by henry a. harbury department gation with a leeds and northrup type r galvanometer. in the ... at...
TRANSCRIPT

OXIDATION-REDUCTION POTENTIALS OF HORSERADISH PEROXIDASE*
BY HENRY A. HARBURY
(From the Department of Biochemistry, Yale University, New Haven, Connecticut)
(Received for publication, October 15, 1956)
Among the most widely applicable, and potentially most ‘incisive, studies of the linked functions (1) of heme proteins are those based on the investi- gation of reversible oxidation-reduction reactions. The determination of linked functions by oxidation-reduction potential measurements (2, 3) has been reported thus far for two different heme proteins, hemoglobin and cytochrome c. Measurements were made in each case of the variation of the potentials with pH. The data of Taylor and Hastings (4) on horse hemoglobin have played a key role in analyses of the heme-heme and heme- protein interactions in that system (1, 5, 6). The measurements by Rod- key and Ball (7) and by Paul (8) of the oxidation-reduction potentials of cytochrome c from heart muscle (cow and horse) have provided evidence for the existence in their preparations of a linked proton function, operating near the physiological range of pH, that had escaped detection by other modes of investigation.
Effects of variables other than pH on the oxidation-reduction potentials of heme proteins have received little attention thus far. A systematic investigation of the influence of a selected group of variables seems desir- able, for it may be expected to yield quantitative information about the interaction between a protein and its prosthetic group and the changes in this interaction which attend the occurrence of various linked reactions. Horseradish peroxidase is at present the heme protein of choice for such a systematic study. Its properties permit, in principle, a systematic evalua- tion of the effects on the oxidation-reduction potential of factors such as pH, temperature, ionic strength, dissociation into free prosthetic group and protein, interaction of the protein-bound prosthetic group with small molecules and ions, and specific modifications of the prosthetic group and the protein.
An investigation of horseradish peroxidase has been undertaken, with the first objective the evaluation of the free energy, enthalpy, and entropy changes of the oxidation-linked proton equilibria in the system ferri HRP-
* This work has been supported by grants from the National Science Foundation, the Rockefeller Foundation, the James Hudson Brown Memorial Fund of the Yale University School of Medicine, and Eli Lilly and Company.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1010 HORSERADISH PEROXIDASE
ferro HRP.’ The purpose of this report is to present the results of the first measurements, which have been of the ferri HRP-ferro HRP system at 30”. Data are reported for the range of pH from 4 to 11. Preliminary measurements, made at values of pH near 7, were presented earlier (9).
EXPERIMENTAL
MaterialsRecrystallized HRP was prepared according to the procedure of Theorell (10). The preparation was carried out by Dr. Karl-Gustav Paul in the laboratories of the Biochemical Department of the Medical Nobel Institute, Stockholm. The very generous gift by Dr. Paul and Pro- fessor Hugo Theorell of the material used in this study is deeply appre- ciated.
Methyl viologen, benzyl viologen, and snthraquinone-2,6disulfonate were obtained from The British Drug Houses, Ltd., 2,6dichlorophenol- indophenol from The Matheson Company, Inc., and phenosafranine from the National Aniline Division, Allied Chemical and Dye Corporation. These dyes were all used without further purification. The rosinduline 2G used was the kind gift of Professor W. Mansfield Clark.
The following buffers have been used: citrate, phosphate, tris(hydroxy- methyl)aminomethane, glycine, @-alanine, and L-lysine. Ionic strengths have varied from 0.07 to 0.19. The use of sodium ions was avoided in the preparation of alkaline buffers.
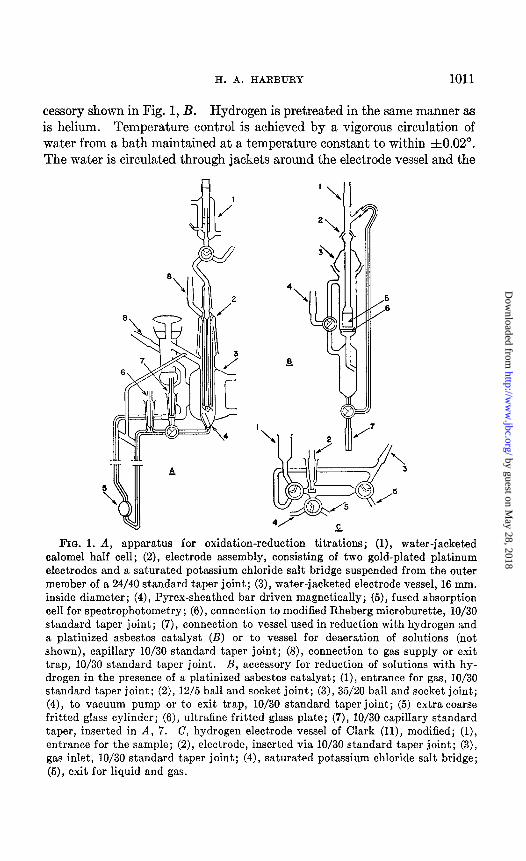
Apparatus-The titration assembly shown in Fig. 1 has proved par- ticularly suitable for the studies reported in this paper. Its use permits the concurrent collection of potentiometric and spectrophotometric data throughout the course of a titration. Both oxidative and reductive titra- tions can be performed conveniently with 5 to 10 ml. samples of solutions as dilute as 1O-6 M. Each of two gold-plated platinum electrodes may be read against the saturated calomel reference electrode. The titrating agent is delivered from a microburette. Deaeration and movement of the solution from one part of the apparatus to another are effected by the use of helium. Prior to its entry into the titration assembly, the helium is passed first through a column of copper maintained at 400”, then through water. A magnetically driven, Pyrex-sheat,hed bar is used to stir the solu- tion in the electrode vessel. Reduction of a solution with hydrogen in the presence of a platinized asbestos catalyst is performed by the use of the ac-
1 The term horseradish peroxidase is abbreviated in this paper as HRP. The pre- fixes ferri and ferro are used to designate states of oxidation only. Ferri HRP repre- sents the state of the free enzyme in the absence of strong oxidizing or reducing agents. Ferro HRP represents what is often termed reduced peroxidase. The pre- fixes ferri and ferro and the terms “ferric” and “ferrous” used occasionally to desig- nate states of oxidation are not intended to indicate a particular electronic configura- tion.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
H. A. HARBURY 1011
