) con abundantes
tricomas muertos que forman una capa protectora.
Criptas estomáticas en Nerium oleander, laurel rosa
Vista superficial, MEB Transcorte de hoja mostrando dos criptas
Detalle de una cripta
Las hojas de Littorella (Plantaginaceae) carecen de estomas y tienen cutículas
gruesas, de manera que sus raíces transportan CO2 para la fotosíntesis
(Moore, 1995).
La epidermis puede tener idioblastos diversos, por ejemplo Maclura tinctoria
presenta cistolitos en la epidermis.

Las hojas pueden ser glabras, o pilosas; la variabilidad de los pelos es
amplísima, pueden presentarse pelos tectores y/o glandulares. Cannabis
sativa, la marihuana, presenta numerosos tricomas glandulares y no
glandulares.
VII. DIFERENCIAS ENTRE PLANTAS MONOCOTILEDONEAS Y
DICOTILEDONEAS.
Monocotiledóneas Dicotiledóneas
Un cotiledón (a veces el embrión
indiferenciado)
Dos cotiledones (a veces 1, 3 Ó4, ó el
embrión indiferenciado)
Arboles (cuando hay crecimiento
arbóreo) poco frecuentes. formados
por gigantismo primario
Arboles frecuentes, formados por
acción de un meristema secundario
Crecimiento simpodial frecuente Crecimiento simpodial poco frecuente
Raíces sin crecimiento secundario Raíces usualmente con crecimiento
secundario
Homorrizia Alorrizia por lo general manifiesta
Hojas usualmente simples Hojas compuestas de común
ocurrencia
Profilos adaxiales Promos usualmente laterales
Una hoja por nudo Una, dos o varias hojas por nudo
Venación paralela (o estriada) Venación por lo general reticulada
Extremos de las venas cerrados Extremos de las venas abiertos (o
libres)
Base foliar ancha Base foliar por lo general angosta y
no envolvente

Estípulas ausentes Estípulas ausentes o presentes
Meristema intercalar en el entrenudo y
la base foliar por lo general presente
Meristema intercalar por lo general
ausente
Haces vasculares esparcidos,
dispuestos radialmente, es decir,
presentes en la médula y la corteza; a
veces formando dos o más anillos
Haces vasculares en un anillo simple,
no penetrando radialmente, rara vez
presentes en la médula y/o la corteza
Haces vasculares cerrados Haces vasculares abiertos
Cambium interfascicular ausente Cambium interfascicular usualmente
presente
Plástidos tipo P en los elementos
crihosos
Plástidos tipo S en los elementos
cribasos
Perianto y androceo usualmente
trímeros
Perianto y androceo usualmente
tetrámeros o pentámeros
Desarrollo de la pared de la antera
tipo "monocotiledóneas"
Desarrollo de la pared de la antera
tipo "dicotiledóneas"
Nectarios septales frecuentes Nectarios septales ausentes
Polen uniaperturado o derivado de
polen uniaperturado
Polen usualmente triaperturado o
derivado de polen triaperturado
Microsporogénesis usualmente
suceslva
Microsporogéllesis usualmente
simultánea

VIII. PLANTAS CON FOTOSINTESIS C4
Las plantas C4 tienen modificado el mesófilo de las hojas y la incorporación del CO2 en
moléculas intermedias de cuatro carbonos (de ahí viene su nombre) como el malato y el
aspartato, a fin de evitar la fotorespiración a baja tensiones de CO2 (menos de 50 ppm)debido
a que la enzima PEP-carboxilasa muestra una mayor afinidad por el CO2 que la
rubisco.y reducen la pérdida de vapor de agua por vía estomática.
Aquí el primer producto fotosintético contiene 4 carbonos: el ácido oxalacético
y sus sales. Las plantas se han adaptado lentamente a la naturaleza en los
climas áridos y cálidos. Han desarrollado una estrategia para optimizar la
capacidad de asimilación del CO2 atmosférico, e incorporarlo a la planta. Para
ello modificaron convenientemente su genoma.
Son de origen principalmente tropical, habitan en condiciones de alta
luminosidad y altas temperaturas (ambientes xerofitos). Esto les permite
competir más eficientemente con las plantas C-3, al tener que cerrar los
estomas para economizar agua y evitar la desecación. Se caracterizan por que
en ellas el producto inmediato de la fijación del CO2 es una molécula de 4
carbonos, lo que da el nombre a este grupo. Se conocen 18 familias de
monocotiledóneas y dicotiledóneas: Amaranthaceae, Aizoaceae,
Chenopodiaceae, Compositeae, Cyperaceae, Euphorbiaceae, Gramineae,
Nyctaginaceae, Portulacaceae, Zigophyllaceae. Son numerosas entre las
gramíneas, Las más conocidas son los cereales originarios de países calientes:
maíz, sorgo, mijo y la caña de azúcar.
Secuencia básica del ciclo fotosintético de las plantas C4.

