comparative analysis of light-harvesting antennae and ... · comparative analysis of...
TRANSCRIPT

Comparative Analysis of Light-Harvesting Antennae andState Transition in chlorina and cpSRP Mutants1[OPEN]
Peng Wang and Bernhard Grimm*
Department of Plant Physiology, Humboldt University Berlin, 10115 Berlin, Germany
ORCID ID: 0000-0002-9730-1074 (B.G.).
State transitions in photosynthesis provide for the dynamic allocation of a mobile fraction of light-harvesting complex II (LHCII)to photosystem II (PSII) in state I and to photosystem I (PSI) in state II. In the state I-to-state II transition, LHCII isphosphorylated by STN7 and associates with PSI to favor absorption cross-section of PSI. Here, we used Arabidopsis(Arabidopsis thaliana) mutants with defects in chlorophyll (Chl) b biosynthesis or in the chloroplast signal recognition particle(cpSRP) machinery to study the flexible formation of PS-LHC supercomplexes. Intriguingly, we found that impaired Chl bbiosynthesis in chlorina1-2 (ch1-2) led to preferentially stabilized LHCI rather than LHCII, while the contents of both LHCI andLHCII were equally depressed in the cpSRP43-deficient mutant (chaos). In view of recent findings on the modified state transitions inLHCI-deficient mutants (Benson et al., 2015), the ch1-2 and chaos mutants were used to assess the influence of varying LHCI/LHCIIantenna size on state transitions. Under state II conditions, LHCII-PSI supercomplexes were not formed in both ch1-2 and chaosplants. LHCII phosphorylation was drastically reduced in ch1-2, and the inactivation of STN7 correlates with the lack of statetransitions. In contrast, phosphorylated LHCII in chaos was observed to be exclusively associated with PSII complexes, indicatinga lack of mobile LHCII in chaos. Thus, the comparative analysis of ch1-2 and chaos mutants provides new evidence for the flexibleorganization of LHCs and enhances our understanding of the reversible allocation of LHCII to the two photosystems.
In oxygenic photosynthesis, PSII and PSI function inseries to convert light energy into the chemical energythat fuels multiple metabolic processes. Most of thislight energy is captured by the chlorophyll (Chl) andcarotenoid pigments in the light-harvesting antennacomplexes (LHCs) that are peripherally associated withthe core complexes of both photosystems (Wobbe et al.,2016). However, since the two photosystems exhibitdifferent absorption spectra (Nelson and Yocum, 2006;Nield and Barber, 2006; Qin et al., 2015), PSI or PSII ispreferentially excited under naturally fluctuating lightintensities and qualities. To optimize photosyntheticelectron transfer, the excitation state of the two photo-systems must be rebalanced in response to changes inlighting conditions. To achieve this, higher plants andgreen algae require rapid and precise acclimatorymechanisms to adjust the relative absorption cross-sections of the two photosystems.
To date, the phenomenon of state transitions is one ofthewell-documented short-termacclimatorymechanisms.
It allows a mobile portion of the light-harvesting antennacomplex II (LHCII) to be allocated to either photosystem,dependingon the spectral composition and intensity of theambient light (Allen and Forsberg, 2001; Rochaix, 2011;Goldschmidt-Clermont and Bassi, 2015; Gollan et al.,2015). State transitions are driven by the redox state of theplastoquinone (PQ) pool (Vener et al., 1997; Zito et al.,1999). When PSI is preferentially excited (by far-red light),the PQ pool is oxidized and all the LHCII is associatedwith PSII. This allocation of antenna complexes is definedas state I. When light conditions (blue/red light or lowlight) favor exciton trapping of PSII, the transition fromstate I to state II occurs. The over-reduced PQpool triggersthe activation of the membrane-localized Ser-Thr kinaseSTN7,which phosphorylates anN-terminal Thr on each oftwo major LHCII proteins, LHCB1 and LHCB2 (Allen,1992; Bellafiore et al., 2005; Shapiguzov et al., 2016).Phosphorylation of LHCII results in the dissociation ofLHCII from PSII and triggers its reversible relocation toPSI (Allen, 1992; Rochaix, 2011). Conversely, when the PQpool is reoxidized, STN7 is inactivated and the constitu-tively active, thylakoid-associated phosphatase TAP38/PPH1 dephosphorylates LHCII, which then reassociateswith PSII (Pribil et al., 2010; Shapiguzov et al., 2010). Thephysiological significance of state transitions has beendemonstrated by the reduction in growth rate seen in thestn7 knock-out mutant under fluctuating light conditions(Bellafiore et al., 2005; Tikkanen et al., 2010).
The canonical state transitions model implies spatialand temporal regulation of the allocation of LHCbetweenthe two spatially segregated photosystems (Dekker andBoekema, 2005). PSII-LHCII supercomplexes are orga-nized in a tightly packed form in the stacked grana
1 This workwas supported by the Alexander vonHumboldt Foun-dation (P.W.) and by the Deutsche Forschungsgemeinschaft FOR2092(grant no. GR 936/18-1 to B.G.).
* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Bernhard Grimm ([email protected]).
P.W. and B.G. designed the research; P.W. performed the experi-ments; P.W. and B.G. analyzed the data and wrote the article.
[OPEN] Articles can be viewed without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.16.01009
Plant Physiology�, November 2016, Vol. 172, pp. 1519–1531, www.plantphysiol.org � 2016 American Society of Plant Biologists. All Rights Reserved. 1519 www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

regions of thylakoid membranes, while PSI-LHCI super-complexes are mainly localized in the nonstacked stro-mal lamellae and grana margin regions (Dekker andBoekema, 2005; Haferkamp et al., 2010). It has been pro-posed that, in the grana margin regions, which harborLHCII and both photosystems, LHCII can migrate rap-idly between them (Albertsson et al., 1990; Albertsson,2001). This idea is supported by the recent discovery ofmega complexes containing both photosystems in thegrana margin regions (Yokono et al., 2015). Furthermore,phosphorylation of LHCII was found to increase not onlythe amount of PSI found in the grana margin region ofthylakoid membranes (Tikkanen et al., 2008a), but also tomodulate the pattern of PSI-PSII megacomplexes underchanging light conditions (Suorsa et al., 2015). Nonethe-less, open questions remain in relation to the physiolog-ical significance of the detection of phosphorylated LHCIIin all thylakoid regions, even under the constant lightconditions (Grieco et al., 2012; Leoni et al., 2013; Wientjeset al., 2013), although LHCII phosphorylation has beenshown to modify the stacking of thylakoid membranes(Chuartzman et al., 2008; Pietrzykowska et al., 2014).
State I-to-state II transition is featured by the forma-tion of LHCII-PSI-LHCI supercomplexes, in whichLHCII favors the light-harvesting capacity of PSI. Re-cently, LHCII-PSI-LHCI supercomplexes have beensuccessfully isolated and purified using various deter-gents (Galka et al., 2012; Drop et al., 2014; Crepin andCaffarri, 2015) or a styrene-maleic acid copolymer (Bellet al., 2015). These findings yielded further insights intothe reorganization of supercomplexes associated withstate transitions, and it was suggested that phospho-rylation of LHCB2 rather than LHCB1 is the essentialtrigger for the formation of state transition super-complexes (Leoni et al., 2013; Pietrzykowska et al.,2014; Crepin and Caffarri, 2015; Longoni et al., 2015).Furthermore, characterization of mutants deficient inindividual PSI core subunits indicates that PsaH, L, andI are required for docking of LHCII at PSI (Lunde et al.,2000; Zhang and Scheller, 2004; Kouril et al., 2005;Plöchinger et al., 2016).
Recently, the state transition capacity has beencharacterized in the Arabidopsis (Arabidopsis thaliana)mutants with missing LHCI components. Although theArabidopsis knock-out mutants lacking one of the fourLHCI proteins (LHCA1-4) showed enhanced accumu-lation of LHCII-PSI complexes, the absorption cross-section of PSI under state II conditions was stillcompromised in the lhca1-4mutants, and it is suggestedthat LHCI mediates the detergent-sensitive interactionbetween ‘extra LHCII’ and PSI (Benson et al., 2015;Grieco et al., 2015). Furthermore, the Arabidopsis mu-tant DLhca lacking all LHCA1-4 proteins was shown tobe compensated for the deficiency of LHCI by bindingLHCII under state II conditions (Bressan et al., 2016). Inspite of this finding, the significant reduction in theabsorption cross-section of PSI was still observed in theDLhca mutant, suggesting a substantial role of LHCIin light absorption under canopy conditions (Bressanet al., 2016). However, these findings emphasize the
acclimatory function of state transitions in balancinglight absorption capacity between the two photosys-tems bymodifying their relative antenna size and implythe dynamic and variable organization of PS-LHCsupercomplexes.
LHC proteins are encoded by the nuclear Lhc super-family (Jansson, 1994). The biogenesis of LHCs includesthe cytoplasmic synthesis of the LHC precursor pro-teins, their translocation into chloroplasts via the TOC/TIC complex, and their posttranslational targeting andintegration into the thylakoid membranes by meansof the chloroplast signal recognition particle (cpSRP)machinery (Jarvis and Lopez-Juez, 2013). The post-translational cpSRP-dependent pathway for the finaltranslocation of LHC proteins into the thylakoidmembrane includes interaction of cpSRP43 with LHCapo-proteins and recruitment of cpSRP54 to form atransit complex. Then binding of this tripartite cpSRPtransit complex to the SRP receptor cpFtsY follows,which supports docking of the transit complex to thy-lakoid membranes and its association with the LHCtranslocase ALB3. Ultimately, ALB3 inserts LHC apo-proteins into the thylakoid membrane (Richter et al.,2010). Importantly, stoichiometric amounts of newlysynthesized Chl a and Chl b as well as carotenoid areinserted into the LHC apo-proteins by unknownmechanisms to form the functional LHCs that associatewith the core complexes of both photosystems in thethylakoid membranes (Dall’Osto et al., 2015; Wang andGrimm, 2015).
