clonidine-specific antisera recognize an endogenous...
TRANSCRIPT

341
Clonidine-Specific AntiseraRecognize an Endogenous Clonidine-Displacing
Substance in Brain
Mary P. Meeley, Andrew C. Towle, Paul Ernsberger, Lynette K. Char,
Phillip M. McCauley, and Donald J. Reis
An endogenous substance in brain, clonidine-displacing substance, binds to the same receptorpopulations as clonidine and is biologically active. Since receptor binding sites can be modeledby using specific antiligand antibodies, we tested the hypothesis that polyclonal antibodies raisedin rat and rabbit against the clonidine analog p-aminoclonidine coupled to hemocyanin wouldrecognize compounds structurally related to clonidine, including clonidine-displacing sub-stance. Binding to anti-p-aminoclonidine antibodies was examined by using a competitiveradioimmunoassay with tritiated p-aminoclonidine as the radioligand. Central vasodepressoragents that, like clonidine, are known to bind with high affinity to both imidazole sites anda2-adrenergic receptors in brain inhibited radioligand binding to anti-p-aminoclonidine anti-bodies. All of these agents contain imidazol(in)e and phenyl ring moieties as part of theirchemical structures (e.g., oxymetazoline); a number of other compounds without one or both ofthese rings failed to cross-react with the antisera. Clonidine-displacing substance, partiallypurified from bovine brain, also inhibited specific radioligand binding to antf-p-aminoclonidineantibodies. The inhibition was dose dependent and high affinity (ICg,, 4 Units). The endogenoussubstance had no effect on the apparent affinity of the antibodies for the radioligand, butblocked a specific number of binding sites. Immunoprecipitation experiments showed thatauthentic clonidine-displacing substance, that which displaces tritiated p-aminoclonidine bind-ing to membrane receptors, is recognized by anti-p-aminoclonidine antibodies. We concludethat a unique subset of structural determinants required for ligand interaction with bothimidazole and a2-adrenergic receptors is critical for binding to anti-p-aminoclonidine antibod-ies, and that since clonidine-displacing substance is recognized by highly clonidine-specificantisera, it may also contain these determinants within its structure, namely the imidazol(in)eand phenyl ring systems. (Hypertension 1989;13:341-351)
Clonidine, a synthetic phenyl imidazolidine, isa highly potent antihypertensive agent thatlowers arterial pressure largely by its action
within the C, area of the rostral ventrolateral medullaoblongata (VLM).1-4 It has been widely assumedthat the vasodepressor and other central actions ofclonidine, such as sedation5-6 and antiwithdrawaleffects,7-8 occur as a result of its role as an a2-adrenergic receptor partial agonist. However, wehave shown recently that clonidine binds with high
From the Division of Neurobiology and the Laboratory ofMolecular Neurobiology (A.C.T.), Cornell University MedicalCollege, New York, New York.
Supported in part by Public Health Service Grant HL-18974from the National Heart, Lung, and Blood Institute.
Address for correspondence: Mary P. Meeley, PhD, Divisionof Neurobiology, Cornell University Medical College, 411 East69th Street, New York, NY 10021.
Received May 17, 1988; accepted November 7, 1988.
affinity, not only to a2-adrenergic receptors, but to anewly described population of sites in brain that aredistinct from both adrenergic and histaminergicreceptors.9 These imidazole binding sites are pres-ent in the VLM in rats910 and in humans" and, infact, may be functional receptors that mediate thehypotensive actions of clonidine and other imida-zole agents in this region.312
The natural ligand for imidazole receptors isunknown. One possibility is an endogenous clonidine-displacing substance (CDS) isolated from bovinebrain.1314 CDS appears to be a relatively hydropho-bic, positively charged, low molecular weight sub-stance that displaces the binding of tritiated cloni-dine13 or the high-affinity clonidine analogue tritiated/7-aminoclonidine ([3H]PAC)14-16 to membranes frombovine and rat brains. Like clonidine, CDS binds toboth a2-adrenergic and imidazole receptors in VLM,with a 30-fold relative selectivity for the latter.1516
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

342 Hypertension Vol 13, No 4, April 1989
This clonidine-like substance is biologically active.Microinjection of CDS into the rostral VLM of therat14 and the cat17 results in dramatic changes inarterial pressure. In addition, CDS inhibits the twitchresponse in the vas deferens of the rat,18 facilitatesaggregation of human platelets,19 and causes a dose-related contraction of rat fundic smooth muscle.20
Recently, the endogenous substance itself has beenmeasured in some peripheral tissues, such as theadrenal gland, gastric fundus, and serum of the rat21
and in sera from both normotensive patients andpatients with pregnancy-induced hypertension.22 Thechemical structure of this clonidine-like substance is,as yet, undetermined.
Recently, it has been shown that antibodies raisedagainst selective drugs can be used as models forspecific recognition sites of receptor proteins.23-26
We reasoned, therefore, that antibodies to a conju-gate of p-aminoclonidine (PAC) may serve as probesfor the recognition sites of a2-adrenergic or imida-zole receptors, or both. Since CDS binds to thesesame receptor populations, the endogenous sub-stance may also be recognized by anti-PAC antibod-ies. If so, then the results of studying the ligandspecificity of the antibodies may yield clues as tothe chemical structure of CDS. Thus, we sought toproduce polyclonal anti-PAC antisera in rats and inrabbits, examine the ligand-binding specificity ofthe antisera relative to receptors labeled by [3H]PACin the VLM, and to determine whether anti-PACsbind CDS from brain.
A preliminary report of related findings with adifferent antiserum has appeared.27
Materials and MethodsProduction of Polyclonal Antisera
Immunogen was prepared as described previ-ously.27 The clonidine analogue PAC was coupledto hemocyanin (from Limulus polyphemus) withglutaraldehyde as the cross-linking agent. Sampling ofcombined media from exhaustive dialysis of the reac-tion product showed that nearly 40% of the [3H]PAC(New England Nuclear, Boston, Massachusetts, 43Ci/mmol) added as tracer was incorporated into thenondialyzable hemocyanin conjugate under the con-ditions used (i.e., coupling ratio, 0.4). The PAC-glutaraldehyde-hemocyanin conjugate (immunogen)was stored at —20° C until just before immunization.
At the time of injection, the immunogen wasthawed and emulsified with an equal volume ofcomplete Freund's adjuvant (1 mg immunogen/ml,final). The emulsion was injected into anesthetizedrats (0.050 ml, i.p. and 0.075 ml, s.c.) and rabbits(1.0 ml, s.c). Booster injections were adminis-tered to rats at week 2, 3, and 4 and to rabbits atweek 4 and 8. Ten days after the final injection,animals were anesthetized with halothane (2% inO2 blown over the nose) and bled via cardiacpuncture (rat) or femoral artery catheterization(rabbit). Blood samples were allowed to clot over-
night at 4° C; the antisera were centrifuged at 300gand stored as 1 ml aliquots at -20° C.
Preparation of Immunoglobulin FractionsFrom Antisera
For most experiments, a total protein fractionwas precipitated from rat (anti-PAC i) and rabbit(anti-PAC^ antisera with 50% (wt/vol) ammoniumsulfate.27 Precipitates were reconstituted in a mini-mum amount of 0.9% saline, dialyzed, adjusted toserum-equivalent volumes, and then stored frozenat -20° C until radioimmunoassay.
Alternatively, anti-PAC2 immunoglobulin Gs wereaffinity purified from serum by Protein A Sepharosechromatography. Anti-PAC2 serum (12 ml) wasloaded onto a Protein A Sepharose (PharmaciaBiotechnology Prods., Piscataway, New Jersey)column (10x0.9 cm, i.d.) that had been equilibratedat 4° C with phosphate-buffered saline (pH 7.0). Thecolumn was washed with phosphate-buffered salineat a flow rate of approximately 20 ml/hr until theoptical density at 280 nm (OD280) of the effluent wasless than 0.1. Immunoglobulin Gs were eluted in a0.1 M glycine buffer, pH 3.0, and fractions of 1 mlwere collected and immediately neutralized with 0.1M Tris base. Elution fractions containing greaterthan 0.1 OD280 were pooled and the immunoglobulinGs concentrated by precipitating with 50% ammo-nium sulfate. Saline-dialyzed aliquots of the anti-PAC2 immunoglobulin Gs were stored as 3x con-centrates (relative to original serum immunoglobulinG content) at -20° C.
