computational neuroscience rezeptive felder - uni … · computational neuroscience rezeptive...
TRANSCRIPT

Computational NeuroscienceRezeptive Felder
Jutta Kretzberg
Master-Grundmodul 2009
Download der pdf Version:http://www.uni-oldenburg.de/sinnesphysiologie/

Computational Neuroscience
Themen:Mechanosensoren des BlutegelsRezeptive Felder im visuellen SystemEntstehung rezeptiver Felder
Analysemethode:Tuning-Kurven
Modelle:Grundlegende VerschaltungsmusterBewegungsdetektion
DatenanalyseModell-
simulationen
Experimentelle Daten
formale Beschreibung
Heute: Rezeptive Felder

Definition: Rezeptives Feld
Das rezeptive Feld einer Sensorzelle ist der räumliche Bereich, in dem das Vorhandensein eines physikalischen Reizes die elektrische Aktivität der Sensorzelle beeinflusst.
Die rezeptiven Felder aller Sensorzellen, die synaptisch mit einem nachgeschalteten Neuron des Nervensystems verbunden sind, bilden zusammen das rezeptive Feld dieses Neurons.

Das Nervensystem des Blutegels
21 sehr ähnliche Körpersegmente
In jedem Ganglion ca. 400 individuell bekannte Neurone
Überschaubare Neuronennetze steuern Verhalten
Weisblatt & Kristan 1985

Mechanosensoren des Blutegels
3 Typen von Mechanorezeptoren, die auf Berührung der Haut reagieren:
T-Zellen (touch)P-Zellen (pressure)N-Zellen (noxious)
Soma im Ganglion, Zellprozesse in der Haut

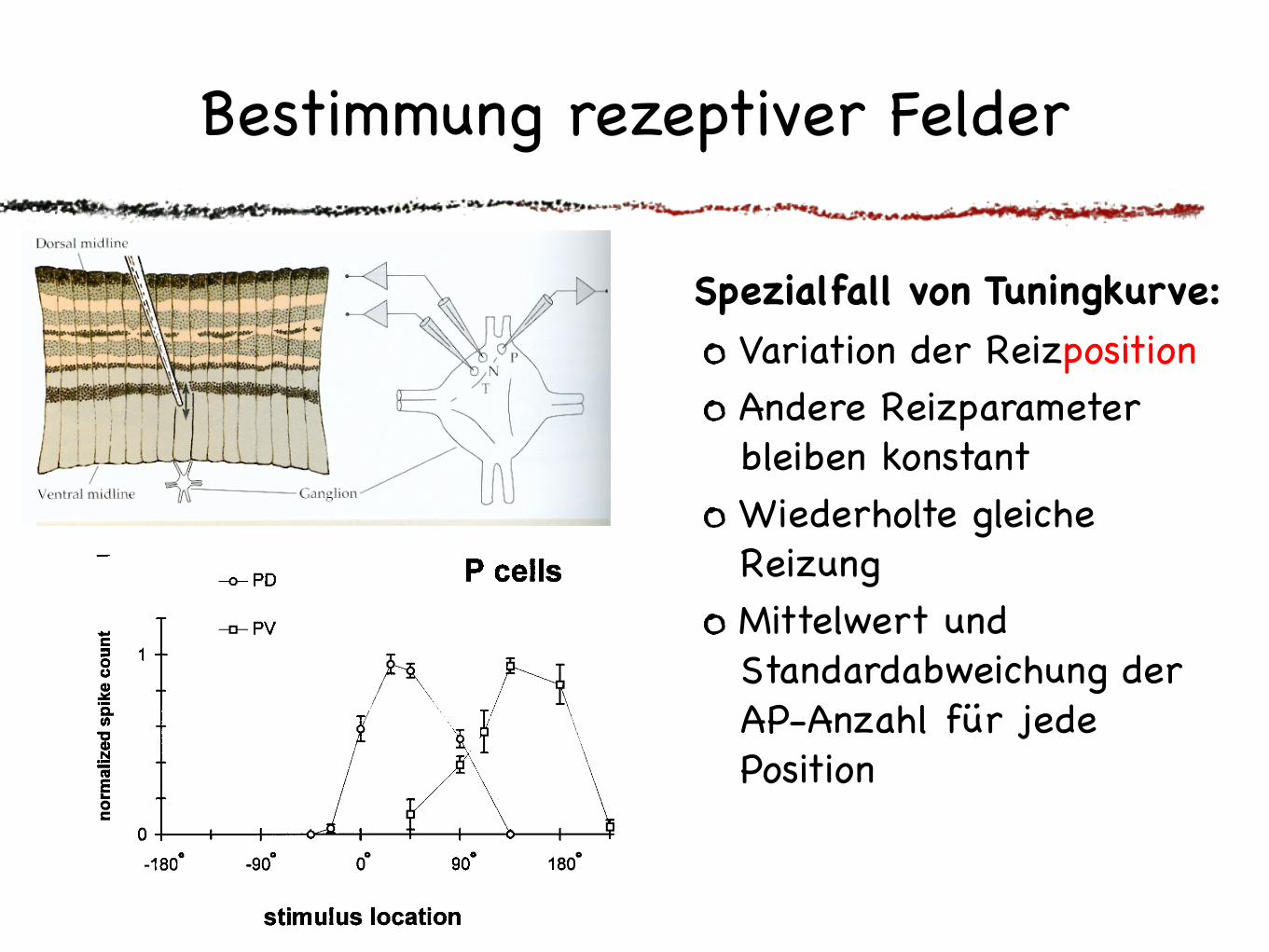
Methode: Tuningkurve
1g
7g
21g100 101 102
0
5
10
15
20
Stimulusstaerke [g]
P−Ze
ll Ant
wort
[AP−
Anza
hl]
Nichollset al. 2001
Variation der Stärke einer ReizeigenschaftAndere Reizparameter bleiben konstant

