chemotaxis in yersinia arthritis
TRANSCRIPT

655
CHEMOTAXIS IN YERSINIA ARTHRITIS
In Vitro Stimulation of Neutrophil Migration by HLA-B27 Positive and Negative Sera
HEIKKI REPO, MARJATTA LEIRISALO, ANJA TIILIKAINEN, and OSSI LAITINEN
Chemotactic and chemokinetic migration of poly- morphonuclear leukocytes in sera from patients with previous yersinia arthritis and from healthy subjects with or without HLA-B27 were studied by the leading front method. Irrespective of yersinia arthritis, zymo- san-activated sera from subjects who were HLA-B27 positive were significantly more chemokinetic than were HLA-B27 negative zymosan-activated sera from healthy control subjects. The chemotactic activities of zymosan-activated sera that were HLA-B27 positive or negative, as determined by chemotactic increments, were much the same. The results suggest that zymosan- activated serum that is HLA-B27 positive stimulates random migration of polymorphonuciear leukocytes, but not their directional migration, more than does HLA-B27 negative serum that has been zymosan acti- vated. This may contribute to accumulation of polymor- phonuclear leukocytes at the site of inflammation in vivo and thereby to inflammatory symptoms in yersinia arthritis patients with HLA-B27.
From the Department of Bacteriology and Immunology, University of Helsinki, and the Second Department of Medicine, Helsinki University Central Hospital, Helsinki, Finland.
Supported by grants from the Finnish National Council for Medical Sciences and from the Paulo Foundation.
Heikki Repo, MD: Postdoctoral fellow, Department of Bacteriology and Immunology, University of Helsinki; Majatta Leirisalo, MD: Lecturer of Medicine, Second Department of Medi- cine, Helsinki University Central Hospital; Anja Tiilikainen, MD: Professor, Department of Medical Microbiology, University of Oulu, Oulu; Ossi Laitinen, MD: Professor, Department of Rheuma- tology, University of Tampere, Tampere.
Address reprint requests to Heikki Repo, MD, Department of Bacteriology and Immunology, University of Helsinki, Haart- maninkatu, 3, SF-00290 Helsinki 29, Finland.
Submitted for publication October 13, 1981; accepted in revised form January 4, 1982.
Reactive arthritis, a form of nonpurulent joint inflammation, can follow an enteric infection caused by Salmonella, Shigella, Campylobacter, or Yersinia enterocolitica, among others. Reactive enteroarthritis is associated with the histocompatibility antigen HLA-B27 ( I ) , and the majority of patients with yer- sinia arthritis have HLA-B27 (2 ,3) .
Clinical studies have indicated that the inflam- matory symptoms, such as arthritis, carditis, and iritis, are more prolonged and more frequent in HLA-B27 positive (B27+) patients with yersinia arthritis than in B27 negative (B27-l patients (2-4). The reason for the difference is not known. However, the finding that B27+ polymorphonuclear leukocytes show high chemo- taxis in vitro suggests that phagocyte function may play a role ( 5 ) in this phenomenon,
Accumulation of such leukocytes at the site of inflammation in vivo is the result of both the liberation of chemotactic agents, such as the complement-de- rived polypeptide C5a, and the responsiveness of the cells to the chemotactic stimulus. We studied the former aspect by testing the chemotactic and chemo- kinetic activities of sera from patients with previous yersinia arthritis and from healthy control subjects. The results indicated that, irrespective of yersinia arthritis, the chemokinetic migration of neutrophils in a membrane filter was significantly higher in B27' than in B27- zymosan-activated serum.
MATERIALS AND METHODS Patients. The diagnosis of yersinia arthritis was based
on a typical clinical picture, significantly elevated antibody titers for Y enterocolitica (serotype 3 or 9) with at least a 4- fold seroconversion during the course of the disease, and exclusion of other causes of arthritis (6,7). We HLA typed
Arthritis and Rheumatism, Vol. 25, No. 6 (June 1982)
656 REP0 ET AL
both the patients and the control subjects. who ranged in age from 20 to 45 years, using several antisera to 30 HLA A. B. and C antigens.
Sera. Samples of sera collected from B27' and B27- patients after complete recovery and from healthy control subjects were stored at -40°C. None of the subjects had any drug treatment at the time of bleeding. Zymosan-activated serum was prepared as described (8). and zymosan particles were removed by centrifugation. In the experiments indicat- ed, fresh serum, heated (30 minutes a t 56°C) serum, zymo- san-activated serum, and heated zymosan-activated serum were tested in parallel.