Tritiated p-Aminoclonidine RadioimmunoassayTo examine the binding properties of anti-PAC
polyclonal antibodies, a radioimmunoassay withunconjugated [3H]PAC as radioligand was adaptedas described27 from the method of Sangameswaranand deBlas.28 Briefly, an aliquot of ammoniumsulfate-precipitated anti-PAC immunoglobulins wasthawed, then diluted to a 10x stock solution withbovine serum albumin (1 mg/ml). The anti-PACstock was added to 50 mM Tris-HCl (pH 7.4)containing the compound to be tested or CDS (seebelow) to give the final dilution indicated and waspreincubated at 25° C for 15 minutes. To definenonspecific binding, unlabeled PAC (20 .M) wasincluded in parallel preincubations. [3H]PAC (1 nM,or as specified) was added to all samples (incubationvolume, 0.5 ml) that were then vortexed and incu-bated for 15 minutes at 25° C, followed by 1 hour onice. The reaction was terminated by precipitatingwith carrier bovine gamma globulin (0.125% wt/vol)and ice-cold polyethylene glycol 8000 (7.5% wt/vol,final volume, 1.0 ml). Samples were vacuum-filteredover Whatman GF/B filters using a modified cellharvester (Brandel, Gaithersburg, Maryland). Thefilters were washed with 6-8 ml of 8% polyethyleneglycol in Tris-HCl, pH 7.4, then transferred toscintillation minivials, covered with 5 ml of scintil-lation cocktail, and counted at 45-50% efficiency.
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Meeley et al Antibodies to p-Aminoclonidine Recognize CDS 343
Protein was measured by the Coomassie Bluedye binding method of Bradford,29 as modified byRead and Northcote,30 with bovine gamma globu-lin as standard.
Isolation of Clonidine-Displacing SubstanceExtracts of calf brain containing CDS were
prepared according to methods described else-where,14-1627 with slight modification. Fresh brains,obtained from a local slaughterhouse (maximum 2hours postmortem), were cleaned of major bloodvessels and membranes, and the cerebellum wasremoved. The tissue was chopped into small pieces,then homogenized in three volumes (vol/wt) ofboiling distilled water using a Polytron (BrinkmanInstruments, Westbury, New York). The homoge-nate was centrifuged at 100,000g for 30 minutes at4° C. The supernatant then was denatured in amicrowave oven by heating just to boiling andrecentrifuged for 15 minutes; the resulting superna-tant was freeze-dried overnight. The solids werereconstituted in distilled water (50-100 ml/2 brainequivalents) and were transferred to a dialysis bagmade of H45 membrane tubing (Thomas Scientific,Philadelphia, Pennsylvania) with a cutoff of 3,500MW. Dialysis was carried out overnight at 4° Cagainst 3x20 volumes (vol/vol) of distilled water.The dialysis media (diffusates) were combined andfreeze-dried. The low molecular weight solids wereextracted with 20 volumes (vol/wt) of high-puritymethanol by bath sonicating for 3 minutes at roomtemperature. The extract was decanted and theresidue re-extracted with 5-10 volumes of freshmethanol. The extracts were pooled, filtered overWhatman (Clifton, New Jersey) No. 1 paper, andthen aliquoted and dried in a Speed-Vac (SavantInstruments, Farmingdale, New York). CDS-containing aliquots were stored dry at -70° C untilimmediately before use when they were reconsti-tuted in distilled water.
The amount of CDS in these preparations wasassayed by competition in radioreceptor bindingexperiments. Inhibition of the binding of [3H]PAC tomembrane receptors by CDS was measured by usingwashed P2 membranes prepared from bovine frontalcortex as previously described1416; a Unit of CDSactivity has been defined as the amount of extractrequired to inhibit [3H]PAC binding by 50%.14
Data AnalysisCompetition curves from studies of radioligand
binding to antibody recognition sites were analyzedby linear regression of Hill plots. More complexdata and data from multiple experiments were ana-lyzed with programs originally designed for quanti-fication of membrane receptor-ligand interactions:the Equilibrium Binding Data Analysis (EBDA) pro-gram of McPherson,31 which provides initial param-eter estimates, followed by subsequent input intothe LIGAND program32 for the final linear and non-linear multivariate least-squares analysis.
Materials[3H]PAC (New England Nuclear; 43 Ci/mmol) was
stored in ethanol at -20° C; subsequently, it wasdiluted with distilled water and stored at 4° C for upto 2 weeks before use. Detomidine, medetomidine,MPV 295, and MPV 830 were the generous gift ofJ.-M. Savola, Department of Pharmacology, Univer-sity of Oulu, Finland. Chloroethylclonidine was sup-plied by Research Biochemicals Inc. (South Natick,Massachusetts), idazoxan by Reckitt & Coleman(Kingston-upon-Hull, United Kingdom), and phen-tolamine by Ciba-Geigy (Summit, New Jersey). Ben-zimidazole and 1-benzylimidazole were from Chem-ical Dynamics Corporation (South Plainfield, NewJersey); all other compounds were purchased fromSigma Chemical Co. (St. Louis, Missouri).
ResultsBinding of Tritiated p-Aminoclonidineto Anti-p-Aminoclonidine Immunoglobulins
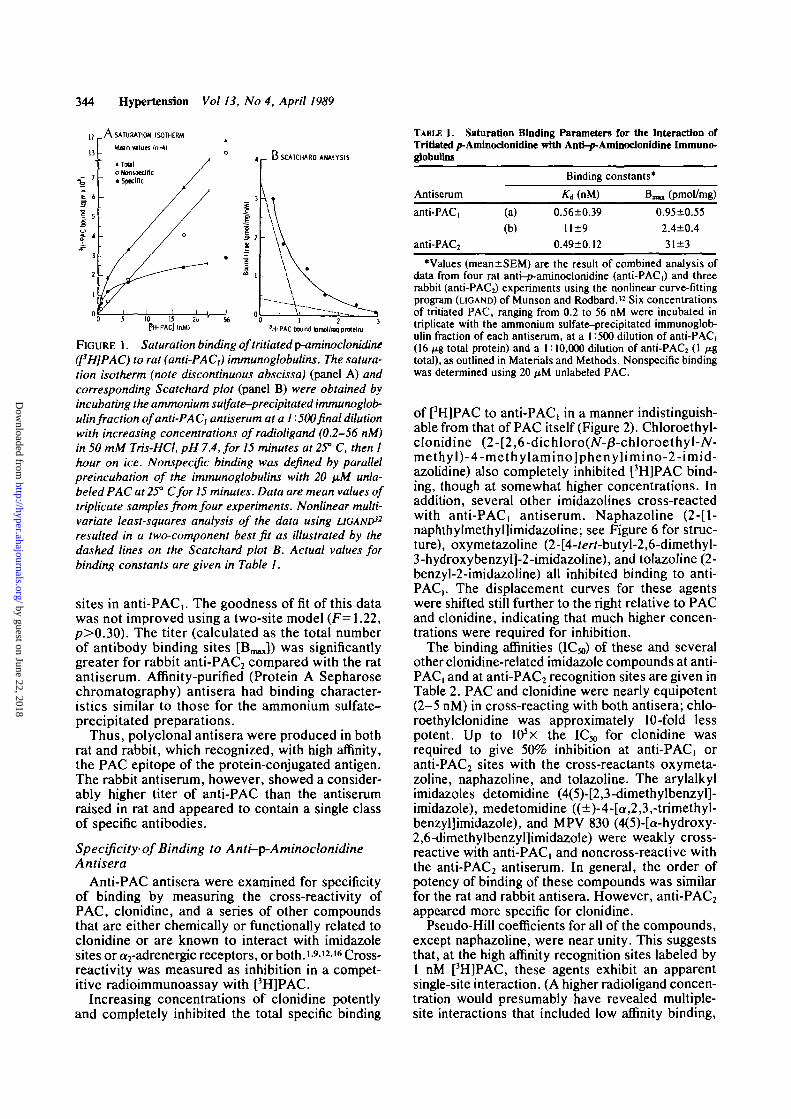
To determine whether antibodies to PAC wereproduced in rat or rabbit after immunization withthe synthetic PAC-glutaraldehyde-hemocyanin con-jugate, the specific binding of unconjugated [3H]PACto anti-PACs was examined. Saturation bindingexperiments were carried out by using the rapid-filtration radioimmunoassay technique detailed inMaterials and Methods. Saturation data for bindingto rat anti-PACi (1:500 final dilution) at [3H]PACconcentrations ranging over nearly three orders ofmagnitude are shown in Figure 1 A. The total amountof [3H]PAC bound increased with radioligand con-centration. Nonspecific binding, measured in thepresence of 20 ^M unlabeled PAC, accounted for29±2% of the total binding at 0.6 nM [3H]PAC(n=3). Specific [3H]PAC binding to anti-PAC, wasof high affinity and saturable; as the radioligandconcentration was more than tripled from 17 to 56nM, specific binding increased by only 20% (Figure1A; note discontinuous abscissa for complete rep-resentation). A Scatchard plot of this saturationdata (Figure IB) was distinctly curvilinear. Non-linear computerized curve-fitting analysis (LIGAND)32
for anti-PACi and for anti-PAC2 is summarized inTable 1.
