coding of vocalizations by single neurons in ventrolateral prefrontal cortex
TRANSCRIPT

lable at ScienceDirect
Hearing Research xxx (2013) 1e9
Contents lists avai
Hearing Research
journal homepage: www.elsevier .com/locate/heares
Review
Coding of vocalizations by single neurons in ventrolateral prefrontalcortex
Bethany Plakke, Mark D. Diltz, Lizabeth M. Romanski*
Dept. Neurobiology & Anatomy, Univ. of Rochester, Box 603, Rochester, NY 14642, USA
a r t i c l e i n f o
Article history:Received 26 December 2012Received in revised form20 June 2013Accepted 16 July 2013Available online xxx
* Corresponding author. University of RochesterNeurobiology and Anatomy, Box 603, 601 Elmwood AvTel.: þ1 585 273 1469; fax: þ1 585 442 8766.
E-mail address: [email protected]
0378-5955/$ e see front matter � 2013 Published byhttp://dx.doi.org/10.1016/j.heares.2013.07.011
Please cite this article in press as: Plakke, B.,(2013), http://dx.doi.org/10.1016/j.heares.20
a b s t r a c t
Neuronal activity in single prefrontal neurons has been correlated with behavioral responses, rules, taskvariables and stimulus features. In the non-human primate, neurons recorded in ventrolateral prefrontalcortex (VLPFC) have been found to respond to species-specific vocalizations. Previous studies have foundmultisensory neurons which respond to simultaneously presented faces and vocalizations in this region.Behavioral data suggests that face and vocal information are inextricably linked in animals and humansand therefore may also be tightly linked in the coding of communication calls in prefrontal neurons. Inthis study we therefore examined the role of VLPFC in encoding vocalization call type information.Specifically, we examined previously recorded single unit responses from the VLPFC in awake, behavingrhesus macaques in response to 3 types of species-specific vocalizations made by 3 individual callers.Analysis of responses by vocalization call type and caller identity showed that w19% of cells had a maineffect of call type with fewer cells encoding caller. Classification performance of VLPFC neurons wasw42% averaged across the population. When assessed at discrete time bins, classification performancereached 70 percent for coos in the first 300 ms and remained above chance for the duration of theresponse period, though performance was lower for other call types. In light of the sub-optimal classi-fication performance of the majority of VLPFC neurons when only vocal information is present, and therecent evidence that most VLPFC neurons are multisensory, the potential enhancement of classificationwith the addition of accompanying face information is discussed and additional studies recommended.Behavioral and neuronal evidence has shown a considerable benefit in recognition and memory per-formance when faces and voices are presented simultaneously. In the natural environment both facialand vocalization information is present simultaneously and neural systems no doubt evolved to integratemultisensory stimuli during recognition.
This article is part of a Special Issue entitled <Vocalizations and Hearing>.� 2013 Published by Elsevier B.V.
Previous work has identified an auditory region in the ventrallateral prefrontal cortex (VLPFC) that is responsive to complexsounds including species-specific vocalizations (Romanski andGoldman-Rakic, 2002). Species-specific vocalizations are complexsound stimuli with varying temporal and spectral features whichcan provide unique information to the listener. The type of calldelivered can indicate different behavioral contexts (food call vs.alarm call) which each elicit a unique response. Vocalizations canalso provide information about the individual uttering the call, andthus would include information on gender, social status, body size,and reproductive status (Hauser and Marler, 1993; Hauser, 1996;
Medical Center, Dept. ofe., Rochester, NY 14642, USA.
(L.M. Romanski).
Elsevier B.V.
et al., Coding of vocalizations13.07.011
Bradbury and Vehrencamp, 1998; Owings and Morton, 1998).Knowing how the brain encodes species-specific vocalizations cancontribute to our understanding of language and communicationprocessing.
Neurophysiological studies have begun to examine the specificacoustic features which neurons in the auditory cortex encode.Work by Wang and colleague’s has demonstrated that neurons inmarmoset auditory cortex lock to the temporal envelope of naturalmarmoset twitter calls (Wang et al., 1995). In addition, it has beenshown that the auditory cortex in marmosets has a specializedpitch processing region (Bendor and Wang, 2005) and that mar-mosets use both temporal and spectral cues to discriminate pitch(Bendor et al., 2012). Neurons in primary auditory cortex that arenot responsive to pure tones are highly selective to complex fea-tures of sounds, features that are found in vocalizations (Sadagopanand Wang, 2009).
by single neurons in ventrolateral prefrontal cortex, Hearing Research

B. Plakke et al. / Hearing Research xxx (2013) 1e92
Other key areas involved in vocalization processing include thebelt and parabelt regions of auditory cortex. While neurons inauditory core areas are responsive to simple stimuli, complexsounds including noise bursts and vocalizations activate regions ofthe belt and parabelt (Rauschecker et al., 1995, 1997; Rauschecker,1998; Kikuchi et al., 2010). Recently, it has been reported that theleft belt and parabelt are more active to complex spectral temporalpatterns (Joly et al., 2012). Furthermore, the selectivity of singleneurons in the anterolateral belt for vocalizations is similar to thatof VLPFC (Romanski et al., 2005; Tian et al., 2001) which arereciprocally connected (Romanski et al., 1999a,b).
This hierarchy of complex sound processing continues along thesuperior temporal plane (STP) (Kikuchi et al., 2010; Poremba et al.,2003) to the temporal pole (Poremba et al., 2004). An area on thesupratemporal plane has also been identified as a vocalization area inmacaque monkeys (Petkov et al., 2008) and cells in this region aremoreselective to individualvoices thancall type(Perrodinetal.,2011).These auditory regions including the belt, parabelt, and STP all projectto VLPFC (Hackett et al., 1999; Romanski et al., 1999a), which is pre-sumed to be the apex of complex auditory processing in the brain.
Previous work has shown that VLPFC neurons are preferentiallydriven by species-specific vocalizations compared to pure tones,noise bursts and other complex sounds (Romanski and Goldman-Rakic, 2002). In terms of vocalization coding, when non-humanprimates are tested in passive listening conditions without spe-cific discrimination tasks, single unit recordings from VLPFC neu-rons show similar responses to calls that are similar in acousticmorphology (Romanski et al., 2005). Examination of VLPFCneuronal responses using a small set of call type categories with anoddball-type task, has suggested that auditory cells in VLPFC mightencode semantic category information (Gifford et al., 2005),including the discrimination between vocalizations that indicatefood vs. non-food (Cohen et al., 2006).
Importantly, it has been shown that VLPFC cells are multisen-sory (Sugihara et al., 2006). These multisensory neurons are esti-mated to represent more than half of the population of theanterolateral VLPFC, including areas 12/47 and 45 (Romanski, 2012;Sugihara et al., 2006). Vocalization call type coding by cells maythen be more dependent upon an integration of not only acousticfeatures but also the appropriate facial gesture ormouthmovementthat accompanies the vocalization. Relying on responses to only theauditory component of a vocalization might then lead to less ac-curate recognition and categorization in regards to call type butalso especially to caller identity.
We asked how well VLPFC neurons encoded the vocalization calltype or caller identity of a species-specific vocalizations using previ-ously recorded data (Romanski et al., 2005) to 3 specific vocalizationcall types from three individual callers, (one coo, grunt, and girneyfrom each of 3 macaque monkeys). We then analyzed the neuralresponse using linear discriminate analysis to assess how accuratelycells classified the different call types (coos, grunts, and girneys).
1. Materials and methods
This analysis and review relies on neurophysiological recordingsof 110 cells from Romanski et al. (2005). Methods are described indetail in Romanski et al. (2005) with modifications to the dataanalysis described here.
1.1. Neurophysiological recordings
As previously described (Romanski et al., 2005) we madeextracellular recordings in two rhesus monkeys (Macaca mulatta)during the performance of a fixation task during which auditorystimuli including species-specific vocalizations were presented. All
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
methods were in accordance with NIH standards and wereapproved by the University of Rochester Care and Use of ResearchAnimals committee. Ventrolateral prefrontal areas 12/47 and 45were targeted (Preuss and Goldman-Rakic, 1991; Romanski andGoldman-Rakic, 2002; Petrides and Pandya, 2002).