Cells. Heparinized blood (10 unitsiml, preservative- free heparin, Medica Co., Helsinki) was collected from healthy volunteers with blood group 0. Buffy coat cells were separated by dextran sedimentation, washed 3 times in Hanks' balanced salt solution, and resuspended in the cell medium at 1 .O x lo6 polymorphonuclear leukocytesiml.
Membrane filter assay. The leading front modifica- tion (9) of the Boyden chamber assay (10) was used accord- ing to the method of Wilkinson ( 1 I ) . Buffy coat cells contain- ing 2.0 x 10' neutrophils were placed on a 3-p pore-size membrane filter (Millipore Corp.). To evaluate both direc- tional migration (chemotaxis) and enhanced random migra- tion (chemokinesis), a checkerboard was made of zymosan- activated serum by varying its concentration (0.75, I .5, 3.0, 6.0, and 12.0%, v/v, in Hanks' balanced salt solution) on both sides of the filter. After the filters were incubated for 50 minutes a t 37°C in a mixture of 2% carbon dioxide in air, they were processed for microscopy. Distance of the cell front from the starting point was determined as an average of 5 microscopic fields (x400) of each of the triplicate filters. In the checkerboard assays, we calculated the theoretical esti- mates for the cells assumed to detect the absolute concentra- tion of zymosan-activated serum and not the gradient (9). Chemotactic responsiveness of neutrophils was evaluated by chemotactic increments:
Chemotactic increment (76) =
observed value ( p ) - theoretical estimate ( p )
theoretical estimate ( p ) x 100
Statistical analysis. The statistical significance of the difference between the groups was evaluated by the Stu- dent's t-test.
RESULTS In the preliminary experiments, chemokinetic
migration of neutrophils was studied simultaneously in fresh serum, heated serum, zymosan-activated serum, and heated zymosan-activated serum from B27+ and B27- healthy subjects without previous yersinia arthri- tis (A-). Two samples of sera, 1 from B27+ donors and the other from B27- donors, were divided on the same day into 4 portions. The first portion was used as fresh serum; the second was heated for 30 minutes at 56°C;
the third and fourth portions were mixed with zymo- san and incubated for 60 minutes at 37°C to prepare zymosan-activated serum, after which the fourth por- tion was heated as above.
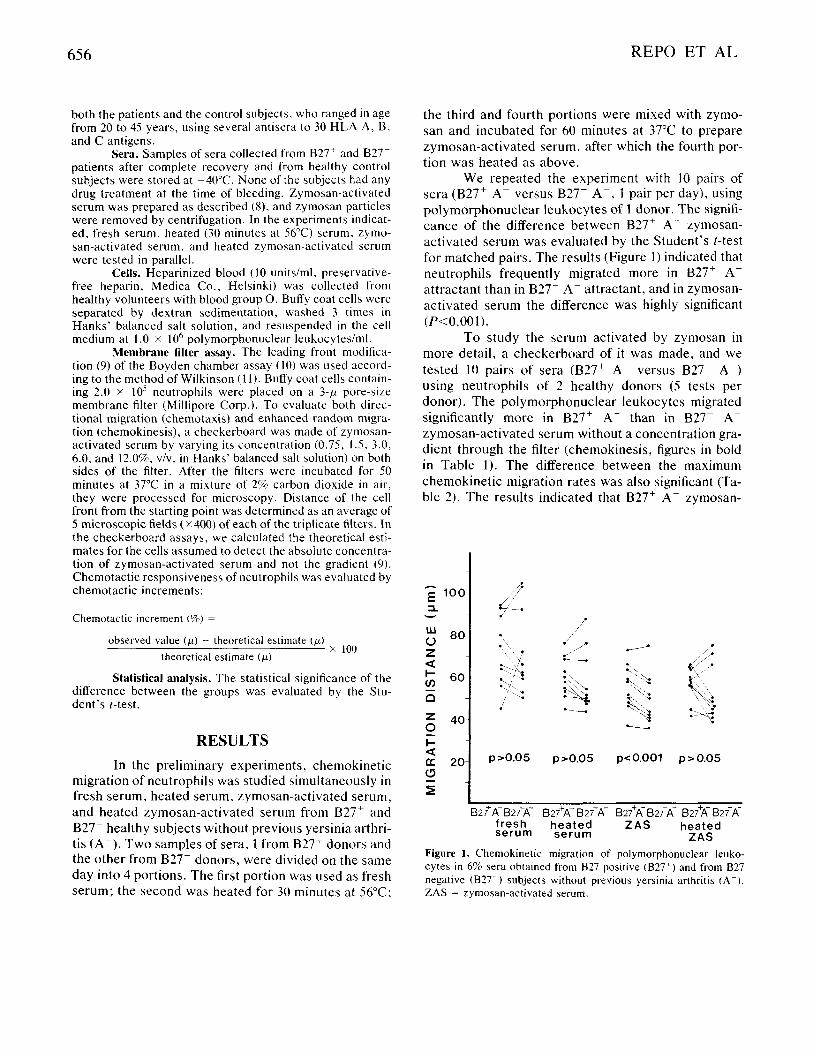
We repeated the experiment with 10 pairs of sera (B27+ A- versus B27- A - , I pair per day), using polymorphonuclear leukocytes of 1 donor. The signifi- cance of the difference between B27+ A- zymosan- activated serum was evaluated by the Student's t-test for matched pairs. The results (Figure I ) indicated that neutrophils frequently migrated more in B27+ A- attractant than in B27- A- attractant, and in zymosan- activated serum the difference was highly significant (P<O.001).
