blocking promiscuous activation at cryptic promoters...
TRANSCRIPT

RESEARCH ARTICLE◥
CELL FATE
Blocking promiscuous activation atcryptic promoters directs celltype–specific gene expressionJongmin Kim,1,2 Chenggang Lu,2* Shrividhya Srinivasan,2* Stephan Awe,3
Alexander Brehm,3 Margaret T. Fuller2,4†
To selectively express cell type–specific transcripts during development, it is critical tomaintain genes required for other lineages in a silent state. Here, we show in the Drosophilamale germline stem cell lineage that a spermatocyte-specific zinc finger protein, Kumgang(Kmg), working with the chromatin remodeler dMi-2 prevents transcription of genesnormally expressed only in somatic lineages. By blocking transcription from normallycryptic promoters, Kmg restricts activation by Aly, a component of the testis-meioticarrest complex, to transcripts for male germ cell differentiation. Our results suggest thatas new regions of the genome become open for transcription during terminaldifferentiation, blocking the action of a promiscuous activator on cryptic promoters is acritical mechanism for specifying precise gene activation.
Highly specialized cell types such as redblood cells, intestinal epithelium, andspermatozoa are produced throughout lifefrom adult stem cells. In such lineages, mi-totically dividing precursors commonly stop
proliferation and initiate a cell type–specific tran-scription program that sets up terminal differen-tiation of the specialized cell type. In theDrosophilamale germ line, stem cells at the apical tip of thetestis self-renew and produce daughter cells thateach undergo four rounds of spermatogonial mi-totic transit amplifying (TA) divisions, after whichthe germ cells execute a final round of DNA syn-thesis (premeiotic S-phase) and initiate terminaldifferentiation as spermatocytes (Fig. 1A) (1). Tran-sition to the spermatocyte state is accompaniedby transcriptional activation of more than 1500genes, many of which are expressed only in malegerm cells (2). Expression of two-thirds of thesedepends both on a testis-specific version of theMMB (Myb-Muv B)/dREAM (Drosophila RBF,dE2F2, and dMyb-interacting proteins) complextermed the testis meiotic arrest complex (tMAC)and on testis-specific paralogs of TATA-bindingprotein–associated factors (tTAFs) (3–5). Althoughthis is one of the most dramatic changes in geneexpression in Drosophila (6), it is not yet under-stood how the testis-specific transcripts are se-
lectively activated during the 3-day spermatocyteperiod.
Identification of an early differentiationgene, kumgang
To identify the first transcripts up-regulated atonset of spermatocyte differentiation (fig. S1A,arrow), we genetically manipulated germ cells tosynchronously differentiate from spermatogoniato spermatocytes in vivo using bam−/− testes,which contain large numbers of overproliferat-ing spermatogonia (fig. S1, B and C) (7, 8). Briefrestoration of Bam expression under heat shockcontrol in hs-bam;bam−/− flies induced synchro-nous differentiation of bam−/− spermatogonia, re-sulting in completion of a finalmitosis, premeioticDNA synthesis, and onset of spermatocyte differ-entiation by 24 hours after Bam expression, even-tually leading to production of functional sperm(Fig. 1B; figs. S1, D to O, and S2; and supplemen-tary text). Comparison bymeans ofmicroarray oftranscripts expressed before versus 24 hours afterheat shock of hs-bam;bam−/− testes identified27 early transcripts that were significantly up-regulated more than twofold in testes fromhs-bam;bam−/− (Fig. 1C, red) but not from bam−/−
flies subjected to the same heat shock regime(fig. S3A). Among these was the early spermato-cyte marker RNA binding protein 4 (Rbp4) (Fig.1C and table S1) (9). At this early time point, thetranscript for CG5204—now named kumgang(kmg), from the Korean name of mythologicalguardians at the gate of Buddhist temples—hadthe greatest increase among all 754 Drosophilapredicted transcription factors (Fig. 1C and tableS1) (10).Kumgang (CG5204) encodes a 747–amino acid
protein with six canonical C2H2-type zinc finger
domains (fig. S4A) expressed in testes but not inovary or carcass (fig. S3B). Kmg protein was ex-pressed independently from the tMAC componentAlways early (Aly) or the tTAF SpermatocyteArrest (Sa) (fig. S3C), and both kmgmRNA andprotein were up-regulated before Topi, anothercomponent of tMAC (fig. S3, D and E) (3, 11). Im-munofluorescence staining of wild-type testesrevealed Kmg protein expressed specifically indifferentiating spermatocytes (Fig. 1D and fig. S3,F toH), where it wasnuclear and enriched on thepartially condensed bivalent chromosomes (Fig. 1,E to E′′). Consistent with dramatic up-regulationof kmgmRNA after the switch from spermatogoniato spermatocyte (Fig. 1C and fig. S3D), expressionofKmgwas first detectedwith immunofluorescencestaining after completion of premeiotic S-phasemarked by down-regulation of Bam (fig. S3, I to I′′)(12), coinciding with expression of Rbp4 protein(fig. S3, J to J′′) and before expression of the tTAFSa (fig. S3, K to K′′).