For the range of radioligand concentrations exam-ined, rat anti-PACi appeared to contain at least twoclasses of antibody binding sites for [3H]PAC: onewith an apparent equilibrium binding constant (/Cd)of 0.56 nM and one with somewhat lower affinity,having a Kd of 11 nM. Thus, immunoglobulins withdifferent affinities for free PAC were present in theammonium sulfate-precipitated fraction of the poly-clonal antiserum. Anti-PAC2 from rabbit was assayedusing the same radioligand concentrations as withanti-PAC,, but at a 1:10,000 final antiserum dilu-tion to give a similar level of total specific [3H]PACbinding at 1 nM. Anti-PAC2 exhibited a singleclass of antibody binding sites, with an affinitynearly identical (Kd, 0.49 nM) to the high-affinity
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from
344 Hypertension Vol 13, No 4, April 1989
_ A SATURATION ISOTHERM
Mean values (n-414 B SCATCHARD ANALYSIS
TABLE 1. Saturation Binding Parameters for the Interaction ofTritiated p-Aminodonidine with Anti-p-Amlnoclonidine Imrauno-globullns
0 1 2 3*H-PAC bound ipmoJ/iag protein]
FIGURE 1. Saturation binding of tritiated p-aminoclonidine(I3H]PAC) to rat (anti-PAC,) immunoglobulins. The satura-tion isotherm (note discontinuous abscissa) (panel A) andcorresponding Scatchard plot (panel B) were obtained byincubating the ammonium sulfate-precipitated immunoglob-ulin fraction of anti-PAC) antiserum atal '• 500 final dilutionwith increasing concentrations of radioligand (0.2-56 nM)in 50 mM Tris-HCl, pH 7.4, for 15 minutes at 25° C, then 1hour on ice. Nonspecific binding was defined by parallelpreincubation of the immunoglobulins with 20 (JLM unla-beled PAC at 25° Cfor 15 minutes. Data are mean values oftriplicate samples from four experiments. Nonlinear multi-variate least-squares analysis of the data using LIGAND32
resulted in a two-component best fit as illustrated by thedashed lines on the Scatchard plot B. Actual values forbinding constants are given in Table 1.
sites in anti-PAC,. The goodness of fit of this datawas not improved using a two-site model (F=1.22,p>0.30). The titer (calculated as the total numberof antibody binding sites [B^]) was significantlygreater for rabbit anti-PAC2 compared with the ratantiserum. Affinity-purified (Protein A Sepharosechromatography) antisera had binding character-istics similar to those for the ammonium sulfate-precipitated preparations.
Thus, polyclonal antisera were produced in bothrat and rabbit, which recognized, with high affinity,the PAC epitope of the protein-conjugated antigen.The rabbit antiserum, however, showed a consider-ably higher titer of anti-PAC than the antiserumraised in rat and appeared to contain a single classof specific antibodies.
Specificity-of Binding to Anti-p-AminoclonidineAntisera
Anti-PAC antisera were examined for specificityof binding by measuring the cross-reactivity ofPAC, clonidine, and a series of other compoundsthat are either chemically or functionally related toclonidine or are known to interact with imidazolesites or a2-adrenergic receptors, or both. •-91216 Cross-reactivity was measured as inhibition in a compet-itive radioimmunoassay with [3H]PAC.
Increasing concentrations of clonidine potentlyand completely inhibited the total specific binding
Antiserum
anti-PAC i
anti-PAC2
(a)(b)
Binding
Ki (nM)
0.56±0.39ll±9
0.49±0.12
constants*
B, m (pmol/mg)
0.95+0.552.4±0.431±3
*Values (mean±SEM) are the result of combined analysis ofdata from four rat anti-p-aminoclonidine (anti-PAC,) and threerabbit (anti-PACJ experiments using the nonlinear curve-fittingprogram (LIGAND) of Munson and Rodbard.32 Six concentrationsof tritiated PAC, ranging from 0.2 to 56 nM were incubated intriplicate with the ammonium sulfate-precipitated immunoglob-ulin fraction of each antiserum, at a 1:500 dilution of anti-PAC,(16 /ig total protein) and a 1:10,000 dilution of anti-PAC2 (1 n%total), as outlined in Materials and Methods. Nonspecific bindingwas determined using 20 jtM unlabeled PAC.
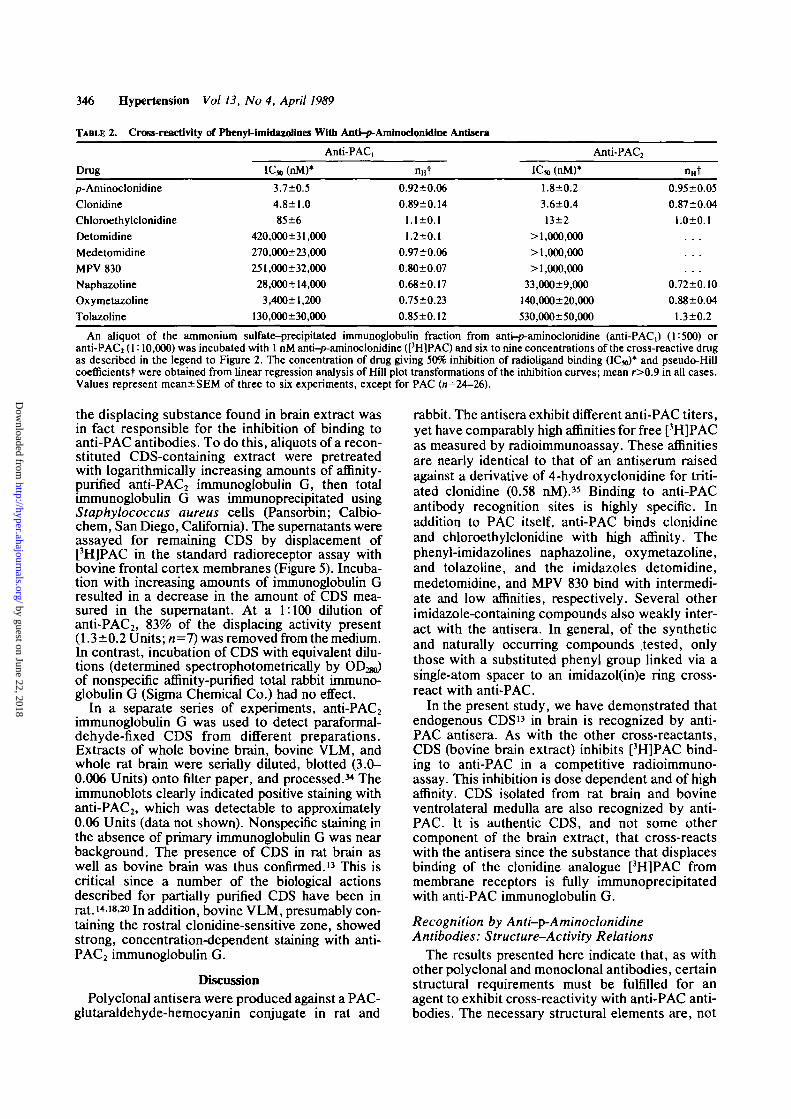
of [3H]PAC to anti-PACi in a manner indistinguish-able from that of PAC itself (Figure 2). Chloroethyl-clonidine (2-[2,6-dichloro(vV-0-chIoroethyl-A/-methyl)-4-methylamino]phenylimino-2-imid-azolidine) also completely inhibited [3H]PAC bind-ing, though at somewhat higher concentrations. Inaddition, several other imidazolines cross-reactedwith anti-PAC, antiserum. Naphazoline (2-[l-naphthylmethyljimidazoline; see Figure 6 for struc-ture), oxymetazoline (2-[4-terf-butyl-2,6-dimethyl-3-hydroxybenzyl]-2-imidazoline), and tolazoline (2-benzyl-2-imidazoline) all inhibited binding to anti-PAC,. The displacement curves for these agentswere shifted still further to the right relative to PACand clonidine, indicating that much higher concen-trations were required for inhibition.