To study the serum activated by zymosan in more detail, a checkerboard of it was made, and we tested 10 pairs of sera (B27+ A- versus B27- A-) using neutrophils of 2 healthy donors (5 tests per donor). The polymorphonuclear leukocytes migrated significantly more in B27+ A- than in B27- A- zymosan-activated serum without a concentration gra- dient through the filter (chemokinesis, figures in bold in Table 1). The difference between the maximum chemokinetic migration rates was also significant (Ta- ble 2) . The results indicated that B27+ A- zymosan-
p< 0.001 p > 0.05
fresh heated ZAS heated Serum serum ZAS
Figure 1. Chemokinetic migration of polymorphonuclear leuko- cytes in 6% sera obtained from 827 positive (B27') and from B27 negative (B27T) subjects without previous yersinia arthritis (A-) . ZAS = zymosan-activated serum.
CHEMOTAXIS IN YERSINIA ARTHRITIS 657
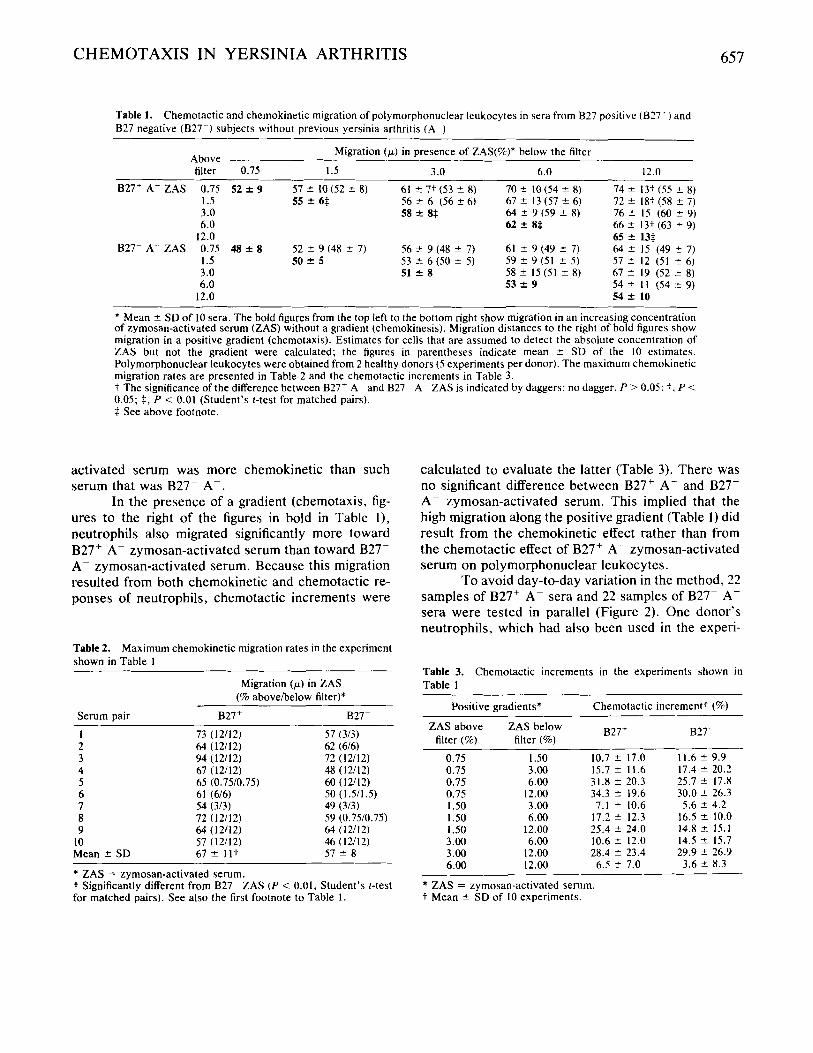
Table 1. Chemotactic and chemokinetic migration of polymorphonuclear leukocytes in sera from B27 positive (B27 ' ) and 827 negative (B27--) subjects without previous yersinia arthritis (A )
~
Migration (p) in presence of ZAS(%)* below the filter Above - filter 0.75 1.5 3.0 6.0 12.0
827' A- ZAS 0.75 52 f 9 57 ? 1 0 0 2 2 8) 61 f 7 t (53 f 8) 70 2 lO(54 f 8) 74 f 13t (55 c 8 ) 1.5 72 ? 18: (58 c 7) 3.0 76 f I5 (60 c 9) 6.0 62 2 8$ 66 ? 13t (63 f 9)
B27- A ZAS 0.75 48 f 8 52 f 9(48 f 7) 56 -e 9 (48 f 7) 61 f 9 (49 2 7) 64 2 I5 (49 t 7) 57 f 12 (51 5 6) I .5 50 f 5
3.0 51 f 8 5 8 ? 1 5 ( 5 1 ? 8 ) 6 7 f 1 9 ( 5 2 ? 8 ) 6.0 53 2 9 54 ? I 1 (54 c_ 9)
12.0 54 f 10
55 f 6$ 56 2 6 (56 2 6) 58 5 8$
67 2 13 (57 ? 6) 64 2 9 (59 ? 8)
12.0 65 2 13$
53 2 6 (50 ? 5 ) 59 2 9 (51 ? 5 )
