biogeography and microhabitat variation in temperate algal-invertebrate symbioses: zooxanthellae and...
TRANSCRIPT

Biogeography and Microhabitat Variation in Temperate Algal-Invertebrate Symbioses:Zooxanthellae and Zoochlorellae in Two Pacific Intertidal Sea Anemones, Anthopleuraelegantissima and A. xanthogrammicaAuthor(s): David Secord and Leon AugustineSource: Invertebrate Biology, Vol. 119, No. 2 (Spring, 2000), pp. 139-146Published by: Wiley on behalf of American Microscopical SocietyStable URL: http://www.jstor.org/stable/3227210 .
Accessed: 09/06/2014 17:42
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Wiley and American Microscopical Society are collaborating with JSTOR to digitize, preserve and extendaccess to Invertebrate Biology.
http://www.jstor.org
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions

Invertebrate Biology 119(2): 139-146. ( 2000 American Microscopical Society, Inc.
Biogeography and microhabitat variation in temperate algal-invertebrate symbioses: zooxanthellae and zoochlorellae in two Pacific intertidal
sea anemones, Anthopleura elegantissima and A. xanthogrammica
David Secordl,2,a and Leon Augustine2
University of Washington at Tacoma, Interdisciplinary Arts and Sciences, Environmental Science, Tacoma, WA 98402-3100, USA
2Friday Harbor Laboratories, Friday Harbor, WA 98250, USA
Abstract. Temperate sea anemones in the genus Anthopleura are unique among cnidarians in harboring two phylogenetically distinct symbiotic algae, zooxanthellae (golden-brown dino- phytes, Symbiodinium) and zoochlorellae (green chlorophytes). To determine whether their
physiological differences generate patterns in anemone habitat and biogeographic distribution, we sampled symbiotic algae in the small clonal A. elegantissima and the large solitary A. xanthogrammica at 8 field sites (and the other large solitary Anthopleura species at one site) spanning 18? of latitude along 2500 km of the Pacific coast of North America. We found that zoochlorellae predominate in low intertidal habitats and northerly latitudes and in A. xantho-
grammica, while zooxanthellae constitute the majority of symbionts in high intertidal habitats and more southerly latitudes and in A. elegantissima. These data are consistent with published predictions based on photosynthetic efficiency of the two algae under varied temperature and
light regimes in the laboratory. This anemone-algal system provides a potential biological signal of benthic intertidal communities' responses to El Nino events and long-term climate changes in the Pacific.
Additional key words: latitudinal gradient, climate change, El Nino, Symbiodinium, Chlorophyta, Cnidaria, Anthozoa
Hundreds of aquatic invertebrate species in at least a dozen phyla engage in symbiosis with unicellular algae (Buchner 1965). Historically, biologists have thought of these endosymbioses in terms of a single algal species or type inhabiting one or more host spe- cies (e.g., zooxanthellae in widespread symbioses with hermatypic corals). However, we now recognize a great deal of diversity among dinophyte symbionts in different invertebrate taxa (Blank & Trench 1985). Molecular genetic techniques have revealed that dif- ferent genotypes, if not taxa, of zooxanthellae co-occur in the same host species (Rowan 1991; Rowan & Pow- ers 1991a,b, 1992; Rowan & Knowlton 1995; Rowan et al. 1997). Moreover, these different algal types ap- pear to be correlated with different microhabitats and associated physical conditions on reefs (Rowan & Knowlton 1995), implying plastic responses by coral- algal associations to environmental change (Budde- meier & Fautin 1993; Rowan et al. 1997).
a Author for correspondence. E-mail: [email protected]
In contrast to hermatypic corals, whose obligate symbionts are simply genetic variants of zooxanthel- lae, some temperate sea anemones in the genus Antho- pleura harbor two distinct symbionts in different algal phyla. The large solitary anemone A. xanthogrammica BRANDT 1835 and its small clonal congener A. elegan- tissima BRANDT 1835 can both contain zooxanthellae (Symbiodinium sp., golden-brown members of the Di- nophyta) or zoochlorellae, unicellular green chloro- phytes unidentified below the phylum level (Muscatine 1971; O'Brien 1980; O'Brien & Wyttenbach 1980; Engebretson & Muller-Parker 1999; R.K. Trench, pers. comm.). The two symbiont species may occur together in the same clone of A. elegantissima, as mixed cells in the same polyp (Muscatine 1971; O'Brien & Wyt- tenbach 1980), and even inside the same host cell (B. Timmerman, pers. comm.). Cells of zooxanthellae and zoochlorellae differ in color and size and are readily distinguishable by means of light microscopy (this does not rule out invisible genetic differences within each algal taxon, however). Therefore, they potentially offer an unusually accessible system for studying how
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions

Secord & Augustine
0so
.Eugene,
/ Coos Bay
. t
.. 'b ,
Euret
40
I' e
3S ..