1.2. Apparatus and stimuli
All training and recording was performed in a sound-attenuatedroom, lined with Sonex (Acoustical Solutions, Inc). Auditory stimuliwere presented to the monkeys by either a pair of Audix PH5-vsspeakers (frequency response �3 dB, 75e20,000 Hz) located oneither side of a center monitor, or a centrally located Yamaha MSP5monitor speaker (frequency response 50 Hz e 40 kHz), located 30inches from the monkey’s head. The auditory stimuli ranged from65 to 80 dB SPLmeasured at the level of themonkey’s ear with a B &K sound level meter, and a Realistic audio monitor.
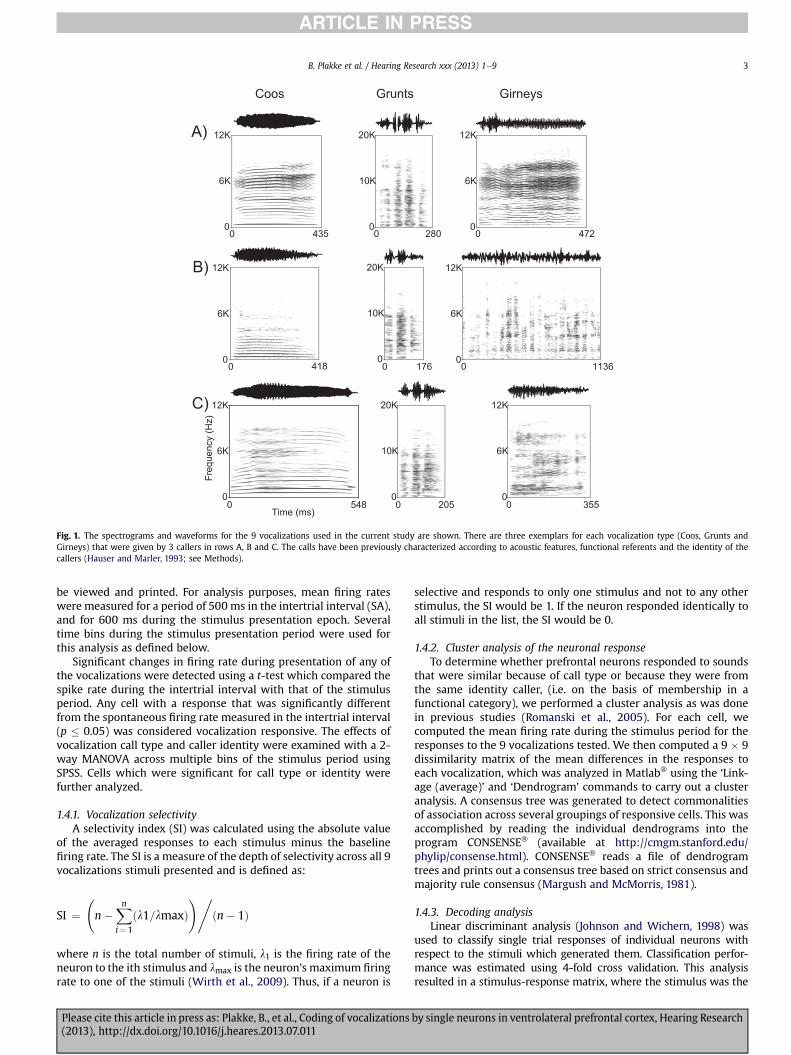
In this report we have focused on prefrontal responses to 3Macaque Vocalizations (Coo, Grunt and Girney) recorded on theisland of Cayo Santiago by Dr. Marc Hauser (Hauser and Marler,1993). The three vocalizations are all normally given during socialexchanges and were uttered by 3 adult female rhesus macaques onCayo Santiago. Thus, in this experiment 3 call types (CO, GT and GY)by three female callerswere used yielding a 3� 3 call matrix (Fig.1).Coos (CO) are given during social interactions including grooming,upon finding food of low value andwhen separated from the group.Grunts (GT) are given during social interaction such as an approachto groom, and upon the discovery of a low value food item andgirneys (GY) are given during grooming and when females attemptto handle infants. In addition to social context, vocalization types inthe macaque vocal repertoire have also been categorized accordingto the presence (or absence) of particular acoustic features (Fig. 1;Hauser and Marler, 1993; Hauser, 1996). Grunts are recognized asNoisy calls along with growls and pant threats while coos aremarked by the presence of a harmonic stack, oftenwith a dominantfundamental frequency and some evidence of vocal tract filtering.Girneys are considered tonal calls and have dominant energy at asingle or narrow band of frequencies. Brain regions which respondto species-specific vocalizations could do so on the basis of acoustic,behavioral context or emotional features of various calls.
1.3. Experimental procedure
Each day the monkeys were brought to the experimentalchamber and were prepared for extracellular recording as previ-ously published (Romanski et al., 2005). Each isolated unit wastested with a battery of auditory and visual stimuli which havebeen discussed in additional studies (Romanski et al., 2005;Sugihara et al., 2006). For this analysis cells were tested with the9 auditory stimuli (3 call types� 3 callers) which were repeated 8e12 times in a randomized design. During each trial a central fixationpoint appeared and subjects fixated for a 500 ms pretrial fixationperiod. Then the vocalization was presented and fixation wasmaintained. After the vocalization terminated a 500 ms post-stimulus fixation period occurred. A juice reward was delivered atthe termination of the post-stimulus fixation period and the fixa-tion requirement was then released. Losing fixation at any timeduring the task resulted in an aborted trial. There was a 2e3 s inter-trial interval, after which the fixation point would appear andsubjects could voluntarily begin a new trial.
1.4. Data analysis
The unit activity was read into Matlab� (Mathworks) and SPSSwhere rasters, histograms and spike density plots of the data could
by single neurons in ventrolateral prefrontal cortex, Hearing Research
Coos Grunts Girneys
A)
B)
C)
12K
6K
0
Freq
uenc
y (H
z)
12K
6K
0
12K
6K
0
12K
6K
0
12K
6K
0
12K
6K
0
20K
10K
0
20K
10K
0
20K
10K
00 548
Time (ms)0 0
0 0 0
0 0 0435 280 472
418 176 1136
205 355
Fig. 1. The spectrograms and waveforms for the 9 vocalizations used in the current study are shown. There are three exemplars for each vocalization type (Coos, Grunts andGirneys) that were given by 3 callers in rows A, B and C. The calls have been previously characterized according to acoustic features, functional referents and the identity of thecallers (Hauser and Marler, 1993; see Methods).
B. Plakke et al. / Hearing Research xxx (2013) 1e9 3
be viewed and printed. For analysis purposes, mean firing rateswere measured for a period of 500 ms in the intertrial interval (SA),and for 600 ms during the stimulus presentation epoch. Severaltime bins during the stimulus presentation period were used forthis analysis as defined below.
Significant changes in firing rate during presentation of any ofthe vocalizations were detected using a t-test which compared thespike rate during the intertrial interval with that of the stimulusperiod. Any cell with a response that was significantly differentfrom the spontaneous firing rate measured in the intertrial interval(p � 0.05) was considered vocalization responsive. The effects ofvocalization call type and caller identity were examined with a 2-way MANOVA across multiple bins of the stimulus period usingSPSS. Cells which were significant for call type or identity werefurther analyzed.
1.4.1. Vocalization selectivityA selectivity index (SI) was calculated using the absolute value
of the averaged responses to each stimulus minus the baselinefiring rate. The SI is a measure of the depth of selectivity across all 9vocalizations stimuli presented and is defined as:
SI ¼ n�
Xni¼1
ðl1=lmaxÞ!,
ðn� 1Þ
where n is the total number of stimuli, l1 is the firing rate of theneuron to the ith stimulus and lmax is the neuron’s maximum firingrate to one of the stimuli (Wirth et al., 2009). Thus, if a neuron is
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
selective and responds to only one stimulus and not to any otherstimulus, the SI would be 1. If the neuron responded identically toall stimuli in the list, the SI would be 0.