cessory shown in Fig. 1, B. Hydrogen is pretreated in the same manner as is helium. Temperature control is achieved by a vigorous circulation of water from a bath maintained at a temperature constant to within ~1~0.02”. The water is circulated through jackets around the electrode vessel and the
FIG. 1. A, apparatus for oxidation-reduction titrations; (l), water-jacketed calomel half cell; (2), electrode assembly, consisting of two gold-plated platinum electrodes and a saturated potassium chloride salt bridge suspended from the outer member of a 24/40 standard taper joint; (3), water-jacketed electrode vessel, 16 mm. inside diameter; (4), Pyrex-sheathed bar driven magnetically; (5), fused absorption cell for spectrophotometry; (6), connection to modified Rheberg microburette, lo/30 standard taper joint; (7), connection to vessel used in reduction with hydrogen and a platinized asbestos catalyst (B) or to vessel for deaeration of solutions (not shown), capillary lo/30 standard taper joint; (S), connection to gas supply or exit trap, lo/30 standard taper joint. B, accessory for reduction of solutions with hy- drogen in the presence of a platinized asbestos catalyst; (l), entrance for gas, IO/30 standard taper joint; (2), 12/5 ball and socket joint; (3), 35/20 ball and socket joint; (4), to vacuum pump or to exit trap, IO/30 standard taper joint; (5) extra coarse fritted glass cylinder; (6), ultrafine fritted glass plate; (7), IO/30 capillary standard taper, inserted in A, 7. C, hydrogen electrode vessel of Clark (ll), modified; (l), entrance for the sample; (2), electrode, inserted via lo/30 standard taper joint; (3), gas inlet, lo/30 standard taper joint; (4), saturated potassium chloride salt bridge; (5), exit for liquid and gas.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1012 HORSERADISH PEROXIDASE
calomel half cell and through coils placed in the cuvette compartment of the spectrophotometer.
Spectrophotometric measurements have been made with a Beckman model DU spectrophotometer, operated both manually and as a recording spectrophotometer by use of the Spectracord recording attachment manu- factured by the Perkin-Elmer Corporation.
Oxidation-reduction potentials have been measured both manually and on a semiautomatic basis with recording equipment. In the manual meas- urements, a Leeds and Northrup type Kl potentiometer has been used in conjunction with a Leeds and Northrup type R galvanometer. In the self-recording arrangement, the type Kl potentiometer has been used in conjunction with an Applied Physics Corporation model No. 30 vibrating reed electrometer and a Varian Associates model No. G-10 recorder. The recorder has been calibrated at frequent intervals with the use of the type Kl potentiometer and a Weston standard cell. Standard cells have been kept in an insulated chamber, and the standard cell used with the potenti- ometer has been measured periodically against two other Weston standard cells maintained for purposes of reference.
For the determination of pH, both hydrogen and glass electrodes have been used. Hydrogen electrode measurements have been made with a slightly modified version (Fig. 1, C) of the electrode developed by Clark (11). Glass electrode measurements have been performed with a Mac- Innes-Belcher (12) type electrode manufactured by the Cambridge Instru- ment Company, Inc. Both electrode systems have been kept in an air bath maintained at 30” f 0.03”, with the glass electrode assembly carefully shielded. Hydrogen electrode potentials have been measured in the same manner as those of other oxidation-reduction systems. Potentials from the glass electrode system have been measured either manually with the type Kl potentiometer, the vibrating reed electrometer serving as a null- point instrument, or have been determined with the use of recording equip- ment in the same fashion as were oxidation-reduction potentials.
Procedures-All of the oxidation-reduction potentials reported in this paper are based on determinations of complete titration curves. Both oxidative and reductive titrations have been performed. The samples titrated have normally been between 5 and 10 ml. in volume and between 2 X 1O-6 M and 4 X 1O-6 M in concentration of peroxidase. Mediator was present at a total molarity one-fortieth that of the peroxidase. In experiments performed below pH 10.5, solutions were brought to the de- sired pH prior to their entry into the titration assembly. In -the case of two reductive titrations performed at higher pH values, the solution under study was kept at neutral pH until after completion of the preliminary deoxygenation procedure, in order that the period of exposure of the en- zyme to a strongly alkaline solution might be kept to a minimum.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

H. A. HARBURY 1013
Special care had to be taken to reduce trace concentrations of oxygen to levels consonant with the accurate determination of potentials. Helium was used to deaerate solutions and to maintain an inert atmosphere during titration. Both oxidative and reductive titrations were preceded by ex- haustive deaeration of the apparatus and of the sample to be titrated. In oxidative titrations, oxygen remaining in solution was removed in the course of the preparatory reduction of ferri HRP with hydrogen in the presence of a platinieed asbestos catalyst. Residual oxygen on the sur- faces of the apparatus was exposed to action by the reduced solution, which subsequently was again reduced, prior to titration. In reductive titrations, residual oxygen was subjected to preliminary titration with reducing agent, under close potentiometric and spectrophotometric control, before titration of HRP was begun. A stationary atmosphere of helium was used in both oxidative and reductive titrations, fresh helium being admitted into the titration apparatus only to effect transport of the solution.
