characterization of rabbit lactate dehydrogenase …characterization of rabbit lactate...
TRANSCRIPT

THE JOURNAL 0 1989 by The American Society for Biochemistry and Molecular Biology,
OF BIOLOGICAL CHEMISTRY Inc.
Vol. 264, No. 7, Issue of March 5, pp. 4076-4081,1989 Printed in U.S.A.
Characterization of Rabbit Lactate Dehydrogenase" and Lactate Dehydrogenase-H cDNAs CONTROL OF LACTATE DEHYDROGENASE EXPRESSION IN RABBIT MUSCLE*
(Received for publication, July 7, 1988)
C. Sass, M. Briand, S. Benslimane, M. Renaud, and Y. BriandS From the Laboratoire de Biochimie, Uniuersite Blaise Pascal-Clermont-Ferrand, F 631 77 Aubiere Cedex, France
Two cDNA clones were isolated, one corresponding to the mRNA coding for lactate dehydrogenase" (LDH-M), the other to the mRNA coding for lactate dehydrogenase-H (LDH-H). The cDNA inserts consist of the entire open reading frame for LDH-M and a partial sequence, from amino acid 117 to 332, for LDH-H. Using these two clones as probes we demon- strate that: (a) the abundance of mRNA is muscle-type dependent; (b) the ratio M/H subunit for protein and mRNA is well related in the muscles studied; and (c) the M + H mRNA level is not relative to the total LDH activity.
Lactic dehydrogenase (LDH),' which is involved in the final step of anaerobic glycolysis, consists of a system of five tetrameric isoenzymes composed of the random association of two subunits (H and M) that are encoded by different genes (1). A sperm-specific isoenzyme encoded by a third gene is found only in testes and spermatozoa (2). The expression of these genes is tissue-specific and shows developmental regulation (3). It has been shown that in muscle, type I fibers are rich in the H subunit, whereas in type I1 fibers the M subunit is predominant (4). We previously demonstrated a clear correlation between fiber type and the percentage LDH- M or LDH-H subunit in rabbit muscle (5).
In order to study the developmental regulation of the mus- cle-specific expression of LDH genes, we isolated and identi- fied the cDNA for the M and H subunits of rabbit LDH. The isolation was made from two cDNA libraries: one constructed from rabbit heart, in which 98% of the LDH isoenzyme is of the H-form, the other from rabbit psoas major muscle in which 93% of the LDH isoenzyme is of M-form.
The complete amino acid sequence of rabbit LDH-M and the partial amino acid sequence of rabbit LDH-H was deter- mined by structural analyses of the cDNAs.
These two clones were used to analyze the expression of LDH genes in muscles. The results show that the expression
* This work was supported by grants from the Institut National de la Recherche Agronomique, from the Conseil Regional Auvergne, and from Association FranCaise contre les Myopathies. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "aduer- tisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in this paper has been submitted to the GenBankTM/EMBL Data Bank with accession number($ 504595.
$To whom correspondence and reprint requests should be ad- dressed.
The abbreviations used are: LDH, lactate dehydrogenase; bp, base pairs; kb, kilobase pairs.
of the LDH isoenzyme is muscle-type specific, that the per- centage of "subunit protein is similar to the percentage of LDH-M mRNA in the studied muscle, but that there is no relation between total activity and M + H mRNA content suggesting different levels of regulation.
MATERIALS AND METHODS
Animals and Muscles-Heart, psoas major and semimembranosus proprius muscle were excised from adult New Zealand rabbits and immediately extracted or frozen in liquid nitrogen. Protein and RNA determinations were done on the same samples.
LDH Actiuity and Isoenzyme Determination-LDH activity was assayed according to Briand et al. (6) in 50 mM triethanolamine buffer, pH 7.3, in the presence of 5 mM pyruvate and 0.234 mM NADH + H+.
Lactic dehydrogenase isoenzymes were separated by polyacryl- amide gel electrophoresis according to the procedure of Fritz et al. (7). The gels were stained at 25 "C for 30 min in a solution of the following composition: DL-lithium lactate, 141 mg; NAD+, 29 mg; nitro blue tetrazolium, 15 mg; phenazine methosulfate, 0.6 mg; sodium glycinate, 0.05 M pH 10, 70 ml.
The gels were scanned in an integrating densitometer (LKB) for the determination of isoenzyme profiles and the percentage of M and H subunit.
RNA Preparation, Northern Blot, and Dot Spots-Total RNA was extracted from the rabbit muscles by modifying the guanidinium isothiocyanate procedure of Cathala et al. (8). RNA poly(A+) was selected through oligo(dT)-cellulose chromatography as described (9).
Northern blots of RNA samples, electrophoresed on denaturing 1% agarose gels (IO), were made on genescreen membranes (Du Pont- New England Nuclear) and hybridized using nick-translated probes as specified by the supplier.
RNA samples for dot-spots were denatured for 15 min at 60 "C in 7.5% formaldehyde, 10 X SSC (1 X SSC = 0.15 M NaCl, 0.015 M sodium citrate) and chilled in ice. The RNAs were then filtered through a 10 X SSC prewetted nitrocellulose membrane by using the Bethesda Research Laboratories hybridot system. DNA samples used as standard hybridization probes were denaturated for 30 min in 0.1 N NaOH. The nitrocellulose membranes were hybridized with probe labels using random oligonucleotides as primers (Boehringer Mann- heim).