1.4.2. Cluster analysis of the neuronal responseTo determine whether prefrontal neurons responded to sounds
that were similar because of call type or because they were fromthe same identity caller, (i.e. on the basis of membership in afunctional category), we performed a cluster analysis as was donein previous studies (Romanski et al., 2005). For each cell, wecomputed the mean firing rate during the stimulus period for theresponses to the 9 vocalizations tested. We then computed a 9 � 9dissimilarity matrix of the mean differences in the responses toeach vocalization, which was analyzed in Matlab� using the ‘Link-age (average)’ and ‘Dendrogram’ commands to carry out a clusteranalysis. A consensus tree was generated to detect commonalitiesof association across several groupings of responsive cells. This wasaccomplished by reading the individual dendrograms into theprogram CONSENSE� (available at http://cmgm.stanford.edu/phylip/consense.html). CONSENSE� reads a file of dendrogramtrees and prints out a consensus tree based on strict consensus andmajority rule consensus (Margush and McMorris, 1981).
1.4.3. Decoding analysisLinear discriminant analysis (Johnson and Wichern, 1998) was
used to classify single trial responses of individual neurons withrespect to the stimuli which generated them. Classification perfor-mance was estimated using 4-fold cross validation. This analysisresulted in a stimulus-response matrix, where the stimulus was the
by single neurons in ventrolateral prefrontal cortex, Hearing Research
B. Plakke et al. / Hearing Research xxx (2013) 1e94
vocalization which was presented on an individual trial, and theresponse was the vocalization to which each single trial neuralresponsewas classified. Each cell of thematrix contained the count ofthe number of times that a particular vocalization (the stimulus) wasclassified as a particular response by the algorithm. Thus, the diagonalelementsof thismatrixcontainedcountsof correct classifications, andthe off-diagonal elements of the matrix contained counts of theincorrect classifications. Percent correct performance for each stim-ulus classwascalculatedbydividing thenumberof correctlyclassifiedtrials for a particular stimulus (the diagonal element of a particularrow)by the total numberof times a particular stimuluswaspresented(usually 8e12, the sum of the off-diagonal elements in a particularrow).
2. Results
We analyzed neurons recorded in macaque VLPFC with species-specific vocalizations grouped by call type and caller identity. 123cells (110 from previous Romanski et al. (2005) plus 13 additionalfrom the same animals) were tested with a balanced list of 3vocalization call types vocalized by 3 female callers (one coo, grunt,and girney from each caller). Recorded neurons, from awakebehaving macaques, were analyzed off line to first determine theresponse to the 9 vocalization stimuli. 81/123 cells were defined asvocalization responsive since the firing rate during early and lateportions of the response period were significantly different frombaseline with a t-test (p < 0.5), for any of the 9 vocalizations.
0
40
20
0
SI= 0.74034
SI= 0.52327B)
A)
Coo 2Coo
3
Grunt 3
Grunt 2
Girney
1
Girney
2
Girney
3Coo
1
Grunt 1
Coo 2Coo
3
Grunt 3
Grunt 2
Girney
1
Girney
2
Girney
3Coo
1
Grunt 1
Spik
es/s
ecSp
ikes
/sec
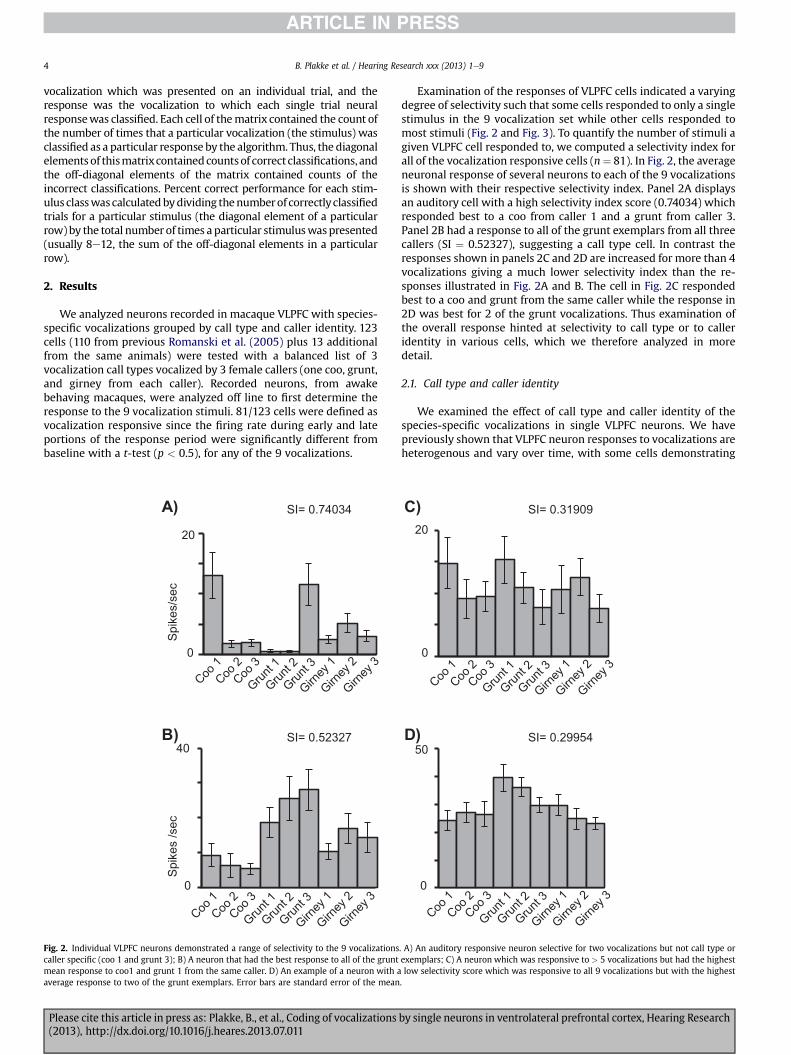
Fig. 2. Individual VLPFC neurons demonstrated a range of selectivity to the 9 vocalizationscaller specific (coo 1 and grunt 3); B) A neuron that had the best response to all of the gruntmean response to coo1 and grunt 1 from the same caller. D) An example of a neuron with aaverage response to two of the grunt exemplars. Error bars are standard error of the mean
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
Examination of the responses of VLPFC cells indicated a varyingdegree of selectivity such that some cells responded to only a singlestimulus in the 9 vocalization set while other cells responded tomost stimuli (Fig. 2 and Fig. 3). To quantify the number of stimuli agiven VLPFC cell responded to, we computed a selectivity index forall of the vocalization responsive cells (n¼ 81). In Fig. 2, the averageneuronal response of several neurons to each of the 9 vocalizationsis shown with their respective selectivity index. Panel 2A displaysan auditory cell with a high selectivity index score (0.74034) whichresponded best to a coo from caller 1 and a grunt from caller 3.Panel 2B had a response to all of the grunt exemplars from all threecallers (SI ¼ 0.52327), suggesting a call type cell. In contrast theresponses shown in panels 2C and 2D are increased for more than 4vocalizations giving a much lower selectivity index than the re-sponses illustrated in Fig. 2A and B. The cell in Fig. 2C respondedbest to a coo and grunt from the same caller while the response in2D was best for 2 of the grunt vocalizations. Thus examination ofthe overall response hinted at selectivity to call type or to calleridentity in various cells, which we therefore analyzed in moredetail.