? 7 . .
Fig. 1. Arrows indicate collecting sites on the west coast of North America. See Table 1 and Methods for details.
environmental change influences intimate species in- teractions in the sea.
Members of the genus Anthopleura are perhaps the most abundant rocky intertidal anemones in the north- east Pacific Ocean (Kozloff 1983; Ricketts et al. 1985), occupying significant amounts of intertidal space (Dayton 1971), preying on zooplankton and benthic invertebrates (Sebens 1981), and, as symbiotic asso- ciations, contributing to primary productivity on a par with macroalgae (Fitt et al. 1982). The two host anem- one species considered here are continuously sympat- ric from southeast Alaska to central California, and patchily so south to northern Baja California, Mexico
(Hand 1955; Francis 1979; Kozloff 1983). Since algal symbionts comprise 15-50% of the volume and pro- tein biomass of their cnidarian hosts (Shick 1991), zo- oxanthellae and zoochlorellae in Anthopleura may be among the most prevalent rocky intertidal organisms on the west coast of North America.
Spatial patterns in the association between these two hosts and their two symbionts were assessed in a rapid, geographically extensive field sampling regime (Fig. 1; but see Table 1). We sampled symbionts on a local spatial scale (high and low intertidal habitats, and large-bodied and small-bodied host species within sites) and on a biogeographic one (the majority of the latitudinal range of both host species). A rapid assess- ment allowed us to generate a "snapshot" view of symbiont ratios, without measuring the possible con- founding effects of seasonal or longer-term variation not addressed in this study. We sought to test the hy- pothesis that the field distribution of the two algae in hospite reflects known differences in their rates of pho- tosynthesis and cell division in the laboratory (Mc- Closkey et al. 1996; Verde & McCloskey 1996; Saun- ders & Muller-Parker 1997).
Specifically, these laboratory data suggest that zoochlorellae photosynthesize more efficiently and grow faster at lower temperatures and light, whereas zooxanthellae do so under higher temperature or light regimes (O'Brien & Wyttenbach 1980; McCloskey et al. 1996; Verde & McCloskey 1996; Saunders & Mull- er-Parker 1997; G. Muller-Parker, unpubl. data). Dur- ing daytime low tides, internal body temperatures are higher in smaller anemones and in those living in the upper intertidal (Dingman 1998). Therefore, we pre- dicted that our field survey of the two algae would find greater proportions of zoochlorellae in cooler hab- itats (low intertidal, larger host species) and at more northerly latitudes, and greater proportions of zooxan- thellae in warmer habitats (high intertidal, smaller host species) and at more southerly latitudes (Secord 1995; Dingman 1998).
Methods
We surveyed endosymbiotic algal cells by collecting anemone tissue at 9 exposed rocky intertidal sites along a 2500 km transect, spanning 18? of latitude along the Pacific coast from northern Washington State to northern Baja California, Mexico (Table 1, Fig. 1). To exclude the effects of seasonal and interannual var- iation on the relative abundance of the two algal spe- cies (expressed as percentages of symbiont cells that are zooxanthellae), we sampled all sites except one on subsequent mornings of a single summer low-tide se- ries (21-29 June, 1994). Given the relative inaccessi-
140
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions

Dual symbiosis in sea anemones
Table 1. Collection sites for a latitudinal survey of algal symbionts in the sea anemones Anthopleura elegantissima and A. xanthogrammica. Except as noted, 40 anemones were sampled at each site (10 high intertidal and 10 low intertidal for each species). See text for details of sampling protocols.
Latitude Sampling site Notes
48?20'N Tatoosh Island, WA A.e. collected July 1994 A.x. collected July 1995
44?20'N Strawberry Hill, OR 42?50'N Cape Blanco, OR 40?30'N Cape Mendocino, CA A.e. not found 38?05'N Dillon Beach, CA 36?25'N Pacific Grove, CA 34?40'N Point Sal, CA 32?30'N Point Loma, CA High A.x. not found 30?25'N Punta Banda, BC, Mexico Only large solitary A. sp. sampled
bility of Tatoosh Island (48?N), the northernmost site, we sampled Anthopleura elegantissima there at the be-
ginning of the following low tide series, -10 d later in July 1994; A. xanthogrammica at Tatoosh were sam- pled in July 1995.