Construction of the cDNA Libraries-Double strand cDNAs were synthesized from rabbit psoas and heart poly(A+) RNA as described by Lapeyre and Amalric (11). Double strand cDNA cloning in XGTll bacteriophage, packaging in uitro, and amplification were carried out according to Huynh et al. (12).
Identification of Rabbit LDH-M and LDH-H-The psoas XGTll library was plated at high density and screened on Plaquescreen membranes (Du Pont-New England Nuclear) as described by the manufacturer, using the nick-translated purified insert from a rat LDH-M cDNA supplied by Dr. L. M. Matrisian (13).
The heart XGTll library was screened on cellulose membranes with sheep specific polyclonal anti rabbit LDH-H sera. The screening procedure was optimized by using the cross-dot system (14). To reduce the background, antibodies were first absorbed with nitrocellulose immobilized protein from XGTll Escherichia coli Y 1090 lysed. POS- itive clones from the two cDNA libraries were prepared according to Maniatis et al. (15). The longest LDH-M and LDH-H cDNAs were
4076

Rabbit Lactate Dehydrogenase Isoenzyme cDNAs 4077
- - - I
- = = - - - - c - - - 6
”
FIG. 1. Sequencing strategy used : 2 E E u - c
x m 4 - 0- . u r n ; E - In- =:
for rabbit LDH-M cDNA and partial : 4 restriction endonuclease map. Ar- b 4 rows indicate the direction and length of 4 each sequencing run.
1 1
100 b o
subcloned in pGEM blue vector (Promega Biotech).
method (16) in Ml3mplO or M13mpll. The DNAs were sequenced by the dideoxy chain termination
RESULTS AND DISCUSSION
Characterization of a LDH-M cDNA from Rabbit Psoas Major Muscle-Twelve positive clones were obtained from 5 X lo5 plaques from the rabbit psoas muscle cDNA library. The longest (pGRTM12) showed an insert of approximately 1600 bp. Sequencing of the cDNA was done according to the strategy outlined in Fig. 1.
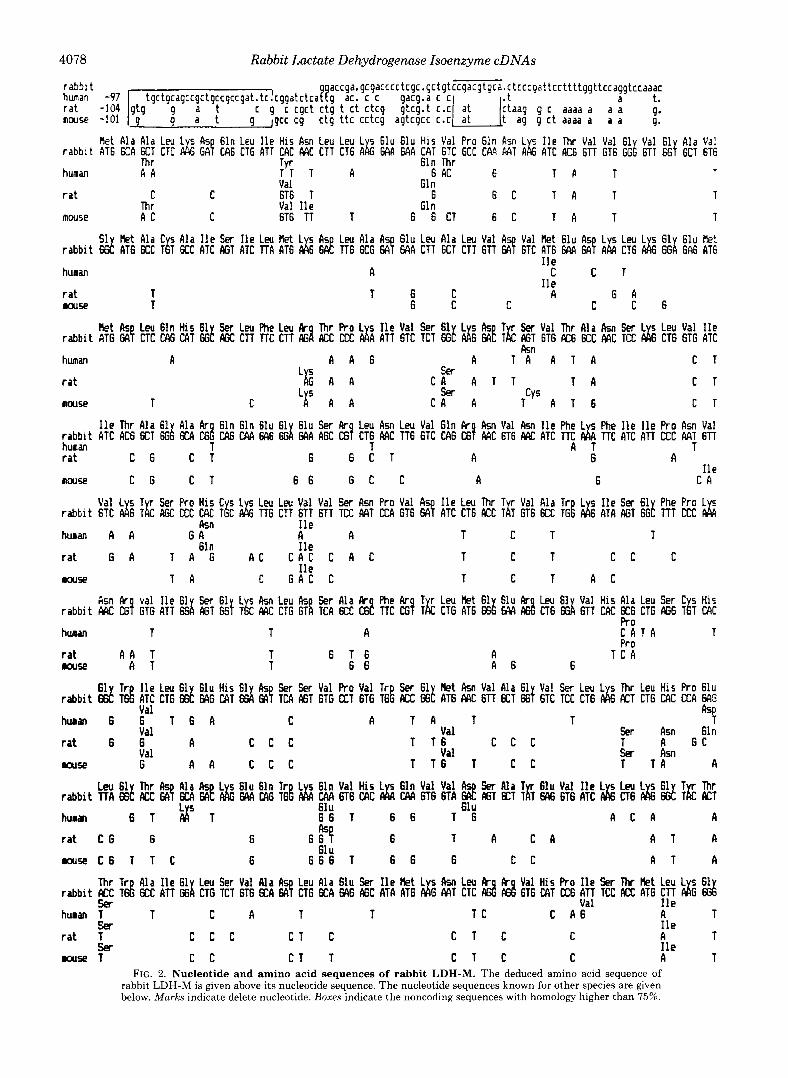
The nucleotide and protein sequences are shown in Fig. 2. The sequence of 1614 nucleotides contains a single 332-amino acid open reading frame.