2.1. Call type and caller identity
We examined the effect of call type and caller identity of thespecies-specific vocalizations in single VLPFC neurons. We havepreviously shown that VLPFC neuron responses to vocalizations areheterogenous and vary over time, with some cells demonstrating
D)
Coo 2Coo
3
Grunt 3
Grunt 2
Girney
1
Girney
2
Girney
3Coo
1
Grunt 1
50
20
0
0
SI= 0.31909C)
SI= 0.29954
Coo 2Coo
3
Grunt 3
Grunt 2
Girney
1
Girney
2
Girney
3Coo
1
Grunt 1
. A) An auditory responsive neuron selective for two vocalizations but not call type orexemplars; C) A neuron which was responsive to > 5 vocalizations but had the highestlow selectivity score which was responsive to all 9 vocalizations but with the highest.
by single neurons in ventrolateral prefrontal cortex, Hearing Research
Coos Grunts Girneys
120
800 8000 08000
A)
B)
C)0
-250
Coo 1 Coo 3 Grunt 1 Grunt 2
90
0800 8000 08000-250 8000
30
0
80008000
Grunt 3Coo 2
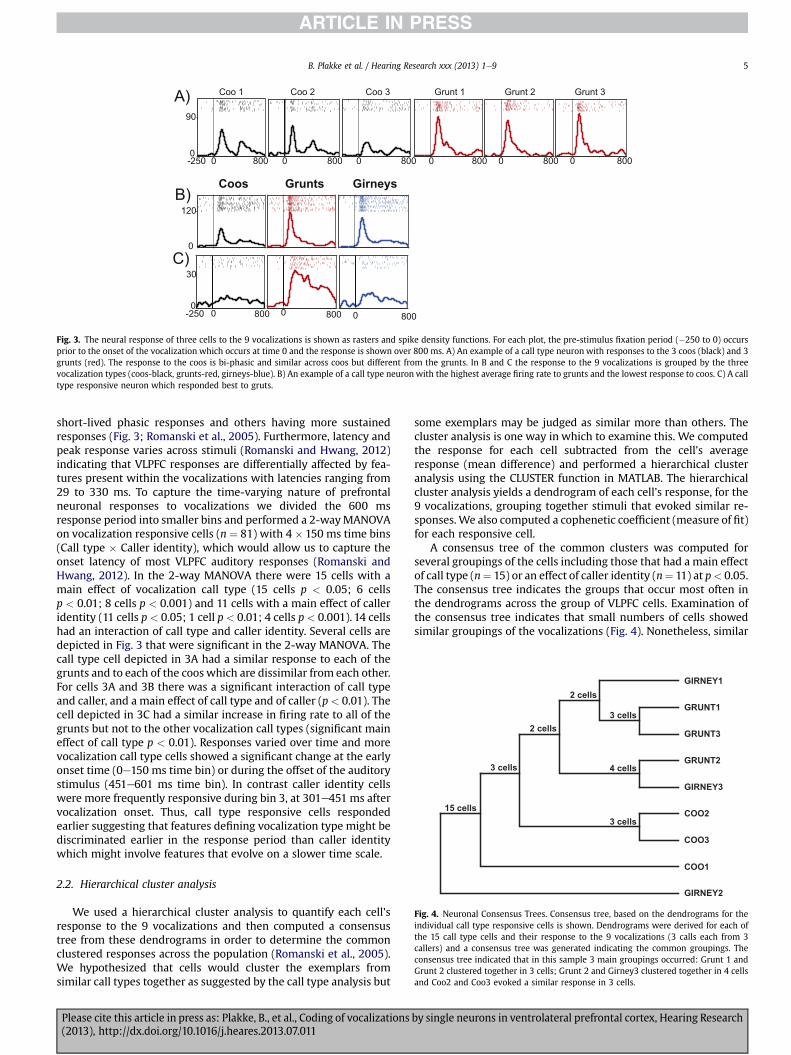
Fig. 3. The neural response of three cells to the 9 vocalizations is shown as rasters and spike density functions. For each plot, the pre-stimulus fixation period (�250 to 0) occursprior to the onset of the vocalization which occurs at time 0 and the response is shown over 800 ms. A) An example of a call type neuron with responses to the 3 coos (black) and 3grunts (red). The response to the coos is bi-phasic and similar across coos but different from the grunts. In B and C the response to the 9 vocalizations is grouped by the threevocalization types (coos-black, grunts-red, girneys-blue). B) An example of a call type neuron with the highest average firing rate to grunts and the lowest response to coos. C) A calltype responsive neuron which responded best to gruts.
GIRNEY1
GIRNEY3
GIRNEY2
COO1
GRUNT1
COO3
COO2
GRUNT2
GRUNT3
15 cells
3 cells
2 cells
2 cells
3 cells
3 cells
4 cells
Fig. 4. Neuronal Consensus Trees. Consensus tree, based on the dendrograms for theindividual call type responsive cells is shown. Dendrograms were derived for each ofthe 15 call type cells and their response to the 9 vocalizations (3 calls each from 3callers) and a consensus tree was generated indicating the common groupings. Theconsensus tree indicated that in this sample 3 main groupings occurred: Grunt 1 andGrunt 2 clustered together in 3 cells; Grunt 2 and Girney3 clustered together in 4 cellsand Coo2 and Coo3 evoked a similar response in 3 cells.
B. Plakke et al. / Hearing Research xxx (2013) 1e9 5
short-lived phasic responses and others having more sustainedresponses (Fig. 3; Romanski et al., 2005). Furthermore, latency andpeak response varies across stimuli (Romanski and Hwang, 2012)indicating that VLPFC responses are differentially affected by fea-tures present within the vocalizations with latencies ranging from29 to 330 ms. To capture the time-varying nature of prefrontalneuronal responses to vocalizations we divided the 600 msresponse period into smaller bins and performed a 2-wayMANOVAon vocalization responsive cells (n ¼ 81) with 4 � 150 ms time bins(Call type � Caller identity), which would allow us to capture theonset latency of most VLPFC auditory responses (Romanski andHwang, 2012). In the 2-way MANOVA there were 15 cells with amain effect of vocalization call type (15 cells p < 0.05; 6 cellsp < 0.01; 8 cells p < 0.001) and 11 cells with a main effect of calleridentity (11 cells p < 0.05; 1 cell p < 0.01; 4 cells p< 0.001). 14 cellshad an interaction of call type and caller identity. Several cells aredepicted in Fig. 3 that were significant in the 2-way MANOVA. Thecall type cell depicted in 3A had a similar response to each of thegrunts and to each of the coos which are dissimilar from each other.For cells 3A and 3B there was a significant interaction of call typeand caller, and a main effect of call type and of caller (p < 0.01). Thecell depicted in 3C had a similar increase in firing rate to all of thegrunts but not to the other vocalization call types (significant maineffect of call type p < 0.01). Responses varied over time and morevocalization call type cells showed a significant change at the earlyonset time (0e150 ms time bin) or during the offset of the auditorystimulus (451e601 ms time bin). In contrast caller identity cellswere more frequently responsive during bin 3, at 301e451 ms aftervocalization onset. Thus, call type responsive cells respondedearlier suggesting that features defining vocalization type might bediscriminated earlier in the response period than caller identitywhich might involve features that evolve on a slower time scale.
2.2. Hierarchical cluster analysis
We used a hierarchical cluster analysis to quantify each cell’sresponse to the 9 vocalizations and then computed a consensustree from these dendrograms in order to determine the commonclustered responses across the population (Romanski et al., 2005).We hypothesized that cells would cluster the exemplars fromsimilar call types together as suggested by the call type analysis but
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
some exemplars may be judged as similar more than others. Thecluster analysis is one way in which to examine this. We computedthe response for each cell subtracted from the cell’s averageresponse (mean difference) and performed a hierarchical clusteranalysis using the CLUSTER function in MATLAB. The hierarchicalcluster analysis yields a dendrogram of each cell’s response, for the9 vocalizations, grouping together stimuli that evoked similar re-sponses.We also computed a cophenetic coefficient (measure of fit)for each responsive cell.