0 50 100 150 200 2500
5
10
15
20
Zeit [ms]
Reizp
raes
enta
tione
n
AP-Zeitpunkte bei Reizung mit 4g
100 101 1020
5
10
15
20
Stimulusstaerke [g]
P−Ze
ll Ant
wort
[AP−
Anza
hl]
Methode: Tuningkurve
Variation der Stärke einer ReizeigenschaftAndere Reizparameter bleiben konstant

Achtung! Variabilität!
Wiederholte gleiche Reizung führt zu unterschiedlichen Antworten!
0 50 100 150 200 2500
5
10
15
20
Zeit [ms]
Reizp
raes
enta
tione
n
100 101 1020
5
10
15
20
Stimulusstaerke [g]
P−Ze
ll Ant
wort
[AP−
Anza
hl]
Raste
rplot

Variation der Stärke einer ReizeigenschaftAndere Reizparameter bleiben konstantWiederholte gleiche Reizung Mittelwert und Standard-abweichung der AP-Anzahl für jede Reizstärke
TOUCH REPRESENTATION BY LEECH SENSORY NEURONS 2589
a greater extent and produce other behavioral responses.] Byusing the same set of stimuli in a different group of fiveanimals, we found that the mean tension response increaseswith stimulus intensity in a manner similar to P-cell activa-tion but quite different from that of T-cell activation. TheT-cell spike count saturated at stimulus intensities ú1 mN;at this intensity, the behavioral response was only 25% ofthe response produced by the 37-mN stimulus. In agreementwith a previous study with a different approach (Kristan1982), this result suggests that P cells provide the majorinput for controlling the magnitude of the local bend re-sponse.However, it is still possible that the T cells provide spatial
FIG. 6. Mechanosensory neuron and behavioral tuning to stimulus inten- information, but their influence is gated by the P cells ( i.e.,sity. Stimuli of different intensities were delivered at S Å 45!. Normalized the information is not manifest in the behavior unless the Presponses (mean { SE; 5 animals) are plotted vs. stimulus intensity. Re-
cells are sufficiently active) . To test this possibility, wesponses of the T and P cells were measured by the normalized spike count,attempted to bias the tension response (measured at 45!)and the behavioral response was measured by normalized peak body wall
tension. evoked by a mechanical stimulus (S Å 45!; 22 mN) withsimultaneous intracellular stimulation of the TV cell (ú20Hz, 1-s trains) . If the T cells provide spatial information,extent to which their tuning curves overlap. In another study,the evoked tension should be diminished by TV stimulationwe characterized the local bend response as a function ofcaused by an effective shift in the bend direction to a morestimulus location (Lewis and Kristan 1998a). The RMSventral location (toward the preferred location of the TVdifference between stimulus location and the direction of thecell) . If the T cells provide nonspecific amplification to theevoked bend (i.e., behavioral error) was 8% and is shownlocal bend response, the tension response should increase.in Fig. 5B for comparison. With the stimulus encoding byWe found that TV stimulation had no effect on the tensionthe sensory neurons more than twice as accurate as the be-response measured at 45! (P ú 0.05, 2 animals, 8 trials) ,havior, it is clear that a neural code for stimulus locationindicating that the T cells play no detectable role in thisinvolving only the spike count contains more than enoughresponse. In similar experiments, intracellular P-cell stimula-information to account for the behavior.tion produced a significant and predictable effect on thebehavior (Lewis and Kristan 1998b). From these and previ-Relative importance of T and P cellsous results, we conclude that the P cells are the major carriersof spatial information to the local bend network.Because both T and P cells independently encode stimulus