Comparison with previously sequenced LDH-M protein from other species unambiguously demonstrated that this sequence encodes the entire LDH-M protein.
The amino acid and nucleotide sequences are very highly conserved among different mammalian species such as rabbit, human (18), rat (13), and mouse (19) and show 94% and 80% homology for these species, respectively (Table I). These results agree with those previously reported from Li et al. (20) for pig, chicken, and dogfish.
The 5’-noncoding region of our cDNA seems to lack about 35 nucleotides compared to the full length cDNA from rat (104 nucleotides) (13) and human (97 nucleotides) (18).
The 3’-noncoding region lacks about 20 to 30 nucleotides and does not contain any polyadenylation signal and poly(A) tail (Fig. 2). This is probably due to S1 nuclease treatment of the cDNA during the cloning process or to the existence of potential EcoRI sites that could cause the loss of a fragment during the subcloning of the cDNA in pGEM blue. This second hypothesis is supported by the observation that the sequences of the 3‘ and the 5‘ EcoRI sites are different from the sequences of the EcoRI linkers used and seem also to come from the cDNA itself.
The noncoding regions are more divergent than the coding region (Table I). The similarities are not randomly distributed but are localized for the 5‘-end in essentially three regions (21,12, and 8 nucleotides) with more than 80% similarity and for the 3‘-end in several clusters with more than 75% simi- larity. Some clusters of nucleotide deletions can be found for the 3’ noncoding region. Clusters seem to be specific for the LDH-M mRNA, they are not found in mouse LDH-X (21) or in human (22) and rabbit (this paper) LDH-H. The role and significance of this region should be determined.
The 3’- and 5”noncoding regions appear very highly con- served (80.9% and 81% similarity) in the evolutionary rela- tively closely related rat and mice, and, more surprisingly, also in humans and rabbits (60% and 77.9% similarity).
Thus, rabbit LDH-M mRNA appears to be more closely related to human than t o rat or mouse mRNA. This bears out the observations of Karlin et al. (23) for the immunoglobulin K gene, that the identity of the flanking and intron regions is stronger between humans and rabbits than between humans and mice. However, they found a more pronounced homology between human and mouse sequences with respect to the coding domain. This does not hold for the LDH-M cDNA (Table I).
Characterization of a LDH-H cDNA from the Rabbit Heart Library-Immunoscreening of 6 X 10‘ bacteriophages of the rabbit heart library identified seven positive clones. The long- est (pGRTHc) contains a cDNA insert of about 800 bp, while the six other ones contain only a fragment of about 180 bp. Sequencing of the longest cDNA, pGRTHc, was done accord- ing to the strategy in Fig. 3.
The nucleotide and deduced amino acid sequences are shown in Fig. 4 where there is a unique open reading frame of 654 bp. The LDH-H cDNA lacked a large 5’ portion of the mRNA, which probably resulted from the immunological screening which only detected translatable proteins. We have not rescreened the library with this probe to obtain a full length LDH-H cDNA. By comparison with the pig LDH-H amino acid sequence, it was deduced that this cDNA corre- sponds to residues 117 to 332. This 216-amino acid sequence has 95.4% similarity with pig LDH-H and 85% similarity with chicken LDH-H. Like the LDH-M mRNA, the LDH-H nu- cleotide coding region is very highly conserved in humans and rabbits. The nucleotide sequence shows 89% identity with the human LDH-H coding sequence (22) (Table 11). The coding sequence is followed by 126 bp of 3”untranslated sequence that appears incomplete. It lacks the polyadenylation signal and the poly(A) tail. Sakai et al. (22) demonstrated that the human LDH-H mRNA has a 203-nucleotide 3‘-noncoding region that is shorter than those of the human LDH-M mRNA. Like the LDH-M mRNA, the human and rabbit LDH-H 3”untranslated regions are more divergent than the coding regions (respectively, 56% and 89% similarity). As described by Li et al. (20) for pig and chicken, there are fewer differences between the same type of isoenzyme in different species than between different types of isoenzyme within a given species; from residues 117 to 332, rabbit LDH-M and LDH-H show 71.5% protein homology and 57.7% nucleotide sequence homology and the 3”noncoding regions differ to such an extent that the different sequences could not be properly aligned. Identical results have been reported with mouse LDH-X and LDH-M by Sakai et al. (21) and with LDH-H and LDH-M (22).
Characterization of Rabbit LDH-H and LDH-M mRNA- Using pGRTM12 and pGRTHc as probes, we analyzed the rabbit LDH mRNA by Northern blot (Fig. 5).
The cDNA pGRTM12 hybridized to an mRNA of about 1.8 kb in psoas muscle and the cDNA pGRTHc hybridized to a mRNA of about 1.4 kb in heart. These results are in good agreement with the RNA sizes predicted from the length of human LDH-M and LDH-H cDNA (18,22).
Level of LDH-H and LDH-M mRNA in Rabbit Muscle- The levels of LDH-M and LDH-H mRNA were estimated on dot-spots of total RNA from different rabbit muscles with specific nick-translated probes of rabbit LDH-M and LDH- H cDNAs. The two probes showed no cross-hybridization in Northern blot analyses.’