A consensus tree of the common clusters was computed forseveral groupings of the cells including those that had a main effectof call type (n¼ 15) or an effect of caller identity (n¼ 11) at p< 0.05.The consensus tree indicates the groups that occur most often inthe dendrograms across the group of VLPFC cells. Examination ofthe consensus tree indicates that small numbers of cells showedsimilar groupings of the vocalizations (Fig. 4). Nonetheless, similar
by single neurons in ventrolateral prefrontal cortex, Hearing Research
CoosGruntsGirneys
100
50
Per
cent
Cor
rect
B. Plakke et al. / Hearing Research xxx (2013) 1e96
call types were not grouped together by a high percentage of cellseven in this small call type responsive group. From the consensustree it is apparent that coos were grouped together byw20% of celltype responsive cells. 2 of the grunt exemplars were also clusteredby 20% of the cells (Fig. 4; Grunt 1 and grunt 3). There were 4 cells(27%) that responded in a similar manner to a grunt from speaker 2and girney from speaker 3. Overall, there were a small number ofcells which showed grouping by a similar call type. Cluster analysisand consensus across the significant caller cells resulted in only afew cells showing a grouping by caller (not shown).
0150 300 450 600Time (ms)
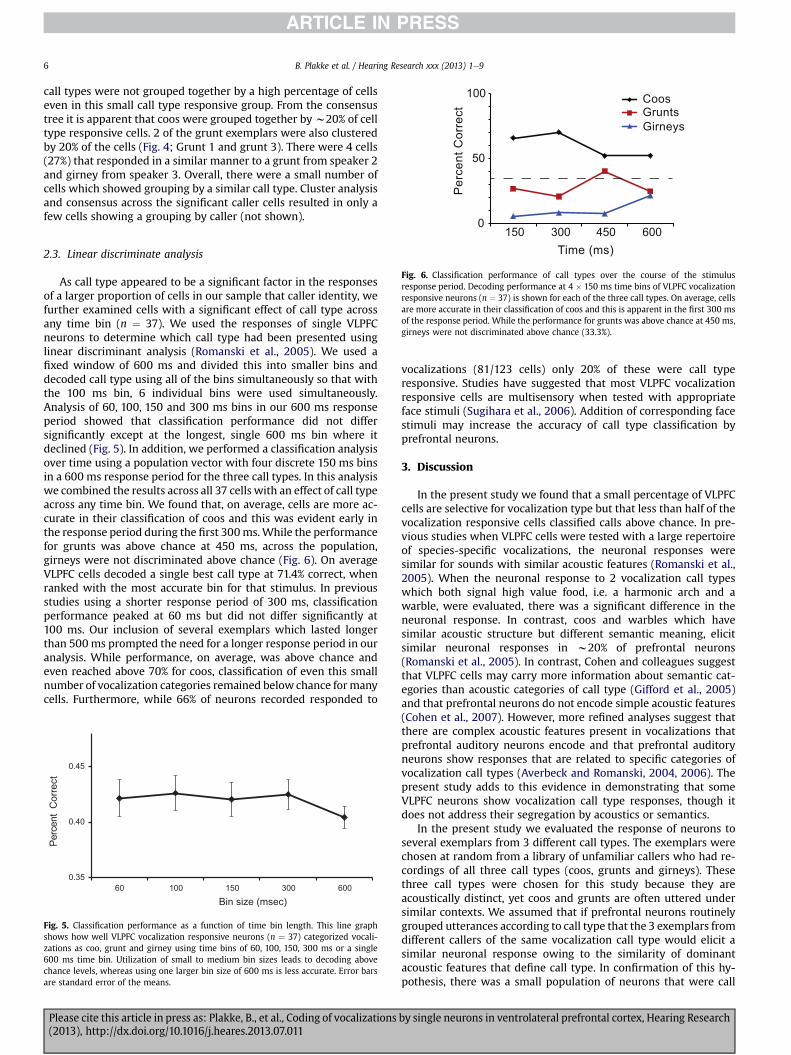
Fig. 6. Classification performance of call types over the course of the stimulusresponse period. Decoding performance at 4 � 150 ms time bins of VLPFC vocalizationresponsive neurons (n ¼ 37) is shown for each of the three call types. On average, cellsare more accurate in their classification of coos and this is apparent in the first 300 msof the response period. While the performance for grunts was above chance at 450 ms,girneys were not discriminated above chance (33.3%).
2.3. Linear discriminate analysis
As call type appeared to be a significant factor in the responsesof a larger proportion of cells in our sample that caller identity, wefurther examined cells with a significant effect of call type acrossany time bin (n ¼ 37). We used the responses of single VLPFCneurons to determine which call type had been presented usinglinear discriminant analysis (Romanski et al., 2005). We used afixed window of 600 ms and divided this into smaller bins anddecoded call type using all of the bins simultaneously so that withthe 100 ms bin, 6 individual bins were used simultaneously.Analysis of 60, 100, 150 and 300 ms bins in our 600 ms responseperiod showed that classification performance did not differsignificantly except at the longest, single 600 ms bin where itdeclined (Fig. 5). In addition, we performed a classification analysisover time using a population vector with four discrete 150 ms binsin a 600 ms response period for the three call types. In this analysiswe combined the results across all 37 cells with an effect of call typeacross any time bin. We found that, on average, cells are more ac-curate in their classification of coos and this was evident early inthe response period during the first 300ms.While the performancefor grunts was above chance at 450 ms, across the population,girneys were not discriminated above chance (Fig. 6). On averageVLPFC cells decoded a single best call type at 71.4% correct, whenranked with the most accurate bin for that stimulus. In previousstudies using a shorter response period of 300 ms, classificationperformance peaked at 60 ms but did not differ significantly at100 ms. Our inclusion of several exemplars which lasted longerthan 500ms prompted the need for a longer response period in ouranalysis. While performance, on average, was above chance andeven reached above 70% for coos, classification of even this smallnumber of vocalization categories remained below chance formanycells. Furthermore, while 66% of neurons recorded responded to
0.35
0.40
0.45
60 100 150 300 600
Bin size (msec)
Perc
ent
Cor
rect
Fig. 5. Classification performance as a function of time bin length. This line graphshows how well VLPFC vocalization responsive neurons (n ¼ 37) categorized vocali-zations as coo, grunt and girney using time bins of 60, 100, 150, 300 ms or a single600 ms time bin. Utilization of small to medium bin sizes leads to decoding abovechance levels, whereas using one larger bin size of 600 ms is less accurate. Error barsare standard error of the means.
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
vocalizations (81/123 cells) only 20% of these were call typeresponsive. Studies have suggested that most VLPFC vocalizationresponsive cells are multisensory when tested with appropriateface stimuli (Sugihara et al., 2006). Addition of corresponding facestimuli may increase the accuracy of call type classification byprefrontal neurons.
3. Discussion
In the present study we found that a small percentage of VLPFCcells are selective for vocalization type but that less than half of thevocalization responsive cells classified calls above chance. In pre-vious studies when VLPFC cells were tested with a large repertoireof species-specific vocalizations, the neuronal responses weresimilar for sounds with similar acoustic features (Romanski et al.,2005). When the neuronal response to 2 vocalization call typeswhich both signal high value food, i.e. a harmonic arch and awarble, were evaluated, there was a significant difference in theneuronal response. In contrast, coos and warbles which havesimilar acoustic structure but different semantic meaning, elicitsimilar neuronal responses in w20% of prefrontal neurons(Romanski et al., 2005). In contrast, Cohen and colleagues suggestthat VLPFC cells may carry more information about semantic cat-egories than acoustic categories of call type (Gifford et al., 2005)and that prefrontal neurons do not encode simple acoustic features(Cohen et al., 2007). However, more refined analyses suggest thatthere are complex acoustic features present in vocalizations thatprefrontal auditory neurons encode and that prefrontal auditoryneurons show responses that are related to specific categories ofvocalization call types (Averbeck and Romanski, 2004, 2006). Thepresent study adds to this evidence in demonstrating that someVLPFC neurons show vocalization call type responses, though itdoes not address their segregation by acoustics or semantics.