Methyl viologen and sodium dithionite were used as reducing agents and potassium ferricyanide and 2,6-dichlorophenol-indophenol as oxidiz- ing agents. The semiquinone of methyl viologen was prepared in unbuf- fered solution of approximately pH 10, by reduction of the oxidized form with hydrogen in the presence of platinized asbestos. Solutions of sodium dithionite were prepared by dissolving the solid in 1O-3 M sodium hydroxide which had previously been deaerated by boiling and by treatment with helium. Solutions of the oxidizing agents were thoroughly deaerated with helium before storage in the microburette.
Mediators were selected from among electromotively active systems with mid-point potentials close to those of the system under study. In a few experiments, two mediators were present in the sample subjected to titra- tion. Their total concentration was the same as that used in the other titrations.
Spectrophotometric observations were made as desired throughout the course of an experiment. Enzymic activity was estimated by determina- tion of the “purpurogallin number” (13). It was not routinely determined at the conclusion of every titration, but was measured sufficiently often to show that, except at the extremes of the pH range of these studies, the conditions to which HRP was exposed did not lead to significant change in its activity.
The pH of the solutions titrated was determined both before and after titration. Measurements were made with the glass electrode. Readings were interpolated between those obtained with buffer solutions of approxi- mately the same ionic strength as that of the sample titrated, the pH values of which had been established by measurements with the hydrogen elec- trode.
The standard of reference was a 0.05 M solution of potassium hydrogen
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1014 HORSERADISH PEROXIDASE
phthalate, to which was assigned a pH value of 4.016 at 30” (14). All calomel electrodes were maintained continuously at 30”. Their potentials were determined periodically by measurement against the reference stand- ard.
Results
Ferri HRP-ferro HRP was found to be an “electromotively sluggish” system, and in this respect it resembles the other heme protein systems that have been studied. Satisfactory potentials have been obtained only in the presence of a mediator. Four systems have served as mediator in the studies reported here: phenosafranine, rosinduline 2G, anthraquinone- 2,6-disulfonate, and benzyl viologen. All display a pH dependency of potentials (15-18) quite different from that found for ferri HRP-ferro HRP, but at various values of pH have potentials sufficiently close to that of the ferri HRP-ferro HRP system to afford effective mediator action. The one most used has been phenosafranine, which has been found to be suitable over a wide range of pH.
In the presence of an appropriate mediator, equilibrium between elec- trodes and the ferri HRP-ferro HRP system is usually rapidly established. Along the central portion of a titration curve, stable potentials may nor- mally be obtained in a matter of minutes. At very low and high states of reduction, however, equilibration is a slow process, often requiring hours.
The semiquinone of methyl viologen (18) has proved to be an effective and convenient agent for the reductive titration of ferri HRP. Over the range of pH in which there is a sufficient difference between the potentials of the half reduced methyl viologen-semiquinone of methyl viologen and ferri HRP-ferro HRP systems, its use has represented the technique of choice. A stronger reducing agent, sodium dithionite, has long been used to prepare ferro HRP, and, with proper precautions, its use in the reductive titration of ferri HRP has proved to be satisfactory. Methyl viologen has been used in reductive titrations at values of pH ranging from 4 to 10. Titrations with sodium dithionite have been performed near pH 7 and in solutions of high pH. An advantage in the use of these two reducing agents is the fact that there is no interference with the spectrophotometric ob- servation of the HRP system in the visible region of the spectrum.
Oxidative titrations have been performed with both 2,6-dichlorophenol- indophenol and potassium ferricyanide as oxidizing agents. The samples titrated were prepared by reduction with hydrogen in the presence of platinized asbestos. The solutions reduced contained ferri HRP and mediator in the 40: 1 ratio of concentrations that has been used in all ti- trations. Theorell (19) has reported that, in contrast to the reduction of ferrihemoglobin with hydrogen, which could be caused to proceed rapidly
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
H. A. HARBURY 1015
by addition to the reduction system of phenosafranine at 0.04 the molarity of the hemoglobin, the reduction of ferri HRP with hydrogen and platin- ized asbestos was found to require at least an equimolar concentration of
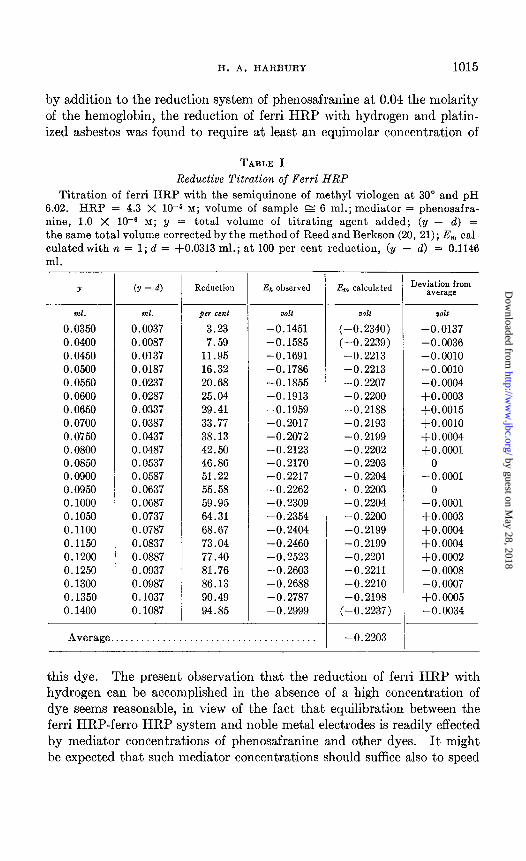
TABLE I
Reductive Titration of Ferri HRP Titration of ferri HRP with the semiquinone of methyl viologen at 30” and pH
6.02. HRP = 4.3 X low5 M; volume of sample E 6 ml.; mediator = phenosafra- nine, 1.0 X 1O-B M; y = total volume of titrating agent added; (y - d) = the same total volume corrected by the method of Reed and Berkson (20, 21); Em cal- culatedwith n = 1; d = +0.0313 ml.; at 100 per cent reduction, (y - d) = 0.1146 ml.