The three chosen muscles are representative of muscle type in rabbit (5): the psoas major muscle, a glycolytic fast twitch muscle composed of type I1 fibers; the semimembranosus
C. Sass, M. Briand, S. Benslimane, M. Renaud, and Y. Briand, unpublished experiments.
4078 Rabbit Lactate Dehydrogenase Isoenzyme cDNAs
r abbl t tgctgcagccgctgccgccgat. tc.cggatctcaPqg ac. c c gacg.a c c
a c c g a . g c g a c c c c t c g c . g c t g t c c g a c g t g c ~ . c t c c c g a t t c c t t t t g g t t c c a g g t c c a a a c a t.
m o w -101 g Jgcc cg c t g t t c c c t c g agtcgcc C.C g c t aaaa a a a g.
Ret Ala A la Leu L s As 61n Leu lle His A s n Leu Leu L 5 61u 61u His Val Pro 61n A M L s I l e Thr Val Val 61 Val 61 Ala Va! r a b b i t AT6 6CA KT CTC d6 GA! cA6 CTG ATT CAC M C C T T CT6 ALG MA 6AA CAT 6TC GCC CAA MT A16 ATC AC6 6TT GT6 GGi 6TT 6Gfl 6CT 6T6
human A A :'; T A 6 AC 6 Val Gln
T A T T
r a t C C 6T6 T 6 6 C Val lle G1 n
T A T T
mouse A C C 6T6 TT T 6 6 CT 6 C T A T T
c g c cgct c t g t c t c tc9 g tcg. t c.c g c aaaa a a a g.
Thr 61n Thr
Thr
r a b b i t 6d AT6 6CC TlT K C ATC ffil ATC T T A AT6 d6 6AE TT6 6CG 68 GAR CTT 6CT CTT 6TT 68 6TC AT6 6M SA! A h CT6 d6 6Gi 6A6 AT6 51 Bet Ala C s Ala l l e Sw I l e Leu Bet L s A5 Leu Ala A 61u Leu Ala Leu Val A- Val Bet 61u A s L s Leu L 5 61 61u k!
huran
r a t T Muse T
A I l e
T G C A I l e
6 A
C C T
G C C C C 6
r a b b i t A T 6 GAf CTC CAG CAT GG! f f i C CTT TTC CTT AG%i KC CCC d A A T T GTC TCT ffi! d G EA! Tk f f i T 6 T G X6 6CC MC TCC d G CT6 GTG ATC k t b Leu 61n His 61 Ser Leu Phe Leu k Thr Pro L s I l e Val Ser 61 L s b T r Ser Val Thr Ala Ikn Sw L s Leu Val l l e
human A A A 6 A T A A T A C T
r a t C A A T T T A C T
k n
Ll(c' A A ser sw
Douse T C Lr A A C A A TcYs A T 6 C T
I l e Thr Ala 61 Ala k 61n 61n 61u 61 61u Ser Rr Leu A s n Leu Val 61n k A s n Val Asn I l e Phe L 5 Phe l l e I l e Pro A s n Val r a b b i t ATE ACG 6CT 6G! 6cll Id CA6 MA EA6 6Gi MA AGE CS! CT6 M C TT6 6TC CA6 E! M C 6T6 ARC ATC TTC &I TTC ATC A T l CCC A4T 6TT human
C 6 T
C T r a t 6 6 C T A
nouse C G C T 6 G 6 C C A 6 C A I l e
r a b b i t 6 T C d6 TE AGC CCC CAC TEC d6 T16 CTT 6TT 6TT TCC ACIT CCA GT6 3 ATC CT6 K C T f i GT6 6CC TGE d6 ATA ffil 6GE llT CM: & h u m A A G A A A T C T T
r a t 6 A T A G A C C A C c A c T C T c c C
T A muse C G A C C T C T A C
Ftsn t val I l e 61 Sw 61 L s kn Leu As Ser Ala k Phe Ar T r Leu k t 61 61u t Leu 61 Val His Ala Leu Ser C 5 His r a b b i t MC CG! GTG AT1 6d f f i T 6G! T i c ME CTG GT! TCA 6M: CS! TTC CG! T k CTG AT6 6Gb 6rw d CT6 6Gi GTT CAC 6C6 CTG f f i G T i T CFIC
hunan T T A C A T A T
r a t A A T T 6 1 6 A T T 6 G muse A 6 6
61 Tr Ile Leu 61 61u His 61 Ser Ser Val Pro Val T r Ser 61 k t Ikn Val Ala 61 Val Ser Leu L s Thr Leu Hi5 Pro 61u r a b b i t 6d T& ATC CT6 d 6AG CAT Ssi 3 TCA f f i T 6 T G CCT 6TG T6i KC 6 G E ATG ME 6TT KT 6G! 6TC TCC CTG p);s K T CTG CAE EA €4':
T A T T 6 A
Val L s T Ser Pro His C s L s Leu Leu Val Val Ser A s n Pro Val I l e Leu Thr T Val Ala Tr L s I l e Ser 61 Phe Pro L
b n
61n
I l e
I l e
I1 e
Pro
Pro A T C A
Val hum 6 6 T 6 A C A T A T T 9 r a t 6 6
Val A c c c T 1 6 c c c T A 6 C
Val Sw Asn 61n
Val G A A c c c T 1 6 T
Val Ser A m Muse c c T T A A
r a b b i t l T A Ed llcc k! % & MA CAG T d Ah CM 6T6 CAC & CM 6T6 6TA 2 ffiT KT T 6 I 3 6 616 ATC d6 ET6 I& 6G! T E KT Leu 61 Thr L s 61u 61n T r L s 6111 Val His L s 61n Val Val SW Ala T 61u Val I l e L s Leu L s 61 T Thr
h u m 6 1 2 1 6 6 1 6 A C A A 61u 6 6 T
r a t C 6 6 6 6 E! 6 T A C A A T A
61 u