The binding affinities (IC50) of these and severalother clonidine-related imidazole compounds at anti-PAC, and at anti-PAC2 recognition sites are given inTable 2. PAC and clonidine were nearly equipotent(2-5 nM) in cross-reacting with both antisera; chlo-roethylclonidine was approximately 10-fold lesspotent. Up to lO^x the IC50 for clonidine wasrequired to give 50% inhibition at anti-PAC, oranti-PAC2 sites with the cross-reactants oxymeta-zoline, naphazoline, and tolazoline. The arylalkylimidazoles detomidine (4<5)-[2,3-dimethylbenzyl]-imidazole), medetomidine ((±)-4-[a,2,3,-trimethyl-benzyl]imidazole), and MPV 830 (4(5)-[a-hydroxy-2,6-dimethylbenzyl]imidazole) were weakly cross-reactive with anti-PAC, and noncross-reactive withthe anti-PAC2 antiserum. In general, the order ofpotency of binding of these compounds was similarfor the rat and rabbit antisera. However, anti-PAC2appeared more specific for clonidine.
Pseudo-Hill coefficients for all of the compounds,except naphazoline, were near unity. This suggeststhat, at the high affinity recognition sites labeled by1 nM [3H]PAC, these agents exhibit an apparentsingle-site interaction. (A higher radioligand concen-tration would presumably have revealed multiple-site interactions that included low affinity binding,
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Meeley et al Antibodies to p-Aminodonidine Recognize CDS 345
• p-Ajnfnodonidnso Cbnldtne• ChJofoethyfdonMlnfl• Oxymetazoina» NaphazoBneA Tolazoflne
Mean values (n»4-24)
FIGURE 2. Inhibition of tritiated p-aminoclonidine(['HIPAC) binding to rat (anti-PAC,) immunoglobulins byclonidine-related compounds. The immunoglobulin-containing fraction of anti-PAC) (1 -500) waspreincubatedwith the compounds indicated at a series of concentrationsin Tris-HCl (pH 7.4) at 25° C for 15 minutes. Parallelpreincubations were carried out in the presence of 20 fiMunlabeled PAC to define nonspecific binding. Radioligand(I nM) was added and the samples incubated for anadditional 15 minutes, followed by 1 hour on ice. Thereaction was terminated, samples were rapid filtered, andthe filters counted. Data are reported as the averagepercent total specific binding for four to 11 experiments.
for example, with anti-PAC,.) Naphazoline appearedto discriminate among more than one class of siteswith high affinity for the radioligand in both anti-sera. Thus, the multiple species of anti-PAC anti-bodies undoubtedly present in the two antiseraexhibit indistinguishable, apparently single-classaffinities for all of the cross-reacting ligands (exceptnaphazoline) under the radioimmunoassay condi-tions specified.
Other compounds that did not effectively cross-react (i.e., inhibit 50% of specific [3H]PAC binding)with either of the anti-PAC antisera are listed inTable 3. A number of naturally occurring and syn-thetic substances had no significant effect on bind-ing. These included the neurotransmitters acetyl-choline, GABA, norepinephrine, and serotonin.Some compounds, most of which were imidazoles[e.g., idazoxan ([l,4-benzodioxan-2-yl]-2-imid-azoline); see Figure 6 for structure], histamine,histidyl-tyrosine, cimetidine (N-Cyano-N-me\hy\-N-[2-[[(5-methyl-1 H-imidazol-4-yDmethyl]thio]ethyriguan-idine), and MPV 295 (4(5)-2-[2,6-dimethylphenyl]-
ethylimidazole) significantly inhibited binding to eitheranti-PAC, or anti-PAC2, or both, at 10-4-l(r3 Mconcentration. However, none of these agents inhib-ited binding by 50% or more. Finally, althoughpolyclonal antisera may be expected to containantibodies produced to the protein portion of hapten-protein conjugates, hemocyanin (1 mg/ml, final con-centration), the carrier protein used in synthesizingthe PAC immunogen, did not interfere with [3H]PACbinding to the antibodies (data not shown), that is, ithad no effect in this assay system.
Thus, binding to polyclonal anti-PAC antibodiesraised in two different species was highly specific.Only those agents that, like clonidine, contain phenyland imidazol(in)e ring moieties in their structurescompeted for binding to antibody recognition sites.
Recognition of Clonidine-Displacing Substance byAnti-p-Aminoclonidine Antibodies
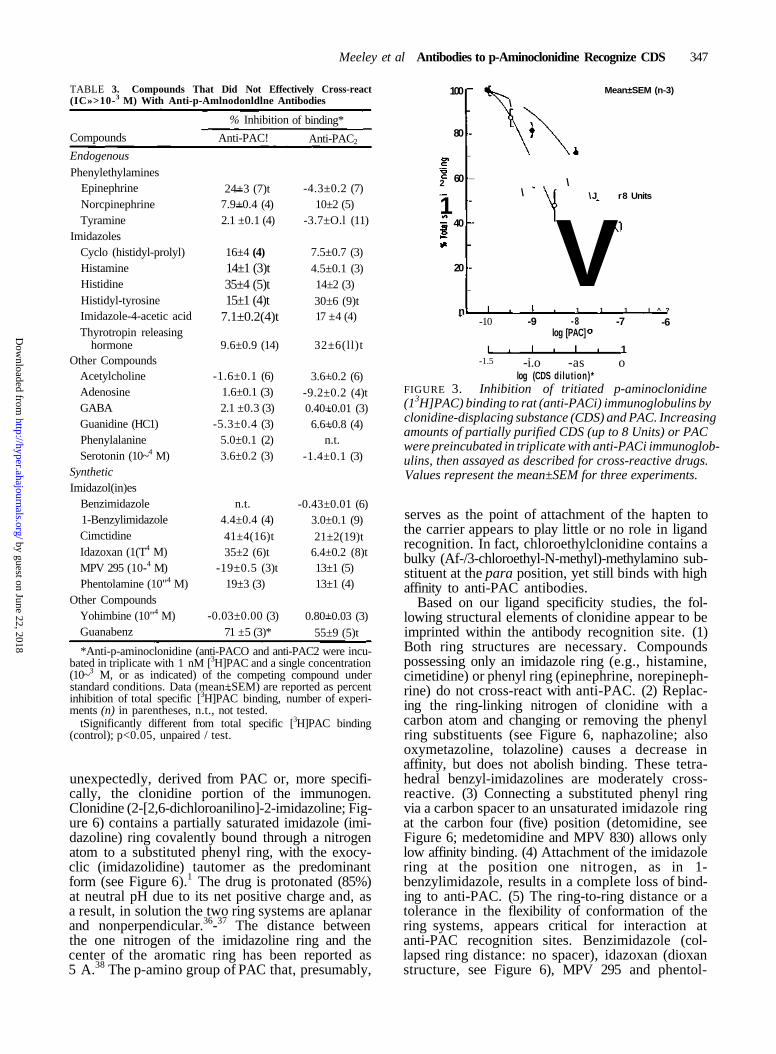
Given the strict structural requirements for cross-reactivity, we sought to determine whether CDSwas truly clonidine-like in that it also was recog-nized by anti-PAC antibodies. As shown in Figure3, increasing amounts of bovine brain extract con-taining CDS inhibited [3H]PAC binding to anti-PACi in a concentration-dependent manner. TheIC50 value for the interaction of CDS with theantiserum was 4.2±0.2 Units. As with PAC itself,this interaction appeared to occur at a single class of[3H]PAC recognition sites since the pseudo-Hillcoefficient was 0.88±0.32 (r>0.9; n=3). Experi-ments carried out with the rabbit antiserum gavesimilar results (data not shown).
To further characterize the apparent cross-reactivity of CDS with anti-PAC antibodies, anti-PAC2 immunoglobulin G that had been affinitypurified on Protein A Sepharose (see Materials andMethods) was used. The immunoglobulin G waspreincubated either in the presence or absence of afixed amount of CDS (approximating its ICJO atanti-PAC, sites: average, 3.2 Units), and then satu-ration binding of [3H]PAC was examined. Scatchardanalysis of the results (Figure 4) showed that CDShad no effect on the radioligand binding affinity ofanti-PAC2: the slopes for data obtained in eithercondition were parallel, giving nearly identical K6values. However, a significant number of [3H]PACbinding sites were blocked by CDS as indicated bythe shift in Scatchard x intercept (B^ in thepresence of CDS. The results are typical of anoncompetitive interaction and are expected forantibody binding, which is an essentially irrevers-ible process, that is, primarily association and notdissociation dependent.33 Nonspecific binding wasunaffected by coincubation with CDS.