1. DIFERENCIAS MORFOLOGICAS
A nivel de la estructura de las hojas: En las c3 su anatomía clásica, los
cloroplastos se localizan en el mesófilo; en las C4, anatomía tipo KRANZ ya
que se caracteriza por la presencia de 2 coronas de células clorenquimáticas
concéntricas que rodean a los haces vasculares: células del mesófilo (externas)
y células de la vaina vascular (internas).
A nivel celular: Las células de las coronas son distintas. Las células de la vaina
vascular son cilíndricas, sin espacios de aire entre ellas; Las células del
mesófilo tienen un abundante contacto una con otra, y con espacios de aire
entre sí, pero mucho más pequeño que los que se encuentran en el mesófilo
lagunoso en las hojas de las C3.
Las plantas C4 necesitan relativamente menos agua para la producción de materia orgánica que las plantas C3

A nivel de los cloroplastos: dimorfismo de los cloroplastos de estas 2 coronas
debido a la especialización. Este dimorfismo se debe aúndimorfismo
estructural y/o dimorfismo en el tamaño.
El dimorfismo estructural: los cloroplastos de la capa del mesófilo
tienen gran cantidad de discos tilacoideos apilados en granas, lo
cual les permite llevar a cabo la Fase luminosa de la fotosíntesis; los
cloroplastos de la corona perivascular carecen de grana y presentan
escasos tilacoides, encontrándose en el estroma de los mismos gran
cantidad de gránulos de almidón, producto de su especialización en
la Fase oscura de la fotosíntesis.
El dimorfismo en el tamaño: varíaen el tamaño del estroma de los
cloroplastos de la vaina vascular en relación a los cloroplastos del
mesófilo.
Las plantas C4 necesitan relativamente menos agua para la
producción de materia orgánica que las plantas C3
Las características fundamentales de los vegetales C4 que constituyen, en su
conjunto, lo que se ha dado en denominar síndrome C4 son las siguientes:
Secciones transversales de una hoja mostrando las diferencias
anatómicas entre las plantas C3 y C4.

1. Diferentes actividades enzimáticas (por ejemplo, PEPCasa
fotosintética C4) para la asimilación del CO2,NO3 y SO4.
2. Anatomía foliar general tipo kranz o en corona, como mínimo, por
dos tipos de tejidos fotosintéticos diferentes.
3. Altas relaciones chl a/b situación típica habitual de las plantas
heliófilas y de las hojas de sol.
4. Elevadas tasas de crecimiento y de fotosíntesis neta cuando las
plantas se hallan situadas en sus ambientes óptimos de elevada
temperatura, irradiancia y déficit hídrico.
5. Bajos niveles de las tasas mensurables de fotorrespiracion, incluso
en condiciones favorables para esta.
6. Insensibilidad de las tasas de fotosíntesis a la variación de las
concentraciones de O2.
7. Insensibilidad de la fotosíntesis a la oscilación de las
concentraciones de CO2.
8. Mínima concentración de compensación fotosintética (punto de
compensación) de CO2.
9. Nulo desprendimiento de O2 a la luz.
10. Insaturacion de las tasas de fotosíntesis, incluso a las máximas
intensidades solares registradas en las zonas más iluminadas de la
superficie de nuestro planeta.
11. Óptimos térmicos elevados de sus enzimas.
12. Valores elevados de C.
13. Marcada reducción de las tasas de consumo hídrico por unidad de
materia seca producida.
Características Plantas C3 Plantas C4
saturación luminosa saturación a < 300 W/m2
no se satura
tasa máxima de fotosíntesis neta (mg CO2/dm2 de área de hoja/h)
15-35 40-30
temperatura óptima para la fotosíntesis (°C) 15-25 30-45concentración de CO2 en la fotosíntesis (ppm CO2)
30-70 0-10

