complejos ligando-proteÍna
DESCRIPTION
MUESTREO Y CONVERGENCIAS EN LOS CÁLCULOS DE ENERGÍA LIBRE DE LA INTERACCIÓN PROTEÍNA-LIGANDO. EL ENLACE DE LOS DERIVADOS DE LA TRIFENOXIPIRIDINA CON LA FACTOR Xa Y LA TRIPSINA. - PowerPoint PPT PresentationTRANSCRIPT

MUESTREO Y CONVERGENCIAS EN LOS CÁLCULOS DE ENERGÍA
LIBRE DE LA INTERACCIÓN PROTEÍNA-LIGANDO.
EL ENLACE DE LOS DERIVADOS DE LA TRIFENOXIPIRIDINA CON LA FACTOR Xa Y LA TRIPSINA
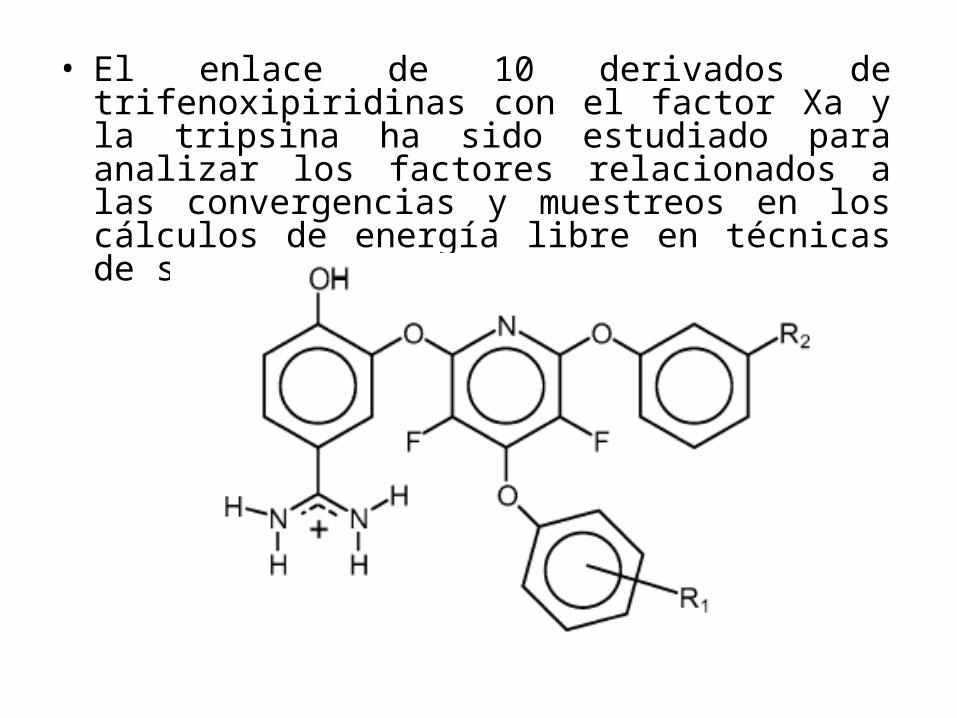
• El enlace de 10 derivados de trifenoxipiridinas con el factor Xa y la tripsina ha sido estudiado para analizar los factores relacionados a las convergencias y muestreos en los cálculos de energía libre en técnicas de simulación de dinámica molecular.

• Los inhibidores investigados estaban inicialmente propuestos en el Proyecto de Valoración Crítica de las Técnicas para la Evaluación de la Energía Libre ( CATFEE ); proyecto en el cual ni los resultados experimentales ni la evaluación de las predicciones de varios grupos han sido publicadas alguna vez.
• Los inhibidores estudiados representan un severo desafío para explicar los cálculos de energía libre. Las mutaciones de un compuesto a otro involucran:
* 19 átomos * la creación y aniquilación de la carga neta * varios modos de enlace alternativos.
• A pesar de eso, se mostrará que es posible obtener resultados favorablemente convergentes ( ±5-10 kJ/mol ) incluso para tal complejo poltiatómico simulando una escala de tiempo de un nanosegundo.