muse C 6 T T C 6 6 6 6 T 6 6 6 c c A T A
r a b b i t XC T& KC A T T ffii CTG TCT 6TG KA 8 CT6 6CA GAG cyjc ATA AT6 d G MT CTC I%! I%! 6T6 CAT E6 A T T TCC KC AT6 Cll d6 Ed huran 1 T C A T T T C C A 6 A T
61 u
Thr T r Ala I l e 61 Leu Ser Val Ala Leu Ala 61u Ser I l e k t L 5 Asn Leu k k Val His Pro I l e Ser Thr k t Leu L 5 61
SB Val I l e
ser
Ser r a t T
muse T
c c c C T C
c c C T T
C T c C
C T C C
n I l e
T I l e A T
FIG. 2. Nucleotide and amino acid sequences of rabbit LDH-M. The deduced amino acid sequence of rabbit LDH-M is given above its nucleotide sequence. The nucleotide sequences known for other species are given below. Marks indicate delete nucleotide. Boxes indicate the noncoding sequences with homology higher than 75%.

Rabbit Lactate Dehydrogenase Isoenzyme cDNAs 4079
rabbit CTC T h GGI ATA dG GAG GA! GTC TTC CTT AGT GTT CCT Tk GTC TT6 G G i CAIl AAT 6Gi ATC TCR 6AF GTT 6TG d G 6 T 6 ACT CTA ACC TCT LEU T r 61 Ile L s 61u As Val Phe Leu S E ~ Val Pro C s Val Leu 61 G l n A w l 61 Ile Ser As Val Val L s Val Thr Leu Thr SP
Ile human T C A T 6 A C G T
Ile rat C C C C A T A C A T A G T C
C T Asn I l e
nouse C C A T A C A G T A G T C
As! LeU
Pr :
frc
rabbit MA MA M G GCC CAC CTG d G d G AGC GCG GAE ACG CTG T6E GGL ATC CAA dA GAG CTG CA6 TTT 61u 61u Glu Ala His L e u L s L s Ser Ala As Thr Leu Tr 61 l l e 61n L s G l u L e u 61n h e
Ar human G 6! T T A T A T 6 G A
T A T C C
A c c G G
G G
C
C - rabbit taaagtcttggataatbtcctaccacttcactgtctaggc acaacaggg. tt.t~gttggaggttgtgcatgtggtcctttaatctgatccatgatt~, . , . , . , , .aagcagtga at human I..C q I a t t 9 .a c I 9 I t tQ I . . ....... a ? ......... .. 9 c tg g t tgg c cal ;;g g ti g Jgtccagttg c tg c tg g c c gca ...... c ... cac g t g g ......... gta .. g rabbit cct..gatggcctgggaggggaaaacat. .gtttcctaaagttagaaat.agaatggtttataaaatccaacagctacatggaggtactgggtgacactgactttgtgtcctgagctggt h u m tt.a. a ........ c.a 9 a
louse gt ga tg ..... .g ctcac ............................... ctgccct c g cad gt ... ..c
- rat gt ga g . . . . . . g . ctcag . . . . . . . . . . . . . . . . . . . . . . . . . . . . ..... 1 btgccct c
... a: gt !.?.!E/ ii rabbit cqcctaagata agttc.cccacctctgaggtgccactgcctgtgt.. . .tgcacgagctcca ttacccct.ccgactagacgtgcgtC actgtgtgctacg aactt.ctggt.. . human t ... rat IJ:: $ .? c a c ca tgt ca aca. 9 t9 c ca t ta 9 g .tgag c gt gat 9 c t ta? c .. ftga . gtgag ct ct cam.. . . . . . . . . . . . ........ g !:;G louse t . . gt .. c c ca tgt a m . .. t ga gtgag ct ct ca. ..................... g ..... rabbit ... human .... ... rat cgca . a . . . . ...... g . . . . . rabbit cttattttgtgtatgatgta. ... tga~$agtgtacatt.gcacataatgtaaaaagaaagatctacata. tagcaatgcaactcgctatccatgtgtcatacccc 1614 human
gca t t c g :[it1 ; J" a ac t t rat .. c ..t a tgtg ct tggatg g caa c ct a
human ctaaaacccccaataaaccttgaacag g 1661 rat g a 1609 ... ........... 1590
g caa a t aa 9 aa 9 a. muse .. a a ..t a tgtg ct g c a t t g g t t tggatg g caa c c a
FIG. 2-continued
TABLE I Comparison of the amino acid and nucleotide sequences of
mammalian LDH-M (% similarity) Amino acid Coding 3"Noncoding 5"Noncoding
sequence region region region
Rabbit/human 93.6 87.7 60 77.9 Rabbit/rat 94.3 84.7 60 56.2 Rabbit/mouse 93.7 85.3 46.3 51.3 Human/rat 94.3 84.5 50.8 56 Human/mouse 93.7 85.3 53.2 Rat/mouse 96.4 92.7 80.9 81
- - - " - a
- 0 " .- >
Y
0
- - a m c = E I O VI VI "
t A
____, +"---
b
100 bp 1 4
I b
FIG. 3. Sequencing strategy used for rabbit LDH-H cDNA and partial restriction endonuclease. Arrows indicate the direc- tion and length of each sequencing run.
