caractérisation par sérotypage et pulsotypage des souches de … · utilisant l’enzyme de...
TRANSCRIPT

Pôle Viandes Fraîches et Produits Transformés Juin 2008
Caractérisation par sérotypage et pulsotypage des souches de Salmonella isolées dans le secteur abattage-découpe porc
Carole FEURER, Gwénaël PIAUDEL

3
Sommaire
Sommaire ................................................................................................................... 3
Lexique....................................................................................................................... 4
Résumé ...................................................................................................................... 5
Introduction................................................................................................................. 6
Matériel et Méthodes .................................................................................................. 7
Isolats et commémoratifs ........................................................................................ 7
Sérotypage.............................................................................................................. 7
Pulsotypage ............................................................................................................ 7
Antibiorésistance..................................................................................................... 8
Traitement statistique des données ........................................................................ 8
Résultats................................................................................................................... 10
Isolats et commémoratifs ...................................................................................... 10
Sérotypage............................................................................................................ 10
Pulsotypage .......................................................................................................... 12
Antibiorésistance................................................................................................... 17
Conclusions et Perspectives..................................................................................... 19
Bibliographie............................................................................................................. 21

4
Lexique
AFSSA Agence Française de Sécurité Sanitaire des Aliments
CNR Centre National de Référence
PCR Polymerase chain reaction – Réaction d’amplification en chaîne
PFGE Pulsed Field Gel Electrophoresis – Electrophorèse en Champ Pulsé
TIAC Toxi-infection alimentaire collective
TSAYE Trypton Soja Agar plus Yeast Extract
Antibiotiques AM Ampicilline
AMC Amoxicilline + acide clavulanique
C Chloramphénicol
CAZ Ceftazidime
CF Céfalotine
CS Colistine
CTX Céfotaxime
ENR Enrofloxacine
GM Gentamicine
K Kanamycine
NA Acide nalidixique
OFX Ofloxacine
S Streptomycine
SSS Sulfamides
SXT Sulfaméthoxazole-triméthoprime
TE Tétracycline

5
Résumé
L’objectif de ce projet est d’enrichir en souches de Salmonella une base de données filière
mise en place lors d’une précédente étude financée par l’OFIVAL.
Un total de 200 souches ont été analysées lors de cette étude. Elles proviennent de deux
entreprises partenaires du projet et sont issues des maillons abattage/découpe et
transformation. Deux techniques de typage ont été utilisées : le sérotypage et le
pulsotypage. En effet, le sérotype et le pulsotype ont été définis lors de la précédente étude
comme informations essentielles à obtenir pour toute souche intégrant la base de données.
Un antibiogramme a également été réalisé sur certaines souches de sérotype Typhimurium
présentant un pulsotype pouvant être associé à des souches DT104 chez lesquelles ont
retrouve communément de nombreuses résistances aux antibiotiques.
Les résultats obtenus montrent une forte prévalence de deux sérotypes (Derby et
Typhimurium), avec au sein de ces 2 sérotypes une prédominance importante de deux
pulsotypes (Ty01 et Der03), représentant à eux seuls plus de la moitié des isolats collectés
lors de cette étude. Les sérotypes obtenus pendant ce travail sont au nombre de 12, avec
une plus grande diversité obtenue dans l’entreprise de transformation, résultats en accord
avec ceux obtenus dans des études similaires et confirmés par ceux du pulsotypage (46
pulsotypes identifiés dont 39 pour l’entreprise de transformation et seulement 12 pulsotypes
identifiés pour l’entreprise d’abattage-découpe).
La très forte prévalence du pulsotype Ty-01 (28%) montre, dans ce cas particulier,
l’importance et l’utilité d’une base de données. Ce pulsotype est en effet régulièrement
associé aux souches de type DT104. Ces dernières sont impliquées dans de nombreux cas
humains de salmonellose, avec de surcroît une tendance à présenter de nombreuses
antibiorésistances, tendance qui semble être à la hausse depuis quelques années. Le suivi
de ce type de souches est donc d’un grand intérêt au sein de la filière porcine, de part son
impact avéré en santé humaine, mais nécessite l’emploi de méthodes d’analyse plus fines
que la seule sérotypie.
Les analyses concernant l’antibiorésistance de souches pouvant être assimilées au type
DT104 montrent qu’aucune des souches étudiées n’était sensible à tous les antibiotiques
testés, la plupart présentant même plusieurs résistances (jusqu’à sept résistances pour deux
souches). Cependant l’échantillonnage testé pour l’antibiorésistance ne permet pas de tirer
de conclusions compte-tenu du faible nombre de souches étudié (20).
Afin de mieux utiliser cette base de données Salmonella et pouvoir tirer des conclusions
utilisables pour les acteurs de la filière, il est nécessaire de continuer à incrémenter le
souchier (300 souches prévues sur l’année 2008) et si possible de pérenniser l’apport de
souches dans l’avenir. L’utilisation d’une telle base n’a en effet d’intérêt que si elle est
régulièrement alimentée par l’ensemble de la filière afin de pouvoir jouer son rôle
d’observatoire et donc de gestion du risque Salmonella.

6
Introduction
Salmonella représente à l’heure actuelle une des principales causes de toxi-infections
alimentaires d’origine bactérienne en France et dans le monde. Leur présence dans les
matières premières est critique compte-tenu des exigences réglementaires nationales ou
communautaires en vigueur. Le contexte réglementaire actuel vise à réduire la
séroprévalence de Salmonella dans la filière porcine et par conséquent le risque pour le
consommateur par la mise en place de programmes de contrôles nationaux à partir de 2009
(directive 2003/99/CE et règlement 2160/2003 du 17 novembre 2003). La création d’une
base de données Salmonella recouvrant spécifiquement les différents maillons de la filière
porc s’inscrit donc dans ce cadre en offrant à terme un outil permettant d’aider à remplir ces
objectifs.
Au niveau européen, différentes bases de données ont été constituées pour réaliser la
surveillance de plusieurs bactéries pathogènes (Listeria avec le réseau Listernet, Salmonella
avec les réseaux Salmgene et Enternet, Campylobacter avec le réseau Campynet). Des
collaborations entre l’Ifip et les laboratoires responsables de ces bases de données sont
menées depuis plusieurs années pour Listeria et Salmonella ce qui a permis d’obtenir de
nombreuses données de pulsotypage pour Listeria ainsi que pour Salmonella mais dans une
moindre mesure .
Dans le cadre de ce projet, l’objectif était d’alimenter la base de données existante en
souches de Salmonella isolées dans les différents maillons de la filière à partir
d’autocontrôles réglementaires ou volontaires. Une analyse de ces souches par des
méthodes fines (sérotypage et pulsotypage) a ensuite été réalisée.
La compilation des résultats devrait permettre de mieux comprendre la transmission de
Salmonella de l’élevage jusqu’au produit fini, ainsi que d’établir les liens ou les spécificités
des souches tout au long de la filière, dans le but de déterminer les meilleures pratiques qui
pourront permettre la maîtrise du danger Salmonella dans l’ensemble de la filière porc.
Le projet consistait à :
-identifier et démarcher les entreprises partenaires pouvant fournir des souches de
Salmonella issus de leurs autocontrôles.
-déterminer le pulsotype et le sérotype de ces souches ainsi que le profil
d’antibiorésistance pour certaines souches d’intérêt.
L’obtention de ces données nous a permis d’obtenir un premier aperçu de la diversité des
souches de Salmonella au sein de la filière ainsi que de leur transmission du maillon
abattage/découpe au maillon transformation.