COMPLEJOS LIGANDO-PROTEÍNA• Un prerrequisito para obtener la correcta
relación de energía libre para el complejo inhibidor-proteína es que el inhibidor forme las interacciones apropiadas dentro del sitio de enlace.
• La figura 9 y la figura 10 muestran las estructuras después de 2ns de simulación con el inhibidor I1 enlazado al factor Xa y a la tripsina respectivamente. Se nota que en ambos casos la numeración de los residuos es la misma que se adopta por archivos PDB de rayos-X.
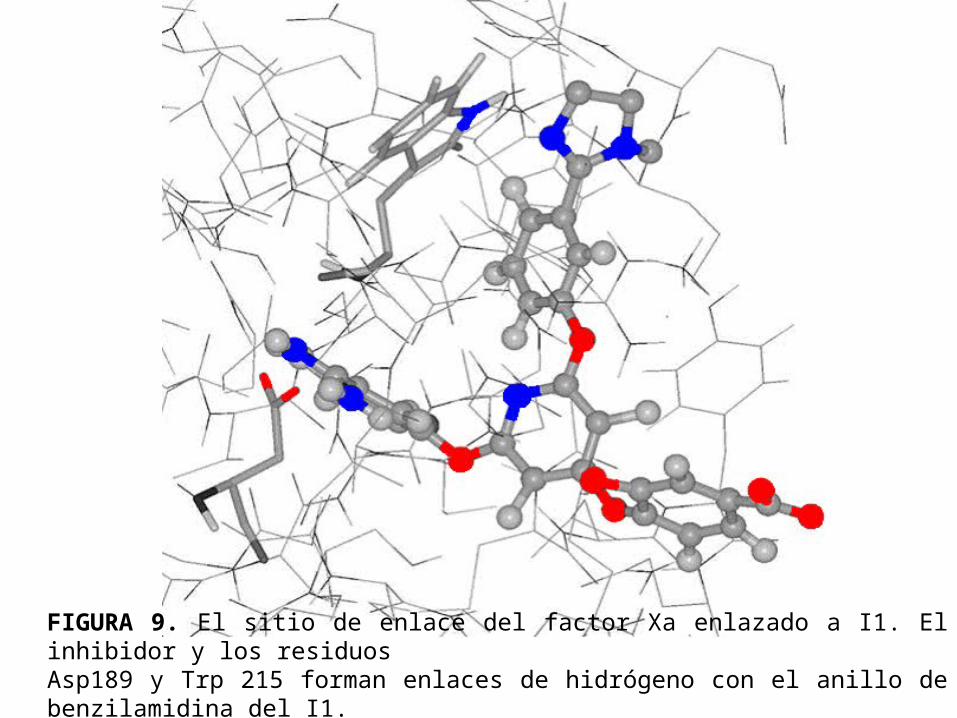
FIGURA 9. El sitio de enlace del factor Xa enlazado a I1. El inhibidor y los residuos Asp189 y Trp 215 forman enlaces de hidrógeno con el anillo de benzilamidina del I1.