Y (Y - 4 Reduction .&, observed Em calculated Deviation from average
Td. ml. per cent VOll VOl2 volt
0.0350 0.0037 3.23 -0.1451 (-0.2340) -0.0137 0.0400 0.0087 7.59 -0.1585 (-0.2239) -0.0036 0.0450 0.0137 11.95 -0.1691 -0.2213 -0.0010 0.0500 0.0187 16.32 -0.1786 -0.2213 -0.0010 0.0550 0.0237 20.68 -0.1855 -0.2207 -0.0004 0.0600 0.0287 25.04 -0.1913 -0.2200 +0.0003 0.0650 0.0337 29.41 -0.1959 -0.2188 $0.0015 0.0700 0.0387 33.77 -0.2017 -0.2193 +0.0010 0.0750 0.0437 38.13 -0.2072 -0.2199 +0.0004 0.0800 0.0487 42.50 -0.2123 -0.2202 +0.0001 0.0850 0.0537 46.86 -0.2170 -0.2203 0 0.0900 0.0587 51.22 -0.2217 -0.2204 -0.0001 0.0950 0.0637 55.58 -0.2262 -0.2203 0 0.1000 0.0687 59.95 -0.2309 -0.2204 -0.0001 0.1050 0.0737 64.31 -0.2354 -0.2200 +0.0003 0.1100 0.0787 68.67 -0.2404 -0.2199 $0.0004 0.1150 0.0837 73.04 -0.2460 -0.2199 $0.0004 0.1200 0.0887 77.40 -0.2523 -0.2201 +o .0002 0.1250 0.0937 81.76 -0.2603 -0.2211 -0.0008 0.1300 0.0987 86.13 -0.2688 -0.2210 -0.0007 0.1350 0.1037 90.49 -0.2787 -0.2198 +0.0005 0.1400 0.1087 94.85 -0.2999 (-0.2237) -0.0034
Average. -0.2203
--
this dye. The present observation that the reduction of ferri HRP with hydrogen can be accomplished in the absence of a high concentration of dye seems reasonable, in view of the fact that equilibration between the ferri HRP-ferro HRP system and noble metal electrodes is readily effected by mediator concentrations of phenosafranine and other dyes. It might be expected that such mediator concentrations should suffice also to speed
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
1016 HORSERADISH PEROXIDASE
the reduction of ferri HRP with hydrogen, provided the dye is not de- stroyed (15) or immobilized under the conditions at which reduction is attempted.
The potentials determined in the course of both oxidative and reductive titrations agree well with those predicted for a reversible 1 electron oxida-
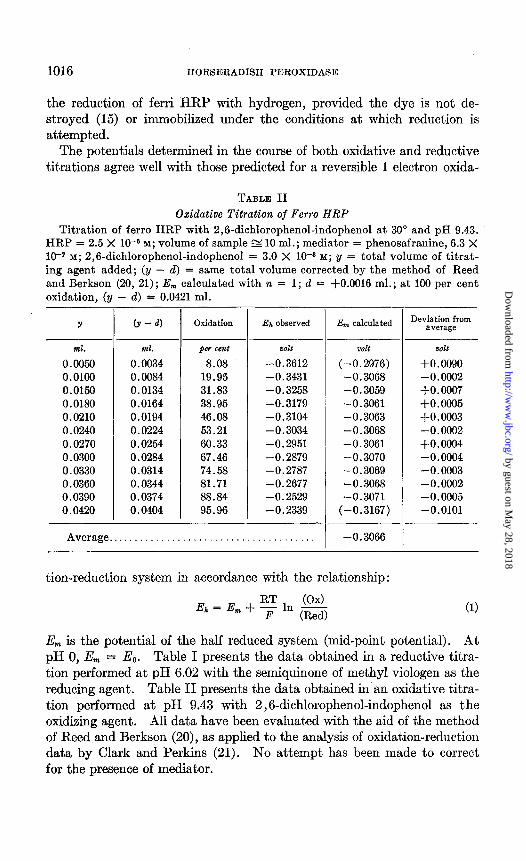
TABLE II Oxidative Titration of Ferro HRP
Titration of ferro HRP with 2,6-dichlorophenol-indophenol at 30” and pH 9.43. HRP = 2.5 X 1O-6 M; volume of sample g 10 ml.; mediator = phenosafranine, 6.3 X 10-r M; 2,6-dichlorophenol-indophenol = 3.0 X 10-S M; y = total volume of titrat- ing agent added; (y - d) = same total volume corrected by the method of Reed and Berkson (20, 21); E,,, calculated with n = 1; d = +O.OOlS ml.; at 100 per cent oxidation, (y - d) = 0.0421 ml.
Y (Y - 4 Oxidation Ej, observed
ml. ml. per cent oolt 0.0050 0.0034 8.08 -0.3612 0.0100 0.0084 19.95 -0.3431 0.0150 0.0134 31.83 -0.3258 0.0180 0.0164 38.95 -0.3179 0.0210 0.0194 46.08 -0.3104 0.0240 0.0224 53.21 -0.3034 0.0270 0.0254 60.33 -0.2951 0.0300 0.0284 67.46 -0.2879 0.0330 0.0314 74.58 -0.2787 0.0360 0.0344 81.71 -0.2677 0.0390 0.0374 88.84 -0.2529 0.0420 0.0404 95.96 -0.2339
-
Average.......................................