At each site, we excised tissue from 20 polyps of A. elegantissima and 20 polyps of A. xanthogrammica (but see Table 1), with samples for each species equal- ly divided between low intertidal (MLLW -0.5 to +0.5 m) and high intertidal (MLLW +1.5 to +2.5 m) habitats. These two anemone species are easily distin- guishable on several morphological features, and be- cause A. elegantissima is small and clonal and A. xan-
thogrammica is large and solitary. An additional symbiotic anemone, Anthopleura sp. (McFadden et al. 1997), formerly known as the large solitary form of A. elegantissima, occurs at sites south of Bodega Bay, California (Hand 1955; Francis 1979; McFadden et al. 1997), but was sampled only at the southernmost site, Punta Banda, where clonal individuals of A. elegan- tissima were not found.
To minimize the pseudoreplication that would result from repeatedly sampling clonemates of A. elegantis- sima, we gathered anemones from clonal aggregations on boulders at least 5 m apart and surrounded by un- inhabitable soft-sediment substrate. On shorelines without isolated boulders, we collected from aggre- gations separated by a distance of at least 5 m. When- ever possible, we collected the individual of A. xan- thogrammica nearest the clone of A. elegantissima sampled (methods as in McFadden et al. 1997). When sampling anemones >5 cm in basal diameter (A. xan- thogrammica and, at the southernmost site only, the large solitary Anthopleura sp.), we used dive shears to remove several (4-10) tentacles and placed them to- gether into individually labeled 2-ml vials. Small clon- al anemones (A. elegantissima, typically 1-3 cm in basal diameter) were gently pried from the substrate
and cut into halves, each of which was placed into an individually labeled 2-ml vial. Anemone tissues were flash-frozen in liquid nitrogen at the end of each day's tide (within 2-4 hr of collection) and stored frozen pending subsequent processing.
To estimate percentages of symbiotic zooxanthellae and zoochlorellae, field-collected anemone tissues con- taining endosymbionts were homogenized in their vi- als by hand pestle in 1-2 ml of filtered seawater per vial. For each sample, we mixed the resultant homog- enate thoroughly before counting algal cells at 400X magnification on a stage hemocytometer. Six replicate microscope fields of homogenate were counted from each vial, representing at least 1000 algal cells per anemone. Zooxanthellae and zoochlorellae were visu- ally scored, and mean relative percentages of each alga were calculated for all combinations of species, inter- tidal height, and latitude. Algal cells are readily distin- guished under a light microscope: zoochlorellae are bright green and smaller (6-8 jLm cell diameter), whereas zooxanthellae are olive brown and larger (10- 12 jim cell diameter).
Results
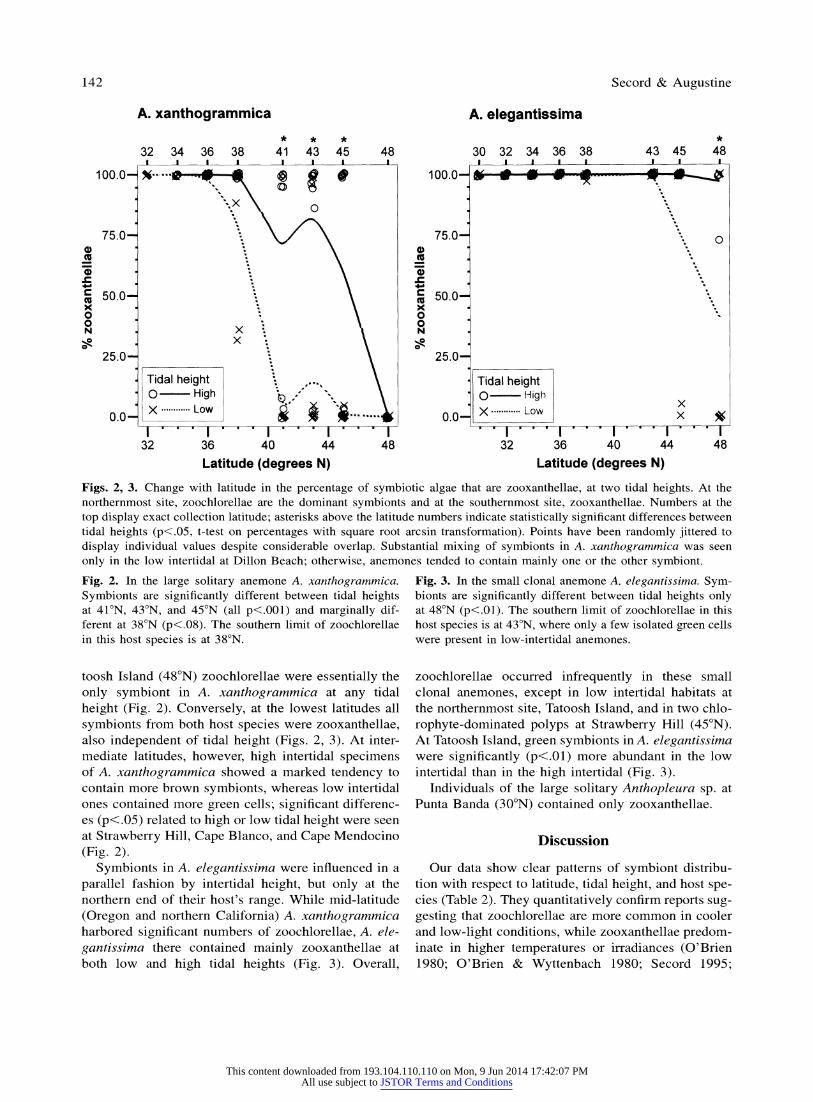
Zooxanthellae and zoochlorellae were not distrib- uted randomly with respect to latitude, tidal height, and anemone host species (Figs. 2, 3). Relative abun- dances of the two algae are expressed as mean per- centages of total endosymbiont cells that are zooxan- thellae. In both host species, zooxanthellae were the exclusive symbiont at the lowest latitudes (south of 38?N), but zoochlorellae extended much farther south in Anthopleura xanthogrammica (38?N) than in A. ele-