location with an accuracy greater than the behavioral accu-racy, we tested whether the encoded information from each Varying encoding timecell type is actually used by the local bend networks.The thresholds and specific modalities differ for each of In Fig. 5 we showed that the decoding error with P-cell
spike trains wasÇ3%. These estimates were made for spikesthese cell types; T cells have a low threshold and are mostsensitive to the velocity of indentation of the body wall by occurring for an entire 700-ms interval after stimulus onset.
The latency for a behavioral response (muscle tension) ismechanical stimulation (Carlton and McVean 1995),whereas P cells have a higher threshold and respond best to typically Ç200 ms. How much information is available at
this short latency? To address this question, we calculatedchanges in the magnitude of body wall indentation. To testthe relative contributions of T and P cells to local bending, the mean decoding error for different encoding times (i.e.,
time since stimulus onset) . An encoding time of 700 mswe quantified the behavioral output and response of the sen-sory cells to mechanical stimuli of different intensities. We corresponds with the control condition (Fig. 5) . We mea-
sured the tuning curves and spike count variability again butlimited our studies to one stimulation site and one body walltension measurement site (45! for both) . At this stimulus only considered spikes that occurred in the time window
corresponding to a particular encoding time. For all encodinglocation, one P cell (right PD) and two T cells ( the right TDand TL) are activated to ú70% of their maximum level, and times tested, the tuning curves, normalized to maximum re-
sponse, were not significantly different from the controla third T cell ( the left TD) is activated to Ç25% of itsmaximum (see Fig. 5A) . The remaining cells are not acti- (Pú 0.05). In addition, the variability in the normalized
spike count for the different integration times was not sig-vated.Figure 6 shows the normalized spike count of the PD and nificantly different from control (P ú 0.05). This means
that the tuning curves in all cases differ only by a scalingTD (mean { SE, 5 animals) . For stimulus intensities ú1mN, the TD-cell spike count does not change significantly. factor ( i.e., the total number of spikes; see Fig. 7A) ; for
longer encoding times, more spikes are available to contrib-The PD-cell spike count increases substantially over the rangeof 1–37 mN (Fig. 6) . [A third type of mechanosensory ute information.
Again, with the tuning curves and the variability of theneuron, the high-threshold N (nociceptive) cell, never pro-duced more than one spike for the largest stimulus intensity responses, we estimated the decoding error for different encod-
ing times (Fig. 7B). At 100 ms, the error decreases belowshown here. We did not investigate stimulus intensitiesgreater than these because the N cells would be activated to that found in the behavior, and interestingly counting spikes
J131-8/ 9k2e$$no30 10-26-98 22:35:29 neupa LP-Neurophys
Lewis & Kristan 1998
Methode: Tuningkurve
J. E. LEWIS AND W. B. KRISTAN2586
stimulus location around the body perimeter. Figure 2 showstuning curves of normalized spike count versus stimuluslocation for the right T and P cells. These curves representthe averages over six different animals. Both T and P cellsexhibit broad and overlapping tuning curves, with peaksdistributed around the body perimeter. By using the commonconvention, we refer to the stimulus location that producesthe maximum response for a given cell as that cell’s pre-ferred location. The preferred locations serve to identify thedifferent cells; TD and PD are activated most by dorsal stim-uli, TV and PV by ventral stimuli, and TL by lateral stimuli.The shapes of these tuning curves, along with the periodic