7
Matériel et Méthodes
Isolats et commémoratifs
Pour permettre la création d’un souchier Salmonelle, des entreprises correspondant aux
différents maillons de la filière porc ont été contactées afin de pouvoir collecter sur la base
du volontariat des isolats provenant de leurs procédures d’autocontrôles volontaires ou
réglementaires. Deux entreprises ont été sollicitées et ont répondu positivement à cette
requête, un abattoir industriel (entreprise E1) et une entreprise de transformation (E2).
Un commémoratif complet pour chaque échantillon expédié était demandé, de manière à
disposer de l’information la plus complète concernant les isolats à intégrer dans la base de
données (date et lieu de prélèvement, origine et stade du prélèvement).
Les souches collectées ont ensuite été isolées sur gélose Hektoen pour vérifier leur pureté.
A partir de colonies isolées, les isolats ont été mis en gélose de conservation en vue de leur
analyse par sérotypage et pulsotypage.
Sérotypage
Pour chaque souche, le sérotypage a été effectué au laboratoire LERQAP de l’Afssa de
façon anonyme. La détermination antigénique a été réalisée selon le schéma de Kauffmann-
White (2001 8ème édition).
Pulsotypage
Le pulsotypage a été réalisé au laboratoire de microbiologie de l’IFIP de Maisons-Alfort en
utilisant l’enzyme de restriction XbaI qui offre une bonne discrimination entre les souches de
Salmonella. L’extraction, la digestion et la migration de l’ADN des souches ont été réalisées
selon le protocole élaboré par le CDC (Center of Disease Control) d’Atlanta.
Une culture sur gélose TSAYE de 24 heures est inondée de tampon CSB (Tris-Hcl 100mM,
EDTA 100mM), puis la densité optique (DO) de la suspension obtenue est mesurée avant
d’être ajustée à une unité de DO. Un volume de 240µl de la suspension ajustée est prélevé
puis mélangé à de l’agarose (1% au final) en surfusion contenant du SDS et de la protéinase
K, puis disposé dans un moule de manière à obtenir des mini blocs d’agarose. Une lyse
cellulaire est ensuite effectuée pendant 2 heures à 52°C dans un tampon de lyse (Tris
0,05M, EDTA 0,05M, N-lauryol sarcosine 1%, Protéinase K 0,156g/ml). Une fois l’incubation
réalisée, les blocs d’agarose sont lavés avec du tampon TE (Tris-Hcl 10mM, EDTA 1mM) à 6
reprises, puis conservés à 4°C dans du tampon TE jusqu’à utilisation.

8
La digestion de l’ADN chromosomique est réalisée au moyen de l’enzyme de restriction XbaI
pendant 4 heures à 37°C dans le tampon optimal de l’enzyme.
Une fois la digestion effectuée les blocs d’agarose sont transférés dans un gel d’agarose à
1% TBE 0,5X (Agarose Seablock), puis la migration est réalisée dans les conditions
suivantes (Bloc 1 : 2 s–64 s, 20h, 6 V/cm, 120°) au moyen d’un appareil CHEF DR® III de
Biorad. A la fin de la migration, le gel est coloré au moyen d’une solution de bromure
d’éthidium (BET) avant d’être révélé sous UV.
Les profils obtenus sont numérisés avant d’être analysés et comparés au moyen du logiciel
Molecular Analyst de Biorad, en utilisant le profil de la souche S.enterica sérotype
Brandeburg H4229 comme référence externe (nécessaire pour la comparaison de gels
indépendants). Les données seront progressivement analysées avec le logiciel Bionumerics
(Applied Maths) dont l’acquisition fonctionnelle est en cours.
Antibiorésistance
La détermination de l’antibiorésistance a été réalisée à l’Afssa de manière anonyme. La
recherche d’antibiorésistances a porté sur 16 antibiotiques parmi les plus courants (Tableau I) et interprétée selon les recommandations du Comité Antibiotiques de la Société Française
de Microbiologie. Cette recherche de résistance a porté spécifiquement sur les souches de
sérotype Typhimurium, du fait de l’implication clinique de ces souches, ainsi que de la forte
prévalence de multirésistances chez celles-ci.
Traitement statistique des données
Afin de valider ou d’infirmer le fait que les résultats obtenus (sérotypes, pulsotypes) étaient
ou non corrélés à certains paramètres (entreprise d’origine, type de matrice d’origine), des
hypothèses d’indépendance ont été testées à l’aide du test d’indépendance du khi2 (χ²). Le
principe est de calculer l’écart entre la distribution obtenue et une distribution théorique que
l’on obtiendrait si les deux variables étaient totalement indépendantes. Cet écart permet
d’accepter ou de rejeter l’hypothèse d’indépendance H0 (données indépendantes).
Les résultats de cette étude étant de nature qualitative et l’échantillonnage suffisant (> 50), il
est donc possible de vérifier si la distribution des types identifiés est liée ou non au
paramètre pris en considération. Dans ce rapport, l’hypothèse de départ (indépendance des
données entre elles) est acceptée ou refusée au seuil de 5%.

9
Tableau I : Récapitulatif des antibiotiques utilisés dans cette étude, les doses testées et les
diamètres critiques utilisés pour l’interprétation.
Famille d’antibiotique Panel I (charge, diamètres critiques selon le CA-SFM*, mm)
PÉNICILLINES AM : Ampicilline (10 µg, 14-19)
AMC: Amoxicilline + acide clavulanique (20 µg + 10 µg, 14-21)
CEPHALOSPORINES CF : Céfalotine (30 µg, 12-18)
CTX : Céfotaxime (30 µg, 15-21)
CAZ : Ceftazidime (30 µg, 15-21)
AMINOSIDES S : Streptomycine (10 UI, 13-15)
GM : Gentamicine (10 UI, 14-16)
K : Kanamycine (30 UI, 15-17)
PHENICOLES C : Chloramphénicol (30 µg, 19-23)
TETRACYCLINES TE : Tétracycline (30 UI, 17-19)
SULFAMIDES-
TRIMETHOPRIMES
SXT : Sulfaméthoxazole-triméthoprime (23.75 µ+1.25 µg, 10-16)
SSS : Sulfamides (200 µg, 12-17) ouSSS250 : Sulfonamides (300 µg, 12-17)
QUINOLONES NA : Acide nalidixique (30 µg, 15-20)
OFX : Ofloxacine (5 µg, 16-22)
ENR : Enrofloxacine (5 µg, 17-22)
POLYPEPTIDES CS : Colistine (50 µg, 15)
: Critères du CA-SFM, 2007 sauf pour l’enrofloxacine (Bayer).