fotorrespiracíón alta bajatasa de transpiración (g H2O/g de peso seco)
450-950 250-350
tasa de crecimiento (g peso seco/dm2 de área de hoja)
0,5-2 3-5
2. DIFERENCIAS FISIOLOGICAS
En las plantas C4 los cloroplastos de la corona perivascular, sus tilacoides
carecen del fotosistema II por lo tanto están incapacitados para llevar a cabo la
fotofosforilación no cíclica.
En las plantas tipo C3 el proceso de fotosíntesis se lleva a cabo en sus 2
fases en un mismo cloroplasto de una misma célula.En el caso de las plantas
tipo C4, las fases del procesos de la fotosíntesis se lleva a cabo en
cloroplastos de células diferentes, es decir los cloroplastos de las células del
mesófilo se encargan principalmente de la fase luminosa, mientras que los
cloroplasto de la vaina perivascular se encargan principalmente de la fase
oscura.
Las plantas C4 se diferencian de las C3porque en ellas previo al Ciclo de
Calvin se lleva a cabo una ruta bioquímica conocida como Ciclo de Hatch y
Slack; el cual se caracteriza porque en este ciclo la molécula aceptora del
CO2(C1) es una molécula de 3C dando lugar a un compuesto de 4 Carbonos lo
que da el nombre al grupo.
En el Ciclo de Hatch y Slack, el CO2 es unido a la molécula de PEP (C3) por
una enzima diferente a la que se presenta en el Ciclo de Calvin y que
corresponde a la PEP carboxilasa, la cual da lugar a OA.
3. DIFERENCIAS BIOQUIMICAS
El oxalacetato se reduce a malato por acción de la NADP+ malato
deshidrogenasa o se convierte en aspartato por acción de la aspartato amino
transferasa.

Los ácidos dicarboxílicos abandonan los cloroplastos de las células del
mesófilo, y difunden hacia las células de la vaina o corona perivascular y a
nivel del citoplasma de estas células se descarboxilan por acción de una de un
total de 3 enzimas descarboxilasas diferentes:
a) la enzima málica NADP+ dependiente
b) la enzima málica NAD+ dependiente
c) la enzima PEP carboxiquinasa.
Por este motivo las plantas C4 han sido divididas en 3 subgrupos según el tipo
de enzima utilizado. La acción de cualquiera de estas enzimas permite la
liberación del CO2 y aumenta su concentración en las células de la vaina
vascular, allí el CO2 es utilizado por los cloroplastos de éstas células siendo
incorporado al Ciclo de Calvin.
A este nivel el ATP necesario en el Ciclo de Calvin proviene de la
fotofosforilación cíclica, y el NADP+H+ proviene de las deshidrogenación
descarboxilativa del ácido málico.
La acción de las 2 descarboxilasas málicas producen además piruvato a nivel
de las células de la vaina vascular, el cual difunde a las células del mesófilo en
donde se trasforma en PEP por acción de la enzima piruvato fosfato dikinasa.
Esta separación de las 2 fases de la fotosíntesis en sus compartimientos
determina que en las plantas tipo C4 la magnitud del proceso conocido como
fotorespiración sea prácticamente no detectable.
Estas diferencias a nivel anatómico, fisiológico y bioquímico determinan:
Que las plantas C4 presentan una mayor eficiencia fotosintética que
las plantas C3.
Que las plantas C4 tiene un mayor costo de energía que las C3.
Por cada molécula de CO2 fijado en la ruta C4, una molécula de PEP
debe de ser regenerada a expensas de 2 enlaces de alta energía, de
los grupos fosfato del ATP. Por lo cual las plantas C4 requieren un