Kmg prevents misexpression of genesnormally expressed in somatic cells
Function of Kmg in spermatocytes was requiredfor male germ cell differentiation. Reducing func-tion of Kmg in spermatocytes—either by meansof cell type–specific RNA interference (RNAi)knockdown (KD) (Fig. 2, A and B, and fig. S4, Aand B) or in flies trans-heterozygous for a CRISPR(clustered regularly interspaced short palindromicrepeats)–induced kmg frameshift mutant and achromosomal deficiency (kmgD7/Df) (fig. S3, A, C,E, and F)—resulted in accumulation of matureprimary spermatocytes arrested just before theG2/M transition for meiosis I and lack of sper-matid differentiation. A 4.3-kb genomic rescuetransgene containing the 2.3-kb kmg open read-ing frame (fig. S4A) fully rescued the differentia-tion defects and sterility of kmgD7/Df flies (fig.S4D), confirming that themeiotic arrest phenotypewas due to loss of function of Kmg. In both kmgKD and kmgD7/Df, Kmg protein levels were lessthan 5% that of wild type (fig. S4G and supplemen-tary text). kmgD7/Dfmutant animals were adult-viable and female-fertile but male-sterile, whichis consistent with the testis-specific expression.Function of Kmg was required in germ cells for
repression of more than 400 genes not normallyexpressed in wild-type spermatocytes. Althoughthe differentiation defects caused by loss of func-tion of kmg appeared, bymeans of phase contrastmicroscopy, to be similar to the meiotic arrestphenotype of testis-specific tMAC componentmutants, analysis of gene expression in kmg KDtestes showed that many Aly (tMAC)–dependentspermatid differentiation genes were expressed,although some at a lower level than that in wildtype. Among the 652 genes with more than 99%lower expression in aly−/− mutant as comparedwith wild-type testes (Fig. 2D, green dots, andtable S2), only four showed similar reducedexpression in kmg KD as compared with that ofsibling control (no Gal4 driver) testes (Fig. 2C,green dots). In contrast, transcripts from morethan 500 geneswere strongly up-regulated inkmgKD testes, with almost no detectable expression
RESEARCH
Kim et al., Science 356, 717–721 (2017) 19 May 2017 1 of 5
1Department of Chemical and Systems Biology, StanfordUniversity School of Medicine, Stanford, CA 94305-5329,USA. 2Department of Developmental Biology, StanfordUniversity School of Medicine, Stanford, CA 94305-5329,USA. 3Institut für Molekularbiologie und Tumorforschung,Philipps-Universität Marburg, Marburg, Germany.4Department of Genetics, Stanford University School ofMedicine, Stanford, CA 94305-5329, USA.*These authors contributed equally to this work. †Correspondingauthor. Email: [email protected]
on June 8, 2019
http://science.sciencemag.org/
Dow
nloaded from
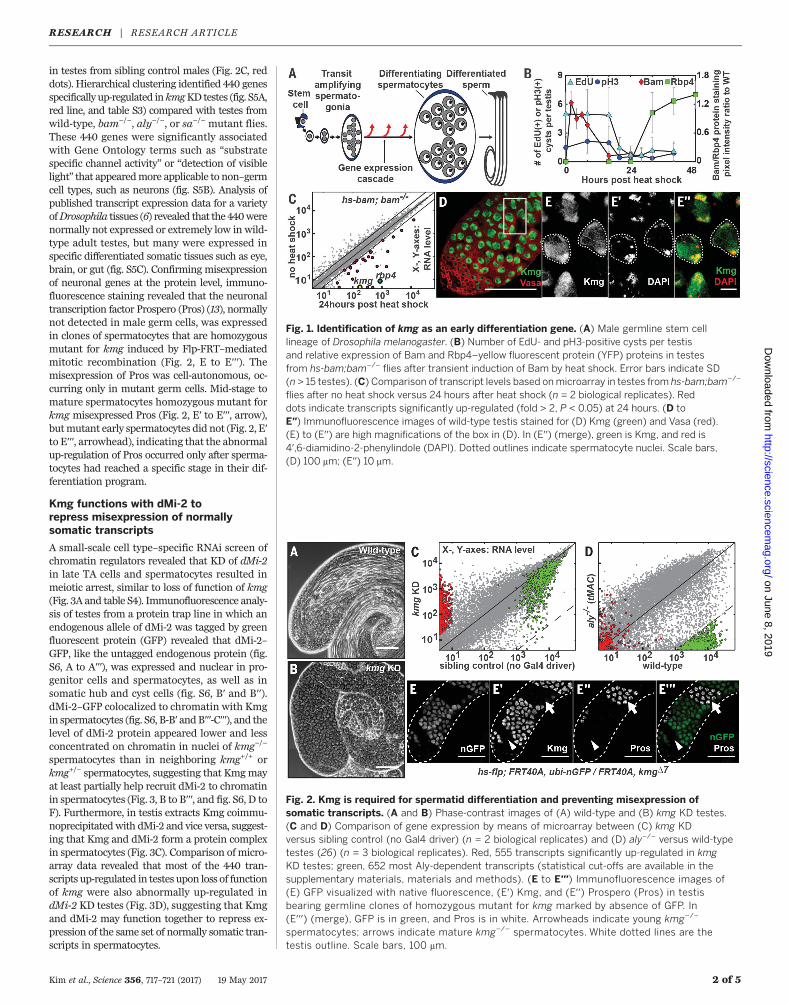
in testes from sibling control males (Fig. 2C, reddots). Hierarchical clustering identified 440 genesspecifically up-regulated inkmgKD testes (fig. S5A,red line, and table S3) compared with testes fromwild-type, bam−/−, aly−/−, or sa−/− mutant flies.These 440 genes were significantly associatedwith Gene Ontology terms such as “substratespecific channel activity” or “detection of visiblelight” that appearedmore applicable to non–germcell types, such as neurons (fig. S5B). Analysis ofpublished transcript expression data for a varietyofDrosophila tissues (6) revealed that the 440werenormally not expressed or extremely low in wild-type adult testes, but many were expressed inspecific differentiated somatic tissues such as eye,brain, or gut (fig. S5C). Confirming misexpressionof neuronal genes at the protein level, immuno-fluorescence staining revealed that the neuronaltranscription factor Prospero (Pros) (13), normallynot detected in male germ cells, was expressedin clones of spermatocytes that are homozygousmutant for kmg induced by Flp-FRT–mediatedmitotic recombination (Fig. 2, E to E′′′). Themisexpression of Pros was cell-autonomous, oc-curring only in mutant germ cells. Mid-stage tomature spermatocytes homozygous mutant forkmg misexpressed Pros (Fig. 2, E′ to E′′′, arrow),butmutant early spermatocytes did not (Fig. 2, E′to E′′′, arrowhead), indicating that the abnormalup-regulation of Pros occurred only after sperma-tocytes had reached a specific stage in their dif-ferentiation program.