gantissima (43?N). Green symbionts occurred more frequently at higher
latitudes. This pattern was accentuated low in the in- tertidal and also in large-bodied hosts; indeed, at Ta-
141
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions
Secord & Augustine
A. xanthogrammica A. elegantissima *
32 34 36 38 41 i . . . i
*
43 !
* 45
! 48
! 30 32 34 36 38
i . . . I
*
43 45 48 1 * 1
O 0
Tidal height 0- High X ............ Low
X
:
32 36 40 44
Latitude (degrees N)
32 36 40 44
Latitude (degrees N)
Figs. 2, 3. Change with latitude in the percentage of symbiotic algae that are zooxanthellae, at two tidal heights. At the northernmost site, zoochlorellae are the dominant symbionts and at the southernmost site, zooxanthellae. Numbers at the top display exact collection latitude; asterisks above the latitude numbers indicate statistically significant differences between tidal heights (p<.05, t-test on percentages with square root arcsin transformation). Points have been randomly jittered to display individual values despite considerable overlap. Substantial mixing of symbionts in A. xanthogrammica was seen only in the low intertidal at Dillon Beach; otherwise, anemones tended to contain mainly one or the other symbiont.
Fig. 2. In the large solitary anemone A. xanthogrammica. Symbionts are significantly different between tidal heights at 41?N, 43?N, and 45?N (all p<.001) and marginally dif- ferent at 38?N (p<.08). The southern limit of zoochlorellae in this host species is at 38?N.
toosh Island (48?N) zoochlorellae were essentially the
only symbiont in A. xanthogrammica at any tidal
height (Fig. 2). Conversely, at the lowest latitudes all
symbionts from both host species were zooxanthellae, also independent of tidal height (Figs. 2, 3). At inter- mediate latitudes, however, high intertidal specimens of A. xanthogrammica showed a marked tendency to contain more brown symbionts, whereas low intertidal ones contained more green cells; significant differenc- es (p<.05) related to high or low tidal height were seen at Strawberry Hill, Cape Blanco, and Cape Mendocino
(Fig. 2). Symbionts in A. elegantissima were influenced in a
parallel fashion by intertidal height, but only at the northern end of their host's range. While mid-latitude
(Oregon and northern California) A. xanthogrammica harbored significant numbers of zoochlorellae, A. ele-
gantissima there contained mainly zooxanthellae at both low and high tidal heights (Fig. 3). Overall,
Fig. 3. In the small clonal anemone A. elegantissima. Sym- bionts are significantly different between tidal heights only at 48?N (p<.01). The southern limit of zoochlorellae in this host species is at 43?N, where only a few isolated green cells were present in low-intertidal anemones.
zoochlorellae occurred infrequently in these small clonal anemones, except in low intertidal habitats at the northernmost site, Tatoosh Island, and in two chlo-
rophyte-dominated polyps at Strawberry Hill (45?N). At Tatoosh Island, green symbionts in A. elegantissima were significantly (p<.01) more abundant in the low intertidal than in the high intertidal (Fig. 3).
