blood characteristics for high altitude adaptation in tibetan chickens
TRANSCRIPT

Blood Characteristics for High Altitude Adaptation in Tibetan Chickens1
H. Zhang,* C. X. Wu,*2 Y. Chamba,† and Y. Ling*
*College of Animal Science and Technology, China Agricultural University, Bejing, China 100094;and †College of Agriculture and Animal Husbandry, Tibet University, Linzhi, Tibet, China 860000
ABSTRACT Tibetan chickens, a unique chicken breednative to high altitude, have good adaptation to hypoxia.The experiment was conducted to determine the adaptiveblood characteristics in Tibetan chickens. Fertile eggsfrom Tibetan and Dwarf Recessive White chickens wereincubated, and the chicks were reared until 10 wk of ageat low altitude (100 m) and high altitude (2,900 m). At 1d and 2, 6, and 10 wk of age, the hematological characteris-tics, blood gas value, and blood volume were measured.Tibetan chickens had more red blood cells (RBC), smallermean cell volume, lower pH and partial pressure of oxy-gen, and higher partial pressure of carbon dioxide at high
Key words: Tibetan chicken, high altitude adaptation, hematological characteristic, blood gas, blood volume
2007 Poultry Science 86:1384–1389
INTRODUCTION
The Tibetan chicken (Gallus gallus), a montane chickenbreed, has a very wide distribution at altitudes of 2,200to 4,100 m in the Qinghai-Tibet Plateau. Although thebirds are exposed to hypobaric hypoxia at these alti-tudes, this breed, with a history of domestication morethan 1,000 yr at high altitude, breeds successfully andhas good resistance to chronic mountain sickness (Zhanget al., 2005, 2006). Tibetan chickens have good adapta-tions to high altitude and therefore are good subjectsfor investigating the mechanisms of adaptation.
Monge and Leon-Velarde (1991) pointed out, in anexcellent review about physiological adaptation to highaltitude in mammals and birds, that a high O2-hemoglo-bin (Hb) affinity, a moderate or absent polycythemicresponse, a low venous partial pressure of oxygen (pO2),a thin-walled pulmonary vascular tree, and the absenceof chronic mountain sickness are characteristics of a ge-notypically adapted high-altitude mammal or bird, andthese conditions are maintained at sea level and aretransmitted to the descendants. Bar-headed geese (Anserindicus), a species that breeds on the Tibetan Plateau and
©2007 Poultry Science Association Inc.Received January 17, 2007.Accepted March 21, 2007.1The research was supported by the 973 Project (2006CB102101) of
China and the Project of National Fundamental Platform for ScientificWork (2005DKA21100-02).
2Corresponding author: [email protected]
1384
altitude and had lower blood volume, erythrocyte vol-ume, and plasma volume at low and high altitude thanDwarf Recessive White chickens. Tibetan chickens rearedat high altitude retained a high level of RBC and a stablelevel of hematocrit from younger to older, but DwarfRecessive White chickens reared at high altitude pre-sented an increase in RBC and hematocrit values. It wasconcluded the adaptation was achieved in Tibetan chick-ens by increase in RBC and blood oxygen affinity, de-crease in mean cell volume, and reducing susceptivityto hypocapnia.
migrates from near sea-level conditions to elevations ashigh as 9,200 m, had a high oxygen affinity and invari-able values of red blood cells (RBC), hematocrit (Hct),Hb, and mean cell volume (MCV) in conditions of sealevel or high altitude (Black and Tenney, 1980). Ye et al.(1994) reported that highland native animals had higherRBC numbers and smaller MCV, which are advanta-geous for oxygen transport. Blood gases are sensitive tothe hypoxic environment, with reductions in the arterialpO2 matching the reduction in atmospheric pO2 in Pe-king ducks and Bar-headed geese (Faraci et al., 1985).Increases in blood volume (BV) also are important inincreasing hypoxic tolerance (Birchard and Tenney,1990), but potential changes in BV in chickens at differentaltitudes have not been reported previously.
Several facets of high altitude adaptation have beenevaluated in lowland chickens (Leon-Velarde andMonge, 2004), but there are few evaluations of chickensthat are native to high altitude. The question addressedin the present study concerns how hematological charac-teristics, blood gases, and BV compare between highaltitude native chickens and lowland chickens, both atlow and high altitudes, and how the Tibetan chickensadapt to high altitude hypoxia in blood characteristics.