Kmg functions with dMi-2 torepress misexpression of normallysomatic transcripts
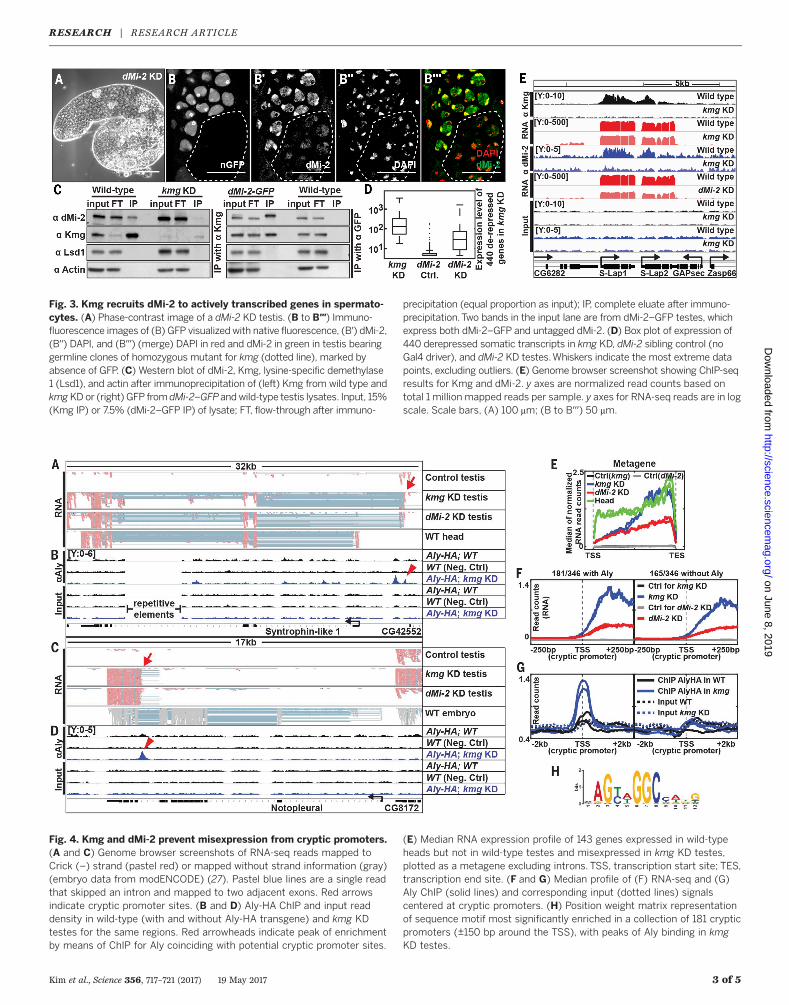
A small-scale cell type–specific RNAi screen ofchromatin regulators revealed that KD of dMi-2in late TA cells and spermatocytes resulted inmeiotic arrest, similar to loss of function of kmg(Fig. 3A and table S4). Immunofluorescence analy-sis of testes from a protein trap line in which anendogenous allele of dMi-2 was tagged by greenfluorescent protein (GFP) revealed that dMi-2–GFP, like the untagged endogenous protein (fig.S6, A to A′′′), was expressed and nuclear in pro-genitor cells and spermatocytes, as well as insomatic hub and cyst cells (fig. S6, B′ and B′′).dMi-2–GFP colocalized to chromatin with Kmgin spermatocytes (fig. S6, B-B′ andB′′′-C′′′), and thelevel of dMi-2 protein appeared lower and lessconcentrated on chromatin in nuclei of kmg−/−
spermatocytes than in neighboring kmg+/+ orkmg+/− spermatocytes, suggesting that Kmgmayat least partially help recruit dMi-2 to chromatinin spermatocytes (Fig. 3, B to B′′′, and fig. S6, D toF). Furthermore, in testis extracts Kmg coimmu-noprecipitatedwith dMi-2 and vice versa, suggest-ing that Kmg and dMi-2 form a protein complexin spermatocytes (Fig. 3C). Comparison of micro-array data revealed that most of the 440 tran-scripts up-regulated in testes upon loss of functionof kmg were also abnormally up-regulated indMi-2 KD testes (Fig. 3D), suggesting that Kmgand dMi-2 may function together to repress ex-pression of the same set of normally somatic tran-scripts in spermatocytes.