Individuals of the large solitary Anthopleura sp. at Punta Banda (30?N) contained only zooxanthellae.
Discussion
Our data show clear patterns of symbiont distribu- tion with respect to latitude, tidal height, and host spe- cies (Table 2). They quantitatively confirm reports sug- gesting that zoochlorellae are more common in cooler and low-light conditions, while zooxanthellae predom- inate in higher temperatures or irradiances (O'Brien 1980; O'Brien & Wyttenbach 1980; Secord 1995;
0
100.0-
75.0- 0
m
c 50.0- x O 0 N
25.0-
100.0-
75.0-
x x
Tidal height O-- High X ............ Low
0 m
.: c 50.0- x 0
o N
25.0-
0.0- 0.0-
48
I I a I I s
. . a I a . a . . . . . a . a I
142
I 48
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions

Dual symbiosis in sea anemones
Table 2. Summary of factors influencing the distribution of two symbionts in sea anemone hosts.
Factors that favor Factors that favor zoochlorellae zooxanthellae
Laboratory Low temperature and light High temperature or light (Verde & McCloskey 1996; Saunders & Muller-Parker 1997)
Field Low intertidal height High intertidal height (Secord 1995; Dingman 1998) High latitude Low latitude
Larger (cooler) hosts Smaller (warmer) hosts
Saunders & Muller-Parker 1997). Specifically, zooxan- thellae were associated with southern latitudes, where they were the exclusive symbiont (south of 38?N), most likely due to physical conditions that promote higher photosynthetic rates and faster growth of that alga. Smaller polyps and those in the high intertidal tend to reach warmer temperatures (see below; Ding- man 1998). Anemones are also likely to experience higher temperatures at more southerly latitudes. Under all these conditions, anemones tended to contain more zooxanthellae (Figs. 2, 3). This is consistent with the hypothesis that physical conditions play a significant role in determining the field distribution of the two symbionts in Anthopleura. We cannot rule out the pos- sibility of biological interactions (different host spe- cies) contributing to the observed patterns, however.
The paradigm of light or temperature effects on the two symbionts is consistent with patterns observed in A. elegantissima along a light gradient within an in- tertidal cave at Tatoosh Island, WA, where zooxan- thellae occur near the exposed mouth of the cave, zoochlorellae in its shaded midsection, and aposym- biotic (alga-free) anemones in its darkest reaches (Se- cord 1995). Indeed, symbiont replacement in aposym- biotic anemones of this species or in those with a mixture of both symbionts, under altered light or tem- perature regimes, occurs on a time scale of weeks in controlled laboratory conditions (Saunders & Muller- Parker 1997) and of months in field transplant exper- iments (Secord 1995).
Data presented here suggest that patterns which hold in intertidal caves and in the laboratory (for symbionts of A. elegantissima) also apply to the effects of host body size, latitude, and tidal height as correlates of light and temperature conditions experienced by en- dosymbionts. Specifically, small clonal polyps of A. elegantissima average 1-2 orders of magnitude smaller by weight or volume than their solitary congeners (A. elegantissima, ash free dry weights 0.006-1.0 g; A. xanthogrammica, 3.0-60.0 g: Sebens 1981). Internal body temperature of host anemones varies in turn with body size, with smaller animals heating up faster dur-
ing extreme spring and summer low tides than larger ones, especially when exposed to direct sunlight (Dingman 1998). Therefore, symbionts in small clonal polyps of A. elegantissima are more likely to experi- ence extreme temperatures than those in A. xantho- grammica, whose polyps are large and solitary.
Of the three interacting conditions for algal growth examined here (host taxonomy, tidal height, and lati- tude), those expected to be associated with greater pro- portions of zoochlorellae (which grow faster and pho- tosynthesize more efficiently at low light and temperatures) are large host size, high latitude, and low intertidal position. Conversely, small host size, high intertidal habitat, and low latitudes (implying warmer temperatures and perhaps more incident light) should lead to more zooxantheilae. Indeed, the south- ern limit of zoochlorellae appears to be Dilion Beach, California, at about 38?N; south of there, only zooxan- thellae were found. Furthermore, at the southernmost sites where zoochlorellae occur, they persist only in large solitary anemones (A. xanthogrammica) and at low tidal heights. Similarly, at the southernmost site where zoochlorellae occur in A. elegantissima (farther north at 43?N) they occur only in the low intertidal.