MATERIALS AND METHODS
Tibetan chickens (Gallus gallus) from Linzhi, Tibet(2,900 m altitude) were transported to Beijing (100 m
at National C
hung Hsing U
niversity Library on A
pril 10, 2014http://ps.oxfordjournals.org/
Dow
nloaded from
BLOOD CHARACTERISTICS OF HIGH-ALTITUDE NATIVE CHICKEN 1385
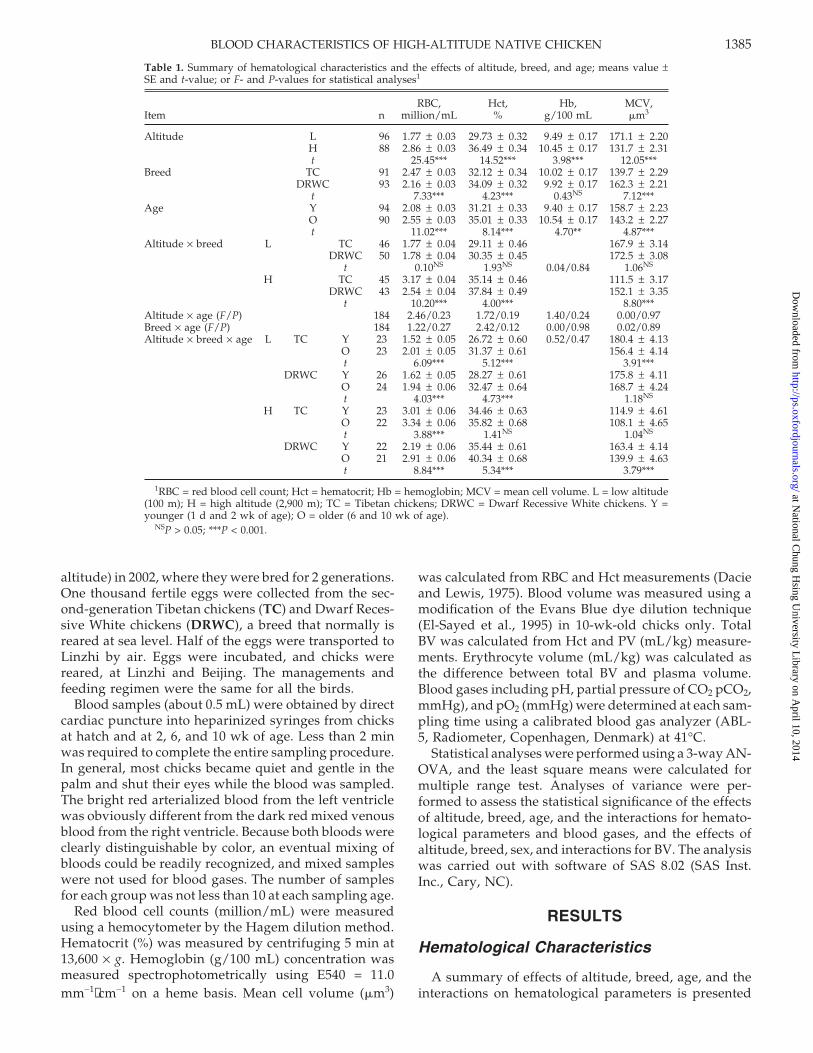
Table 1. Summary of hematological characteristics and the effects of altitude, breed, and age; means value ±SE and t-value; or F- and P-values for statistical analyses1
RBC, Hct, Hb, MCV,Item n million/mL % g/100 mL �m3
Altitude L 96 1.77 ± 0.03 29.73 ± 0.32 9.49 ± 0.17 171.1 ± 2.20H 88 2.86 ± 0.03 36.49 ± 0.34 10.45 ± 0.17 131.7 ± 2.31t 25.45*** 14.52*** 3.98*** 12.05***
Breed TC 91 2.47 ± 0.03 32.12 ± 0.34 10.02 ± 0.17 139.7 ± 2.29DRWC 93 2.16 ± 0.03 34.09 ± 0.32 9.92 ± 0.17 162.3 ± 2.21
t 7.33*** 4.23*** 0.43NS 7.12***Age Y 94 2.08 ± 0.03 31.21 ± 0.33 9.40 ± 0.17 158.7 ± 2.23
O 90 2.55 ± 0.03 35.01 ± 0.33 10.54 ± 0.17 143.2 ± 2.27t 11.02*** 8.14*** 4.70** 4.87***
Altitude × breed L TC 46 1.77 ± 0.04 29.11 ± 0.46 167.9 ± 3.14DRWC 50 1.78 ± 0.04 30.35 ± 0.45 172.5 ± 3.08
t 0.10NS 1.93NS 0.04/0.84 1.06NS
H TC 45 3.17 ± 0.04 35.14 ± 0.46 111.5 ± 3.17DRWC 43 2.54 ± 0.04 37.84 ± 0.49 152.1 ± 3.35
t 10.20*** 4.00*** 8.80***Altitude × age (F/P) 184 2.46/0.23 1.72/0.19 1.40/0.24 0.00/0.97Breed × age (F/P) 184 1.22/0.27 2.42/0.12 0.00/0.98 0.02/0.89Altitude × breed × age L TC Y 23 1.52 ± 0.05 26.72 ± 0.60 0.52/0.47 180.4 ± 4.13
O 23 2.01 ± 0.05 31.37 ± 0.61 156.4 ± 4.14t 6.09*** 5.12*** 3.91***
DRWC Y 26 1.62 ± 0.05 28.27 ± 0.61 175.8 ± 4.11O 24 1.94 ± 0.06 32.47 ± 0.64 168.7 ± 4.24t 4.03*** 4.73*** 1.18NS
H TC Y 23 3.01 ± 0.06 34.46 ± 0.63 114.9 ± 4.61O 22 3.34 ± 0.06 35.82 ± 0.68 108.1 ± 4.65t 3.88*** 1.41NS 1.04NS
DRWC Y 22 2.19 ± 0.06 35.44 ± 0.61 163.4 ± 4.14O 21 2.91 ± 0.06 40.34 ± 0.68 139.9 ± 4.63t 8.84*** 5.34*** 3.79***
1RBC = red blood cell count; Hct = hematocrit; Hb = hemoglobin; MCV = mean cell volume. L = low altitude(100 m); H = high altitude (2,900 m); TC = Tibetan chickens; DRWC = Dwarf Recessive White chickens. Y =younger (1 d and 2 wk of age); O = older (6 and 10 wk of age).