The first committed steps in Chl synthesis occur inthe Mg branch of the tetrapyrrole biosynthesis path-way. 5-Aminolevulinic acid synthesis provides theprecursor for the formation of protoporphyrin IX,which is directed into the Mg branch (Tanaka andTanaka, 2007; Brzezowski et al., 2015). Chl synthesisends with the conversion of Chl a to Chl b catalyzed byChl a oxygenase (CAO; Tanaka et al., 1998; Tomitaniet al., 1999). It has been hypothesized that coordinationbetween Chl synthesis and the posttranslational cpSRPpathway is a prerequisite for the efficient integration ofChls into LHC apo-proteins.
In this study, we intend to characterize the assemblyof LHCs when the availability of Chl molecules or theintegration of LHC apo-proteins into thylakoid mem-branes is limiting. To this end, we compared the as-sembly of LHCs and the organization of PS-LHCcomplexes in two different sets of Arabidopsis mutants.Firstly, we used the chlorina1-2 (ch1-2) mutant, which isdefective in the CAO gene. The members of the secondset of mutants carry knock-out mutations in genes in-volved in the chloroplast SRP pathway (Richter et al.,2010).
Our studies revealed distinct accumulation ofPS-LHC supercomplexes between the two sets of mu-tant relative to wild-type plants. In spite of the defect insynthesis of Chl b, ch1-2 retains predominantly intactPSI-LHCI supercomplexes but has strongly reducedamounts of LHCII. In contrast, the chaos (cpSRP43)mutant exhibits synchronously reduced contents of
1520 Plant Physiol. Vol. 172, 2016
Wang and Grimm
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
both LHCI and LHCII, which results in the accumula-tion of PS core complexes without accompanyingLHCs. Thus, the distribution of LHCs in the thylakoidmembranes of the two mutants, ch1-2 and chaos, wereexplored under varying light conditions with the aim ofelucidating the influence of modified LHCI/LHCIIantenna size on state transitions. Our results contributeto an expanding view on the variety of photosyntheticcomplexes, which can be observed in Arabidopsisplants with specified mutations in LHC biogenesis.
RESULTS
Reduced Contents of LHCs in ch1-2 and cpsrp Mutants
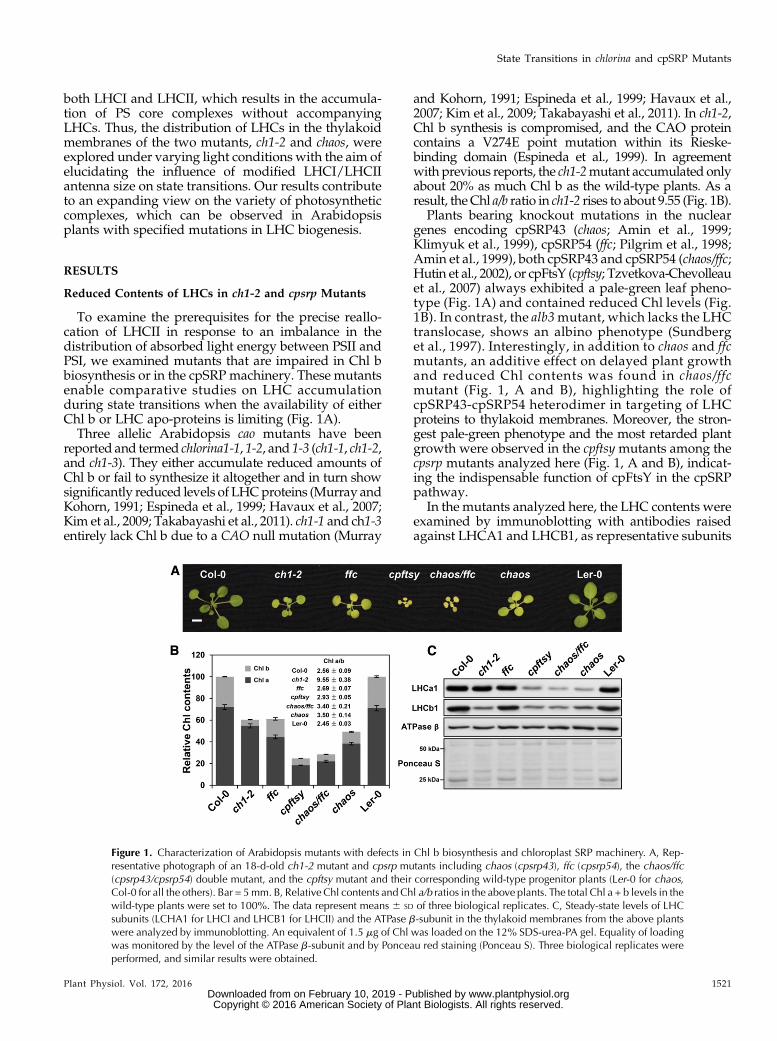
To examine the prerequisites for the precise reallo-cation of LHCII in response to an imbalance in thedistribution of absorbed light energy between PSII andPSI, we examined mutants that are impaired in Chl bbiosynthesis or in the cpSRP machinery. These mutantsenable comparative studies on LHC accumulationduring state transitions when the availability of eitherChl b or LHC apo-proteins is limiting (Fig. 1A).Three allelic Arabidopsis cao mutants have been
reported and termed chlorina1-1, 1-2, and 1-3 (ch1-1, ch1-2,and ch1-3). They either accumulate reduced amounts ofChl b or fail to synthesize it altogether and in turn showsignificantly reduced levels of LHCproteins (Murray andKohorn, 1991; Espineda et al., 1999; Havaux et al., 2007;Kim et al., 2009; Takabayashi et al., 2011). ch1-1 and ch1-3entirely lack Chl b due to a CAO null mutation (Murray
and Kohorn, 1991; Espineda et al., 1999; Havaux et al.,2007; Kim et al., 2009; Takabayashi et al., 2011). In ch1-2,Chl b synthesis is compromised, and the CAO proteincontains a V274E point mutation within its Rieske-binding domain (Espineda et al., 1999). In agreementwith previous reports, the ch1-2mutant accumulated onlyabout 20% as much Chl b as the wild-type plants. As aresult, the Chl a/b ratio in ch1-2 rises to about 9.55 (Fig. 1B).
Plants bearing knockout mutations in the nucleargenes encoding cpSRP43 (chaos; Amin et al., 1999;Klimyuk et al., 1999), cpSRP54 (ffc; Pilgrim et al., 1998;Amin et al., 1999), both cpSRP43 and cpSRP54 (chaos/ffc;Hutin et al., 2002), or cpFtsY (cpftsy; Tzvetkova-Chevolleauet al., 2007) always exhibited a pale-green leaf pheno-type (Fig. 1A) and contained reduced Chl levels (Fig.1B). In contrast, the alb3mutant, which lacks the LHCtranslocase, shows an albino phenotype (Sundberget al., 1997). Interestingly, in addition to chaos and ffcmutants, an additive effect on delayed plant growthand reduced Chl contents was found in chaos/ffcmutant (Fig. 1, A and B), highlighting the role ofcpSRP43-cpSRP54 heterodimer in targeting of LHCproteins to thylakoid membranes. Moreover, the stron-gest pale-green phenotype and the most retarded plantgrowth were observed in the cpftsymutants among thecpsrp mutants analyzed here (Fig. 1, A and B), indicat-ing the indispensable function of cpFtsY in the cpSRPpathway.
In the mutants analyzed here, the LHC contents wereexamined by immunoblotting with antibodies raisedagainst LHCA1 and LHCB1, as representative subunits
Figure 1. Characterization of Arabidopsis mutants with defects in Chl b biosynthesis and chloroplast SRP machinery. A, Rep-resentative photograph of an 18-d-old ch1-2 mutant and cpsrp mutants including chaos (cpsrp43), ffc (cpsrp54), the chaos/ffc(cpsrp43/cpsrp54) double mutant, and the cpftsy mutant and their corresponding wild-type progenitor plants (Ler-0 for chaos,Col-0 for all the others). Bar = 5mm. B, Relative Chl contents and Chl a/b ratios in the above plants. The total Chl a + b levels in thewild-type plants were set to 100%. The data represent means 6 SD of three biological replicates. C, Steady-state levels of LHCsubunits (LCHA1 for LHCI and LHCB1 for LHCII) and the ATPase b-subunit in the thylakoid membranes from the above plantswere analyzed by immunoblotting. An equivalent of 1.5 mg of Chl was loaded on the 12% SDS-urea-PA gel. Equality of loadingwas monitored by the level of the ATPase b-subunit and by Ponceau red staining (Ponceau S). Three biological replicates wereperformed, and similar results were obtained.
Plant Physiol. Vol. 172, 2016 1521
State Transitions in chlorina and cpSRP Mutants
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

of LHCI and LHCII, respectively. As shown before(Espineda et al., 1999), LHCB1 was strongly reduced inch1-2 (Fig. 1C), while the LHCA1 content was unex-pectedly slightly diminished (Fig. 1C). In contrast toch1-2 mutant, the cpsrp mutants contained severely re-duced contents of the LHCPs of both photosystems.Combining previous detailed descriptions of the effectsof cpsrpmutations on levels of various LHCI and LHCIIsubunits (Pilgrim et al., 1998; Espineda et al., 1999;Hutin et al., 2002; Tzvetkova-Chevolleau et al., 2007;Ouyang et al., 2011), we concluded that, in each of thethree cpsrpmutants studied here, steady-state amountsof LHCI and LHCII proteins are equally affected (Fig.1C). Interestingly, levels of LHC proteins were clearlyhigher in ffc than in chaos, chaos/ffc, and cpftsy (Fig. 1C),indicating that cpSRP43 functions predominantly andindependently from cpSRP54 in targeting of LHC pro-teins to the thylakoidmembranes (Tzvetkova-Chevolleauet al., 2007; Liang et al., 2016). In summary, our initialresults suggest that malfunction of the cpSRP pathwaydepresses steady-state levels of both LHCI and LHCII,while strongly reduced Chl b biosynthesis preferentiallyaffects LHCII.