total de 5 ATP para fijar una molécula de CO2, mientras que las
plantas c3 requieren de sólo 3 ATP.
La asimilación de CO2 a nivel de hexosa tiene un requerimiento de
ATP/NADPH de 18/12 es decir 1.5 en plantas c3 y de 30/12 o sea 2.5
en plantas C4.
El Descubrimiento De La Vía Bioquímica C4
Los biólogos a veces se alejan del o que sucede en la naturalezaporque
trabajan solo con algunos tipos de organismos que crecen fácilmente ene l
laboratorio. Los primeros experimentos sobre las reacciones
fotoindependientes del a fotosíntesis son un buen ejemplo de tal error. Estos
experimentos se realizaron sobre algas unicelulares y espinaca en los últimos
años de la década de 1940, principalmente en la Universidad de California,
Berkeley. Se reportó que las algas y la espinaca usaban la misma vía
bioquímica para construir azucares a partir de dióxido de carbono. Esta vi ase
llamo el ciclo de Calvin-Benson en honor a sus descubridores.
Las algas y la espinaca no están estrechamente relacionadas. Las primeras
son organismos unicelulares acuáticos y la segunda, un organismo multicelular
terrestre. Dado que estos organismos muy diferentes tenían la misma vía
bioquímica, los biólogos supusieron que todas la salgas y las plantas tienen la
misma vía bioquímica. La hipótesis parecía tan razonable que nadie la
cuestiono ene se momento.
Investigaciones adicionales sobre el ciclo de Calvin-Benson, sin embargo,
revelaron un fenómeno peculiar: el ciclo es inhibido por el oxígeno liberado del
as reacciones dependientes del a luz. Esta observación no tenía sentido: ¿por
qué un producto del a primer aparte de la fotosíntesis inhibiría una reacción en l
asegunda parte?.
El gas oxígeno inhibía el ciclo de Calvin-Benson especialmente cuando las
condiciones son cálidas y secas. La razón radica en que las plantas cierran
parcialmente las aperturas minúsculas, llamadas estomas, bajo estas
condiciones para reducir la evaporación del agua. Un efecto adiciona les que
muy poco dióxido de carbono puede entrare n las hojas y muy poco gas

oxígeno puede salir. La concentración de dióxido de carbono disminuye y la
concentración de gas oxígeno aumenta.
Cuando el oxígeno llega a ser más abundante que el dióxido de carbono dentro
del cloroplasto, bloque a la enzima que normalmente une el dióxido de carbono
con RuBP. El oxígeno entra, en lugar del dióxido de carbono, al ciclo de Calvin-
Benson, el ciclo se cierra y la síntesis de azúcar se detiene.
Si todas las plantas tienen el ciclo de Calvin-Benson y si el gas oxigeno inhibe
el ciclo, ¿Cómo sintetizan las plantase l azúcar mientras sus estomas están
cerrados? ¿Cómo, por ejemplo, crece el maíz tan rápidamente durante los
veranos secos y calientes?
Algunos investigadores sugirieron que quizás las plantas en los climas secos y
calientes no usan el ciclo de Calvin-Benson. Cuestionaron la hipótesisde que
todas las plantas y la salgas usan el ciclo y comenzaron a busca runa vía
bioquímica diferente: una que no fuera inhibida por el gas oxígeno. Tal vía fue
encontrada por en la década de 1960 por M. D. Hatch en Australia y C. R.
Slack en Inglaterra. Ellos descubrieron que la caña de azúcar, una planta que
crece en climas secos y calientes, fija carbono (es decir, une el dióxido de
carbono con una molécula orgánica) de una forma diferente del a fijación de
carbono e nalgas y espinacas.
La vía de Hatch-Slack evita la inhibición del oxígeno al fijare l carbono en las
células del a hoja que no capturan la energía lumínica, no desarrollando así
altas concentraciones de oxígeno. Además, la vía de Hatch-Slack usa una
enzima diferente para obtener dióxido de carbono y unirlo al RuBP. Esta
enzima se une únicamente con el dióxido de carbono, aunque sea mínima su
cantidad en l ahoja; y no se une con el gas oxígeno. El dióxido de carbono
fijado en las células no fotosintetizadoras se bombea en cantidades masivas a
las células fotosintetizadoras, donde monopoliza la enzima que la une con el
RuBP. Por consiguiente, el oxígeno, en vez del dióxido de carbono, se une al
RuBP y el ciclo de Calvin-Benson se activa rápidamente, aunque el resto
estuviera prácticamente cerrado.
Estas dos maneras de recobrare l dióxido de carbono son conocidas como las
vías C3 y C4. En la vía C3, la primera molécula estable (ácido fosfoglicérico)