NSP > 0.05; ***P < 0.001.
altitude) in 2002, where they were bred for 2 generations.One thousand fertile eggs were collected from the sec-ond-generation Tibetan chickens (TC) and Dwarf Reces-sive White chickens (DRWC), a breed that normally isreared at sea level. Half of the eggs were transported toLinzhi by air. Eggs were incubated, and chicks werereared, at Linzhi and Beijing. The managements andfeeding regimen were the same for all the birds.
Blood samples (about 0.5 mL) were obtained by directcardiac puncture into heparinized syringes from chicksat hatch and at 2, 6, and 10 wk of age. Less than 2 minwas required to complete the entire sampling procedure.In general, most chicks became quiet and gentle in thepalm and shut their eyes while the blood was sampled.The bright red arterialized blood from the left ventriclewas obviously different from the dark red mixed venousblood from the right ventricle. Because both bloods wereclearly distinguishable by color, an eventual mixing ofbloods could be readily recognized, and mixed sampleswere not used for blood gases. The number of samplesfor each group was not less than 10 at each sampling age.
Red blood cell counts (million/mL) were measuredusing a hemocytometer by the Hagem dilution method.Hematocrit (%) was measured by centrifuging 5 min at13,600 × g. Hemoglobin (g/100 mL) concentration wasmeasured spectrophotometrically using E540 = 11.0mm−1�cm−1 on a heme basis. Mean cell volume (�m3)
was calculated from RBC and Hct measurements (Dacieand Lewis, 1975). Blood volume was measured using amodification of the Evans Blue dye dilution technique(El-Sayed et al., 1995) in 10-wk-old chicks only. TotalBV was calculated from Hct and PV (mL/kg) measure-ments. Erythrocyte volume (mL/kg) was calculated asthe difference between total BV and plasma volume.Blood gases including pH, partial pressure of CO2 pCO2,mmHg), and pO2 (mmHg) were determined at each sam-pling time using a calibrated blood gas analyzer (ABL-5, Radiometer, Copenhagen, Denmark) at 41°C.
Statistical analyses were performed using a 3-way AN-OVA, and the least square means were calculated formultiple range test. Analyses of variance were per-formed to assess the statistical significance of the effectsof altitude, breed, age, and the interactions for hemato-logical parameters and blood gases, and the effects ofaltitude, breed, sex, and interactions for BV. The analysiswas carried out with software of SAS 8.02 (SAS Inst.Inc., Cary, NC).
RESULTS
Hematological Characteristics