It would be instructive to do more detailed sampling at this apparent transition zone in central and northern California, to identify some of the ecological factors that allow zoochlorellae to persist at low latitudes. For example, do wholly zoochlorellate anemones occur subtidally in central California, or undetected in caves or shaded microhabitats? Do isolated populations of zoochlorellae exist even farther south in appropriate microhabitats, a condition that could present hosts with an alternative symbiont if environmental conditions changed? The southernmost site where we collected zoochlorellae is about 4? north of the putative bound- ary between biogeographic provinces at Point Concep- tion, CA (Ricketts et al. 1985). Coincidentally, 38?N is also the approximate northern range limit of the large solitary Anthopleura sp., which apparently lacks zoochlorellae throughout its range (Francis 1979; McFadden et al. 1997; V. Pearse, pers. comm.). Final-
143
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions

Secord & Augustine
ly, sampling north of Tatoosh Island (48?N) could identify any complementary transition zone from anemones with mixed symbionts to those with only zoochlorellae, perhaps reflecting the combined effects of symbiont geographic and tidal distribution shown here.
A. xanthogrammica demonstrates most clearly the interplay between the effects of latitude and tidal height on symbiont ratios (Fig. 2). At the highest and lowest latitudes, there was no relationship between tid- al height and symbiont ratios; at these latitudes, anem- one populations contained either zooxanthellae or zoochlorellae, but not both. We suggest, however, that at intermediate latitudes, the warming effects of high tidal height may mimic those of low latitude, and the cooling effects of low tidal height produce the pattern seen farther north. Indeed, at low tide large anemones may act as algal insulators, maintaining internal tem- peratures closer to that of ambient seawater (Dingman 1998), thereby buffering their symbionts from temper- ature extremes. Green symbionts, which experience expulsion from anemones at high temperatures (O'Brien & Wyttenbach 1980; Saunders & Muller- Parker 1997), may thus persist in A. xanthogrammica even in relatively warm (high intertidal or southern) habitats.
We have not yet addressed the prospect of zonation of algae within host anemones as well as with changes in microhabitat and latitude. If we had sampled iden- tical body regions for both host species, we may have seen a different pattern of symbiont distribution. In Washington State, a higher proportion of zoochlorellae occur in the column of anemones than in the tentacles (B. Timmerman, pers. comm.); a gradient of increasing zooxanthella prevalence may even occur from the proximal to distal portions of individual tentacles (J. Ly, pers. comm.). Since we sampled entire small pol- yps and only tentacles for large ones, our sampling of A. xanthogrammica may not be truly representative of symbiont distribution in the entire animal.
Under changing oceanic conditions, hosts may lose one symbiont species and gain another that may be physiologically adapted to the altered environment (Buddemeier & Fautin 1993; Ware et al. 1996). Her- matypic corals and sponges apparently do this with different zooxanthella genotypes (Buddemeier 1997; Rowan et al. 1997; Hill & Wilcox 1998). Some natural flatworm populations contain two or more symbiont taxa, and so have the potential to change their host- symbiont associations as well (Douglas 1985). Among cnidarians, only Anthopleura species harbor two mi- croscopically distinguishable taxa of symbionts. These symbiotic associations are environmentally sensitive (e.g., Engebretson & Martin 1994; Martin et al. 1996)
and are unusual among documented marine symbioses in that they vary along geographic gradients (but see for terrestrial examples: Longino 1989; Montchamp- Moreau et al. 1991; Rollinger & Langenheim 1993; Parker & Spoerke 1998). Clear population-level bio- logical signals of regional climate change are relatively rare (Paine 1986; Barry et al. 1995; Parmesan 1996), especially when coupled with sensitive multiple sym- bioses (Secord & Kareiva 1996; Secord 1998). To the extent that algal ratios in Anthopleura spp. may shift with changing environmental conditions, the symbio- ses described here constitute intimate species interac- tions that may measurably respond to intermediate- term (benthic signals of El Ninio events; Paine 1986) or long-term (coastal warming; Barry et al. 1995) en- vironmental change.
While our data suggest that the type of symbionts found in anemones is strongly associated with physical conditions correlated with latitude and tidal height, they do not rule out the possibility that other factors play a role. For example, hosts may vary either genetically or taxonomically in their symbiont specificity, or symbi- onts may vary genetically in their affinity for host spe- cies. Our data also do not settle the question of whether it is the averages or the extremes of physical conditions (e.g., temperature and irradiance) covarying with sym- biont distribution which explain the patterns reported here, nor do they address the possibility that UV light is a factor. Overall, however, our data support the idea that symbiont species' habitat distributions should mir- ror their physiological tolerances. In addition, symbiont distributions appear to reflect the combined effects of local physical factors (tidal height and within-host con- ditions) and geographically determined ones (latitude). This four-species host-symbiont system builds a strong case for physical regulation of symbiont populations, reflecting algal biology rather than strict control by anemone hosts.