Kim et al., Science 356, 717–721 (2017) 19 May 2017 2 of 5
Fig. 1. Identification of kmg as an early differentiation gene. (A) Male germline stem celllineage of Drosophila melanogaster. (B) Number of EdU- and pH3-positive cysts per testisand relative expression of Bam and Rbp4–yellow fluorescent protein (YFP) proteins in testesfrom hs-bam;bam−/− flies after transient induction of Bam by heat shock. Error bars indicate SD(n > 15 testes). (C) Comparison of transcript levels based onmicroarray in testes from hs-bam;bam−/−
flies after no heat shock versus 24 hours after heat shock (n = 2 biological replicates). Reddots indicate transcripts significantly up-regulated (fold > 2, P < 0.05) at 24 hours. (D toE′′) Immunofluorescence images of wild-type testis stained for (D) Kmg (green) and Vasa (red).(E) to (E′′) are high magnifications of the box in (D). In (E′′) (merge), green is Kmg, and red is4′,6-diamidino-2-phenylindole (DAPI). Dotted outlines indicate spermatocyte nuclei. Scale bars,(D) 100 mm; (E′′) 10 mm.
Fig. 2. Kmg is required for spermatid differentiation and preventing misexpression ofsomatic transcripts. (A and B) Phase-contrast images of (A) wild-type and (B) kmg KD testes.(C and D) Comparison of gene expression by means of microarray between (C) kmg KDversus sibling control (no Gal4 driver) (n = 2 biological replicates) and (D) aly−/− versus wild-typetestes (26) (n = 3 biological replicates). Red, 555 transcripts significantly up-regulated in kmgKD testes; green, 652 most Aly-dependent transcripts (statistical cut-offs are available in thesupplementary materials, materials and methods). (E to E′′′) Immunofluorescence images of(E) GFP visualized with native fluorescence, (E′) Kmg, and (E′′) Prospero (Pros) in testisbearing germline clones of homozygous mutant for kmg marked by absence of GFP. In(E′′′) (merge), GFP is in green, and Pros is in white. Arrowheads indicate young kmg−/−
spermatocytes; arrows indicate mature kmg−/− spermatocytes. White dotted lines are thetestis outline. Scale bars, 100 mm.
RESEARCH | RESEARCH ARTICLEon June 8, 2019
http://science.sciencemag.org/
Dow
nloaded from
Kim et al., Science 356, 717–721 (2017) 19 May 2017 3 of 5
Fig. 3. Kmg recruits dMi-2 to actively transcribed genes in spermato-cytes. (A) Phase-contrast image of a dMi-2 KD testis. (B to B′′′) Immuno-fluorescence images of (B) GFP visualized with native fluorescence, (B′) dMi-2,(B′′) DAPI, and (B′′′) (merge) DAPI in red and dMi-2 in green in testis bearinggermline clones of homozygous mutant for kmg (dotted line), marked byabsence of GFP. (C) Western blot of dMi-2, Kmg, lysine-specific demethylase1 (Lsd1), and actin after immunoprecipitation of (left) Kmg from wild type andkmgKDor (right) GFP from dMi-2–GFPandwild-type testis lysates. Input, 15%(Kmg IP) or 7.5% (dMi-2–GFP IP) of lysate; FT, flow-through after immuno-
precipitation (equal proportion as input); IP, complete eluate after immuno-precipitation.Two bands in the input lane are from dMi-2–GFP testes, whichexpress both dMi-2–GFP and untagged dMi-2. (D) Box plot of expression of440 derepressed somatic transcripts in kmg KD, dMi-2 sibling control (noGal4 driver), and dMi-2 KD testes.Whiskers indicate the most extreme datapoints, excluding outliers. (E) Genome browser screenshot showing ChIP-seqresults for Kmg and dMi-2. y axes are normalized read counts based ontotal 1 million mapped reads per sample. y axes for RNA-seq reads are in logscale. Scale bars, (A) 100 mm; (B to B′′′) 50 mm.
Fig. 4. Kmg and dMi-2 prevent misexpression from cryptic promoters.(A and C) Genome browser screenshots of RNA-seq reads mapped toCrick (–) strand (pastel red) or mapped without strand information (gray)(embryo data from modENCODE) (27). Pastel blue lines are a single readthat skipped an intron and mapped to two adjacent exons. Red arrowsindicate cryptic promoter sites. (B and D) Aly-HA ChIP and input readdensity in wild-type (with and without Aly-HA transgene) and kmg KDtestes for the same regions. Red arrowheads indicate peak of enrichmentby means of ChIP for Aly coinciding with potential cryptic promoter sites.
(E) Median RNA expression profile of 143 genes expressed in wild-typeheads but not in wild-type testes and misexpressed in kmg KD testes,plotted as a metagene excluding introns. TSS, transcription start site; TES,transcription end site. (F and G) Median profile of (F) RNA-seq and (G)Aly ChIP (solid lines) and corresponding input (dotted lines) signalscentered at cryptic promoters. (H) Position weight matrix representationof sequence motif most significantly enriched in a collection of 181 crypticpromoters (±150 bp around the TSS), with peaks of Aly binding in kmgKD testes.