E, calculated
piozt
(-0.2976) -0.3068 -0.3059 -0.3061 -0.3063 -0.3068 -0.3061 -0.3070 -0.3069 -0.3068 -0.3071
(-0.3167)
-0.3066
tion-reduction system in accordance with the relationship :
RT (OX) Eh = Em -I- 7 In (led)
Deviation from average
oolt
+0.0090 -0.0002 +0.0007 +0.0005 +0.0003 -0.0002 +0.0004 -0.0004 -0.0003 -0.0002 -0.0005 -0.0101
(1)
Em is the potential of the half reduced system (mid-point potential). At pH 0, Em = E,. Table I presents the data obtained in a reductive titra- tion performed at pH 6.02 with the semiquinone of methyl viologen as the reducing agent. Table II presents the data obtained in an oxidative titra- tion performed at pH 9.43 with 2,6-dichlorophenol-indophenol as the oxidizing agent. All data have been evaluated with the aid of the method of Reed and Berkson (20), as applied to the analysis of oxidation-reduction data by Clark and Perkins (21). No attempt has been made to correct for the presence of mediator.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
H. A. HARBURY 1017
Potentials have been determined over the range of pH 4.0 to 11.3. Beyond the limits of this range, HRP has been found to undergo irreversible change under the conditions of the present studies. The results of the two experiments conducted at the extremes of the range here reported must be considered to represent approximations only. Table III summarizes the results of the present series of measurements.
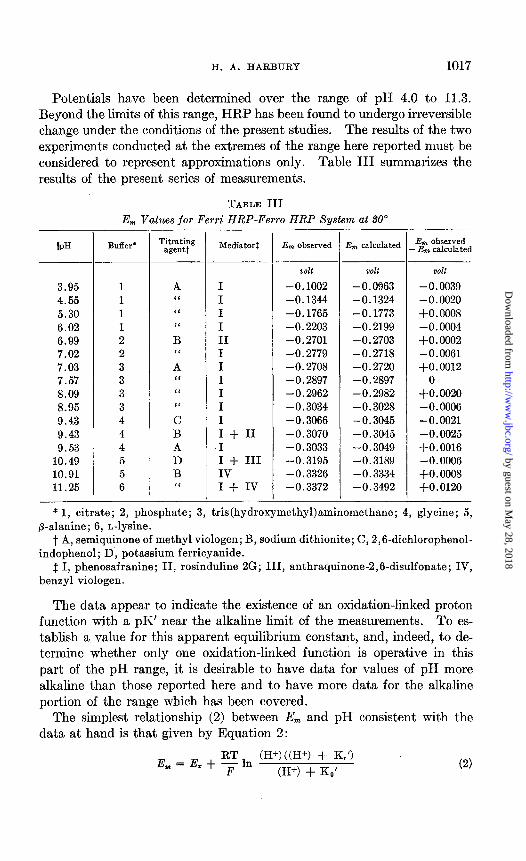
TABLE III E,,, Values for Ferri HRP-Ferro HRP System at SO0
IPH B&X*
3.95 1 4.55 1 5.30 1 6.02 1 6.99 2 7.02 2 7.03 3 7.57 3 8.09 3 8.95 3 9.43 4 9.43 4 9.53 4
10.49 5 10.91 5 11.25 6
Titrating agentt
A ‘I “ I‘ B “ A ‘I I‘ “ C B A D B “
Mediatorf
I I I I II I I I I I I I + II I I + III IV I + IV
-
Em observed Em calculated Em observed -Em calculated
oolt volt volt
-0.1002 -0.0963 -0.0039 -0.1344 -0.1324 -0.0020 -0.1765 -0.1773 +0.0008 -0.2203 -0.2199 -0.0004 -0.2701 -0.2703 +0.0002 -0.2779 -0.2718 -0.0061 -0.2708 -0.2720 +0.0012 -0.2897 -0.2897 0 -0.2962 -0.2982 +0.0020 -0.3034 -0.3028 -0.0006 -0.3066 -0.3045 -0.0021 -0.3070 -0.3045 -0.0025 -0.3033 -0.3049 $0.0016 -0.3195 -0.3189 -0.0006 -0.3326 -0.3334 +O.OOOS -0.3372 -0.3492 +0.0120
- * 1, citrate; 2, phosphate; 3, tris(hydroxymethyl)aminomethane; 4, glycine; 5,
p-&nine; 6, n-lysine. t A, semiquinone of methyl viologen; B, sodium dithionite; C, 2,6-dichlorophenol-
indophenol; D, potassium ferricyanide. $ I, phenosafranine; II, rosinduline 2G; III, anthraquinone-2,6-disulfonate; IV,
benzyl viologen.
The data appear to indicate the existence of an oxidation-linked proton function with a pK’ near the alkaline limit of the measurements. To es- tablish a value for this apparent equilibrium constant, and, indeed, to de- termine whether only one oxidation-linked function is operative in this part of the pH range, it is desirable to have data for values of pH more alkaline than those reported here and to have more data for the alkaline portion of the range which has been covered.
The simplest relationship (2) between E,,, and pH consistent with the data at hand is that given by Equation 2:
E,,, = E, + F In (H+) (@I+) + K,‘) (II+) + KS’
(2)
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
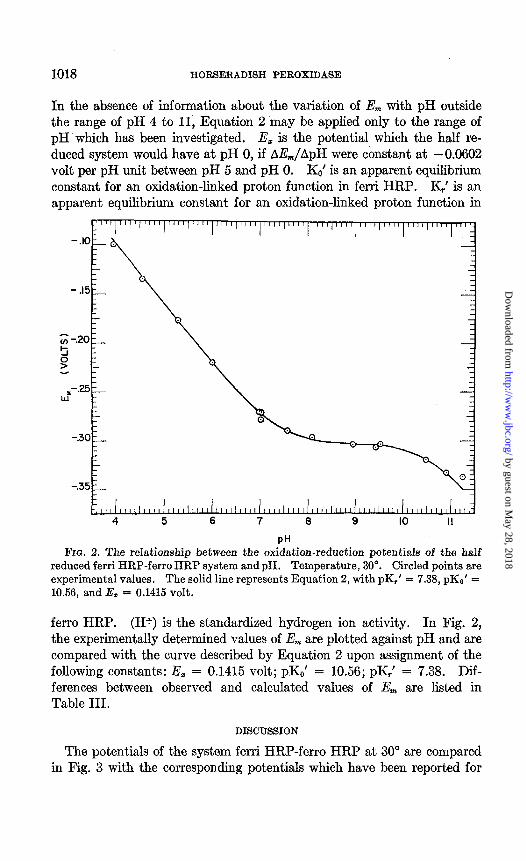
1018 HORSERADISH PEROXIDASE
In the absence of information about the variation of E,,, with pH outside the range of pH 4 to 11, Equation 2 may be applied only to the range of pH which has been investigated. E, is the potential which the half re- duced system would have at pH 0, if AE,,JApH were constant at -0.0602 volt per pH unit between pH 5 and pH 0. KO’ is an apparent equilibrium constant for an oxidation-linked proton function in ferri HRP. K,’ is an apparent equilibrium constant for an oxidation-linked proton function in
4 5 6 7 8 9 IO II
PH FICA 2. The relationship between the oxidation-reduction potentials of the half
reduced ferri HRP-ferro HRP system andpH. Temperature, 30”. Circled points are experimental values. The solid line represents Equation 2, with pK,’ = 7.38, pKo’ = 10.56, and I& = 0.1415 volt.