proprius muscle, an oxidative slow twitch muscle composed of type I fibers; and the heart. The three muscles each have a specific pattern of LDH activity and LDH isoenzyme subunit distribution as shown in Table 111. In the psoas major muscle,
GGC MG TTC A l T RlT CCC U I G RTT 61C RAG TRC ffiT CCC 1 C TGC RTC ATC AIC GTG Gll ICC 1 C C C A GIG WIC AIT 76 G I ~ L ~ S h e 11e I I~ RO GI" 11a 'vd L ~ S ~ y r Ser R o kn Cyr Ile Ile Ile Val V a l 5er Rsn Pro V a l kp Ile
LRI nr 1yr V a l mr irp t y s LW 5er ~ l y teu R O tys H I S Rp V a l l l e G l y Ser G l y Cys kn Lw A s p 5er Rla
PGR T I T C6C lR7 C l C AT6 KT G%C E I G CTT GGC AI1 CAT CCC RGC f f i C TGC 021 G S A TGG All T I G EA W A UIl !%A 231 kg h e Rq Tyr LW k t A l a G1u ~ y s Leu Gly Ile HIS Ro Ser Ser Cy5 His 61y lrp Ile Leu Gly Glu Hi5 G l y
WC TCA R6G TTG GCI G I C 166 ffil 6wi GTG RRT GI6 GCA 661 611 IC1 C I C U I G GRA CTG ART CXA O I G R I G GGR Rcfi 512 asp 5er ~9 ieu RIA V a l 1rp 5er 61y Val asn vai Ala G1y V a l Ser Leu Gln G 1 u Leu kn Rn Glu k t 61y Thr
WIC MT WT I T ERA MC I66 1 6 W G GIG UIC M G RIG GTG GTG UVI f f i T KC 1Al W R GIC RIC M E CIA MR 666 396 kp Asn asp Ser G l u kn Irp Lys Glu V a l lhs Lys kt V a l Val G1u Ser Rla Tyr Glu Val Ile Ly5 Leu Lys G l y
TRC ACT RRC 166 GCT All GM\ T I R 161 GTG ET W l CTC R T I MA TCC ATG C16 M 1 T CTC TCC AGG Rll UIC CCA 4b8 Tyr mr kn lrp Rla Ile G l y Leu 5er Val A l a kp Leu l le Glu Ser k t Leu Lys kn Leu Ser kg IIP Hli h o
V a l Ser Thr kt Val R9 Gly kt lyr G l y lie Glu Ser G l u Val R e Leu Ser Leu Fro Cys l ie Leu kn A l a k9
6% T I A KC f f i C GTC AIC MC CAG W6 CTC lyKj GRC WIT UKi G I T KC CRG ClC RAG RAR A61 6cR W C Acc I l G 166 b24 G 1 y Leu Thr Ser V a l Ile kn G l n lyr Leu Lys kp kp G l u Val Ala 61n Leu L y i Lys Ser Ala R s p mr Leu Irp
WC RTC CAG RAG WC ClC MI WC C16 t q a c t r q r q a g t p c t ~ g p c t q t a q ~ a ~ ~ c ~ a a q c t c t ~ ~ t g t ~ a t a t q t ~ c t ~ a q c c ~ a c a q c 714 k p l i e G l n Lys kg Leu Lys kp Leu
q c t c a t g r c ~ q q g t q t g g g g c ~ ~ t q t t t q ~ t ! t t ~ ~ t ~ ~ t ~ ~ t t ~ ~ t ~ g q t q a ~ t t t g ~ c c c q c c 780
CTT ACA TAC G l G ACC T6G iY IG Clfi 161 6 6 A CTG CCC RAG CAC c 6 T GIG RIT f f i A AGT 666 16C iYI1 CTG W T TCT GCT 1%
GT6 TCC AUI A N GTG ffi6 GGC A16 TRT 66C ATT WG M W G I C 1 1 C CTG A6c C I T CCG 16C AIC CTC 1 1 KC C6G 54b
FIG. 4. Partial nucleotide and amino acid sequences of rab- bit LDH-H. The deduced amino acid sequence is given below its nucleotide sequence.