Acknowledgments. We are grateful to D. Brumbaugh, R. Brodie, and B. Pernet for assistance in the field, and to R. Grosberg, B. Helmuth, M. Kalton, C. McFadden, G. Muller- Parker, R. Paine, D. St. Clair, P. St. Clair, and the directors and staffs of Friday Harbor Laboratories, Shannon Point Ma- rine Center, and Hopkins Marine Station, for logistical or other support. We thank the United States Coast Guard, Big Salmon Resort, and the Makah Tribal Nation for access to study sites at Tatoosh Island. Comments from A. Adams, C. Greengrove, B. Kalikoff, M. Kucher, G. Muller-Parker, R. Paine, V. Pearse, B. Pughe, and two anonymous reviewers substantially improved earlier versions of this manuscript. G. Muller-Parker and R.K. Trench clarified the taxonomic status of the symbionts, and S. Perdue helped improve the
figures. Funding was provided by grants to D.S. from Sigma Xi, the Lerner Gray Fund of the American Museum of Nat-
144
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions

Dual symbiosis in sea anemones
ural History, and the Phycological Society of America; a National Science Foundation Graduate Fellowship to D.S.; a Mellon Foundation fellowship to L.A.; and National Sci- ence Foundation grants to R.T Paine. This is contribution No. 1001 of the Center for the Study of Invertebrates and Society at the University of Washington, Tacoma.
References
Barry JP, Baxter CH, Sagarin RD, & Gilman SE 1995. Climate-related, long-term faunal changes in a California rocky intertidal community. Science 267: 672-675.
Blank RJ & Trench RK 1985. Speciation and symbiotic di- noflagellates. Science 229: 656-658.
Buchner P 1965. Endosymbiosis of Animals with Plant Mi- croorganisms. Wiley Interscience Publ., New York. 909 PP.
Buddemeier RW 1997. Making light work of adaptation. Na- ture 388: 229-230.
Buddemeier RW & Fautin DG 1993. Coral bleaching as an adaptive mechanism: a testable hypothesis. BioScience 43: 320-326.
Dayton PK 1971. Competition, disturbance, and community organization: the provision and subsequent utilization of space in a rocky intertidal community. Ecol. Monogr. 41: 351-389.
Dingman HC 1998. Environmental influence on algal sym- biont populations in the sea anemone Anthopleura elegan- tissima. Master's thesis, Western Washington University. 92 pp.
Douglas AE 1985. Growth and reproduction of Convoluta roscoffensis containing different naturally occurring algal symbionts. J. Mar. Biol. Assoc. UK 65: 871-879.
Engebretson H & Martin KLM 1994. Effects of decreased salinity on expulsion of zooxanthellae in the symbiotic sea anemone Anthopleura elegantissima. Pac. Sci. 48: 446- 457.
Engebretson HP & Muller-Parker G 1999. Translocation of photosynthetic carbon from two algal symbionts to the sea anemone Anthopleura elegantissima. Biol. Bull. 197: 72- 81.
Fitt WK, Pardy RL, & Littler MM 1982. Photosynthesis, respiration, and contribution to community productivity of the symbiotic sea anemone Anthopleura elegantissima (Brandt 1835). J. Exp. Mar. Biol. Ecol. 61: 213-232.
Francis L 1979. Contrast between solitary and clonal life- styles in the sea anemone Anthopleura elegantissima. Amer. Zool. 19: 669-681.
Hand C 1955. The sea anemones of central California. Part II. The endomyarian and mesomyarian anemones. Was- mann J. Biol. 13: 37-99.
Hill M & Wilcox T 1998. Unusual mode of symbiont re- population after bleaching in Anthosigmella varians: ac- quisition of different zooxanthellae strains. Symbiosis 25: 279-289.
Kozloff EN 1983. Seashore Life of the Northern Pacific Coast, 2nd edition. University of Washington Press, Se- attle. 370 pp.
Longino J 1989. Geographic variation and community struc-
ture in an ant-plant mutualism: Azteca and Cecropia in Costa Rica. Biotropica 21: 126-132.
Martin KLM, Lawson MC, & Engebretson H 1996. Adverse effects of hyposalinity from stormwater runoff on the ag- gregating anemone, Anthopleura elegantissima, in the ma- rine intertidal zone. Bull. So. Cal. Acad. Sci. 95: 46-51.