In the present study we evaluated the response of neurons toseveral exemplars from 3 different call types. The exemplars werechosen at random from a library of unfamiliar callers who had re-cordings of all three call types (coos, grunts and girneys). Thesethree call types were chosen for this study because they areacoustically distinct, yet coos and grunts are often uttered undersimilar contexts. We assumed that if prefrontal neurons routinelygrouped utterances according to call type that the 3 exemplars fromdifferent callers of the same vocalization call type would elicit asimilar neuronal response owing to the similarity of dominantacoustic features that define call type. In confirmation of this hy-pothesis, there was a small population of neurons that were call
by single neurons in ventrolateral prefrontal cortex, Hearing Research

B. Plakke et al. / Hearing Research xxx (2013) 1e9 7
type responsive in our analysis and clearly showed a similarresponse across exemplars from different callers to each call type(Figs. 2c and 3a,b). However, in our sample of 81 cells, w20% had amain effect of vocalization call type, 14% with an effect of caller and17% cells had an interaction of call type and caller. Thus the percentof cells that appear to encode vocalization call type is somewhatsmall and those encoding the identity of the caller that issued thecall are even smaller, considering the robust response of theseneurons to vocalizations.
Cells which were responsive to the vocalizations but did nothave an effect of call type or caller often were selective andresponded to 1e3 vocalizations (Fig. 2). VLPFC cells have beenpreviously shown to be selective in their responses not only tovocalizations (Romanski and Goldman-Rakic, 2002; Romanskiet al., 2005) but also to face stimuli (O’Scalaidhe et al., 1997, 1999). Itis not clear whether the selectivity of VLPFC neurons is based onsome combination of acoustic features in a particular vocalizationwhich evoke a response or some other aspect of the sound such asits meaning or association (Cohen et al., 2006). However, therewere no cells which responded with a similar response to all cooand grunt vocalizations which represent a similar functional cate-gory but which are acoustically dissimilar. Responses across theVLPFC population are heterogenous and correlation of the pre-frontal neuronal response with complex features of the vocaliza-tions may be necessary to provide evidence of vocalization call typecoding (Averbeck and Romanski, 2006).
We further examined vocalization call type classification usingthe responses of single VLPFC neurons on individual trials usinglinear discriminant analysis. Analysis across different bins in the600ms response period showed that classification performance didnot differ significantly from 60 to 300 ms (Fig. 6). Examination ofthe temporal aspects of call type categorization across the popu-lation yielded interesting results. We found that, on average cellswere more accurate in their classification of coos and this wasevident early in the response period during the first 300 m. Clas-sification performance of coos reached 70 percent in the first300 ms and remained above chance for the duration of theresponse period. In previous studies using a shorter responseperiod of 300 ms, classification performance peaked at 60 ms butdid not differ significantly at 100 ms (Averbeck and Romanski,2006). Our inclusion of several exemplars which lasted longerthan 500ms prompted our need for a longer response period in ouranalysis. Interestingly, Rendall et al. (1998) examined coos, grunts,and screams for spectral features and using a discriminate analysisfound that coos were discriminated more accurately than grunts orscreams (Rendall et al., 1998). In addition, it was found that cooshad a spectral peak which was stable and distinctive (Rendall et al.,1998). Ceugniet and Izumi (2004) demonstrated that macaquescould discriminate coos from different individuals and that callduration, pitch, and harmonics were used to make discriminationsbetween calls. Thus, it is possible that these features (call duration,pitch, harmonics, and spectral cues) led to the greater accuracy ofclassification of coos found here for VLPFC neurons. In fact, evenhumans are better at discriminating coos from different individualmonkeys than screams from those same individuals (Owren andRendall, 2003). If coos themselves have more cues that makediscrimination easier, then this information may be parsed out inearlier auditory regions (belt, parabelt, STG) and routed to VLPFC.Perhaps having multiple coos from the same individual would haveincreased our discrimination accuracy for identity, or increased thenumber of identity responsive cells.
The overall classification performance of VLPFC neurons in ouranalysis of a small number of vocalization categories was w42%(Fig. 5). There are several possibilities as to why the percent correctwas not higher. First, our data is based only on a small population of
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
neurons recorded singly which may not be adequate to support thecomplex function of vocalization call type processing. Classificationacross simultaneously recorded cells would most likely yield su-perior accuracy (Georgopoulos and Massey, 1988; Averbeck et al.,2003). Second, studies have shown higher classification perfor-mance when assessing responses to categories with clear bound-aries such as vocalization and non-vocalization stimuli. Forexample, population decoding in early auditory processing area R(rostral field), found classification accuracy of natural sounds(monkey calls and environmental sounds) to exceed 90% correct80 msec after stimulus onset (Kusmierek et al., 2012). Finally, oursample of neurons included cells from both areas 12/47 and 45which may be specialized for different functions with regard tovocalizations. Area 12/47 receives more input from rostral belt andparabelt regions while area 45 receives a denser projection fromcaudal parabelt, the STS and from inferotemporal cortex (Romanskiet al., 1999; Romanski and Goldman-Rakic, 2002; Barbas, 1988;Webster et al., 1994). In addition, area 12/47 has more unimodalauditory responsive cells. Both regions receive input from the STGand STS and have multisensory responsive neurons. Thus, vocali-zations may be processed differently in the more auditoryresponsive area of 12/47 compared to area 45, which may be morespecialized for face and object processing (Romanski, 2012, 2007).Focusing our analysis on a larger sample of cells which have simi-larly tuned responses to specific complex features may increase thecategorization performance of cells in VLPFC.
Possibly the most important reason for the modest classificationperformance of VLPFC cells to vocalization stimuli is the fact thatmost of these cells are multisensory and may typically processvocalizations in a multisensory manner. In the current study wehave examined responses only to vocalizations which may be onlyhalf of the stimulus that elicits the best response for many pre-frontal neurons. In previous work where VLPFC neurons weretested with separate and simultaneous combinations of vocaliza-tions and the accompanying dynamic facial gesture it has beenshown that more than half the recorded population was multi-sensory (Sugihara et al., 2006). VLPFC cells show multisensoryenhancement or suppression to face-vocalization combinations.Moreover, neuronal responses demonstrate more information forcombined face-vocalization stimuli than for the unimodal compo-nents alone (Sugihara et al., 2006). These strong responses to face-vocalization pairs suggest that this region of VLPFC may bespecialized for face-vocalization integration (Romanski, 2012). It islikely that VLPFC neurons would show significantly enhancedclassification of vocalization call types when both face and vocali-zation information is present. Even neural responses in auditorycortex are enhanced when appropriate visual information is pre-sent (Kayser et al., 2010). Importantly, it has been shown thatdiscrimination of vocalizations improves when a congruent facestimulus accompanies the vocalization (Chandrasekaran et al.,2011). In this study, behavioral performance of rhesus macaquesdiscriminating vocalizations improved when congruent avatar facestimuli accompanied the vocalizations.
It is well known that combining auditory and visual stimuli canenhance accuracy and decrease reaction time (Stein and Meredith,1993). This has been demonstrated in many brain regions includingthe inferior frontal gyrus in the human brain during neuroimagingwhere the PFC accumulates information about auditory and visualstimuli to be used in recognition (Noppeney et al., 2010). Moreover,training of faceevoice pairs can enhance activity in VLPFC for voicerecognition (von Kriegstein and Giraud, 2006) and the interactionbetween face and voice processing involves connections betweenface areas, voice areas, and VLPFC (von Kriegstein and Giraud, 2006).
Voice recognition in the human brain is also a multisensoryprocess and includes voice regions found in the superior temporal
by single neurons in ventrolateral prefrontal cortex, Hearing Research

B. Plakke et al. / Hearing Research xxx (2013) 1e98
sulcus STS (Belin et al., 2000) and VLPFC (Fecteau et al., 2005). In anfMRI study with face and voice recognition, Brodmann’s areas 45 and47 were activated during the delay period in these working memorytasks (Rama and Courtney, 2005). Our current data showing vocali-zation identity cells in VLPFC, which included areas 45 and 47, couldprovide the underlying neuronal mechanism for these effects.