ferro HRP. (H+) is the standardized hydrogen ion activity. In Fig. 2, the experimentally determined values of E,,, are plotted against pH and are compared with the curve described by Equation 2 upon assignment of the following constants: E, = 0.1415 volt; pK,’ = 10.56; pK,’ = 7.38. Dif- ferences between observed and calculated values of Em are listed in Table III.
DISCUSSION
The potentials of the system ferri HRP-ferro HRP at 30” are compared in Fig. 3 with the corresponding potentials which have been reported for
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
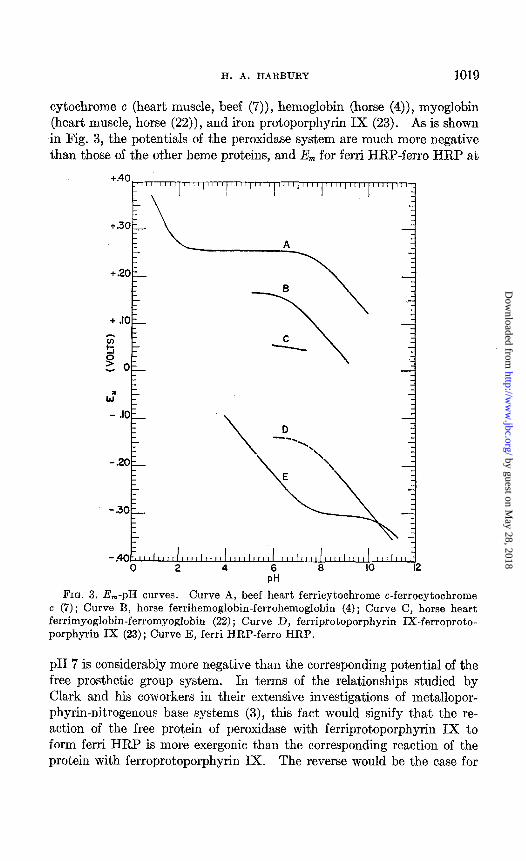
H. A. HARBURY 1019
cytochrome c (heart muscle, beef (Y’)), hemoglobin (horse (4)), myoglobin (heart muscle, horse (22)), and iron protoporphyrin IX (23). As is shown in Fig. 3, the potentials of the peroxidase system are much more negative than those of the other heme proteins, and E,,, for ferri HRP-ferro HRP at
+.40
t \ t
e.30;
+201
+ .lOL.
5; - 5 - E OL
b.7 - - .10-L
-.201
-30;
A
B 5 -c
PH FIG. 3. E,-pH curves. Curve A, beef heart ferricytochrome c-ferrocytochrome
c (7); Curve B, horse ferrihemoglobin-ferrohemoglobin (4); Curve C, horse heart ferrimyoglobin-ferromyoglobin (22) ; Curve D, ferriprotoporphyrin IX-ferroproto- porphyrin IX (23); Curve E, ferri HRP-ferro HRP.
pH 7 is considerably more negative than the corresponding potential of the free prosthetic group system. In terms of the relationships studied by Clark and his coworkers in their extensive investigations of metallopor- phyrin-nitrogenous base systems (3), this fact would signify that the re- action of the free protein of peroxidase with ferriprotoporphyrin IX to form ferri HRP is more exergonic than the corresponding reaction of the protein with ferroprotoporphyrin IX. The reverse would be the case for
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1020 HORSERADISH PEROXIDASE
the analogous (hypothetical) reactions in the myoglobin, hemoglobin, and cytochrome c systems.
The very negative potentials of the ferri HRP-ferro HRP system appear not unreasonable in relation to other available information. The reduction of ferri HRP has been accomplished only through the application of rela- tively strong reducing agents, and ferro HRP in vitro is oxidized by air at an exceedingly high rate. The existence of ferro HRP in vivo has yet to be observed. Freshly prepared enzyme has always been found to be in the “ferric” state. It is well established that peroxidatic activity does not in- volve formation of ferro HRP, but rather the formation of a complex or compound containing 2 more oxidizing equivalents than does ferri HRP (24, 25). Although at one time it was suggested that cyclic reduction to the “ferrous” state occurs in the action of peroxidase as an oxidase in the presence of dihydroxymaleic acid (26)) it is now known that here also there is no formation of ferro HRP (27).