RESEARCH | RESEARCH ARTICLEon June 8, 2019
http://science.sciencemag.org/
Dow
nloaded from

Chromatin immunoprecipitation followed bysequencing (ChIP-seq) revealed that Kmg proteinlocalized along the bodies of genes actively tran-scribed in the testis. ChIP-seq with antibody toKmg identified 798 genomic regions strongly en-riched by immunoprecipitation of Kmg (peaks,q value < 10−10) fromwild-type but not from kmgKD testes. Of the 798 robust Kmg ChIP-seq peaks,698 overlapped with exonic regions of 680 dif-ferent genes actively transcribed in testes (Fig. 3E;fig. S7, A, C, E, G, and I; and table S5). The en-richment was often strongest just downstream ofthe transcription start site (TSS), but with sub-stantial enrichment along the gene body as well(Fig. 3E and fig. S7, A, C, E, and G).ChIP-seq with antibody to dMi-2 also showed
enrichment along the gene bodies of the same680 genes bound by Kmg, with a similar bias justdownstream of the TSS (Fig. 3E and fig. S7, B, D,F, and H). The dMi-2 ChIP signal along thesegenes was partially reduced in kmg KD testes,suggesting that Kmg may recruit dMi-2 to thebodies of genes actively transcribed in the testis(Fig. 3E and fig. S7, B, D, F, and H).RNA-seq analysis revealed that the 680 genes
bound by Kmgwere strongly expressed in testes(Fig. 3E and fig. S7, I to K) and most stronglyenriched in the GO term categories “spermato-genesis” and “male gamete generation” (fig. S8A).One-third of the genes bound by Kmg were ro-bustly activated as spermatogonia differentiateinto spermatocytes and were much more highlyexpressed in the testes than in other tissues (fig.S8B, red rectangle). The median levels of tran-script expression of most of the 680 Kmg boundgenes did not show appreciable change upon lossof Kmg (fig. S7, J and K).Genes that are normally transcribed in somat-
ic cells that became up-regulated upon loss ofKmg function in spermatocytes for the most partdid not appear to be bound byKmg.Only 3 of the440 genes up-regulated in kmg KD overlappedwith the 680 genes with robust Kmg peaks, sug-gesting that Kmg may prevent misexpression ofnormally somatic transcripts either indirectly orby acting at a distance.
Kmg and dMi-2 prevent misexpressionfrom cryptic promoters
Inspection of RNA-seq reads from kmg and dMi-2KD testes mapped onto the genome showed that
~80% of the transcripts that were detected withmicroarray analysis as misexpressed in KD ascompared with wild-type testes did not initiatefrom the promoters used in the somatic tissues inwhich the genes are normally expressed (Fig. 4, Aand C, arrows). Metagene analysis (Fig. 4E), aswell as visualization of RNA expression centeredon the TSSs annotated in the Ensembl database(fig. S9A), showed that most of the 143 genesthat are normally expressed in wild-type headsbut not in wild-type testes weremisexpressed inkmg or dMi-2 KD testes from a start site differ-ent from the annotated TSS used in heads. Tran-script assembly from our RNA-seq data by usingCufflinks for the 143 genes also showed that thetranscripts that aremisexpressed in kmg or dMi-2KD testesmost often initiate fromdifferent TSSsthan the transcripts fromthe samegeneassembledfromwild-type heads (fig. S9, B to D, and supple-mentary text).Of the 440 genes scored via microarray as
derepressed in kmg KD testes, 346 could be as-signed with TSSs in kmg KD testes based onvisual inspection of our RNA-seq data mappedonto the genome browser (table S6 and supple-mentary text). Of these, only 67 produced tran-scripts in kmg KD testes that started within 100basepairs (bp) of theTSSannotated in theEnsembldatabase, based on the tissue(s) in which thegene was normally expressed. In contrast, for therest of the 346 genes, the transcripts expressed inkmgKD testes started from either a TSS upstream(131 of 346) or downstream (148 of 346) of the an-notated TSSs. Of the 346 genes, 262 were misex-pressed starting from nearly identical positionsin dMi-2 KD as in kmg KD testes (table S6), sug-gesting that Kmg and dMi-2 function together toprevent misexpression from cryptic promoters.