10
Résultats
Isolats et commémoratifs
Lors de cette étude, 200 isolats de Salmonella ont été collectés. Parmi ces isolats, 88
provenaient de l’entreprise E1 (abattage découpe) et 112 de l’entreprise E2 (transformation).
Les origines de prélèvement étaient diverses : environnement (2), animaux (13), abats (10),
matières premières (épaule, poitrine, onglets, longe, panne … ; 104), produits partiellement
transformés (hachés, découpés-désossés … ; 26) ou transformés (saucisses, saucisson,
merguez, jambon cuit, jambon sec… ; 45). Le panel de produits couvert dans cette étude est
donc relativement important. Cependant, certaines catégories de prélèvements sont très peu
représentées ce qui montre la nécessité d’implémenter la base en souches issues de ces
catégories afin de tirer des conclusions plus pertinentes.
Sérotypage
Le sérotypage des différents isolats collectés a été réalisé par l’unité de Caractérisation et
Epidémiologie Bactérienne (CEB) de l’Afssa LERQAP de Maisons-Alfort. Dans le cadre de
notre étude, nous n’avons pas pu obtenir de résultats de sérotypage pour 7 isolats par les
méthodes de référence. Quatre souches n’étaient pas agglutinables et 3 présentaient un
sérotype dit incomplet dans le schéma de Kauffmann-White (SI 21:i ; SI 1,3,19:- ; SI 4,12:-
:1,2). Les résultats de sérotypage des isolats collectés lors de ce travail ont été intégrés dans
le cadre du réseau Salmonella auquel l’IFIP collabore depuis plusieurs années. Ce réseau
recense les prévalences des sérotypes de Salmonella présents dans les différentes filières
alimentaires en France. La surveillance réalisée les années passées montre une très faible
représentation de souches issues de la filière porcine, en particulier par rapport aux souches
d’origine aviaire, d’où un intérêt supplémentaire de la constitution d’une base de données
filière.
Les résultats obtenus pour l’ensemble des isolats collectés lors de cette étude sont
présentés dans le diagramme figure 1. Les figures 2 et 3 présentent respectivement la
diversité des sérotypes obtenue dans les entreprises E1 et E2.
Sur l’ensemble de l’étude, le sérotypage a mis en évidence 12 sérotypes différents. Les
principaux sérotypes retrouvés dans le panel de souches étudié sont le sérotype Derby
(44%) et le sérotype Typhimurium (39%), les autres sérotypes se partageant les 17%
restants, avec en troisième place Infantis (3%), puis Rissen et Kedougou (2%), London
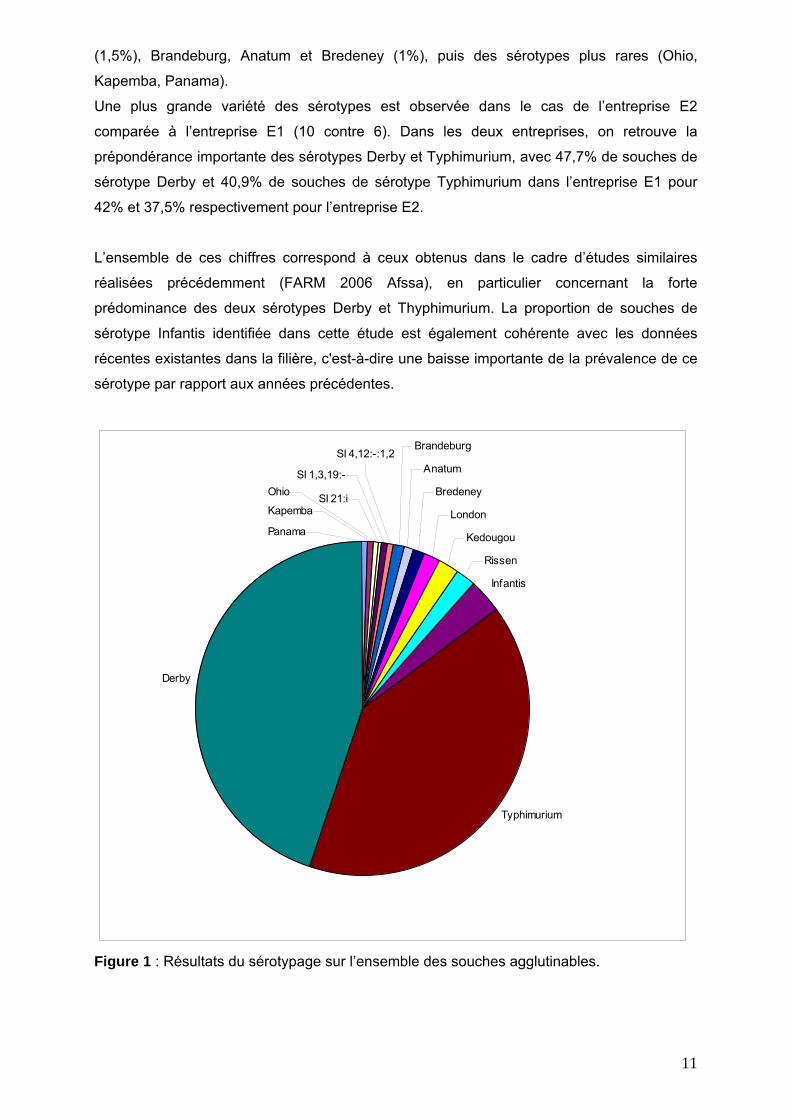
11
(1,5%), Brandeburg, Anatum et Bredeney (1%), puis des sérotypes plus rares (Ohio,
Kapemba, Panama).
Une plus grande variété des sérotypes est observée dans le cas de l’entreprise E2
comparée à l’entreprise E1 (10 contre 6). Dans les deux entreprises, on retrouve la
prépondérance importante des sérotypes Derby et Typhimurium, avec 47,7% de souches de
sérotype Derby et 40,9% de souches de sérotype Typhimurium dans l’entreprise E1 pour
42% et 37,5% respectivement pour l’entreprise E2.
L’ensemble de ces chiffres correspond à ceux obtenus dans le cadre d’études similaires
réalisées précédemment (FARM 2006 Afssa), en particulier concernant la forte
prédominance des deux sérotypes Derby et Thyphimurium. La proportion de souches de
sérotype Infantis identifiée dans cette étude est également cohérente avec les données
récentes existantes dans la filière, c'est-à-dire une baisse importante de la prévalence de ce
sérotype par rapport aux années précédentes.
Brandeburg
Anatum
Bredeney
London
Kedougou
Rissen
Infantis
Typhimurium
Derby
SI 1,3,19:-
SI 4,12:-:1,2
Ohio SI 21:iKapemba
Panama
Figure 1 : Résultats du sérotypage sur l’ensemble des souches agglutinables.