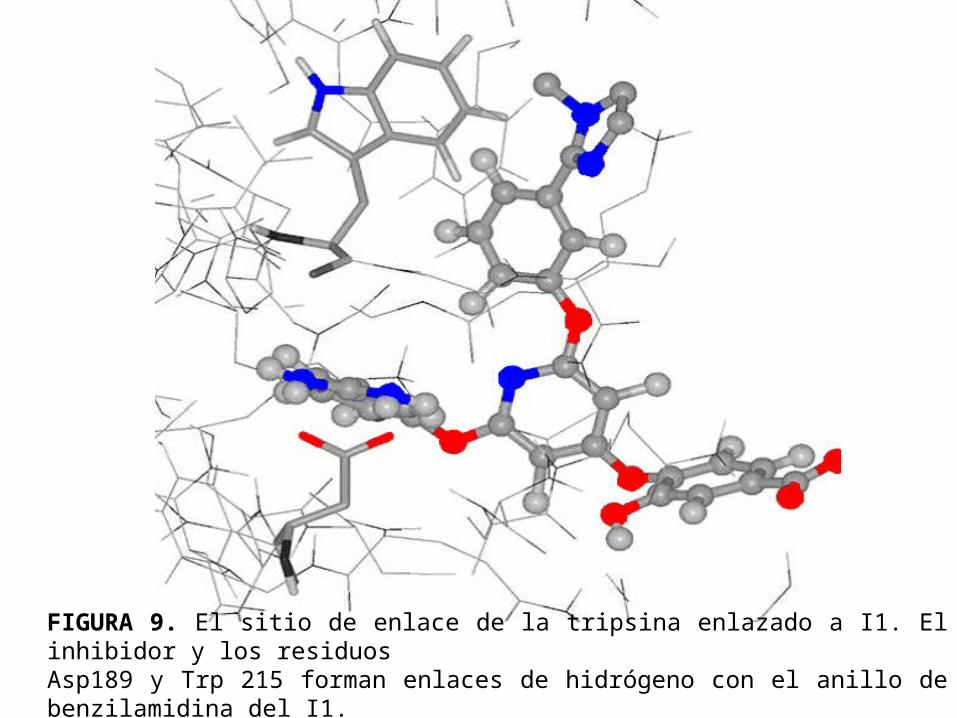
FIGURA 9. El sitio de enlace de la tripsina enlazado a I1. El inhibidor y los residuos Asp189 y Trp 215 forman enlaces de hidrógeno con el anillo de benzilamidina del I1.

• El anillo de bencilamidina se ubica en la cavidad S1 haciendo puentes hidrógeno con el Asp189.
• La distancia promedio entre el grupo amidínico del inhibidor y los oxígenos carbonílicos de este residuo es 2.1Å para el factor Xa y la tripsina.
• El grupo hidroxilo en la posición para del anillo de benzilamidina forma un enlace de hidrógeno con la Ser195; las distancias de enlace promedios en el factor Xa y la tripsina son 2.3Å y 2.1Å respectivamente.
• Las interacciones del sitio de enlace en S1 en las simulaciones son consistentes con los resultados obtenidos del estudio de la cristalografía de rayos X de la tripsina y del factor Xa acomplejado con la bisfenoxipiridina.

• El anillo fenílico del inhibidor I1, enlazado al sustituyente 1-methil-2-(2H)-imidazol se encuentra en la vecindad del Trp215 en la cavidad S4.
• La distancia promedio entre los centro de masas de estos dos grupos es 6.0Å en el factor Xa y 5.0Å en la tripsina.
• En la cristalografía de rayos X del factor Xa acomplejado con la bisfenoxipiridina, el grupo fenóxido se ubica cómodamente en la cavidad S4 cerca del anillo indólico del Trp215 ( 4.0Å ).

MUTACIONES EN EL FACTOR Xa Y LA TRIPSINA
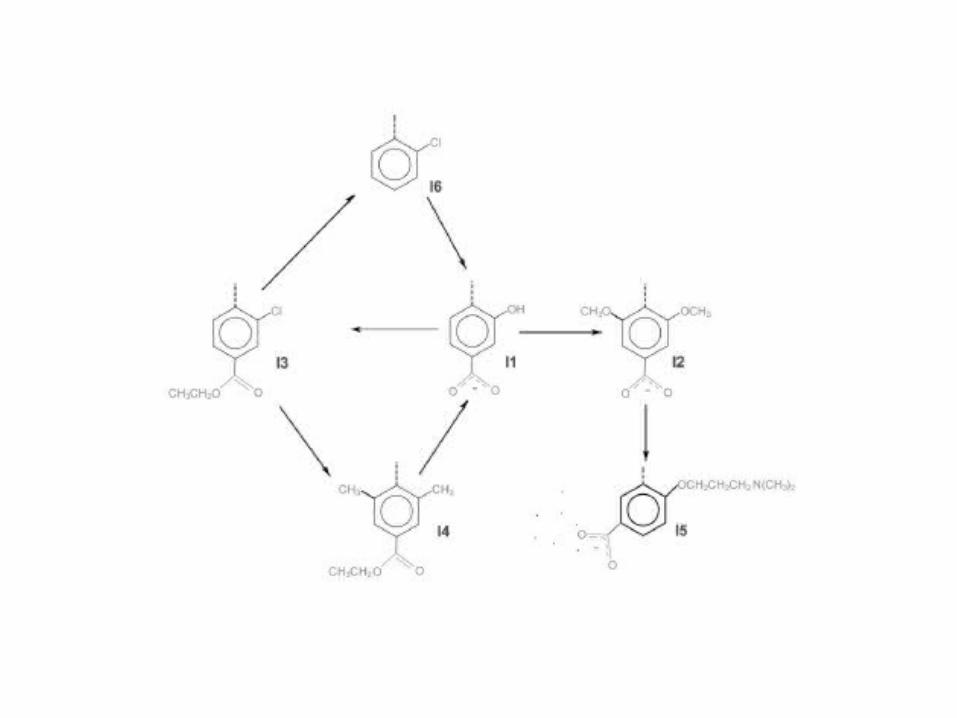
• Comenzando con la estructura equilibrada del complejo proteína-I1, todas las mutaciones mostrada en la figura 2 y 3 se realizaron, excepto para la transformación I2→I5.