después del a fijación del carbono tiene tres átomos de carbono. La mayoría
del as plantas usan esta vía.
En la vía C4, la primera molécula estable tiene cuatro átomos de carbono.
Muchas plantas en los climas secos y calientes usan esta vía (una tercer
amanera de fijar carbono, conocida como metabolismo ácido crasuláceo
(CAM), se ha descubierto en cactus yo tras plantas que viven en desiertos. Es
muy similar a la vía C4, pero el CAM del as plantas reduce la perdida de agua
al abrir sus estomas únicamente en l anoche.
La vía C4, es particularmente común el pastos, una categoría de plantas de
incluyen el maíz y la caña de azúcar. La ventaja del a vía C4 en los climas
secos y cálidos es que permite a la planta conservare l agua al cerrar
parcialmente sus estomas mientras se construye el azúcar.
IX. PLANTAS C3
1. Definición.
Este tipo de plantas poseen el mecanismo fotosintético más básico. Plantas en
las que el CO2 se fija inicialmente mediante el ciclo de Calvin en la fotosíntesis.
La mayoría de las especies, en especial aquellas que pertenecen a ambientes
húmedos y templados, son de este grupo. Ejemplos típicos lo constituyen la
mayoría de los árboles y cultivos tales como el trigo, el arroz, la cebada, la
papa, la soya y la yuca.
TRIGO SOYA
YUCA PAPA

2. Características
La ruta metabólica C3 se encuentra en los organismos
fotosintéticos como las cianobacterias, algas verdes y en la
mayoría de las plantas vasculares.
Requieren de mayor agua para la producción de materia
inorgánica.

X. LA BIOINGENIERÍA PUEDE CONVERTIR LAS C3 EN C4
Las mayores empresas de bioingeniería hacen fuertes inversiones para
lograrlo. En 2007 Monsanto y BASF reunieron 1500 millones de dólares, para
lograr semillas que resistan la sequía. Se trata de transferir a la semilla un gen
tomado de una bacteria. La planta industrial será operativa en 2012. Empresas
con miles de empleados, que manipulan un gen microscópico.
Luego vendrá la nueva semilla de soja, que producirá más aceite que la actual.
Y algodón resistente a la sequía, y cereales que usen con más eficiencia el
Nitrógeno. Syngenta, Du Pont y Bayer Crop Science se han sumado a la
investigación.
No hacen más que imitar la evolución de la naturaleza en dotar del mecanismo
C4 a ciertas plantas C3. No podemos reprocharles nada. Los gobiernos
legislan sobre los alimentos transgénicos, precisamente porque son
necesarios. Hacen falta mejores cosechas con menos agua. ¿Tan difícil es
añadir un gen de una célula donante al genoma de una célula receptora, un
cereal?
Recordemos algo elemental: desde principios de los años 70 disponemos de
herramientas moleculares, que permiten cortar el ADN. Se llaman enzimas de
restricción, y cortan el ADN en zonas específicas del genoma. Además
disponemos de otras enzimas llamadas Ligasas ADN, que permiten pegar
trozos de ADN cortados. Por lo tanto podemos cortar el genoma de una célula
donante con un determinado enzima de restricción, tomar el fragmento de ADN
que contenga el gen que buscamos, y pegarlo en el genoma de la célula
receptora, formando un ADN recombinante.