* Mean 2 SD of 10 sera. The bold figures from the top left to the bottom right show migration in an increasing concentration of zymosan-activated serum (ZAS) without a gradient (chemokinesis). Migration distances to the right of bold figures show migration in a positive gradient (chemotaxis). Estimates for cells that are assumed to detect the absolute concentration of ZAS but not the gradient were calculated; the figures in parentheses indicate mean ? SD of the 10 estimates. Polymorphonuclear leukocytes were obtained from 2 healthy donors ( 5 experiments per donor). The maximum chemokinetic migration rates are presented in Table 2 and the chemotactic increments in Table 3. t The significance of the difference between B27' A and 827 0.05; $, P < 0.01 (Student's &test for matched pairs). $ See above footnote.
activated serum was more chemokinetic than such serum that was B27- A-.
In the presence of a gradient (chemotaxis, fig- ures to the right of the figures in bold in Table l) , neutrophils also migrated significantly more toward B27+ A- zymosan-activated serum than toward B27-- A- zymosan-activated serum. Because this migration resulted from both chemokinetic and chemotactic re- ponses of neutrophils, chemotactic increments were
Table 2. shown in Table I
Maximum chemokinetic migration rates in the experiment
Migration (p) in %AS (% abovebelow filter)*
Serum pair B27' 827-
I 73 (12112) 57 (313) 2 64 (12112) 62 (616) 3 94 (12112) 72 (12112) 4 67 (12112) 48 (12112) 5 65 (0.7510.75) 60 (12112) 6 61 (W6) 50 ( I S11.5) 7 54 (313) 49 (313) 8 72 (12112) 59 (0.75/0.75) 9 64 (12/12) 64 (12112)
10 57 (12/12) 46 (12112) Mean 2 SD 67 ? I l t
* ZAS = zymosan-activated serum. t Significantly different from B27 ZAS ( P < 0.01. Student's I-test for matched pairs). See also the first footnote to Table I .
57 ? 8 --
A- ZAS is indicated by daggers: no dagger. P > 0.05: t , P <
calculated to evaluate the latter (Table 3). There was no significant difference between B27' A- and B27- A- zymosan-activated serum. This implied that the high migration along the positive gradient (Table I ) did result from the chemokinetic effect rather than from the chemotactic effect of B27+ A-. zymosan-activated serum on polymorphonuclear leukocytes.