A summary of effects of altitude, breed, age, and theinteractions on hematological parameters is presented
at National C
hung Hsing U
niversity Library on A
pril 10, 2014http://ps.oxfordjournals.org/
Dow
nloaded from
ZHANG ET AL.1386
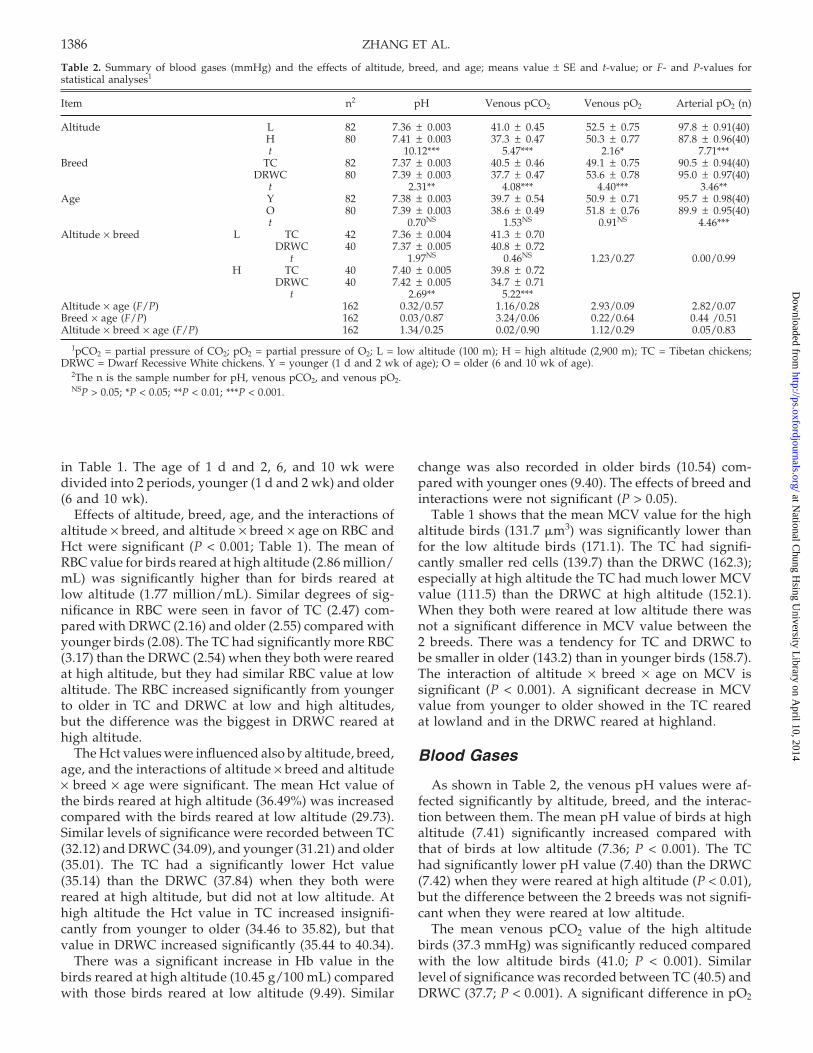
Table 2. Summary of blood gases (mmHg) and the effects of altitude, breed, and age; means value ± SE and t-value; or F- and P-values forstatistical analyses1
Item n2 pH Venous pCO2 Venous pO2 Arterial pO2 (n)
Altitude L 82 7.36 ± 0.003 41.0 ± 0.45 52.5 ± 0.75 97.8 ± 0.91(40)H 80 7.41 ± 0.003 37.3 ± 0.47 50.3 ± 0.77 87.8 ± 0.96(40)t 10.12*** 5.47*** 2.16* 7.71***
Breed TC 82 7.37 ± 0.003 40.5 ± 0.46 49.1 ± 0.75 90.5 ± 0.94(40)DRWC 80 7.39 ± 0.003 37.7 ± 0.47 53.6 ± 0.78 95.0 ± 0.97(40)
t 2.31** 4.08*** 4.40*** 3.46**Age Y 82 7.38 ± 0.003 39.7 ± 0.54 50.9 ± 0.71 95.7 ± 0.98(40)
O 80 7.39 ± 0.003 38.6 ± 0.49 51.8 ± 0.76 89.9 ± 0.95(40)t 0.70NS 1.53NS 0.91NS 4.46***
Altitude × breed L TC 42 7.36 ± 0.004 41.3 ± 0.70DRWC 40 7.37 ± 0.005 40.8 ± 0.72
t 1.97NS 0.46NS 1.23/0.27 0.00/0.99H TC 40 7.40 ± 0.005 39.8 ± 0.72
DRWC 40 7.42 ± 0.005 34.7 ± 0.71t 2.69** 5.22***
Altitude × age (F/P) 162 0.32/0.57 1.16/0.28 2.93/0.09 2.82/0.07Breed × age (F/P) 162 0.03/0.87 3.24/0.06 0.22/0.64 0.44 /0.51Altitude × breed × age (F/P) 162 1.34/0.25 0.02/0.90 1.12/0.29 0.05/0.83
1pCO2 = partial pressure of CO2; pO2 = partial pressure of O2; L = low altitude (100 m); H = high altitude (2,900 m); TC = Tibetan chickens;DRWC = Dwarf Recessive White chickens. Y = younger (1 d and 2 wk of age); O = older (6 and 10 wk of age).
2The n is the sample number for pH, venous pCO2, and venous pO2.NSP > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001.
in Table 1. The age of 1 d and 2, 6, and 10 wk weredivided into 2 periods, younger (1 d and 2 wk) and older(6 and 10 wk).
Effects of altitude, breed, age, and the interactions ofaltitude × breed, and altitude × breed × age on RBC andHct were significant (P < 0.001; Table 1). The mean ofRBC value for birds reared at high altitude (2.86 million/mL) was significantly higher than for birds reared atlow altitude (1.77 million/mL). Similar degrees of sig-nificance in RBC were seen in favor of TC (2.47) com-pared with DRWC (2.16) and older (2.55) compared withyounger birds (2.08). The TC had significantly more RBC(3.17) than the DRWC (2.54) when they both were rearedat high altitude, but they had similar RBC value at lowaltitude. The RBC increased significantly from youngerto older in TC and DRWC at low and high altitudes,but the difference was the biggest in DRWC reared athigh altitude.