Kmg prevents promiscuous activityof Aly
Many of the ectopic promoters from which themisexpressed transcripts originated appeared tobe bound by Aly, a component of tMAC, in kmgKD testes (Fig. 4, B and D, arrowheads, and Fand G). ChIP for Aly was performed by usingantibody to hemagglutinin (HA) on testis extractsfrom flies bearing an Aly-HA genomic transgeneable to fully rescue the aly−/− phenotype (fig. S10and materials and methods). Of 346 genes withnew TSSs assigned via visual inspection, 181 had
a region of significant enrichment for Aly as de-tected with ChIP, with its peak summit locatedwithin 100 bp of the cryptic promoter (Fig. 4, Fand G, and supplementary text). Motif analysisby means of MEME revealed that these regionswere enriched for the DNA sequence motif(AGYWGGC) (Fig. 4H and fig. S11). This motifwas not significantly enriched in the set of 165cryptic promoters at which Aly was not detectedin kmgKD testes (fig. S11B). Enrichment of Aly atthe cryptic promoters wasmuch stronger in kmgKD as compared with wild-type testes (Fig. 4, B,D, andG), suggesting that in the absence of Kmg,Aly may bind to and activate misexpression fromcryptic promoters.Genetic tests revealed that the misexpression
of somatic transcripts in kmg KD spermatocytesindeed required function of Aly. The neuronaltranscription factor Pros, abnormally up-regulatedin kmg KD or mutant spermatocytes (Fig. 2, E toE′′′), was no longer misexpressed if the kmg KDspermatocytes were also mutant for aly (Fig. 5, Aand B, and fig. S12, A to F), even though germcells in kmg KD;aly−/− testes appear to reach thedifferentiation stage at which Pros turned on inthe kmg KD germ cells (fig. S13 and supplemen-tary text). Assessment by means of quantitativereverse transcription polymerase chain reaction(RT-PCR) revealed that misexpression of five outof five transcripts in kmgKD testes also requiredfunction of Aly (fig. S12G). Global transcriptomeanalysis via microarray of kmg KD versus kmgKD;aly−/− testes showed that the majority of the440 genes that were derepressed because of lossof function of kmg in spermatocyteswere no longerabnormally up-regulated in kmgKD;aly−/− testes(Fig. 5C). Even genes without noticeable bindingof Aly at their cryptic promoters were suppressedin kmg KD;aly−/−, suggesting that Aly may regu-late this group of genes indirectly.Together, our ChIP and RNA-seq data show
that Kmg and dMi-2 bind actively transcribedgenes (Fig. 3E and fig. S7) (14) but are requiredto block expression of aberrant transcripts fromother genes that are normally silent in testes. Themammalian ortholog of dMi-2, CHD4 (Mi-2b), hasbeen shown to bind active genes in mouse embry-onic stem cells (15) or T lymphocyte precursors(16) but also plays a role in ensuring lineage-specific gene expression in other contexts (17, 18).We cannot rule out that Kmg and dMi-2 mightalso act directly at the cryptic promoter sites butthat our ChIP conditions did not capture theirtransient or dynamic binding because severalchromatin remodelers or transcription factors,such as the thyroid hormone receptor, have beendifficult to detect with ChIP (19, 20). Kmg anddMi-2may repressmisexpression from cryptic pro-moters indirectly by activating as-yet-unidentifiedrepressor proteins. However, it is also possiblethat Kmg and dMi-2 act at a distance by modu-lating chromatin structure or confining transcrip-tional initiation or elongation licensing machineryto normally active genes.Changes in the genomic localization of Aly
protein in wild-type versus kmg KD testes raisedthe possibility that Kmg may in part prevent
Kim et al., Science 356, 717–721 (2017) 19 May 2017 4 of 5
Fig. 5. Aly is required for misexpression of aberrant transcripts in kmg KD testes. (A and B)Immunofluorescence images of (A) kmg KD and (B) kmg KD;aly−/− testes stained for Vasa (red) andPros (light blue). Scale bars, 100 mm. (C) Box plot of expression of 440 somatic transcriptsderepressed in kmg KD testes, showing transcript levels in kmg sibling control (no Gal4 driver), kmgKD, and kmg KD;aly−/− testes. Whiskers indicate the most extreme data points, excluding outliers.
RESEARCH | RESEARCH ARTICLEon June 8, 2019
http://science.sciencemag.org/
Dow
nloaded from
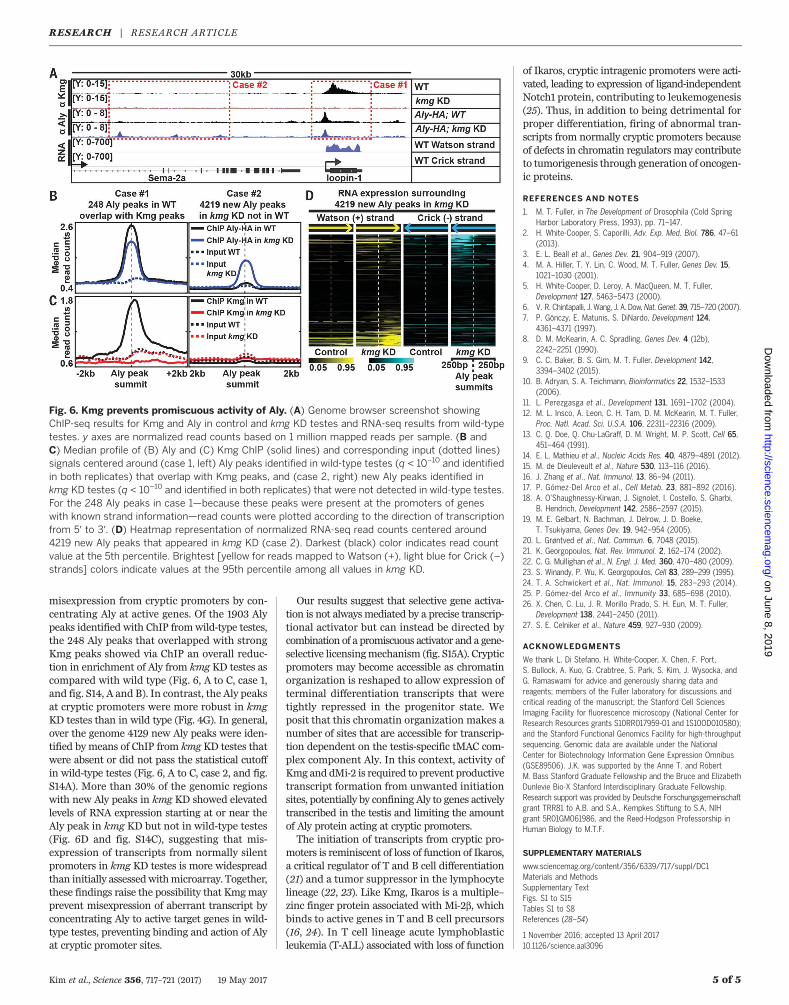