Data from behavioral studies in animals and humans shows thatrecognition and memory are enhanced when both face and vocalinformation are present. In fact, use of auditory information aloneresults in inferior performance on memory tasks compared toperformance with visual stimuli alone (Fritz et al., 2005). Trainingnon-human primates to perform auditory working memory (WM)paradigms typically takes longer than for visual WM tasks and ithas been shown that performance accuracy is greater for visualWM than for auditory WM tasks in non-human primates (Scottet al., 2012). Surprisingly, despite our superior language abilities,human subjects also perform better when given faces than whengiven voices as memoranda (Yarmey et al., 1994; Hanley et al.,1998). It has been demonstrated that recognition of speaker iden-tity from a single utterance only resulted in 35% accuracy, withfemales outperforming males (Skuk, and Schweinberger, 2013).Even when face and voice stimuli are familiar and the content andfrequency of presentation is carefully controlled in a similarmanner as our animal experiments, faces are more easily remem-bered and more semantic information is retrieved from humanfaces compared to their voices (Barsics and Brédart, 2012). Thus, thesub-optimal performance of our population of VLPFC responsiveneurons in categorizing vocalization call types is perhaps expectedwhen only vocal information is available.
Finally, our present study relies on data collected during stim-ulus presentation and it has been shown that prefrontal neuronsare most active during working memory, categorization and goaldirected behavior (Freedman et al., 2001; Russ et al., 2007). Inhumans it has been found that task demands can activate differentfrontal regions (Owen et al., 1996) and that modality and task de-mands can interact within VLPFC (Protzner and McIntosh, 2009). Inour lab, context during a non-match-to-sample task has beenshown to modulate firing rates, with some cells showing an effectof task conditions and others maintaining a preference for aparticular stimulus (Hwang and Romanski, 2009). An analysis ofneuronal responses when face and vocal information are activelyused in a recognition paradigm and carefully compared with per-formance using only auditory or only visual information is neededto fully understand the processing of communication stimuli andthe impact of face and vocalization integration on recognition.
Acknowledgments
This research was funded by the National Institute for Deafnessand Communication Disorders (DC 04845, LMR and DC05409,Center for Navigation and Communication Sciences).
References
Averbeck, B.B., Crowe, D.A., Chafee, M.V., Georgopoulos, A.P., 2003. Neural activity inprefrontal cortex during copying geometrical shapes. II. Decoding shape seg-ments from neural ensembles. Exp. Brain Res. 150 (2), 142e153.
Averbeck, B.B., Romanski, L.M., 2006. Probabilistic encoding of vocalizations inmacaque ventral lateral prefrontal cortex. J. Neurosci. 26, 11023e11033.
Averbeck, B.B., Romanski, L.M., 2004. Principal and independent components ofmacaque vocalizations: constructing stimuli to probe high-level sensory pro-cessing. J. Neurophysiol. 91, 2897e2909.
Barbas, H., 1988. Anatomic organization of basoventral and mediodorsal visualrecipient prefrontal regions in the rhesus monkey. J. Comp. Neurol. 276, 313e342.
Barsics, C., Bredart, S., 2012. Recalling semantic information about newly learnedfaces and voices. Memory 20 (5), 527e534.
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
Belin, P., Zatorre, R.J., Lafaille, P., Ahad, P., Pike, B., 2000. Voice-selective areas inhuman auditory cortex. Nature 403, 309e312.
Bendor, D., Osmanski, M.S., Wang, X., 2012. Dual-pitch Processing Mechanisms inPrimate Auditory Cortex, vol. 32, pp. 16149e16161.
Bendor, D., Wang, X., 2005. The neuronal representation of pitch in primate audi-tory cortex. Nature 436 (7054), 1161e1165.
Bradbury, J.W., Vehrencamp, S.L., 1998. Principles of Animal Communication.Blackwell, Oxford.
Ceugniet, M., Izumi, A., 2004. Vocal Individual Discrimination in Japanese Monkeys,vol. 45, pp. 119e128.
Chandrasekaran, C., Lemus, L., Trubanova, A., Gondan, M., Ghazanfar, A.A., 2011.Monkeys and Humans Share a Common Computation for Face/voice Integra-tion, vol. 7, p. e1002165.
Cohen, Y.E., Hauser, M.D., Russ, B.E., 2006. Spontaneous processing of abstractcategorical information in the ventrolateral prefrontal cortex. Biol. Lett. 2 (2),261e265.
Cohen, Y.E., Theunissen, F., Russ, B.E., Gill, P., 2007. Acoustic features of rhesus vo-calizations and their representation in the ventrolateral prefrontal cortex.J. Neurophysiol. 97 (2), 1470e1484.
Fecteau, S., Armony, J.L., Joanette, Y., Belin, P., 2005. Sensitivity to voice in humanprefrontal cortex. J. Neurophysiol. 94, 2251e2254.
Freedman, D.J., Riesenhuber, M., Poggio, T., Miller, E.K., 2001. Categorical represen-tation of visual stimuli in the primate prefrontal cortex. Science 291, 312e316.
Fritz, J., Mishkin, M., Saunders, R.C., 2005. In search of an auditory engram. Proc.Natl. Acad. Sci. U. S. A 102 (26), 9359e9364.
Georgopoulos, A.P., Massey, J.T., 1988. Cognitive spatial-motor processes. 2. Infor-mation transmitted by the direction of two-dimensional arm movements andby neuronal populations in primate motor cortex and area 5. Exp. Brain Res. 69(2), 315e326.
Gifford III, G.W., Maclean, K.A., Hauser, M.D., Cohen, Y.E., 2005. The neurophysiologyof functionally meaningful categories: macaque ventrolateral prefrontal cortexplays a critical role in spontaneous categorization of species-specific vocaliza-tions. J. Cogn. Neurosci. 17, 1471e1482.
Hackett, T.A., Stepniewska, I., Kaas, J.H., 1999. Prefrontal connections of the parabeltauditory cortex in macaque monkeys. Brain Res. 817, 45e58.
Hanley, J.R., Smith, S.T., Hadfield, J., 1998. I recognise you but I can’t place you: aninvestigation of familiar-only experiences during tests of voice and facerecognition. Q. J. Exp. Psychol. 51 (1), 179e195.
Hauser, M.D., 1996. The Evolution of Communication. MIT Press, Cambridge, MA.Hauser, M.D., Marler, P., 1993. Food associated calls in rhesus macaques (Macaca
mulatta) I. Socioecological factors. Behav. Ecol. 4, 194e205.Hwang, J., Romanski, L.M., 2009. Comparison of Face and Non-face Stimuli in an
Audio-visual Discrimination Task, 578.5.Johnson, R.A., Wichern, D.W., 1998. Applied Multivariate Statistical Analysis. Pren-
tice Hall, Saddle River, NJ.Joly, O., Ramus, F., Pressnitzer, D., Vanduffel, W., Orban, G.A., 2012. Interhemispheric
differences in auditory processing revealed by fMRI in awake rhesus monkeys.Cereb. Cortex 22 (4), 838e853.
Kayser, C., Logothetis, N.K., Panzeri, S., 2010. Visual enhancement of the informationrepresentation in auditory cortex. Curr. Biol. 20 (1), 19e24.
Kikuchi, Y., Horwitz, B., Mishkin, M., 2010. Hierarchical auditory processing directedrostrally along the monkey’s supratemporal plane. J. Neurosci. 30 (39), 13021e13030.
Kusmierek, P., Ortiz, M., Rauschecker, J.P., 2012. Sound-identity Processing in EarlyAreas of the Auditory Ventral Stream in the Macaque, vol. 107, pp. 1123e1141.