12
Derby
Ohio
Typhimurium
Non sérotypable
Brandenburg
Kedougou
Derby
Typhimurium
Rissen
LondonPanama
Kapemba
Bredeney
BrandeburgAnatumNon
sérotypables
Infantis
Figure 2 : Résultats du sérotypage pour
l’entreprise E1 Figure 3 : Résultats du sérotypage pour
l’entreprise E2
Pulsotypage
Le pulsotypage des isolats de Salmonella avec l’enzyme XbaI, a permis une bonne
discrimination entre les souches de même sérotype. Un nombre faible d’isolats (4) n’a
cependant pas pu être typé par cette méthode, le plus souvent s’agissant de sérotypes rares
et peu représentés lors de cette étude (Panama, Kapemba, Ohio).
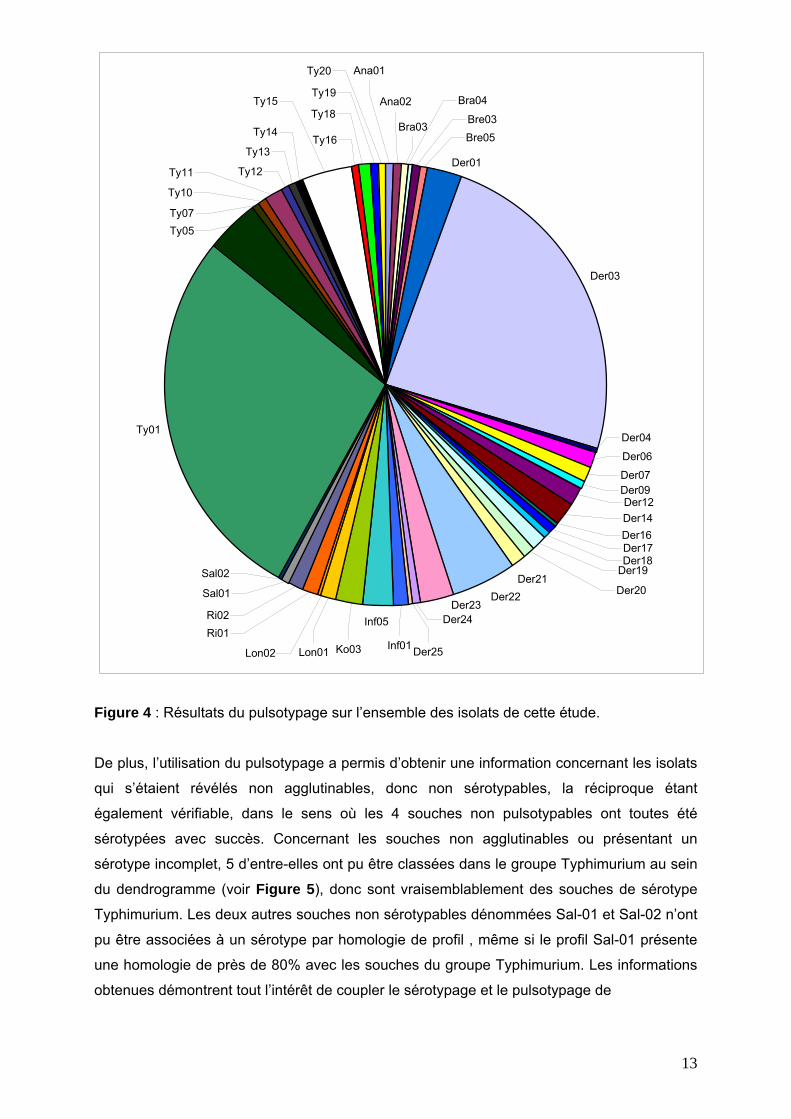
La Figure 4 résume les résultats obtenus pour l’ensemble des isolats reçus et analysés. Les
résultats du pulsotypage sont en accord avec les résultats du sérotypage, la quasi majorité
des pulsotypes pouvant facilement être classés dans les sérotypes précédemment obtenus
(voir Figure 5). Seule une souche identifiée comme Brandenburg par sérotypage n’a pu être
regroupée par la méthode du pulsotypage avec les autres souches Brandenburg. Un
deuxième sérotypage de cette souche sera réalisé afin de vérifier qu’il n’y ait pas eu d’erreur
lors du traitement de l’échantillon.
La méthode de pulsotypage se montre cependant beaucoup plus discriminante que le
sérotypage, puisque 46 pulsotypes ont été identifiés contre 12 pour la sérotypie. En
particulier, 18, 13, 2, 2, 2, 2, 2 et 2 pulsotypes différents ont été identifiés au sein des
sérotypes Derby, Typhimurium, Anatum, Bredeney, Brandenburg, Rissen, London et Infantis
respectivement. Un seul pulsotype a été identifié parmi les 4 souches de sérotype Kedougou
du panel.
Une autre information apportée par le dendrogramme obtenu dans cette étude révèle la forte
clonalité des isolats. En effet, on peut constater que les profils obtenus par exemple pour les
pulsotypes Der-04, , Der-06, et Der-16 sont très proches de celui obtenu pour le pulsotype
Der-03, révélant une parenté génétique importante (supérieure à 80%) donc peu de
mutations séparant un pulsotype d’un autre.
13
Der22
Inf05
Ty01
Ty12
Ty13
Ty14
Der03
Ty05
Ty15
Ty11
Ty07
Ty10
Ana01
Ana02 Bra04
Bra03Bre03
Der01
Bre05Ty16
Ty18
Ty19
Ty20
Der04
Der06
Der07
Der12Der09
Der16Der14
Der17Der18
Der19
Der20Sal02
Sal01
Ri02Ri01
Lon02 Ko03Lon01 Inf01
Der23
Der25
Der24
Der21
Figure 4 : Résultats du pulsotypage sur l’ensemble des isolats de cette étude.
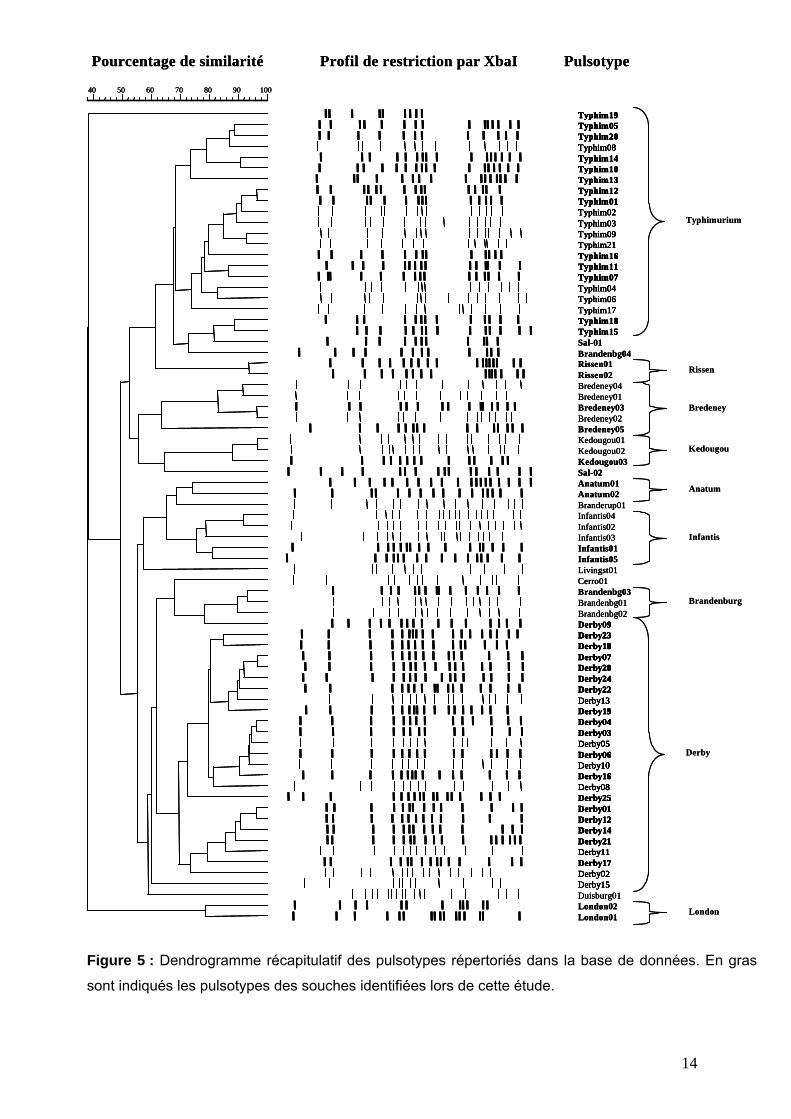
De plus, l’utilisation du pulsotypage a permis d’obtenir une information concernant les isolats
qui s’étaient révélés non agglutinables, donc non sérotypables, la réciproque étant
également vérifiable, dans le sens où les 4 souches non pulsotypables ont toutes été
sérotypées avec succès. Concernant les souches non agglutinables ou présentant un
sérotype incomplet, 5 d’entre-elles ont pu être classées dans le groupe Typhimurium au sein
du dendrogramme (voir Figure 5), donc sont vraisemblablement des souches de sérotype
Typhimurium. Les deux autres souches non sérotypables dénommées Sal-01 et Sal-02 n’ont
pu être associées à un sérotype par homologie de profil , même si le profil Sal-01 présente
une homologie de près de 80% avec les souches du groupe Typhimurium. Les informations
obtenues démontrent tout l’intérêt de coupler le sérotypage et le pulsotypage de
14
Figure 5 : Dendrogramme récapitulatif des pulsotypes répertoriés dans la base de données. En gras
sont indiqués les pulsotypes des souches identifiées lors de cette étude.
Typhim19Typhim05Typhim20Typhim08Typhim14Typhim10Typhim13Typhim12Typhim01Typhim02Typhim03Typhim09Typhim21Typhim16Typhim11Typhim07Typhim04Typhim06Typhim17Typhim18Typhim15Sal-01Brandenbg04Rissen01Rissen02Bredeney04Bredeney01Bredeney03Bredeney02Bredeney05Kedougou01Kedougou02Kedougou03Sal-02Anatum01Anatum02Branderup01Infantis04Infantis02Infantis03Infantis01Infantis05Livingst01Cerro01Brandenbg03Brandenbg01Brandenbg02Derby09Derby23Derby18Derby07Derby20Derby24Derby22Derby13Derby19Derby04Derby03Derby05Derby06Derby10Derby16Derby08Derby25Derby01Derby12Derby14Derby21Derby11Derby17Derby02Derby15Duisburg01London02London01
100908070605040
Pourcentage de similarité PulsotypeProfil de restriction par XbaI
Derby
Typhimurium
Bredeney
Rissen
Brandenburg
Infantis
Kedougou
Anatum
London
Typhim19Typhim05Typhim20Typhim08Typhim14Typhim10Typhim13Typhim12Typhim01Typhim02Typhim03Typhim09Typhim21Typhim16Typhim11Typhim07Typhim04Typhim06Typhim17Typhim18Typhim15Sal-01Brandenbg04Rissen01Rissen02Bredeney04Bredeney01Bredeney03Bredeney02Bredeney05Kedougou01Kedougou02Kedougou03Sal-02Anatum01Anatum02Branderup01Infantis04Infantis02Infantis03Infantis01Infantis05Livingst01Cerro01Brandenbg03Brandenbg01Brandenbg02Derby09Derby23Derby18Derby07Derby20Derby24Derby22Derby13Derby19Derby04Derby03Derby05Derby06Derby10Derby16Derby08Derby25Derby01Derby12Derby14Derby21Derby11Derby17Derby02Derby15Duisburg01London02London01
Typhim19Typhim05Typhim20Typhim08Typhim14Typhim10Typhim13Typhim12Typhim01Typhim02Typhim03Typhim09Typhim21Typhim16Typhim11Typhim07Typhim04Typhim06Typhim17Typhim18Typhim15
Typhim19Typhim05Typhim20Typhim08Typhim14Typhim10Typhim13Typhim12Typhim01Typhim02Typhim03Typhim09Typhim21Typhim16Typhim11Typhim07Typhim04Typhim06Typhim17Typhim18
Typhim19Typhim05Typhim20Typhim08Typhim14Typhim10Typhim13Typhim12Typhim01Typhim02Typhim03Typhim09Typhim21Typhim16Typhim11Typhim07Typhim04Typhim06Typhim17Typhim18Typhim15Sal-01Brandenbg04Rissen01Rissen02Bredeney04Bredeney01Bredeney03Bredeney02Bredeney05Kedougou01Kedougou02Kedougou03Sal-02Anatum01Anatum02Branderup01Infantis04Infantis02Infantis03Infantis01Infantis05Livingst01Cerro01Brandenbg03Brandenbg01Brandenbg02Derby09Derby23Derby18Derby07Derby20Derby24Derby22Derby13Derby19Derby04Derby03Derby05Derby06Derby10Derby16Derby08Derby25Derby01Derby12Derby14Derby21Derby11Derby17Derby02Derby15
Derby09Derby23Derby18Derby07Derby20Derby24Derby22Derby13Derby19Derby04Derby03Derby05Derby06Derby10Derby16Derby08Derby25Derby01Derby12Derby14Derby21Derby11Derby17Derby02Derby15
Derby23Derby18Derby07Derby20Derby24Derby22Derby13Derby19Derby04Derby03Derby05Derby06Derby10Derby16Derby08Derby25Derby01Derby12Derby14Derby21Derby11Derby17Derby02
Derby23Derby18Derby07Derby20Derby24Derby22Derby13Derby19Derby04Derby03Derby05Derby06Derby10Derby16Derby08Derby25Derby01Derby12Derby14Derby21Derby11Derby17Derby02Derby15Duisburg01London02London01
100908070605040 100908070605040
Pourcentage de similarité PulsotypeProfil de restriction par XbaI
Derby
Typhimurium
Bredeney
Rissen
Brandenburg
Infantis
Kedougou
Anatum
London