The Hct values were influenced also by altitude, breed,age, and the interactions of altitude × breed and altitude× breed × age were significant. The mean Hct value ofthe birds reared at high altitude (36.49%) was increasedcompared with the birds reared at low altitude (29.73).Similar levels of significance were recorded between TC(32.12) and DRWC (34.09), and younger (31.21) and older(35.01). The TC had a significantly lower Hct value(35.14) than the DRWC (37.84) when they both werereared at high altitude, but did not at low altitude. Athigh altitude the Hct value in TC increased insignifi-cantly from younger to older (34.46 to 35.82), but thatvalue in DRWC increased significantly (35.44 to 40.34).
There was a significant increase in Hb value in thebirds reared at high altitude (10.45 g/100 mL) comparedwith those birds reared at low altitude (9.49). Similar
change was also recorded in older birds (10.54) com-pared with younger ones (9.40). The effects of breed andinteractions were not significant (P > 0.05).
Table 1 shows that the mean MCV value for the highaltitude birds (131.7 �m3) was significantly lower thanfor the low altitude birds (171.1). The TC had signifi-cantly smaller red cells (139.7) than the DRWC (162.3);especially at high altitude the TC had much lower MCVvalue (111.5) than the DRWC at high altitude (152.1).When they both were reared at low altitude there wasnot a significant difference in MCV value between the2 breeds. There was a tendency for TC and DRWC tobe smaller in older (143.2) than in younger birds (158.7).The interaction of altitude × breed × age on MCV issignificant (P < 0.001). A significant decrease in MCVvalue from younger to older showed in the TC rearedat lowland and in the DRWC reared at highland.
Blood Gases
As shown in Table 2, the venous pH values were af-fected significantly by altitude, breed, and the interac-tion between them. The mean pH value of birds at highaltitude (7.41) significantly increased compared withthat of birds at low altitude (7.36; P < 0.001). The TChad significantly lower pH value (7.40) than the DRWC(7.42) when they were reared at high altitude (P < 0.01),but the difference between the 2 breeds was not signifi-cant when they were reared at low altitude.
The mean venous pCO2 value of the high altitudebirds (37.3 mmHg) was significantly reduced comparedwith the low altitude birds (41.0; P < 0.001). Similarlevel of significance was recorded between TC (40.5) andDRWC (37.7; P < 0.001). A significant difference in pO2
at National C
hung Hsing U
niversity Library on A
pril 10, 2014http://ps.oxfordjournals.org/
Dow
nloaded from
BLOOD CHARACTERISTICS OF HIGH-ALTITUDE NATIVE CHICKEN 1387
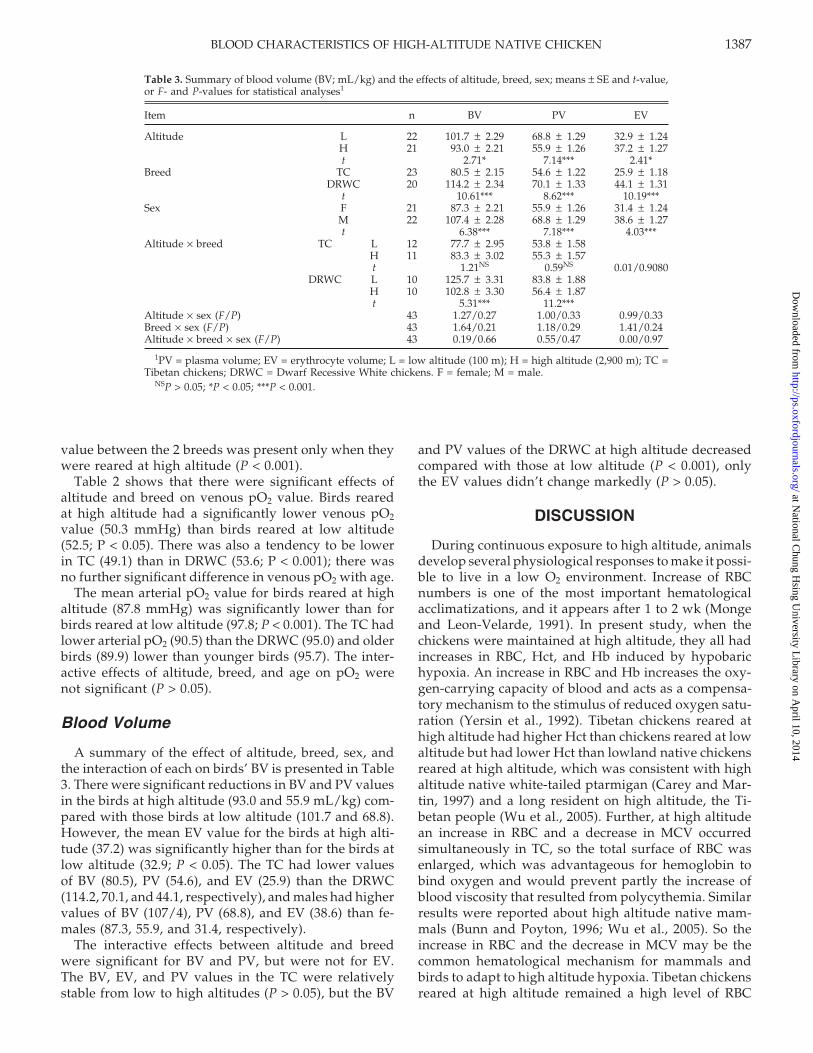
Table 3. Summary of blood volume (BV; mL/kg) and the effects of altitude, breed, sex; means ± SE and t-value,or F- and P-values for statistical analyses1