Accumulation of Photosynthetic Apparatus in ch1-2 andcpsrp Mutants
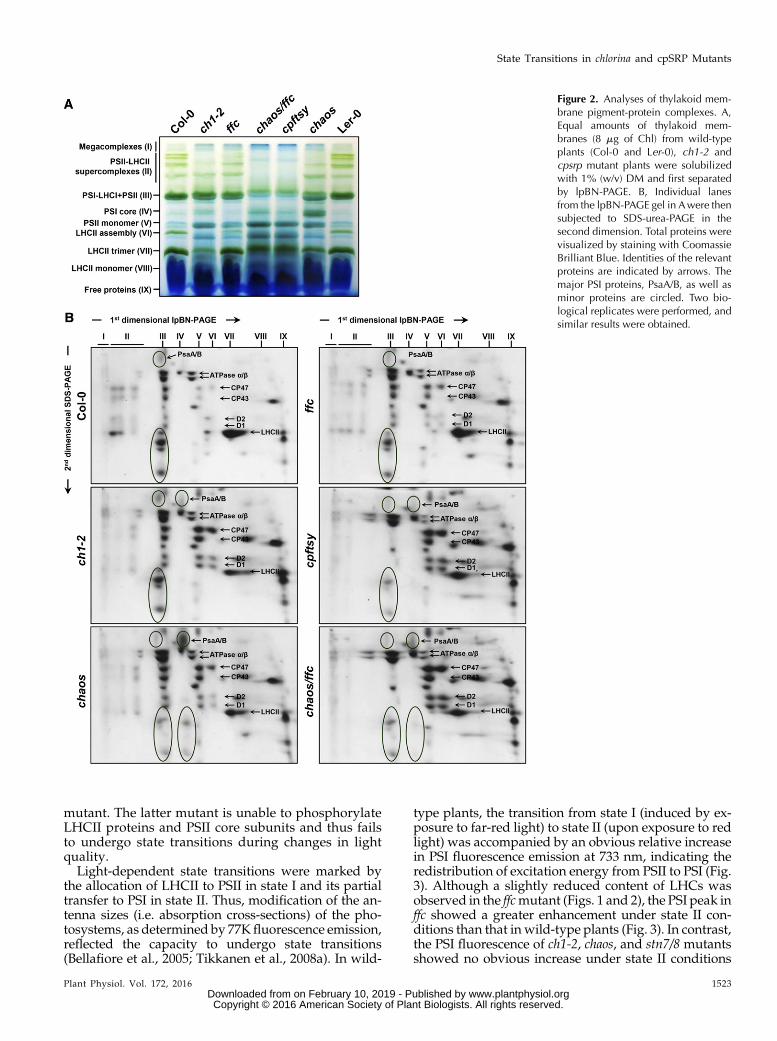
The diminished LHC contents observed in ch1-2 andcpsrp mutants enabled us to examine the consequencesof each mutation for the assembly of PS-LHC com-plexes in the thylakoid membranes. For this pur-pose, the isolated thylakoid membranes were treatedwith the nonionic detergent n-dodecyl-b-D-maltoside(b-DM) to efficiently solubilize both grana and non-stacked regions (Jarvi et al., 2011; Grieco et al., 2015).The thylakoid membranes were then fractionated bylarge-pore Blue-Native polyacrylamide gel electropho-resis (lpBN-PAGE) (Jarvi et al., 2011) followed by SDS-PAGE in the second dimension to determine the proteincomposition of each of the various photosyntheticcomplexes.
In the thylakoid membranes, LHCII is peripherallyassociated with PSII to form a PSII-LHCII super-complex, which is mainly localized in the grana coreregions (Dekker and Boekema, 2005). Depending on thebinding strength of LHCII to PSII, four variants of PSII-LHCII supercomplexes (II) were observed on thelpBN-PA gel (Fig. 2, A and B). Apart from the PSII-LHCII supercomplexes, several PSII subcomplexes, in-cluding the PSII dimer (III), PSII monomer (V), LHCIIassembly complex (VI), trimeric and monomeric LHCII(VII and VIII) could be detected (Fig. 2, A and B), whichis consistent with previous reports (Jarvi et al., 2011).The ch1-2, ffc, and chaos mutants were characterized byreduced amounts of the PSII-LHCII supercomplexesand LHCII trimers, which are in turn associated withelevated levels of the PSII monomer and LHCII as-sembly complex (Fig. 2, A and B). The chaos/ffc andcpftsymutants showed a more severe reduction in PSII-LHCII supercomplexes and PSII dimers (Fig. 2, A and
B), suggesting that simultaneous loss of cpSRP43 andcpSRP54 or deficiency of the cpFtsY receptor affects notonly the stability of antenna proteins but also the as-sembly of the PSII core complex in the thylakoidmembranes. This observation is supported by the ear-lier finding that cpSRP54 and cpFtsY cooperate in thecotranslational integration of plastid-encoded PSII coresubunits (Richter et al., 2010).
In contrast to the various PSII-LHCII supercomplexesseen in wild-type plants, only a single PSI-LHCIsupercomplex (III) was observed in control plants,whichmigrates close to PSII dimers on lpBN-PA gels. Inmutants defective in LHCI formation, only PSI corecomplexes are observed (Havaux et al., 2007; Wientjeset al., 2009; Takabayashi et al., 2011; Benson et al., 2015).A dominant band of PSI core complexes was observedin the chaos mutant (Fig. 2, A and B), confirming re-duced accumulation of LHCI subunits in chaos relativeto ch1-2 (Fig. 1C). In addition to drastically disruptedassembly and/or reduced stability of PSII-LHCIIsupercomplexes and PSII dimers in chaos/ffc and cpftsymutants, accumulation of both PSI-LHCI and PSI corecomplexes was strongly impaired (Fig. 2, A and B). Incontrast to these observations, the slight reductions inLHCI proteins seen in ch1-2 and ffc are consistent with aminor perturbation of PSI-LHCI supercomplex forma-tion (Fig. 2, A and B).
In summary, based on the accumulation of PS-LHCcomplexes in the thylakoid membranes, ch1-2 and thedifferent cpsrp mutants can be classified into threegroups: (1) ch1-2 exhibited a drastically reduced contentof LHCII and only a slightly impaired LHCI content; (2)chaos and ffc were both characterized by impaired ac-cumulation of both LHCI and LHCII, with levels ofboth complexes being more severely affected in thechaos mutant than in ffc; and (3) the chaos/ffc and cpftsymutants showed the greatest reductions in LHC con-tent, and accumulated photosystem core complexes.
Impaired State Transitions in ch1-2 and cpsrp Mutants
Short-term state transitions enable the reversible al-location of LHCII to PSI when PSII rather than PSI ispreferentially activated (Allen and Forsberg, 2001;Rochaix, 2011; Goldschmidt-Clermont and Bassi, 2015;Gollan et al., 2015). The observations that ch1-2 andchaos mutants exhibited distinct accumulation of PSI-LHCI complexes (Figs. 1 and 2) led to further explora-tion of the association of LHCII with PSI or PSII duringstate transitions. It was recently shown that an intactLHCI complex is required for a complete state I-to-stateII transition (Benson et al., 2015). To explore thesefindings further, we compared state transitions in ch1-2and chaoswith control seedlings. We hypothesized thatthe defects in formation of PSI-LHCI supercomplexesobserved in chaos would lead to an aberrant transitionrelative to ch1-2 and control plants under state II con-ditions (Figs. 1 and 2). As additional controls, we ex-amined the ffc mutant, in which levels of both LHCswere only slightly reduced, and the stn7/8 double
1522 Plant Physiol. Vol. 172, 2016
Wang and Grimm
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
mutant. The latter mutant is unable to phosphorylateLHCII proteins and PSII core subunits and thus failsto undergo state transitions during changes in lightquality.Light-dependent state transitions were marked by
the allocation of LHCII to PSII in state I and its partialtransfer to PSI in state II. Thus, modification of the an-tenna sizes (i.e. absorption cross-sections) of the pho-tosystems, as determined by 77K fluorescence emission,reflected the capacity to undergo state transitions(Bellafiore et al., 2005; Tikkanen et al., 2008a). In wild-
type plants, the transition from state I (induced by ex-posure to far-red light) to state II (upon exposure to redlight) was accompanied by an obvious relative increasein PSI fluorescence emission at 733 nm, indicating theredistribution of excitation energy from PSII to PSI (Fig.3). Although a slightly reduced content of LHCs wasobserved in the ffcmutant (Figs. 1 and 2), the PSI peak inffc showed a greater enhancement under state II con-ditions than that in wild-type plants (Fig. 3). In contrast,the PSI fluorescence of ch1-2, chaos, and stn7/8 mutantsshowed no obvious increase under state II conditions
Figure 2. Analyses of thylakoid mem-brane pigment-protein complexes. A,Equal amounts of thylakoid mem-branes (8 mg of Chl) from wild-typeplants (Col-0 and Ler-0), ch1-2 andcpsrp mutant plants were solubilizedwith 1% (w/v) DM and first separatedby lpBN-PAGE. B, Individual lanesfrom the lpBN-PAGE gel in Awere thensubjected to SDS-urea-PAGE in thesecond dimension. Total proteins werevisualized by staining with CoomassieBrilliant Blue. Identities of the relevantproteins are indicated by arrows. Themajor PSI proteins, PsaA/B, as well asminor proteins are circled. Two bio-logical replicates were performed, andsimilar results were obtained.