Since the experiments described here involvedthe use of partially purified extracts of brain con-taining CDS and since CDS is defined in all of ourstudies i*-i6,27 ^ a substance that displaces thebinding of [3H]PAC to bovine frontal cortex mem-brane receptors, it was critical to determine whether
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from
346 Hypertension Vol 13, No 4, April 1989
TABLE 2. Cross-reactivity of Pbtnyi-imidazoUnes With Anti-p-AminodonidiiK Antisera
Drug
p-Aminoclonidine
Clonidine
Chloroethylclonidine
Detomidine
Medetomidine
MPV830
Naphazoline
Oxymetazoline
Tolazoline
Anti-PAC,
ICJO (nM)»
3.7±0.5
4.8±1.0
85±6
42O,OOO±31,0OO
270,000+23,000
251,000+32,000
28,000+14,000
3,400±l,200
130,0O0±3O,0O0
nHt0.92±0.06
0.89±0.14
l . l±0.1
1.2±0.1
0.97±0.06
0.80±0.07
0.68±0.17
0.75±0.23
0.85±0.12
Anti-PAC2
IC50 (nM)*
1.8±0.2
3.6+0.4
13±2
> 1,000,000
> 1,000,000
> 1,000,000
33,OO0±9,O00
14O,0OO±20,0O0
53O,0O0±50,OOO
nHt0.95+0.05
0.87±0.04
1.0±0.1
0.72±0.10
0.88+0.04
1.3±0.2
An aliquot of the ammonium sulfate-precipitated immunoglobulin fraction from anti-p-aminoclonidine (anti-PACi) (1:500) oranti-PAC2 (1:10,000) was incubated with 1 nM anti-p-aminoclonidine ([3H]PAC) and six to nine concentrations of the cross-reactive drugas described in the legend to Figure 2. The concentration of drug giving 50% inhibition of radioligand binding (ICJO)* and pseudo-Hillcoefficientst were obtained from linear regression analysis of Hill plot transformations of the inhibition curves; mean r>0.9 in all cases.Values represent mean+SEM of three to six experiments, except for PAC (n=24—26).
the displacing substance found in brain extract wasin fact responsible for the inhibition of binding toanti-PAC antibodies. To do this, aliquots of a recon-stituted CDS-containing extract were pretreatedwith logarithmically increasing amounts of affinity-purified anti-PAC2 immunoglobulin G, then totalimmunoglobulin G was immunoprecipitated usingStaphylococcus aureus cells (Pansorbin; Calbio-chem, San Diego, California). The supernatants wereassayed for remaining CDS by displacement of[3H]PAC in the standard radioreceptor assay withbovine frontal cortex membranes (Figure 5). Incuba-tion with increasing amounts of immunoglobulin Gresulted in a decrease in the amount of CDS mea-sured in the supernatant. At a 1:100 dilution ofanti-PAC 2, 83% of the displacing activity present(1.3±0.2 Units; n=l) was removed from the medium.In contrast, incubation of CDS with equivalent dilu-tions (determined spectrophotometrically by ODjgo)of nonspecific affinity-purified total rabbit immuno-globulin G (Sigma Chemical Co.) had no effect.
In a separate series of experiments, anti-PAC2immunoglobulin G was used to detect paraformal-dehyde-fixed CDS from different preparations.Extracts of whole bovine brain, bovine VLM, andwhole rat brain were serially diluted, blotted (3.0-0.006 Units) onto filter paper, and processed.34 Theimmunoblots clearly indicated positive staining withanti-PAC2, which was detectable to approximately0.06 Units (data not shown). Nonspecific staining inthe absence of primary immunoglobulin G was nearbackground. The presence of CDS in rat brain aswell as bovine brain was thus confirmed.13 This iscritical since a number of the biological actionsdescribed for partially purified CDS have been inrat i4,i8,2o j n addition, bovine VLM, presumably con-taining the rostral clonidine-sensitive zone, showedstrong, concentration-dependent staining with anti-PAC2 immunoglobulin G.
DiscussionPolyclonal antisera were produced against a PAC-
glutaraldehyde-hemocyanin conjugate in rat and
rabbit. The antisera exhibit different anti-PAC titers,yet have comparably high affinities for free [3H]PACas measured by radioimmunoassay. These affinitiesare nearly identical to that of an antiserum raisedagainst a derivative of 4-hydroxyclonidine for triti-ated clonidine (0.58 nM).35 Binding to anti-PACantibody recognition sites is highly specific. Inaddition to PAC itself, anti-PAC binds clonidineand chloroethylclonidine with high affinity. Thephenyl-imidazolines naphazoline, oxymetazoline,and tolazoline, and the imidazoles detomidine,medetomidine, and MPV 830 bind with intermedi-ate and low affinities, respectively. Several otherimidazole-containing compounds also weakly inter-act with the antisera. In general, of the syntheticand naturally occurring compounds tested, onlythose with a substituted phenyl group linked via asingle-atom spacer to an imidazol(in)e ring cross-react with anti-PAC.
In the present study, we have demonstrated thatendogenous CDS13 in brain is recognized by anti-PAC antisera. As with the other cross-reactants,CDS (bovine brain extract) inhibits [3H]PAC bind-ing to anti-PAC in a competitive radioimmuno-assay. This inhibition is dose dependent and of highaffinity. CDS isolated from rat brain and bovineventrolateral medulla are also recognized by anti-PAC. It is authentic CDS, and not some othercomponent of the brain extract, that cross-reactswith the antisera since the substance that displacesbinding of the clonidine analogue [3H]PAC frommembrane receptors is fully immunoprecipitatedwith anti-PAC immunoglobulin G.
Recognition by Anti-p-AminoclonidineAntibodies: Structure-Activity Relations
The results presented here indicate that, as withother polyclonal and monoclonal antibodies, certainstructural requirements must be fulfilled for anagent to exhibit cross-reactivity with anti-PAC anti-bodies. The necessary structural elements are, not
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from
Meeley et al Antibodies to p-Aminoclonidine Recognize CDS 347
TABLE 3. Compounds That Did Not Effectively Cross-react(IC»>10-3 M) With Anti-p-Amlnodonldlne Antibodies
Compounds
Endogenous
PhenylethylaminesEpinephrineNorcpinephrineTyramine
ImidazolesCyclo (histidyl-prolyl)HistamineHistidineHistidyl-tyrosineImidazole-4-acetic acidThyrotropin releasing
hormoneOther Compounds
AcetylcholineAdenosineGABAGuanidine (HC1)PhenylalanineSerotonin (10~4 M)
SyntheticImidazol(in)es
Benzimidazole1-BenzylimidazoleCimctidineIdazoxan (1(T4 M)MPV 295 (10-4 M)Phentolamine (10"4 M)
Other CompoundsYohimbine (10"4 M)Guanabenz
% Inhibition
Anti-PAC!
24+3 (7)t7.9+0.4 (4)2.1 ±0.1 (4)
16±4 (4)14±1 (3)t35±4 (5)t15±1 (4)t
7.1±0.2(4)t
9.6±0.9 (14)
-1.6±0.1 (6)1.6±0.1 (3)2.1 ±0.3 (3)
-5.3±0.4 (3)5.0±0.1 (2)3.6±0.2 (3)
n.t.4.4±0.4 (4)41±4(16)t35±2 (6)t
-19±0.5 (3)t19±3 (3)
-0.03±0.00 (3)71 ±5 (3)*
of binding*
Anti-PAC2
-4.3±0.2 (7)10±2 (5)
-3.7±O.l (11)
7.5±0.7 (3)4.5±0.1 (3)14±2 (3)30±6 (9)t17 ±4 (4)
32±6(ll)t
3.6+0.2 (6)-9.2±0.2 (4)t0.40+0.01 (3)6.6+0.8 (4)
n.t.-1.4±0.1 (3)
-0.43±0.01 (6)3.0±0.1 (9)21±2(19)t
6.4±0.2 (8)t13±1 (5)13±1 (4)
0.80+0.03 (3)55±9 (5)t
*Anti-p-aminoclonidine (anti-PACO and anti-PAC2 were incu-bated in triplicate with 1 nM [3H]PAC and a single concentration(10~3 M, or as indicated) of the competing compound understandard conditions. Data (mean+SEM) are reported as percentinhibition of total specific [3H]PAC binding, number of experi-ments (n) in parentheses, n.t., not tested.
tSignificantly different from total specific [3H]PAC binding(control); p<0.05, unpaired / test.
unexpectedly, derived from PAC or, more specifi-cally, the clonidine portion of the immunogen.Clonidine (2-[2,6-dichloroanilino]-2-imidazoline; Fig-ure 6) contains a partially saturated imidazole (imi-dazoline) ring covalently bound through a nitrogenatom to a substituted phenyl ring, with the exocy-clic (imidazolidine) tautomer as the predominantform (see Figure 6).1 The drug is protonated (85%)at neutral pH due to its net positive charge and, asa result, in solution the two ring systems are aplanarand nonperpendicular.36-37 The distance betweenthe one nitrogen of the imidazoline ring and thecenter of the aromatic ring has been reported as5 A.38 The p-amino group of PAC that, presumably,
2
1
100
80
60
40
20
n
-
_
-
-
\
\
Mean±SEM (n-3)
\\J r8 Units
V1 1 1 I ^ ?