misexpression from cryptic promoters by con-centrating Aly at active genes. Of the 1903 Alypeaks identified with ChIP fromwild-type testes,the 248 Aly peaks that overlapped with strongKmg peaks showed via ChIP an overall reduc-tion in enrichment of Aly from kmg KD testes ascompared with wild type (Fig. 6, A to C, case 1,and fig. S14, A and B). In contrast, the Aly peaksat cryptic promoters were more robust in kmgKD testes than in wild type (Fig. 4G). In general,over the genome 4129 new Aly peaks were iden-tified by means of ChIP from kmg KD testes thatwere absent or did not pass the statistical cutoffin wild-type testes (Fig. 6, A to C, case 2, and fig.S14A). More than 30% of the genomic regionswith new Aly peaks in kmg KD showed elevatedlevels of RNA expression starting at or near theAly peak in kmg KD but not in wild-type testes(Fig. 6D and fig. S14C), suggesting that mis-expression of transcripts from normally silentpromoters in kmg KD testes is more widespreadthan initially assessedwithmicroarray. Together,these findings raise the possibility that Kmgmayprevent misexpression of aberrant transcript byconcentrating Aly to active target genes in wild-type testes, preventing binding and action of Alyat cryptic promoter sites.
Our results suggest that selective gene activa-tion is not alwaysmediated by a precise transcrip-tional activator but can instead be directed bycombination of a promiscuous activator and a gene-selective licensingmechanism (fig. S15A). Crypticpromoters may become accessible as chromatinorganization is reshaped to allow expression ofterminal differentiation transcripts that weretightly repressed in the progenitor state. Weposit that this chromatin organization makes anumber of sites that are accessible for transcrip-tion dependent on the testis-specific tMAC com-plex component Aly. In this context, activity ofKmg and dMi-2 is required to prevent productivetranscript formation from unwanted initiationsites, potentially by confining Aly to genes activelytranscribed in the testis and limiting the amountof Aly protein acting at cryptic promoters.The initiation of transcripts from cryptic pro-
moters is reminiscent of loss of function of Ikaros,a critical regulator of T and B cell differentiation(21) and a tumor suppressor in the lymphocytelineage (22, 23). Like Kmg, Ikaros is a multiple–zinc finger protein associated with Mi-2b, whichbinds to active genes in T and B cell precursors(16, 24). In T cell lineage acute lymphoblasticleukemia (T-ALL) associated with loss of function
of Ikaros, cryptic intragenic promoters were acti-vated, leading to expression of ligand-independentNotch1 protein, contributing to leukemogenesis(25). Thus, in addition to being detrimental forproper differentiation, firing of abnormal tran-scripts from normally cryptic promoters becauseof defects in chromatin regulators may contributeto tumorigenesis through generation of oncogen-ic proteins.
REFERENCES AND NOTES
1. M. T. Fuller, in The Development of Drosophila (Cold SpringHarbor Laboratory Press, 1993), pp. 71–147.
2. H. White-Cooper, S. Caporilli, Adv. Exp. Med. Biol. 786, 47–61(2013).
3. E. L. Beall et al., Genes Dev. 21, 904–919 (2007).4. M. A. Hiller, T. Y. Lin, C. Wood, M. T. Fuller, Genes Dev. 15,
1021–1030 (2001).5. H. White-Cooper, D. Leroy, A. MacQueen, M. T. Fuller,
Development 127, 5463–5473 (2000).6. V. R. Chintapalli, J. Wang, J. A. Dow,Nat. Genet. 39, 715–720 (2007).7. P. Gönczy, E. Matunis, S. DiNardo, Development 124,
4361–4371 (1997).8. D. M. McKearin, A. C. Spradling, Genes Dev. 4 (12b),
2242–2251 (1990).9. C. C. Baker, B. S. Gim, M. T. Fuller, Development 142,
3394–3402 (2015).10. B. Adryan, S. A. Teichmann, Bioinformatics 22, 1532–1533
(2006).11. L. Perezgasga et al., Development 131, 1691–1702 (2004).12. M. L. Insco, A. Leon, C. H. Tam, D. M. McKearin, M. T. Fuller,
Proc. Natl. Acad. Sci. U.S.A. 106, 22311–22316 (2009).13. C. Q. Doe, Q. Chu-LaGraff, D. M. Wright, M. P. Scott, Cell 65,
451–464 (1991).14. E. L. Mathieu et al., Nucleic Acids Res. 40, 4879–4891 (2012).15. M. de Dieuleveult et al., Nature 530, 113–116 (2016).16. J. Zhang et al., Nat. Immunol. 13, 86–94 (2011).17. P. Gómez-Del Arco et al., Cell Metab. 23, 881–892 (2016).18. A. O’Shaughnessy-Kirwan, J. Signolet, I. Costello, S. Gharbi,
B. Hendrich, Development 142, 2586–2597 (2015).19. M. E. Gelbart, N. Bachman, J. Delrow, J. D. Boeke,
T. Tsukiyama, Genes Dev. 19, 942–954 (2005).20. L. Grøntved et al., Nat. Commun. 6, 7048 (2015).21. K. Georgopoulos, Nat. Rev. Immunol. 2, 162–174 (2002).22. C. G. Mullighan et al., N. Engl. J. Med. 360, 470–480 (2009).23. S. Winandy, P. Wu, K. Georgopoulos, Cell 83, 289–299 (1995).24. T. A. Schwickert et al., Nat. Immunol. 15, 283–293 (2014).25. P. Gómez-del Arco et al., Immunity 33, 685–698 (2010).26. X. Chen, C. Lu, J. R. Morillo Prado, S. H. Eun, M. T. Fuller,
Development 138, 2441–2450 (2011).27. S. E. Celniker et al., Nature 459, 927–930 (2009).