McCloskey LR, Cove TG, & Verde EA 1996. Symbiont ex-
pulsion from the anemone Anthopleura elegantissima (Brandt). J. Exp. Mar. Biol. Ecol. 195: 173-186.
McFadden CS, Grosberg RK, Cameron BB, Karlton DP, & Secord D 1997. Genetic relationships within and between clonal and solitary forms of the sea anemone Anthopleura elegantissima revisited: evidence for the existence of two
species. Mar. Biol. 128: 127-139.
Montchamp-Moreau C, Ferveur J-F, & Jacques M 1991.
Geographic distribution and inheritance of three cytoplas- mic incompatibility types in Drosophila simulans. Genet- ics 129: 399-407.
Muscatine L 1971. Experiments on green algae coexistent with zooxanthellae in sea anemones. Pac. Sci. 25: 13-21.
O'Brien TL 1980. The symbiotic association between intra- cellular zoochlorellae (Chlorophyceae) and the coelenter- ate Anthopleura xanthogrammica. J. Exp. Zool. 211: 343- 355.
O'Brien TL & Wyttenbach CR 1980. Some effects of tem-
perature on the symbiotic association between zoochlorel- lae (Chlorophyceae) and the sea anemone Anthopleura xanthogrammica. Trans. Am. Microsc. Soc. 99: 221-225.
Paine RT 1986. Benthic community-water column coupling during the 1982-1983 El Nifio. Are community changes at high latitudes attributable to cause or coincidence? Lim- nol. Oceanogr. 31: 351-360.
Parker MA & Spoerke JM 1998. Geographic structure of
lineage associations in a plant-bacterial mutualism. J. Evol. Biol. 11: 549-562.
Parmesan C 1996. Climate and species' range. Nature 382: 765-766.
Ricketts EF, Calvin J, Hedgpeth JW, & Phillips DW 1985. Between Pacific Tides, 5th edition. Stanford University Press, Stanford. 652 pp.
Rollinger JL & Langenheim JH 1993. Geographic survey of
fungal endophyte community composition in leaves of coastal redwood. Mycologia 85: 149-156.
Rowan R 1991. Molecular systematics of symbiotic algae. J. Phycol. 27: 661-666.
Rowan R & Knowlton N 1995. Intraspecific diversity and
ecological zonation in coral-algal symbiosis. Proc. Natl. Acad. Sci. USA 92: 2850-2853.
Rowan R & Powers DA 1991a. Molecular genetic identifi- cation of symbiotic dinoflagellates (zooxanthellae). Mar. Ecol. Prog. Ser. 71: 65-73.
199 b. A molecular genetic classification of zooxan- thellae and the evolution of animal-algal symbioses. Sci- ence 251: 1348-1351.
1992. Ribosomal RNA sequences and the diversity of symbiotic dinoflagellates (zooxanthellae). Proc. Natl. Acad. Sci. USA 89: 3639-3643.
Rowan R, Knowlton N, Baker A, & Jara J 1997. Landscape
145
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions

Secord & Augustine
ecology of algal symbionts creates variation in episodes of coral bleaching. Nature 388: 265-269.
Saunders BK & Muller-Parker G 1997. The effects of tem-
perature and light on two algal populations in the temper- ate sea anemone Anthopleura elegantissima (Brandt, 1835). J. Exp. Mar. Biol. Ecol. 211: 213-224.
Sebens KP 1981. The allometry of feeding, energetics, and
body size in three sea anemone species. Biol. Bull. 161: 152-171.
Secord DL 1995. Host specificity and symbiotic interactions in sea anemones. Ph.D. dissertation, University of Wash-
ington. 89 pp.
1998. Symbioses as species interactions in basic and
applied ecology. Symbiosis News 1(3): 4-9. Secord D & Kareiva P 1996. Perils and pitfalls in the host
specificity paradigm. BioScience 46: 448-453. Shick JM 1991. A Functional Biology of Sea Anemones.
Chapman & Hall, London. 395 pp. Verde EA & McCloskey LR 1996. Photosynthesis and res-
piration of two species of algal symbionts in the anemone Anthopleura elegantissima (Brandt, 1835). J. Exp. Mar. Biol. Ecol. 195: 187-202.
Ware JR, Fautin DG, & Buddemeier RW 1996. Patterns of coral bleaching: modeling the adaptive bleaching hypoth- esis. Ecol. Modeling 84: 199-214.
146
This content downloaded from 193.104.110.110 on Mon, 9 Jun 2014 17:42:07 PMAll use subject to JSTOR Terms and Conditions