• Para esta mutación, los resultados en medio acuoso ( Tabla 4 ) habían indicado que probando un tiempo mínimo a cada valor de del orden de nanosegundos era requerido.
• Como las pruebas del complejo demoran mucho más tiempo para lograr la convergencia del sistema proteína-inhibidor, el sistema demanda mucha capacidad computacional, por lo que esta mutación no fue estudiada en el factor Xa o tripsina.

• Tabla 4. Las energías libres ( KJ/mol ) para las mutaciones I2 I5 y I2 I5* en agua obtenidas usando diferentes muestreos de tiempos.

• Como se mostró antes la rotación del anillo aromático en agua, en general, se prueba adecuadamente dentro de 150ps.
• Sin embargo, dentro de la proteína, las pruebas se restringen ya que los anillos aromáticos no pueden adoptar todas las orientaciones posibles espontáneamente.
• Por consiguiente, para casos en que la rotación alrededor de los anillos aromáticos rinde diferentes estados, los cálculos fueron realizados en ambas orientaciones.
• Los resultados de las mutaciones en el factor Xa y en la tripsina son mostrados en las figuras 11 y 12 respectivamente.
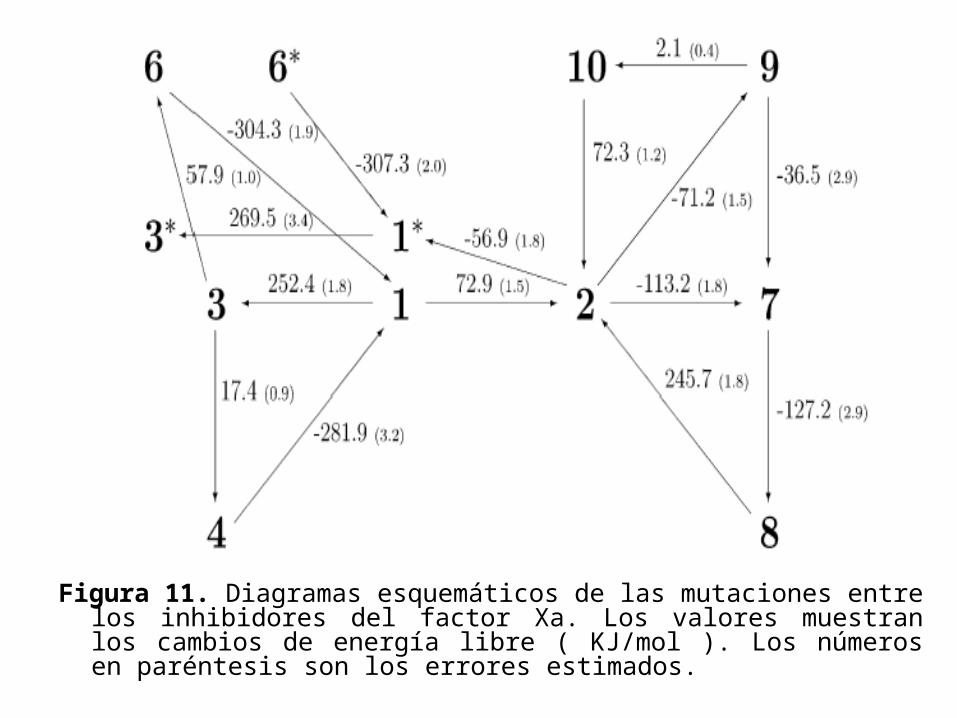
Figura 11. Diagramas esquemáticos de las mutaciones entre los inhibidores del factor Xa. Los valores muestran los cambios de energía libre ( KJ/mol ). Los números en paréntesis son los errores estimados.