Este último paso se suele llevar a cabo con la ayuda de una molécula de ADN
transportadora, a la que llamamos vector. Para ello precisaremos disponer de
una técnica, genéricamente llamada transformación genética, que permita
introducir la información genética manipulada in Vitro en la célula receptora. Si
hemos tomado el gen donante desde un organismo determinado, y lo
expresamos en otro, por ejemplo, el gen de un animal-microbio en un cereal, al
organismo resultante se le llama transgénico.
Durante miles de años el biotecnólogo de alimentos estuvo limitado en sus
labores de selección por los cruces sexuales. Siempre y cuando dispongamos
de los genes, podemos expresar los de una especie en otra. Necesitamos usar
de un método de transformación genética que permita introducir ADN foráneo.
En el caso de los vegetales (cereales) no es una labor sencilla, ya que las
células vegetales están recubiertas de una gruesa capa de celulosa y
hemicelulosa, llamada pared celular, que actúa como una barrera impidiendo la
entrada del ADN exógeno. Para introducirlo usamos una bacteria:
Agrobacterium tumefaciens. Con ella saltamos la barrera de la pared celular.
Desgraciadamente algunas variedades vegetales comestibles, como la banana,
el maíz, o el trigo no aceptan el Agrobacterium. En estos casos se utilizan
enzimas líticos, que degradan la celulosa y la hemicelulosa.
Favorecido por la ausencia de pared celular, el vector entra dentro de la célula,
y se integra en el genoma de un protoplasto receptor. Posteriormente en unas
condiciones adecuadas de cultivo, los protoplastos pueden regenerar la pared
celular, e incluso dan lugar a una planta adulta transformada.
Copiar, cortar y coser. La sastrería genética es un arte que requiere el uso de
centenares de herramientas: son proteínas enzimáticas (enzimas) que
catalizan reacciones químicas en la célula viva. En los tiempos heroicos el
sastre tenía que prepararse sus propias herramientas, pero hoy se compran en
el comercio, que las ofrece en catálogos muy extensos. Comercializar un
alimento transgénico requiere una autorización del gobierno. Es un alimento
seguro.

Es urgente lograr mejores cosechas con menos agua, para alimentar a una
población mundial en aumento. Las especies vegetales C3 que nos sirven de
alimento requieren demasiada agua. Conseguir su conversión a C4 es una
meta difícil, pero asequible, no es una quijotada.
XI. CONCLUSIONES.

XII. Bibliografía.
Albert, A.: Libro verde de la biotecnología en la agricultura. Ed. Soc. Esp.
de Biotecnología 1998.
Casal, I.: La biotecnología aplicada a la agricultura. Ed. Soc. Esp. de
Biotecnología 2000.
Evans, J.: Food security is all in the genes. Chemistry and Industry 10
May 2010.
Evans, J.: Better than Nature? Photosynthesis process. Chemistry and
Industry 11 may 2010.
Ferro, A.: Alimentos transgénicos. Ed. McGrawHill 2000.
García Olmedo, F.: La tercera revolución verde. Ed. Debate SA 1998.
Gómez, L.: Biotecnología y Alimentos. Ed. Soc. Esp. de Biotecnología
2003.
Martínez-Gil, F. (1995). Elementos de Fisiología Vegetal. MP. México
Ramón, D.: Los genes que comemos. Ed.Algar 1996.
Salisbury, F.B. y Ross C.W. (1994) Fisiología Vegetal. Grupo Editorial
Iberoamérica. México.
XIII. Linkografía.
http://www.xtec.cat/~cgarci38/ceta/tecnologia/fotosintesis.htm.
http://www.plantprotection.hu/modulok/spanyol/maize/index.htm.
http://biologia-jct.iespana.es/curtis/libro/c36b.htm.