Item n BV PV EV
Altitude L 22 101.7 ± 2.29 68.8 ± 1.29 32.9 ± 1.24H 21 93.0 ± 2.21 55.9 ± 1.26 37.2 ± 1.27t 2.71* 7.14*** 2.41*
Breed TC 23 80.5 ± 2.15 54.6 ± 1.22 25.9 ± 1.18DRWC 20 114.2 ± 2.34 70.1 ± 1.33 44.1 ± 1.31
t 10.61*** 8.62*** 10.19***Sex F 21 87.3 ± 2.21 55.9 ± 1.26 31.4 ± 1.24
M 22 107.4 ± 2.28 68.8 ± 1.29 38.6 ± 1.27t 6.38*** 7.18*** 4.03***
Altitude × breed TC L 12 77.7 ± 2.95 53.8 ± 1.58H 11 83.3 ± 3.02 55.3 ± 1.57t 1.21NS 0.59NS 0.01/0.9080
DRWC L 10 125.7 ± 3.31 83.8 ± 1.88H 10 102.8 ± 3.30 56.4 ± 1.87t 5.31*** 11.2***
Altitude × sex (F/P) 43 1.27/0.27 1.00/0.33 0.99/0.33Breed × sex (F/P) 43 1.64/0.21 1.18/0.29 1.41/0.24Altitude × breed × sex (F/P) 43 0.19/0.66 0.55/0.47 0.00/0.97
1PV = plasma volume; EV = erythrocyte volume; L = low altitude (100 m); H = high altitude (2,900 m); TC =Tibetan chickens; DRWC = Dwarf Recessive White chickens. F = female; M = male.
NSP > 0.05; *P < 0.05; ***P < 0.001.
value between the 2 breeds was present only when theywere reared at high altitude (P < 0.001).
Table 2 shows that there were significant effects ofaltitude and breed on venous pO2 value. Birds rearedat high altitude had a significantly lower venous pO2
value (50.3 mmHg) than birds reared at low altitude(52.5; P < 0.05). There was also a tendency to be lowerin TC (49.1) than in DRWC (53.6; P < 0.001); there wasno further significant difference in venous pO2 with age.
The mean arterial pO2 value for birds reared at highaltitude (87.8 mmHg) was significantly lower than forbirds reared at low altitude (97.8; P < 0.001). The TC hadlower arterial pO2 (90.5) than the DRWC (95.0) and olderbirds (89.9) lower than younger birds (95.7). The inter-active effects of altitude, breed, and age on pO2 werenot significant (P > 0.05).
Blood Volume
A summary of the effect of altitude, breed, sex, andthe interaction of each on birds’ BV is presented in Table3. There were significant reductions in BV and PV valuesin the birds at high altitude (93.0 and 55.9 mL/kg) com-pared with those birds at low altitude (101.7 and 68.8).However, the mean EV value for the birds at high alti-tude (37.2) was significantly higher than for the birds atlow altitude (32.9; P < 0.05). The TC had lower valuesof BV (80.5), PV (54.6), and EV (25.9) than the DRWC(114.2, 70.1, and 44.1, respectively), and males had highervalues of BV (107/4), PV (68.8), and EV (38.6) than fe-males (87.3, 55.9, and 31.4, respectively).
The interactive effects between altitude and breedwere significant for BV and PV, but were not for EV.The BV, EV, and PV values in the TC were relativelystable from low to high altitudes (P > 0.05), but the BV
and PV values of the DRWC at high altitude decreasedcompared with those at low altitude (P < 0.001), onlythe EV values didn’t change markedly (P > 0.05).