Plant Physiol. Vol. 172, 2016 1523
State Transitions in chlorina and cpSRP Mutants
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
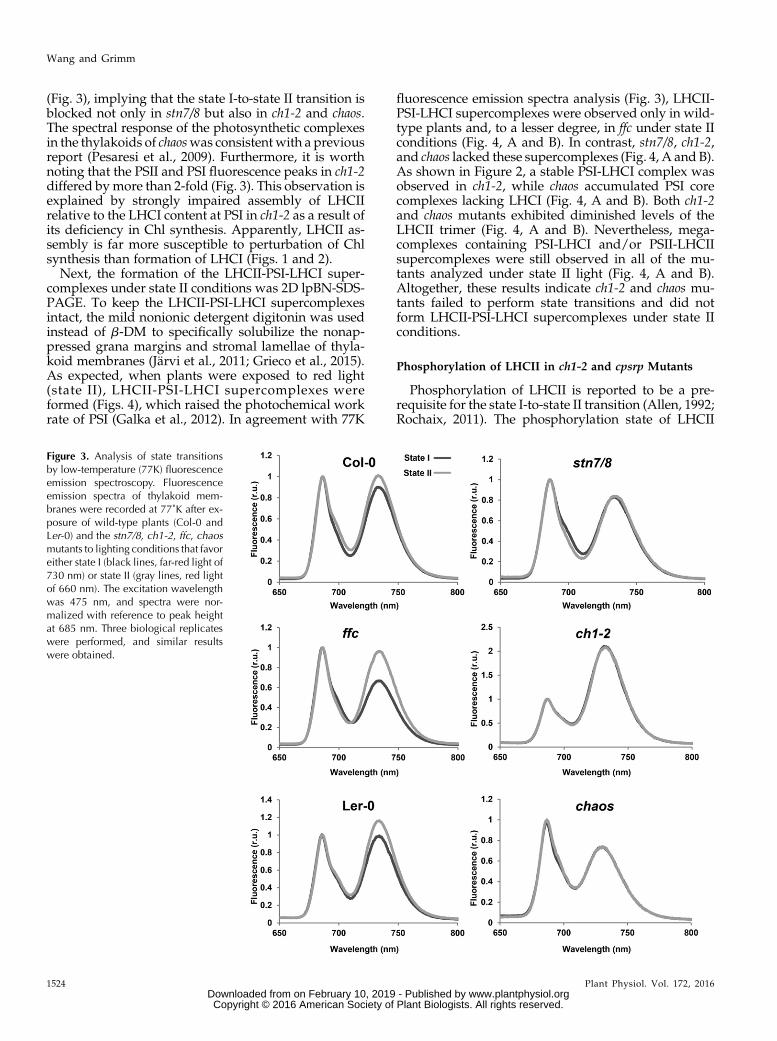
(Fig. 3), implying that the state I-to-state II transition isblocked not only in stn7/8 but also in ch1-2 and chaos.The spectral response of the photosynthetic complexesin the thylakoids of chaoswas consistent with a previousreport (Pesaresi et al., 2009). Furthermore, it is worthnoting that the PSII and PSI fluorescence peaks in ch1-2differed bymore than 2-fold (Fig. 3). This observation isexplained by strongly impaired assembly of LHCIIrelative to the LHCI content at PSI in ch1-2 as a result ofits deficiency in Chl synthesis. Apparently, LHCII as-sembly is far more susceptible to perturbation of Chlsynthesis than formation of LHCI (Figs. 1 and 2).
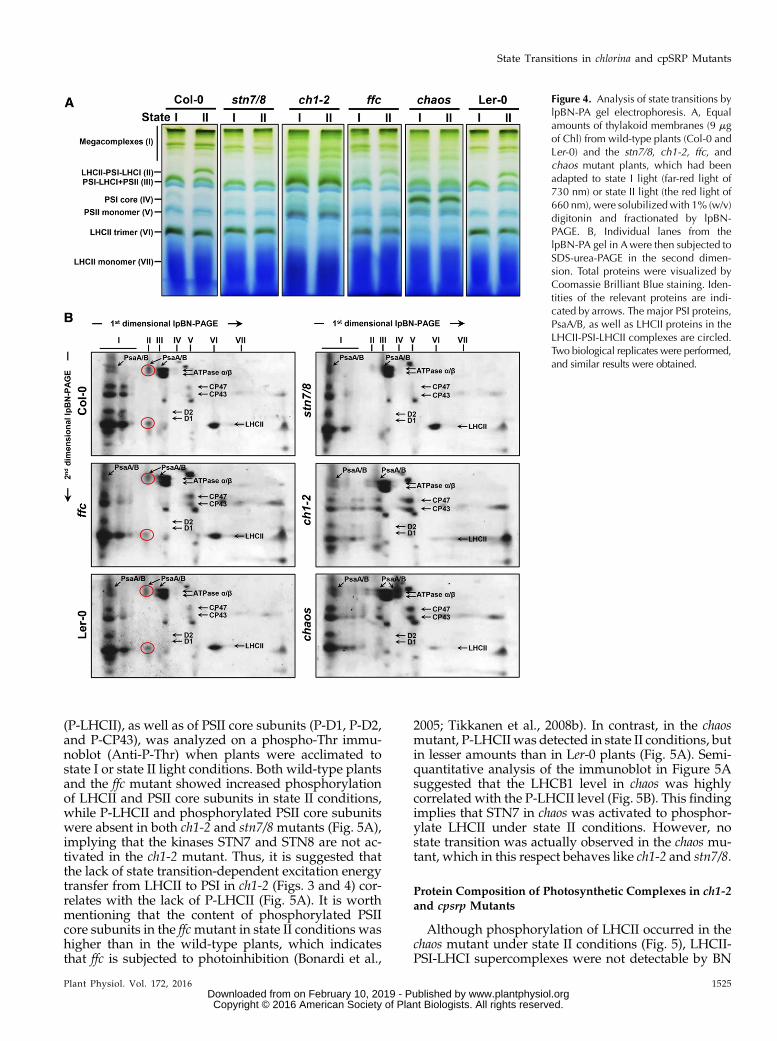
Next, the formation of the LHCII-PSI-LHCI super-complexes under state II conditions was 2D lpBN-SDS-PAGE. To keep the LHCII-PSI-LHCI supercomplexesintact, the mild nonionic detergent digitonin was usedinstead of b-DM to specifically solubilize the nonap-pressed grana margins and stromal lamellae of thyla-koid membranes (Järvi et al., 2011; Grieco et al., 2015).As expected, when plants were exposed to red light(state II), LHCII-PSI-LHCI supercomplexes wereformed (Figs. 4), which raised the photochemical workrate of PSI (Galka et al., 2012). In agreement with 77K
fluorescence emission spectra analysis (Fig. 3), LHCII-PSI-LHCI supercomplexes were observed only in wild-type plants and, to a lesser degree, in ffc under state IIconditions (Fig. 4, A and B). In contrast, stn7/8, ch1-2,and chaos lacked these supercomplexes (Fig. 4, A and B).As shown in Figure 2, a stable PSI-LHCI complex wasobserved in ch1-2, while chaos accumulated PSI corecomplexes lacking LHCI (Fig. 4, A and B). Both ch1-2and chaos mutants exhibited diminished levels of theLHCII trimer (Fig. 4, A and B). Nevertheless, mega-complexes containing PSI-LHCI and/or PSII-LHCIIsupercomplexes were still observed in all of the mu-tants analyzed under state II light (Fig. 4, A and B).Altogether, these results indicate ch1-2 and chaos mu-tants failed to perform state transitions and did notform LHCII-PSI-LHCI supercomplexes under state IIconditions.
Phosphorylation of LHCII in ch1-2 and cpsrp Mutants
Phosphorylation of LHCII is reported to be a pre-requisite for the state I-to-state II transition (Allen, 1992;Rochaix, 2011). The phosphorylation state of LHCII
Figure 3. Analysis of state transitionsby low-temperature (77K) fluorescenceemission spectroscopy. Fluorescenceemission spectra of thylakoid mem-branes were recorded at 77˚K after ex-posure of wild-type plants (Col-0 andLer-0) and the stn7/8, ch1-2, ffc, chaosmutants to lighting conditions that favoreither state I (black lines, far-red light of730 nm) or state II (gray lines, red lightof 660 nm). The excitation wavelengthwas 475 nm, and spectra were nor-malized with reference to peak heightat 685 nm. Three biological replicateswere performed, and similar resultswere obtained.
1524 Plant Physiol. Vol. 172, 2016
Wang and Grimm
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
(P-LHCII), as well as of PSII core subunits (P-D1, P-D2,and P-CP43), was analyzed on a phospho-Thr immu-noblot (Anti-P-Thr) when plants were acclimated tostate I or state II light conditions. Both wild-type plantsand the ffc mutant showed increased phosphorylationof LHCII and PSII core subunits in state II conditions,while P-LHCII and phosphorylated PSII core subunitswere absent in both ch1-2 and stn7/8 mutants (Fig. 5A),implying that the kinases STN7 and STN8 are not ac-tivated in the ch1-2 mutant. Thus, it is suggested thatthe lack of state transition-dependent excitation energytransfer from LHCII to PSI in ch1-2 (Figs. 3 and 4) cor-relates with the lack of P-LHCII (Fig. 5A). It is worthmentioning that the content of phosphorylated PSIIcore subunits in the ffcmutant in state II conditions washigher than in the wild-type plants, which indicatesthat ffc is subjected to photoinhibition (Bonardi et al.,
2005; Tikkanen et al., 2008b). In contrast, in the chaosmutant, P-LHCII was detected in state II conditions, butin lesser amounts than in Ler-0 plants (Fig. 5A). Semi-quantitative analysis of the immunoblot in Figure 5Asuggested that the LHCB1 level in chaos was highlycorrelated with the P-LHCII level (Fig. 5B). This findingimplies that STN7 in chaos was activated to phosphor-ylate LHCII under state II conditions. However, nostate transition was actually observed in the chaos mu-tant, which in this respect behaves like ch1-2 and stn7/8.
Protein Composition of Photosynthetic Complexes in ch1-2and cpsrp Mutants
Although phosphorylation of LHCII occurred in thechaos mutant under state II conditions (Fig. 5), LHCII-PSI-LHCI supercomplexes were not detectable by BN
Figure 4. Analysis of state transitions bylpBN-PA gel electrophoresis. A, Equalamounts of thylakoid membranes (9 mgof Chl) from wild-type plants (Col-0 andLer-0) and the stn7/8, ch1-2, ffc, andchaos mutant plants, which had beenadapted to state I light (far-red light of730 nm) or state II light (the red light of660 nm), were solubilizedwith 1% (w/v)digitonin and fractionated by lpBN-PAGE. B, Individual lanes from thelpBN-PA gel in Awere then subjected toSDS-urea-PAGE in the second dimen-sion. Total proteins were visualized byCoomassie Brilliant Blue staining. Iden-tities of the relevant proteins are indi-cated by arrows. The major PSI proteins,PsaA/B, as well as LHCII proteins in theLHCII-PSI-LHCII complexes are circled.Two biological replicates were performed,and similar results were obtained.