nature of the dependent variable (i.e., stimulus location, S ,on the body perimeter varies between 0180 and /180!) ,suggest that a cosine function might be an appropriate sum-
FIG. 1. Typical mechanosensory spike trains. Intracellular recordings mary of the data from both cell types. We fit the means forfrom the T and P mechanosensory neurons during a 500-ms mechanical each cell from each individual animal to the cosine functionstimulus to the body wall (22 mN). The T cell exhibits rapidly adapting
in Eq. 2responses at both stimulus onset and offset, whereas the P cell shows acomparatively tonic response.
f(S) Å Rmax
1 0 A{cos (S 0 So) 0 A} (2)
in the population of sensory neurons. Theunissen and Miller deriveThis formulation was used previously (e.g., Miller et al.
an expression that describes the transinformation, T , as a function1991) and is convenient because Rmax is the maximum re-of the RMS error, e (see Eq. 1 following, and Eq. 11 in Theunissen
and Miller 1991)
T Å log2 !!2p
e " 0 1
2 ln (2)(1)
The wind-sensitive interneurons in the cricket cercal system de-scribed by Theunissen and Miller are completely analogous to themechanosensory neurons described in this study. Here we use Eq.1 to provide a quantitative measure of the information content inthe population responses of the sensory neurons and thus providean additional interpretation of the encoding error.
R E S U L T S
A mechanical stimulus that elicits the local bend behavioractivates two types of mechanosensory neurons, T cells andP cells (Kristan et al. 1982). In a single segment there aresix T cells and four P cells. Figure 1 shows typical spiketrains produced by the mechanical stimulus used in ourstudy. The T cell adapted rapidly and fired at the onset andoffset of the stimulus. Recently, Carlton and McVean (1995)proposed that the T cells encode the velocity of skin indenta-tion. The T-cell firing dynamics are similar to those shownby rapidly adapting (RA) afferents in humans (Vallbo andJohansson 1984; Wheat et al. 1995). The P-cell spike trainshowed a slowly adapting (SA) response to a mechanicalstimulus, which is similar to responses of the human SAafferents (Vallbo and Johansson 1984; Wheat et al. 1995).To quantify the responses of the T and P neurons, we simplycounted the number of spikes occurring in the 700-ms periodafter onset of a 500-ms stimulus (unless otherwise noted).In doing this, we ignore the temporal pattern of the neuralresponses.
Spatial tuning: response as a function of stimulus locationFIG. 2. Mechanosensory neuron tuning curves to stimulus location.
Mean normalized spike count is plotted for different stimulus locations forTo assess the ability of the T and P cells to encode the
the (A) 3 types of T cells and (B) 2 types of P cells (mean { SE, 6location of a mechanical stimulus on the skin, we first char- animals) . The mean spike counts at the tuning curve peaks were 15.1 (T
cells) and 7.2 (P cells) .acterized their average response properties as a function of
J131-8/ 9k2e$$no30 10-26-98 22:35:29 neupa LP-Neurophys
Variation der Reizposition Andere Reizparameter bleiben konstantWiederholte gleiche Reizung Mittelwert und Standardabweichung der AP-Anzahl für jede Position
Bestimmung rezeptiver Felder
Spezialfall von Tuningkurve:

Wie entstehen räumlich inhomogene rezeptive Felder?
Häufig löst der gleiche Reiz an verschiedenen Stellen im rezeptiven Feld unterschiedlich starke Reaktionen aus. Ein Grund dafür ist die inhomogene dendritische Stuktur (z.B. dichtere Verzweigung in der Mitte) => Entstehung auf zellulärer Ebene Blackshaw
1981
Dendritische Verzweigungen der T-Zelle in
der Haut

Wie kodieren Blutegel den Ort einer Berührung?
An jeder Stelle des Körpers überlappen sich die rezeptiven Felder von zwei P-Zellen.
P1 P2 P3 P4
P1
P2 P3
P4
Thomson &Kristan 2006

Wie kodieren Blutegel den Ort einer Berührung?
Rate und Latenz hängen vom Ort der Berührung ab.
P1 P2 P3 P4
P2
P3
Thomson &Kristan 2006

Wie kodieren Blutegel den Ort einer Berührung?
Vier Kodierungsarten zeigen eine signifikante Abhängigkeit der Antworten vom Berührungsort.
absolute Latenz
relative Latenz
Rate einer Zelle
relative Rate
stärkste Abhängigkeit Thomson &Kristan 2006

Ein idealer Beobachter kann den Ort einer Berührung am besten aus der relativen Latenz der P-Zell-Antworten schliessen.
Um zu wissen, was der Egel tut, muss man die nachgeschalteten Zellantworten kennen.
latencybehaviorspike count
Wie kodieren Blutegel den Ort einer Berührung?
Thomson &Kristan 2006

Rezeptive Felder im visuellen System
Retinale Ganglienzellen und LGN: meist runde Zentrum-Umfeld Struktur
Visueller Cortex:Simple cells: Meist langgestreckte Struktur mit erregendem und hemmendem AnteilComplex cells: Empfindlich für bestimmte BewegungsrichtungHypercomplex cells: Empfindlich für bestimmte Bewegungsrichtung und Größe des bewegten Objekts

Tuning für Bewegungsreize
Complex Cell:Bewegung in Vorzugsrichtung löst viele APs aus, Bewegung in Gegenrichtung keine
nach Hubel & Wiesel
Retina:Bei manchen Tieren gibt es Bewegungstuning schon bei retinalen Ganglienzellen
!! !" # " !#
$
"#
%&'()*+,-.//012
34*5&-67+&-."012
a
!! !" # " !#
$
"#
%&'()*+,-.//012
34*5&-67+&-."012
b
!! !" # " !#
"##
!##
8##
9##
$##
:&7;-'7+&;),-./12
%&'()*+,-.//012
c
!! !" # " !#
"##
!##
8##
9##
$##
:&7;-'7+&;),-./12
%&'()*+,-.//012
d
RGC ohne Vorzugsrichtung:
RGC mit Vorzugsrichtung:
Schildkröte, eigene Daten

Hubel & Wiesel:Rezeptive Felder
http://www.science.smith.edu/departments/NeuroSci/courses/bio330/
0.00 ON2.08 OFF4.10 simple7.36 complex10.11 complex12.11 hyper-complex14.50 depth-sensitive

http://www.yorku.ca/eye/recepfld.htm
z.B. Retinale Ganglienzellen:Häufig löst der gleiche Reiz an verschiedenen Stellen im rezeptiven Feld unterschiedlich starke Reaktionen aus.
Ein Grund ist die Kombination excitatorischer und inhibitorischer synaptischer Eingänge => Entstehung auf Netzwerkebene
Wie entstehen räumlich inhomogene rezeptive Felder?