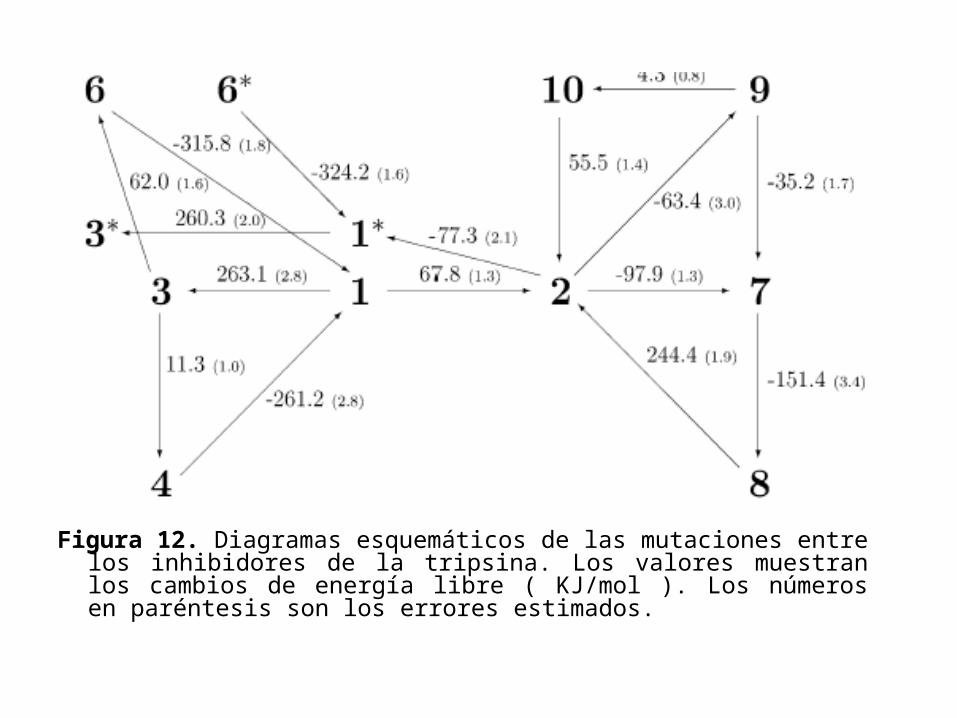
Figura 12. Diagramas esquemáticos de las mutaciones entre los inhibidores de la tripsina. Los valores muestran los cambios de energía libre ( KJ/mol ). Los números en paréntesis son los errores estimados.

• La diferencia energética de la energía libre en la misma mutación, pero con distinta conformación del inhibidor es de 2.8 a 17.1KJ/mol.
• Así, 250ps para cada punto no es suficiente para probar una distribución de equilibrio de las orientaciones de los anillos bencénicos dentro de la cavidad de enlace.
• En efecto es probable que debido al impedimento estérico cada rotación no es posible mientras el inhibidor este dentro de la cavidad de enlace; y si el tratamiento de las conformaciones es independiente, se requerirá considerar independiente al muestreo de tiempo.
• Así, ambas orientaciones se considerarán separadamente, y la orientación preferida se considerará que será la de menor energía libre.