Plant Physiol. Vol. 172, 2016 1525
State Transitions in chlorina and cpSRP Mutants
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
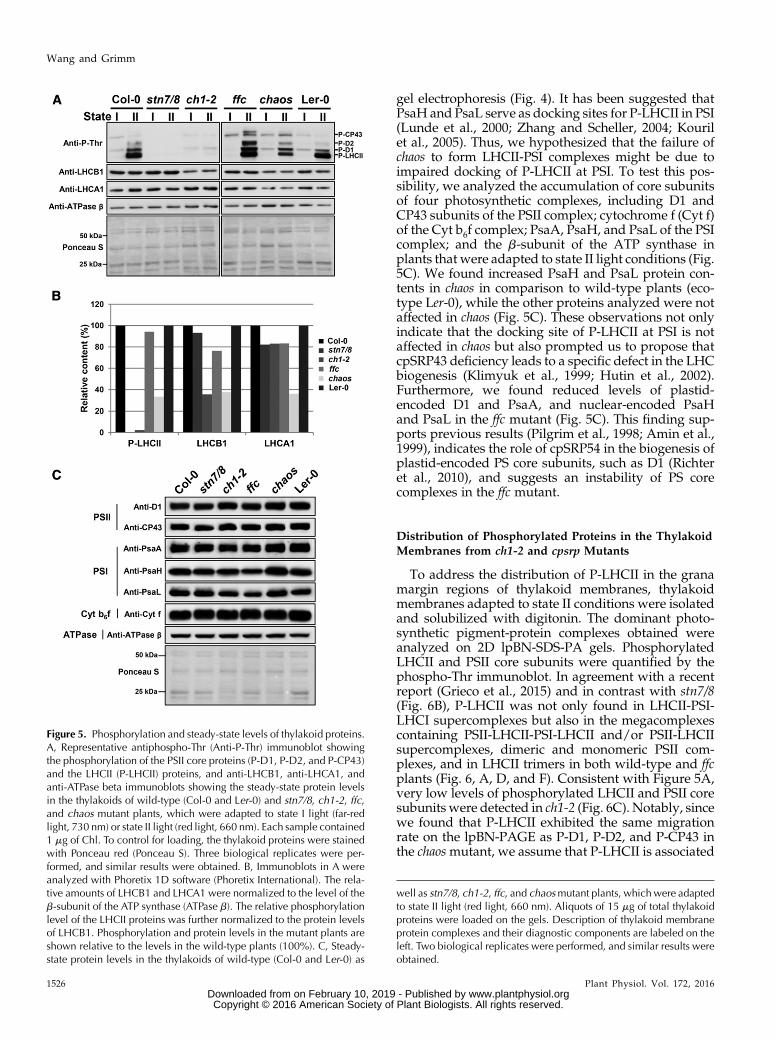
gel electrophoresis (Fig. 4). It has been suggested thatPsaH and PsaL serve as docking sites for P-LHCII in PSI(Lunde et al., 2000; Zhang and Scheller, 2004; Kourilet al., 2005). Thus, we hypothesized that the failure ofchaos to form LHCII-PSI complexes might be due toimpaired docking of P-LHCII at PSI. To test this pos-sibility, we analyzed the accumulation of core subunitsof four photosynthetic complexes, including D1 andCP43 subunits of the PSII complex; cytochrome f (Cyt f)of the Cyt b6f complex; PsaA, PsaH, and PsaL of the PSIcomplex; and the b-subunit of the ATP synthase inplants that were adapted to state II light conditions (Fig.5C). We found increased PsaH and PsaL protein con-tents in chaos in comparison to wild-type plants (eco-type Ler-0), while the other proteins analyzed were notaffected in chaos (Fig. 5C). These observations not onlyindicate that the docking site of P-LHCII at PSI is notaffected in chaos but also prompted us to propose thatcpSRP43 deficiency leads to a specific defect in the LHCbiogenesis (Klimyuk et al., 1999; Hutin et al., 2002).Furthermore, we found reduced levels of plastid-encoded D1 and PsaA, and nuclear-encoded PsaHand PsaL in the ffc mutant (Fig. 5C). This finding sup-ports previous results (Pilgrim et al., 1998; Amin et al.,1999), indicates the role of cpSRP54 in the biogenesis ofplastid-encoded PS core subunits, such as D1 (Richteret al., 2010), and suggests an instability of PS corecomplexes in the ffc mutant.
Distribution of Phosphorylated Proteins in the ThylakoidMembranes from ch1-2 and cpsrp Mutants
To address the distribution of P-LHCII in the granamargin regions of thylakoid membranes, thylakoidmembranes adapted to state II conditions were isolatedand solubilized with digitonin. The dominant photo-synthetic pigment-protein complexes obtained wereanalyzed on 2D lpBN-SDS-PA gels. PhosphorylatedLHCII and PSII core subunits were quantified by thephospho-Thr immunoblot. In agreement with a recentreport (Grieco et al., 2015) and in contrast with stn7/8(Fig. 6B), P-LHCII was not only found in LHCII-PSI-LHCI supercomplexes but also in the megacomplexescontaining PSII-LHCII-PSI-LHCII and/or PSII-LHCIIsupercomplexes, dimeric and monomeric PSII com-plexes, and in LHCII trimers in both wild-type and ffcplants (Fig. 6, A, D, and F). Consistent with Figure 5A,very low levels of phosphorylated LHCII and PSII coresubunits were detected in ch1-2 (Fig. 6C). Notably, sincewe found that P-LHCII exhibited the same migrationrate on the lpBN-PAGE as P-D1, P-D2, and P-CP43 inthe chaosmutant, we assume that P-LHCII is associated
Figure 5. Phosphorylation and steady-state levels of thylakoid proteins.A, Representative antiphospho-Thr (Anti-P-Thr) immunoblot showingthe phosphorylation of the PSII core proteins (P-D1, P-D2, and P-CP43)and the LHCII (P-LHCII) proteins, and anti-LHCB1, anti-LHCA1, andanti-ATPase beta immunoblots showing the steady-state protein levelsin the thylakoids of wild-type (Col-0 and Ler-0) and stn7/8, ch1-2, ffc,and chaos mutant plants, which were adapted to state I light (far-redlight, 730 nm) or state II light (red light, 660 nm). Each sample contained1 mg of Chl. To control for loading, the thylakoid proteins were stainedwith Ponceau red (Ponceau S). Three biological replicates were per-formed, and similar results were obtained. B, Immunoblots in A wereanalyzed with Phoretix 1D software (Phoretix International). The rela-tive amounts of LHCB1 and LHCA1 were normalized to the level of theb-subunit of the ATP synthase (ATPase b). The relative phosphorylationlevel of the LHCII proteins was further normalized to the protein levelsof LHCB1. Phosphorylation and protein levels in the mutant plants areshown relative to the levels in the wild-type plants (100%). C, Steady-state protein levels in the thylakoids of wild-type (Col-0 and Ler-0) as
well as stn7/8, ch1-2, ffc, and chaosmutant plants, whichwere adaptedto state II light (red light, 660 nm). Aliquots of 15 mg of total thylakoidproteins were loaded on the gels. Description of thylakoid membraneprotein complexes and their diagnostic components are labeled on theleft. Two biological replicates were performed, and similar results wereobtained.
1526 Plant Physiol. Vol. 172, 2016
Wang and Grimm
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
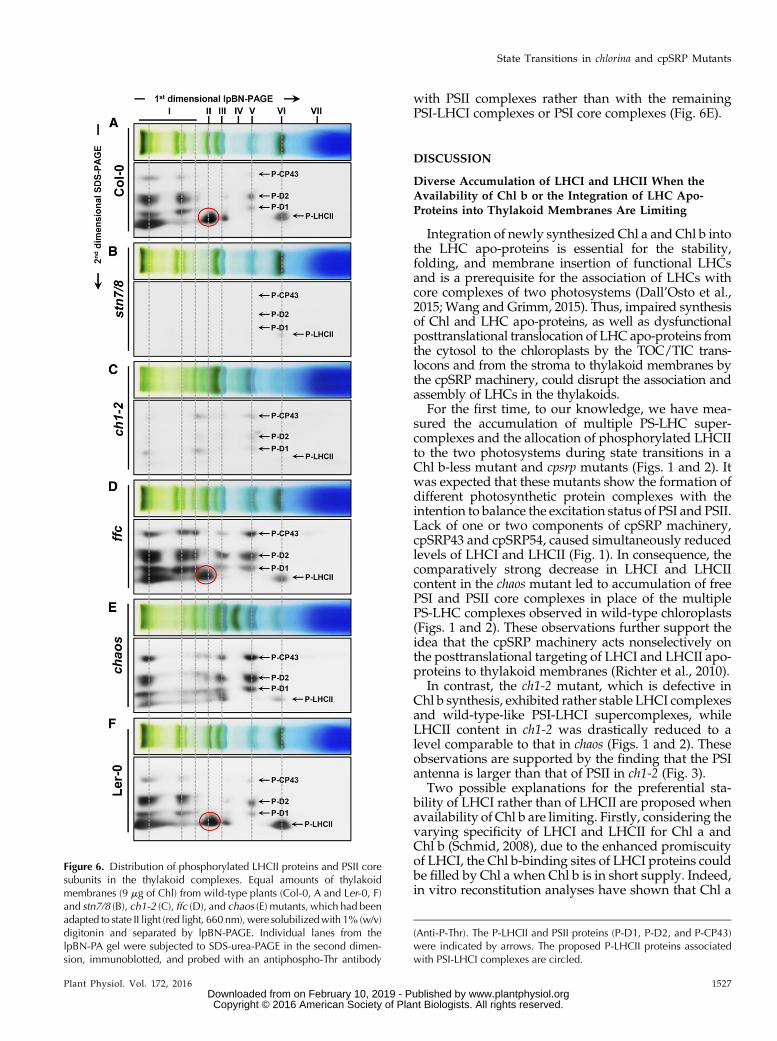
with PSII complexes rather than with the remainingPSI-LHCI complexes or PSI core complexes (Fig. 6E).
DISCUSSION
Diverse Accumulation of LHCI and LHCII When theAvailability of Chl b or the Integration of LHC Apo-Proteins into Thylakoid Membranes Are Limiting
Integration of newly synthesized Chl a and Chl b intothe LHC apo-proteins is essential for the stability,folding, and membrane insertion of functional LHCsand is a prerequisite for the association of LHCs withcore complexes of two photosystems (Dall’Osto et al.,2015; Wang and Grimm, 2015). Thus, impaired synthesisof Chl and LHC apo-proteins, as well as dysfunctionalposttranslational translocation of LHC apo-proteins fromthe cytosol to the chloroplasts by the TOC/TIC trans-locons and from the stroma to thylakoid membranes bythe cpSRP machinery, could disrupt the association andassembly of LHCs in the thylakoids.