-10 -9 -8log [PAC]
-7
1
-6
-1.5 -i.o -as olog (CDS dilution)*
FIGURE 3. Inhibition of tritiated p-aminoclonidine(13H]PAC) binding to rat (anti-PACi) immunoglobulins byclonidine-displacing substance (CDS) and PAC. Increasingamounts of partially purified CDS (up to 8 Units) or PACwere preincubated in triplicate with anti-PACi immunoglob-ulins, then assayed as described for cross-reactive drugs.Values represent the mean±SEM for three experiments.
serves as the point of attachment of the hapten tothe carrier appears to play little or no role in ligandrecognition. In fact, chloroethylclonidine contains abulky (Af-/3-chloroethyl-N-methyl)-methylamino sub-stituent at the para position, yet still binds with highaffinity to anti-PAC antibodies.
Based on our ligand specificity studies, the fol-lowing structural elements of clonidine appear to beimprinted within the antibody recognition site. (1)Both ring structures are necessary. Compoundspossessing only an imidazole ring (e.g., histamine,cimetidine) or phenyl ring (epinephrine, norepineph-rine) do not cross-react with anti-PAC. (2) Replac-ing the ring-linking nitrogen of clonidine with acarbon atom and changing or removing the phenylring substituents (see Figure 6, naphazoline; alsooxymetazoline, tolazoline) causes a decrease inaffinity, but does not abolish binding. These tetra-hedral benzyl-imidazolines are moderately cross-reactive. (3) Connecting a substituted phenyl ringvia a carbon spacer to an unsaturated imidazole ringat the carbon four (five) position (detomidine, seeFigure 6; medetomidine and MPV 830) allows onlylow affinity binding. (4) Attachment of the imidazolering at the position one nitrogen, as in 1-benzylimidazole, results in a complete loss of bind-ing to anti-PAC. (5) The ring-to-ring distance or atolerance in the flexibility of conformation of thering systems, appears critical for interaction atanti-PAC recognition sites. Benzimidazole (col-lapsed ring distance: no spacer), idazoxan (dioxanstructure, see Figure 6), MPV 295 and phentol-
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

348 Hypertension Vol 13, No 4, April 1989
400
300
f
•5
1 2 0 0
o Without CDSKo-0.45±.03nM
• With CDSIgG
IgG
Mean values (n-3)
25 50 75Bound (pmol/mg IgG)
100
FIGURE 4. Scatchard plot showing saturation binding oftritiated p-aminoclonidine ([3H]PAC) to rabbit (anti-PAC2) antibodies in the presence and absence ofclonidine-displacing substance (CDS). Affinity-purified anti-PACjimmunoglobulin G (IgG) was preincubated at a 1 '• 10,000final dilution with and without CDS (3.2 units of methanol-extracted CDS per tube). Radioligand was added at eightincreasing concentrations, and the assay carried outunder standard conditions. The Scatchard analysis shownhere of the saturation data from three experiments indi-cates no change in affinity constant (Kd) in the presenceof CDS, but a decrease in the apparent total number ofbinding sites (B^). Nonspecific binding, defined in par-allel incubations with 20 fiM unlabeled PAC, was unaf-fected by CDS.
amine (extended ring distances: two-atom spacers),and histidyl-tyrosine (seven-atom spacers) fail tocross-react effectively and exhibit only a weakinteraction with anti-PAC antibodies. Thus, in gen-eral, the spatial orientation of the phenyl and imi-dazol(in)e rings, and perhaps their degree of substi-tution, constitute the critical chemical determinantsof the clonidine epitope. Since they are not readilyavailable for study, it is unclear whether unsatur-ated (imidazole) analogues of the partially saturatedimidazolines, linked at ring position two (i.e., sub-stituted 2-benzylimidazoles), would be as effectiveas clonidine in binding to anti-PAC antibodies.
Recognition by Anti-p-AminoclonidineAntibodies: Antihypertensive Agents andTheir Receptors
Two types of receptors are localized in the VLMthat are labeled by the clonidine derivative [3H]PAC.It is at 'er2-adrenergic receptors in the C, area of thisregion that clonidine has traditionally been thoughtto act as a partial agonist to lower arterial pres-sure.1-39 However, clonidine binds with even higher
1.6
1.4
1.2
I 1.0
- 0.8o
a6
0.4
0.2
0 L
oAnti-PAC2lgG• Normal rabbit IgG
Mean values ±SEM(n-H>
4
no IgG
1:100000 UQOOO 1:1,000IgG dilution
1:100
FIGURE 5. Immunoprecipitation of clonidine-displacingsubstance (CDS) receptor binding activity with rabbitanti-p-aminoclonidine (anti-PAC2) immunoglobulin G(IgG). Triplicate samples of CDS were incubated withaffinity-purified anti-PAC2 at 4 dilutions (1 • 100-100,000)and a buffer-only control, for 90 minutes at 25° C (0.2 mltotal volume). Equivalent dilutions of nonspecific affinity-purified total rabbit IgG (Sigma) were also incubated withCDS (three of seven experiments). A standardized sus-pension of Pansorbin cells (Calbiochem; 0.05 ml), with abinding capacity of 2.1 mg IgGlml, was added, and thesamples incubated for an additional 5 minutes. IgG-complexed cells were removed by centrifugation, and thesupernatants assayed by [3H]PAC radioreceptor assay todetermine Units of CDS activity. Values represent themean±SEM of the Units remaining after IgG treatment.
affinity to a newly discovered class of putative imi-dazole receptors in VLM, which are distinct fromadrenergic and histaminergic receptors.9 Recent evi-dence from our laboratory suggests that the vasode-pressor potency of clonidine and related compoundscorrelates best with binding affinity at imidazolereceptor sites, not a2-adrenergic receptors.12 Alter-natively, vasodepressor potency may result fromconcerted effects via both receptor types. This dualaction would be structurally represented by thephenyl moiety of phenylethylamine adrenergic agentsalong with the imidazol(in)e moiety of histaminergicagents, both of which are present in clonidine-likeagents. These possibilities raise the question as towhether the recognition sites of clonidine-specificantibodies resemble classic a2-adrenergic sites, imi-dazole sites, or a hybrid of both receptors.
Specific anti-PAC antibodies cross-react with clo-nidine and only a few other agents, all of which bind
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Meeley et al Antibodies to p-Aminodonidine Recognize CDS 349
CROSSSTRONG (ICso<100nM)
Cl N v '
/ \ / H
In-AmlnolclonldJne
REACTIVE
MODERATE
aNaphazollnei
WEAK
HjC CH3 N J
— H H
Detomidine
NON CROSS-REACTIVE
oo—( ^
Idazoran
FIGURE 6. Chemical structures of clonidineand related compounds. Clonidine exists insolution chiefly as the exocyclic imidazolidinetautomer, which is protonated at physiologi-cal pH. Thus, the phenyl and imidazoline ringsystems are nonperpendicular and aplanar.It is because of this arrangement of theclonidine epitope, presumably, that clonidine-specific antibodies can recognize other apla-nar phenyl imidazoline derivatives, such asnaphazoline and, to a lesser extent, detomi-dine. Antibody interaction with detomidinemay be weak because the unsaturated imida-zole ring is attached at position fourifive)rather than position two. Idazoxan containsboth the phenyl and imidazoline rings; how-ever, it is not well-recognized by anti-p-aminoclonidine antibodies. This may be dueto the presence of the stoically rigid dioxangroup or an inappropriate inter-ring dis-tance, or both.
with high affinity (nanomolar range) to both imida-zole and a2-adrenergic receptor populations (seeReference 9; detomidine, medetomidine, MPV 830,and tolazoline, unpublished observations). Thus,anti-PAC sites resemble a hybrid of imidazole anda2-adrenergic receptor sites. Significantly, of theagents tested thus far in vivo, all of those thatcross-react with anti-PAC antibodies elicit hypoten-sion when administered directly into the centralnervous system. Specifically, clonidine and PACboth bind with high affinity to anti-PAC recognitionsites, and both are exquisitely potent central vaso-depressor agents.'34- >2-40 Naphazoline and oxymeta-zoline also lower arterial pressure on centraladministration,41 but only at doses that exceedthose of clonidine by two orders of magnitude.These two compounds also exhibit a substantiallylower affinity for anti-PAC sites compared withclonidine. In addition, detomidine, medetomidine,and MPV 830 are potent sedatives that act asvasodepressor agents via central mechanisms.42-43
These sedative agents are a2-selective (P. Erns-berger, unpublished observation) and only weaklycross-reactive with anti-PAC antibodies. Thus,binding to anti-PAC antibodies defines a uniquesubset of agents with affinities for anti-PAC sitesthat appear to be qualitatively related to theireffectiveness in lowering arterial pressure within thecentral nervous system. Although several noncross-reactive compounds show moderate central hypo-tensive actions, such as guanabenz (a2-adrenergic),and idazoxan and cimetidine (imidazole),12 bindingto anti-PAC antibodies may be useful in providinginformation for the design of novel centrally actingantihypertensive agents.