DISCUSSION
During continuous exposure to high altitude, animalsdevelop several physiological responses to make it possi-ble to live in a low O2 environment. Increase of RBCnumbers is one of the most important hematologicalacclimatizations, and it appears after 1 to 2 wk (Mongeand Leon-Velarde, 1991). In present study, when thechickens were maintained at high altitude, they all hadincreases in RBC, Hct, and Hb induced by hypobarichypoxia. An increase in RBC and Hb increases the oxy-gen-carrying capacity of blood and acts as a compensa-tory mechanism to the stimulus of reduced oxygen satu-ration (Yersin et al., 1992). Tibetan chickens reared athigh altitude had higher Hct than chickens reared at lowaltitude but had lower Hct than lowland native chickensreared at high altitude, which was consistent with highaltitude native white-tailed ptarmigan (Carey and Mar-tin, 1997) and a long resident on high altitude, the Ti-betan people (Wu et al., 2005). Further, at high altitudean increase in RBC and a decrease in MCV occurredsimultaneously in TC, so the total surface of RBC wasenlarged, which was advantageous for hemoglobin tobind oxygen and would prevent partly the increase ofblood viscosity that resulted from polycythemia. Similarresults were reported about high altitude native mam-mals (Bunn and Poyton, 1996; Wu et al., 2005). So theincrease in RBC and the decrease in MCV may be thecommon hematological mechanism for mammals andbirds to adapt to high altitude hypoxia. Tibetan chickensreared at high altitude remained a high level of RBC
at National C
hung Hsing U
niversity Library on A
pril 10, 2014http://ps.oxfordjournals.org/
Dow
nloaded from

ZHANG ET AL.1388
and a stable level of Hct from younger to older, butDRWC reared at high altitude presented an increasefrom younger to older in RBC and Hct values. The resultsindicated that at low O2 environment the higher levelof RBC in TC was genetic adaptation but in DRWC wasa physiological compensatory response.
Reduced environmental oxygen availability at highaltitude stimulates ventilation, which plays an importantrole in maintaining an adequate oxygen transfer to theblood for mammals and birds (Monge and Leon-Vel-arde, 1991), but also increases the exhalation of CO2 andresults in hypocapnia (Besch et al., 1971). So the highaltitude natives blunt hypoxic ventilatory response,serving to counteract acid-base problems arising fromhyperventilation (Hochachka et al., 1999). At high alti-tude, TC exhibited significant lower value in pH andhigher value in pCO2 than DRWC. The results indicatedTC were less susceptible to hypocapnia than DRWC un-der conditions of hypoxia. According to Powell (1990),the most important consequence of acclimatization inducks was hypocapnia and not enhanced O2 delivery.During acute severe hypoxia, the pO2 declined in bar-headed geese of the Himalayas and lowland Pekin ducks(Faraci et al., 1984, 1985). Consistent with the results, weobserved that pO2 of TC and DRWC were lower at highaltitude than that at low altitude. Furthermore, TC hadlower pO2 than DRWC, which might be because TChad higher hemoglobin-oxygen affinity of blood thanlowland chickens. The higher affinity may increase oxy-gen saturation of blood and acts as to compensate forthe reduced partial pressure of oxygen and is consideredto be characteristic of genotypically adapted high alti-tude mammals or birds (Monge and Leon-Velarde,1991). A number of montane species, such as bar-headedand Andean geese (Weber et al., 1993) and Tufted ducks(Lutfullah et al., 2005), had special mutations that alteredthe amino acid residues in Hb and increased O2 affinity.We found in the other experiment that the Hb aminoacid sequences of TC had a functional mutation of Met-32(B13)-Leu in αD globin chain, which evidently causesthe increase in oxygen affinity (Gou et al., 2005).
Chronic exposure to hypoxia elicited a decrement inBV and PV for microswine (Durkot et al., 1996) andhuman (Robach et al., 2000; Heinicke et al., 2003). Weobserved a similar result in DRWC. However, in TC thevalues for BV, PV, and EV did not vary when comparedat low and high altitudes. Furthermore, we found thatTC had lower BV, PV, and EV than DRWC, but therewere some reports that many high altitude native ani-mals had higher BV or EV (Birchard and Tenney, 1990;Claydon et al., 2004). There were higher BV, PV, andEV in males than in females, but the effect of sex wasindependent of altitude, so the difference in BV, PV,and EV between males and females was not related tohypoxic response. Whether the amount of BV is in someway related to oxygen transport and hypoxic adaptationrequires further study.
REFERENCES
Besch, E. L., R. R. Burton, and A. H. Smith. 1971. Influence ofchronic hypoxia on blood gas tensions and pH in domesticfowl. Am. J. Physiol. 220:1379–1382.
Birchard, G. F., and S. M. Tenney. 1990. Relationship betweenblood-oxygen affinity and blood volume. Resp. Physiol.83:365–374.
Black, C. P., and S. M. Tenney. 1980. Oxygen transport duringprogressive hypoxia in high-altitude and sea-level water-fowl. Resp. Physiol. 39:217–239.
Bunn, H. F., and R. O. Poyton. 1996. Oxygen sensing andmolecular adaptation to hypoxia. Physiol. Rev. 76:839–885.
Carey, C., and K. Martin. 1997. Physiological ecology of incuba-tion of ptarmigan eggs at high and low altitude. Wildl. Biol.3:211–218.
Claydon, V. E., L. J. Norcliffe, J. P. Moore, M. Rivera-Ch, F.Leon-Velarde, O. Appenzeller, and R. Hainsworth. 2004.Orthostatic tolerance and blood volumes in Andean highaltitude dwellers. Exp. Physiol. 89:565–571.