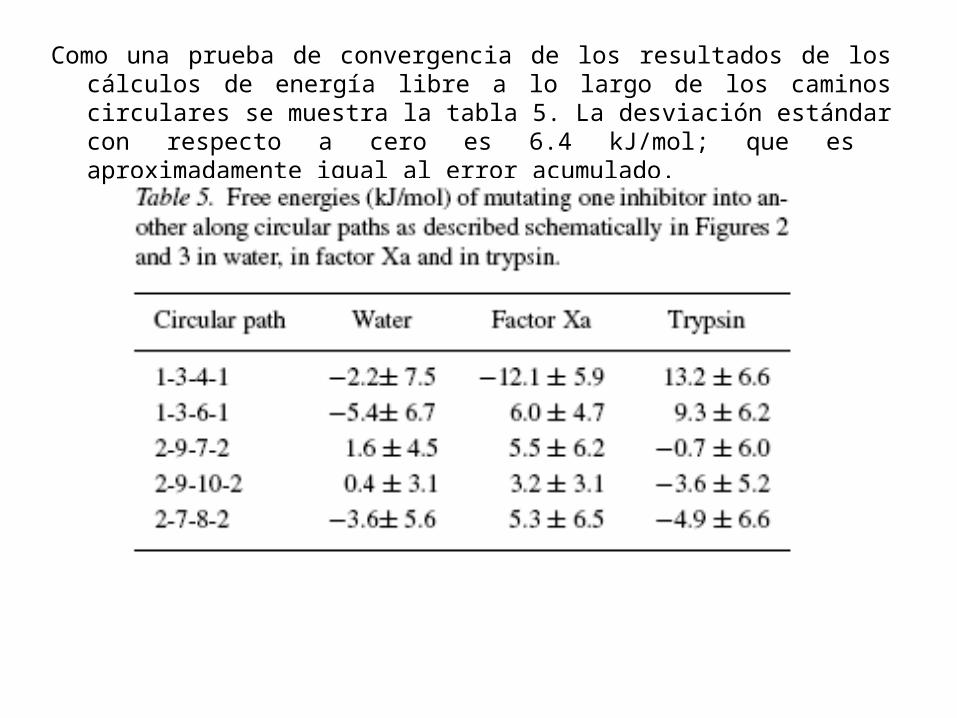
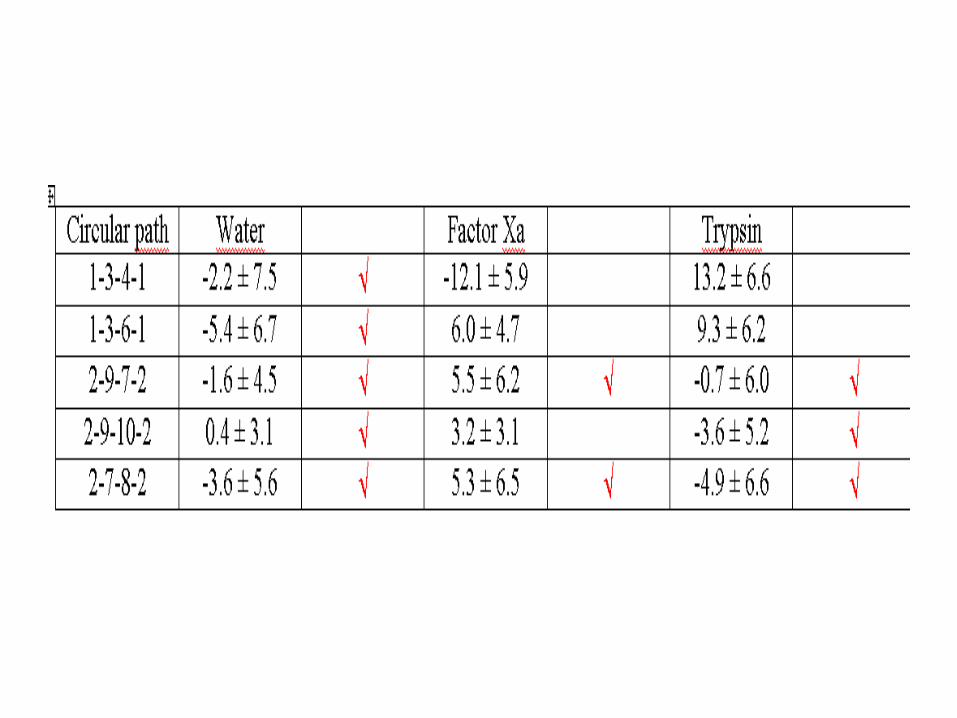
Como una prueba de convergencia de los resultados de los cálculos de energía libre a lo largo de los caminos circulares se muestra la tabla 5. La desviación estándar con respecto a cero es 6.4 kJ/mol; que es aproximadamente igual al error acumulado.