Grundlegende Verschaltungen:1:1-Verschaltung
Die Aktivität eines präsynap-tischen Neurons wird gewichtet an ein postsynaptisches Neuron über-tragen
Dabei kann durch Inhibition eine Vorzeichenumkehr stattfinden
Bedeutung:1. Prä- und postsynaptische Zelle haben gleiche rezeptive Felder2. Topologie-erhaltende Abbildung des Reizortes
prä-synaptisch
post-synaptisch
+ - + -

Grundlegende Verschaltungen:Konvergenz
Die Aktivität mehrerer präsynap-tischer Neuronen wird durch ein postsynaptisches Neuron integriertDie synaptischen Verbindungen können unterschiedlich stark seinExcitation und Inhibition (auch shunting inhibition) können gemischt auftretenDas postsynaptische Neuron nimmt eine “Mittelung” der gewichteten präsynaptischen Aktivitäten vorIn biologischen neuronalen Netzen ist die Interaktion nichtlinear
Bedeutung:1. Räumliche Vergrößerung des rezeptiven Feldes2. Komplexe Struktur des rezeptiven Feldes3. Zeitlicher Vergleich der Inputs möglich4. Erhöhte Zuverlässigkeit bei redundanten Inputs
prä-synaptisch
post-synaptisch
+ +- -

Konvergenz:Integration vs Koinzidenzdetektion
prä-synaptisch
post-synaptisch
+ +- -
Zusätzlich zur räumlichen hat Konvergenz auch eine zeitliche KomponenteWichtiger Parameter: Zeitskala der Konvergenz
lange Integrationszeiten bedeuten zeitliche Integration
kurze Integrationszeiten ermöglichen Koinzidenz-detektion
=> Ratencode
=> Zeitcode
z.B. Ermittlung von Reizintensität durch zeitlicheIntegration im visuellen Cortex
z.B. Vergleich zwischen Ohren bei Richtungshörenim auditorischen Cortex

Grundlegende Verschaltungen:Divergenz
Die Aktivität eines präsynaptischen Neurons wird an mehrere postsyn-aptische Neurone weitergegebenDie synaptischen Verbindungen können unterschiedlich stark seinEs ist abhängig von den Rezeptoren des postsynaptichen Neurons, ob sich präsynaptische Aktivität excitatorisch oder inhibitorisch auswirkt. Glutamat kann sowohl erregende als auch hemmende Synapsen bilden
Bedeutung:1. Mehrere postsynaptische Neurone mit überlappenden rezeptiven Feldern2. Erhöhte Zuverlässigkeit der Populationsantwort bei unzuverlässigen Synapsen
prä-synaptisch
post-synaptisch
+ +- -

Grundlegende Verschaltungen:Vorzeichenumkehr
Die Aktivität eines präsynaptischen Neurons wird sowohl direkt an postsynaptische Neurone, als auch an postynaptische Interneurone weitergegeben, die bei der weiteren Verschaltung das Vorzeichen umkehren
prä-synaptisch
post-synaptisch +
+
-
Bedeutung:1. Möglichkeit, Excitation und Inhibition zu kombi-nieren2. Hemmung eines “Gegen-spielers” bewirkt indirekte Verstärkung

Grundlegende Verschaltungen:komplette Vernetzung
Alle präsynaptischen Neuron sind mit allen postsynaptischen verbundenSynaptische Gewichte können unterschiedlich und Excitation und Inhibition gemischt sein (soweit Transmitterausstattung biologischer Neurone das zulässt)Normalfall für künstliche neuronale Netze, die häufig mit zufällig gewählten Gewichten einer kompletten Vernetzung initialisiert werden
Bedeutung:1. Postsynaptische Zellen können komplexe rezeptive Felder haben2. Vorteile von Konvergenz und Divergenz werden kombiniert
prä-synaptisch
post-synaptisch

Grundlegende Verschaltungen:Reziproke Vernetzung (Feedback)
Zwei Neurone sind wechselseitig miteinander verschaltetInformation fließt im Kreis, Neurone sind also gleichzeitig prä- und postsynaptisch zu sich selbst
Bedeutung:1. Aktivität hängt nicht nur von sensorischen Inputs ab2. Es können sich typische Aktivitätsmuster entwickeln3. Besonders wichtig bei Taktgeber-Netzwerken
++
--
-
Selbst-erregungSelbst-erregungSelbst-hemmung
+
z.B. StomatogastrischesGanglion:

Grundlegende Verschaltungen:Rekurrente Vernetzung (Feedback)
Allgemeiner Fall des FeedbacksMehrere Neurone sind im Kreis miteinander verbundenEs gibt also keine eindeutig geschichtete StrukturAlle Kombinationen aus Erregung und Hemmung sind möglichNeurone können an mehreren Feedback-Schleifen beteiligt seinNormalfall kortikaler VerarbeitungBei künstlichen neuronalen Netzen sehr mächtig, aber schwer analysierbar
Bedeutung:1. Netzwerk-intrinsische Aktivität überlagert stimulus-getriebene2. Definition von rezeptiven Feldern wird schwierig (oder unmöglich)

Der Normalfall: Kombination vieler Möglichkeiten

!! !" # " !#
$
"#
%&'()*+,-.//012
34*5&-67+&-."012
a
!! !" # " !#
$
"#
%&'()*+,-.//012
34*5&-67+&-."012
b
!! !" # " !#
"##
!##
8##
9##
$##
:&7;-'7+&;),-./12
%&'()*+,-.//012
c
!! !" # " !#
"##
!##
8##
9##
$##
:&7;-'7+&;),-./12
%&'()*+,-.//012
d
Asymmetrisches Geschwindigkeitstuningbei RGC der Schildkröte:
Idee von Reichardt (1961):Netzwerkinteraktion
http://www.rcs.ei.tum.de/RCS/research/mvnm/index_html/reichardt_detector
Verzögerung
Korrelation
Integration
Wie könnte Empfindlichkeit für Bewegung entstehen?

!! !" # " !#
$
"#
%&'()*+,-.//012
34*5&-67+&-."012
a
!! !" # " !#
$
"#
%&'()*+,-.//012
34*5&-67+&-."012
b
!! !" # " !#
"##
!##
8##
9##
$##
:&7;-'7+&;),-./12
%&'()*+,-.//012
c
!! !" # " !#
"##
!##
8##
9##
$##
:&7;-'7+&;),-./12
%&'()*+,-.//012
d
Asymmetrisches Geschwindigkeitstuningbei RGC der Schildkröte:
Wie könnte Empfindlichkeit für Bewegung entstehen?
Alternative: Dendritische Verarbeitungz.B. Spannungsabhängige Nichtlinearitäten in Starburst Amakrinzellen
Hausselt et al.2006

Sensorische Reize können viele Dimensionen haben. Z.B. im visuellen System
Vieldimensionale Parameterräume
Position Räumliche StrukturHelligkeit Zeitliche StrukturKontrast BewegungsrichtungWellenlänge Bewegungsgeschwindigkeit...
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Kombination der variierten Parameter: Mehrdimensionale Tuningkurven
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich
Programm
Arbeitsweise von Sensorzellen - Kodierung sensorischer Reize durch elektrische Signale
Stimulus-Response Kurven
Wie modelliert man Sensorzellen?
Integrate and Fire Modell
Neuronenmodelle im Vergleich

Zusammenfassung
Rezeptive Felder beschreiben den räumlichen Bereich, in dem ein Neuron auf physikalische Reize reagiert.
Die Antworten von Neuronen auf Reize innerhalb ihres rezeptiven Feldes können in komplexer Weise von verschiedenen Stimuluseigenschaften abhängen. Zur Beschreibung dieser Abhängigkeiten verwendet man Tuningkurven.
Die Struktur rezeptiver Felder wird sowohl durch Netzwerkinteraktionen als auch durch dendritische Verarbeitung hervorgerufen.

Literatur
pdf-Version der Folien:http://www.sinnesphysiologie.uni-oldenburg.de/40426.html
Überblick über Lehrbücher zum Thema:http://www.sinnesphysiologie.uni-oldenburg.de/20349.html
Abbildungen verwendet aus:Kandel, Schwartz, Jessel. Principles of neural science. 4th ed. McGraw-Hill Medical, 2000.Nicholls et al. From Neuron to brain. 4th ed. Sinauer, 2001.Blackshaw. Journal of Physiology 320:219-228, 2000.Thomson & Kristan. Journal of Neuroscience 30:8009-8016, 2006Hausselt et al. PLoS Biology 5:1474-1493, 2007