For the first time, to our knowledge, we have mea-sured the accumulation of multiple PS-LHC super-complexes and the allocation of phosphorylated LHCIIto the two photosystems during state transitions in aChl b-less mutant and cpsrp mutants (Figs. 1 and 2). Itwas expected that these mutants show the formation ofdifferent photosynthetic protein complexes with theintention to balance the excitation status of PSI and PSII.Lack of one or two components of cpSRP machinery,cpSRP43 and cpSRP54, caused simultaneously reducedlevels of LHCI and LHCII (Fig. 1). In consequence, thecomparatively strong decrease in LHCI and LHCIIcontent in the chaos mutant led to accumulation of freePSI and PSII core complexes in place of the multiplePS-LHC complexes observed in wild-type chloroplasts(Figs. 1 and 2). These observations further support theidea that the cpSRP machinery acts nonselectively onthe posttranslational targeting of LHCI and LHCII apo-proteins to thylakoid membranes (Richter et al., 2010).
In contrast, the ch1-2 mutant, which is defective inChl b synthesis, exhibited rather stable LHCI complexesand wild-type-like PSI-LHCI supercomplexes, whileLHCII content in ch1-2 was drastically reduced to alevel comparable to that in chaos (Figs. 1 and 2). Theseobservations are supported by the finding that the PSIantenna is larger than that of PSII in ch1-2 (Fig. 3).
Two possible explanations for the preferential sta-bility of LHCI rather than of LHCII are proposed whenavailability of Chl b are limiting. Firstly, considering thevarying specificity of LHCI and LHCII for Chl a andChl b (Schmid, 2008), due to the enhanced promiscuityof LHCI, the Chl b-binding sites of LHCI proteins couldbe filled by Chl a when Chl b is in short supply. Indeed,in vitro reconstitution analyses have shown that Chl a
Figure 6. Distribution of phosphorylated LHCII proteins and PSII coresubunits in the thylakoid complexes. Equal amounts of thylakoidmembranes (9 mg of Chl) from wild-type plants (Col-0, A and Ler-0, F)and stn7/8 (B), ch1-2 (C), ffc (D), and chaos (E) mutants, which had beenadapted to state II light (red light, 660 nm),were solubilizedwith 1% (w/v)digitonin and separated by lpBN-PAGE. Individual lanes from thelpBN-PA gel were subjected to SDS-urea-PAGE in the second dimen-sion, immunoblotted, and probed with an antiphospho-Thr antibody
(Anti-P-Thr). The P-LHCII and PSII proteins (P-D1, P-D2, and P-CP43)were indicated by arrows. The proposed P-LHCII proteins associatedwith PSI-LHCI complexes are circled.
Plant Physiol. Vol. 172, 2016 1527
State Transitions in chlorina and cpSRP Mutants
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

can in fact be integrated into LHCI apo-proteins, suchas LHCA2 and LHCA4, to form the stable LHCI(Schmid et al., 2002). Furthermore, the Chl a-containingLHCI has been characterized in the ch1-1mutant, whichlacks Chl b altogether (Havaux et al., 2007; Takabayashiet al., 2011). However, the Chl a-containing LHCI wasless tightly associated with PSI core complexes, indi-cating Chl b is essential for the efficient energy transferand stable assembly of PSI-LHCI supercomplexes(Takabayashi et al., 2011). Thus, the residual amount ofChl b in ch1-2 (20% of the wild-type level) might well besufficient for the organization of functional PSI-LHCIsupercomplexes (Figs. 1–3).
Secondly, we suggest that newly synthesized Chl bmight be preferentially integrated into LHCI ratherthan LHCII, particularly when only limited amounts ofChl b are available. This hypothesis can be supportedby our finding of the preferential accumulation of LHCIin ch1-2 (Figs. 1 and 2). So far, very little attention hasbeen paid to the mechanisms that determine the dis-tribution of newly synthesized Chl to the various LHCand PS-LHC complexes. What knowledge we dohave is based on radioactive labeling with 14C. Theradiolabeling experiments with Chl precursor mole-cules carried out on organisms exposed to high-lightlevels confirmed ongoing Chl synthesis in both higherplants (Beisel et al., 2010) and cyanobacteria (Kopecnaet al., 2012). Interestingly, in the cyanobacteria, thefreshly synthesized Chl was localized predominantly inPSI and to a lesser extent in PSII (Kopecna et al., 2012).In contrast, most of the fresh Chls produced in thechloroplasts of higher plants were suggested to supportthe PSII repair cycle, since PSI is very stable underhigh-light stress (Feierabend and Dehne, 1996). In thiscontext, ch1-2 could be a useful tool for assessing therelative affinities of LHCI and LHCII for Chl b duringtheir biogenesis.
Modified State Transitions Observed in ch1-2 and chaosImply the Flexible Association of LHCs withTwo Photosystems
To balance the excitation status of PSI and PSII, statetransitions enable the rapid and efficient modificationof the relative antenna size of the two photosystems inresponse to fluctuating light conditions (Allen andForsberg, 2001; Rochaix, 2011; Goldschmidt-Clermontand Bassi, 2015; Gollan et al., 2015). In the state I-to-stateII transition, phosphorylated LHCII proteins associatewith PSI-LHCI to favor the absorption cross-section ofPSI. However, increased absorption cross-section of PSIand formation of LHCII-PSI complexes were notdetected in ch1-2 and chaos upon exposure to PSII-favoring light (Figs. 3 and 4), suggesting a block ofthe state I-to-state II transition in both ch1-2 and chaosmutants.
According to the canonical model of state transitions,phosphorylation of LHCII is an essential prerequisitefor state I-to-state II transition and triggers the disso-ciation of LHCII from PSII and promotes its lateral
migration to PSI-LHCI-enriched regions of thylakoidmembranes (Allen, 1992; Rochaix, 2011). In this way,the missing formation of LHCII-PSI-LHCI complexes inch1-2 is associated with the lack of P-LHCII under stateII light conditions (Figs. 5A and 6C), which is explainedby repression of STN7 activity. It is striking that incomparison with wild-type and cpsrp mutant plants,ch1-2 exhibited 2-fold larger antenna size of PSI thanthat of PSII (Fig. 3). In contrast, chaos exhibited theLHCII antenna similar to that in ch1-2 but less LHCIantenna (Figs. 1 and 2). This leads to the comparablybalanced excitation state of PSI and PSII. The phos-phorylation of LHCII was observed in chaos in the stateII conditions (Figs. 5A and 6E), suggesting the moreactivated STN7 in chaos mutant than that in ch1-2 mu-tant. We assume that the electron transfer chain and thePQ pool were more oxidized in ch1-2 than in wild-typeand chaosmutants. In turn, oxidation of PQ pool in ch1-2will lead to inactivation of STN7.
As phosphorylation of LHCII is observed in chaosupon exposure to PSII light, balanced distribution ofexcitation energy between PSI and PSII is likely to berequired under state II conditions. However, P-LHCIIof chaoswas associated with PSII complexes rather thanwith PSI core complexes or a residual amount of intactPSI-LHCI supercomplexes (Fig. 6E). The localization ofP-LHCII could result from a failure to dissociate fromPSII or an inability to dock at PSI. The latter prospect ischallenged by the elevated accumulation of LHCII-PSIcore complexes in the lhca4mutant (Benson et al., 2015)and the DLhcamutant (Bressan et al., 2016), which wereadapted to state II conditions. Structural studies ofLHCII-PSI-LHCI complexes showed an opposite lo-calization of P-LHCII and LHCI within the state-transition supercomplexes (Kouril et al., 2005; Dropet al., 2014). It is very unlikely that LHCI contributes tostable docking of LHCII at PSI complexes.
We propose that an impaired dissociation of P-LHCIIfrom PSII results from a strongly reduced content ofLHCII in chaos (Figs. 1 and 2). It is expected that amobile P-LHCII pool is limited to migrate toward PSIcomplexes and form LHCII-PSI complexes. Althoughfree phosphorylated LHCII trimers could be detected inchaos (Fig. 6E), the majority of LHCII trimers were as-sociated with dimeric PSII core complexes (Figs. 2 and4). According to the binding affinity of LHCII trimer tothe PSII homodimer, S (strong), M (medium), and L(loose) variants of LHCII trimers were found in thethylakoid membranes of higher plants (Dekker andBoekema, 2005). It was indicated that the L-LHCIItrimers could be associated with PSI, while the S-LHCIIand M-LHCII are unlikely to be involved in state tran-sitions (Pietrzykowska et al., 2014). Thus, the failure toform P-LHCII-PSI complexes in chaos is proposed to bedue to the lack of a mobile LHCII pool.
In summary, the distinct accumulation of LHCI andLHCII complexes in ch1-2 and cpsrp mutants not onlyunderlines the requirement for coordination of Chl bi-osynthesis and the posttranslational integration ofLHC apo-proteins into thylakoid membranes (Dall’Osto
1528 Plant Physiol. Vol. 172, 2016
Wang and Grimm
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

et al., 2015; Wang and Grimm, 2015), but also indicatesthe variable accumulation of LHCI and LHCII com-plexes, when Chl b synthesis is compromised in ch1-2mutant. Furthermore, the detailed comparative analy-sis of state transitions in ch1-2 and chaos mutants pro-vides evidence for the flexible association of LHCs withthe two photosystems.
MATERIALS AND METHODS
Plant Materials, Growth Conditions, and Light Treatment
The following Arabidopsis (Arabidopsis thaliana) mutants were used in thisstudy: ch1-2, which contains a V274E mutation in the Rieske binding site(CS3120; Espineda et al., 1999), the maize Ds transposon-containing cpsrp43mutant chaos (Klimyuk et al., 1999), the T-DNA insertion lines ffc (cpsrp54,CS850421; Pilgrim et al., 1998), cpftsy (SALK_049077; Tzvetkova-Chevolleauet al., 2007) and stn7/8 (Bonardi et al., 2005), and the chaos/ffc double mutant(Hutin et al., 2002), together with the wild-type Arabidopsis Col-0 and Ler-0.Wild-type and mutant Arabidopsis plants were routinely grown at 22°C and70% humidity with 100 mM photons m22 s21 on a 16-h-light/8-h-dark photo-period.