Recordando :
• Ecuaciones de Gibbs

• Las energías libres de enlace son obtenidas restando la energía del inhibidor en agua de la correspondiente energía libre del inhibidor en la proteína. La energía libre de enlace relativa de dos inhibidores a través de 2 proteínas fue obtenida restando la energía libre del inhibidor en la proteína de la correspondiente energía libre en otra proteína.
• = _ = _• Es conveniente tener un simple estado de referencia
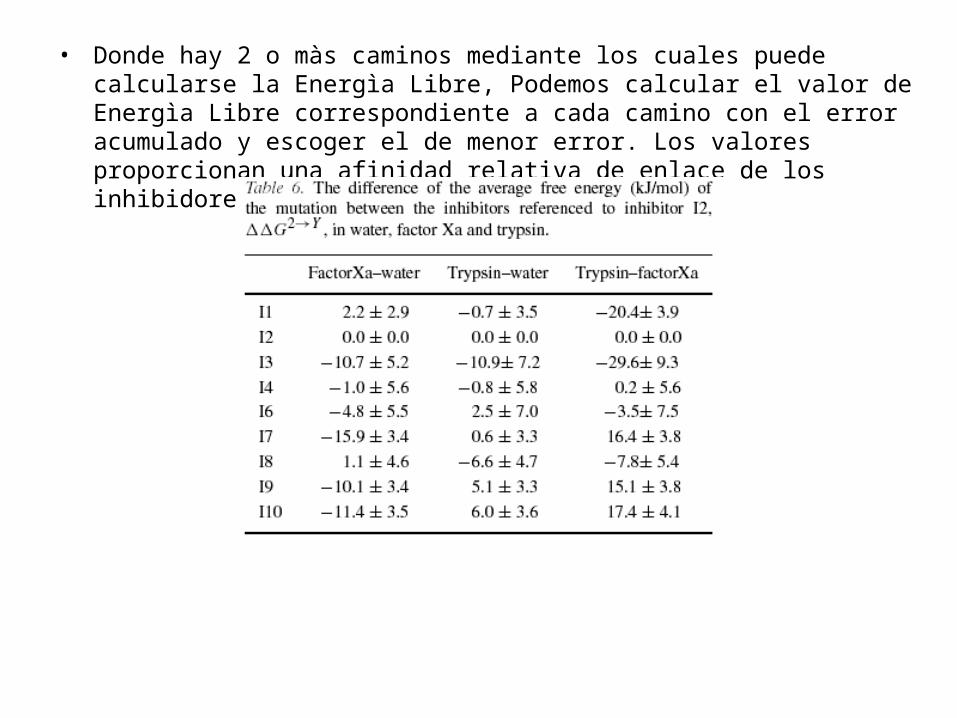
para los inhibidores, por eso en la tabla 6 las Energìas Libres relativas de Enlace son dadas con respecto a I2
• • Donde hay 2 o màs caminos mediante los cuales
puede calcularse la Energìa Libre, Podemos calcular el valor de Energìa Libre correspondiente a cada camino con el error acumulado y escoger el de menor error. Los valores proporcionan una afinidad relativa de enlace de los inhibidores.
• Donde hay 2 o màs caminos mediante los cuales puede calcularse la Energìa Libre, Podemos calcular el valor de Energìa Libre correspondiente a cada camino con el error acumulado y escoger el de menor error. Los valores proporcionan una afinidad relativa de enlace de los inhibidores.