Margush, T., McMorris, F.R., 1981. Consensus n-trees. Bull. Math. Biol. 43, 239e244.Noppeney, U., Ostwald, D., Werner, S., 2010. Perceptual decisions formed by accu-
mulation of audiovisual evidence in prefrontal cortex. J. Neurosci. 30 (21),7434e7446.
O’Scalaidhe, S.P., Wilson, F.A., Goldman-Rakic, P.S., 1997. Areal segregation of face-processing neurons in prefrontal cortex. Science 278, 1135e1138.
O’Scalaidhe, S.P.O., Wilson, F.A.W., Goldman-Rakic, P.G.R., 1999. Face-selectiveneurons during passive viewing and working memory performance of rhesusmonkeys: evidence for intrinsic specialization of neuronal coding. Cereb. Cortex9, 459e475.
Owen, A.M., Evans, A.C., Petrides, M., 1996. Evidence for a two-stage model ofspatial working memory processing with the lateral frontal cortex: a positronemission tomography study. Cereb. Cortex 6, 31e38.
Owings, D.H., Morton, E.S., 1998. Animal Vocal Communication: a New Approach.Cambridge University Press, Cambridge.
Owren, M.J., Rendall, D., 2003. Salience of caller identity in rhesus monkey (Macacamulatta) coos and screams: perceptual experiments with human (Homo sapi-ens) listeners. J. Comp. Psychol. 117 (4), 380e390.
Perrodin, C., Kayser, C., Logothetis, N.K., Petkov, C.I., 2011. Voice cells in the primatetemporal lobe. Curr. Biol. 21 (16), 1408e1415.
Petkov, C.I., Kayser, C., Steudel, T., Whittingstall, K., Augath, M., Logothetis, N.K.,2008. A voice region in the monkey brain. Nat. Neurosci. 11 (3), 367e374.
Petrides, M., Pandya, D.N., 2002. Comparative cytoarchitectonic analysis of thehuman and the macaque ventrolateral prefrontal cortex and corticocorticalconnection patterns in the monkey. Eur. J. Neurosci. 16 (2), 291e310.
Poremba, A., Malloy, M., Saunders, R.C., Carson, R.E., Herscovitch, P., Mishkin, M.,2004. Species-specific calls evoke asymmetric activity in the monkey’s tem-poral poles. Nature 427 (6973), 448e451.
Poremba, A., Saunders, R.C., Crane, A.M., Cook, M., Sokoloff, L., Mishkin, M., 2003.Functional mapping of the primate auditory system. Science 299, 568e572.
by single neurons in ventrolateral prefrontal cortex, Hearing Research

B. Plakke et al. / Hearing Research xxx (2013) 1e9 9
Preuss, T.M., Goldman-Rakic, P.S., 1991. Ipsilateral cortical connections of granularfrontal cortex in the strepsirhine primate Galago, with comparative commentson anthropoid primates. J. Comp. Neurol. 310, 507e549.
Protzner, A.B., McIntosh, A.R., 2009. Modulation of Ventral Prefrontal CortexFunctional Connections Reflects the Interplay of Cognitive Processes andStimulus Characteristics, vol. 19, pp. 1042e1054.
Rama, P., Courtney, S.M., 2005. Functional topography of working memory for faceor voice identity. Neuroimage 24 (1), 224e234.
Rauschecker, J.P., 1998. Cortical processing of complex sounds. Curr. Opin. Neuro-biol. 8 (4), 516e521.
Rauschecker, J.P., Tian, B., Hauser, M., 1995. Processing of complex sounds in themacaque nonprimary auditory cortex. Science 268 (5207), 111e114.
Rauschecker, J.P., Tian, B., Pons, T., Mishkin, M., 1997. Serial and parallel processingin rhesus monkey auditory cortex. J. Comp. Neurol. 382 (1), 89e103.
Rendall, D., Owren, M.J., Rodman, P.S., 1998. The role of vocal tract filtering inidentity cueing in rhesus monkey (Macaca mulatta) vocalizations. J. Acoust. Soc.Am. 103 (1), 602e614.
Romanski, L.M., 2012. Integration of faces and vocalizations in ventral prefrontalcortex: implications for the evolution of audiovisual speech. Proc. Natl. Acad.Sci. U. S. A 109 (Suppl. 1), 10717e10724.
Romanski, L.M., 2007. Representation and integration of auditory and visual stimuli inthe primate ventral lateral prefrontal cortex. Cereb. Cortex 17 (Suppl. 1), i61ei69.
Romanski, L.M., Averbeck, B.B., Diltz, M., 2005. Neural representation of vocaliza-tions in the primate ventrolateral prefrontal cortex. J. Neurophysiol. 93 (2),734e747.
Romanski, L.M., Bates, J.F., Goldman-Rakic, P.S., 1999a. Auditory belt and parabeltprojections to the prefrontal cortex in the rhesus monkey. J. Comp. Neurol. 403,141e157.
Romanski, L.M., Goldman-Rakic, P.S., 2002. An auditory domain in primate pre-frontal cortex. Nat. Neurosci. 5, 15e16.
Romanski, L.M., Hwang, J., 2012. Timing of audiovisual inputs to the prefrontalcortex and multisensory integration. Neuroscience 214, 36e48.
Please cite this article in press as: Plakke, B., et al., Coding of vocalizations(2013), http://dx.doi.org/10.1016/j.heares.2013.07.011
Romanski, L.M., Tian, B., Fritz, J., Mishkin, M., Goldman-Rakic, P.S., Rauschecker, J.P.,1999b. Dual streams of auditory afferents target multiple domains in the pri-mate prefrontal cortex. Nat. Neurosci. 2 (12), 1131e1136.
Russ, B.E., Lee, Y.S., Cohen, Y.E., 2007. Neural and behavioral correlates of auditorycategorization. Hear. Res. 229 (1e2), 204e212.
Sadagopan, S., Wang, X., 2009. Nonlinear spectrotemporal interactions underlyingselectivity for complex sounds in auditory cortex. J. Neurosci. 29 (36), 11192e11202.
Scott, B.H., Mishkin, M., Yin, P., 2012. Monkeys have a limited form of short-termmemory in audition. Proc. Natl. Acad. Sci. U. S. A 109 (30), 12237e12241.
Skuk, B.G., Schweinberger, S.R., 2013. Gender differences in familiar voice identifi-cation. Hear. Res. 296, 131e140.
Stein, B.E., Meredith, M.A., 1993. The Merging of the Senses. MIT Press, Cambridge.Sugihara, T., Diltz, M.D., Averbeck, B.B., Romanski, L.M., 2006. Integration of auditory
and visual communication information in the primate ventrolateral prefrontalcortex. J. Neurosci. 26, 11138e11147.
Tian, B., Reser, D., Durham, A., Kustov, A., Rauschecker, J.P., 2001. Functionalspecialization in rhesus monkey auditory cortex. Science 292, 290e293.
von Kriegstein, K., Giraud, A.L., 2006. Implicit multisensory associations influencevoice recognition. PLoS Biol. 4 (10), e326.
Wang, X., Merzenich, M.M., Beitel, R., Schreiner, C.E., 1995. Representation of aspecies-specific vocalization in the primary auditory cortex of the commonmarmoset: temporal and spectral characteristics. J. Neurophysiol. 74 (6), 2685e2706.
Webster, M.J., Bachevalier, J., Ungerleider, L.G., 1994. Connections of inferior tem-poral areas TEO and TE with parietal and frontal cortex in macaque monkeys.Cereb. Cortex 4, 470e483.
Wirth, S., Avsar, E., Chiu, C.C., Sharma, V., Smith, A.C., Brown, E., Suzuki, W.A., 2009.Trial outcome and associative learning signals in the monkey hippocampus.Neuron 61 (6), 930e940.
Yarmey, A.D., Yarmey, A.L., Yarmey, M.J., 1994. Face and voice identifications inshowups and lineups. Appl. Cogn. Psychol. 8 (5), 453e464.
by single neurons in ventrolateral prefrontal cortex, Hearing Research