TABLE I1 Comparison of the amino acid and nucleotide sequences of human
and rabbit LDH (% similarity) Amino acid Coding 3'-Noncoding
sequence region region
Rabbit H/human H 89 89 56 Rabbit H/rabbit M 71.5 57.5 Human H/human M 75 68

4080 Rabbit Lactate Dehydrogenase Isoenzyme cDNAs
1 .8 K b
1 . 4 K b
FIG. 5. Northern blot analysis of RNA from rabbit muscle. Northern blot of RNA from psoas major (1 pg-a) and heart (1 pg-b), hybridized with the nick-translated cDNA probes pGRTM12 (A) or pGRTHc (B) .
TABLE 111 Lactic dehydrogenase activity and mRNA in rabbit muscle
PM, psoas major; SP, semimembranosus proprius; H, heart. Values are means + S.D. (N = 3).
Muscle T~~~~~ + H subunit LDH mRNA/g LDH M + H % LDH-M/M Picomoles total % LDH-M mRNA/
proteinb muscle X lo2‘ mRNAd
PM 1100+50 9 3 2 4 38.1 97.6 SP 80 rt 60 20rt 8 98.4 12.2 H 6 0 f 1 0 3 + 2 167.1 0.7 “Total activity is expressed in pmol substrate/min/g of fresh
muscle. bThe ratio LDH-M/M + H subunit protein was calculated from
electrophoresis as described under “Materials and Methods.” The molar concentration of total LDH mRNA is calculated from
the values of Table IV, Footnote 6. dPercentage of LDH-M mRNA is expressed as a molar ratio to
the H + M LDH mRNA and calculated from values of Table IV, Footnote 6.
the LDH-M subunit represents 93% of the total LDH subunit, whereas in the semimembranosus proprius muscle and the heart, we find, respectively, only 20% and 3% LDH-M sub- unit.
Table IV shows for the same muscle samples used for LDH protein determination that the amounts of LDH-M or LDH- H mRNA are quite different in the three muscles studied. The highest amount of LDH-M mRNA is found in the psoas major muscle, followed by the semimembranosus proprius muscle, then the heart. The opposite distribution is observed for the amount of LDH-H mRNA.
We have not been able to compare the amount of LDH-M or LDH-H mRNA with those of the respective protein sub- units, since we have only calculated the relative LDH subunit distribution by the electrophoretic profile of LDH isoenzymes. Nevertheless, our results (Table 111) show a rather good relationship between the ratio of LDH-M mRNA/total LDH mRNA with the ratio of LDH-M subunit/LDH-M + LDH-H subunit, suggesting that the muscle-specific ratio of LDH-M and -H subunit is essentially determined by the amount of LDH-M and -H mRNA.
These results are in agreement with Miles et ~ l . (25) and Jungmann et ~ l . (26) who demonstrated that the cyclic-AMP induction of LDH-M in the C6 rat glioma cell line is controlled at the transcriptional and post transcriptional level. Con- versely Piechaczyk et ~ l . (24) suggest that the transcriptional level of LDH-M mRNA is similar in different tissues (kidney,
TABLE IV Lactic dehydrogenase isoenzyme and mRNA in rabbit muscles
PM, psoas major; SP, semimembranosus proprius; H, heart. Values are means + S.D. (N = 3).
% mRNA/total RNA pmol mRNA/g Muscle x 102” PP total RNA/g muscle x lozb
muscle LDH-M LDH-H LDH-M LDH-H
PM 7.02 + 2.7 0.13 k 0.01 287 f 60.2 37.2 0.89 SP 0.9 + 0.15 5 + 2.1 726 f 82.9 12.1 86.3 H 0.12 + 0.06 15.3 f 4.8 457 f 44.6 1.1 166 ?6 mRNA/total RNA is expressed as percentage weight.
*The molar concentration of LDH-M and LDH-H mRNA is calculated from the values determined in columns 1 and 2 and corrected with the expected size of the respective mRNA (1.8 kb for LDH-M and 1.4 kb for LDH-H mRNA).
liver, heart, and skeletal muscle). To determine if the amount of total LDH mRNA is related
to the total LDH, we have compared the amount of total LDH-mRNA with the LDH activity in the different muscles studied. Table I11 shows that there is no relationship between them. This result cannot be explained by the specific activity of H or M LDH because, under our experimental conditions, the specific activity of LDH-M4 was only twice that of LDH- H4 (not shown). This result suggests two hypotheses. Firstly, the turnover of LDH is different for the different muscles and secondly there is a translational control depending on the tissue.
Nadal-Ginard (27) showed that the LDH turnover is differ- ent in mouse heart and mouse skeletal muscle (T!,,, respec- tively, 8 days and 43 days) and proposes that the amount of total LDH is regulated at the degradation level while the H and M subunit ratio is regulated at the synthesis level. Our results support this theory.