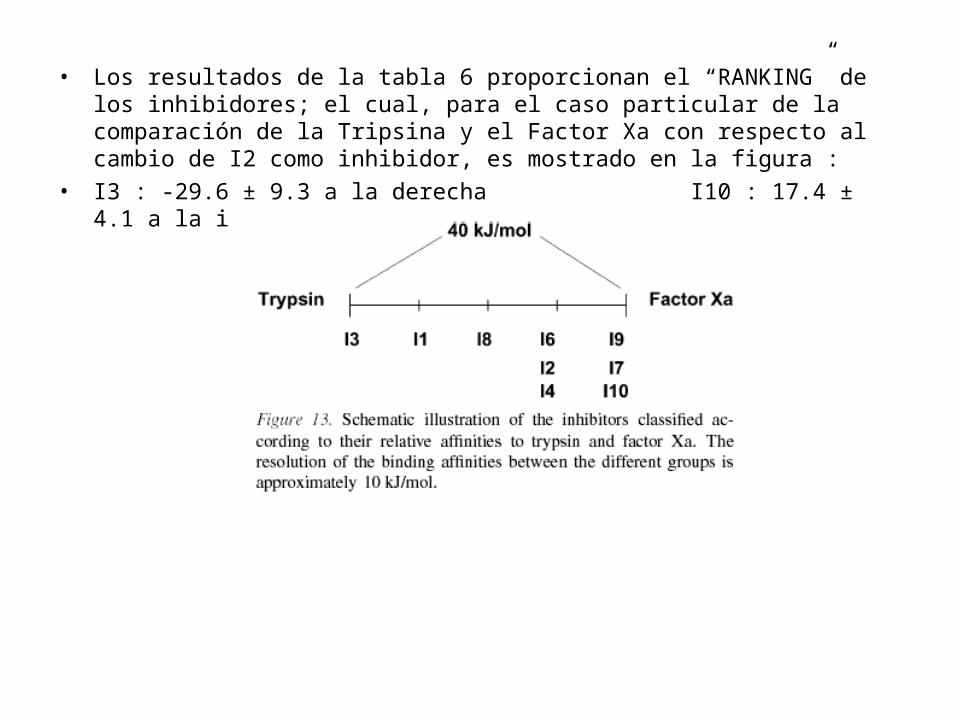
• Los resultados de la tabla 6 proporcionan el “RANKING” de los inhibidores; el cual, para el caso particular de la comparación de la Tripsina y el Factor Xa con respecto al cambio de I2 como inhibidor, es mostrado en la figura :
• I3 : -29.6 ± 9.3 a la derecha I10 : 17.4 ± 4.1 a la izquierda

• La distinción entre los inhibidores es sòlo hecha si la diferencia de las Energias Libres de Enlace es mucho màs grande que el error asociado.
• error• El error promedio, estimado del cierre del ciclo
termodinámico para la tripsina y el factor Xa es 5.4 kJ/mol. Por consiguiente dividamos a los inhibidores en dos grupos que difieren en aproximadamente 10.0 kJ/mol. Se observa que usar el inhibidor I3 es màs apropiado para la Tripsina y usar el inhibidor I10 es màs apropiado para el factorXa.

• El error promedio, estimado del cierre del ciclo termodinámico para la tripsina y el factor Xa es 5.4 kJ/mol. Por consiguiente dividamos a los inhibidores en dos grupos que difieren en aproximadamente 10.0 kJ/mol. Se observa que usar el inhibidor I3 es màs apropiado para la Tripsina y usar el inhibidor I10 es màs apropiado para el factorXa.

CONCLUSIONES• En este trabajo, no se contó con ningún dato
experimental de las constantes de enlace de cualquiera de los 10 inhibidores. Tal información serìa la prueba contundente de nuestros calculos de Energia Libre.
• La exactitud de losa cálculos de Energìa Libre dependen de 2 factores :
• Exactitud de los ForceField.• El grado de muestreo y la convergencia.• Para los casos en que las mutaciones eran simples
( cambiaban pocos átomos y no había cambio de carga ) las muestras en agua mostraban una razonable convergencia que fue obtenida durante 150ps para cada valor de de todas las transformaciones estudiadas, salvo I2 I5. Las ligeras mejoras al aumentar la duraciòn del tiempo tiempo.