15
Salmonella pour constituer une base de données de souches finement caractérisées.
Lors de cette étude, nous avons remarqué la prépondérance de deux pulsotypes, Ty-01
(28% du total des souches analysées) et Der-03 (23.5%). Même si ces deux pulsotypes sont
retrouvés de manière majoritaire dans les deux entreprises, des disparités apparaissent
entre l’entreprise E1 et l’entreprise E2. Statistiquement, le profil Ty-01 serait plus présent
dans l’entreprise 1 que dans l’entreprise E2 (36,36% contre 19,64%), phénomène que l’on
ne retrouve pas pour le pulsotype Der-03. Cette donnée pourrait montrer une sensibilité plus
grande des souches de type Ty-01 aux contraintes présentes dans les ateliers de
transformation par rapport aux souches de type Der-03.
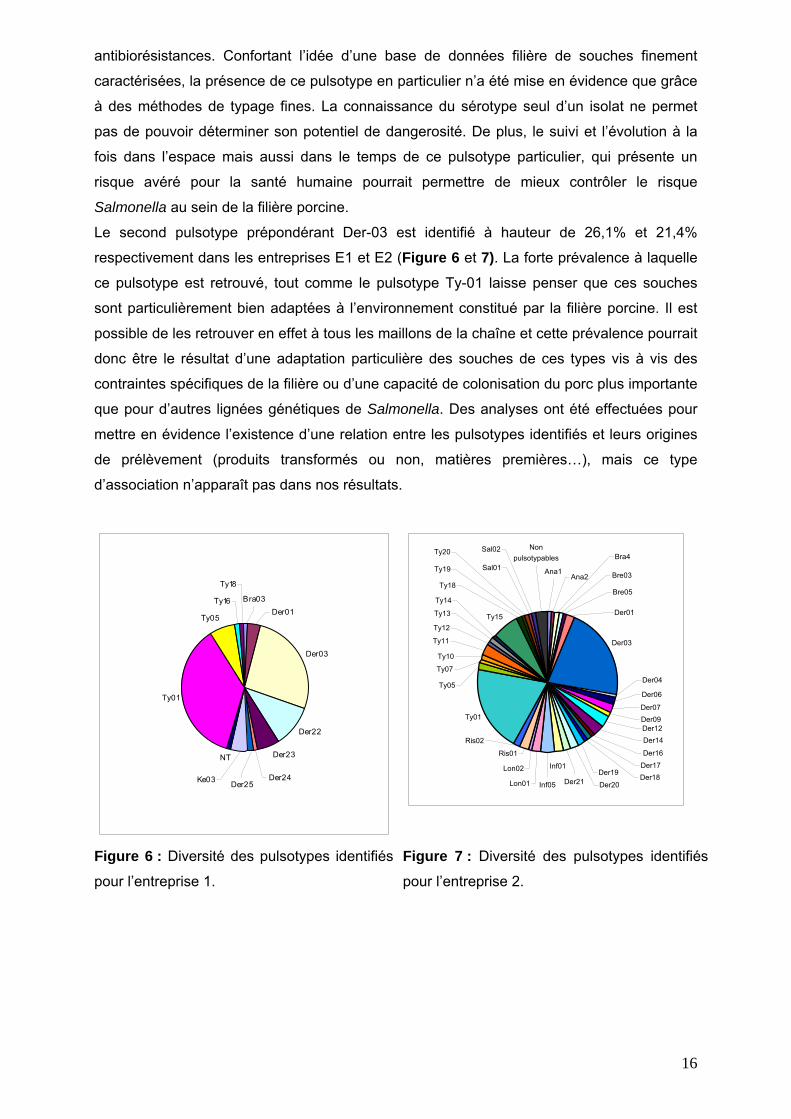
Les autres pulsotypes identifiés ont une incidence comprise entre 0,5% et 10% en moyenne
dans les deux entreprises (Figures 6 et 7). De manière encore plus flagrante que pour le
sérotypage, une différence notable entre les deux types d’ateliers se retrouve dans la
diversité des pulsotypes identifiés dans chaque entreprise : 12 types dans l’entreprise E1 et
39 dans l’entreprise E2. Cette différence importante peut s’expliquer de différentes
manières :
- Une plus grande variété dans les apports de matières premières au sein des ateliers de
transformation, comparativement à l’entreprise 1 d’abattage-découpe qui abat des animaux
provenant toujours des mêmes élevages .
- D’autres sources d’entrée propres à une entreprise de transformation (emballages,
ingrédients…).
Cependant, quantitativement, 75% des souches identifiées dans l’entreprise E1 sont
également identifiées dans l’entreprise E2, montrant ainsi que la matière première resterait la
principale source de contamination dans l’entreprise. Ces résultats devront être confirmés
par l’analyse d’un panel plus important de souches.
Le profil dénommé Ty-01 est très souvent associé aux souches Salmonella enterica
sérotype Typhimurium lysotype DT104 (type déterminé par l’utilisation de virus bactériens
spécifiques). Les souches Salmonella Typhimurium de lysotype DT104 sont fréquemment
impliquées dans l’apparition de salmonelloses multirésistantes aux antibiotiques chez
l’homme. Les études portant sur ce lysotype en particulier ont montré une augmentation
importante dans l’apparition de résistances à de nombreux antibiotiques, la majorité vis à vis
d’antibiotiques classiques (ampicilline, chloramphénicol, tétracyline, sulfonamides,
streptomycine), certaines souches arborant une résistance vis à vis de 16 antibiotiques différents (Malorny, 2001).
Ce pulsotype est retrouvé pour 36,4% des souches analysées dans l’entreprise 1 et 19.6%
des souches issues de l’entreprise 2. Il se pose donc le problème de la gestion du risque vis
à vis de ces isolats potentiellement pathogènes et porteurs de plus de nombreuses
16
antibiorésistances. Confortant l’idée d’une base de données filière de souches finement
caractérisées, la présence de ce pulsotype en particulier n’a été mise en évidence que grâce
à des méthodes de typage fines. La connaissance du sérotype seul d’un isolat ne permet
pas de pouvoir déterminer son potentiel de dangerosité. De plus, le suivi et l’évolution à la
fois dans l’espace mais aussi dans le temps de ce pulsotype particulier, qui présente un
risque avéré pour la santé humaine pourrait permettre de mieux contrôler le risque
Salmonella au sein de la filière porcine.
Le second pulsotype prépondérant Der-03 est identifié à hauteur de 26,1% et 21,4%
respectivement dans les entreprises E1 et E2 (Figure 6 et 7). La forte prévalence à laquelle
ce pulsotype est retrouvé, tout comme le pulsotype Ty-01 laisse penser que ces souches
sont particulièrement bien adaptées à l’environnement constitué par la filière porcine. Il est
possible de les retrouver en effet à tous les maillons de la chaîne et cette prévalence pourrait
donc être le résultat d’une adaptation particulière des souches de ces types vis à vis des
contraintes spécifiques de la filière ou d’une capacité de colonisation du porc plus importante
que pour d’autres lignées génétiques de Salmonella. Des analyses ont été effectuées pour
mettre en évidence l’existence d’une relation entre les pulsotypes identifiés et leurs origines
de prélèvement (produits transformés ou non, matières premières…), mais ce type
d’association n’apparaît pas dans nos résultats.
Der03
Der22
Der23NT
Ty01
Ty05
Ty16
Ty18
Der24Ke03
Der01Bra03
Der25
Der03
Lon01
Lon02
Ris01
Ty01
Ty15
Der18Der17
Der16
Der14Der12Der09Der07
Der04
Bra4
Der06
Der21
Inf01
Inf05
Ty14
Ty13
Ty12
Ty11
Ty10
Ty07
Ty05
Ris02
Ty20
Ty19
Ty18
Non pulsotypables
Sal02
Der01
Bre03
Bre05
Ana1Ana2
Sal01
Der20
Der19
Figure 6 : Diversité des pulsotypes identifiés
pour l’entreprise 1. Figure 7 : Diversité des pulsotypes identifiés
pour l’entreprise 2.
17
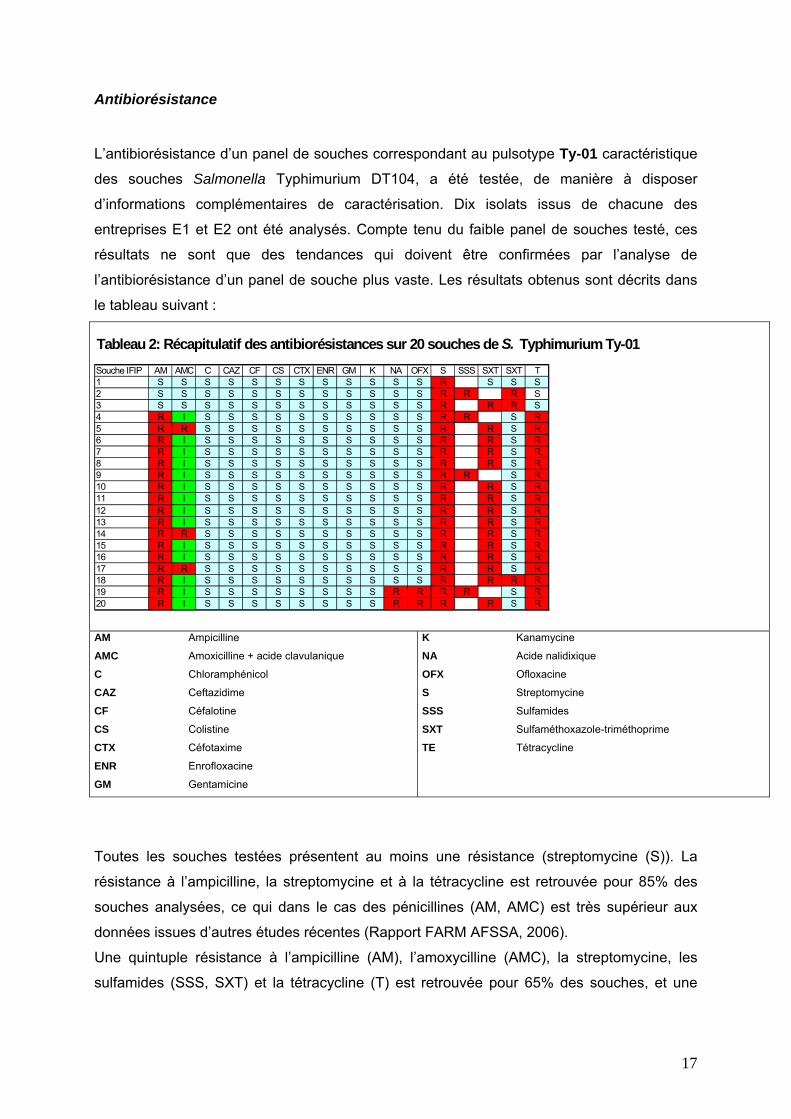
Antibiorésistance
L’antibiorésistance d’un panel de souches correspondant au pulsotype Ty-01 caractéristique
des souches Salmonella Typhimurium DT104, a été testée, de manière à disposer
d’informations complémentaires de caractérisation. Dix isolats issus de chacune des
entreprises E1 et E2 ont été analysés. Compte tenu du faible panel de souches testé, ces
résultats ne sont que des tendances qui doivent être confirmées par l’analyse de
l’antibiorésistance d’un panel de souche plus vaste. Les résultats obtenus sont décrits dans
le tableau suivant :
Tableau 2: Récapitulatif des antibiorésistances sur 20 souches de S. Typhimurium Ty-01Souche IFIP AM AMC C CAZ CF CS CTX ENR GM K NA OFX S SSS SXT SXT T1 S S S S S S S S S S S S R S S S2 S S S S S S S S S S S S R R R S3 S S S S S S S S S S S S R R R S4 R I S S S S S S S S S S R R S R5 R R S S S S S S S S S S R R S R6 R I S S S S S S S S S S R R S R7 R I S S S S S S S S S S R R S R8 R I S S S S S S S S S S R R S R9 R I S S S S S S S S S S R R S R10 R I S S S S S S S S S S R R S R11 R I S S S S S S S S S S R R S R12 R I S S S S S S S S S S R R S R13 R I S S S S S S S S S S R R S R14 R R S S S S S S S S S S R R S R15 R I S S S S S S S S S S R R S R16 R I S S S S S S S S S S R R S R17 R R S S S S S S S S S S R R S R18 R I S S S S S S S S S S R R R R19 R I S S S S S S S S R R R R S R20 R I S S S S S S S S R R R R S R
AM Ampicilline
AMC Amoxicilline + acide clavulanique
C Chloramphénicol
CAZ Ceftazidime
CF Céfalotine
CS Colistine
CTX Céfotaxime
ENR Enrofloxacine
GM Gentamicine
K Kanamycine
NA Acide nalidixique
OFX Ofloxacine
S Streptomycine
SSS Sulfamides
SXT Sulfaméthoxazole-triméthoprime
TE Tétracycline
Toutes les souches testées présentent au moins une résistance (streptomycine (S)). La
résistance à l’ampicilline, la streptomycine et à la tétracycline est retrouvée pour 85% des
souches analysées, ce qui dans le cas des pénicillines (AM, AMC) est très supérieur aux
données issues d’autres études récentes (Rapport FARM AFSSA, 2006).
Une quintuple résistance à l’ampicilline (AM), l’amoxycilline (AMC), la streptomycine, les
sulfamides (SSS, SXT) et la tétracycline (T) est retrouvée pour 65% des souches, et une