To avoid day-to-day variation in the method, 22 samples of B27+ A- sera and 22 samples of B27- A- sera were tested in parallel (Figure 2). One donor's neutrophils, which had also been used in the experi-
Table 3. Table 1
Chemotactic increments in the experiments shown in
Chemotactic incrementt (7%)
B27+ B27-
Positive gradients*
ZAS above ZAS below filter (5%) filter (%)
---
0.75 0.75 0.75 0.75 I SO 1.50 I S O 3.00 3.00 6.00
1 S O 3.00 6.00
12.00 3.00 6.00
12.00 6.00
12.00 12.00
10.7 ? 17.0 11.6 2 9.9 15.7 ? 11.6 17.4 ? 20.2 31.8 2 20.3 25.7 ? 17.8
7.1 ? 10.6 5.6 ? 4.2 17.2 f 12.3 16.5 2 10.0 25.4 ? 24.0 14.8 2 15.1
28.4 f 23.4 29.9 2 26.9
34.3 2 19.6 30.0 ? 26.3
10.6 2 12.0 14.5 2 15.7
6.5 ? 7.0 3.6 f 8.3
* ZAS = zymosan-activated serum. t Mean 2 SD of 10 experiments.

658 REP0 ET AL
100
80
- 5 60 aI 0 c m v) c
a .- 5 40 e 5
L
W .-
20
0
B27+A- B27-A- ZAS
Figure 2. Chemokinetic migration of polymorphonuclear feuko- cytes (PMN) in 6% zymosan-activated serum (ZAS) obtained from 22 B27 positive (B27+) and 22 B27 negative (B27-) subjects without previous yersinia arthritis (A-). Samples of ZAS were tested in parallel with PMN of 1 donor; each point indicates the mean of 15 microscopic fields of triplicate filters. The difference between B27' A- and B27- A- ZAS was significant (P < 0.05, Student's 1-test). Shaded areas denote mean 2 SE.
ments of Figure 1, migrated significantly more in B27+ A- than in B27- A- zymosan-activated seqm. To see whether the phenomenon was associated with yersinia arthritis and whether it could be demonstrated with cells of different donors, chemokinetic migration of neutrophils of 6 healthy volunteers were tested in 100 sera from subjects with or without previous yersinia arthritis (Figure 3). Irrespective of yersinia arthritis, B27+ zymosan-activated serum was significantly more chemokinetic than such serum that was B27T.
When the groups were combined on the basis of HLA-B27, chemokinetic migration of polymorphonu- clear leukocytes was significantly higher in B27+ (74 2 0.9p, mean ? SE, number of sera = 60) than in B27- zymosan-activated serum (69 5 I .2 p, number of sera = 40, P < 0.01). The results indicated that the high
chemokinetic activity was associated with HLA-B27 antigen. Although the B27+ A+ zymosan-activated serum was more chemokinetic than that which was B27+ A-, the difference was not significant (Figure 3). Furthermore, migration in B27- A+ serum activated by zymosan was very similar to that in B27- A- zymosan-activated serum, suggesting that the high chemokinetic activity was not associated with the previous yersinia arthritis.
To see whether the high chemokinetic activity and the low chemokinetic activity were reproducible, we selected, on the basis of the results in Figure 3, 5 B27+ subjects with high chemokinetic activity and 5 B27- subjects with low chemokinetic activity. The subjects were rebled, and samples of sera were com- bined to make B27' pooled serum and B27- pooled serum. Migration of polymorphonuclear leukocytes was then tested in fresh pooled serum, in heated
_. .- t-p<0.05+
n
e c I- pc0.024 0 .-
4-
I-. peo.01- M .- 2 t-pco.1-i
I B27+A+ B27-Af B27+A- B27-A-
N=34 N=14 N=26 N=26 U S
Figure 3. Chemokinetic migration of polymorphonuclear leuko- cytes (PMN) in 6% zymosan-activated serum (ZAS) obtained from B27 positive (B27+) and from B27 negative (B27-) subjects with previous yersinia arthritis (A+) or without it (A-). N = number of samples of ZAS. The 100 samples of ZAS were tested in parallel with PMN of 1 donor, and the experiment was repeated with cells of 5 other donors. Each point indicates the mean migration of PMN of 6 donors. The significance of the difference between the groups was evaluated by Student's t-test. Shaded areas denote mean 2 SE.

CHEMOTAXIS IN YERSINIA ARTHRITIS 659
pooled serum, in zymosan-activated serum, and in such serum that had been heated (Table 4).
The results indicated that the differences be- tween fresh B27+ pooled serum and fresh B27- pooled serum and between B27+ and B27- zymosan-activat- ed serum were significant.