Recognition by Anti-p-AminoclonidineAntibodies: Clonidine-Displacing Substance
The recognition of CDS by antibodies raisedagainst the clonidine analog PAC is added evidence
that this endogenous substance from brain is trulyclonidine-like. The fact that anti-PAC antibodiesexhibit such strict structural requirements for bind-ing, and yet. CDS is also recognized by theseantibodies, strongly suggests that the natural sub-stance shares some of the same chemical elementsas clonidine and the other cross-reactants, namely,the phenyl and imidazol(in)e ring moieties. Thereported finding that CDS is relatively hydrophobicand positively charged at neutral pH13 is consistentwith the presence of these structural elements. Thisobservation may provide useful clues for the struc-tural analysis and identification of purified CDS.
Binding to clonidine-specific antibodies also sup-ports our proposal that CDS is a unique substancein brain.16 CDS appears to be distinct from anumber of endogenous compounds, including thecatecholamines and other neurotransmitters such asacetylcholine, GABA, histamine, and serotonin,since these do not bind effectively to anti-PAC.Furthermore, CDS is one of a small and unique setof agents that bind to the imidazole and a2-adrenergic receptor populations in VLM1516 as wellas to anti-PAC antibodies. This finding may accountfor the clonidine-related central cardiovascularactions of CDS.1417 We cannot, of course, rule outthe possibility that more than one substance in ourpartially purified CDS preparation combines in theantibody recognition site to result in a unique profileof inhibition of [3H]PAC binding. However, thedose-response relation for clonidine and for CDSappear to be similar, based on pseudo-Hill coeffi-cients. In addition, receptor binding experimentsusing the same CDS preparation described hereindicate that these extracts contain a single activespecies.16 Furthermore, our data show that morethan 85% of this inhibitory species can be immuno-precipitated with anti-PAC antibodies.
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

350 Hypertension Vol 13, No 4, April 1989
Recognition of endogenous CDS by anti-PACantibodies is probably the result of a relatively weakinteraction. That is, CDS binds with lower affinityto antibody recognition sites (IC50, 4 Units) than toa2-adrenergic (1 Unit)14 or imidazole (0.03 Units)16
receptor sites labeled by [3H]PAC. This pattern issimilar to that of the low affinity cross-reactantsnaphazoline and oxymetazoline, but contrasts withthat for the high affinity clonidine epitope. Cloni-dine binds with sixfold higher affinity to anti-PACantibodies than to receptors in VLM.9 Once identi-fied, the chemical structure of CDS as it compareswith clonidine will help to explain these observeddifferences in binding affinities. For example, CDSmay contain an imidazole ring and not the partiallysaturated imidazoline ring that is present in syn-thetic compounds such as clonidine and naphazo-line. The degree of saturation of this ring may beimportant for high affinity binding to a clonidine-specific antibody and less critical for interactionwith receptors, which have a broader specificity.
Recognition by Anti-p-AminoclonidineAntibodies: Quantifying Clonidine-DisplacingSubstance Using Radioimmunoassay
The production of polyclonal antisera againstPAC has provided us with a tool for further evalu-ating the ligand binding specificities of imidazoleand a2-adrenergic receptors and with indirect infor-mation on the chemical structure of endogenousCDS, since it also binds to anti-PAC. In addition,we now have a highly specific means of measuringrelative amounts of CDS in brain extracts or otherbiological samples. Despite being somewhat lesssensitive than the [3H]PAC radioreceptor assay, the[3H]PAC radioimmunoassay has the advantage ofbeing a more accurate method for determining lev-els of CDS. Various salts or other factors that areknown to interfere with or displace [3H]PAC bind-ing to receptors,944 which may be present in par-tially purified preparations of CDS, do not affectbinding to anti-PAC antibodies. Thus, it is likelythat radioimmunoassay values represent a truermeasure of CDS activity in crude preparations thanthe radioreceptor assay estimates of CDS Unitsused heretofore. Furthermore, used in tandemwith radioreceptor1416 or bioassays1820 for CDS,the radioimmunoassay ensures positive identifica-tion and accurate tracking of the substance duringpurification.
Furthermore, in addition to determining relativelevels of CDS activity based on extent of inhibition,absolute amounts of CDS can be estimated usingsaturation binding data from the radioimmunoas-say. For example, the results of the saturationbinding of [3H]PAC to anti-PAC2 immunoglobulin Gcarried out in the presence of CDS (refer to Figure4) can be used to estimate the molar quantities ofCDS in bovine brain. As its IC50 concentration, onlyapproximately 2% of the [3H]PAC that was added tothe assay tubes bound to the antibodies. CDS, at a
concentration near its IC^, blocked 29 pmol of[3H]PAC binding sites per milligram immunoglob-ulin G. Assuming that, as with the radioligand,about 2% of the CDS added was bound to thesesites and that the interaction of CDS at anti-PACsites is at least a thousandfold weaker than that ofPAC itself (see above), then, to a first approxima-tion, no more than 0.002% of the known amount ofCDS added was bound to recognition sites. If theassumptions are valid, then 1 Unit of CDS isequivalent to 110 pmol of CDS. On this basis, givena molecular mass of 590 Daltons,45 a yield of 130 ngof CDS per gram of bovine brain tissue is obtainedroutinely in our preparations. This estimated levelis on the order of that of several transmitter sub-stances known to be localized in discrete regions ofthe brain, such as norepinephrine,46 epinephrine47
histamine,48 and a number of neuropeptides (forreviews, see Reference 49).
We conclude that highly specific antisera pro-duced against the clonidine analogue PAC bind anendogenous CDS from brain and may serve asuseful immunological tools for localization, purifi-cation, and structural identification of this sub-stance. In addition, anti-PAC, along with otheranti-drug antibodies, may be useful as highly spe-cific models for the receptor subpopulations respon-sible for mediating the antihypertensive, sedative,and antiwithdrawal effects of clonidine.
AcknowledgmentsThe expert aid and advice of Dr. Corinne Abate,
Ms. Janie L. Callaway, and Dr. Teresa A. Milner isgratefully acknowledged.