Pigment Analysis
Chls were extracted from leaves with alkaline acetone (100% acetone:0.2 M
NH4OH, 9:1) and analyzed using reversed-phase chromatography on an Agi-lent HPLC system as described (Schlicke et al., 2014).
Isolation of Thylakoid Membranes
Thylakoidmembraneswere isolated fromArabidopsis plants grown inwell-controlled phytochambers or adapted to state I or state II conditions in thepresence of 10 mM sodium fluoride NaF as described (Järvi et al., 2011). Chlconcentration was determined as described (Wellburn, 1994).
77K Fluorescence Emission Spectroscopy
Freshly isolated thylakoids equivalent to 10 mg Chl mL21 were resuspendedin Chl fluorescence buffer (20 mM HEPES, pH 7.8, 60% glycerol, 300 mM Suc,5 mM MgCl2) and frozen in liquid nitrogen. Chl a fluorescence emission wasdetected using a F-6500 fluorometer (Jasco). The samplewas excited at a 475-nmwavelength. The emission spectra between 655 nm and 800 nm were recordedwith a bandwidth of 10 nm.
Second Dimensional lpBN-SDS-PAGE
lpBN-PAGE was performed essentially according to Järvi et al. (2011). Tocomprehensively analyze the PSI-LHC supercomplexes present in grana andunstacked thylakoids, freshly isolated thylakoids equivalent to 0.5 mg Chl mL21 weresolubilized with 1% b-DM at 4°C for 5 min. To detect the LHCII-PSI-LHCIsupercomplexes formed during state transitions, freshly isolated thylakoids werepartially solubilized with 1% (w/v) digitonin at room temperature for 15 min. Forthe second dimensional SDS-PAGE analysis, the excised lpBN-PAGE lanes weresoaked in SDS sample buffer (50 mM Tris-HCl, pH 6.8, 2% SDS, 10% glycerol,0.002%bromophenol blue, and 50mMDTT) for 0.5 h at room temperature and thenlayered onto 12% SDS-PAGE gels containing 6 M urea. The gels were stained withCoomassie Brilliant Blue G250 or used for immunoblot analyses.
Immunoblot Analyses
For immunoblot analysis, total thylakoid proteins normalized to equal Chlcontents were separated on 12% SDS-PAGE gels containing 6 M urea. Afterelectrophoresis, proteins were transferred to nitrocellulose membranes (GEHealthcare) and probed with specific antibodies directed against the light-harvesting antenna proteins LHCA1 and LHCB1 (Agrisera); the PSI core subu-nits D1 and CP43 (Agrisera); the Cyt b6f subunit Cyt f (Agrisera); the PSI coresubunits PsaA, PsaH, and PsaL (Agrisera); the ATP synthase b-subunit (Agrisera);
and phosphorylated thylakoid proteins (anti-P-Thr,NewEngland Biolabs). Signalswere detected with the SuperSignal West Pico Chemiluminescent Substrate(Thermo Scientific).
ACKNOWLEDGMENTS
We thank Dr. Danja Schünemann for all cpsrp mutants and discussion ofimpaired PSI-LHCI complexes in chaos mutant, and Dr. Dario Leister for thestn7/8 mutant.
Received June 24, 2016; accepted September 21, 2016; published September 23,2016.
LITERATURE CITED
Albertsson P (2001) A quantitative model of the domain structure of thephotosynthetic membrane. Trends Plant Sci 6: 349–358
Albertsson PA, Andreasson E, Svensson P (1990) The domain organizationof the plant thylakoid membrane. FEBS Lett 273: 36–40
Allen JF (1992) Protein phosphorylation in regulation of photosynthesis.Biochim Biophys Acta 1098: 275–335
Allen JF, Forsberg J (2001) Molecular recognition in thylakoid structureand function. Trends Plant Sci 6: 317–326
Amin P, Sy DA, Pilgrim ML, Parry DH, Nussaume L, Hoffman NE (1999)Arabidopsis mutants lacking the 43- and 54-kilodalton subunits of thechloroplast signal recognition particle have distinct phenotypes. PlantPhysiol 121: 61–70
Beisel KG, Jahnke S, Hofmann D, Köppchen S, Schurr U, Matsubara S(2010) Continuous turnover of carotenes and chlorophyll a in matureleaves of Arabidopsis revealed by 14CO2 pulse-chase labeling. PlantPhysiol 152: 2188–2199
Bell AJ, Frankel LK, Bricker TM (2015) High yield non-detergent isolationof photosystem i-light-harvesting chlorophyll II membranes from spin-ach thylakoids: implications for the organization of the PS I antennae inhigher plants. J Biol Chem 290: 18429–18437
Bellafiore S, Barneche F, Peltier G, Rochaix JD (2005) State transitions andlight adaptation require chloroplast thylakoid protein kinase STN7.Nature 433: 892–895
Benson SL, Maheswaran P, Ware MA, Hunter CN, Horton P, Jansson S,Ruban AV, Johnson MP (2015) An intact light harvesting complex Iantenna system is required for complete state transitions in Arabidopsis.Nat Plants 1: 15176
Bonardi V, Pesaresi P, Becker T, Schleiff E, Wagner R, Pfannschmidt T,Jahns P, Leister D (2005) Photosystem II core phosphorylation andphotosynthetic acclimation require two different protein kinases. Nature437: 1179–1182
Bressan M, Dall’Osto L, Bargigia I, Alcocer MJP, Viola D, Cerullo G,D’Andrea C, Bassi R, Ballottari M (2016) LHCII can substitute for LHCIas an antenna for photosystem I but with reduced light-harvesting ca-pacity. Nat Plants 2: 16131
Brzezowski P, Richter AS, Grimm B (2015) Regulation and function oftetrapyrrole biosynthesis in plants and algae. Biochim Biophys Acta1847: 968–985
Chuartzman SG, Nevo R, Shimoni E, Charuvi D, Kiss V, Ohad I,Brumfeld V, Reich Z (2008) Thylakoid membrane remodeling duringstate transitions in Arabidopsis. Plant Cell 20: 1029–1039
Crepin A, Caffarri S (2015) The specific localizations of phosphorylatedLhcb1 and Lhcb2 isoforms reveal the role of Lhcb2 in the formation ofthe PSI-LHCII supercomplex in Arabidopsis during state transitions.Biochim Biophys Acta 1847: 1539–1548
Dall’Osto L, Bressan M, Bassi R (2015) Biogenesis of light harvestingproteins. Biochim Biophys Acta 1847: 861–871
Dekker JP, Boekema EJ (2005) Supramolecular organization of thylakoidmembrane proteins in green plants. Biochim Biophys Acta 1706: 12–39
Drop B, Yadav K N S, Boekema EJ, Croce R (2014) Consequences of statetransitions on the structural and functional organization of photosystemI in the green alga Chlamydomonas reinhardtii. Plant J 78: 181–191
Espineda CE, Linford AS, Devine D, Brusslan JA (1999) The AtCAO gene,encoding chlorophyll a oxygenase, is required for chlorophyll b syn-thesis in Arabidopsis thaliana. Proc Natl Acad Sci USA 96: 10507–10511
Feierabend J, Dehne S (1996) Fate of the porphyrin cofactors during thelight-dependent turnover of catalase and of the photosystem II re-actioncenter protein D1 in mature rye leaves. Planta 198: 413–422
Plant Physiol. Vol. 172, 2016 1529
State Transitions in chlorina and cpSRP Mutants
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Galka P, Santabarbara S, Khuong TT, Degand H, Morsomme P, JenningsRC, Boekema EJ, Caffarri S (2012) Functional analyses of the plantphotosystem I-light-harvesting complex II supercomplex reveal thatlight-harvesting complex II loosely bound to photosystem II is a veryefficient antenna for photosystem I in state II. Plant Cell 24: 2963–2978
Goldschmidt-Clermont M, Bassi R (2015) Sharing light between twophotosystems: mechanism of state transitions. Curr Opin Plant Biol 25:71–78
Gollan PJ, Tikkanen M, Aro EM (2015) Photosynthetic light reactions:integral to chloroplast retrograde signalling. Curr Opin Plant Biol 27:180–191
Grieco M, Suorsa M, Jajoo A, Tikkanen M, Aro EM (2015) Light-harvesting II antenna trimers connect energetically the entire photo-synthetic machinery including both photosystems II and I. Biochim BiophysActa 1847: 607–619
Grieco M, Tikkanen M, Paakkarinen V, Kangasjärvi S, Aro EM (2012)Steady-state phosphorylation of light-harvesting complex II proteinspreserves photosystem I under fluctuating white light. Plant Physiol160: 1896–1910
Haferkamp S, Haase W, Pascal AA, van Amerongen H, Kirchhoff H(2010) Efficient light harvesting by photosystem II requires an optimizedprotein packing density in grana thylakoids. J Biol Chem 285: 17020–17028
Havaux M, Dall’osto L, Bassi R (2007) Zeaxanthin has enhanced antioxi-dant capacity with respect to all other xanthophylls in Arabidopsisleaves and functions independent of binding to PSII antennae. PlantPhysiol 145: 1506–1520
Hutin C, Havaux M, Carde JP, Kloppstech K, Meiherhoff K, Hoffman N,Nussaume L (2002) Double mutation cpSRP43–/cpSRP54– is necessaryto abolish the cpSRP pathway required for thylakoid targeting of thelight-harvesting chlorophyll proteins. Plant J 29: 531–543
Jansson S (1994) The light-harvesting chlorophyll a/b-binding proteins.Biochim Biophys Acta 1184: 1–19
Järvi S, Suorsa M, Paakkarinen V, Aro EM (2011) Optimized native gelsystems for separation of thylakoid protein complexes: novel super- andmega-complexes. Biochem J 439: 207–214
Jarvis P, López-Juez E (2013) Biogenesis and homeostasis of chloroplastsand other plastids. Nat Rev Mol Cell Biol 14: 787–802
Kim EH, Li XP, Razeghifard R, Anderson JM, Niyogi KK, Pogson BJ,ChowWS (2009) The multiple roles of light-harvesting chlorophyll a/b-proteincomplexes define structure and optimize function of Arabidopsis chloro-plasts: a study using two chlorophyll b-less mutants. Biochim Biophys Acta1787: 973–984