Heat treatment (30 minutes at 56°C) markedly decreased migration of neutrophils in both sera. Heat- ed B27+ serum was more chemokinetic than heated B27- serum (P < 0.05), whereas in heated zymosan- activated serum (60 minutes at 37°C followed by 30 minutes at 56"C), the corresponding difference was not significant (P > 0.05). The results suggested that the difference between B27+ sera and B27- sera has its origin in heat-labile chemokinetic factor(s) in fresh serum.
DISCUSSION The results in the present study show that,
irrespective of previous yersinia arthritis, B27+ zymo- san-activated serum stimulated random migration of neutrophils in the membrane filter more than did such serum that was B27T. Normal human serum contains chemotactic factors, chemokinetic factors, cell-direct-
ed inhibitors of chemotaxis, and chemotactic-factor inactivators. The effect of serum on neutrophils results from the balance between stimulators and inhibitors of cell migration. The finding that the chemotactic re- sponses of neutrophils to B27+ and B27- zymosan- activated serum, as determined by the chemotactic increments (Table 3), were much the same suggests that the balance involved was between chemokinetic factors and cell-directed inhibitors rather than be- tween chemotactic factors and their inactivators. This probably means that the difference between B27+ and B27- zymosan-activated serum does not have its origin in the complement-derived chemotactic pep- tides, such as CSa, which show both chemotactic and chemokinetic activities, whereas the neutrophil re- sponses to the B27+ sera were chemokinetic without a detectable enhanced chemotactic component (Tables I and 3).
With the use of a membrane filter assay, a leukokinesis-enhancing factor in normal human serum has been described (12). This factor is heat labile, it stimulates both random migration and phagocytic function of neutrophils, and reduced levels of it were detected in 3 patients with recurrent bacterial infec- tions. Furthermore, increased levels of activity similar
Table 4. Chemokinetic migration of polymorphonuclear leukocytes in pooled sera from B27 positive (B27+) and B27 negative (B27-) subjects*
Migration (p)t in 3% pooled serum on both sides of filter
Fresh sera Heated sera ZAS Heated ZAS Neutrophil donor B27+ B27- B27' B27- B27+ B27- B27+ B27-
1 113 2 18 85 t 11 52 t 8 51 t 16 63 t 10 50 t 9 62 t 12 46 t 7 2 132 ? 13 126 t 14 62 2 10 47 t 8 7 7 t 1 1 57 t 10 72 t 7 64 f 6 3 132 2 9 92 2 17 61 f 4 45 t 7 75 2 10 48 t 6 54 t 10 51 t 8 4 93 t 11 53 2 10 47 ? 5 42 t 4 51 t 7 47 t 4 41 ? 4 43 5 5 5 114 2 19 75 t 25 50 t 5 45 2 6 57 ? 6 45 t 6 50 t 5 40 2 4 6 128 ? 12 106 t 32 43 t 6 51 t 9 61 t 10 52 t 6 54 f 9 54 ? 10 7 65 f 16 51 2 5 41 ? 8 42 t 4 45 t 4 45 t 7 48 2 7 35 t 6 8 85 5 17 45 2 11 51 t 8 40 t 5 47 t 4 36 t 6 49 2 10 49 2 5 9 72 t 13 69 2 4 43 ? 5 36 t 7 48 t 5 29 t 4 47 t 4 46 2 3
10 86 ? 9 59 t 11 57 " 4 54 t 5 46 2 6 46 ? 4 42 f 10 49 2 7
Mean t SD 102 t 251 76 t 26 51 5 8$ 45 ? 6 57 t 128 46 t 8 52 t 9 48 t 8
* On the basis of the results shown in Figure 3 ,5 B27+ subjects with highly chemokinetic sera (distances of migration in the experiments shown in Figure 1 were 76,79, and l l p f o r 3 B27+ A- subjects, as well as 82 and 83p for 2 B27+ A + subjects) and 5 healthy B27- subjects with low che- mokinetic sera (distances of migration 57, 62,62,64, and 68p) were contacted for rebleeding. In 1 of the 2 subjects, the diagnosis of yersinia ar- thritis was made in 1974 and in the other in 1977; both subjects had completely recovered from the disease. Samples of sera were combined to make B27+ pooled serum and B27- pooled serum. Fresh sera, heated sera, zymosan-activated sera (ZAS), and heated ZAS were tested on the same day in parallel with neutrophils of 10 healthy donors. t Mean 2 SD of 15 microscopic fields (5 fields per filter). $ The difference between B27' and B27- pooled sera was significant at the level of P < 0.05 by the Student's t-test for matched pairs. 9 The difference between B27' and B27- pooled sera was significant at the level of P < 0.01 by the Student's t-test for matched pairs. TI The difference between B27+ and B27- pooled sera was significant at the level of P < 0.001 by the Student's t-test for matched pairs.