References1. Kobinger W: Central a-adrenergic systems as targets for
hypotensive drugs. Rev Physiol Biochem Pharmacol 1978;81:39-100
2. Bousquet P, Schwartz J: a-Adrenergic drugs: Pharmacolog-ical tools for the study of the central vasomotor control.Biochem Pharmacol 198332:1459-1465
3. Bousquet P, Feldman J, Schwartz J: Central cardiovasculareffects of a-adrenergic drugs: Differences between catechol-amines and imidazolines. J Pharmacol Exp Ther 1984;230:232-236
4. Granata AR, Numao Y, Kumada M, Reis DJ: Al noradren-ergic neurons tonically inhibit sympathoexcitatory neuronsof the Cl area in rat brainstem. Brain Res 1986;377:127-146
5. Delbarre B, Schmitt H: Sedative effects of a-sympathomi-metic drugs and their antagonism by adrenergic and cholin-ergic blocking drugs. Eur J Pharmacol 1971 ;13:356-363
6. Delbarre B, Schmitt H: A further attempt to characterizesedative receptors activated by clonidine in chickens andmice. Eur J Pharmacol 1973;22:355-359
7. Fielding S, Wilker J, Hynes M, Szewczak M, Novick WJ Jr,Lai H: A comparison of clonidine with morphine for anti-nociceptive and antiwithdrawal actions. J Pharmacol ExpTher 1978^07:899-905
8. Gold MS, Redmond DE Jr, Kleber HD: Clonidine in opiatewithdrawal. Lancet I978;2:599-6O2
9. Ernsberger P, Meeley MP, Mann JJ, Reis DJ: Clonidinebinds to imidazole binding sites as well as aradrenoceptorsin the ventrolateral medulla. Eur J Pharmacol 1987;134:1-13
10. Emsberger P, Arango V, Meeley MP, Reis DJ: Selectivebinding of an endogenous clonidine-like substance to imida-
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Meeley et al Antibodies to p-Aminoclonldine Recognize CDS 351
zole binding sites and distribution of these sites in medullaoblongata of the rat (abstract). Soc Neurosci Abstr 1986;12:1334
11. BriccaG, Dontenwill M, Molines A, FeldmanJ, Belcourt A,Bousquet P: Evidence for the existence of a homogenouspopulation of imidazoline receptors in the human brainstem.Eur J Pharmacol 1988; 150:401-402
12. Emsberger P, Giuliano R, Willette RN, Granata AR, ReisDJ: Hypertensive action of clonidine analogs correlates withbinding affinity at imidazole and not aradrenergic receptorsin the rostral ventrolateral medulla. J Hypertens 1988;6(suppl 4):S554-S557
13. Atlas D, Burstein Y: Isolation and partial purification of aclonidine-displacing endogenous brain substance. Eur J Bio-chem 1984;144:287-293
14. Meeley MP, Emsberger PR, Granata AR, Reis DJ: Anendogenous clonidine-displacing substance from bovine brain:Receptor binding and hypotensive actions in the ventrolat-eral medulla. Life Sci 1986,38:1119-1126
15. Emsberger P, Meeley MP, Reis DJ: An endogenous clonidine-like substance binds preferentially to imidazole binding sitesin the ventrolateral medulla labeled by 3H-p-aminoclonidine.J Hypertens 1986;4(suppl 5):S1O9-S111
16. Emsberger P, Meeley MP, Reis DJ: An endogenous sub-stance with clonidine-like properties: Selective binding toimidazole sites in the ventrolateral medulla. Brain Res 1988;441:309-318
17. Bousquet P, Feldman J, Atlas D: An endogenous, non-catecholamine clonidine antagonist increases mean arterialblood pressure. Eur J Pharmacol 1986;124:167-170
18. Diamant S, Atlas D: An endogenous brain substance, CDS(clonidine-displacing substance), inhibits the twitch responseof rat vas deferens. Biochem Biophys Res Comm 1986;134:184-190
19. Diamant S, Eldor A, Atlas D: A low molecular weight brainsubstance interacts, similarly to clonidine, with a2-adrenoceptors of human platelets. Eur J Pharmacol 1987;144:247-255
20. Felsen D, Emsberger P, Meeley MP, Reis DJ: Clonidine-displacing substance is biologically active on smooth mus-cle. Eur J Pharmacol 1987;142:453-455
21. Meeley MP, Hensley ML, McCauley PM, Emsberger P,Reis DJ: Clonidine-displacing substance is present in periph-eral tissues of the rat. Am J Hypertens 1988 (in press)
22. Kreisberg GA, Diamant S, Diamant YZ, Atlas D: Raisedlevels of an endogenous nonadrenergic substance in theserum of pregnancy-induced hypertension patients. Isr JMedSci 1987 ;23:1194-1197
23. Rockson S, Homey CJ, Haber E: Anti-alprenolol antibodiesin the rabbit: A new probe for the study of 0-adrenergicreceptor interaction. Circ Res 1980;46:808-813
24. Graham RM, Hess H-J, Haber E, Homey CJ: Antibodies tothe alpha,- and alpharselective antagonists prazosin andyohimbine as probes of the alpha-adrenergic binding sites.Hypertension 1982;4(suppl II):II-183-II-187
25. Gainer MW, Nathanson NM: Recognition of muscarinicacetylcholine receptor ligands by monoclonal antibodiesagainst propylbenzilylcholine mustard. Biochem Pharamcol1986;35:1209-1212
26. Sherman MA, Linthicum DS, Bolger MB: Haloperidol bind-ing to monoclonal antibodies: Conformational analysis andrelationships to D-2 receptor binding. Mol Pharmacol 1986;29:589-598
27. Meeley MP, Towle AC, Emsberger P, Reis DJ: A specificantiserum recognizes clonidine'-displacing substance: Impli-cations for the structure of the brain's own clonidine.Neurosci Lett 1988;84:84-90
28. Sangameswaran L, deBlas AL: Demonstration ofbenzodiazepine-like molecules in the mammalian brain witha monoclonal antibody to benzodiazepines. Proc Nail AcadSci USA 1985;82:5560-5564
29. Bradford M: A rapid and sensitive method for the quantifi-cation of microgram quantities of protein utilizing the prin-ciple of protein-dye binding. Analyt Biochem 1976;72:248-254
30. Read SM, Northcote DH: Minimization of variation in theresponse to different proteins of the Coomassie Blue Gdye-binding assay for protein. Analyt Biochem 1981; 116:53-64
31. McPherson GA: A practical computer-based approach to theanalysis of radioligand binding experiments. Comp ProgBiomed 1983;17:107-114
32. Munson PJ, Rodbard D: LIGAND: A versatile computerizedapproach for characterization of ligand binding systems.Anal Biochem 198O;107:22O-239
33. Davis GC, Hein MB, Chapman DA: Evaluation of immu-nosorbents for the analysis of small molecules. Isolation andpurification of cytokinins. J Chromatog 1986;366:171-189
34. Milner TA, Pickel VM: Ultrastructural localization andafferent sources of substance P in the rat parabrachialregion. Neuroscience 1986; 17:687-707
35. Jarrott B, Spector S: Disposition of clonidine in rats asdetermined by radioimmunoassay. J Pharmacol Exp Ther1978 ;207:195-207
36. Rouot D, Leclerc G, Wermuth C-G: Structure de la cloni-dine. Chim Ther 1973,5:545-551
37. Meerman-van Benthem CM, van der Meer K, Mulden JJC,Timmermans PBMWM, van Zwieten PA: Clonidine base:Evidence for conjugation of both ring systems. Mol Phar-macol 1975 ;11:667-670
38. Wermuth CG, Schwartz J, Leclerc G, Gamier JP, Rouot B:Conformation de la clonidine et hypotheses sur son interac-tion avec un recepteur a-adrenergique. Chim Ther 1973;1:115-116.
39. Timmermans PBMWM, Schoop A, Kwa H, van ZwietenPA: Characterization of a-adrenoceptors participating in thecentral hypotensive and sedative effects of clonidine usingyohimbine, rauwolscine, and corynanthine. Eur J Pharma-col 1981;70:7-15
40. LeClerc G, Rouot B, Schwartz J, Velly J, Wermuth CG:Studies on some para-substituted clonidine derivatives thatexhibit a-adrenoceptor stimulant activity. Br J Pharmacoll980;71:5-9
41. Kobinger W, Pichler L: Centrally induced reduction in sym-pathetic tone—a postsynaptic a-adrenoceptor-stimulatingaction of imidazolines. Eur J Pharmacol 1976;4O:311-320
42. Savola J-M, Ruskoaho H, Puurunen J, Karki NT: Cardio-vascular action of detomidine, a sedative and analgesicimidazole derivative with a-agonistic properties. Eur J Phar-macol 1985;118:69-76
43. Savola J-M, Ruskoaho H, Salonen JS, Puurunen J, Kar-jalainen A, Kurkela K, Karki NT: Cardiovascular andsedative a-adrenoceptor effects of detomidine-like arylalkylimidazoles and associated derivatives. Arzneim Forsch 1988;38:29-35
44. Bylund DB, U'Prichard DC: Characterization of o,- anda2-adrenergic receptors. Int Rev Neurobiol 1983;24:343-431
45. Atlas D, Diamant S, Fales HM, Pannell L: The brain's ownclonidine: Purification and characterization of endogenousclonidine displacing substance from brain. J CardiovascPharmacol 1987;10(suppl 12):SI22-S127
46. Versteeg DHG, van der Gugten J, de Jong W, Palkovits M:Regional concentrations of noradrenaline and dopamine inrat brain. Brain Res 1976;113:563-574
47. Anton AH, Sayre DF: A study of the factors affecting thealuminum oxide-trihydroxyindole procedure for the analysisof catecholamines. J Pharmacol Exp Ther 1962;138:360-375
48. Steinbusch HWM, Mulder AH: Localization and projectionsof histamine-immunoreactive neurons in the central nervoussystem of the rat, in Ganellin CR, Schwartz JR (eds):Frontiers in Histamine Research. Oxford, Pergamon, 1985,pp 119-136
49. Iversen LL, Iversen SD, Snyder SH (eds): Handbook ofPsychopharmacology I, Section IV: Basic Neuropharmacol-ogy, vol 16, Neuropeptides. New York, Plenum Press, 1983
KEY WORDS • clonidine-displacing substance • clonidine •polyclonal antiserum • ligand specificity • imidazole receptors• cardiovascular control
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from

M P Meeley, A C Towle, P Ernsberger, L K Char, P M McCauley and D J ReisClonidine-specific antisera recognize an endogenous clonidine-displacing substance in brain.
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 1989 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/01.HYP.13.4.341
1989;13:341-351Hypertension.
http://hyper.ahajournals.org/content/13/4/341World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer process is available in the
Request Permissions in the middle column of the Web page under Services. Further information about thisOffice. Once the online version of the published article for which permission is being requested is located, click
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertension Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on June 22, 2018http://hyper.ahajournals.org/
Dow
nloaded from