Keilin and Hartree (28) have recently made a study of the cyanide com- pound of ferro HRP. They fmd that between pH 5.5 and 6.0 ferri HRP- cyanide cannot be reduced by sodium dithionite to form ferro HRP-cya- nide, although such reduction can be accomplished between pH 8.5 and 9.5. Moreover, upon acidification of a solution of ferro HRP-cyanide pre- pared at pH 9, there occurs, below pH 8.5, a progressive oxidation to ferri HRP-cyanide. At pH 6, no ferro HRP-cyanide remains. At pH 9 (tem- perature approximately 20” (?)), the dissociation constant of ferro HRP- cyanide was found to be approximately lo3 times larger than that of ferri HRP-cyanide. On the basis of these observations and our preliminary data on the potentials of the ferri HRP-ferro HRP system near pH 7 (9)) Keilin and Hartree conclude that cyanide must lower markedly the already negative potentials of the cyanide-free system, and that, as a result, at pH 5 to 6 sodium dithionite can no longer effect the reduction of the “ferric” cyanide compound.
A lOOO-fold difference in the magnitudes of the dissociation constants of the cyanide compounds, the “ferrous” compound being the more highly dissociable, would be expected to lower the oxidation-reduction potential of a 1 electron system by about 0.18 volt. If we assume the ratio of the dissociation constants to be the same at pH 6 as that at pH 9, and assume the ratio to be lo3 at 30”) then at pH 9 and 30” the potential of the ferri HRP-cyanide-ferro HRP-cyanide system would be approximately - 0.48 volt, whereas at pH 6 it would be approximately -0.40 volt. The poten- tial at pH 9 would thus be about 0.06 volt more positive, and that at pH 6 about 0.04 volt more negative, than the corresponding potentials of the hydrogen system. It is interesting, furthermore, that according to this very approximate calculation the potentials of the HRP-cyanide and
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

H. A. HARBURY 1021
hydrogen systems should be about equal at pH 8, the potentials of the HRP-cyanide system becoming more negative than those of the hydrogen system upon further acidification. Keilin and Hartree find ferro HRP-cy- anide to be stable in the range pH 8.5 to 9.5, oxidation setting in upon acidification below pH 8.5.
The oxidation-reduction potentials for the different heme proteins at pH 7 range from very positive to very negative values. Beef heart ferricyto- chrome c-ferrocytochrome c is among the most positive biological oxidation- reduction systems known, and ferri HRP-ferro HRP is among the most negative. It might be (9) that the resistance to reduction displayed by native catalase results from very negative oxidation-reduction potentials for the ferricatalase-ferrocatalase system, potentials so negative as to pre- clude the existence of free ferrocatalase under the conditions at which re- duction of ferricatalase has been attempted.
From the data on the variation of the peroxidase potentials with pH (Fig. 2), it will be seen that Em changes little with pH in the range pH 8.5 to 9.5, becomes less negative as the pH decreases below pH 8.5, and be- comes more negative as the pH increases above pH 9.5. The simplest interpretation (2) of the variation in AE,/ApH indicated by these data is to ascribe the change in slope below pH 8.5 to an oxidation-linked proton equilibrium in ferro HRP and the change above pH 9.5 to an oxidation- linked proton equilibrium in ferri HRP.
It is generally accepted that in ferri HRP in neutral solution the sixth coordination position about the iron of the prosthetic group is occupied by a molecule of water (29, 30). The spectrum of ferri HRP in alkaline solu- tion differs from that in neutral solution (31). This difference has been ascribed to the dissociation of the water molecule coordinated to the iron, with the resultant formation of “alkaline peroxidase,” a form of ferri HRP in which the sixth coordination position is thought to be occupied by a hydroxyl ion (29,32). Theorell(32) has studied this reaction both spectro- photometrically and magnetometrically and has reported a value of 11.27 for pK’ as measured magnetometrically at 20” and a value of 10.91 as de- termined spectrophotometrically. The two determinations were made at different ionic strengths. It seems likely that the provisional value 10.56 reported in the present communication represents the same equilibrium. The higher temperature at which the present experiments were conducted would be expected to result in a smaller value for pK’. Lower ionic strengths would result in a similar effect.
The observation of the existence of an oxidation-linked proton equili- brium with a pK’ of 7.38 at 30” is worthy of special comment. Like the equilibrium in ferrihemoglobin with pK’ 6.65 at 30” (4), and the equilib- rium in ferricytochrome c with pK’ 7.8 at 30” as reported by Rodkey and
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1022 HORSERADISH PEROXIDASE
Ball (7) and pK’ 6.86 at 20” as reported by Paul (8), this oxidation-linked function has escaped detection by spectrophotometric and magnetometric means. Differential titration data for the pertinent range of pH have not been fully reported, In the only study which has been published (19), technical difficulties were encountered below pH 8, but it was concluded that the titration difference between ferri and ferro HRP remains approxi- mately constant over the range of pH 4 to 8. In a later review (33), however, Theorell, referring to unpublished data, states that the titration difference decreases below pH 8 and mentions the possibility of the existence of an oxidation-linked proton equilibrium in ferro HRP with a pK’ near 7. This later conclusion is thus in qualitative agreement with the results of the present investigation.
On the basis of the information now available, no hypothesis will be ad- vanced as to the nature of the proton equilibrium with pK’ 7.38 in ferro HRP. It may be noted, however, that the present data need not be con- sidered to contradict the hypothesis that the fifth and sixth coordination positions about the iron of the prosthetic group are occupied by a carboxyl group of the protein (19) and a water molecule (29, 30). A heme-linked proton equilibrium in a protein system need not involve the dissociation of a group occupying a coordination position about the iron of the prosthetic group*
There can be no doubt of the importance of learning the identity of the groups in the protein which display first order effects in the interaction with the prosthetic group, and the great interest which has been evinced in the identity of what have usually been presumed to be the foremost of these groups, those occupying the available coordination positions about the iron of the prosthetic group, is readily understandable. It is apparent, however, that the reactivity and functional specificity displayed by a heme protein may be expected not only to depend upon the nature of the groups occupy- ing the coordination positions about the heme iron, but to reflect the interaction of the prosthetic group with the larger number of groups in- cluded within the original meaning (5) of the term “heme-linked groups.” This interaction is likely to be highly dependent on factors of solvation and configuration, factors which may differ greatly not only between one heme protein and another, but also between reversibly interconvertible states and compounds of the same heme protein. Insight into the very sensitive re- lationship between function and structure which is displayed by the heme proteins requires further information not only about the interaction between the prosthetic group and protein in each of the heme proteins, but also, most importantly, about the changes in this interaction which accompany various linked reactions. The immediate need is for the compilation of a body of quantitative information on pertinent entropy, enthalpy, and free energy changes.