18
résistance à 7 antibiotiques a été constatée pour 2 souches (ampicilline, amoxycilline,
streptomycine, sulfamides, tétracycline, acide nalidixique (NA) et ofloxacine (OFX)).
La sensibilité aux céfalosporines (CAZ, CF, CTX ) (100%) correspond à ce qui a été mis en
évidence précédemment dans la filière porc. De la même manière, la sensibilité aux
fluoroquinolones (ENR, NA, OFX) reste très élevée, ainsi que pour la gentamicine (GM), la
kanamycine (K) et le chloramphénicol (C). Dans le cas de la tétracyline, la résistance
retrouvée correspond également aux chiffres publiés précédemment et il semblerait qu’une
augmentation de la sensibilité aux sulfamides soit détectable par rapport aux années
précédentes (FARM, AFSSA, 2006).
Au contraire, la prévalence des résistances observées s’avère supérieure dans le cas de la
streptomycine (100% contre 57,6% en 2004 dans la filière porc) ainsi que pour les
pénicillines (85% contre 21% pour l’ampicilline et 1% pour l’amoxyciline).
Cependant nous n’avons pas mis en évidence le profil de pentarésistance « classique »
ASCTSu arboré par un grand nombre d’isolats de lysotype DT104 (prévalence de 15,4% en
2004, en augmentation par rapport aux années précédentes), qui correspond à la résistance
à l’ampicilline, la streptomycine, le chloramphénicol, la tétracycline et aux sulfamides.
Ces profils de résistance peuvent être associés à une utilisation importante de ces
antibiotiques dans la filière porcine, ce qui entraîne l’émergence de résistances bactériennes
vis-à-vis de ces additifs et/ou médicaments vétérinaires dans l’ensemble de la filière avec les
problèmes subséquents en thérapeutique humaine.
De plus, il serait intéressant d’étendre la surveillance des antibiorésistances à des souches
de pulsotypes prépondérants dans la filière (en particulier le pulsotype Der-03). Cela
permettrait de déterminer s’il existe une corrélation entre les antibiorésistances arborées par
les souches bactériennes les plus représentées, donc de potentielles voies de transmission
des résistances entre souches dans la filière. Pour répondre à ces interrogations et valider
les tendances décrites ici, un panel plus important d’isolats devra être analysé.