660 REP0 ET AL
to that caused by leukokinesis-enhancing factor were detected in a patient with episodes of sterile pyarthro- sis and pyoderma gangrenosum; these levels may have contributed to the patient’s increased inflammatory response (13). In this context, it is tempting to pro- pound that B27+ zymosan-activated serum contains more of this factor than does such serum that is B27T.
The similarity of our serum factor(s) to the leukokinesis-enhancing one is supported by the find- ings that, first, B27+ zymosan-activated serum stimu- lated random migration rather than directional migra- tion of neutrophils and, second, the difference in chemokinetic activity between B27+ serum and B27- serum was evident in fresh sera and in zymosan- activated serum but markedly lost in heated sera and in heated sera that had been zymosan activated (Table 4). Conversely, in the preliminary experiments the difference between B27+ and B27- subjects was sig- nificant in the zymosan-activated serum but not in the corresponding samples of fresh sera (Figure 1). This is different from the leukokinesis-enhancing factor, which can be demonstrated without preincubation of serum with zymosan (12). Thus, the present results did not show unambiguously whether the chemokinetic activity was associated with the leukokinesis-enhanc- ing factor or other factors. In future, detailed studies on the physicochemical properties of the serum fac- tor(s) in the present study will be needed to see their relationship to the leukokinesis-enhancing factor, to human serum albumin that is chemokinetic rather than chemotactic (14), and also to cell-directed inhibitors (12).
Neutrophil migration through a 3-p pore-size membrane filter is affected by several factors, such as deformability (15-18) of the cells and their adhesive- ness to the filter substance (19). Immunoglobulin G is one of the cell-directed inhibitors in normal human serum and increases adherence of neutrophils to nylon fibers (20), whereas chemotactic factors increase de- formability of these cells (21). The modes of action of the leukokinesis-enhancing factor and the chemokine- tic factors in the present study are not known, but they may increase deformability of the neutrophils and/or modify their adhesiveness to be more appropriate for cell migration. This would increase the distances of both random migration and directional migration in the membrane filter but not the directional responses of the leukocytes, as determined by chemotactic incre- ments. There was a remarkable overlapping in the results obtained with the B27+ and the B27- zymosan- activated sera (Figures 2 and 3), suggesting that the
high chemokinetic activity is not unique to the B27 haplotype. At present, the reason for the difference in the chemokinetic activity between B27+ and B27- zymosan-activated serum is not known, and it may be that the HLA-B27 antigen, itself, or some factor(s) associated with this antigen are responsible.
We recently found that B27+ neutrophils show high chemotaxis in the membrane filter (5 ) . Although the cells were thoroughly washed, serum factors could still have remained on the cell surfaces. However, the chemotactic increments of B27+ neutrophils were significantly higher than those of B27- ones (9, whereas in B27+ and B27- zymosan-activated serum, they were much the same (Table 3), suggesting that the cell-associated phenomenon and the serum-associated phenomenon were distinct.
At present, the biologic implications of the results in our present study remain uncertain. Poly- morphonuclear leukocytes are considered to accentu- ate inflammation by releasing lysosomal enzymes dur- ing phagocytosis of foreign invaders or damaged tissue components (22). Such an enzyme release also occurs when neutrophils are exposed to chemotactic agents in vitro (23). Furthermore, neutrophils play an important role in the control of vascular permeability in inflam- mation (24). Both the high chemokinetic activity of B27+ zymosan-activated serum demonstrated in the present study and the high responsiveness of B27+ neutrophils to serum thus activated (5 ) may contribute to accumulation of neutrophils at the site of inflamma- tion in vivo, and, thereby, to the inflammatory symp- toms in B27+ patients with yersinia arthritis.
ACKNOWLEDGMENT The skillful technical assistance of Mrs. Eine Viro-
lainen is gratefully acknowledged.