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

H. A. HARBURY 1023
SUMMARY
1. Oxidation-reduction potentials of the ferri horseradish peroxidase (ferri HRP)-ferro horseradish peroxidase (ferro HRP) system at 30” have been determined potentiometrically over the range of pH 4 to 11.
2. Throughout the range of pH investigated, the potentials of the ferri HRP-ferro HRP system are much more negative than the corresponding potentials which have been reported for beef cytochrome c, horse hemo- globin, and horse myoglobin. Em at pH 7 and 30” is -0.271 volt. This is more negative than the corresponding potential of half reduced iron pro- toporphyrin IX, which is estimated to be -0.14 volt.
3. The simplest interpretation of the observed variation of Em with pH is that there exists an oxidation-linked proton equilibrium in ferro HRP with pK,’ at 30” = 7.38, and an oxidation-linked proton equilibrium in ferri HRP with pKO’ at 30” = 10.56. The latter value is a provisional one. In accordance with this interpretation, the relationship between Em and pH over the range of pH 4 to 11 is expressed by the equation
Et,, = Ez + y In (H+) ((H+) + K,‘) (XI+) + Ko’
4. A titration assembly is described which has proved suitable for the study of small samples of dilute, highly autoxidieable systems. Its use permits the concurrent collection of potentiometric and spectrophotometric data throughout the course of a titration.
I should like to express my gratitude for the encouragement, interest, and support of Professor W. Mansfield Clark and Professor Joseph S. Fruton. I am deeply indebted also to Dr. Karl-Gustav Paul and Professor Hugo Theorell for their interest in this work, and for the most generous contri- bution of enzyme which has speeded it.
BIBLIOGRAPHY
1. Wyman, J., Jr., Advances in Protein Chem., 4,407 (1948). 2. Clark, W. M., and Cohen, B., Pub. Health Rep., U. S. P. H. S., 38, 666 (1923). 3. Clark, W. M., Taylor, J. F., Davies, T. H., and Vestling, C. S., J. Biol. Chem.,
136, 643 (1940). 4. Taylor, J. F., and Hastings, A. B., J. Biol. Chem., 131, 649 (1939). 5. Coryell, C. D., and Pauling, L., J. Biol. Chem., 132,769 (1940). 6. Wyman, J., Jr., and Ingalls, E. N., J. Biol. Chem., 139,877 (1941). 7. Rodkey, F. L., and Ball, E. G., J. Biol. Chem., 182, 17 (1950). 8. Paul, K.-G., Arch. Biochem., 12,441 (1947). 9. Harbury, H. A., J. Am. Chem. Sot., 76,4625 (1953).
10. Theorell, H., Ark. Kemi, Mineral. o. Geol., 16 A, No. 2 (1942). 11. Clark, W. M., J. Biol. Chem., 23,475 (1915). 12. MacInnes, D. A., and Belcher, D., Ind. and Eng. Chem., Anal. Ed., 6, 199 (1933). 13. Willstatter, R., and Stoll, A., Ann. Chem., 416, 21 (1918).
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1024 HORSERADISH PEROXIDASE
14. Hitchcock, D. I., and Taylor, A. C., J. Am. Chem. SCJC., 60,271O (1938). 15. Stiehler, R. D., Chen, T.-T., and Clark, W. M., J. Am. Chem. Sot., 66,891 (1933). 16. Michaelis, L., J. BioZ. Chem., 91,369 (1931). 17. Conant, J. B., Kahn, H. M., Fieser, L. F., and Kurtz, S. S., J. Am. Chem. Sac.,
44, 1382 (1922). 18. Michaelis, L., and Hill, E. S., J. Gen. Physiol., 16,859 (1933). 19. Theorell, H., Ark. Kemi, Mineral. o. Geol., 16 A, No. 14 (1943). 20. Reed, L. J., and Berkson, J., J. Phys. Chem., 33, 760 (1929). 21. Clark, W. M., and Perkins, M. E., J. Am. Chem. Sot., 64, 1228 (1932). 22. Taylor, J. F., and Morgan, V. E., J. BioZ. Chem., 144, 15 (1942). 23. Shack, J., and Clark, W. M., J. BioZ. Chem., 171,143 (1947). 24. George, P., Advances in Catalysis, 4, 367 (1952). 25. Chance, B., Arch. Biochem. and Biophys., 37,235 (1952). 26. Swedin, B., and Theorell, H., Nature, 146,71 (1940). 27. Chance, B., J. BioZ. Chem., 197, 577 (1952). 28. Keilin, D., and Hartree, E. F., Biochem. J., 61, 153 (1955). 29. Keilin, D., and Hartree, E. F., Biochem. J., 49, 88 (1951). 30. Chance, B., Arch. Biochem. and Biophys., 40, 153 (1952). 31. Keilin, D., and Mann, T., Proc. Roy. Sot. London, Series B, 122, 119 (1937). 32. Theorell, H., Ark. Kemi, Mineral. o. Geol., 16 A, No. 3 (1942). 33. Theorell, H., Advances in Enzymol., 7, 265 (1947).
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Henry A. HarburyPEROXIDASE
POTENTIALS OF HORSERADISH OXIDATION-REDUCTION
1957, 225:1009-1024.J. Biol. Chem.
http://www.jbc.org/content/225/2/1009.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
html#ref-list-1
http://www.jbc.org/content/225/2/1009.citation.full.accessed free atThis article cites 0 references, 0 of which can be
by guest on May 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from