19
Conclusions et Perspectives
L’intérêt d’établir une base de données Salmonella pour la filière est multiple. Une fois
implémenté, cet outil peut permettre de mieux appréhender l’épidémiologie du germe dans
l’ensemble de la filière, et par conséquent réduire la prévalence de Salmonella afin de
satisfaire aux exigences de la directive zoonose d’une part et du règlement 2073/2005
concernant les critères microbiologiques applicables aux denrées alimentaires d’autre part.
Lors de cette étude, les résultats obtenus en sérotypie sont conformes avec ceux présentés
dans des études récentes (Rapport AFSSA 2006) avec deux sérotypes majeurs dans la
filière porc qui sont Derby et Typhimurium.
La mise en évidence de deux pulsotypes fortement représentés dans l’ensemble des isolats
collectés (Der-03 et Ty-01) montre une potentielle adaptation de ces deux types à la filière
porcine, ce qui expliquerait leur sur-représentation dans le cadre de cette étude.
Le point critique dans les analyses effectuées est que le profil Ty-01 identifié fait partie des
profils généralement associés aux souches de type DT104. Ces dernières correspondent à
des souches de plus en plus identifiées en médecine humaine depuis quelques années et
qui présentent généralement de nombreuses résistances à différentes familles
d’antibiotiques.
L’analyse de l’antibiorésistance d’un petit panel de souches Ty-01 isolées dans cette étude a
confirmé un taux important de résistance aux antibiotiques les plus courants, même si
l’échantillonnage testé n’était pas suffisamment représentatif pour tirer d’autres conclusions
et qu’il faudra mener cette analyse sur un panel plus large pour confirmer les tendances qui
ont été observées.
Il ressort de ce travail que certaines souches de Salmonella qui sont d’intérêt majeur en
médecine humaine sont retrouvées à une prévalence très forte ( 28% des souches sont de
type Ty-01). Leur présence pouvant être considérée comme un danger avéré, la surveillance
de ce type de souches en particulier présente un intérêt fort pour la gestion du risque
Salmonella au sein de la filière.
Pour cela, il est nécessaire de continuer à incrémenter cette base de données afin d’estimer
l’évolution de la représentativité du type Ty-01 au sein de la filière et de son profil
d’antibiorésistance. Par ailleurs, il serait utile d’analyser un plus grand panel de souches afin
d’obtenir une meilleure représentativité de la base de données concernant les souches de
sérotypes et/ou pulsotypes rarement identifiés ou les souches issues d’origines de
prélèvement peu représentées. L’implémentation pérenne de la base de données permettra
d’obtenir une vision précise de la prévalence de Salmonella dans la filière à un moment