Klimyuk VI, Persello-Cartieaux F, Havaux M, Contard-David P, SchuenemannD, Meiherhoff K, Gouet P, Jones JD, Hoffman NE, Nussaume L (1999) Achromodomain protein encoded by the arabidopsis CAO gene is a plant-specific component of the chloroplast signal recognition particle pathwaythat is involved in LHCP targeting. Plant Cell 11: 87–99
Kopecná J, Komenda J, Bucinská L, Sobotka R (2012) Long-term accli-mation of the cyanobacterium Synechocystis sp. PCC 6803 to high lightis accompanied by an enhanced production of chlorophyll that is pref-erentially channeled to trimeric photosystem I. Plant Physiol 160: 2239–2250
Kouril R, Zygadlo A, Arteni AA, de Wit CD, Dekker JP, Jensen PE,Scheller HV, Boekema EJ (2005) Structural characterization of a com-plex of photosystem I and light-harvesting complex II of Arabidopsisthaliana. Biochemistry 44: 10935–10940
Leoni C, Pietrzykowska M, Kiss AZ, Suorsa M, Ceci LR, Aro EM, JanssonS (2013) Very rapid phosphorylation kinetics suggest a unique role forLhcb2 during state transitions in Arabidopsis. Plant J 76: 236–246
Liang FC, Kroon G, McAvoy CZ, Chi C, Wright PE, Shan SO (2016)Conformational dynamics of a membrane protein chaperone enablesspatially regulated substrate capture and release. Proc Natl Acad SciUSA 113: E1615–E1624
Longoni P, Douchi D, Cariti F, Fucile G, Goldschmidt-Clermont M (2015)Phosphorylation of the light-harvesting complex II isoform Lhcb2 iscentral to state transitions. Plant Physiol 169: 2874–2883
Lunde C, Jensen PE, Haldrup A, Knoetzel J, Scheller HV (2000) The PSI-Hsubunit of photosystem I is essential for state transitions in plant pho-tosynthesis. Nature 408: 613–615
Murray DL, Kohorn BD (1991) Chloroplasts of Arabidopsis thaliana ho-mozygous for the ch-1 locus lack chlorophyll b, lack stable LHCPII andhave stacked thylakoids. Plant Mol Biol 16: 71–79
Nelson N, Yocum CF (2006) Structure and function of photosystems I andII. Annu Rev Plant Biol 57: 521–565
Nield J, Barber J (2006) Refinement of the structural model for the photo-system II supercomplex of higher plants. Biochim Biophys Acta 1757:353–361
Ouyang M, Li X, Ma J, Chi W, Xiao J, Zou M, Chen F, Lu C, Zhang L(2011) LTD is a protein required for sorting light-harvesting chlorophyll-binding proteins to the chloroplast SRP pathway. Nat Commun 2: 277
Pesaresi P, Hertle A, Pribil M, Kleine T, Wagner R, Strissel H, IhnatowiczA, Bonardi V, Scharfenberg M, Schneider A et al (2009) ArabidopsisSTN7 kinase provides a link between short- and long-term photosyn-thetic acclimation. Plant Cell 21: 2402–2423
Pietrzykowska M, Suorsa M, Semchonok DA, Tikkanen M, Boekema EJ,Aro EM, Jansson S (2014) The light-harvesting chlorophyll a/b bindingproteins Lhcb1 and Lhcb2 play complementary roles during state tran-sitions in Arabidopsis. Plant Cell 26: 3646–3660
Pilgrim ML, van Wijk KJ, Parry DH, Sy DA, Hoffman NE (1998) Ex-pression of a dominant negative form of cpSRP54 inhibits chloroplastbiogenesis in Arabidopsis. Plant J 13: 177–186
Plöchinger M, Torabi S, Rantala M, Tikkanen M, Suorsa M, Jensen PE,Aro EM, Meurer J (2016) The low molecular weight protein PsaI sta-bilizes the light-harvesting complex II docking site of photosystem I.Plant Physiol 172: 450–463
Pribil M, Pesaresi P, Hertle A, Barbato R, Leister D (2010) Role of plastidprotein phosphatase TAP38 in LHCII dephosphorylation and thylakoidelectron flow. PLoS Biol 8: e1000288
Qin X, Suga M, Kuang T, Shen JR (2015) Photosynthesis. Structural basisfor energy transfer pathways in the plant PSI-LHCI supercomplex. Sci-ence 348: 989–995
Richter CV, Bals T, Schünemann D (2010) Component interactions, reg-ulation and mechanisms of chloroplast signal recognition particle-dependent protein transport. Eur J Cell Biol 89: 965–973
Rochaix JD (2011) Assembly of the photosynthetic apparatus. Plant Physiol155: 1493–1500
Schlicke H, Hartwig AS, Firtzlaff V, Richter AS, Glässer C, Maier K,Finkemeier I, Grimm B (2014) Induced deactivation of genes encodingchlorophyll biosynthesis enzymes disentangles tetrapyrrole-mediatedretrograde signaling. Mol Plant 7: 1211–1227
Schmid VH (2008) Light-harvesting complexes of vascular plants. Cell MolLife Sci 65: 3619–3639
Schmid VH, Potthast S, Wiener M, Bergauer V, Paulsen H, Storf S (2002)Pigment binding of photosystem I light-harvesting proteins. J Biol Chem277: 37307–37314
Shapiguzov A, Chai X, Fucile G, Longoni P, Zhang L, Rochaix JD (2016)Activation of the Stt7/STN7 kinase through dynamic interactions withthe cytochrome b6f complex. Plant Physiol 171: 82–92
Shapiguzov A, Ingelsson B, Samol I, Andres C, Kessler F, Rochaix JD,Vener AV, Goldschmidt-Clermont M (2010) The PPH1 phosphatase isspecifically involved in LHCII dephosphorylation and state transitionsin Arabidopsis. Proc Natl Acad Sci USA 107: 4782–4787
Sundberg E, Slagter JG, Fridborg I, Cleary SP, Robinson C, Coupland G(1997) ALBINO3, an Arabidopsis nuclear gene essential for chloroplastdifferentiation, encodes a chloroplast protein that shows homology toproteins present in bacterial membranes and yeast mitochondria. PlantCell 9: 717–730
Suorsa M, Rantala M, Mamedov F, Lespinasse M, Trotta A, Grieco M,Vuorio E, Tikkanen M, Järvi S, Aro EM (2015) Light acclimation in-volves dynamic re-organization of the pigment-protein megacomplexesin non-appressed thylakoid domains. Plant J 84: 360–373
Takabayashi A, Kurihara K, Kuwano M, Kasahara Y, Tanaka R, Tanaka A(2011) The oligomeric states of the photosystems and the light-harvestingcomplexes in the Chl b-less mutant. Plant Cell Physiol 52: 2103–2114
Tanaka A, Ito H, Tanaka R, Tanaka NK, Yoshida K, Okada K (1998)Chlorophyll a oxygenase (CAO) is involved in chlorophyll b formationfrom chlorophyll a. Proc Natl Acad Sci USA 95: 12719–12723
Tanaka R, Tanaka A (2007) Tetrapyrrole biosynthesis in higher plants.Annu Rev Plant Biol 58: 321–346
Tikkanen M, Grieco M, Kangasjärvi S, Aro EM (2010) Thylakoid proteinphosphorylation in higher plant chloroplasts optimizes electron transferunder fluctuating light. Plant Physiol 152: 723–735
Tikkanen M, Nurmi M, Kangasjärvi S, Aro EM (2008b) Core proteinphosphorylation facilitates the repair of photodamaged photosystem IIat high light. Biochim Biophys Acta 1777: 1432–1437
1530 Plant Physiol. Vol. 172, 2016
Wang and Grimm
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Tikkanen M, Nurmi M, Suorsa M, Danielsson R, Mamedov F, Styring S,Aro EM (2008a) Phosphorylation-dependent regulation of excitationenergy distribution between the two photosystems in higher plants.Biochim Biophys Acta 1777: 425–432
Tomitani A, Okada K, Miyashita H, Matthijs HC, Ohno T, Tanaka A(1999) Chlorophyll b and phycobilins in the common ancestor of cya-nobacteria and chloroplasts. Nature 400: 159–162
Tzvetkova-Chevolleau T, Hutin C, Noël LD, Goforth R, Carde JP, Caffarri S,Sinning I, Groves M, Teulon JM, Hoffman NE, et al (2007) Canonical signalrecognition particle components can be bypassed for posttranslational proteintargeting in chloroplasts. Plant Cell 19: 1635–1648
Vener AV, van Kan PJ, Rich PR, Ohad I, Andersson B (1997) Plastoquinolat the quinol oxidation site of reduced cytochrome bf mediates signaltransduction between light and protein phosphorylation: thylakoidprotein kinase deactivation by a single-turnover flash. Proc Natl AcadSci USA 94: 1585–1590
Wang P, Grimm B (2015) Organization of chlorophyll biosynthesis andinsertion of chlorophyll into the chlorophyll-binding proteins in chlo-roplasts. Photosynth Res 126: 189–202
Wellburn AR (1994) The spectral determination of chlorophyll a andchlorophyll b, as well as total carotenoids, using various solvents withspectrophotometers of different resolution. J Plant Physiol 144: 307–313
Wientjes E, Oostergetel GT, Jansson S, Boekema EJ, Croce R (2009) Therole of Lhca complexes in the supramolecular organization of higherplant photosystem I. J Biol Chem 284: 7803–7810
Wientjes E, van Amerongen H, Croce R (2013) LHCII is an antenna of bothphotosystems after long-term acclimation. Biochim Biophys Acta 1827:420–426
Wobbe L, Bassi R, Kruse O (2016) Multi-level light capture control inplants and green algae. Trends Plant Sci 21: 55–68
Yokono M, Takabayashi A, Akimoto S, Tanaka A (2015) A megacomplexcomposed of both photosystem reaction centres in higher plants. NatCommun 6: 6675
Zhang S, Scheller HV (2004) Light-harvesting complex II binds to severalsmall subunits of photosystem I. J Biol Chem 279: 3180–3187
Zito F, Finazzi G, Delosme R, Nitschke W, Picot D, Wollman FA (1999)The Qo site of cytochrome b6f complexes controls the activation of theLHCII kinase. EMBO J 18: 2961–2969
Plant Physiol. Vol. 172, 2016 1531
State Transitions in chlorina and cpSRP Mutants
www.plantphysiol.orgon February 10, 2019 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.