CONCLUSION
By screening the two rabbit cDNA libraries from heart, containing 97% LDH-H protein, and from psoas major mus- cle, containing 93% LDH-M protein, we are able to isolate cDNAs coding for LDH-H and LDH-M. The amino acid and nucleotide sequences of rabbit LDH-M are 93.6% and 87.7% homologous with human LDH-M (18), the amino acid se- quence 117 to 332, and the corresponding nucleotide sequence of rabbit LDH-H are 89% homologous with human LDH-H (22), whereas the similarity between rabbit LDH-H and -M is lower (71.5% and 57.5%). A similar result is described for humans. This result confirms that similarities between M or H subunits are closer between species than between the M and H subunits of the same species.
Isolation of the cDNAs for both H and M LDH subunits opens the way for further studies of LDH regulation during the process of myofibrillar differentiation. We have already demonstrated a correlation between LDH isoenzyme profile and muscle fiber type (16). Alternatively, LDH isoenzymes represent a marker for studies of muscle differentiation.
The expression of LDH-M and LDH-H of adult rabbit muscle was also investigated in this study. A good relationship was found between the percentage M to H mRNA and the percentage M to H subunit protein, which indicates a tran- scriptional and/or a post-transcriptional control. However, as there was no relationship between the level of H + M mRNA and the total enzyme activity, this probably indicates the presence of an already suggested tissue-dependent transla- tional control (27).
Acknowledgments-We thank Dr. Fiszman from the Institut Pas- teur, Paris, in whose laboratory the sequences were made. We are

Rabbit Lactate Dehydrogenase Isoenzyme cDNAs 4081
grateful to Dr. R. Breatnach for his gift of plasmid pLDH2, t.0 J. C. 49-78, IRL Press, Arlington, VA Barlet from INRA, Theix, for the preparation of sheep antisera, and 13. Matrisian, id. M., Rautmann, G., Magun, B. E., and BreathnaCh, to Mr. Lacoste from Elevage de la Font des Escuroux for providing R. (1985) Nucleic Acids Res. 13, 711-726 the rabbit. 14. Alric, M., Cheyvialie, D., and Renaud, M. (1986) Anal. Biochem.
REFERENCES 15. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular 1. Cahn, R. D., Kaplan, N. O., Levine, L., and Zwilling, E. (1962)
Cloning: A ~ b o r a t o r y Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
2. Goldberg, E. (1963) Science 139,602-603 16. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Nutl.
3. Markert, C. L., Shakelee, J. B., and Whitt, G. S. (1975) Science 17. Messing, J. (1983) Methods Enzymol. 109, 20-78 Acad. Sci. U. S. A. 74,5463-5476
4. Leberer, E., and Pette, D. (1984) Histochemistry 80,295-298 147,9-15 5. Briand, M., and Briand, Y. (1986) Comp. Biochem. Physiol. 85B, 19. Fukasawa, K. M., and Li, S. S.-L. (1987) Genetics 116,99-105
6. Briand, M., Talmant, A., Briand, Y., Monin, B., and Durand, R. J. Biol Chem. 258,7029-7032 (1981) Eur. J . Biochem. 46, 359-365 21. Sakai, I., Sharief, F. S., and Li, S. S.-L. (1987) Biochem. J. 242,
7. Fritz, P., Morrisson, W. J., White, E. L., and Wessel, E. S. (1970) 619-622 Anal. Biochem. 36,443-452 22. Sakai, I., Sharief, F. S., Pan, Y.4. E., and Li, S. S.-L. (1987)
8. Cathafa, G., Savouret, J. F., Mendez, B., West, B. L., Karin, M., 23. Karlin, s. M,, Ghandour, G., and Fou,ser, D. E. (1985j Mol. Bioi. Martial, J . A., and Baxtar, J. M. (1983) DNA ( N . Y . ) 2, 329- 335
9. Aviv, H., and Leder, P. (1972) Proc. Natl. Acad. Sci. U. 5'. A. 69, 24. Piechaczyk, M., Blanchard, J. M., Marty, L., Dani, C., Pana- bieres, F., El Sabouty, S., Fort, P., and Jeanteur, P. (1984)
1408-1412 10. Lehrach, H., Diamond, D., Wozney, J. M., and Boedtker, H. 25. Miles, M. F., Hung, P., and Jungmann, R. A. (1981) J. ~ & l .
Nucleic Acids Res. 1 2 , 6951-6963
11. Lapeyre, B., and Amalric, F. (1985) Gene (Amst.) 37, 215-220 26. Jungmann, R. A., Kelley, D. C., Miles, M. F., and Milkowski, D. 12. Huynh, T. V., Young, R. A., and Davis, R. N. (1984) in DNA M. (1983) J. Biol. Chem. 258,5312-5318
155,328-334
Science 136,962-969
189, 102-114 18. Tsujibo, H., Tiano, H. F., and Li, S. S.-L. (1985) Eur. J. Biochem.
481-485 20. Li, S. S.-L., Fitch, W. M., Pan, Y.-C. E., and Sharief, F. S. (1983)
Biochem J. 248,933-936
Evo~. 2, 35-52
(1977) Biochemistry 16,4743-4751 Chem. 256,12545-12552
Cloning Techniques. A Practical Approach (Glover, D., ed) pp. 27. Nadal-Ginard, B. (1978) J. Biol. Chem. 253, 170-177