20
donné, tout en permettant un meilleur suivi dans le temps de l’évolution de sa prévalence,
donc un suivi de l’efficacité des mesures visant à améliorer la situation dans la filière.
Cette base de données filière permet également de se placer de manière complémentaire
par rapports aux autres bases existantes constituées soit à partir d’isolats humains (CNR
Pasteur) soit à partir d’isolats majoritairement issus de la filière aviaire (AFSSA)
(Swanenburg, 2001 ; Fravalo, 2002), et donc de contribuer à une meilleure connaissance de
l’épidémiologie de cette bactérie dans la filière porcine.

21
Bibliographie
Brisabois, A., Frémy, S., Moury, F., Oudart, C., Piquet, C., and Pires Gomes, C. (1997).
Inventaires des Salmonella 1994-1995, Maisons-Alfort : CNEVA Edition.
Brisabois, A., Danan, C., Frémy, S., Granier, S., Moury, F., Oudart, C., Piquet, C., and Pires
Gomes, C. (2006). Inventaire du réseau Salmonella 2004, Sérotypage et sensibilité aux
antibiotiques.
Fisher, I.S.T., and Gill, O.N. (2001). Réseaux de surveillance internationaux et principes de
collaboration. Eurosurveillance 6 (2), 17-21.
FARM 2003-2004, Rapport du programme français de surveillance de l’antibiorésistance
des bactéries d’origine animale, AFSSA août 2006.
Fravalo, P., Beloeil, P.A., Fablet, C., Hascouet, Y., Salvat, G., and Madec, F. (2002). Risk
factors of Salmonella excretion by finishing pigs. Salmonella and salmonellosis, 289-296.
Giovannacci, I., Queguiner, S., Ragimbeau, C., Salvat, G., Vendeuvre, J.L., Carlier, V., and
G. Ermel. (2001). Tracing of Salmonella spp. in two pork slaughter and cutting plants using
serotyping and macrorestriction genotyping. Journal of Applied Microbiology 90, 131-147.
Malorny, B, Schroeters, A, Bunge, C, Hoog, B, Steinbeck, A, R.Helmuth. (2001). Evaluation
of molecular typing methods for Salmonella enterica serovar Typhimurium DT104 isolated in
Germany from healthy pigs. Vetenary Research 32, 119-219.
Swaminathan, B., Barrett, J., Hunter, S.B., Tauxe, R.V., and the CDC Pulsenet Task Force.
(2001). PulseNet: the molecular subtyping network for foodborne bacterial disease
surveillance, United States. Emerging Infectious Diseases 7 (3), 382-389.
Swanenburg, M., Urlings, H.A.P.,. Snijders, J.M.A, Keuzenkamp, D.A., and F. van Knapen
(2001). Salmonella in slaughter pigs: prevalence, serotypes and critical control points during
slaughter in two slaughterhouses. International Journal of Food Microbiology. 70 (3), 243-254
Weill, F.-X. Lailler, R., ans Brisabois, A. 2004. Tendances récentes de la résistance aux
antibiotiques des Salmonella d’origines animale et humaine. BEH n°32-33.

22
Antenne Rennes – Le Rheu La Motte au Vicomte B.P. 35104 - 35651 Le Rheu Cedex Tél. : + 33 (0)2 99 60 98 20 - Fax : + 33 (0)2 99 60 93 55
[email protected] - www.ifip.asso.fr
Antenne Toulouse 34 bd de la Gare - 31500 TOULOUSE
Tél. : + 33 (0)5 62 16 61 70 - Fax : + 33 (0)5 61 54 32 63
Antenne Maisons-Alfort 7, avenue du Général de Gaulle – 94704 MAISONS-ALFORT
Tél. : + 33 (0)1 43 68 57 85 - Fax : + 33 (0)1 43 76 07 20
Avec le concours financier de INAPORC
Étude financée au titre du programme national de développement agricole et rural
© IFIP 2008