Dacie, J. V., and S. M. Lewis. 1975. Practical Haematology.Churchill Livingston, Edinburgh, UK.
Durkot, M. J., R. W. Hoyt, A. Darrigrand, L. J. Hubbard, G.H. Kamimori, and A. Cymerman. 1996. Chronic hypobarichypoxia decreases intracellular and total body water in mi-croswine. Comp. Biochem. Physiol. A 114:117–121.
El-Sayed, H., S. R. Goodau, and R. Hainsworth. 1995. Re-evalu-ation of Evans blue dye dilution method of plasma volumemeasurement. Clin. Lab. Haematol. 17:189–194.
Faraci, F. M., D. L. Kilgore Jr, and M. R. Fedde. 1984. Oxygendelivery to the heart and brain during hypoxia: Pekin duckvs. bar-headed goose. Am. J. Physiol. Reg. I. 247: 69-75.
Faraci, F. M., D. L. Kilgore Jr., and M. R. Fedde. 1985. Blood flowdistribution during hypocapnic hypoxia in Pekin ducks andBar-headed geese. Resp. Physiol. 61:21–30.
Gou, X., N. Li, L. Lian, D. Yan, H. Zhang, and C. Wu. 2005.Hypoxia adaptation and hemoglobin mutation in Tibetchick embryo. Sci. China Ser. C 48:616–623.
Heinicke, K., N. Prommer, J. Cajigal, T. Viola, C. Behn, and W.Schmidt. 2003. Long-term exposure to intermittent hypoxiaresults in increased hemoglobin mass, reduced plasma vol-ume, and elevated erythropoietin plasma levels in man.Eur. J. Appl. Physiol. 88:535–543.
Hochachka, P. W., J. L. Rupert, and C. Monge. 1999. Adaptationand conservation of physiological systems in the evolutionof human hypoxia tolerance. Comp. Biochem. Physiol. AMol. Integr. Physiol. 124:1–17.
Leon-Velarde, F., and C. Monge. 2004. Avian embryos in hyp-oxic enviroments. Resp. Physiol. Neurobiol. 141:331–343.
Lutfullah, G., S. A. Ali, and A. A. Abbasi. 2005. Molecularmechanism of high altitude respiration: Primary structure ofa minor hemoglobin component from Tufted duck (Aythyafuligula, Anseriformes). Biochem. Biophys. Res. Commun.326:123–130.
Monge, C., and F. Leon-Velarde. 1991. Physiological adapta-tion to high altitude: Oxygen transport in mammals andbirds. Physiol. Rev. 71:1135–1172.
Powell, F. 1990. Acclimatization to high altitude. Page 41–44in Hypoxia: The Adaptations, J. R. Sutton, G. Coates, andJ. E. Remmers, ed. Decker, Toronto, Canada.
Robach, P., M. Dechele, S. Jarrot, J. Vaysse, J. C. Schneider, N.P. Mason, J. P. Herry, B. Gardette, and J. P. Richalet. 2000.Operation Everest III: Role of plasma volume expansion onO2 max during prolonged high-altitude exposure. J. Appl.Physiol. 89:29–37.
Weber, R. E., T. H. Jessen, H. Malte, and J. Tame. 1993. Mutanthemoglobins (alpha 119-Ala and beta 55-Ser): Functions re-lated to high-altitude respiration in geese. J. Appl. Physiol.75:2646–2655.
Wu, T., X. Wang, C. Wei, H. Cheng, X. Wang, Y. Li, Ge-Dong,H. Zhao, P. Young, G. Li, and Z. Wang. 2005. Hemoglobin
at National C
hung Hsing U
niversity Library on A
pril 10, 2014http://ps.oxfordjournals.org/
Dow
nloaded from

BLOOD CHARACTERISTICS OF HIGH-ALTITUDE NATIVE CHICKEN 1389
levels in Qinghai-Tibet: Different effects of gender for Tibetsvs. Han. J. Appl. Physiol. 98:598–604.
Ye, R., Y. Cao, and Q. Bai. 1994. Blood indices of plateau pikaand relationship with hypoxia adaptation. Acta Anim. Sci.Sinica 2:114–119.
Yersin, A. G., W. E. Huff, L. F. Cubena, M. H. Elissalde, R. B.Harvey, D. A. Witzel, and L. E. Giroir. 1992. Change inhematological, blood gas, and serum biochemical variables
during exposure to simulated high altitude. Avian Dis.36:189–196.
Zhang, H., C. Wu, Y. Chamba, Y. Ling, and Z. Luo. 2006.Adaptability to high altitude and NOS activity of lung inTibetan chicken. J. China Agric. Univ. 11:35–38.
Zhang, H., C. Wu, Y. Chamba, X. Ma, X. Tang, and Pobu.2005. Curve analysis of embryonic mortality in chickensincubated at high altitude. J. China Agric. Univ. 10:109–114.
at National C
hung Hsing U
niversity Library on A
pril 10, 2014http://ps.oxfordjournals.org/
Dow
nloaded from