REFERENCES 1. Brewerton DA: HLA-B27 and the inheritance of sus-
ceptibility to rheumatic disease. Arthritis Rheum
2. Aho K, Ahvonen P, Lassus A, Sievers K, Tiilikainen A: HL-A 27 in reactive arthritis: a study of yersinia arthritis and Reiter’s disease. Arthritis Rheum 17521-526, 1974
3. Laitinen 0, Leirisalo M, Skylv G: Relation between HLA-B27 and clinical features in patients with yersinia arthritis. Arthritis Rheum 20: 1121-1 124, 1977
4. Aho K, Ahvonen P, Laitinen 0, Leirisalo M: Arthritis associated with Yersinia enterocolitica infection, Yer- sinia Enterocolitica. Edited by EJ Bottone. Boca Raton, Florida, CRC Press Inc., 1981, pp 113-124
19: 656-668, 1976

CHEMOTAXIS IN YERSINIA ARTHRITIS 66 1
5. Leirisalo M, Rep0 H, Tiilikainen A, Kosunen TU, Laitinen 0: Chemotaxis in yersinia arthritis: HLA-B27 positive neutrophils show high stimulated motility in vitro. Arthritis Rheum 23:1036-1044, 1980
6. Ahvonen P, Severs K, Aho K: Arthritis associated with Yersinia enterocolitica infection. Acta Rheum Scand
7. Ahvonen P: Human yersiniosis in Finland. 11. Clinical features. Ann Clin Res 4:39-48, 1972
8. Rep0 H: Leukocyte migration agarose test for the as- sessment of human neutrophil chemotaxis. I. Effects of environmental factors on neutrophil migration under agarose. Scand J Immunol6:203-209, 1977
9. Zigmond SH, Hirsch JG: Leukocyte locomotion and chemotaxis: new methods for evaluation, and demon- stration of a cell-derived chemotactic factor. J Exp Med
10. Boyden S: The chemotactic effect of mixtures of anti- body and antigen on polymorphonuclear leucocytes. J Exp Med 115:453-466, 1962
11. Wilkinson PC: Chemotaxis and Inflammation. Edin- burgh, Churchill Livingstone, 1974, pp 168-172
12. Maderazo E, Woronick CL, Ward PA: Leukokinesis- enhancing factor in human serum: partial characteriza- tion and relationship to disorders of leukocyte migra- tion. Clin Immunol Immunopathol 12:382-395, 1979
13. Jacobs JC, Goetzl EJ: “Streaking leukocyte factor,” arthritis, and pyoderma gangrenosum. Pediatrics 56:570-578, 1975
14. Keller HU, Wissler JH, Hess MW, Cottier H: Distinct chemokinetic and chemotactic responses in neutrophil granulocytes. Eur J Immunol 8:l-7, 1978
15. Miller ME: Developmental maturation of human neutro- phi1 motility and its relationship to membrane deforma- bility, The Phagocytic Cell in Host Resistance. Edited
15 : 232-253, 1969
137~387-4 10, 1973
by JA Bellanti, DH Dayton. New York, Raven Press,
16. Rep0 H, Vuopio P, Leirisalo M, Jansson S-E, Kosunen TU: Impaired neutrophil chemotaxis in Pelger-Huet anomaly. Clin Exp Immunol 36:326-333, 1979
17. Miller ME: Cell elastimetry in the study of normal and abnormal movement of human neutrophils. Clin Im- rnunol Immunopathol 14:503-510, 1979
18. Rep0 H, Jokipii AMM, Leirisalo M, Kosunen TU: Leucocyte motility in the newborn: determination of spontaneous movement is essential in the in vitro assess- ment of neutrophil chemotaxis. Clin Exp Immunol
19. Wilkinson PC, Lackie JM: The adhesion, migration and chemotaxis of leucocytes in inflammation. Curr Top Pathol 68:47-88, 1979
20. Woronick LC, Malnick J, Maderazo EG: Cell-directed inhibitor of human leukocyte locomotion: identification of immunoglobulin G as a cell-directed inhibitor in normal human serum. J Lab Clin Med 98:58-67, 1981
21. Kawaoka EJ, Miller ME, Cheung ATW: Chemotactic factor-induced deformability of human polymorphonu- clear leukocytes. J Clin Immunol 1:41-44, 1981
22. Wasi S, Movat H: Phlogistic substances in neutrophil leukocyte lysosomes: their possible role in vivo and their in vitro properties. Curr Top Pathol 68:213-237, 1979
23. Showell HJ, Freer RJ, Zigmond SH, Schiffman S, Aswankumar S, Corcoran B, Becker EL: The structure activity relations of synthetic peptides as chemotactic factors and inducers of lysosomal enzyme secretion for neutrophils. J Exp Med 143:1154-1169, 1976
24. Wedmore CV, Williams TJ: Control of vascular perme- ability by polymorphonuclear leukocytes in inflamma- tion. Nature 289:646-650, 1981
1975, pp 295-302
40:620-626, 1980