ACKNOWLEDGMENTS
We thank L. Di Stefano, H. White-Cooper, X. Chen, F. Port,S. Bullock, A. Kuo, G. Crabtree, S. Park, S. Kim, J. Wysocka, andG. Ramaswami for advice and generously sharing data andreagents; members of the Fuller laboratory for discussions andcritical reading of the manuscript; the Stanford Cell SciencesImaging Facility for fluorescence microscopy (National Center forResearch Resources grants S10RR017959-01 and 1S10OD010580);and the Stanford Functional Genomics Facility for high-throughputsequencing. Genomic data are available under the NationalCenter for Biotechnology Information Gene Expression Omnibus(GSE89506). J.K. was supported by the Anne T. and RobertM. Bass Stanford Graduate Fellowship and the Bruce and ElizabethDunlevie Bio-X Stanford Interdisciplinary Graduate Fellowship.Research support was provided by Deutsche Forschungsgemeinschaftgrant TRR81 to A.B. and S.A., Kempkes Stiftung to S.A, NIHgrant 5R01GM061986, and the Reed-Hodgson Professorship inHuman Biology to M.T.F.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/356/6339/717/suppl/DC1Materials and MethodsSupplementary TextFigs. S1 to S15Tables S1 to S8References (28–54)
1 November 2016; accepted 13 April 201710.1126/science.aal3096
Kim et al., Science 356, 717–721 (2017) 19 May 2017 5 of 5
Fig. 6. Kmg prevents promiscuous activity of Aly. (A) Genome browser screenshot showingChIP-seq results for Kmg and Aly in control and kmg KD testes and RNA-seq results from wild-typetestes. y axes are normalized read counts based on 1 million mapped reads per sample. (B andC) Median profile of (B) Aly and (C) Kmg ChIP (solid lines) and corresponding input (dotted lines)signals centered around (case 1, left) Aly peaks identified in wild-type testes (q < 10−10 and identifiedin both replicates) that overlap with Kmg peaks, and (case 2, right) new Aly peaks identified inkmg KD testes (q < 10−10 and identified in both replicates) that were not detected in wild-type testes.For the 248 Aly peaks in case 1—because these peaks were present at the promoters of geneswith known strand information—read counts were plotted according to the direction of transcriptionfrom 5′ to 3′. (D) Heatmap representation of normalized RNA-seq read counts centered around4219 new Aly peaks that appeared in kmg KD (case 2). Darkest (black) color indicates read countvalue at the 5th percentile. Brightest [yellow for reads mapped to Watson (+), light blue for Crick (–)strands] colors indicate values at the 95th percentile among all values in kmg KD.
RESEARCH | RESEARCH ARTICLEon June 8, 2019
http://science.sciencemag.org/
Dow
nloaded from

expressionspecific gene−Blocking promiscuous activation at cryptic promoters directs cell type
Jongmin Kim, Chenggang Lu, Shrividhya Srinivasan, Stephan Awe, Alexander Brehm and Margaret T. Fuller
DOI: 10.1126/science.aal3096 (6339), 717-721.356Science
, this issue p. 717Scienceprevented aberrant gene expression and enabled proper differentiation in the adult sperm stem cell lineage.
factorsprotein and a chromatin remodeler were found to act together to block transcription from cryptic promoters. These zinc finger− male germ line. A multipleDrosophila identified such a mechanism in the et al.other cell lineages. Kim
specific gene expression. However, mechanisms are also needed to block expression of genes that specify−cell typeTo generate cells with a specific identity, only a subset of genes is used. Most studies focus on factors that turn on
Blocking somatic genes to make sperm
ARTICLE TOOLS http://science.sciencemag.org/content/356/6339/717
MATERIALSSUPPLEMENTARY http://science.sciencemag.org/content/suppl/2017/05/17/356.6339.717.DC1
REFERENCES
http://science.sciencemag.org/content/356/6339/717#BIBLThis article cites 53 articles, 20 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.Sciencelicensee American Association for the Advancement of Science. No claim to original U.S. Government Works. The title Science, 1200 New York Avenue NW, Washington, DC 20005. 2017 © The Authors, some rights reserved; exclusive
(print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
on June 8, 2019
http://science.sciencemag.org/
Dow
nloaded from